Embed Size (px)
Citation preview

Vol. 46, No. 5APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Nov. 1983, P. 1118-11240099-2240/83/111118-07$02.00/0Copyright © 1983, American Society for Microbiology
Denitrification and Assimilatory Nitrate Reduction inAquaspirillum magnetotacticumDENNIS A. BAZYLINSKI AND R. P. BLAKEMORE*
Department of Microbiology, University ofNew Hampshire, Durham, New Hampshire 03824
Received 13 April 1983/Accepted 15 August 1983
Aquaspirillum magnetotacticum MS-1 grew microaerobically but not anaerobi-cally with N03- or NH4' as the sole nitrogen source. Nevertheless, cell yieldsvaried directly with N03- concentration under microaerobic conditions. Productsof N03- reduction included NH4', N20, NO, and N2. N02- and NH2OH, eachtoxic to cells at 0.2 mM, were not detected as products of cells growing on NO3-.NO3- reduction to NH4' was completely repressed by the addition of 2 mMNH4' to the growth medium, whereas NO3- reduction to N20 or to N2 was not.C2H2 completely inhibited N20 reduction to N2 by growing cells. These resultsindicate that A. magnetotacticum is a microaerophilic denitrifier that is versatilein its nitrogen metabolism, concomitantly reducing NO3 by assimilatory anddissimilatory means. This bacterium appears to be the first described denitrifierwith an absolute requirement for 02. The process of N03 reduction appears welladapted for avoiding accumulation of several nitrogenous intermediates that aretoxic to cells.
Motile bacteria whose principal swimming di-rections are influenced by magnetic fields, in-cluding the geomagnetic field, are common insediments of diverse aquatic habitats (4, 27).Cells of the bipolarly flagellated, obligate mi-croaerophile, Aquaspirillum magnetotacticum(5, 14, 26), synthesize magnetosomes (intracellu-lar, enveloped, iron-rich crystals) consisting ofmagnetite (Fe3O4). Magnetosomes impart toeach cell a permanent magnetic dipole moment(3, 13). Cells synthesizing Fe3O4 from soluble(chelated) iron accumulate the hydrous ferricoxide, ferrihydrite (R. B. Frankel, G. C. Pa-paefthymiou, R. P. Blakemore, and W. O'Brien,Biochim. Biophys. Acta, in press). Thus, bacte-rial magnetite synthesis appears to parallel theprocess of magnetite biomineralization in chi-tons (class Mollusca), involving iron reductionand dehydration of a ferrihydrite precursor (23).Iron reduction by nitrate reductase has beensuggested for soil microorganisms (29, 30).Moreover, Sorensen (35) obtained evidence thatoxidized iron may replace N03 as a terminalelectron acceptor in microorganisms found insurface sediments.
This study was undertaken to clarify the bio-chemistry of NO3 reduction in A. magnetotac-ticum as a prelude to establishing whether en-zymes of NO3 reduction are involved in itsability to synthesize magnetite through iron re-duction under microaerobic conditions.N03- reduction in Aquaspirillum species is
poorly understood. Within this genus, A. iter-sonii and A. psychrophilum also reduce N03beyond the NO2 stage, but only the latterspecies forms visible gas (15, 20). N20 is theterminal product of N03 reduction in A. iter-sonii (7). A. fasciculus, A. gracile, and A. poly-morphum appear to reduce NO3 to N02 only(16, 20). A. dispar (ATCC 27510 and 27650) wasfound to grow anaerobically with NO3, reduc-ing it beyond the NO2 stage (21).
Cells of A. magnetotacticum grow microaero-bically with N03 or NH4' as a sole N source.N03 is reduced, forming NH3 and nitrousoxide (N20) but no detectable N02 (5, 11);Bazylinski and Blakemore, Abstr. Annu. Meet.Am. Soc. Microbiol. 1982, I 53, p. 103). Thus,this bacterium appears to assimilate products ofN03 reduction while denitrifying.True denitrifiers typically reduce 90% or more
of the available N oxide (NO3- or N02) to Ngas and couple this reduction to electron trans-port phosphorylation (6, 7). Certain non-denitri-fying N03 reducers, including strains of Esche-richia coli, produce N20 in amounts less than30% of the N oxide (6; J. M. Tiedje, personalcommunication). Because the gaseous productsof N03 reduction in Aquaspirillum specieshave not been quantified, the role of theseorganisms in denitrification is still unclear.Moreover, some non-denitrifying bacteria pro-duce N20 during NO3 reduction to NH4' (6,33, 34). Thus, another goal of this study was to
1118
on Septem
ber 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

N03- REDUCTION IN A. MAGNETOTACTICUM 1119
establish whether A. magnetotacticum can beconsidered a denitrifier by currently acceptedcriteria despite its absolute requirement for 02.
MATERIALS AND METHODS
Bacteria and growth conditions. The organism usedin this study was A. magnetotacticum MS-1. Magneto-tactic cells of this strain and those of a non-magneto-tactic variant (see below) were routinely cultured in a
growth medium containing the following (grams per
liter): tartaric acid, 0.75; KH2PO4, 0.69; NaNO3, 0.17;and sodium thioglycolate, 0.06. To each liter of thismedium were added 2.0 ml of 10mM ferric quinate (5),10 ml of vitamin mixture (36), 5 ml of mineral solution(36), and 0.1 ml of 1% (wt/vol) aqueous resazurin. Themineral solution was modified by the addition of 0.4 g
of Na2MoO4 * 2H20 per liter. Ammonium ion was
added to the medium as required, either as (NH4)2SO4or NH4Cl, as indicated. NaNO2 or NH2OH * HCI was
added to the medium as indicated. The pH of themedium was adjusted to 6.75 with NaOH added beforesterilization.
Experiments were carried out with cells culturedmicroaerobically at 30°C in stoppered 160-ml serum
vials each containing 60 ml of culture medium. 02-freeN2 or He was bubbled through the medium (ca. 500ml/min) for 15 min at room temperature before eachvial was sealed. The headspace gas of each was thenreplaced with either N2 or He after the vials were
repeatedly evacuated with the use of a vacuum mani-fold, as described previously by Balch et al. (2). Afterbeing autoclaved, the medium was anaerobic (color-less). Just before inoculation, sterile air or 02 was
added to the bottles to obtain an initial headspaceconcentration of 0.2 to 1.0% (vol/vol) 02 (200 to 1,000Pa 02)- Cells were also mass cultured in 10- to 15-literbatch cultures, as previously described (5).
Estimation of cell yield. Cell numbers were deter-mined by direct cell counts with a Petroff-Hausser cell-counting chamber. Dry cell weights were determinedby filtering culture samples through 0.2-,um polycar-bonate filters (Nuclepore Corp., Pleasanton, Calif.),which were then dried to constant weight at 60°C.
Cell magnetism. Cultures were assessed for theirmagnetism by microscopically noting the fraction ofcells, living or dead, that reversed direction when asmall magnetic stirring bar 5 to 10 cm away from themicroscope stage was rotated 180° from its initialposition. Occasionally, cells were negatively stainedwith 0.5% uranyl acetate (wt/vol; pH 4.2) and exam-
ined by electron microscopy for the presence of mag-netosomes.Use of acetylene to block N20 reduction. We used
established methods to inhibit N20 reduction withacetylene (C2H2) (12, 38). C2H2 was generated fromdistilled water and CaC2 (granular; Fisher ScientificCo., Pittsburgh, Pa.). All cultures grown with C2H2were incubated on a shaker at 30°C.
Chemical analyses. N03- was determined with a
Beckman SelectIon 2000 ion analyzer (Beckman In-struments, Inc., Irvine, Calif.). N02 was analyzedwith sulfanilamide-N-1-naphthylethylene-diamine di-hydrochloride (1). NH4' was determined by the reduc-tive amination of a-ketoglutarate (Sigma TechnicalBulletin no. 170-UV, Sigma Chemical Co., St. Louis,Mo.). Bound and free NH2OH was assayed by the
Csaky procedure (10) and by the method of Magee andBums (25).N2O, NO, and 02 were measured by gas chromatog-
raphy on a Varian series 2400 gas chromatographequipped with a 63Ni electron capture detector (VarianInstruments, Walnut Creek, Calif.). Two Porapak Qcolumns (3 mm by 1.8 m) were arranged in a series, themeshes being 80/100 and 60/80, respectively. 02-freeN2 at a flow rate of 25 ml/min was the carrier gas. Theoperating temperatures were as follows: detector,300°C; column oven, 55C; injector, 70°C. Under theseconditions, H2, He, 02, NO, C02, N20, C2H2, andwater were separated.N2 was determined with a Perkin-Elmer model
3920A gas chromatograph equipped with a thermalconductivity detector. A molecular sieve 5A column(60/80 mesh; 3 mm by 1.8 m) was used for thestationary phase. 02-free helium at a flow rate of 30ml/min was the carrier gas. The bridge current was 225mA, and the operating temperatures were as follows:detector, 130°C; injector, 120°C; column oven, 40°C.Peak areas were determined with a Hewlett-Packard
model 3390A computing integrator. For each analysis,standard curves were prepared with pure gases (ScottEnvironmental Technology, Inc.).Samples of the culture headspace gas were removed
with a gas-tight syringe (series A-2; Precision Scien-tific Co.) that had been previously flushed at leastthree times with 02-free N2 or He; the samples werethen immediately injected into the gas chromatograph.To determine the total concentration of a gaseous
product, we calculated the amount present in thesolution by using Henry's Law and published values ofsolubility coefficients (22).
Cells grown to late-exponential phase (10 liters; 8 x10' cells per ml) were harvested by continuous flowcentrifugation in a CEPA-model LE (New BrunswickScientific Co., Edison, N.J.) electrically driven centri-fuge equipped with water cooling. Cells were washedseveral times with 50 mM potassium phosphate buffer(pH 6.90) by centrifugation (11,000 x g for 15 min at5°C) and dried to constant weight in vacuo over CaSO4at 110°C. Dried cells were analyzed for total protein,amino acids, and their elemental composition.For amino acid analysis, samples of whole cells
were hydrolyzed with HCl and treated with 10.74 mMaqueous Na2 EDTA to remove iron. Amino acids andintracellular NH4' were determined by using a single-column acid-hydrolysate methodology (Spinco Appli-cation Note AN-001, 4/77; Beckman Instruments,Spinco Division) with a Beckman model 118CL aminoacid analyzer equipped with a Varian model CDS-111C peak integrator. The analyzer-integrator systemwas calibrated with a Beckman standard referencemixture.
Total cell protein was determined by the method ofLowry et al. (24), with bovine serum albumin as thestandard.
Cell elemental composition was determined with aPerkin-Elmer model 240B elemental analyzer withacetanilide as the standard.
RESULTSEffect of nitrogenous compounds on growth and
magnetite synthesis. NH4+ and N03- are used assole sources of nitrogen by A. magnetotacticum
VOL. 46, 1983
on Septem
ber 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

1120 BAZYLINSKI AND BLAKEMORE
N20
NaNO2H20H-HCI
I~~~~~~~1 1\0 24 48 72 96 120 144
HoursFIG. 1. Growth response of A. magnetotacticum
to added nitrogen compounds. At 48 h, cultures previ-ously grown without a source of fixed nitrogen wereprovided with NH4Cl, (NH4)2SO4, NaNO3, N20,NaNO2, or NH2OH HCI, each at a final concentra-tion of 2 mM N. The controls received an equalvolume of anaerobic growth medium minus an Nsource. Symbols and bars represent means and stan-dard deviations, respectively, obtained with triplicatecultures.
(26) (Fig. 1). After an initial lag period, thegrowth rate observed with N03 was muchhigher than that with NH4' (Fig. 1). A lag periodwas not observed with either (NH4)2SO4 or
NH4C1. Higher cell yields were obtained withNaNO3- (1.2 x 108 cells per ml) than with(NH4)2SO4 (2.9 x 107 cells per ml) or NH4Cl (2.1X 107 cells per ml). Cell lysis became apparent
72 h after the addition of N03- or NH4' to theculture medium (Fig. 1).The effect of known intermediates of NO3
reduction by other bacteria was determined.Free NH2OH or N02 (2 mM) were toxic tocells and produced lysis (Fig. 1). When added togrowth medium containing no fixed nitrogensource, each compound exhibited toxicity even
at a concentration of 0.2 mM (data not shown).However, N02 at less than 1 mM was not toxicfor cells actively growing on N03 . N20 (120
,umols added to the headspace of shaking cul-tures) had no detectable effect on the growth ofcells in the absence of a combined nitrogensource (Fig. 1).Although cultures grown with NH4+ or NO3
each contained magnetotactic cells, those grownwith NH4' frequently contained a higher pro-portion of nonmagnetotactic cells than thosegrown with N03.
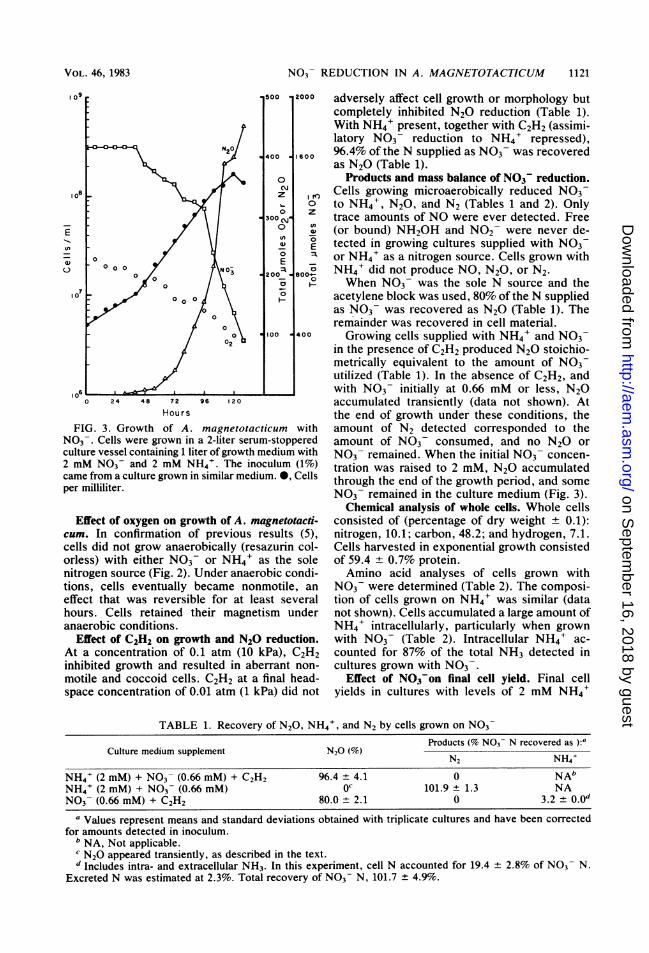
Cultures grown with NO3 under microaero-bic conditions frequently showed a biphasicgrowth pattern (Fig. 1 and 2). Figure 3 shows theutilization of N03 and 02 as well as the produc-tion ofN02 , NQ, and N20 by growing cells. 02steadily disappeared throughout growth. N03-,on the other hand, was utilized most extensivelyafter 40 h. The accumulation of N20 appeared tocorrelate with the extent of NO3 utilization. Atabout 40 h, the cell growth rate increased from aculture doubling time of about 40 h to a doublingtime of 16 h. Traces ofN02 or NO or both wereoccasionally detected during growth of A. mag-netotacticum, but neither of these accumulatedin significant amounts. Of note, cells activelyusing N03 also continued to consume 02-
10 NOj
E7
10 NH
-)o60 24 48 72 96 120 144
HoursFIG. 2. Effect of 02 on the growth of A. magneto-
tacticum with or without NH4+ or N03 . To limit theintroduction of 02, 1% inocula (vol/vol) were fromcultures grown until 02 had completely disappeared.Cells used as inocula came from culture mediumsimilar to that used in the experiment. Data points andbars represent means and standard deviations, respec-tively, obtained with triplicate cultures. Symbols: 0,N03 (2 mM), microaerobic conditions (initial PO2, 0.2kPa); A, N03 (2 mM), anaerobic conditions (res-azurin colorless); O, NH4' (2 mM), microaerobicconditions (initial PO,, 0.2 kPa); 0, NH4+ (2 mM),anaerobic conditions (resazurin colorless).
APPL. ENVIRON. MICROBIOL.
on Septem
ber 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
N03- REDUCTION IN A. MAGNETOTACTICUM 1121
E
a)
0 24 48 72 96 20
Hours
FIG. 3. Growth of A. magnetotacticum withNO3-. Cells were grown in a 2-liter serum-stopperedculture vessel containing 1 liter of growth medium with2 mM N03 and 2 mM NH4'. The inoculum (1%)came from a culture grown in similar medium. 0, Cellsper milliliter.
Effect of oxygen on growth of A. magnetotacti-cum. In confirmation of previous results (5),cells did not grow anaerobically (resazurin col-orless) with either NO3 or NH4' as the solenitrogen source (Fig. 2). Under anaerobic condi-tions, cells eventually became nonmotile, aneffect that was reversible for at least severalhours. Cells retained their magnetism underanaerobic conditions.
Effect of C2H2 on growth and N20 reduction.At a concentration of 0.1 atm (10 kPa), C2H2inhibited growth and resulted in aberrant non-motile and coccoid cells. C2H2 at a final head-space concentration of 0.01 atm (1 kPa) did not
adversely affect cell growth or morphology butcompletely inhibited N20 reduction (Table 1).With NH4' present, together with C2H2 (assimi-latory N03 reduction to NH4' repressed),96.4% of the N supplied as N03 was recoveredas N20 (Table 1).
Products and mass balance of N03 reduction.Cells growing microaerobically reduced N03to NH4', N20, and N2 (Tables 1 and 2). Onlytrace amounts of NO were ever detected. Free(or bound) NH2OH and NO2- were never de-tected in growing cultures supplied with NO3-or NH4' as a nitrogen source. Cells grown withNH4' did not produce NO, N2O, or N2-When NO3 was the sole N source and the
acetylene block was used, 80% of the N suppliedas NO3- was recovered as N20 (Table 1). Theremainder was recovered in cell material.Growing cells supplied with NH4' and N03-
in the presence of C2H2 produced N2O stoichio-metrically equivalent to the amount of N03utilized (Table 1). In the absence of C2H2, andwith N03 initially at 0.66 mM or less, N2Oaccumulated transiently (data not shown). Atthe end of growth under these conditions, theamount of N2 detected corresponded to theamount of N03 consumed, and no N20 orN03 remained. When the initial NO3 concen-tration was raised to 2 mM, N2O accumulatedthrough the end of the growth period, and someN03 remained in the culture medium (Fig. 3).Chemical analysis of whole cells. Whole cells
consisted of (percentage of dry weight + 0.1):nitrogen, 10.1; carbon, 48.2; and hydrogen, 7.1.Cells harvested in exponential growth consistedof 59.4 ± 0.7% protein.Amino acid analyses of cells grown with
N03 were determined (Table 2). The composi-tion of cells grown on NH4' was similar (datanot shown). Cells accumulated a large amount ofNH4' intracellularly, particularly when grownwith NO3 (Table 2). Intracellular NH4' ac-counted for 87% of the total NH3 detected incultures grown with NO3.
Effect of NO3 on final cell yield. Final cellyields in cultures with levels of 2 mM NH4'
TABLE 1. Recovery of N20, NH4', and N2 by cells grown on NO3-Products (to N03- N recovered as ):a
Culture medium supplement N20 (%)N2 NH4+
NH4+ (2 mM) + NO3- (0.66 mM) + C2H2 96.4 + 4.1 0 NAbNH4+ (2 mM) + N03- (0.66 mM) OC 101.9 + 1.3 NAN03- (0.66 mM) + C2H2 80.0 + 2.1 0 3.2 +±.od
a Values represent means and standard deviations obtained with triplicate cultures and have been correctedfor amounts detected in inoculum.
b NA, Not applicable.c N20 appeared transiently, as described in the text.d Includes intra- and extracellular NH3. In this experiment, cell N accounted for 19.4 + 2.8% of N03- N.
Excreted N was estimated at 2.3%. Total recovery of N03- N, 101.7 + 4.9%.
VOL . 46, 1983
091
on Septem
ber 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

1122 BAZYLINSKI AND BLAKEMORE
TABLE 2. Amino acid composition of whole cellsof A. magnetotacticum grown microaerobically with
N03 (2 mM) as the sole N sourceAmino acid nmol/mg of cell (dry wt)
Alanine...................... 560.4Glycine...................... 475.9Aspartic acid................. 421.9Leucine ..................... 405.7Glutamic acid ................ 403.7Valine....................... 337.5Lysine ...................... 291.3Threonine ................... 247.8Serine ....................... 236.9Arginine ..................... 216.6Isoleucine ................... 209.8Proline ...................... 198.9Phenylalanine ................ 177.0Methionine .................. 112.9Tyrosine..................... 101.1Histidine .................... 97.6
% Recovery of proteina.......... 93.9%
Ammonia .................... 1,761.2
a Protein was 59.4% of the cell dry weight.
sufficient to repress assimilatory N03- reduc-tion (Table 1) were higher with larger amounts ofN03 (Fig. 4). A substantial change in cell massoccurred when N03 was raised from 1 to 10mM. No corresponding increase in cell numbersover this range of N03 values was detected.Cells grown with 10 mM N03 or more wereabnormally long (20 to 50 ,um) and poorly motile.At lower NO3 concentrations, cultures con-tained cells that were smaller (2 to 10 ,um),actively motile, and magnetotactic. Cell growthwas inhibited at NO3 levels above 40 mM.
DISCUSSIONA. magnetotacticum MS-1 cells synthesize all
of their required nitrogenous compounds denovo from NH4+ or N03 ions. Because theygrow with N03 as a sole nitrogen source,thereby producing NH4+, this organism is capa-ble of assimilatory NO3 reduction. This capa-bility is widespread among bacteria and fungi(31) but apparently not among members of thegenus Aquaspirillum. A. itersonii and A. delica-tum are the only other members known to growwith N03 as the sole nitrogen source (16). Ofcourse, the inability of some species to growwith N03 may reflect requirements for pep-tides or other constitutents of complex mediaused in culturing them. Although it is uncertainwhether free NH2OH is produced during bacte-rial assimilatory N03 reduction (18, 31, 32, 37),NO2 has definitely been observed as a freeintermediate in other species (19, 31). We didnot detect either of these compounds during thegrowth of A. magnetotacticum with N03 , nor
did they support growth of this organism. Infact, at concentrations similar to those used byothers in culturing bacteria (37), each was toxic,producing cell lysis. Thus, the eight-electrontransfer occurring during NO3 reduction toNH3 in A. magnetotacticum may occur withoutthe production of free intermediates. This possi-bility is supported by data suggesting that a six-electron transfer was involved in reducing NO2to NH3 in E. coli (18), Achromobacter fischeri(32), and Veillonella alcalescens (37). A similarprocess in A. magnetotacticum might precludethe accumulation of NO2 or possibly othertoxic intermediates of assimilatory NO3 reduc-tion.
Assimilatory reduction of NO3- to NH3 wasrepressed by 2 mM NH4' in the culture medium,as evidenced by the complete conversion ofnitrogen supplied as N03 to an N gas (N20 inthe presence of C2H2; N2 in its absence) underthese conditions. Moreover, neither N20 nor N2was detected when N03 was omitted. This isconsistent with the well-recognized repressionof assimilatory NO3- reductase by NH4' and byother reduced nitrogenous compounds (31).These findings also suggest that a dissimilatorypathway of N03 reduction to NH4', recentlyfound to be of significance in the production ofNH4' in soils (9, 34) and in the bovine rumen(17), is not present in cells of A. magnetotacti-cum MS-1.
90
80
108~~~~~~~~~~~~7loB~~~~~~~~~~G
,8. . , o~~~~~.4
106 0
N 0O (mM)FIG. 4. Effect of initial N03- concentration on
final cell yields of A. magnetotacticum. Cells weregrown microaerobically (initial Po29 1 kPa) in 500-,mlbatch cultures. Final cell yields are reported as directcell counts (0) and dry cell weights (0). Symbols andbars represent means and standard deviations, respec-tively, of triplicate analysis.
APPL. ENVIRON. MICROBIOL.
on Septem
ber 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

N03- REDUCTION IN A. MAGNETOTACTICUM 1123
The similar amino acid composition of cellsgrown with either NH4' or N03 suggests asimilar mechanism of NH3 assimilation by eachcell type.
Production of free N02, which characterizesdissimilatory NO3 reduction by many orga-nisms (19, 31), was never apparent during deni-trification by A. magnetotacticum. Thus, cellsof strain MS-1 possess an efficient means ofreducing toxic NO2. It seems likely that therate of N02 reduction may be higher than therate of N03 reduction. Cells of A. magnetotac-ticum produced only trace amounts of NO andaccumulated N20 while reducing NO3. Theseappeared as transient intermediates and weresubsequently reduced to N2.With sufficient NH4' present to repress as-
similatory NO3 reduction, increased concen-trations of N03 resulted in increased finalgrowth yields. This suggests that NO3 reduc-tion under microaerobic conditions is coupled toenergy conservation in this organism.True denitrifiers typically reduce 90% or more
of the N oxide to N gas and couple this reductionto electron transport phosphorylation (6, 7). Bythese criteria, our data confirm that A. magneto-tacticum is indeed a denitrifier. Of the N sup-plied to cells as N03 alone, 80%o was recoveredas N gas. The remainder was recovered in cellmaterial and excreted nitrogenous products, in-cluding NH4'. Thus, under conditions in whichNO3 is the sole N source, cells of A. magneto-tacticum concomitantly carry out denitrificationand assimilatory N03 reduction to NH4+.
Cell growth with NO3 in batch culture isbiphasic. The onset of rapid cell growth ap-peared to correlate with the onset of N20 pro-duction from N03 . The data also suggest thatNO3 dissimilation commenced when the dis-solved 02 reached ca. 4.1 ,umol/liter.
Denitrification is generally associated withanoxic conditions because 02 not only inhibitsdenitrifying enzyme activity but represses syn-thesis of new denitrifying enzymes as well (19).However, some organisms tolerate limited quan-tities of 02 while denitrifying (31). Cells of A.magnetotacticum are obligately microaerophilicand do not grow anaerobically, even with N03(5). Moreover, they consume 02 while denitrify-ing. Thus, this bacterium appears to be the firstdescribed denitrifier which actually requiresrather than tolerates 02. This may reflect aspecific requirement for 02 as a substrate foroxygenases participating in cell biosynthesis(e.g., heme or lipid synthesis). However, wehave been unable to relieve this 02 requirementby growing cells in complex media or by addinghemin. We lack evidence that 02 iS specificallyrequired for N03 reduction. Moreover, ourdata do not enable us to determine whether,
under microaerobic conditions, respiration in-volving 02 and N03 as terminal electron accep-tors occurs simultaneously.
Cultures grown microaerobically with NH4'or N03 as the sole nitrogen source containsome non-magnetotactic cells. However, wehave frequently observed that cultures grownwith NH4', in contrast to those grown withN03 , contain a larger proportion of cells thatare not magnetotactic and do not contain magne-tosomes. This is consistent with the possibleinvolvement of NO3 reducing enzymes in mag-netite synthesis. Dissimilatory NO3 reductaseis an induced enzyme in most bacteria whichsynthesize it (31) and therefore would not besynthesized by cells growing with NH4' as thesole N source unless a suitable inducer (perhapseven Fe+3) was present. However, it is not yetknown whether any of the enzymes involved indenitrification in A. magnetotacticum can re-duce ferric iron.
Alternatively, the formation of bacterial mag-netite might result from the oxidation of ferroushydroxide [Fe(OH)2] coupled with a reductionof N03- or N20. This reaction can occur non-biologically at pH 8 (8, 28), although it has notyet been shown to occur enzymatically.Our results, which confirm and extend those
of Escalante-Semerena et al. (11), clearly estab-lish that A. magnetotacticum is a microaerophi-lic denitrifier. The possibility that denitrificationis a characteristic shared by other magnetotacticbacteria is an interesting one. Knowledge of thisprocess in strain MS-1 can be expected to lead tomore information concerning the ecologicalniche of these interesting organisms. We recent-ly showed that growing cells ofA. magnetotacti-cum reduce C2H2 microaerobically (Bazylinskiand Blakemore, Curr. Microbiol., in press).Thus, in addition to its capacity for assimilatoryand dissimilatory N03 reduction, this speciesalso fixes atmospheric N2. Its versatility withrespect to nitrogen metabolism may play a sig-nificant role in magnetite synthesis and can beexpected to favor its survival in microaerobicaquatic habitats.
ACKNOWLEDGMENTS
We gratefully acknowledge valuable communications, com-ments, and encouragement from J. M. Tiedje and P. Cornell ofMichigan State University and R. B. Frankel of the Massa-chusetts Institute of Technology. We are grateful to N.Blakemore and A. Geshnizgani for valuable technical assist-ance. Amino acid and elemental analyses were performedthrough the Instrumentation Center of the University of NewHampshire.
This work was supported by National Science Foundationgrants PCM 79-22224 and PCM 82-15900 and Office of NavalResearch contract N0014-80-C-0029.
LITERATURE CITED
1. American Public Health Association. 1980. Standard meth-ods for the examination of water and wastewater, 15th
VOL. 46, 1983
on Septem
ber 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

1124 BAZYLINSKI AND BLAKEMORE
ed., p. 380-383. American Public Health Association,Washington, D.C.
2. Balch, W. E., and R. S. Wolfe. 1976. New approach to thecultivation of methanogenic bacteria: 2-mercaptoethane-sulfonic acid (HS-CoM)-dependent growth of Methano-bacterium ruminantium in a pressurized atmosphere.Appl. Environ. Microbiol. 32:781-791.
3. Balkwill, D. L., D. Maratea, and R. P. Blakemore. 1980.Ultrastructure of a magnetotactic spirillum. J. Bacteriol.141:1399-1408.
4. Blakemore, R. P. 1975. Magnetotactic bacteria. Science190:377-379.
5. Blakemore, R. P., D. Maratea, and R. S. Wolfe. 1979.Isolation and pure culture of a freshwater magnetic spiril-lum in chemically defined medium. J. Bacteriol. 140:720-729.
6. Bleakley, B. H., and J. M. Tiedje. 1982. Nitrous oxideproduction by organisms other than nitrifiers or denitri-fiers. Appl. Environ. Microbiol. 44:1342-1348.
7. Bryan, B. A. 1981. Physiology and biochemistry of deni-trification, p. 67-84. In C. C. Delwiche (ed.), Denitrifica-tion, nitrification, and atmospheric nitrous oxide. JohnWiley & Sons, Inc., New York.
8. Buresh, R. J., and J. T. Moraghan. 1976. Chemical reduc-tion of nitrate by ferrous iron. J. Environ. Qual. 5:320-325. '
9. Caskey, W. H., and J. M. Tiedje. 1979. Evidence forClostridium as agents of dissimilatory reduction of nitrateto ammonium in soils. Soil. Sci. Soc. Am. J. 43:931-936.
10. Csaky, T. Z. 1948. On the estimation of bound hydroxyl-amine in biological materials. Acta Chem. Scand. 2:450-454.
11. Escalante-Semerena, J. C., R. P. Blakemore, and R. S.Wolfe. 1980. Nitrate dissimilation under microaerophilicconditions by a magnetic spirillum. Appl. Environ. Micro-biol. 40:429-430.
12. Federova, R. I., E. I. Milekhina, and N. I. Illyukhina.1973. Evaluation of the method of "gas metabolism" fordetecting extra-terrestrial life: identification of nitrogen-fixing microorganisms. Izv. Akad. Nauk SSSR, Ser. Biol.6:797-806.
13. Frankel, R. B., and R. P. Blakemore. 1980. Navigationalcompass in magnetic bacteria. J. Mag. Magn. Mater. 15-18:1562-1564.
14. Frankel, R. B., R. P. Blakemore, and R. S. Wolfe. 1979.Magnetite in freshwater magnetotactic bacteria. Science203:1355-1356.
15. Gauthier, D. K., G. D. Clark-Walker, W. T. Garrard,Jr., and J. Lascelles. 1970. Nitrate reductase and solublecytochrome c in Spirillum itersonii. J. Bacteriol. 102:797-803.
16. Hylemon, P. B., J. S. Wells, Jr., N. R. Krieg, and H. W.Jannasch. 1973. The genus Spirillum: a taxonomic study.Int. J. Syst. Bacteriol. 23:340-380.
17. Kaspar, H. F., and J. M. Tiedje. 1981. Dissimilatoryreduction of nitrate and nitrite in the bovine rumen:nitrous oxide production and effect of acetylene. Appl.Environ. Microbiol. 41:705-709.
18. Kemp, J. D., and D. E. Atkinson. 1966. Nitrite reductase
of Escherichia coli specific for reduced nicotinamideadenine dinucleotide. J. Bacteriol. 92:628-634.
19. Knowles, R. 1981. Denitrification, p. 323-369. In E. A.Paul and J. N. Ladd (ed.), Soil biochemistry, vol. 5.Marcel Dekker, Inc., New York.
20. Krieg, N. R. 1976. Biology of the chemoheterotrophicspirilla. Bacteriol. Rev. 40:55-115.
21. Krieg, N. R., and P. B. Hylemon. 1976. The taxonomy ofthe chemoheterotrophic spirilla. Annu. Rev. Microbiol.30:303-325.
22. Linke, W. F. 1965. Solubilities of inorganic and metal-organic compounds, vol. 2., 4th ed., p. 794-804. Ameri-can Chemical Society, Washington, D.C.
23. Lowenstam, H. L. 1981. Minerals formed by microorga-nisms. Science 211:1126-1131.
24. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folin phenolreagent. J. Biol. Chem. 193:265-275.
25. Magee, W. E., and R. H. Burris. 1954. Fixation of N, andutilization of combined nitrogen by Nostoc muscorum.Am. J. Bot. 41:771-782.
26. Maratea, D., and R. P. Blakemore. 1981. Aquaspirillummagnetotacticum sp. nov., a magnetic spirillum. Int. J.Syst. Bacteriol. 31:452-455.
27. Moench, T. T., and W. A. Konetzka. 1978. A novel meth-od for the isolation and study of a magnetotactic bacteri-um. Arch. Microbiol. 119:203-212.
28. Moraghan, J. T., and R. J. Buresh. 1977. Chemical reduc-tion of nitrite and nitrous oxide by ferrous iron. Soil. Sci.Soc. Am. J. 41:47-50.
29. Ottow, J. C. G. 1969. Mechanisms of iron reduction bynitrate reductase inducible aerobic organisms. Naturwis-senschaften 56:371-372.
30. Ottow, J. C. G. 1970. Selection, characterization, andiron reducing capacity of nitrate reductaseless (nit-) mu-tants of iron reducing bacteria. Z. Allg. Mikrobiol. 10:55-62.
31. Payne, W. J. 1973. Reduction of nitrogenous oxides bymicroorganisms. Bacteriol. Rev. 37:409-452.
32. Prakash, O., and J. C. Sadana. 1972. Purification, charac-terization, and properties of nitrite reductase ofAchromo-bacterfischeri. Arch. Biochem. Biophys. 148:614-632.
33. Smith, M. S. 1982. Dissimilatory reduction of NO2- toNH4' and N20 by a soil Citrobacter sp. Appl. Environ.Microbiol. 43:854-860.
34. Smith, M. S., and K. Zimmerman. 1981. Nitrous oxideproduction by non-denitrifying soil nitrate reducers. SoilSci. Soc. Am. J. 45:865-871.
35. Sorensen, J. 1982. Reduction of ferric iron in anaerobic,marine sediment and interaction with reduction of nitrateand sulfate. Appl. Environ. Microbiol. 43:319-324.
36. Wolin, E. A., M. J. Wolin, and R. S. Wolfe. 1963. Forma-tion of methane by bacterial extracts. J. Biol. Chem.238:2882-2886.
37. Yordy, D. M., and E. A. Delwiche. 1979. Nitrite reductionin Veillonella alcalescens. J. Bacteriol. 137:905-911.
38. Yoshinari, T., R. Hynes, and R. Knowles. 1977. Acetyleneinhibition of nitrous oxide reduction and measurement ofdenitrification and nitrogen-fixation in soil. Soil Biol.Biochem. 9:177-183.
APPL. ENVIRON. MICROBIOL.
on Septem
ber 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from





























