Embed Size (px)
Citation preview

Uranium Transformations in StaticMicrocosmsS H E L L Y D . K E L L Y , * , † , ‡ W E I - M I N W U , §
F A N Y A N G , | C R A I G S . C R I D D L E , §
T E R E N C E L . M A R S H , |
E D W A R D J . O ’ L O U G H L I N , †
B R U C E R A V E L , † , ⊥ D A V I D W A T S O N , 3
P H I L I P M . J A R D I N E , 3 A N DK E N N E T H M . K E M N E R †
Biosciences Division, Argonne National Laboratory,9700 South Cass Avenue, Argonne, Illinois 60439-4843,Department of Civil and Environmental Engineering, StanfordUniversity, Stanford, California 94305-4020, Microbiology andMolecular Genetics, Michigan State University, 2215Biomedical Physical Sciences, East Lansing, Michigan48824-4320, and Environmental Science Division, Oak RidgeNational Laboratory, Oak Ridge, Tennessee 37831
Received July 21, 2009. Revised manuscript receivedOctober 21, 2009. Accepted November 11, 2009.
Elucidation of complex biogeochemical processes and theireffectsonspeciationofUinthesubsurfaceiscritical fordevelopingremediation strategies with an understanding of stability. Wehave developed static microcosms that are similar to bioreductionprocess studies in situ under laminar flow conditions or insediment pores. Uranium L3-edge X-ray absorption near-edgespectroscopy analysis with depth in the microcosms indicatedthat transformation of UVI to UIV occurred by at least two distinctprocesses. Extended X-ray absorption fine structure (EXAFS)analysis indicated that initial UVI species associated with C- andP-containing ligands were transformed to UIV in the form ofuraninite and U associated with Fe-bound ligands. Microbialcommunity analysis identified putative FeIII and sulfate reducersat two different depths in the microcosms. The slow reductionof UVI to UIV may contribute the stability of UIV withinmicrocosms at 11 months after a decrease in bioreducingconditions due to limited electron donors.
1. Introduction
Uranium is a common groundwater contaminant at U.S.Department of Energy (DOE) sites and worldwide. Biore-duction through sequestering U has been investigated sincethe early 1990s (1) and tested in situ in recent years (2-6).Delivering an electron donor, such as ethanol, to U-contaminated subsurface leads to groundwater concentra-tions below the U.S. Environmental Protection Agency (EPA)maximum contaminant level of 0.126 µM (30 µg L-1) fordrinking water (6). Aqueous U is removed rapidly during
conversion of ethanol to acetate and is associated withinterspecies hydrogen transfer (7, 8) as follows:
CH3CH2OH + H2O f CH3COO- + H+ + 2H2
(∆G° ) +9.6 kJ/reaction) (1)
Until recently, reducing UVI to UIV under anaerobicconditions was commonly assumed to sequester U in thesubsurface as uraninite, on the basis of the low solubility ofuraninite (9) and its ease of formation in laboratory experi-ments (1, 10). Many studies attributed the loss of UVI fromthe aqueous phase during microbial stimulation to UIV
precipitation without direct measurement. X-ray absorptionspectroscopy (XAS) measurements for solid-phase productsof biogeochemical systems have shown that the productsare difficult to predict as the complexity of a system increases.Under some conditions, UIV reduction product exists innanoparticulate or colloidal (11, 12) forms, while under otherconditions UIV is associated with Fe or other transition metals(13-15). Other studies have shown that UIV can be solubilizedby interaction with chelating agents (16, 17). In each study,the biostimulation product failed to follow the model of bulkuraninite formation leading to U sequestration.
Our previous measurements (15) of sediment cores fromthe Oak Ridge National Laboratory Integrated Field-ResearchChallenge (IFRC) collected before biostimulation showed UVI
bound through equatorial oxygen to P- and C-containingligands. Sediments surged from wells at the same locationsafter a year of biostimulation showed U complexed with Feand P ligands. No uraninite was detected in these sediments.These results illustrate the heterogeneity of the system andthe complexity of U transformations in field materials.
XAS is uniquely suited to monitoring changes in Uspeciation (U transformations) as they occur in an intact,undisturbed microcosm during the evolution of biogeochem-ical systems, which is similar to the bioreduction process inthe subsurface under laminar flow or in sediment pores.Previous X-ray absorption near-edge spectroscopy (XANES)studies have demonstrated the utility of monitoring intactcolumns of sediment materials (18-20). In this study, wedemonstrate that U and Fe speciation, aqueous chemical,and microbiology can be monitored in microcosms duringthe evolution of biogeochemical conditions over an 11-monthperiod under slowly stimulated conditions that retain thestability of UIV.
2. Materials and MethodsAdditional Materials and Methods are given in the SupportingInformation. The sediment was taken from well FW026 ofArea 3 at the IFRC site adjacent to the former S-3 ponds atthe Y-12 National Security Complex, Oak Ridge, TN. Thegeochemical properties of the sediment and groundwater inmicrocosms are described in Table S1 of Supporting Infor-mation. The sediment contained significant amounts of U(713 mg kg-1), HCl-extractable Fe (47 mg g-1), ferrous iron(7.0 mg g-1), and total organic carbon (7.6 mg g-1). Theaqueous phase contained sulfate, 1.06 mM; bicarbonate, 1.0mM; Ca2+, 1.37 mM; and Mn2+, 0.058 mM.
Two duplicate 650-mL microcosms were prepared understerile anoxic conditions in a Coy anaerobic chamber filledwith 95% nitrogen and 5% H2 and were incubated in the Coychamber (Figure 1). The initial H2 (0.42 mmol) in themicrocosms is well suited to our measurement cycles at thesynchrotron of approximately 3 months, allowing for moni-toring of U transformations. Clone libraries of 16S rRNAsequences were constructed to identify predominant bacterial
* Corresponding author phone: (630) 270-8748; fax: (630) 252-9793; email: [email protected].
† Argonne National Laboratory.‡ Present address: EXAFS Analysis, Bolingbrook, IL 60440.§ Stanford University.| Michigan State University.⊥ Present address: Synchrotron Methods Group at Brookhaven
National Laboratory, National Institute of Standards and Technology,Bldg 535A, Upton, NY 11973.3 Oak Ridge National Laboratory.
Environ. Sci. Technol. 2010, 44, 236–242
236 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 44, NO. 1, 2010 10.1021/es902191s 2010 American Chemical SocietyPublished on Web 12/02/2009

groups (section 1.2, Supporting Information) after 5 and 11months of incubation from sediment removed from the toplayer and from 2-cm depth.
Uranium XANES measurements (section 1.3, SupportingInformation) were performed on the microcosms within 5days of initialization of the microcosms at T0, then again at3 months (T3), 5 months (T5), and 11 months (T11). UraniumEXAFS measurements were made on the microcosms andalso on centrifuged sediment packaged in a plastic holderand sealed with Kapton film and Kapton tape at T0. Becauseof the low U concentrations in the microcosms, U EXAFSmeasurements were performed sparingly at T0 and again onspecified regions of the microcosms at T11. Iron XANES andEXAFS measurements were made on the microcosms at T5.Details are found elsewhere (21). The parameters determinedby the EXAFS modeling are given in Tables S2-S4 inSupporting Information, and the EXAFS spectra �(k) areshown in Figure S1 in Supporting Information. Analyticalmethods are described in section 1.4, Supporting Information.
3. Results and Discussion3.1. Groundwater and Sediment Geochemistry. The mi-crocosms are pictured in Figure 1. The microcosm sedimentswere initially a uniform yellow-tan color. At T3, the top layerturned black, and black regions appeared throughout themicrocosm. By T11, the entire sediment had turned darkgray-green, similar to the color of bioreduced sedimentsamples during field test (5), and the black regions had grown,consistent with bioreduction in both microcosms.
The initial chemical composition of the aqueous phaseand the solids is presented in Table S1 of SupportingInformation. The sediment contained U at 713 mg kg-1 andFe at 47.1 mg g-1, with 15% of the total Fe as FeII. Aqueoussulfate concentrations decreased from 1.06 to less than 0.02
mM by T11 (Figure 2A). During this time, the aqueous UVI
concentration decreased from 11.85 to <0.1 µM (Figure 2B),which is below the EPA drinking water standard. The Ca2+
concentration declined slightly from 1.37 to 1.21 mM. ByT11, the groundwater pH had increased from 6.40 to 7.0,probably because of microbial activities of sulfate reduction,such as
SO42- + 4H2 f HS- + OH- + 3H2O (2)
Aqueous FeII concentrations remained low at <1 µM,suggesting formation of ferrous precipitates following FeIII
reduction. Based on stoichiometry, to reduce aqueous sulfateof each microcosm required about 1.6 mmol of H2 or 3.2mmol of electrons, which was much greater that the amountof H2 in headspace initially (0.42 mmol). Additional amountof electron donor would be required to reduce FeIII and UIV
in sediment. H2 (5%) in the Coy chamber gas can penetratethe polypropylene plastic wall (1 mm thick) of the microcosmsand serve as a dominant electron donor. The sedimentcontained chemical oxygen demand (COD) (22.7 mg g-1),total organic carbon (7.63 mg g-1), and FeII (7 mg g-1). Bothorganics and ferrous compounds contributed to COD. Onthe basis of the stoichiometry of oxidation of Fe2+ to Fe3+,the contribution of COD by FeII was limited (0.143 mg ofCOD/mg of Fe2+), and thus more than 95% of the COD wasfrom organics. Therefore, the organics in the sediments wasestimated as 1476 mg of COD or 184 mmol of electron pereach microcosm. The composition and biodegradability ofthe organic carbon were not determined. The source oforganic carbon could be plant residue in soil and biomassaccumulated during previous in situ biostimulation tests.The contribution of sediment organics as electron donorsmight be limited due to slow anaerobic degradation. However,
FIGURE 1. Microcosm appearance at T0 (left), T3 (top right), and T11 (bottom right).
VOL. 44, NO. 1, 2010 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 237

hydrogen-utilizing reactions, especially sulfate reductioncould significantly stimulate anaerobic microbial activity andaccelerate anaerobic degradation of soil organics.
3.2. Average U Valence State in Sediments. At T0, T3,T5, and T11, U XANES measurements were made at 2-mmintervals from the sediment-groundwater interface anddown into the sediment, with a beam size of 1 mm × 1 mmX-ray. The time evolution of the XANES spectra from a fixedsediment position is shown in Figure 3A. The UIV absorptionedge is shifted to the left of the UVI absorption edge (horizontalarrow in Figure 3A). In addition, the resonance feature abovethe absorption edge (indicated by the vertical arrow)decreases with increasing UIV, because of the loss of axialoxygen atoms of uranyl. These spectra clearly show more UIV
as the system incubates. The summary of linear combinationfitting of the U XANES spectra in Figure 3B illustrates thepercentage of UIV (with the remainder as UVI) with depthbelow the sediment-groundwater interface. At T0, only UVI
was detected in the microcosm. At T3, nearly 75% ( 10% UIV
was detected at the topmost black layer and 60% ( 10% UIV
was detected in the top 15 mm of the microcosm, with lessUIV at greater depths. At T5, nearly 80% ( 10% of the U wastransformed to UIV with an even distribution profile, and atT11 UIV reached a constant value of 80% ( 10% alongsediment depth. This observation indicates that the reductionof UVI occurred mainly in the upper part of the sediment andthen expanded toward the entire sediment profile (Figure3B).
3.3. Iron Valence and Speciation. The Fe XANES andEXAFS measurements were collected at T5. Examples of the
Fe XANES spectra are shown in Figure 4A. Although thespecific nature of the standards is reported, we do not believethat they represent the true Fe species within the samples;rather, the electronic configuration of the standards is similarto the species within the sediments. Figure 4B shows the realpart of the Fourier transform of the EXAFS spectra and themodel and the model components for Fe-O and Fe-S. Boththe XANES and EXAFS spectra require an FeS component.The results of linear combination fitting of the Fe XANESspectra are summarized in Table 1. The FeS component islargest at the sediment-water interface and decreases withdepth. This observation is consistent with the formation ofFeS-containing black top layer of sediment and a decreasein aqueous sulfate. At T0, HCl-extractable Fe in the sedimentcontained 15% FeII. This analysis also reveals that an FeIII
(hydr)oxide phase remained high (>50% ( 10%), even at T5,although up to 80% ( 10% of the UVI had been reduced toUIV.
3.4. Uranium Species. Tables S2 and S3, SupportingInformation, summarize the results of analysis of the U L3-edge EXAFS spectra, and Figure 5 illustrates the predominantU species. The U EXAFS spectra collected from sedimentpacked into a sample holder and measured directly in themicrocosm are nearly identical. At T0, both spectra weremodeled accurately with monodentate P ligands (UVI-P) andbidentate C ligands (UVI-C), as illustrated in Figure 5A. Thesediment samples from the same area had NaOH-extractablePO4
3- as high as 0.94 mg g-1 (22). At T11, the EXAFS spectrawere consistent with U associated with Fe and with UIV in
FIGURE 2. Concentrations of (A) aqueous sulfate and (B) U over the 11-month test period.
FIGURE 3. U L3-edge XANES spectra for microcosm 1 (O) and microcosm 2 (4) as the microcosms aged from T0 to T11. (A) Evolutionof the U spectra, with percentage of UVI (based on linear combination fitting), with the remainder as UIV. Arrows point in thedirection of more U(IV). (B) Percentage of UIV (based on linear combination fitting of the XANES spectra), with depth below thegroundwater-sediment interface at 0 mm. Uncertainty is approximately 10%.
238 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 44, NO. 1, 2010
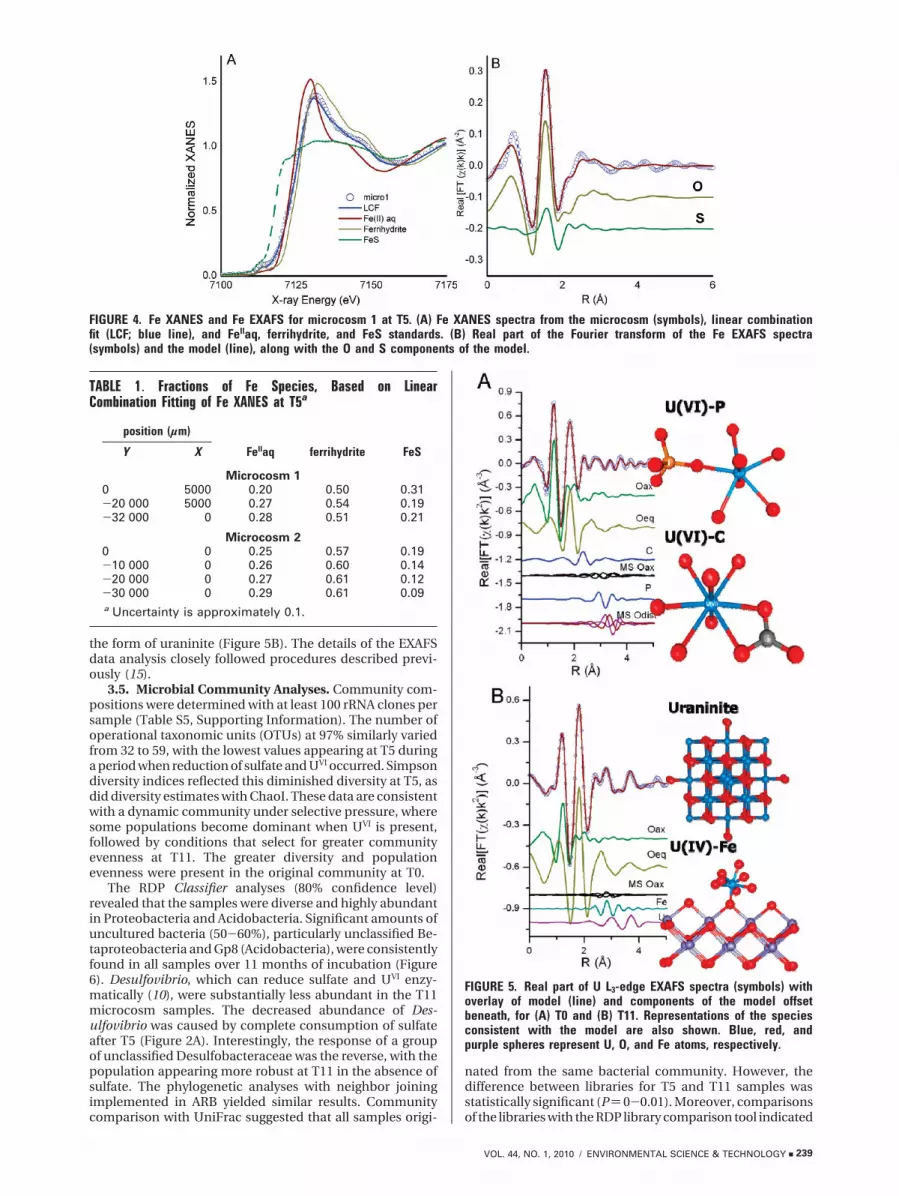
the form of uraninite (Figure 5B). The details of the EXAFSdata analysis closely followed procedures described previ-ously (15).
3.5. Microbial Community Analyses. Community com-positions were determined with at least 100 rRNA clones persample (Table S5, Supporting Information). The number ofoperational taxonomic units (OTUs) at 97% similarly variedfrom 32 to 59, with the lowest values appearing at T5 duringa period when reduction of sulfate and UVI occurred. Simpsondiversity indices reflected this diminished diversity at T5, asdid diversity estimates with ChaoI. These data are consistentwith a dynamic community under selective pressure, wheresome populations become dominant when UVI is present,followed by conditions that select for greater communityevenness at T11. The greater diversity and populationevenness were present in the original community at T0.
The RDP Classifier analyses (80% confidence level)revealed that the samples were diverse and highly abundantin Proteobacteria and Acidobacteria. Significant amounts ofuncultured bacteria (50-60%), particularly unclassified Be-taproteobacteria and Gp8 (Acidobacteria), were consistentlyfound in all samples over 11 months of incubation (Figure6). Desulfovibrio, which can reduce sulfate and UVI enzy-matically (10), were substantially less abundant in the T11microcosm samples. The decreased abundance of Des-ulfovibrio was caused by complete consumption of sulfateafter T5 (Figure 2A). Interestingly, the response of a groupof unclassified Desulfobacteraceae was the reverse, with thepopulation appearing more robust at T11 in the absence ofsulfate. The phylogenetic analyses with neighbor joiningimplemented in ARB yielded similar results. Communitycomparison with UniFrac suggested that all samples origi-
nated from the same bacterial community. However, thedifference between libraries for T5 and T11 samples wasstatistically significant (P) 0-0.01). Moreover, comparisonsof the libraries with the RDP library comparison tool indicated
FIGURE 4. Fe XANES and Fe EXAFS for microcosm 1 at T5. (A) Fe XANES spectra from the microcosm (symbols), linear combinationfit (LCF; blue line), and FeIIaq, ferrihydrite, and FeS standards. (B) Real part of the Fourier transform of the Fe EXAFS spectra(symbols) and the model (line), along with the O and S components of the model.
TABLE 1. Fractions of Fe Species, Based on LinearCombination Fitting of Fe XANES at T5a
position (µm)
Y X FeIIaq ferrihydrite FeS
Microcosm 10 5000 0.20 0.50 0.31-20 000 5000 0.27 0.54 0.19-32 000 0 0.28 0.51 0.21
Microcosm 20 0 0.25 0.57 0.19-10 000 0 0.26 0.60 0.14-20 000 0 0.27 0.61 0.12-30 000 0 0.29 0.61 0.09a Uncertainty is approximately 0.1.
FIGURE 5. Real part of U L3-edge EXAFS spectra (symbols) withoverlay of model (line) and components of the model offsetbeneath, for (A) T0 and (B) T11. Representations of the speciesconsistent with the model are also shown. Blue, red, andpurple spheres represent U, O, and Fe atoms, respectively.
VOL. 44, NO. 1, 2010 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 239

that the Desulfobacteraceae population was statisticallydifferent between the T5 samples, in which they were in lowabundance, and the T0 and T11 samples. Similarly Thioba-cillus, a FeII-oxidizing microorganism, was statistically dif-ferent in the T5 microcosm from the T0 and T11 samples,but only for the sample from the top of the microcosms. Thepresence of unclassified Deltaproteobacteria, which mightinclude unknown FeIII reducers with UVI reduction capability,was observed in the sediments.
3.6. Implications for Bioremediation. Previous studiesconducted on sediments biostimulated to reduce UVI haverevealed shifts in the structure of bacterial communities afterbiostimulation with electron donors (23-30). Denitrificationand metal and sulfate reduction were attributed to theabundance of Beta-, Gamma-, and Deltaproteobacteria,Acidobacteria, and Actinobacteria (23, 24, 29, 31). Forexample, apparent increases in the population size ofGeobacter, Geothrix, and Dechloromonas, all Deltaproteo-bacteria, have been correlated with the reduction of UVI withethanol addition at Oak Ridge (26-28, 30, 32). Similarly thisstudy identified apparent increases in Desulfocapsa, Anaer-omyxobacter, and unclassified Desulfobacteraceae (all Deltap-roteobacteria). But Geobacter remained at a relatively lowabundance throughout this study. Brodie et al. (27) detectedan increase of Betaproteobacteria during biostimulation withlactate addition, followed by a decline upon reoxidation ofU in a column study. We also noticed an initial increasefollowed by a decrease in the abundance of Betaproteobac-teria, although uranium remained in the reduced form (Figure3). As the Betaproteobacteria includes sulfate and metalreducers, the decrease in abundance late in the incubationperiod might be due to the depletion of sulfate and FeIII, aspostulated by others (27, 31). This also suggests that thestability of UIV was not correlated to the abundance change.Suzuki et al. (25) noted that the population size of Thiobacillusdecreased after biostimulation of sediment from a differentIFRC study site. We have found low abundance of Thiobacillus
sp. in the original reduced IFRC sediments and a reappear-ance after sulfate and metals had been reduced. Severalstudies reported the correlation of uranium reduction witha substantial population of Actinobacteria (23, 24, 27). Incontrast, we detected few Actinobacteria throughout theincubation period, suggesting that this population is notessential for UVI reduction. We did find abundant Acido-bacteria throughout our measurements, consistent withHwang et al. (30). Finally, Clostridiaceae were implicated byothers in metal reduction (27-29) but few members ofFirmicutes were detected in our study. Thus, the apparentrise and fall of bacterial groups is frequently, though notalways, consistent with changes in electron donor andacceptor abundances. However, this simple cause/effectrelationship plays out differently in different sediments,suggesting that other selective pressures such as interspeciescompetition, commensalism, predation, and geochemicaldifferences are factors as well.
This study suggests that 80% of UVI was transformed toUIV in the first 5 months by direct or indirect mechanismsof metal- or sulfate-reducing bacteria. This hypothesis issupported by the measurement of UIV, the decrease inaqueous sulfate concentrations, the accumulation of FeSspecies at the top layers of the sediment, and the observationof Desulfuvibrio spp. and other metal reducers in sedimentsamples. The H2 within the microcosm head space, slowdiffusion of H2 from the anoxic chamber into the microcosm,and the organic matter in the sediment could contribute tothe observed reactions. Reduction of UVI to UIV by sulfate-reducing bacteria has been reported previously (25, 33-37).The individual processes occurring during the period fromT0 to T3 and then from T3 to T5 are difficult to identify. Theinitial uranyl species (Figure 5) of UVI associated with C- andP-containing ligands has been previously identified in thesesame sediments (15) and in UVI absorption experiments (38).On the basis of previous studies (15), it may be that the uranylcarbonate species are more readily reduced than the uranyl
FIGURE 6. Top ten most abundant bacterial groups in the original sediment sample (T0) and their abundances in the microcosmsamples. The bacterial community profiles were generated by RDP Classifier at the 80% confidence level.
240 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 44, NO. 1, 2010

phosphate species and that this could create a need for twodifferent reduction mechanisms.
After 5 months, aqueous sulfate was nearly consumedand the population of Desulfovibrio spp. bacteria decreasedas expected; other bacteria known for metal reduction, suchas Geobacter spp., remained at relatively low abundance.The UIV content of the sediment increased only slightly anda relatively even distribution of UIV fraction (0.8 ( 0.1) wasobserved at T11 (Figure 3B). One possible mechanism forthe stability of UIV could be reduction of UVI to UIV by bioticallyproduced FeII oxides such as green rust. The colors of thesediments suggest reduced FeII oxides become dominant inthis stage (Figure 1). The uranium speciation indicated theassociation of U with Fe and U ligands, which was identifiedin another study of UVI reduction by abiotically producedgreen rust (14). The conditions within the microcosms werecapable of sustaining 80% reduction of U within thesediments for 11 months. Previous studies found the reoxi-dation of UIV under reducing conditions in columns filledwith sediments from Oak Ridge site with increased bicar-bonate concentrations (12.7 mM) due to microbial respirationthat caused thermodynamically favorable conditions for UIV
oxidation (39). But our microcosm tests did not experiencethe same increased carbonate concentration. Therefore, thebiogeochemical condition in this study maintained UIV upto 80% in sediment and aqueous U below 30 µg/L in thepresence of 1.21 mM Ca2+ at T11. Proper biogeochemicalconditions are key to maintaining the stability of UIV.
Complete reduction of all UVI to UIV may not be possible,because the sediments from Oak Ridge, TN, are known forcontaining a small fraction of U that is resistant to biore-duction. Over a 2-year in situ bioreduction period, no morethan 82% of UIV was reduced to UIV as determined by UXANES (6). The remaining UVI may be sequestered withinthe sediment matrix, with very poor availability for biologicalor abiotic reduction. The UIV associated with Fe within thesediments (Figure 5B) is likely in a form with low solubility,although the exact forms of UIV-Fe cannot be identified.Uraninite is known to incorporate some UVI, and the U-UEXAFS signal typical of uraninite was detected in thesediments at T11.
Elucidation of complex biogeochemical processes andtheir effects on the chemical speciation of U in the subsurfaceis critical to the development of sound remediation strategiesbased on a fundamental understanding of the system. TheXANES analysis indicates the transformation of UVI to UIV incontaminated sediments exposed to U-rich groundwater formore than 50 years of operation at the IFRC occurred by atleast two distinct processes. The aqueous U concentrationsdecreased below EPA drinking water standards, while adiverse speciation of U was sequestered within the sedimentsfor up to 11 months even though bacteria known for metalor sulfate reduction were at a very low abundance in themicrocosms. Further research is needed to identify whethermore stable forms of sequestered U require slight changesin the geochemical conditions to slowly form stable speciesof U within sediments.
AcknowledgmentsThis work was supported under Contract DE-AC02-06CH11357 by the DOE Office of Science, Office of Biologicaland Environmental Research, Environmental RemediationSciences Program. MRCAT operations are supported by DOEand the MRCAT member institutions. The Advanced PhotonSource is supported by the DOE Office of Science, Office ofBasic Energy Sciences. We thank Tonia Mehlhorn andKenneth Lowe for analysis of groundwater samples.
Supporting Information AvailableDetailed Materials and Methods section, bacterial analysistable, and detailed figures and tables describing the EXAFS
models. This material is available free of charge via theInternet at http://pubs.acs.org.
Literature Cited(1) Lovley, D. R.; Phillips, E. J. P.; Gorby, Y. A.; Landa, E. R. Microbial
reduction of uranium. Nature 1991, 350 (6317), 413–416.(2) Anderson, R. T.; Vrionis, H. A.; Ortiz-Bernad, I.; Resch, C. T.;
Long, P. E.; Dayvault, R.; Karp, K.; Marutzky, S.; Metzler, D. R.;Peacock, A.; White, D. C.; Lowe, M.; Lovley, D. R. Stimulatingthe in situ activity of Geobacter species to remove uranium fromthe groundwater of a uranium-contaminated aquifer. Appl.Environ. Microbiol. 2003, 69 (10), 5884–5891.
(3) Istok, J. D.; Senko, J. M.; Krumholz, L. R.; Watson, D.; Bogle,M. A.; Peacock, A.; Chang, Y. J.; White, D. C. In situ bioreductionof technetium and uranium in a nitrate-contaminated aquifer.Environ. Sci. Technol. 2004, 38, 468–475.
(4) Wu, W. M.; Carley, J.; Fienen, M.; Mehlhorn, T.; Lowe, K.; Nyman,J.; Luo, J.; Gentile, M. E.; Rajan, R.; Wagner, D.; Hickey, R. F.;Gu, B. H.; Watson, D.; Cirpka, O. A.; Kitanidis, P. K.; Jardine,P. M.; Criddle, C. S. Pilot-scale in situ bioremediation of uraniumin a highly contaminated aquifer. 1. Conditioning of a treatmentzone. Environ. Sci. Technol. 2006, 40 (12), 3978–3985.
(5) Wu, W. M.; Carley, J.; Gentry, T.; Ginder-Vogel, M. A.; Fienen,M.; Mehlhorn, T.; Yan, H.; Caroll, S.; Pace, M. N.; Nyman, J.;Luo, J.; Gentile, M. E.; Fields, M. W.; Hickey, R. F.; Gu, B. H.;Watson, D.; Cirpka, O. A.; Zhou, J. Z.; Fendorf, S.; Kitanidis,P. K.; Jardine, P. M.; Criddle, C. S. Pilot-scale in situ bioreme-dation of uranium in a highly contaminated aquifer. 2. Reductionof U(VI) and geochemical control of U(VI) bioavailability.Environ. Sci. Technol. 2006, 40 (12), 3986–3995.
(6) Wu, W. M.; Carley, J.; Luo, J.; Ginder-Vogel, M. A.; Cardenas, E.;Leigh, M. B.; Hwang, C. C.; Kelly, S. D.; Ruan, C. M.; Wu, L. Y.;Van Nostrand, J.; Gentry, T.; Lowe, K.; Mehlhorn, T.; Carroll, S.;Luo, W. S.; Fields, M. W.; Gu, B. H.; Watson, D.; Kemner, K. M.;Marsh, T.; Tiedje, J.; Zhou, J. Z.; Fendorf, S.; Kitanidis, P. K.;Jardine, P. M.; Criddle, C. S. In situ bioreduction of uranium(VI)to submicromolar levels and reoxidation by dissolved oxygen.Environ. Sci. Technol. 2007, 41 (16), 5716–5723.
(7) Schink, B. Energetics of syntrophic cooperation in methanogenicdegradation. Microbiol. Mol. Biol. Rev. 1997, 61, 262–280.
(8) Wu, W.; Hickey, R. F.; Zeikus, J. G. Characterization of metabolicperformance of methanogenic granules treating brewery waste-water: role of sulfate-reducing bacteria. Appl. Environ. Microbiol.1991, 57, 3438–3445.
(9) Langmuir, D. Uranium solution-mineral equilibria at lowtemperatures with applications to sedimentary ore deposits.Geochim. Cosmochim. Acta 1978, 42, 547–569.
(10) Lovley, D. R.; Roden, E. E.; Phillips, E. J. P.; Woodward, J. C.Enzymatic iron and uranium reduction by sulfate-reducingbacteria. Mar. Geol. 1993, 113 (1-2), 41–53.
(11) Suzuki, Y.; Kelly, S. D.; Kemner, K. M.; Banfield, J. F. RadionuclideContamination - Nanometre-size Products of Uranium Biore-duction. Nature 2002, 419 (6903), 134–134.
(12) O’Loughlin, E. J.; Kelly, S. D.; Cook, R. E.; Csencsits, R.; Kemner,K. M. Reduction of uranium(VI) by mixed iron(II)/iron(III)hydroxide (green rust): Formation of UO2 nanoparticles. Environ.Sci. Technol. 2003, 37 (4), 721–727.
(13) Boyanov, M. I.; O’Loughlin, E. J.; Roden, E. E.; Fein, J. B.; Kemner,K. M. Adsorption of Fe(II) and U(VI) to carboxyl-functionalizedmicrospheres: The influence of speciation on uranyl reductionstudied by titration and XAFS. Geochim. Cosmochim. Acta 2007,71 (8), 1898–1912.
(14) O’Loughlin, E. J.; Kelly, S. D.; Kemner, K. M. An XAFS investigationof the interactions of uranium(VI) with secondary mineralizationproducts from the bioreduction of iron(III) oxides. Environ. Sci.Technol. (submitted for publication).
(15) Kelly, S. D.; Kemner, K. M.; Carley, J.; Criddle, C.; Jardine, P. M.;Marsh, T. L.; Phillips, D.; Watson, D.; Wu, W. M. Speciation ofuranium in sediments before and after in situ biostimulation.Environ. Sci. Technol. 2008, 42 (5), 1558–1564.
(16) Burgos, W. D.; Senko, J. M.; Dempsey, B. A.; Roden, E. E.; Stone,J. J.; Kenmer, K. M.; Kelly, S. D. Soil humic acid decreasesbiological uranium(VI) reduction by Shewanella putrefaciensCN32. Environ. Eng. Sci. 2007, 24 (6), 755–761.
(17) Brooks, S. C.; Fredrickson, J. K.; Carroll, S. L.; Kennedy, D. W.;Zachara, J. M.; Plymale, A. E.; Kelly, S. D.; Kemner, K. M.; Fendorf,S. Inhibition of bacterial U(VI) reduction by calcium. Environ.Sci. Technol. 2003, 37 (9), 1850–1858.
(18) Tokunaga, T. K.; Wan, J. M.; Kim, Y. M.; Sutton, S. R.; Newville,M.; Lanzirotti, A.; Rao, W. Real-time X-ray absorption spec-troscopy of uranium, iron, and manganese in contaminated
VOL. 44, NO. 1, 2010 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 241

sediments during bioreduction. Environ. Sci. Technol. 2008, 42(8), 2839–2844.
(19) Tokunaga, T. K.; Wan, J. M.; Pena, J.; Sutton, S. R.; Newville,M. G. Hexavalent uranium diffusion into soils from concentratedacidic and alkaline solutions. Environ. Sci. Technol. 2004, 38(11), 3056–3062.
(20) Komlos, J.; Mishra, B.; Lanzirotti, A.; Myneni, S. C. B.; Jaffe, P. R.Real-time speciation of uranium during active bioremediationand U(IV) reoxidation. J. Environ. Eng.-ASCE 2008, 134 (2), 78–86.
(21) Kelly, S. D.; Hesterberg, D.; Ravel, B. Analysis of soils and mineralsusing X-ray absorption spectroscopy. In Methods of soil analysis,Part 5: Mineralogical methods; Ulery, A. L., Drees, L. R., Eds.;Soil Science Society of America: Madison, WI, 2008; pp 367-463.
(22) Gu, B. H.; Wu, W. M.; Ginder-Vogel, M. A.; Yan, H.; Fields, M. W.;Zhou, J.; Fendorf, S.; Criddle, C. S.; Jardine, P. M. Bioreductionof uranium in a contaminated soil column. Environ. Sci. Technol.2005, 39 (13), 4841–4847.
(23) Akob, D. M.; Mills, H. J.; Kostka, J. E. Metabolically activemicrobial communities in uranium-contaminated subsurfacesediments. FEMS Microbiol. Ecol. 2007, 59 (1), 95–107.
(24) Nevin, K. P.; Finneran, K. T.; Lovely, D. R. Microorganismsassociated with uranium bioremediation in a high-salinitysubsurface sediment. Appl. Environ. Microbiol. 2003, 69 (6),3672–3675.
(25) Suzuki, Y.; Kelly, S. D.; Kemner, K. A.; Banfield, J. F. Microbialpopulations stimulated for hexavalent uranium reduction inuranium mine sediment. Appl. Environ. Microbiol. 2003, 69 (3),1337–1346.
(26) North, N. N.; Dollhopf, S. L.; Petrie, L.; Istok, J. D.; Balkwill,D. L.; Kostka, J. E. Change in bacterial community structureduring in situ biostimulation of subsurface sediment cocon-taminated with uranium and nitrate. Appl. Environ. Microbiol.2004, 70 (8), 4911–4920.
(27) Brodie, E. L.; DeSantis, T. Z.; Joyner, D. C.; Baek, S. M.; Larsen,J. T.; Andersen, G. L.; Hazen, T. C.; Richardson, P. M.; Herman,D. J.; Tokunaga, T. K.; Wan, J. M. M.; Firestone, M. K. Applicationof a high-density oligonucleotide microarray approach to studybacterial population dynamics during uranium reduction andreoxidation. Appl. Environ. Microbiol. 2006, 72 (9), 6288–6298.
(28) Michalsen, M. M.; Peacock, A. D.; Spain, A. M.; Smithgal, A. N.;White, D. C.; Sanchez-Rosario, Y.; Krumholz, L. R.; Istok, J. D.Changes in microbial community composition and geochem-istry during uranium and technetium bioimmobilization. Appl.Environ. Microbiol. 2007, 73 (18), 5885–5896.
(29) Moreels, D.; Crosson, G.; Garafola, C.; Monteleone, D.; Taghavi,S.; Fitts, J. P.; van der Lelie, D. Microbial community dynamicsin uranium contaminated subsurface sediments under bio-stimulated conditions with high nitrate and nickel pressure.Environ. Sci. Pol. Res. 2008, 15 (6), 481–491.
(30) Hwang, C. C.; Wu, W. M.; Gentry, T. J.; Carley, J.; Corbin, G. A.;Carroll, S. L.; Watson, D. B.; Jardine, P. M.; Zhou, J. Z.; Criddle,C. S.; Fields, M. W. Bacterial community succession during insitu uranium bioremediation: spatial similarities along con-trolled flow paths. ISME J. 2009, 3 (1), 47–64.
(31) Fields, M. W.; Yan, T. F.; Rhee, S. K.; Carroll, S. L.; Jardine, P. M.;Watson, D. B.; Criddle, C. S.; Zhou, J. Z. Impacts on microbialcommunities and cultivable isolates from groundwater con-taminated with high levels of nitric acid-uranium waste. FEMSMicrobiol. Ecol. 2005, 53 (3), 417–428.
(32) Mohanty, S. R.; Kollah, B.; Hedrick, D. B.; Peacock, A. D.;Kukkadapu, R. K.; Roden, E. E. Biogeochemical processes inethanol stimulated uranium-contaminated suhsurface sedi-ments. Environ. Sci. Technol. 2008, 42 (12), 4384–4390.
(33) Sani, R. K.; Peyton, B. M.; Amonette, J. E.; Geesey, G. G. Reductionof uranium(VI) under sulfate-reducing conditions in the pres-ence of Fe(III)-(hydr)oxides. Geochim. Cosmochim. Acta 2004,68 (12), 2639–2648.
(34) Sani, R. K.; Peyton, B. M.; Dohnalkova, A.; Amonette, J. E.Reoxidation of reduced uranium with iron(III) (hydr)oxidesunder sulfate-reducing conditions. Environ. Sci. Technol. 2005,39 (7), 2059–2066.
(35) Suzuki, Y.; Kelly, S. D.; Kemner, K. M.; Banfield, J. F. EnzymaticU(VI) reduction by Desulfosporosinus species. Radiochim. Acta2004, 92 (1), 11–16.
(36) Payne, R. B.; Casalot, L.; Rivere, T.; Terry, J. H.; Larsen, L.; Giles,B. J.; Wall, J. D. Interaction between uranium and the cytochromec(3) of Desulfovibrio desulfuricans strain G20. Arch. Microbiol.2004, 181 (6), 398–406.
(37) Beyenal, H.; Sani, R. K.; Peyton, B. M.; Dohnalkova, A. C.; Amonette,J. E.; Lewandowski, Z. Uranium immobilization by sulfate-reducingbiofilms. Environ. Sci. Technol. 2004, 38 (7), 2067–2074.
(38) Bostick, B. C.; Fendorf, S.; Barnett, M. O.; Jardine, P. M.; Brooks,S. C. Uranyl surface complexes formed on subsurface mediafrom DOE facilities. Soil. Sci. Soc. Am. J. 2002, 66, 99–108.
(39) Wan, J. M.; Tokunaga, T. K.; Brodie, E.; Wang, Z. M.; Zheng,Z. P.; Herman, D.; Hazen, T. C.; Firestone, M. K.; Sutton, S. R.Reoxidation of bioreduced uranium under reducing conditions.Environ. Sci. Technol. 2005, 39 (16), 6162–6169.
ES902191S
242 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 44, NO. 1, 2010





























