Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, Feb. 2007, p. 1062–1071 Vol. 81, No. 30022-538X/07/$08.00�0 doi:10.1128/JVI.01558-06Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Targeted Disruption of Kaposi’s Sarcoma-Associated HerpesvirusORF57 in the Viral Genome Is Detrimental for the Expression of
ORF59, K8�, and K8.1 and the Production of Infectious Virus�
Vladimir Majerciak,1 Natalia Pripuzova,1 J. Philip McCoy,2 Shou-Jiang Gao,3 and Zhi-Ming Zheng1*HIV and AIDS Malignancy Branch, Center for Cancer Research, NCI,1 and Flow Cytometry Core Facility, NHLBI,2
NIH, Bethesda, Maryland, and Children’s Cancer Research Institute and Departments of Pediatrics and Microbiology,The University of Texas Health Science Center at San Antonio, San Antonio, Texas3
Received 20 July 2006/Accepted 5 November 2006
Kaposi’s sarcoma-associated herpesvirus (KSHV) ORF57 regulates viral gene expression at the posttranscrip-tional level during viral lytic infection. To study its function in the context of the viral genome, we disrupted KSHVORF57 in the KSHV genome by transposon-based mutagenesis. The insertion of the transposon into the ORF57exon 2 region also interrupted the 3� untranslated region of KSHV ORF56, which overlaps with the ORF57 codingregion. The disrupted viral genome, Bac36-�57, did not express ORF57, ORF59, K8�, K8.1, or a higher level ofpolyadenylated nuclear RNA after butyrate induction and could not be induced to produce infectious viruses in thepresence of valproic acid, a histone deacetylase inhibitor and a novel KSHV lytic cycle inducer. The ectopicexpression of ORF57 partially complemented the replication deficiency of the disrupted KSHV genome and theexpression of the lytic gene ORF59. The induced production of infectious virus particles from the disrupted KSHVgenome was also substantially restored by the simultaneous expression of both ORF57 and ORF56; complemen-tation by ORF57 alone only partially restored the production of virus, and expression of ORF56 alone showed noeffect. Altogether, our data indicate that in the context of the viral genome, KSHV ORF57 is essential for ORF59,K8�, and K8.1 expression and infectious virus production.
Kaposi’s sarcoma-associated herpesvirus (KSHV), alsoknown as human herpesvirus 8, is a gamma-2 herpesvirus.KSHV has a genome of �165 kb that encodes up to 90 viralgenes, and its genome shares significant sequence homologywith that of the Epstein-Barr virus and herpesvirus saimiri (5,33, 37). As with other gammaherpesviruses, KSHV infectioncan be either latent or lytic depending on cellular conditions(39). What makes the virus become latent during the infectionremains largely unknown, but the latent KSHV genome per-sists in a nonintegrated circular episomal form as it does in thelytic stage (1, 2, 53). The latent KSHV can be activated ininfected cells by chemical inducers, such as sodium n-butyrate(31), 12-O-tetradecanoylphorbol-13-acetate (TPA) (36), andvalproic acid (VA) (15, 40), and the resulting lytic gene ex-pression leads to production of infectious virus. Expression ofa viral transactivator, ORF50 (RTA), as a result of chemicalinduction is essential for the lytic switch from KSHV latency(26, 42).
Nuclear transcription of viral lytic genes can be robustlyactivated in trans by ORF50 (25, 42), but the newly synthesizedviral transcripts must be processed correctly at the posttran-scriptional level, including RNA 5� capping, splicing, 3� poly-adenylation, and export from the nucleus to the cytoplasm, toensure their efficient protein translation. The members of theherpesvirus family have evolved a specific mechanism to facil-itate this process, the encoding of a posttranscriptional regu-
latory protein that promotes the expression of specific viraltranscripts (43, 54). KSHV ORF57, which is transactivated byORF50 (24, 48), encodes a viral early nuclear protein of 455amino acid residues (9, 14). ORF57 promotes viral gene ex-pression by mediating the nuclear export of viral RNAs in aCRM1-independent manner, presumably through its interac-tion with the cellular export factor Aly/REF (30), similar to itshomologs in other herpesviruses (12, 16, 49). However, thisfunction of ORF57 has never been studied in the context of theviral genome during lytic infection. Moreover, our recent datashowed that interactions between the three nuclear localiza-tion signals of ORF57 and Aly/REF are not essential forORF57-mediated accumulation of KSHV ORF59 transcriptsin living cells (28).
To study KSHV ORF57 function in the context of theKSHV genome during lytic viral infection, we took the geneticapproach of disrupting ORF57 in a KSHV genome con-structed in a bacterial artificial chromosome, Bac36 (56). Sinceits construction, Bac36 has been widely used to characterizeKSHV gene function by the genetic manipulation of individualviral genes within the viral genome (27, 51, 53). In this study,we demonstrate that when the ORF57 gene is disrupted, theKSHV genome cannot express a subset of viral lytic genes andproduce infectious virions in response to chemical induction.
MATERIALS AND METHODS
Cells. Human 293 cells were obtained from the American Type Culture Col-lection (Manassas, VA) and were cultivated in Dulbecco’s modified Eagle me-dium (Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum(HyClone, Logan, UT), 2 mM L-glutamine, 100 U/ml penicillin, and 100 g/mlstreptomycin (Invitrogen).
Plasmid construction. Construction of the KSHV ORF57 expression vectorpVM7 (pORF57) was described previously (28). To create the ORF56 expres-
* Corresponding author. Mailing address: HIV and AIDS Malig-nancy Branch, Center for Cancer Research, NCI/NIH, 10 Center Dr.,Rm. 10 S255, MSC-1868, Bethesda, MD 20892-1868. Phone: (301)594-1382. Fax: (301) 480-8250. E-mail: [email protected].
� Published ahead of print on 15 November 2006.
1062

sion vector pVM9 (pORF56), the KSHV ORF56 open reading frame (nucleo-tides [nt] 79433 to 81946 of the KSHV genome) was amplified by PCR andcloned into a pFLAG-CMV-5.1 vector (Sigma, St. Louis, MO). The insert wasconfirmed by enzyme digestion and sequencing. The constructed vectors expressORF57 and ORF56 with a C-terminal FLAG tag. An RTA expression vector,pORF50, was a gift from Yan Yuan of the University of Pennsylvania.
Construction of a KSHV ORF57-null mutant. We chose Bac36 to create anORF57-null mutant by random transposon-based mutagenesis with an EZ::TN�KAN-2� kit (Epicenter, Madison, WI). Bac36 contains a full-length KSHVgenome and a hygromycin B resistance gene and expresses green fluorescentprotein as a marker (56). Briefly, purified KSHV Bac36 DNA (Bac36-wt) wasincubated for 2 h with Tn5 transposase, and a 1.2-kb transposon cassette thatcontains a kanamycin resistance gene (Kanr) and was then transformed intoEscherichia coli strain DH10B by electroporation. The recombinant Bac cloneswith transposon insertions were selected on Luria-Bertani medium agar plates inthe presence of kanamycin (50 �g/ml). Individual clones were expanded andsequenced using the KAN-2 FP-1 (5�-ACCTACAACAAAGCTCTCATCAACC-3�) and RP-1 (5�-GCAATGTAACATCAGAGATTTTGAG-3�) primers todetermine the transposon insertion site. A KSHV Bac clone with a transposoninsert in the ORF57 coding region was designated Bac36-�57. All Bac36 DNAswere isolated from E. coli using a QIAGEN Large-Construct kit (QIAGEN,Valencia, CA).
Genetic analysis of the KSHV ORF57-null mutant. Several methods wereemployed to confirm that the transposon was inserted into the ORF57 codingregion without disrupting the integrity of the rest of the viral genome. First, wedesigned a pair of primers flanking the insert, oVM12 (Pr82699; 5�-CTCAGACTCCCTGCGAGCAT-3�) and oVM13 (Pr83151; 5�-TCTGGTAACAAACGCATTGC-3�), and analyzed the PCR products from the Bac36-wt and Bac36-�57genomes by agarose gel electrophoresis. The integrity of the viral genome aftertransposon insertion was determined by restriction enzyme digestion of Bac36DNAs. One microgram of purified Bac36 DNAs (wild type and mutant) wasdigested with KpnI, and the resulting DNA fragments were separated in a 0.7%agarose gel together with 1-kb DNA ladders (Invitrogen) at 40 V for 14 h. Afterseparation, the DNA fragments were transferred onto a nylon membrane andanalyzed by Southern blot analysis. Probe labeling and hybridization were carriedout using an AlkPhos direct labeling kit (Amersham, Piscataway, NJ) accordingto the manufacturer’s instructions. Briefly, 100 ng of purified ORF57-specificPCR products from nt 82699 to 83151 of the KSHV genome or a EZ::TNXhoI-BamHI fragment was directly labeled with alkaline phosphatase using achemical cross-linker, and the chemically labeled probes were then hybridized tothe membrane at 55°C overnight. After washes, the bound probe was detected bychemiluminescence using the CPD-Star detection reagent (Amersham) and ex-posed to X-ray film. After detection, the membrane was stripped in 0.5% sodiumdodecyl sulfate (SDS) at 60°C for 1 h, reconstituted in 100 mM Tris-HCl (pH 8.0)for 5 min at room temperature, and reprobed with another probe.
Transient transfection and establishment of stable cell lines. Purified Bac36DNAs were transfected into 293 cells using Lipofectamine 2000 (Invitrogen).Hygromycin B (Sigma, 150 ng/ml) was then added for selection 24 h aftertransfection. Three weeks after selection, a homogenous population of greenfluorescent protein (GFP)-positive cells harboring KSHV Bac36 DNA was ob-tained. Butyrate (Sigma) at a final concentration of 3 mM, TPA (Sigma) at 20ng/ml, or VA (Sigma) at 1 mM was used for lytic induction. Transient transfec-tion of the stable cell lines with an ORF50 expression vector, pORF50, was alsoused for lytic induction.
Western blot analysis. Protein samples were prepared by direct lysis of cells in2 SDS protein sample buffer containing 5% 2-mercaptoethanol. The lysed cellswere boiled and separated in SDS-polyacrylamide gel electrophoresis (PAGE)gels. The following antibodies were used in Western blot analyses: a rabbitpolyclonal anti-ORF57 antibody against a synthetic peptide (amino acids 119 to132 of ORF57 [unpublished data]; used at a dilution of 1:3,000), a monoclonalimmunoglobulin M-type anti--tubulin antibody (1:1,000; BD Pharmingen, SanDiego, CA), a polyclonal anti-GFP antibody (1:3,000; Clontech, Mountain View,CA), and a monoclonal anti-FLAG M2 antibody (1:2,500; Sigma), together withcorresponding horseradish peroxidase-conjugated secondary antibodies (1:10,000;Sigma). The signal on the Western blot was detected with a West Pico chemi-luminescence substrate (Pierce, Rockford, IL).
RPA. Total cell RNA was prepared using the TRIzol reagent (Invitrogen)according to the manufacturer’s instructions. An antisense ORF59-specific probecovering nt 95870 to 96171 of the KSHV genome and an antisense polyadeny-lated nuclear (PAN) RNA probe covering nt 29400 to 29608 were transcribed invitro. An antisense human cyclophilin probe (Ambion, Austin, TX) was used asan internal control for sample loading. All probes were prepared using theRiboprobe system (Promega, Madison, WI) in the presence of [�-32P]GTP. An
RNase protection assay (RPA) was carried out using an RPA III kit (Ambion)according to the manufacturer’s instructions. Briefly, 30 to 40 �g of total RNAwas hybridized with 4 ng of each probe overnight at 42°C and then digested witha mixture of RNase A and RNase T1 for 30 min at 37°C. Protected fragmentswere separated in an 8% PAGE denaturing gel. The signals were captured usinga Molecular Dynamics Storm 860 PhosphorImager and analyzed with Image-Quant software (Amersham).
Immunofluorescence staining. Before staining, stable Bac36 cells were grownon coverslips and the viral lytic cycle was induced with butyrate. Twenty-fourhours after induction, the monolayers were washed twice with phosphate-buff-ered saline (PBS), fixed with 2% paraformaldehyde in PBS for 20 min at roomtemperature, and quenched twice with 100 mM glycin in PBS (10 min each). Forintracellular staining, the cells were permeabilized with 0.1% Triton X-100 inPBS for 10 min at room temperature. Blocking was performed using blockingsolution (3% bovine serum albumin in PBS with 0.05% Tween 20) for 1 h at37°C. All primary and secondary antibodies were diluted 1:50 in blocking solu-tion and incubated for 1 h at 37°C, followed by three washes (10 min each) withPBS containing 0.05% Tween 20. The following primary antibodies were used:rabbit polyclonal anti-ORF57 antibody (as described above), mouse monoclonalanti-K8 (Promab, Albany, CA), mouse monoclonal anti-ORF59 (Advanced Bio-technologies, Columbia, MD), rat monoclonal anti-ORF73 (anti-LANA; Ad-vanced Biotechnologies), and rabbit polyclonal anti-K8.1 antibody (19), togetherwith corresponding tetramethylrhodamine isothiocyanate (TRITC)-conjugatedsecondary antibodies (1:50; Sigma). Fluorescence images were collected andsaved in TIFF format, and Adobe Photoshop 6.0 software (Adobe Systems, SanJose, CA) was used to process the images into composite figures.
Virus production and complementation assay. To activate the lytic phase andvirion production, the stable Bac36 cells were incubated with 1 mM VA for 72 h.After induction, the free-virus-containing supernatant (30 ml from a T-162 flask)was clarified by low-speed centrifugation (5,000 g, 30 min) to remove cells andcell debris. The virus in the clarified culture supernatant was then concentratedby high-speed centrifugation (25,000 g, 3 h). The virus pellets were resus-pended in an equal volume (700 �l) of serum-free Dulbecco’s modified Eaglemedium, and 300 �l of the resuspension was titrated in duplicate on freshlyplated 293 cells with the addition of 5 �g/ml polybrene (Sigma). The inoculatedcells were examined daily for the appearance of GFP-positive cells as an indi-cation of viral infection. Within 72 to 96 h after viral infection, the cells weretrypsinized, and the cell suspension was analyzed by flow cytometry to determinethe number of GFP-positive cells. The complementation assay was carried out bytransient transfection of Bac36 cells with ORF57 or ORF56 expression vectorsbefore VA induction. The empty vector, pFLAG-CMV-5.1, was used as a con-trol. Twenty-four hours after transfection, the lytic cycle was induced by VA asdescribed above.
RESULTS
Construction of a KSHV Bac36-�57 mutant. To create aKSHV virus lacking ORF57 expression, we utilized Bac36,which contains the wt KSHV genome (Bac36-wt) (56), as atemplate for random transposon-based mutagenesis. The mu-tant KSHV genome was screened by sequencing and showed atransposon insertion at the nt 83001 position in the N-terminalORF57 coding region (Fig. 1A), along with duplication of a9-bp KSHV genome sequence on each end of the insert. Theinsertion took place 998 nt downstream of the ORF57 tran-scription start site and disrupted exon 2 of ORF57. Accord-ingly, the selected mutant was named Bac36-�57. To furtherverify the insertion position, we carried out a PCR assay usinga paired primer set, oVM12 and oVM13, flanking the insertionsite of Bac36-�57. The PCR amplification generated a 453-bpproduct from Bac36-wt with no insertion and a 1.6-kb productfrom Bac36-�57 containing the 1.2-kb transposon cassette(Fig. 1B), confirming the results of the sequencing.
We further analyzed Bac36-�57 by restriction enzyme diges-tion and Southern blotting. Purified Bac36-wt and Bac36-�57DNA digested with KpnI were first separated in an agarose gelfor ethidium bromide staining and then transferred onto anylon membrane for Southern blot analysis. As shown in Fig.
VOL. 81, 2007 DISRUPTION OF KSHV ORF57 REDUCES LYTIC GENE EXPRESSION 1063
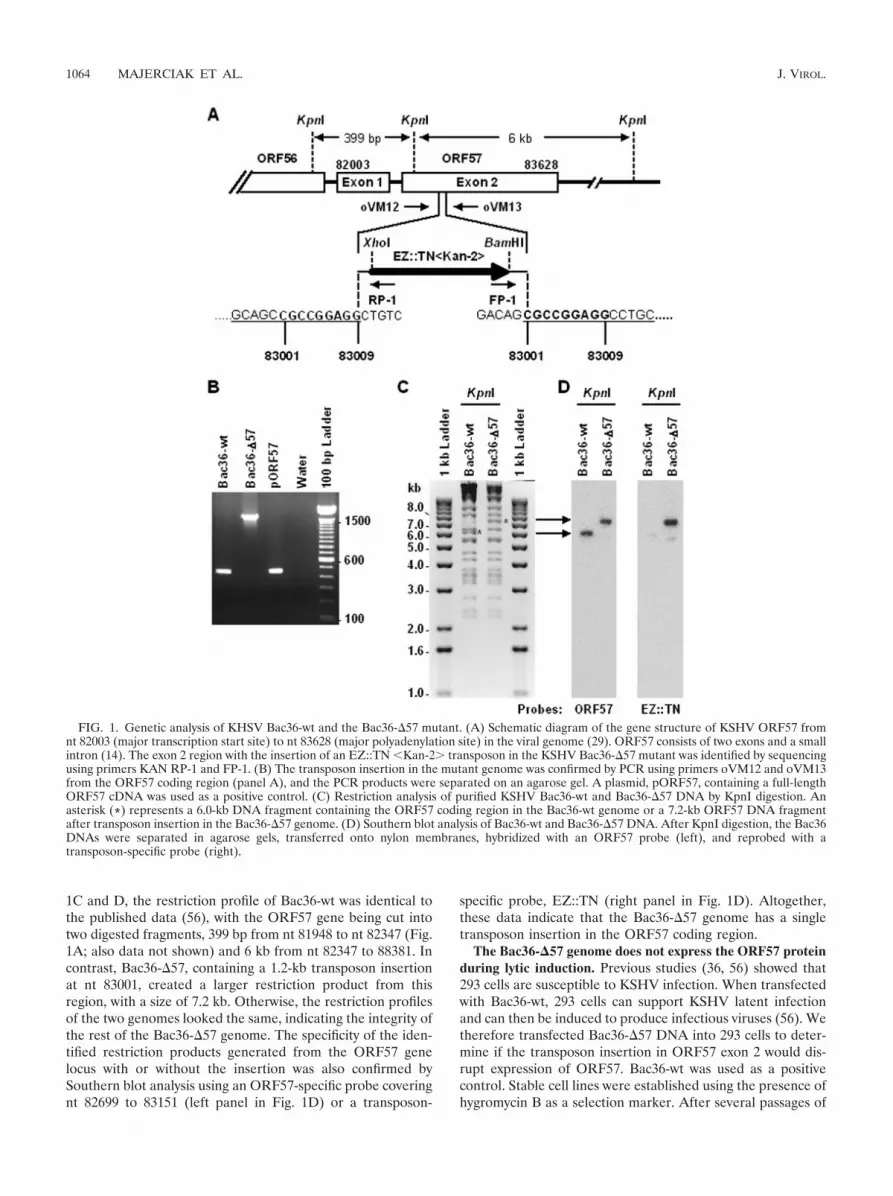
1C and D, the restriction profile of Bac36-wt was identical tothe published data (56), with the ORF57 gene being cut intotwo digested fragments, 399 bp from nt 81948 to nt 82347 (Fig.1A; also data not shown) and 6 kb from nt 82347 to 88381. Incontrast, Bac36-�57, containing a 1.2-kb transposon insertionat nt 83001, created a larger restriction product from thisregion, with a size of 7.2 kb. Otherwise, the restriction profilesof the two genomes looked the same, indicating the integrity ofthe rest of the Bac36-�57 genome. The specificity of the iden-tified restriction products generated from the ORF57 genelocus with or without the insertion was also confirmed bySouthern blot analysis using an ORF57-specific probe coveringnt 82699 to 83151 (left panel in Fig. 1D) or a transposon-
specific probe, EZ::TN (right panel in Fig. 1D). Altogether,these data indicate that the Bac36-�57 genome has a singletransposon insertion in the ORF57 coding region.
The Bac36-�57 genome does not express the ORF57 proteinduring lytic induction. Previous studies (36, 56) showed that293 cells are susceptible to KSHV infection. When transfectedwith Bac36-wt, 293 cells can support KSHV latent infectionand can then be induced to produce infectious viruses (56). Wetherefore transfected Bac36-�57 DNA into 293 cells to deter-mine if the transposon insertion in ORF57 exon 2 would dis-rupt expression of ORF57. Bac36-wt was used as a positivecontrol. Stable cell lines were established using the presence ofhygromycin B as a selection marker. After several passages of
FIG. 1. Genetic analysis of KHSV Bac36-wt and the Bac36-�57 mutant. (A) Schematic diagram of the gene structure of KSHV ORF57 fromnt 82003 (major transcription start site) to nt 83628 (major polyadenylation site) in the viral genome (29). ORF57 consists of two exons and a smallintron (14). The exon 2 region with the insertion of an EZ::TN �Kan-2� transposon in the KSHV Bac36-�57 mutant was identified by sequencingusing primers KAN RP-1 and FP-1. (B) The transposon insertion in the mutant genome was confirmed by PCR using primers oVM12 and oVM13from the ORF57 coding region (panel A), and the PCR products were separated on an agarose gel. A plasmid, pORF57, containing a full-lengthORF57 cDNA was used as a positive control. (C) Restriction analysis of purified KSHV Bac36-wt and Bac36-�57 DNA by KpnI digestion. Anasterisk (*) represents a 6.0-kb DNA fragment containing the ORF57 coding region in the Bac36-wt genome or a 7.2-kb ORF57 DNA fragmentafter transposon insertion in the Bac36-�57 genome. (D) Southern blot analysis of Bac36-wt and Bac36-�57 DNA. After KpnI digestion, the Bac36DNAs were separated in agarose gels, transferred onto nylon membranes, hybridized with an ORF57 probe (left), and reprobed with atransposon-specific probe (right).
1064 MAJERCIAK ET AL. J. VIROL.

selection, all cells contained Bac36 DNAs and became GFPpositive. To examine the expression of ORF57 from bothBac36-wt and Bac36-�57, we developed a rabbit anti-ORF57polyclonal antibody against a synthetic peptide correspondingto amino acids 119 to 132 of the ORF57 protein, upstream ofthe insertion site. In a Western blot analysis, the antibodyrecognized a full-length ORF57 protein with the predicted sizeof 50 kDa in butyrate- or TPA-induced Bac36-wt stable cellsbut not in uninduced Bac36-wt cells (Fig. 2A, upper panel),indicating its specificity for ORF57. In contrast, no protein ofany size was detectable in butyrate- or TPA-induced Bac36-�57 stable cells (Fig. 2A, upper panel), despite detection ofsimilar amounts of tubulin protein in all samples (Fig. 2A,lower panel). To further confirm the Western blot results, weperformed immunofluorescence staining of stable Bac36-wt(Fig. 2B, upper panel) and Bac36-�57 (lower panel) cells 24 hafter butyrate induction. Again, only the stable cells with aBac36-wt genome were found to express ORF57, although allBac36-wt and Bac36-�57 stable cells were GFP positive, anindication that they harbor the KSHV genome. From thesedata, we conclude that the insertion of the transposon in the
Bac36-�57 genome completely disrupted the production of theORF57 protein.
ORF57 is essential for the expression of a subset of virallytic genes. To determine whether the lack of ORF57 expres-sion would affect the expression of other viral lytic genes, weanalyzed five viral genes that are expressed at different stagesof the viral cycle: ORF59, K8�, K8.1, LANA, and PAN RNA.ORF59, K8�, and PAN are viral early lytic genes. ORF59encodes a viral DNA polymerase processivity factor for viralDNA replication (3, 6, 21) and is a downstream target ofORF57 in transient assays (14, 28). K8� encodes a viral K-bZIP protein involved in viral gene transcription and DNAreplication (20, 22, 57). PAN expresses an abundant viral non-coding RNA with unknown function (41, 55) that was alsoshown to be a downstream target of ORF57 in transient trans-fection experiments (14). K8.1 is a viral late gene encoding anenvelope glycoprotein (4, 19, 35, 45). LANA is a latent geneencoding a latency-associated nuclear antigen responsible forKSHV genome segregation (2, 13, 18). Twenty-four hours afterinduction of the lytic viral cycle by butyrate, we compared theexpression of ORF57, ORF59, K8�, K8.1, and LANA in stableBac36-wt and Bac36-�57 cells using indirect immunofluores-cence staining with a specific antibody against each protein. Asubstantial number of Bac36-wt stable cells were positive forORF57, ORF59, and K8�, along with many fewer cells thatwere positive for K8.1, indicating that the lytic cycle was initi-ated by the induction, but the majority of the stable cellsremained at the early stage of viral replication. In contrast, noBac36-�57 cells expressed ORF59, K8�, and K8.1 despite ex-pressing LANA at levels similar to those for Bac36-wt cells(Fig. 3A). Consistent with our findings, a recent study usingVero cells stably transfected with an ORF57-null KSHV ge-nome also showed no ORF59 expression after induction withORF50 (10).
We next compared the two stable cell lines for their expres-sion of ORF59 and PAN RNAs, which had been identified asORF57 targets in another study (14) but never confirmed inthe context of viral replication. We performed an RPA on totalRNA isolated from the stable cell lines 24 h after butyrateinduction. As predicted, we found that both the ORF59 tran-script (Fig. 3B) and PAN RNA (Fig. 4A) were present in theBac36-wt stable cells, but very little was present in the Bac36-�57 stable cells (compare lane 4 to lane 5 in Fig. 3B and lane5 to lane 6 in Fig. 4A) in spite of the fact that a few stable cellsshowed some residual PAN RNA expression in the absence oflytic induction (Fig. 4A, lanes 3 and 4). When RTA (ORF50)was used to transactivate PAN RNA expression from thetwo cell lines, we also found that the amount of PAN RNAexpressed by the Bac36-�57 cells was much less (Fig. 4A),but RTA transactivation was less potent than that by bu-tyrate in induction under our conditions (compare lanes 7and 8 with lanes 5 and 6 in Fig. 4A). Similar results were alsoobtained by transient transfection of 293 cells with the twoBac36 DNAs and butyrate induction (Fig. 4B and C). Dataindicate that PAN RNA is indeed an ORF57 target in thecontext of the KSHV genome. Based on all of those obser-vations, we conclude that the Bac36-�57 genome cannotexpress a subset of viral lytic genes in the absence of theORF57 protein.
However, we found that both the Bac36-�57 genome and
FIG. 2. Expression of the ORF57 protein in 293 cells transfectedwith Ba36-wt and Bac36-�57 mutant DNA. 293 cells were transfectedwith Bac36 DNAs and selected for stable cells in the presence ofhygromycin B. (A) Western blot analysis of protein extracts fromBac36 DNA-harboring stable cell lines with or without 24 h of induc-tion by 3 mM sodium n-butyrate (NB) or 20 ng/ml phorbol ester(TPA). A rabbit polyclonal anti-ORF57 antibody was used to detectthe ORF57 protein in cell extracts, and the sample loading was con-trolled by tubulin. (B) Immunofluorescent staining of butyrate-inducedORF57 protein expression in Bac36-wt (upper panel) and Bac36-�57(lower panel) stable cell lines. All stable cells expressed GFP (middlepanels) independently of induction. Cells grown on coverslips werestained with an anti-ORF57 antibody in combination with a TRITC-conjugated secondary antibody after 24 h of butyrate induction.
VOL. 81, 2007 DISRUPTION OF KSHV ORF57 REDUCES LYTIC GENE EXPRESSION 1065

the Bac36-wt genome, in the absence of any induction, couldspontaneously express PAN RNA to a comparable level intransiently transfected 293 cells independently of ORF57 (Fig.4D and E).
Disruption of ORF57 in the KSHV genome reduces produc-tion of infectious virus. Because ORF57 disruption affects ex-pression of viral early genes important for viral DNA replica-tion, we wanted to determine whether ORF57 is essential forthe production of cell-free infectious virus. Initially, we com-pared the efficiencies of various chemical inducers at inducinginfectious virus production from the Bac36-wt stable cells andfound that 1 mM VA was the most effective (data not shown).VA is a prescription drug that has been used to treat patientswith seizures and bipolar disorders (23, 34). A recent studyshowed that, similar to butyrate, VA inhibits histone deacety-lases (8, 32), and it is very potent for inducing the expression oflytic KSHV genes in BCBL-1 cells (15, 40) but is much lesstoxic than butyrate (8, 32). This minimal toxicity allowed us touse VA for long-term induction to study virus production. Wetreated both the Bac36-wt and the Bac36-�57 stable cells withVA for 72 h, harvested the virus-containing culture superna-tants, clarified the supernatants with low-speed centrifugation,and concentrated the virus with high-speed centrifugation. Theproduction of infectious virus from each stable cell line wastitrated with fresh 293 cells, and the infected cells were exam-ined daily for GFP expression by fluorescence microscopy. Asshown in Fig. 5A, the supernatant collected from the VA-induced Bac36-wt cells was infectious, with the infected 293cells becoming GFP positive 48 h after infection. The titer inthe concentrated virus resuspension was about 3.3 102 in-fectious viruses/ml. In contrast, 293 cells did not express GFP48 h after treatment with the supernatant obtained from theVA-induced Bac36-�57 stable cells, indicating that the Bac36-�57 cells produced few, if any, infectious viruses. To betterdetermine the numbers of infected 293 cells in each sample,the inoculated 293 cells were collected 96 h after infection, andGFP-positive cells were quantified by flow cytometry. Approx-imately 60 cells per 100,000 collected were GFP positive in theBac36-wt sample, but only 15 cells per 100,000 treated with theBac36-�57 supernatant were GFP positive (compare bar 1 tobar 2 in Fig. 5B). These data indicate that disruption of ORF57expression indeed disrupts the production of infectious viruses.
To ensure that the expression of GFP in 293 cells was me-diated solely through infectious virus particles, the concen-trated virus preparations were also treated with DNase I for 30min at 37°C to remove any possible residual cell debris con-taminated with the Bac36-�57 genome that might have trans-fected the fresh 293 cells. After the treatment, a flow cytometryanalysis found no GFP-positive cells among the cells treatedwith Bac36-�57 viral preparations; however, this treatmentalso substantially reduced the number of GFP-positive cellsobserved among the cells infected with Bac36-wt viral prepa-rations (data not shown).
Complementation of ORF57 deficiency in the Bac36-�57genome by ectopic expression of a functional ORF57. Ourrecent analysis of the ORF56 and ORF57 gene locus revealedthat ORF56 encodes a bicistronic RNA and utilizes a poly(A)signal downstream of ORF57 (29). Consequently, our targeteddisruption of the ORF57 coding region would also disrupt the3� untranslated region (UTR) of the ORF56 RNA and affectORF56 expression, because lack of a polyadenylation signalwould make the ORF56 RNA unstable even though its codingregion remained intact. KSHV ORF56 encodes a viral DNAprimase important for viral DNA replication (7, 50), and its
FIG. 3. Expression of ORF57, ORF59, K8�, K8.1, and LANA inBac36-wt and Bac36-�57 stable cell lines after lytic induction. (A) Immuno-fluorescent staining of ORF57, ORF59, K8�, K8.1, and LANA proteins inBac36 stable cells after 24 h of induction with butyrate. The stable cells werestained with specific antibodies (see Materials and Methods) in combinationwith TRITC-conjugated secondary antibodies. (B) The expression of ORF59transcripts from Bac36 stable cells. RNase protection assays using an anti-sense ORF59-specific probe were carried out on 30 �g of total cell RNAisolated from 293 cells with or without stable Bac36 transfection after treat-ment with butyrate for 24 h. Cellular cyclophilin RNA was used as a loadingcontrol. Yeast total RNA was used as a negative control.
1066 MAJERCIAK ET AL. J. VIROL.

expression increases in the presence of ORF57 (29). There-fore, disruption of ORF56 would constitute an additional de-fect in the Bac36-�57 genome, one that has been shown to bedetrimental to infectious virus production (7). To complement
the functions of the deleted sections of the Bac36-�57 genome,we cotransfected Bac36-�57 cells with expression vectors forORF56 and ORF57 FLAG tag fusions 1 day before VA in-duction. Four days after VA induction of the cells, the infec-
FIG. 4. Expression of PAN RNA in Bac36-wt and Bac36-�57 stable cell lines or transiently transfected cells. (A) Substantial PAN RNAexpression depends on ORF57. Bac36-wt and Bac36-�57 stable cells (5 105/well) were plated into a 6-well plate, 3 wells for each. The cells at24 h after seeding were treated with 3 mM of sodium butyrate (NB) or transfected with 3 �g of plasmid pORF50 (p50) and were harvested 24 hafter NB treatment or 48 h after transfection. The cells without treatment were used as uninduced controls. Total cell RNA was prepared, andeach RNA sample at 40 �g was examined by RPA for PAN RNA and cyclophilin expression with a PAN- and cyclophilin-specific probe. (B to E)PAN RNA expression in transiently transfected 293 cells with Bac36 DNAs. 293 cells at 5 105/well were plated into a 6-well plate and transfectedwith 5 �g of purified Bac36 DNAs. The cells in each well were split into two wells 48 h after transfection and were induced by NB (3 mM) 24 hafter the split for an additional 24 h (B and C) or cultivated for an additional 48 h without induction (E). Transfection efficiencies of 293 cells withBac36-wt or Bac36-�57 were compared by appearance of GFP-positive cells (D). The cell lysates and total cell RNA were prepared and examinedby Western blotting for ORF57 or GFP (B) or by RPA for PAN RNA or cyclophilin with a PAN- or cyclophilin-specific probe (C and E).Quantitation of PAN RNA levels in each sample (A, C, and E) was normalized to cyclophilin expression.
VOL. 81, 2007 DISRUPTION OF KSHV ORF57 REDUCES LYTIC GENE EXPRESSION 1067
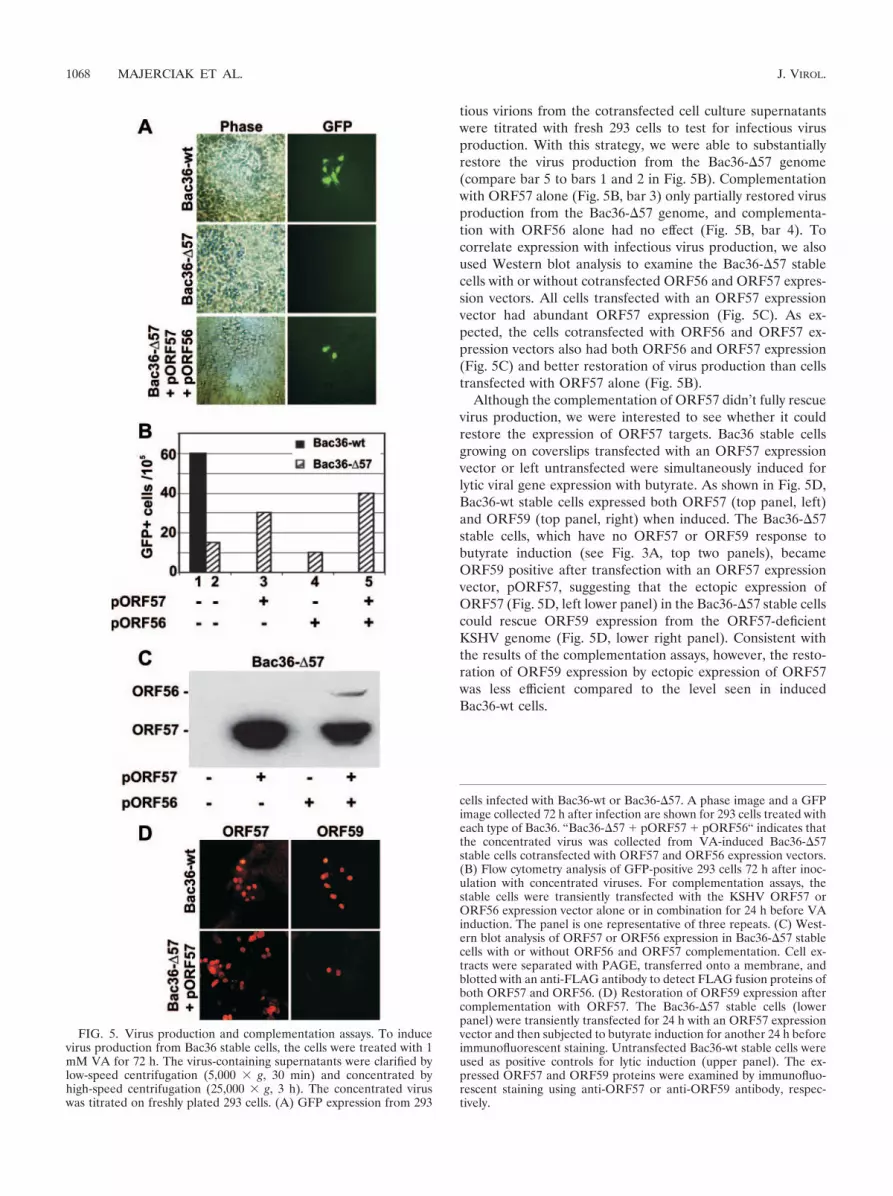
tious virions from the cotransfected cell culture supernatantswere titrated with fresh 293 cells to test for infectious virusproduction. With this strategy, we were able to substantiallyrestore the virus production from the Bac36-�57 genome(compare bar 5 to bars 1 and 2 in Fig. 5B). Complementationwith ORF57 alone (Fig. 5B, bar 3) only partially restored virusproduction from the Bac36-�57 genome, and complementa-tion with ORF56 alone had no effect (Fig. 5B, bar 4). Tocorrelate expression with infectious virus production, we alsoused Western blot analysis to examine the Bac36-�57 stablecells with or without cotransfected ORF56 and ORF57 expres-sion vectors. All cells transfected with an ORF57 expressionvector had abundant ORF57 expression (Fig. 5C). As ex-pected, the cells cotransfected with ORF56 and ORF57 ex-pression vectors also had both ORF56 and ORF57 expression(Fig. 5C) and better restoration of virus production than cellstransfected with ORF57 alone (Fig. 5B).
Although the complementation of ORF57 didn’t fully rescuevirus production, we were interested to see whether it couldrestore the expression of ORF57 targets. Bac36 stable cellsgrowing on coverslips transfected with an ORF57 expressionvector or left untransfected were simultaneously induced forlytic viral gene expression with butyrate. As shown in Fig. 5D,Bac36-wt stable cells expressed both ORF57 (top panel, left)and ORF59 (top panel, right) when induced. The Bac36-�57stable cells, which have no ORF57 or ORF59 response tobutyrate induction (see Fig. 3A, top two panels), becameORF59 positive after transfection with an ORF57 expressionvector, pORF57, suggesting that the ectopic expression ofORF57 (Fig. 5D, left lower panel) in the Bac36-�57 stable cellscould rescue ORF59 expression from the ORF57-deficientKSHV genome (Fig. 5D, lower right panel). Consistent withthe results of the complementation assays, however, the resto-ration of ORF59 expression by ectopic expression of ORF57was less efficient compared to the level seen in inducedBac36-wt cells.
FIG. 5. Virus production and complementation assays. To inducevirus production from Bac36 stable cells, the cells were treated with 1mM VA for 72 h. The virus-containing supernatants were clarified bylow-speed centrifugation (5,000 g, 30 min) and concentrated byhigh-speed centrifugation (25,000 g, 3 h). The concentrated viruswas titrated on freshly plated 293 cells. (A) GFP expression from 293
cells infected with Bac36-wt or Bac36-�57. A phase image and a GFPimage collected 72 h after infection are shown for 293 cells treated witheach type of Bac36. “Bac36-�57 � pORF57 � pORF56“ indicates thatthe concentrated virus was collected from VA-induced Bac36-�57stable cells cotransfected with ORF57 and ORF56 expression vectors.(B) Flow cytometry analysis of GFP-positive 293 cells 72 h after inoc-ulation with concentrated viruses. For complementation assays, thestable cells were transiently transfected with the KSHV ORF57 orORF56 expression vector alone or in combination for 24 h before VAinduction. The panel is one representative of three repeats. (C) West-ern blot analysis of ORF57 or ORF56 expression in Bac36-�57 stablecells with or without ORF56 and ORF57 complementation. Cell ex-tracts were separated with PAGE, transferred onto a membrane, andblotted with an anti-FLAG antibody to detect FLAG fusion proteins ofboth ORF57 and ORF56. (D) Restoration of ORF59 expression aftercomplementation with ORF57. The Bac36-�57 stable cells (lowerpanel) were transiently transfected for 24 h with an ORF57 expressionvector and then subjected to butyrate induction for another 24 h beforeimmunofluorescent staining. Untransfected Bac36-wt stable cells wereused as positive controls for lytic induction (upper panel). The ex-pressed ORF57 and ORF59 proteins were examined by immunofluo-rescent staining using anti-ORF57 or anti-ORF59 antibody, respec-tively.
1068 MAJERCIAK ET AL. J. VIROL.

DISCUSSION
In this study, we genetically analyzed the importance ofKSHV ORF57 for the expression of lytic viral genes in thecontext of the KSHV genome. Insertion of a transposon-con-taining cassette into the ORF57 exon 2 region of the KSHVgenome in Bac36 rendered the KSHV genome incapable ofexpressing ORF57 and producing infectious viruses, demon-strating that KSHV ORF57 is essential for virus production. Inaddition, the mutant genome lacking ORF57 expression wasalso unable to express ORF59, K8�, K8.1, or a higher level ofPAN RNA, thus establishing a subset of lytic viral genes ex-pressed in both the early and late viral life cycles as targets ofORF57. Since transient transfection of an ORF56 expressionvector did not lead to detectable levels of the ORF56 proteinin the absence of ORF57, we hypothesize that ORF56 expres-sion also depends on ORF57. The lack of an available anti-ORF56 antibody prevents us from pursuing this assumptionfurther in the context of the viral genome. LANA is not anORF57 target, because lack of ORF57 expression in the mu-tant KSHV genome had no effect on the expression of LANA.Since LANA expression in latent KSHV infection is indepen-dent of lytic gene expression, it was not surprising that ORF57had no effect on LANA expression in lytic induction.
The finding that substantial expression of PAN RNA is de-pendent on ORF57 was consistent with others’ reports thatKSHV ORF57 acts by itself or synergistically with ORF50 toincrease PAN RNA expression in transient transfection (10,14). The ORF57-enhanced PAN expression was also seen bytransient transfection when an ORF57-null KSHV genomewas used in a recent study (10). Unfortunately, there was nodirect comparison of the ORF57-null genome with the wtKSHV genome for PAN expression during lytic induction inthe reported study (10). By direct comparison of the wt andmutant genome, we concluded in this report that ORF57 pro-motes PAN RNA expression in the context of the virus ge-nome. However, we also found that a small amount of PANRNA can be spontaneously expressed or induced from thevirus genome in the absence of ORF57. Consistently, otherstudies also showed that the ORF57-null KSHV genome itselfcould be induced by infection with Ad50 as a source of ORF50to express PAN RNA to a certain level in the absence ofectopic overexpression of ORF57 (10). Together, these obser-vations suggest that ORF57 promotes PAN RNA expression,most likely at the posttranscriptional level.
Various analyses of gene structure and organization in theKSHV genome have shown that it contains multiple gene clus-ter regions. Several genes within a cluster region will use eithera common promoter or a common poly(A) signal for theirgene expression, and the resulting bicistronic or polycistronictranscripts overlap each other (11, 22, 25, 38, 44, 46, 52, 57).This organization of gene structure makes the genetic manip-ulation of the KSHV genome complex, and the results fromsuch studies could be misleading. It is certainly more difficult tocomplement a disrupted gene within a cluster using comple-mentation assays. For example, disruption of the LANA exon2 coding region by transposon insertion (53) will simulta-neously interrupt an intron within the vCyclin and vFLIP tran-scripts and affect their RNA splicing (38, 54); disruption of aK8.1 coding region by the same approach (27) will inevitably
destroy the 3� UTR of the ORF50 and K8� transcripts and willaffect the expression of all three genes, since they all share thesame poly(A) signal downstream of the K8.1 coding region (46,54, 57). Since the KSHV ORF57 coding region overlaps withthe 3� UTR of ORF56 (29), disruption of the ORF57 codingregion also inevitably damages the structure of ORF56 tran-scripts and simultaneously knocks out two genes. Accordingly,to complement the loss of two gene products, we cotransfectedBac36-�57 stable cells with ORF56 and ORF57 expressionvectors and effectively rescued the production of infectiousviruses from the mutant genome to a level nearly comparableto that from the Bac36-wt genome. Although it was puzzling,the partial restoration of infectious virus production in ourstudy by the ectopic expression of ORF57, but not ORF56alone, might imply that the transposon-mediated insertion didnot abrogate all ORF56 protein expression from Bac36-�57.Alternatively, a fraction of ORF56 transcripts from the Bac36-�57 genome might utilize any one of the three AATAAAhexamers from the inserted EZ::TN �KAN-2� transposonimmediately downstream for polyadenylation, since the virusprimase ORF56 is required for viral DNA synthesis. Anotherreport also indicates that the disruption of ORF57 by insertionof a Kanr cassette in the KSHV genome could be comple-mented by the ectopic expression of ORF57 alone in the pres-ence of ORF50 (10).
Reverse genetics is a powerful tool for dissecting the func-tions of viral genes. Since its development, the Bac36 systemhas been successfully used to define the functions of severalKSHV genes (10, 17, 27, 51, 53, 56). Because of the largegenome size, genomic rearrangements could occur in bothbacteria and mammalian cells. Adaptation of proper screeningprocedures, such as restriction analysis and Southern hybrid-ization, could ensure the selection of the correct clones. Theinduction of lytic replication in 293 cells is also at times notefficient. We observed that only few such induced cells (lessthan 1%) reach the late stage of infection (24 h after induc-tion), when the viral structural protein K8.1 is produced, andan extremely low number of infectious viruses were present inthe culture supernatant. Even after concentration by high-speed centrifugation, only �60 cells in every 100,000 cellsbecame infected in flow cytometry assays. While the selectionof individual high-producer cell clones could significantly en-hance the efficiency, the use of an entire cell population isoften necessary to achieve true representation, and this is re-quired to definitely delineate the function of a viral gene, as isthe case in this work. Alternative cell culture systems, such asVero cells that can support replication efficiency, should helpwider adaptation of the system in the investigation of thefunctions of KSHV genes in the context of the viral genome(10, 47). Nevertheless, determining the mechanism(s) that pre-vents the Bac36 genome from activating lytic infection wouldprovide further insights into the process of KSHV infection.
ACKNOWLEDGMENTS
This research was supported by the Intramural Research Program ofthe NIH, National Cancer Institute, Center for Cancer Research andDivision of Intramural Research, National Heart, Lung, and BloodInstitute.
We thank Jae Jung and Bala Chandran for providing anti-K8.1antibodies and Yan Yuan for the expression vector pORF50.
VOL. 81, 2007 DISRUPTION OF KSHV ORF57 REDUCES LYTIC GENE EXPRESSION 1069

REFERENCES
1. Ballestas, M. E., P. A. Chatis, and K. M. Kaye. 1999. Efficient persistence ofextrachromosomal KSHV DNA mediated by latency-associated nuclear an-tigen. Science 284:641–644.
2. Ballestas, M. E., and K. M. Kaye. 2001. Kaposi’s sarcoma-associated her-pesvirus latency-associated nuclear antigen 1 mediates episome persistencethrough cis-acting terminal repeat (TR) sequence and specifically binds TRDNA. J. Virol. 75:3250–3258.
3. Chan, S. R., and B. Chandran. 2000. Characterization of human herpesvirus8 ORF59 protein (PF-8) and mapping of the processivity and viral DNApolymerase-interacting domains. J. Virol. 74:10920–10929.
4. Chandran, B., C. Bloomer, S. R. Chan, L. Zhu, E. Goldstein, and R. Horvat.1998. Human herpesvirus-8 ORF K8.1 gene encodes immunogenic glyco-proteins generated by spliced transcripts. Virology 249:140–149.
5. Chang, Y., E. Cesarman, M. S. Pessin, F. Lee, J. Culpepper, D. M. Knowles,and P. S. Moore. 1994. Identification of herpesvirus-like DNA sequences inAIDS-associated Kaposi’s sarcoma. Science 266:1865–1869.
6. Chen, Y., M. Ciustea, and R. P. Ricciardi. 2005. Processivity factor of KSHVcontains a nuclear localization signal and binding domains for transportingviral DNA polymerase into the nucleus. Virology 340:183–191.
7. Delecluse, H. J., M. Kost, R. Feederle, L. Wilson, and W. Hammerschmidt.2001. Spontaneous activation of the lytic cycle in cells infected with a re-combinant Kaposi’s sarcoma-associated virus. J. Virol. 75:2921–2928.
8. Gottlicher, M., S. Minucci, P. Zhu, O. H. Kramer, A. Schimpf, S. Giavara,J. P. Sleeman, C. F. Lo, C. Nervi, P. G. Pelicci, and T. Heinzel. 2001. Valproicacid defines a novel class of HDAC inhibitors inducing differentiation oftransformed cells. EMBO J. 20:6969–6978.
9. Gupta, A. K., V. Ruvolo, C. Patterson, and S. Swaminathan. 2000. Thehuman herpesvirus 8 homolog of Epstein-Barr virus SM protein (KS-SM) isa posttranscriptional activator of gene expression. J. Virol. 74:1038–1044.
10. Han, Z., and S. Swaminathan. 2006. Kaposi’s sarcoma-associated herpesvi-rus lytic gene ORF57 is essential for infectious virion production. J. Virol.80:5251–5260.
11. Haque, M., V. Wang, D. A. Davis, Z. M. Zheng, and R. Yarchoan. 2006.Genetic organization and hypoxic activation of the Kaposi’s sarcoma-asso-ciated herpesvirus ORF34-37 gene cluster. J. Virol. 80:7037–7051.
12. Hiriart, E., L. Bardouillet, E. Manet, H. Gruffat, F. Penin, R. Montserret, G.Farjot, and A. Sergeant. 2003. A region of the Epstein-Barr virus (EBV)mRNA export factor EB2 containing an arginine-rich motif mediates directbinding to RNA. J. Biol. Chem. 278:37790–37798.
13. Hu, J., A. C. Garber, and R. Renne. 2002. The latency-associated nuclearantigen of Kaposi’s sarcoma-associated herpesvirus supports latent DNAreplication in dividing cells. J. Virol. 76:11677–11687.
14. Kirshner, J. R., D. M. Lukac, J. Chang, and D. Ganem. 2000. Kaposi’ssarcoma-associated herpesvirus open reading frame 57 encodes a posttran-scriptional regulator with multiple distinct activities. J. Virol. 74:3586–3597.
15. Klass, C. M., L. T. Krug, V. P. Pozharskaya, and M. K. Offermann. 2005.The targeting of primary effusion lymphoma cells for apoptosis by inducinglytic replication of human herpesvirus 8 while blocking virus production.Blood 105:4028–4034.
16. Koffa, M. D., J. B. Clements, E. Izaurralde, S. Wadd, S. A. Wilson, I. W.Mattaj, and S. Kuersten. 2001. Herpes simplex virus ICP27 protein providesviral mRNAs with access to the cellular mRNA export pathway. EMBO J.20:5769–5778.
17. Krishnan, H. H., N. Sharma-Walia, L. Zeng, S. J. Gao, and B. Chandran.2005. Envelope glycoprotein gB of Kaposi’s sarcoma-associated herpesvirusis essential for egress from infected cells. J. Virol. 79:10952–10967.
18. Krithivas, A., M. Fujimuro, M. Weidner, D. B. Young, and S. D. Hayward.2002. Protein interactions targeting the latency-associated nuclear antigen ofKaposi’s sarcoma-associated herpesvirus to cell chromosomes. J. Virol. 76:11596–11604.
19. Li, M., J. MacKey, S. C. Czajak, R. C. Desrosiers, A. A. Lackner, and J. U.Jung. 1999. Identification and characterization of Kaposi’s sarcoma-associ-ated herpesvirus K8.1 virion glycoprotein. J. Virol. 73:1341–1349.
20. Lin, C. L., H. Li, Y. Wang, F. X. Zhu, S. Kudchodkar, and Y. Yuan. 2003.Kaposi’s sarcoma-associated herpesvirus lytic origin (ori-Lyt)-dependentDNA replication: identification of the ori-Lyt and association of K8 bZipprotein with the origin. J. Virol. 77:5578–5588.
21. Lin, K., C. Y. Dai, and R. P. Ricciardi. 1998. Cloning and functional analysisof Kaposi’s sarcoma-associated herpesvirus DNA polymerase and its pro-cessivity factor. J. Virol. 72:6228–6232.
22. Lin, S. F., D. R. Robinson, G. Miller, and H. J. Kung. 1999. Kaposi’ssarcoma-associated herpesvirus encodes a bZIP protein with homology toBZLF1 of Epstein-Barr virus. J. Virol. 73:1909–1917.
23. Loscher, W. 1999. Valproate: a reappraisal of its pharmacodynamic proper-ties and mechanisms of action. Prog. Neurobiol. 58:31–59.
24. Lukac, D. M., L. Garibyan, J. R. Kirshner, D. Palmeri, and D. Ganem. 2001.DNA binding by Kaposi’s sarcoma-associated herpesvirus lytic switch pro-tein is necessary for transcriptional activation of two viral delayed earlypromoters. J. Virol. 75:6786–6799.
25. Lukac, D. M., J. R. Kirshner, and D. Ganem. 1999. Transcriptional activa-
tion by the product of open reading frame 50 of Kaposi’s sarcoma-associatedherpesvirus is required for lytic viral reactivation in B cells. J. Virol. 73:9348–9361.
26. Lukac, D. M., R. Renne, J. R. Kirshner, and D. Ganem. 1998. Reactivationof Kaposi’s sarcoma-associated herpesvirus infection from latency by expres-sion of the ORF 50 transactivator, a homolog of the EBV R protein. Virol-ogy 252:304–312.
27. Luna, R. E., F. Zhou, A. Baghian, V. Chouljenko, B. Forghani, S. J. Gao, andK. G. Kousoulas. 2004. Kaposi’s sarcoma-associated herpesvirus glycopro-tein K8.1 is dispensable for virus entry. J. Virol. 78:6389–6398.
28. Majerciak, V., K. Yamanegi, S. H. Nie, and Z. M. Zheng. 2006. Structuraland functional analyses of Kaposi’s sarcoma-associated herpesvirus (KSHV)ORF57 nuclear localization signals in living cells. J. Biol. Chem. 281:28365–28378.
29. Majerciak, V., K. Yamanegi, and Z. M. Zheng. 2006. Gene structure andexpression of Kaposi’s sarcoma-associated herpesvirus ORF56, ORF57,ORF58, and ORF59. J. Virol. 80:11968–11981.
30. Malik, P., D. J. Blackbourn, and J. B. Clements. 2004. The evolutionarilyconserved Kaposi’s sarcoma-associated herpesvirus ORF57 protein interactswith REF protein and acts as an RNA export factor. J. Biol. Chem. 279:33001–33011.
31. Miller, G., M. O. Rigsby, L. Heston, E. Grogan, R. Sun, C. Metroka, J. A.Levy, S. J. Gao, Y. Chang, and P. Moore. 1996. Antibodies to butyrate-inducible antigens of Kaposi’s sarcoma-associated herpesvirus in patientswith HIV-1 infection. N. Engl. J. Med. 334:1292–1297.
32. Minucci, S., and P. G. Pelicci. 2006. Histone deacetylase inhibitors and thepromise of epigenetic (and more) treatments for cancer. Nat. Rev. Cancer6:38–51.
33. Neipel, F., J. C. Albrecht, and B. Fleckenstein. 1997. Cell-homologous genesin the Kaposi’s sarcoma-associated rhadinovirus human herpesvirus 8: de-terminants of its pathogenicity? J. Virol. 71:4187–4192.
34. Peterson, G. M., and M. Naunton. 2005. Valproate: a simple chemical withso much to offer. J. Clin. Pharm. Ther. 30:417–421.
35. Raab, M. S., J. C. Albrecht, A. Birkmann, S. Yaguboglu, D. Lang, B. Fleck-enstein, and F. Neipel. 1998. The immunogenic glycoprotein gp35-37 ofhuman herpesvirus 8 is encoded by open reading frame K8.1. J. Virol.72:6725–6731.
36. Renne, R., W. Zhong, B. Herndier, M. McGrath, N. Abbey, D. Kedes, and D.Ganem. 1996. Lytic growth of Kaposi’s sarcoma-associated herpesvirus (hu-man herpesvirus 8) in culture. Nat. Med. 2:342–346.
37. Russo, J. J., R. A. Bohenzky, M. C. Chien, J. Chen, M. Yan, D. Maddalena,J. P. Parry, D. Peruzzi, I. S. Edelman, Y. Chang, and P. S. Moore. 1996.Nucleotide sequence of the Kaposi sarcoma-associated herpesvirus (HHV8).Proc. Natl. Acad. Sci. USA 93:14862–14867.
38. Sarid, R., J. S. Wiezorek, P. S. Moore, and Y. Chang. 1999. Characterizationand cell cycle regulation of the major Kaposi’s sarcoma-associated herpes-virus (human herpesvirus 8) latent genes and their promoter. J. Virol. 73:1438–1446.
39. Schulz, T. F. 2006. The pleiotropic effects of Kaposi’s sarcoma herpesvirus.J. Pathol. 208:187–198.
40. Shaw, R. N., J. L. Arbiser, and M. K. Offermann. 2000. Valproic acid induceshuman herpesvirus 8 lytic gene expression in BCBL-1 cells. AIDS 14:899–902.
41. Sun, R., S. F. Lin, L. Gradoville, and G. Miller. 1996. Polyadenylylatednuclear RNA encoded by Kaposi sarcoma-associated herpesvirus. Proc. Natl.Acad. Sci. USA 93:11883–11888.
42. Sun, R., S. F. Lin, L. Gradoville, Y. Yuan, F. Zhu, and G. Miller. 1998. Aviral gene that activates lytic cycle expression of Kaposi’s sarcoma-associatedherpesvirus. Proc. Natl. Acad. Sci. USA 95:10866–10871.
43. Swaminathan, S. 2005. Post-transcriptional gene regulation by gamma her-pesviruses. J. Cell Biochem. 95:698–711.
44. Talbot, S. J., R. A. Weiss, P. Kellam, and C. Boshoff. 1999. Transcriptionalanalysis of human herpesvirus-8 open reading frames 71, 72, 73, K14, and 74in a primary effusion lymphoma cell line. Virology 257:84–94.
45. Tang, S., K. Yamanegi, and Z. M. Zheng. 2004. Requirement of a 12-base-pair TATT-containing sequence and viral lytic DNA replication in activationof the Kaposi’s sarcoma-associated herpesvirus K8.1 late promoter. J. Virol.78:2609–2614.
46. Tang, S., and Z. M. Zheng. 2002. Kaposi’s sarcoma-associated herpesvirusK8 exon 3 contains three 5�-splice sites and harbors a K8.1 transcription startsite. J. Biol. Chem. 277:14547–14556.
47. Vieira, J., and P. M. O’Hearn. 2004. Use of the red fluorescent protein as amarker of Kaposi’s sarcoma-associated herpesvirus lytic gene expression.Virology 325:225–240.
48. Wang, S. E., F. Y. Wu, Y. Yu, and G. S. Hayward. 2003. CCAAT/enhancer-binding protein alpha is induced during the early stages of Kaposi’s sarcoma-associated herpesvirus (KSHV) lytic cycle reactivation and together with theKSHV replication and transcription activator (RTA) cooperatively stimu-lates the viral RTA, MTA, and PAN promoters. J. Virol. 77:9590–9612.
49. Williams, B. J., J. R. Boyne, D. J. Goodwin, L. Roaden, G. M. Hautbergue,S. A. Wilson, and A. Whitehouse. 2005. The prototype gamma-2 herpesvirus
1070 MAJERCIAK ET AL. J. VIROL.

nucleocytoplasmic shuttling protein, ORF 57, transports viral RNA throughthe cellular mRNA export pathway. Biochem. J. 387:295–308.
50. Wu, F. Y., J. H. Ahn, D. J. Alcendor, W. J. Jang, J. Xiao, S. D. Hayward, andG. S. Hayward. 2001. Origin-independent assembly of Kaposi’s sarcoma-associated herpesvirus DNA replication compartments in transient cotrans-fection assays and association with the ORF-K8 protein and cellular PML.J. Virol. 75:1487–1506.
51. Xu, Y., D. P. AuCoin, A. R. Huete, S. A. Cei, L. J. Hanson, and G. S. Pari.2005. A Kaposi’s sarcoma-associated herpesvirus/human herpesvirus 8ORF50 deletion mutant is defective for reactivation of latent virus and DNAreplication. J. Virol. 79:3479–3487.
52. Yamanegi, K., S. Tang, and Z. M. Zheng. 2005. Kaposi’s sarcoma-associatedherpesvirus K8 is derived from a spliced intermediate of K8 pre-mRNAand antagonizes K8� (K-bZIP) to induce p21 and p53 and blocks K8�-CDK2 interaction. J. Virol. 79:14207–14221.
53. Ye, F. C., F. C. Zhou, S. M. Yoo, J. P. Xie, P. J. Browning, and S. J. Gao.2004. Disruption of Kaposi’s sarcoma-associated herpesvirus latent nuclearantigen leads to abortive episome persistence. J. Virol. 78:11121–11129.
54. Zheng, Z. M. 2003. Split genes and their expression in Kaposi’s sarcoma-associated herpesvirus. Rev. Med. Virol. 13:173–184.
55. Zhong, W., H. Wang, B. Herndier, and D. Ganem. 1996. Restricted expres-sion of Kaposi sarcoma-associated herpesvirus (human herpesvirus 8) genesin Kaposi sarcoma. Proc. Natl. Acad. Sci. USA 93:6641–6646.
56. Zhou, F. C., Y. J. Zhang, J. H. Deng, X. P. Wang, H. Y. Pan, E. Hettler, andS. J. Gao. 2002. Efficient infection by a recombinant Kaposi’s sarcoma-associated herpesvirus cloned in a bacterial artificial chromosome: applica-tion for genetic analysis. J. Virol. 76:6185–6196.
57. Zhu, F. X., T. Cusano, and Y. Yuan. 1999. Identification of the immediate-early transcripts of Kaposi’s sarcoma-associated herpesvirus. J. Virol. 73:5556–5567.
VOL. 81, 2007 DISRUPTION OF KSHV ORF57 REDUCES LYTIC GENE EXPRESSION 1071





























