Embed Size (px)
Citation preview

J. N. Am. Benthol. Soc., 2007, 26(3):450–459� 2007 by The North American Benthological Society
Functional bioturbator diversity enhances benthic–pelagic processesand properties in experimental microcosms
Adriano Caliman1, Joao J. F. Leal2, Francisco A. Esteves3,Luciana S. Carneiro4, Reinaldo L. Bozelli5, AND Vinicius F. Farjalla6
Laboratorio de Limnologia, CCS, Universidade Federal do Rio de Janeiro, Rio de Janeiro-RJ, Brazil 68020
Abstract. Widespread declines in biodiversity at both global and local scales have motivated considerableresearch directed toward understanding how changes in biological diversity may affect the stability andfunction of the ecosystems on which we rely. However, the research effort devoted to addressing this question inbenthic systems has been minimal. In laboratory microcosms, we manipulated the number and composition of3 functionally distinct benthic invertebrate freshwater species that are bioturbators of sediment over 3 biomasslevels. Our objective was to test the effects of bioturbator diversity on rates and reliability of total dissolved P(TDP) flux between benthic and pelagic habitats. Both composition and species richness affected TDP flux. TDPflux was highest in the most species-rich community because of functional complementarity rather thanselection effects. Furthermore, species richness enhanced TDP flux reliability by increasing the predictability ofthe biomass–TDP flux relationship by 30%, on average, for each species added. We attributed these nonadditiveeffects of invertebrate diversity to a combination of functionally mediated biogeochemical interactions anddensity-mediated interaction strength. Thus, our results suggest that bioturbator diversity can be important tonutrient cycling in aquatic ecosystems by strengthening benthic–pelagic coupling.
Key words: niche complementarity, species richness, species composition, biodiversity, benthic–pelagiccoupling, reliability, bioturbation, phosphorus cycle.
One of the major challenges in contemporaryecological research is to understand the role ofbiological diversity in ecosystem functioning andservices (Loreau et al. 2002, Hooper et al. 2005, andreferences therein). The fundamental motivation of thisscientific inquiry has been to answer how ecosystemprocesses will be affected by the increasing rates oflocal and global loss of biodiversity (Chapin et al. 2000).
Two underlying mechanisms have been suggestedto explain how increasing biodiversity generally altersecosystem-level processes to an extent that cannot bepredicted from the sum of the roles of individualspecies (i.e., nonadditive effects). First, the magnitudeof individual species’ effects on ecosystem functioningis variable. Thus, species-rich communities may havemore influence on ecosystem functioning than species-poor communities, on average, because they are more
likely to contain species that have large effects onecosystem functioning (i.e., selection effect; Huston1997, Tilman et al. 1997, Loreau 1998). Second,different species use and may facilitate the acquisitionof slightly different resources. Therefore, species-richcommunities may be more efficient than species-poorcommunities at using the overall resource pool,thereby enhancing ecosystem processes (i.e., nichecomplementarity; Tilman et al. 1997, Loreau 1998,Cardinale et al. 2002).
Aquatic ecosystems are more threatened than terres-trial systems by human activities (Ricciardi andRasmussen 1999), but most of our knowledge of theeffects of biodiversity on ecosystem functioning comesfrom studies carried out on terrestrial vegetation andlaboratory microbial systems (e.g., Tilman et al. 1997,Bell et al. 2005, Hooper et al. 2005). Only recently haveexperimental studies in coastal marine sediments andstream ecosystems explored the importance of interac-tions among benthic invertebrates species for ecosys-tem processes (i.e., nutrient fluxes, energy transfer, oractivity of sediment bacteria; Emmerson and Raffaelli2000, Emmerson et al. 2001, Bolam et al. 2002,Cardinale et al. 2002, Cardinale and Palmer 2002,
1 E-mail addresses: [email protected] [email protected] [email protected] [email protected] [email protected] To whom correspondence should be addressed. E-mail:
450

Waldbusser et al. 2004, Jonsson and Malmqvist 2005,Mermillod-Blondin et al. 2005, Ieno et al. 2006). Thesestudies showed multiple effects of benthic diversity onaquatic ecosystem functioning, but the strength anddirection of these effects were highly variable inresponse to number, composition, biomass of interact-ing species, ecosystem process analyzed, and theirunderlying mechanisms. The multitude of findings hasgenerated diverse conclusions about the relativeimportance of benthic invertebrate diversity in aquaticecosystem processes, highlighting the complexity of therelationships and the need to take into account otheraquatic environments, species functional traits, andecological processes (see Covich et al. 2004 for review).
To our knowledge, no effort has been devoted tounderstanding the role of benthic invertebrate diversityin processes and properties that regulate P cycling inshallow lakes, although P is a main limiting nutrient inprimary and microbial productivity in freshwaterenvironments and a major contributor to the processof eutrophication (e.g., Schindler 1977, Wetzel 2001).Hence, understanding the factors that regulate Pcycling in these systems has important ecological andeconomic ramifications. Furthermore, shallow lakeecosystems show higher interhabitat connectivity thandeep ones (Schindler and Scheuerell 2002). Thus, effectsof bioturbator diversity might assume greater impor-tance in subsidizing benthic–pelagic coupling andwhole-ecosystem functioning in shallow lakes than indeep lakes (Palmer et al. 2000, Covich et al. 2004).
Here, the effects of interspecific interactions amongfunctionally different bioturbating species that belongto 3 ubiquitous groups of freshwater benthic inverte-brates were analyzed through a short-term experimentin laboratory microcosms. The rates and reliability oftotal dissolved P (TDP) flux between the benthic andpelagic habitats were measured as ecosystem func-tions. We used Naeem’s (1998) definition of reliabilityto estimate ecosystem stability because it refers to theprobability that a system will provide a consistentlevel of performance over a given unit of change in itsbiotic or abiotic parameters (e.g., community biomass).We hypothesized that aspects of biodiversity, such ascommunity composition or species richness, wouldalter, in a nonadditive way, the effects of sedimentbioturbators (measured as TDP flux) on rates andreliability of benthic–pelagic coupling.
Methods
Sampling and characteristics of sediment and organisms
Sediment and benthic invertebrates were collectedclose to the littoral region of Imboassica lagoon (lat228500S, long 448420W), a tropical, shallow, coastal
freshwater ecosystem located in Rio de Janeiro State,Brazil. Untreated domestic sewage input adds largeloads of N and P to the lagoon and causes eutrophi-cation (Albertoni et al. 2001). Sediment at the samplingsite is predominantly silt and clay with a mean total Pconcentration of 0.25 mg/g (Callisto et al. 1998).
Samples from the upper layer of sediment (0–5 cm)were taken with a core sampler, sieved through a 1-mm mesh to remove macrofauna and larger particles(shells), frozen for 2 wk, and then thawed. Thisprocedure removed all metazoans as well as theirresistant forms (Emmerson et al. 2001). Next, thesediment was homogenized and allowed to settle for10 d in a 30-L aquarium with a 10-cm-deep layer ofprefiltered (25-lm mesh) lagoon water to reduce thenatural heterogeneity of the sediment and to permitrecovery of its biogeochemical depth gradient (e.g.,Emmerson et al. 2001, Stief et al. 2004, Mermillod-Blondin et al. 2005). The aquarium was aeratedconstantly and was kept in the dark to preventphotosynthesis.
Individuals of 3 species—larval Chironomus sp.(Meigen) (Diptera:Chironomidae), Heteromastus similis(Southern) (Polychaeta:Captellidae), and Heleobia aus-tralis (D’Orbigny) (Gastropoda:Hydrobiidae)—werecollected from the field 1 d before the experimentbegan and were conditioned in species-specific aquariafilled with lagoon sediment and water to allow them toacclimatize to laboratory conditions. Together thesespecies constitute .80% of the total benthic inverte-brate biomass in Imboassica lagoon (Callisto et al.1998, Figueiredo-Barros 2004), but they differ clearly inspatial distribution and foraging behavior within thesediment.
Chironomus sp. is a filter feeder and a tube dweller,and its undulations promote a continuous flux ofwater through permanent U-shaped burrows. Thisflux oxygenates deep layers of sediment and pumpslarge amounts of dissolved and particulate materialfrom sediment to the overlying water (Gallepp 1979,Graneli 1979). During periods of hypoxia, Chironomuslarvae may present striking facultative behaviorseither by elongating their U-shaped sediment burrowsinto chimneys that tower above the sediment surfaceor by feeding on surface deposits and ingesting bothsediment particles and microbial organisms (Mermil-lod-Blondin et al. 2002, Stief et al. 2005). Heleobiaaustralis ploughs the sediment surface and feedsmainly on organic detritus (Goncalves-Junior et al.1998). Its behavior as a surface-deposit feeder has littleeffect on vertical sediment geochemistry but cangreatly affect interfacial geochemical kinetics mediatedby aerobic microbes because it reworks the top layer ofsediment (Figueiredo-Barros 2004). Heteromastus similis
2007] 451BENTHIC BIODIVERSITY AND THE P CYCLE

is a head-down subsurface-deposit feeder that buildsextensive semipermanent galleries in the sediment(Rao 1980). It modifies the distribution of sedimentorganic matter and intensifies benthic–pelagic cou-pling by sediment advection and upward egestion offecal pellets (Rao 1980).
Experimental design
Benthic invertebrate species richness (1–3 species inall possible combinations) was manipulated across 3invertebrate biomass (density) levels (150, 300, and 450mg wet mass), in a full factorial-design replacementseries (Jolliffe 2000, Emmerson et al. 2001) in micro-cosms containing a sediment–water interface (seeExperimental setup and sampling procedures below).Thus, the contribution of a species to total invertebratebiomass was decreased to ½ in 2-species mixtures or to1 /
3 in 3-species mixtures when compared with its ownmonocultures. Microcosms without macrofauna wereestablished as controls used to estimate nutrient fluxacross the sediment–water interface in the absence ofbenthic invertebrates. All macroinvertebrate treat-ments and controls were replicated 43 for a total of88 microcosms. This experimental design was chosenfor 3 reasons: 1) to compare the results of mixtureswith results from respective constituent monoculturesand allow us to separate potential confounding effectsbetween species number and composition (Huston1997); 2) to cover the natural range of species–biomassdistributions in Imboassica lagoon; and 3) to provideinformation about the effects of biodiversity onbenthic–pelagic coupling by manipulating speciesrichness and composition over a realistic range ofdensity-mediated interaction strengths.
Experimental setup and sampling procedures
A few hours before starting the experiment, micro-cosms were established by transferring the stabilizedsediment into Plexiglas tubes (20 cm long and 4 cminternal diameter) to a depth of 10 cm. The overlyingwater (9-cm depth) of each microcosm was drainedand replaced by fresh 0.7-lm-filtered (GF/F Whatman,Pleasanton, California) lagoon water to reduce TDPresulting from dead organic material within thesediment and most planktonic organisms. Individualsof a given species and size were weighed to the nearest0.1 mg (wet mass after blotting excess water) andimmediately distributed into respective microcosms.Therefore, biomass was proportional to density.Throughout the experiment, each microcosm wassupplied with its own aerator and kept in the darkto prevent depletion of dissolved O2, stratification ofthe water column, and photosynthesis. Water samples
(20 mL) were taken from each microcosm at thebeginning and the end (24-h incubation) of theexperiment, filtered through 0.7-lm-pore filters (GF/F Whatman), and immediately frozen until TDP wasdetermined.
TDP concentrations were measured by autoclavingwater samples with potassium persulfate oxidant andanalyzing the digested samples photometrically usingthe ascorbic acid method according to Golterman et al.(1978). TDP flux was calculated as the differencebetween initial and final TDP concentrations for eachmicrocosm. TDP can be released by the organismsthemselves as a metabolic by-product, but an earlystudy carried out with the same species from the sameecosystem showed minimal contributions to overallTDP fluxes through this mechanism (Figueiredo-Barros 2004). Therefore, changes in TDP concentra-tions measured in our experiment were assumed torepresent the effects of bioturbation of the sediment.
Data analysis
The effects of biodiversity on ecosystem processescan be assessed in a variety of ways. The mostcommon test is to compare the response (in this case,TDP flux) observed with a mixture of species with thatexpected from the sum of the responses of eachcomponent species in monoculture (Loreau 1998,Wardle 1999). If the response to a mixture of speciesoutperforms the expected response, then species areinteracting synergistically, a situation known as non-transgressive overyielding. However, this test does notallow differentiation among the possible mechanisms(selection effects or niche complementarity) by whichbiodiversity is operating because the contributions ofthe component species to the observed multispeciesresponse cannot be calculated for some ecosystemprocesses, such as nutrient fluxes (Emmerson et al.2001). A more conservative and rigorous test is tocompare the response observed with a mixture ofspecies with the response observed with the best-performing component species in monoculture (Lor-eau 1998, Wardle 1999). A higher response with amixture of species indicates transgressive overyieldingand can be interpreted as evidence of niche comple-mentarity.
These 2 approaches were used to identify nonaddi-tive interactions and to confirm the existence ofcomplementarity effects. The overall effects of inver-tebrate diversity on TDP flux were analyzed byintegrating responses across the whole range ofbiomass with an additive null model according toEmmerson and Raffaelli (2000). This model comparesthe relationship between invertebrate biomass and
452 [Volume 26A. CALIMAN ET AL.

ecosystem response (TDP flux) for species alone and inmixture. Observations derived from single-speciesregressions between biomass and TDP flux were usedto generate an expected mixture regression againstwhich we could compare observed responses frommultispecies mixtures. Further details about this nullmodel can be found in Emmerson and Raffaelli (2000).We used analysis of covariance (ANCOVA) (withspecies richness as a main factor and biomass as acovariate) to compare the observed response from amultispecies mixture against the expected responsefrom the multispecies mixture and the observedresponse to the best-performing component monocul-ture.
Linear regression of TDP flux values as a function ofinvertebrate species richness was used to detect theoverall effect of invertebrate species richness on TDPflux. A separate nested analysis of variance (ANOVA)with species composition nested within species rich-ness, followed by Tukey’s tests of pairwise compari-sons, was used to assess the overall effect of speciescomposition. Regression analysis combined with arestricted nested ANOVA was used instead of a uniquecomplete ANOVA test because it allowed betterexploration of quantitative (species richness) andqualitative (species composition) effects of invertebratebiodiversity on TDP flux (see Cottingham et al. 2005for further details). The effects of invertebrate speciesrichness on TDP flux reliability were evaluated byregressing the coefficients of determination (R2) ob-tained from each biomass–TDP flux linear regressionas a function of benthic invertebrate species richness. Apositive relationship between R2 values and inverte-brate species richness should indicate that species-richbenthic communities have a greater probability thanspecies-poor communities of providing a consistentlevel of TDP flux over a given change in benthiccommunity biomass. This analysis was similar to oneused by Naeem and Li (1997) and Naeem (1998) toassess the effects of microbial biodiversity on temporalreliability of other ecosystem processes. All analyseswere done using STATISTICA (version 6.0; StatSoft,
Tulsa, Oklahoma). Results were considered significantwhen p , 0.05.
Results
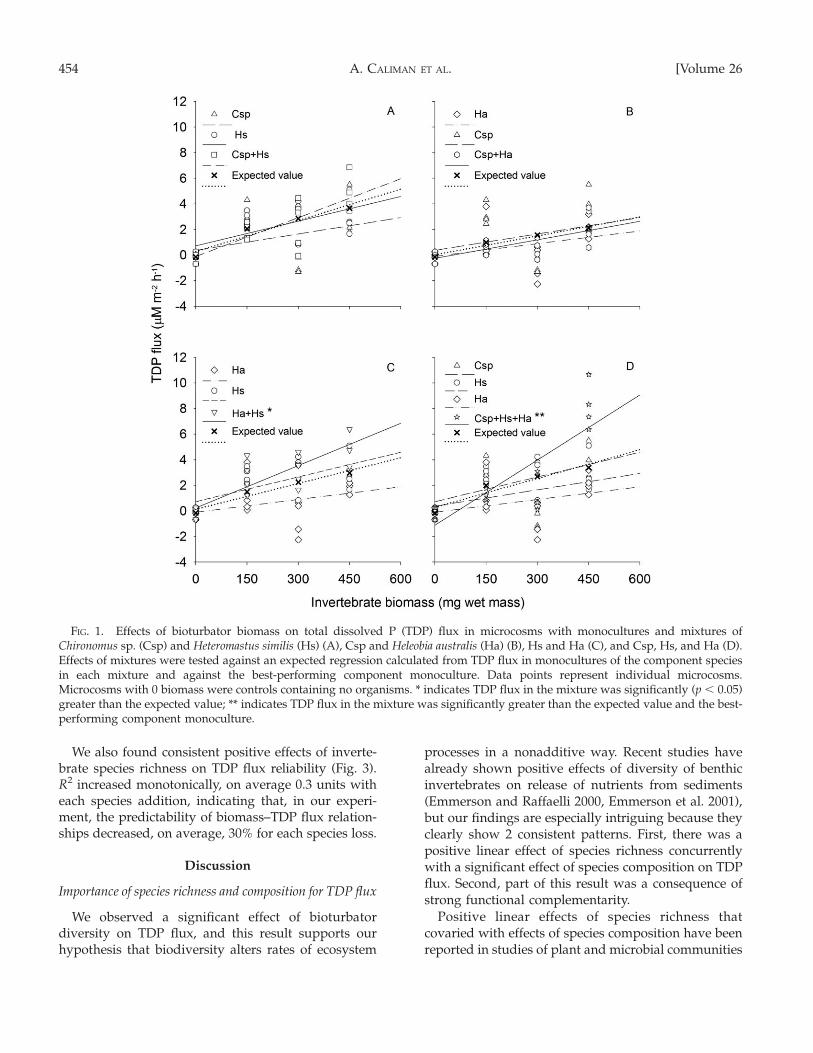
Invertebrate biomass had a significant positivelinear effect on TDP flux for 5 of the 7 speciescombinations (Table 1, Fig. 1A–D). Invertebrate bio-mass had slightly positive but nonsignificant effects onTDP flux in microcosms with Chironomus sp. and H.australis monocultures (Table 1); these results canprobably be attributed to the low TDP fluxes atintermediate (300 mg) biomass for both species (Fig.1B). TDP flux was higher in microcosms with Hetero-mastus similis monocultures than in microcosms withChironomus sp. or H. australis monocultures, highlight-ing the functional differences between the species.
Comparisons between observed TDP fluxes inmicrocosms containing multispecies mixtures andrespective expected TDP fluxes showed significanteffects of invertebrate diversity on TDP flux (Fig. 1A–D). Among 2-species assemblages, a significant posi-tive nonadditive interaction occurred only in themixture composed of H. similis and H. australis. Thismixture yielded TDP fluxes that were higher thanexpected fluxes but not higher than fluxes in the best-performing component monoculture (expected flux:F1,28 ¼ 5.31, p ¼ 0.02; best-performing monoculture:F1,28 ¼ 2.83, p ¼ 0.1; Fig. 1C), indicating nontrans-gressive overyielding. However, in the 3-speciesassemblage, TDP flux was higher than both theexpected flux and the flux in the best-performingcomponent monoculture (expected flux: F1,28¼ 9.9, p¼0.003; best-performing monoculture: F1,28 ¼ 8.73, p ¼0.006; Fig. 1D), indicating functional complementarity.
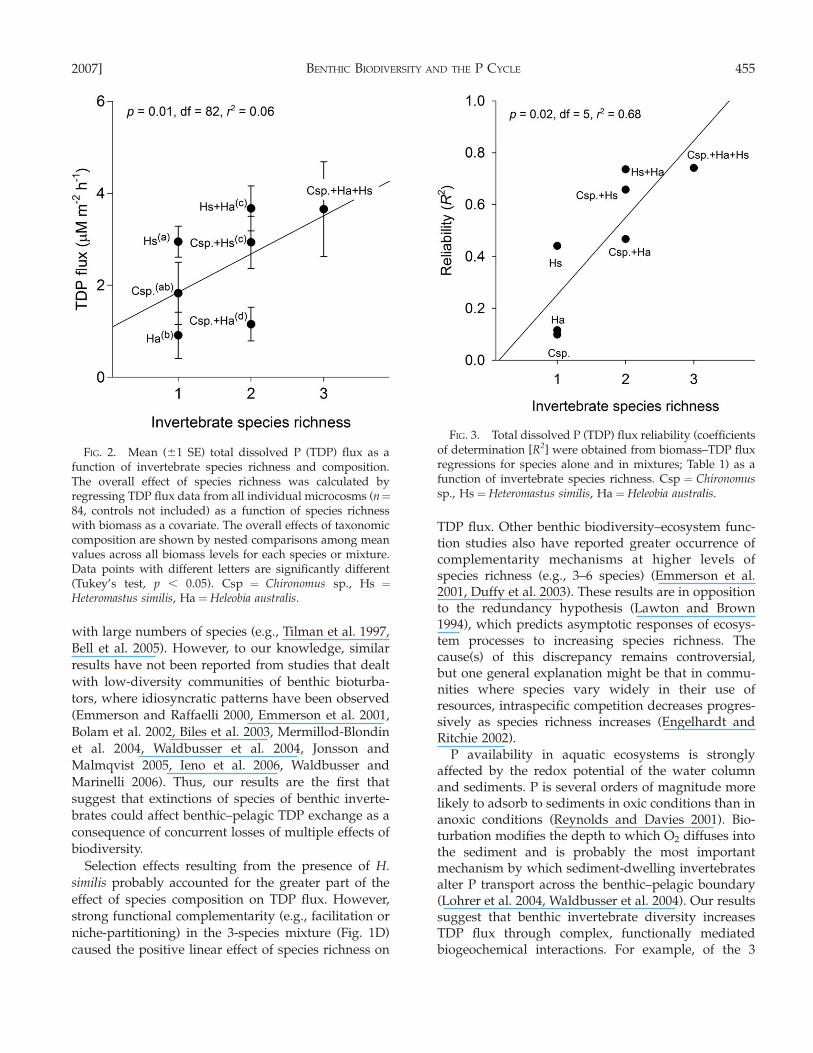
Both invertebrate species richness and compositionaffected TDP flux (Fig. 2). TDP flux was a positive linearfunction of invertebrate species richness. On average,each species addition corresponded to an increase of0.83 lM m�2 h�1 in TDP flux rates from the sediment tothe water column. However, a considerable part of thiseffect could be attributed to the presence of H. similis(nested ANOVA, F¼ 8.45, df¼ 4, p , 0.0001; Fig. 2).
TABLE 1. Statistical parameters from linear regressions of total dissolved P (TDP) flux as a function of invertebrate biomass forbioturbator species in monocultures and in 2- and 3-species mixtures. * indicates significant linear relationship.
Treatments R2 Slope F p
Chironomus sp. 0.11 0.004 1.79 0.2Heteromastus similis 0.42 0.006 10.22 0.006*Heleobia australis 0.13 0.003 2.12 0.16Chironomus sp. þ H. similis 0.64 0.010 25.80 0.0002*Chironomus sp. þ H. australis 0.45 0.004 11.84 0.004*H. similis þ H. australis 0.72 0.011 36.56 ,0.0001*Chironomus sp. þ H. similis þ H. australis 0.70 0.017 33.28 ,0.0001*
2007] 453BENTHIC BIODIVERSITY AND THE P CYCLE
We also found consistent positive effects of inverte-brate species richness on TDP flux reliability (Fig. 3).R2 increased monotonically, on average 0.3 units witheach species addition, indicating that, in our experi-ment, the predictability of biomass–TDP flux relation-ships decreased, on average, 30% for each species loss.
Discussion
Importance of species richness and composition for TDP flux
We observed a significant effect of bioturbatordiversity on TDP flux, and this result supports ourhypothesis that biodiversity alters rates of ecosystem
processes in a nonadditive way. Recent studies havealready shown positive effects of diversity of benthicinvertebrates on release of nutrients from sediments(Emmerson and Raffaelli 2000, Emmerson et al. 2001),but our findings are especially intriguing because theyclearly show 2 consistent patterns. First, there was apositive linear effect of species richness concurrentlywith a significant effect of species composition on TDPflux. Second, part of this result was a consequence ofstrong functional complementarity.
Positive linear effects of species richness thatcovaried with effects of species composition have beenreported in studies of plant and microbial communities
FIG. 1. Effects of bioturbator biomass on total dissolved P (TDP) flux in microcosms with monocultures and mixtures ofChironomus sp. (Csp) and Heteromastus similis (Hs) (A), Csp and Heleobia australis (Ha) (B), Hs and Ha (C), and Csp, Hs, and Ha (D).Effects of mixtures were tested against an expected regression calculated from TDP flux in monocultures of the component speciesin each mixture and against the best-performing component monoculture. Data points represent individual microcosms.Microcosms with 0 biomass were controls containing no organisms. * indicates TDP flux in the mixture was significantly (p , 0.05)greater than the expected value; ** indicates TDP flux in the mixture was significantly greater than the expected value and the best-performing component monoculture.
454 [Volume 26A. CALIMAN ET AL.
with large numbers of species (e.g., Tilman et al. 1997,Bell et al. 2005). However, to our knowledge, similar
results have not been reported from studies that dealt
with low-diversity communities of benthic bioturba-tors, where idiosyncratic patterns have been observed
(Emmerson and Raffaelli 2000, Emmerson et al. 2001,
Bolam et al. 2002, Biles et al. 2003, Mermillod-Blondinet al. 2004, Waldbusser et al. 2004, Jonsson and
Malmqvist 2005, Ieno et al. 2006, Waldbusser and
Marinelli 2006). Thus, our results are the first thatsuggest that extinctions of species of benthic inverte-
brates could affect benthic–pelagic TDP exchange as a
consequence of concurrent losses of multiple effects ofbiodiversity.
Selection effects resulting from the presence of H.similis probably accounted for the greater part of the
effect of species composition on TDP flux. However,strong functional complementarity (e.g., facilitation or
niche-partitioning) in the 3-species mixture (Fig. 1D)
caused the positive linear effect of species richness on
TDP flux. Other benthic biodiversity–ecosystem func-tion studies also have reported greater occurrence ofcomplementarity mechanisms at higher levels ofspecies richness (e.g., 3–6 species) (Emmerson et al.2001, Duffy et al. 2003). These results are in oppositionto the redundancy hypothesis (Lawton and Brown1994), which predicts asymptotic responses of ecosys-tem processes to increasing species richness. Thecause(s) of this discrepancy remains controversial,but one general explanation might be that in commu-nities where species vary widely in their use ofresources, intraspecific competition decreases progres-sively as species richness increases (Engelhardt andRitchie 2002).
P availability in aquatic ecosystems is stronglyaffected by the redox potential of the water columnand sediments. P is several orders of magnitude morelikely to adsorb to sediments in oxic conditions than inanoxic conditions (Reynolds and Davies 2001). Bio-turbation modifies the depth to which O2 diffuses intothe sediment and is probably the most importantmechanism by which sediment-dwelling invertebratesalter P transport across the benthic–pelagic boundary(Lohrer et al. 2004, Waldbusser et al. 2004). Our resultssuggest that benthic invertebrate diversity increasesTDP flux through complex, functionally mediatedbiogeochemical interactions. For example, of the 3
FIG. 2. Mean (61 SE) total dissolved P (TDP) flux as afunction of invertebrate species richness and composition.The overall effect of species richness was calculated byregressing TDP flux data from all individual microcosms (n¼84, controls not included) as a function of species richnesswith biomass as a covariate. The overall effects of taxonomiccomposition are shown by nested comparisons among meanvalues across all biomass levels for each species or mixture.Data points with different letters are significantly different(Tukey’s test, p , 0.05). Csp ¼ Chironomus sp., Hs ¼Heteromastus similis, Ha ¼Heleobia australis.
FIG. 3. Total dissolved P (TDP) flux reliability (coefficientsof determination [R2] were obtained from biomass–TDP fluxregressions for species alone and in mixtures; Table 1) as afunction of invertebrate species richness. Csp ¼ Chironomussp., Hs ¼Heteromastus similis, Ha ¼ Heleobia australis.
2007] 455BENTHIC BIODIVERSITY AND THE P CYCLE

species tested, H. similis promoted the highest TDPflux, probably because its head-down deposit-feedingbehavior promotes upward transport of O2-free fecalpellets through sediment egestion. In species mixturescontaining H. similis and H. australis, H. similis fecalpellets may have been crushed as H. australis ploughedthe sediment surface, increasing soluble P in theoverlying water. Neira and Hopner (1994) observedthat meiofauna activity increased the rate of destruc-tion of Heteromastus filiformis fecal pellets by ;85%.Levinton (1995) suggested that this kind of interactionalso can occur between subsurface- and surface-deposit feeders.
Chironomus sp. did not interact synergistically withH. similis and H. australis in the 2-species mixtures, butit did interact synergistically in the 3-species mixture.We hypothesize that this result might have been aconsequence of a complex combination of functionalcomplementarity and density-mediated effects. Larvaeof Chironomus sp. build permanent, well-irrigated U-shaped burrows through which oxygenated water isconducted to deeper sediment layers and upwardtransport of dissolved and particulate materials ispromoted (Gallepp 1979, Graneli 1979, Mermillod-Blondin et al. 2002). Thus, Chironomus sp. can promoteeither positive (by pumping porewater-rich soluble Pto overlying water) or negative (by increasing Padsorption within the sediment) TDP flux betweenbenthic and pelagic habitats. Mermillod-Blondin et al.(2002) investigated the effects of Chironomus plumosuson sediment nutrient flux and observed that theoutcome of bioturbation by this species was non-linearly density dependent. They attributed this resultto functional responses of larvae to density-mediatedintraspecific interference competition (see also Mari-nelli and Williams 2003, Lohrer et al. 2004).
In our study, TDP flux was not linearly related to thebiomass of Chironomus sp. in monoculture (Table 1,Fig. 1B). Biomass-dependent changes in bioturbationbehavior might dictate the effect of bioturbation byChironomus sp. on TDP flux. Considering the argu-ments already presented, we suggest that the reduc-tion of Chironomus sp. biomass in the 3-speciesmixtures may have resulted in a positive balancebetween downward transport of O2 and upwardtransport of P across the sediment–water interface.This positive balance might, in turn, have interactedcomplementarily with H. similis and H. australis effectsto generate the high TDP flux from sediment tooverlying water observed in the microcosms with the3-species mixture (Fig. 1D). We suggest that, in 3-species mixtures, these interactions might have been aconsequence of complementary niche partitioning (viaupward pumping of TDP-rich pore water stimulated
by H. similis) or facilitation (via oxygenation of deeperlayers of the sediment, which increases the depth towhich H. australis can forage), alone or in combination.On the other hand, we propose that, in the 2-speciesmixtures, Chironomus sp. biomass was high enough tonullify potential synergistic interactions because ofstrong density-dependent effects (in mixtures ofChironomus sp. and H. australis; Fig. 1B) or strongadsorption of TDP to the sediment (in mixtures ofChironomus sp. and H. similis; Fig. 1C). However, weare unable to evaluate which mechanisms generatedthe observed results because we did not monitorspecies interactions in our experiment. Future workshould examine the specific mechanisms underlyingcomplementary interactions in benthic invertebratecommunities.
Importance of species richness for TDP flux reliability
The positive effect of invertebrate species richnesson TDP flux reliability observed in our experimentagreed with results of a similar study in which highmicrobial species diversity supported more reliableaggregated community biomass (Naeem and Li 1997).However, the 2 experiments differ remarkably in theirimplications. According to the insurance hypothesisproposed by Yachi and Loreau (1999), biodiversityinsures ecosystems against declines in function be-cause the presence of many species in an ecosystemincreases the likelihood that some species will main-tain ecosystem functions even if other species fail. Thishypothesis assumes the existence of some redundancyamong species that permits functional or numericalcompensation and buffers declines in ecosystemfunction. Naeem and Li’s (1997) experiment includedredundant species, but our experiment used 3 func-tionally different benthic invertebrate species. There-fore, our study yielded 2 important results: 1)invertebrate species richness promotes more predict-able ecosystem process even without functionalredundancy, and 2) the loss of any species reducesthe reliability of benthic–pelagic coupling whenaccompanied by alterations in benthic communitybiomass.
Our results are presented with 2 caveats. First,functional groups remain a controversial and evenarbitrary concept in ecology (Loreau 2004). We areunable to assert the degree of niche overlap among thespecies used in our experiment. Second, benthicinvertebrate species can change their behavior inresponse to fluctuating biotic (e.g., density; Mermil-lod-Blondin et al. 2002, Marinelli and Williams 2003)and abiotic (e.g., water flow, O2 concentration; Reise2002, Biles et al. 2003, Stief et al. 2005) conditions, and
456 [Volume 26A. CALIMAN ET AL.

these behavioral changes may alter their functionalrole in the ecosystem. Behavioral/functional plasticitymay have accounted for some unknown facultativespecies traits in our experiment, which may have led tosome unrecognized functional redundancy amongspecies in species-rich treatments.
Despite the mismatch between theory and ourresults and the caveats discussed above, we suggestthat the positive effect of bioturbator richness on TDPflux reliability is likely to have occurred even in theabsence of significant functional redundancy becauseof the same complementarity mechanisms that in-creased TDP flux in species-rich treatments. Functionaldifferences (i.e., feeding behavior and vertical spatialdistributions) among species may stabilize their abilityto coexist by relaxing interspecific competition evenwhen community biomass is high. On the other hand,the intensity of intraspecific competition pressureshould increase as biomass increases in monocultures,thereby reducing the biomass–TDP flux reliability.
Consequences of bioturbator diversity for habitat couplingin lake ecosystems
The fact that bioturbator diversity enhances benthic–pelagic processes (nutrient flux rates) and properties(nutrient flux reliability) in freshwater lacustrineecosystems confirms early empirical observations andtheoretical predictions derived from marine andstream systems (Palmer et al. 2000, Emmerson et al.2001, Biles et al. 2003, Covich et al. 2004, Jonsson andMalmqvist 2005). Extrapolations of short-term labora-tory experiments to the field should be made withcaution, but our results support 2 opposing points ofview. High benthic invertebrate diversity should beimportant for nutrient regeneration in oligotrophicecosystems because it can enhance benthic–pelagiccoupling, with potential indirect feedbacks on benthicproduction if more food for invertebrates sinks to thesediment as pelagic production increases. However,high benthic invertebrate diversity could be undesir-able to lake restoration/management efforts in eutro-phic systems if it enhances remobilization of P that hadbeen buried in sediment back into the water column,thereby exacerbating eutrophication. Further researchis needed to elucidate how direct and indirect effects ofbioturbator diversity on nutrient release from thesediment propagate into pelagic and benthic foodwebs under different trophic states.
Our findings are particularly important to ourunderstanding of the functioning of shallow lakeswhere the benthic habitat might subsidize ecosystem-level processes such as nutrient cycling and aquaticproductivity (Schindler and Scheuerell 2002). In these
systems, human-mediated loss of biodiversity throughcontamination, habitat modification, and introductionsof exotic species may severely alter habitat connectionsand, therefore, the fundamental flows of nutrients andenergy among aquatic compartments and their inter-acting communities.
Acknowledgements
We thank Walter Dodds, Nils Okun, Rafael Guar-iento, Albert Suhett, Adriana Rocha, and 2 anonymousreferees for constructive comments that improved thefinal version of this manuscript. We also are indebtedto Joao Marcelo and Alexandre Lopes for fieldassistance. This work was supported by grants fromthe International Foundation for Science to Vinicius F.Farjalla (A/4006–1) and from PETROBRAS. Scholar-ships were provided by Conselho Nacional deDesenvolvimento Cientıfico e Tecnologico, Coordena-cao de Aperfeicoamento de Pessoal de Nıvel Superior,and Fundacao Carlos Chagas Filho de Amparo aPesquisa do Estado do Rio de Janeiro.
Literature Cited
ALBERTONI, E. F., C. PALMA-SILVA, AND F. D. ESTEVES. 2001.Macroinvertebrates associated with Chara in a tropicalcoastal lagoon (Imboassica lagoon, Rio de Janeiro,Brazil). Hydrobiologia 457:215–224.
BELL, T., J. A. NEWMAN, B. W. SILVERMAN, S. L. TURNER, AND A.K. LILLEY. 2005. The contribution of species richness andcomposition to bacterial services. Nature 436:1157–1160.
BILES, C. L., M. SOLAN, I. ISAKSSON, D. M. PATERSON, C. EMES,AND D. G. RAFFAELLI. 2003. Flow modifies the effect ofbiodiversity on ecosystem functioning: an in situ studyof estuarine sediments. Journal of Experimental MarineBiology and Ecology 285:165–177.
BOLAM, S. G., T. F. FERNANDES, AND M. HUXHAM. 2002.Diversity, biomass, and ecosystem processes in themarine benthos. Ecological Monographs 72:599–615.
CALLISTO, M., J. F. GONCALVES-JUNIOR, J. J. F. LEAL, AND M. M.PETRUCIO. 1998. Macroinvertebrados bentonicos naslagoas Imboassica, Cabiunas, e Comprida. Pages 479–486 in F. A. Esteves (editor). Ecologia das LagoasCosteiras do Parque Nacional da Restinga de Jurubatibae do Municıpio de Macae (RJ). Nupem/UFRJ, Rio deJaneiro, Brazil.
CARDINALE, B. J., AND M. A. PALMER. 2002. Disturbancemoderates biodiversity-ecosystem function relationships:experimental evidence from caddisflies in stream meso-cosms. Ecology 83:1915–1927.
CARDINALE, B. J., M. A. PALMER, AND S. L. COLLINS. 2002.Species diversity enhances ecosystem functioningthrough interspecific facilitation. Nature 415:426–429.
CHAPIN, F. S., E. S. ZAVALETA, V. T. EVINER, R. L. NAYLOR, P. M.VITOUSEK, H. L. REYNOLDS, D. U. HOOPER, S. LAVOREL, O. E.SALA, S. E. HOBBIE, M. C. MACK, AND S. DIAZ. 2000.
2007] 457BENTHIC BIODIVERSITY AND THE P CYCLE

Consequences of changing biodiversity. Nature 405:234–242.
COTTINGHAM, K. L., J. T. LENNON, AND B. L. BROWN. 2005.Knowing when to draw the line: designing moreinformative ecological experiments. Frontiers in Ecologyand Environment 3:145–152.
COVICH, A. P., M. C. AUSTEN, F. BARLOCHER, E. CHAUVET, B. J.CARDINALE, C. L. BILES, P. INCHAUSTI, O. DANGLES, M.SOLAN, M. O. GESSNER, B. STATZNER, AND B. MOSS. 2004. Therole of biodiversity in the functioning of freshwater andmarine benthic ecosystems. BioScience 54:767–775.
DUFFY, J. E., J. P. RICHARDSON, AND E. A. CANUEL. 2003. Grazerdiversity effects on ecosystem functioning in seagrassbeds. Ecology Letters 6:637–645.
EMMERSON, M. C., AND D. G. RAFFAELLI. 2000. Detecting theeffects of diversity on measures of ecosystem function:experimental design, null models and empirical obser-vations. Oikos 91:195–203.
EMMERSON, M. C., M. SOLAN, C. EMES, D. M. PATERSON, AND D.RAFFAELLI. 2001. Consistent patterns and the idiosyncraticeffects of biodiversity in marine ecosystems. Nature 411:73–77.
ENGELHARDT, K. A. M., AND M. E. RITCHIE. 2002. The effect ofaquatic plant species richness on wetland ecosystemprocesses. Ecology 83:2911–2924.
FIGUEIREDO-BARROS, M. P. 2004. Bioturbacao na interfacesedimento-agua por tres especies de macroinvertebradosbentonicos com enfase na ciclagem de nutrientes. MSThesis, Instituto de Biofısica Carlos Chagas Filho,Universidade Federal do Rio de Janeiro, Rio de Janeiro,Brazil.
GALLEPP, G. W. 1979. Chironomid influence on phosphorusrelease in sediment-water microcosms. Ecology 60:547–556.
GOLTERMAN, H. L., R. S. CLYMO, AND M. A. M. OHNSTAD. 1978.Methods for physical and chemical analysis of freshwa-ter. Blackwell Scientific Publication, Oxford, UK.
GONCALVES-JUNIOR, J. F., J. J. F. LEAL, AND M. F. P. CALLISTO.1998. Population dynamic of Heleobia australis (Gastro-poda) in a coastal lagoon (Rio de Janeiro, Brazil).Verhandlungen der Internationalen Vereinigung furtheoretische und angewandte Limnologie 26:2056–2057.
GRANELI, W. 1979. Influence of Chironomus plumosus larvae onthe exchange of dissolved substances between sedimentand water. Hydrobiologia 66:149–159.
HOOPER, D. U., F. S. CHAPIN, J. J. EWEL, A. HECTOR, P. INCHAUSTI,S. LAVOREL, J. H. LAWTON, D. M. LODGE, M. LOREAU, S.NAEEM, B. SCHMID, H. SETALA, A. J. SYMSTAD, J. VANDERMEER,AND D. A. WARDLE. 2005. Effects of biodiversity onecosystem functioning: a consensus of current knowl-edge. Ecological Monographs 75:3–35.
HUSTON, M. A. 1997. Hidden treatments in ecologicalexperiments: re-evaluating the ecosystem function ofbiodiversity. Oecologia (Berlin) 110:449–460.
IENO, E. N., M. SOLAN, P. BATTY, AND G. J. PIERCE. 2006. Howbiodiversity affects ecosystem functioning: roles ofinfaunal species richness, identity and density in themarine benthos. Marine Ecology Progress Series 311:263–271.
JOLLIFFE, P. A. 2000. The replacement series. Journal ofEcology 88:378–385.
JONSSON, M., AND B. MALMQVIST. 2005. Species richness andcomposition effects in a detrital processing chain. Journalof the North American Benthological Society 24:798–806.
LAWTON, J. H., AND V. K. BROWN. 1994. Redundancy inecosystems. Pages 255–270 in E. D. Schulze and H. A.Mooney (editors). Biodiversity and ecosystem function.Springer–Verlag, Berlin, Germany.
LEVINTON, J. 1995. Bioturbators as ecosystem engineers:control of the sediment fabric, inter-individual interac-tions, and material fluxes. Pages 29–36 in C. B. Jones andJ. H. Lawton (editors). Linking species and ecosystems.Chapman and Hall, New York.
LOHRER, A. M., S. F. THRUSH, AND M. M. GIBBS. 2004.Bioturbators enhance ecosystem function through com-plex biogeochemical interactions. Nature 431:1092–1095.
LOREAU, M. 1998. Separating chance and other effects inbiodiversity experiments. Oikos 82:600–602.
LOREAU, M. 2004. Does functional redundancy exist? Oikos104:606–611.
LOREAU, M., S. NAEEM, AND P. INCHAUSTI (EDITORS). 2002.Biodiversity and ecosystem functioning: synthesis andperspectives. Oxford University Press, New York.
MARINELLI, R. L., AND T. J. WILLIAMS. 2003. Evidence fordensity-dependent effects of infauna on sediment bio-geochemistry and benthic-pelagic coupling in nearshoresystems. Estuarine, Coastal and Shelf Sciences 57:179–192.
MERMILLOD-BLONDIN, F., F. FRANCOIS-CARCAILLET, AND R. ROSEN-
BERG. 2005. Biodiversity of benthic invertebrates andorganic matter processing in shallow marine sediments:an experimental study. Journal of Experimental MarineBiology and Ecology 315:187–209.
MERMILLOD-BLONDIN, F., M. GERINO, M. CREUZE DES CHATELLIERS,AND V. DEGRANGE. 2002. Functional diversity among 3detritivorous hyporheic invertebrates: an experimentalstudy in microcosms. Journal of the North AmericanBenthological Society 21:132–149.
MERMILLOD-BLONDIN, F., M. GERINO, S. SAUVAGE, AND M. CREUZE
DES CHATELLIERS. 2004. Influence of nontrophic interac-tions between benthic invertebrates on river sedimentprocesses: a microcosm study. Canadian Journal ofFisheries and Aquatic Sciences 61:1817–1831.
NAEEM, S. 1998. Species redundancy and ecosystem reliabil-ity. Conservation Biology 12:39–45.
NAEEM, S., AND S. B. LI. 1997. Biodiversity enhances ecosystemreliability. Nature 390:507–509.
NEIRA, C., AND T. HOPNER. 1994. The role of Heteromastusfiliformis (Capitellidae, Polychaeta) in organic carboncycling. Ophelia 39:55–73.
PALMER, M. A., A. P. COVICH, S. LAKE, P. BIRO, J. J. BROOKS, J.COLE, C. DAHM, J. GIBERT, W. GOEDKOOP, K. MARTENS, AND J.VERHOEVEN. 2000. Linkages between aquatic sedimentbiota and life above sediments as potential drivers ofbiodiversity and ecological processes. BioScience 50:1062–1075.
RAO, D. S. 1980. Ecology of Heteromastus similis Southern 1921(Polychaeta, Capitellidae) in the Vasishta Godovari.
458 [Volume 26A. CALIMAN ET AL.

Proceedings of the Indian Academy of Sciences: AnimalSciences 89:407–414.
REISE, K. 2002. Sediment mediated species interactions incoastal waters. Journal of Sea Research 48:127–141.
REYNOLDS, C. S., AND P. S. DAVIES. 2001. Sources andbioavailability of phosphorus fractions in freshwaters: aBritish perspective. Biological Reviews 76:27–64.
RICCIARDI, A., AND J. B. RASMUSSEN. 1999. Extinction rates ofNorth American freshwater fauna. Conservation Biology13:1220–1222.
SCHINDLER, D. W. 1977. Evolution of phosphorus limitation inlakes. Science 195:260–262.
SCHINDLER, D. E., AND M. D. SCHEUERELL. 2002. Habitatcoupling in lake ecosystems. Oikos 98:177–189.
STIEF, P., D. ALTMANN, D. DE BEER, R. BIEG, AND A. KURECK. 2004.Microbial activities in the burrow environment of thepotamal mayfly Ephoron virgo. Freshwater Biology 49:1152–1163.
STIEF, P., L. NAZAROVA, AND D. DE BEER. 2005. Chimneyconstruction by Chironomus riparius larvae in responseto hypoxia: microbial implications for freshwater sedi-ments. Journal of the North American BenthologicalSociety 24:858–871.
TILMAN, D., J. KNOPS, D. WEDIN, P. REICH, M. RITCHIE, AND E.
SIEMANN. 1997. The influence of functional diversity and
composition on ecosystem processes. Science 277:1300–
1302.
WALDBUSSER, G. G., AND R. L. MARINELLI. 2006. Macrofaunal
modification of porewater advection: role of species
function, species interaction, and kinetics. Marine Ecol-
ogy Progress Series 311:217–231.
WALDBUSSER, G. G., R. L. MARINELLI, R. B. WHITLATCH, AND P. T.
VISSCHER. 2004. The effects of infaunal biodiversity on
biogeochemistry of coastal marine sediments. Limnology
and Oceanography 49:1482–1492.
WARDLE, D. A. 1999. Is ‘‘sampling effect’’ a problem for
experiments investigating biodiversity-ecosystem func-
tion relationships? Oikos 87:403–407.
WETZEL, R. G. 2001. Limnology: lake and river ecosystems.
Academic Press, San Diego, California.
YACHI, S., AND M. LOREAU. 1999. Biodiversity and ecosystem
productivity in a fluctuating environment: the insurance
hypothesis. Proceedings of the National Academy of
Sciences of the United States of America 96:1463–1468.
Received: 5 May 2006
Accepted: 23 February 2007
2007] 459BENTHIC BIODIVERSITY AND THE P CYCLE





























