Embed Size (px)
Citation preview

PistillatamdashDuplications as a Mode for Floral Diversification in (Basal) Asterids
Tom Viaene Dries Vekemans Vivian F Irish Anke Geeraerts Suzy HuysmansSteven Janssens Erik Smets and Koen GeutenLaboratory of Plant Systematics Institute of Botany and Microbiology PO Box 2437 Leuven Belgium Department ofMolecular Cellular and Developmental Biology Yale University New Haven CT and Nationaal Herbarium NederlandUniversiteit Leiden Branch PO Box 9514 RA Leiden The Netherlands
Basal asterid families and to a lesser extent the asterids as a whole are characterized by a high variation in petal andstamen morphology Moreover the stamen number the adnation of stamens to petals and the degree of sympetaly varyconsiderably among basal asterid taxa The B group genes members of the APETALA3 (AP3) and PISTILLATA (PI)gene lineages have been shown to specify petal and stamen identities in several core eudicot species Duplicate genes inthese lineages have been shown in some cases to have diversified in their function for instance in Petunia a PI paralog isrequired for the fusion of stamens to the corolla tube illustrating that such genes belonging to this lineage are not justinvolved in specifying the identity of the stamens and petals but can also specify novel floral morphologies Thismotivated us to study the duplication history of class B genes throughout asterid lineages which comprise approximatelyone-third of all flowering plants The evolutionary history of the PI gene subfamily indicates that the two genes inPetunia result from an ancient duplication event coinciding with the origin of core asterids A second duplication eventoccurred before the speciation of basal asterid Ericales families These and other duplications in the PI lineage are notcorrelated with duplications in the AP3 lineage To understand the molecular evolution of the Ericales PI genes afterduplication we have described their expression patterns using reverse transcription polymerase chain reaction and in situhybridization reconstructed how selection shaped their protein sequences and tested their protein interaction specificitywith other class B proteins We find that after duplication PI paralogs have acquired multiple different expressionpatterns and negative selective pressure on their codons is relaxed whereas substitutions in sites putatively involved inproteinndashprotein interactions show positive selection allowing for a change in the interaction behavior of the PI paralogsafter duplication Together these observations suggest that the asterids have preferentially recruited PI duplicate genes todiverse and potentially novel roles in asterid flower development
Introduction
The MADS- intervening- keratin-like and C-terminaldomain subfamily of plant MADS-box genes (Theissenet al 1996) encodes DNA-binding transcription factors thatare proposed to have played a crucial role in the evolutionand diversification of the angiosperm flower (eg Beckerand Theissen 2003 Irish 2003 Hu and Saedler 2007) Thisgene family has been preferentially retained after genomeduplications (Liu and Wendel 2003 Blanc and Wolfe2004 Maere et al 2005) and various gene duplicationand loss events have created a phylogenetic pattern that in-deed reveals a large potential for gene diversificationthrough shifts in gene expression and protein interactionspecificity Within this family of MADS-box genes GLO-BOSAPISTILLATA (PI) belongs together with DEFI-CIENSAPETALA3 (AP3) to the class B lineage of floralorgan identity genes (Sommer et al 1990 Trobner et al1992) In a number of core eudicots genes in this lineagehave been shown to be responsible for proper developmentof petals and stamens in the second and third whorls of theflower (Schwarz-Sommer et al 1990 Coen andMeyerowitz1991) In Arabidopsis mutations in both PI and AP3 causehomeotic changes in the second and third whorls leading tothe conversion of petals into sepals and stamens into carpels(Jack et al 1992 Goto and Meyerowitz 1994) Moreoverthe protein products of AP3 and PI form obligate hetero-dimers that are required for nuclear localization andDNA binding (McGonigle et al 1996 Riechmann et al1996) and the presence of both gene products is necessaryto cause an ectopic phenotype (Jack et al 1994 Krizek and
Meyerowitz 1996) Furthermore the AP3ndashPI heterodimer islikely to act as part of distinct higher order protein com-plexes that uniquely specify each organ type (Honma andGoto 2001 Pelaz et al 2001 Theissen and Saedler 2001)
In this class B gene lineage several duplication eventshave been documented throughout angiosperm evolutionand functional divergence of some of these duplicates hasbeen demonstrated For instance a major duplication eventin the AP3-lineage at the base of the core eudicots resulted inthe TM6 and the euAP3 lineages (Kramer et al 1998) Thisduplication has allowed for the functional diversification ofthe resulting gene duplicates in members of the Solanaceae(de Martino et al 2006 Rijpkema et al 2006) Within theRanunculales a separate AP3-lineage duplication event hasoccurred (Kramer et al 1998) and functional analyses inPapaver somniferum have demonstrated subfunctionaliza-tion of the resulting AP3 paralogs in specifying petal andstamen identity (Drea et al 2007) Duplication of the PI lin-eage genes has also been identified in various clades withinthe flowering plants (Kramer et al 1998 Stellari et al 2004)In monocots divergence of expression patterns of PI paral-ogs was detected in Oryza sativa and Zea mays (Kang et al1998 Kyozuka et al 2000Munster et al 2001) Divergencein the roles of B-class genes has also been noted with respectto the evolution of novel floral morphologies For instancetwo PI paralogs FBP1 and PMADS2 have been identifiedin Petunia hybrida which act in a redundant fashionin petal and stamen development but FBP1 is also requiredfor the fusion of stamen filaments with the petal tube(Vandenbussche et al 2004) Furthermore in Aquilegiathe B-class genes appear to be required for the modificationof a floral organ the staminodium (Kramer et al 2007)These examples taken together illustrate diversifying evolu-tion in the class B gene lineage where numerous duplicationevents have created opportunities for functional divergence
Key words asterids PISTILLATA duplication
E-mail tomviaenebiokuleuvenbe koengeutenbiokuluvenbe
Mol Biol Evol 26(11)2627ndash2645 2009doi101093molbevmsp181Advance Access publication August 13 2009
The Author 2009 Published by Oxford University Press on behalf ofthe Society for Molecular Biology and Evolution All rights reservedFor permissions please e-mail journalspermissionsoxfordjournalsorg
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
However the extent to which this dynamic pattern ofclass B gene evolution has played a role in core eudicot flo-ral diversification is not well understood Core eudicotflowers in general are organized in a regular manner witha set number of organ types a clear distinction between se-pals and petals and fused carpels Nonetheless significantmorphological variation can be observed within this groupand in particular the asterids display a very wide range offloral diversity in shape form symmetry and color of thefloral organs The asterids contain some 80000 extantspecies and comprise a total of about one-third of all flower-ing plant species (Stevens 2001 onwards) During early as-terid diversification two extant groups branched off theorders Cornales and Ericales which together we denoteas the lsquolsquobasal asteridsrsquorsquo (Albach et al 2001 Bremer et al2002) These early diverging clades have retained severalfloral characters that are present in both rosids and asteridsincluding a high variation in the degree of sympetaly sta-men number adnation of stamens to corolla and ovulemorphology and anatomy (Stevens 2001 onwards Kubitzki2003) Moreover this group is of special interest becausesome floral morphological features that are constant in coreasterids are variable in basal asterids
Although a number of functional and phylogeneticanalyses have been carried out with several members ofthe asterid clade (eg Petunia Vandenbussche et al 2004Rijpkema et al 2006 Solanum de Martino et al 2006Antirrhinum Sommer et al 1990 Trobner et al 1992 Ger-bera Yu et al 1999 and Lamiales Aagaard et al 20052006) the evolutionary history of class B genes in asteridsis not yet well characterized In particular a well-describedphylogenetic framework for the PI lineage is still lackingFurthermore there has been virtually no sampling of thehighly variable basal asterid families except for Hydrangea(Cornales) (Kramer and Irish 2000)
We reconstructed the evolutionary history ofPISTILLATA- APETALA3- and TM6-like genes in asteridsby isolating new sequences from basal asterids and Aqui-foliales and combining these with previously characterizedclass B genes and expressed sequence tag (EST) data Ouranalyses suggest that during the evolution of both basaland core asterids old and recent PI gene duplications haveoccurred and that these are not correlated with duplicationsin the AP3 lineage Furthermore we have examined thepossibility of functional divergence of duplicate genes inbasal asterids by carrying out expression studies includingreverse transcription polymerase chain reaction (RTndashPCR)and in situ hybridization data as well as by assessing se-quence divergence and changes in interaction behaviorthrough yeast-2-hybrid analysis These data together withprevious analyses of class B gene functional capabilities incore asterids suggest that PI gene duplications have oc-curred multiple times during the evolution of the asteridsand may be associated with the recruitment of these novelduplicates to new floral functions in this clade
Material and MethodsSpecies Sampling
Table 1 lists the species used in this study Our ratio-nale was to sample densely across the orders Ericales and
Cornales because the largest variation in floral morphologyof asterids is found here For these orders species from 19 of23Ericales families and four of sevenCornales familieswerestudied The unsampled families were mainly monospecificor low in taxon number From seven larger families morethan one species was sampled In addition we added Ilexaquifolium from the core asterid order Aquifoliales Thenewly generated sequences are indicated in bold in table 1To supplement our sampling with species from core asteridswe performedGenbank Blast searches in both the nucleotideand EST databases In this way sequences from 27 otherasterid species were added of which the EST sequencesare marked with an asterisk in table 1 Naming of sequencesexplicitly follows the priority rule in which newmembers ofa gene lineage are named after the first discoveredmember ofthis cladeDEFICIENS (Sommer et al 1990) andGLOBOSA(Trobner et al 1992) fromAntirrhinummajus and TM6 fromSolanum lycopersicon (Pnueli et al 1991)
Cloning of Class B MADS-Box Genes
Floral buds were frozen in liquid nitrogen and stored at80 C Total RNA was isolated from these using the In-visorb Spin Plant RNA kit (Invitek Berlin Germany) Fromthis total RNA mRNA was isolated using the NucleotrapmRNA kit (Macherey-Nagel Duren DE) and reverse tran-scribed into cDNA using avian myeloblastosis virus (AMV)reverse transcriptase(PromegaMadisonWI)andthe includedoligo-dT primer Our initial strategy was applying 3-rapidamplification of CDNA by PCR (Frohmann et al 1988) usingprimers (MADSBFCT 5-GGGGTACCAAYMGICAR-GTIACITAYTCIAAGMGIMG-3 RQVT 5-CGRCARG-TGACSTTCTSCAARCG-3 oligodT 5-CCGGATCCT-CTAGAGCGGCCGC(T)17-3) and polymerase chain reac-tion (PCR)-programs from the literature (Kramer et al 1998Winter et al 1999) After a provisional alignment of basal as-terid PISTILLATA-sequences the primer from Kramer et al(1998) was modified (5-GGGGTACCAAYMGGCARGT-GACGTAYTCGAAGMGGMG-3) Based on the samealignment new forward and reverse primers (5-GYTYTGG-GATGCTAARCATGAG-3 3-AGMNAGNACRTYTA-AWCCRAC-5) were designed to amplify a second copy ofPI in Galax urceolata Using this forward primer in com-bination with the oligodT as reverse primer PI homologuesfromMaesaargenteaandAcanthogiliagloriosawere isolatedA second copy of Primula denticulata was isolated usinga primer based on a short PI sequence of Primula vulgarisin Genbank (DQ381443) After creating an alignment ofAP3-like genes a reverse primer on the C-terminus (3-YCCRTYRTCVACYARCCCRTARTGWGGATC) was de-signed and combined with the MADSBFCT forward primer(Krameretal1998)toamplifyAP3-likegenesmorespecificallyAll PCR amplifications were carried out using Taq DNAPolymerase (Invitrogen Carlsbad CA) PCR products weregel-purified with the Nucleospin extract 2 kit (Macherey-Nagel) and cloned into the pGEM-T vector (Promega) Aftertransformation between 50 and 100white cloneswere checkedforinsertsinaPCRreactionusingthesameprimersandprogramPlasmidDNA for selected cloneswas extractedwith theNucle-ospinPlasmidkit (Macherey-Nagel) Inaneffort to sampleclassB members exhaustively for each species between 20 and 40
2628 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Table 1List of Class B Homologues Included in This Study
Order Family Species PI AP3 TM6
Cornales Cornaceae Alangium platinifolium ApGLO ApDEF ApTM6Nyssaceae Nyssa sylvatica NsGLO1 NsGLO2 NsDEF NsTM6
Davidia involucrata DiGLO1 DiGLO2 DiTM6Loasaceae Loasa vulcanica LvGLO LvTM6Hydrangeaceae Hydrangea macrophylla HmPI HmAP3 HmTM6
Philadelphus pubescens PhpTM6Ericales Balsaminaceae Impatiens hawkeri IhGLO IhDEF1
IhDEF2Impatiens glandulifera IgGLO1 IgGLO2
Marcgraviaceae Marcgravia umbellata MuGLO MuDEFLecythidaceae Gustavia brasiliensis GbGLO1 GbGLO2 GbTM6
Napoleonaea vogelii NvGLO NvDEFPolemoniaceae Phlox paniculata PpGLO PpDEF PpTM6
Acanthogilia gloriosa AgGLOIpomopsis aggregata IaGLO
Sapotaceae Synsepalum dulcificum SydGLO SydDEFDiapensiaceae Galax urceolata GuGLO1 GuGLO2 GuDEF GuTM6Styracaceae Halesia diptera HdGLO1 HdGLO2 HdDEF
Styrax japonicus SjGLO SjDEF SjTM6Roridulaceae Roridula gorgonias RgGLOActinidiaceae Saurauia zahlbruckneri SzGLO1 SzGLO2
SzGLO3SzTM6
Actinidia chinensis AcGLO1 AcGLO2Clethraceae Clethra tomentosa CtGLO CtDEF CtTM6Theaceae Stewartia pseudocamellia SpGLO1 SpGLO2
SpGLO3SpDEF
Camellia japonica CjGLO1 CjGLO2 CjDEF CjTM6Symplocaceae Symplocos chinensis ScGLO1 ScGLO2 ScTM6Sarraceniaceae Sarracenia drummondii SdGLO SdDEF SdTM6Ericaceae Enkianthus campanulatus EcGLO
Erica hiemalis EhGLO1 EhGLO2 EhDEFEbenaceae Diospyros digyna DdGLO DdDEFPrimulaceae Primula denticulata PdGLO1 PdGLO2 PdDEFMyrsinaceae Cyclamen persicum CpGLO CpDEF
Hymenandra walichii HwGLO HwDEF HwTM6Theophrastaceae Jacquinia auriantaca JaGLO JaDEF JaTM6
Clavija latifolia ClGLO ClDEFMaesaceae Maesa argentea MaGLO MaDEF
Aquifoliales Aquifoliaceae Ilex aquifolium IaGLO1 IaGLO2 IxaAP3 IxaTM6Asterales Asteraceae Gerbera hybrida GGLO1 GDEF2 GDEF1
Lactuca sativa LsGLOTaraxacum officinale ToGLOCichorium intybus CiGLOSenecio vulgaris SvGLOHelianthus annuus HaPI HaAP3 HaTM6Centaurea solstitialis CsGLOHieracium piloselloides HpDEF1
HpDEF2Gentianales Rubiaceae Hedyotis terminalis HtGLO
Apocynaceae Catharanthus roseus CrGLOGentianaceae Eustoma grandiflorum EgGLO1 EgDEF
Solanales Solanaceae Petunia hybrida FBP1 PMADS2 PhDEF PhTM6Solanum lycopersicon SlGLO1 SlGLO2 LeAP3Ipomoea nil InGLONicotiana tabacum NtGLOCestrum elegans CeDEF CeTM6Nicotiana benthamiana NtTM6Solanum pseudolulo SopDEF SpTM6Brunfelsia uniflora BuDEF BuTM6
Lamiales Scrophulariaceae Mimulus guttatus MgGLOMimulus ringens MrDEFA
MrDEFBVerbenaceae Verbena hybrida VhGLOPlantaginaceae Antirrhinum majus GLO DEF
Misopates orontium MoGLO MoDEFChelone glabra CgDEF
Oleaceae Syringa vulgaris SvPI SvAP3Lamiaceae Salvia coccinea ScDEFA
ScDEFB
PISTILLATAmdashDuplications in Asterids 2629
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
plasmid inserts were sequenced using T7 and SP6 universalprimers using the BigDye Terminator 11 kit (Applied Bio-systems Foster City CA) on an Applied Biosystems 310 se-quencer or the plasmids were sent for sequencing (MacroGenInc Seoul South Korea) New sequences were deposited inGenbank (Accession numbers GQ141105ndashGQ141187)
Identification and Sequence Analysis
Obtained nucleotide sequences were translated intothe corresponding amino acid sequence and manuallyaligned using MacClade 4 (Maddison RP and MaddisonWP 2003) together with previously cloned and character-ized class B sequences from Genbank (table 1) After align-ment nucleotide sequences were analyzed with PAUP4b10 (Swofford 2002) MrBayes 312 (Huelsenbeck andRonquist 2001) and PHYML (Guindon and Gascuel2003) (PI matrix only) Paup 4b10 (Swofford 2002)was used for parsimony bootstrap analysis A MaximumParsimony (MP) heuristic search was conducted using1000 random addition sequences with tree bisection andreconnection branch swapping and saving of multiple par-simonous trees (MulTrees on) Branch support was calcu-lated by nonparametric bootstrap analysis on 1000pseudoreplicate data sets (Felsenstein 1985) Parametersfor the Bayesian analysis and maximum likelihood (ML)analysis were estimated using Modeltest 306 (Posadaand Crandall 2001) Modeltest selected the GTR (generaltime reversible) thorn I thorn G substitution model using theAkaike Information criterion MrBayes was run for 5 mil-lion generations where for every 100 generations one treewas saved For both matrices the search reached the sta-tionary phase around 50000 generations This numberwas considered the lsquolsquoburn-in periodrsquorsquo and was excludedwhen the consensus phylogeny was constructed PHYML(Guindon and Gascuel 2003) was used for ML inference ofthe PImatrix Confidence in the clades was estimated by theapproximate likelihood ratio test (aLRT) method from Ani-simova and Gascuel (2006) and traditional bootstrap anal-
ysis with 100 replicates For the aLRT tests we used boththe v2 statistic and the more conservative shimodeiva-Hasqawa (SH)-like test as branch support measures To fur-ther look into the supposed ancient PI duplication duringthe evolution of the Ericales clade we additionally con-structed a matrix with only the duplicated PI genes resultingfrom this duplication Cornales PI genes were used as out-groups as the reconstructed PI phylogeny gives us noclear answers on the exact timing of duplication A similaranalysis strategy was used but Modeltest selected theHasegawandashKishinondashYano (HKY) thorn G substitution modelfor the Bayesian and ML analyses
Semiquantitative RT-PCR
RNA was extracted from each organ type separatelyusing the Invisorb Spin Plant RNA kit (Invitek) and eachRNA sample was DNase treated using TURBO DNA-free(Ambion Austin TX) Total RNA was reverse transcribedinto cDNA using AMV reverse transcriptase (Promega) andthe included oligo-dT primer and random primers Gene-specific primers are listed in supplementary table S1 Sup-plementary Material online Expression of AP3 in Gustaviabrasiliensis and of TM6 in Cyclamen persicum Primulaacaulis Stewartia pseudocamellia andHalesia diptera wereexaminedwith primers fromAP3-like sequences from closelyrelated species Ribosomal 18S RNA was used as a control
In Situ Hybridization
Antisense probes were in vitro transcribed using T7RNApolymerase (NewEnglandBiolabs IpswichAustralia)in thepresence of digoxigenin-labeled uracil-5-triphosphate(Roche Basel Switzerland) from PCR-amplified templatesthat included a T7 promoter Tissues were fixed in 4 para-formaldehydeparaffinembeddedandsectionedat8lmSec-tions were mounted on Probe-On-Plus slides (FisherScientific Pittsburgh PA) Prehybridization hybridizationand detection essentially followed Carr and Irish (1997)
Table 1Continued
Order Family Species PI AP3 TM6
Ranunculales Ranunculaceae Aquilegia vulgaris AqvPITrautvetteria carolinensis TrcAP3
Vitales Vitaceae Vitis vinifera VvDEF VvTM6Saxifragales Grossulariacaea Ribes sanguineum
Saxifragaceae Saxifraga careyana SxcAP3 SxcTM6Malpighiales Salicaceae Populus deltoides
Populus tomentosa PtTM6Myrtales Myrtaceae Melaleuca quinquenerviaRosales Rosaceae Malus domestica MdTM6
Rosa rugosa RrDEF RrTM6Cucurbitales Cucurbitaceae Cucumis sativusFagales Betulaceae Betula pendula
Juglandaceae Juglans regia JrDEFTrochodendrales Trochodendraceae Trochodendron aralioides TraPI1 TroAP3Magnoliales Annonaceae Asimina longifolia AlAP3Dilleniales Dilleniaceae Dillenia indica DiiAP3 DiiTM6Asparagales Orchidaceae Phalaenopsis hybrid PhahAP3
Brassicaceae Arabidopsis thaliana AP3Brassica juncea BjAP3
Newly generated asterid sequences are marked in bold EST data are highlighted with an asterisk
2630 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Hybridizations in Sarracenia rubra and Stewartia rostratawere carried out using heterologous probes of the closely re-lated speciesSarracenia drummondii andS pseudocamellia
Codon Substitution Models
Omega (x or dNdS) is a measure of the amount of nat-ural selection acting on the protein To test whether positiveselection acted after the duplication event at the base of theEricales the previously obtained Bayesian topology of PIgenes was pruned in TreeEdit (indicated in fig 1) and thematching data sets were analyzed using PAML version314 (Yang 1997) We used two data matrices to investigate
selective pressure a matrix in which only the duplicatedgenes that originated after the duplication event at the baseof the Ericales are present (hereafter matrix A) and a secondmatrix in which the paralogous copies from species whohave lost either one of both paralogs are incorporated inmatrix A (hereafter matrix B) The log-likelihood of the se-quence alignments was calculated using different codonsubstitution models and likelihood ratio tests were usedto detect significant differences in fit between nested mod-els Branch models (Yang 1998 Yang and Nielsen 1998)were used to test for episodic adaptive evolution after du-plication either in all lineages combined or in paralogouslineages separately To test for the presence of positive
FIG 1mdashEvolutionary reconstruction of PI gene phylogeny in asterids as inferred from ML and Bayesian analysis Bayesian posterior probabilitiesand nonparametric bootstrap support values are indicated above branches SH-like and X2-like supports are shown below branches Gray circle 5
inferred duplication event Data sets used for assessments of selection pressures are indicated by stars black star 5 sequences included in matrix Awhite star 5 extra sequences added to matrix A to form Matrix B Marcgravia umbellata and Diospyros digyna PI sequences are underlined
PISTILLATAmdashDuplications in Asterids 2631
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
selection on sequence sites site models were applied thathold x constant among all branches but allow x to varyamong codons Site models include M1a M2a M7 andM8 (Nielsen and Yang 1998 Wong et al 2004 Yang et al2005) Positive selection is inferred when M2a and M8 arerespectively significantly better than M1a and M7 Branch-sitemodels allow thex ratio to vary amongboth sites and lin-eages (Zhang et al 2005) and can be used to detect positivelyselected sites in specific lineages within the tree We usedthe modified branch-site model A and compared this modelwith the nested site model M1a (Test 1) and the null modelA (Test 2) to test for positive selection (Zhang et al 2005)
Yeast Two-Hybrid Assays
The intervening- keratin-like and C-terminal domaindomains of the two PI (CjGLO1 and CjGLO2) the AP3(CjDEF) and TM6 genes (CjTM6) from Camellia japonicawere fusedwith theGAL4activationdomain in thepGAD424vector and the GAL4 DNA-binding domain in the pGBT9vector (ClontechMountainView CA) Each possible vectorcombination was transformed in yeast and two colonies pertransformation were used for B-galactosidase liquid assaysusing the protocol available at httpwwwfhcrcorgsciencelabsgottschlingyeast20ProtocolsBgalhtml
Results
In total 83 new B-lineage sequences were amplifiedfrom asterid species Forty-seven new PI and 36 new AP3-like sequences were identified from 31 and 25 asterid speciesrespectively (table 1) All but one of these species belong tothe basal asterid orders Cornales and Ericales In addition PIhomologues were amplified from I aquifolium (Aquifoliales)
Two Ancient and Frequent Recent Duplications HaveOccurred in the Asterid PI Lineage
For the phylogenetic analyses of the PI lineage wecombined the newly identified PI genes with previouslyidentified asterid PI sequences and EST data from Genbankto generate one data set Basal eudicot sequences were usedas outgroups for this data set and AqvPI (Aquilegia Ranun-culales) was used to root the PI tree A nucleotide alignmentof 74 taxa was analyzed using MP ML and Bayesian anal-yses The results are summarized in figure 1
The Bayesian topology which has relatively high pos-terior probabilities for a majority of nodes did not conflictwith the most likely tree from the ML analysis The MPanalysis however did not resolve the relationships betweenthe major clades and high bootstrap values were only ob-tained for nodes at lower taxonomical levels Similar to thebootstrap values from the MP analysis bootstrap supportfor the ML tree is low and we chose to only indicate thebootstrap values from the MP analysis on the phylogenyHowever to provide an additional measure of support ina likelihood framework we show SH-like and v2-like aLRTtest values These provide additional support for the Bayes-ian inferred relationships
The inferred PI phylogeny (fig 1) indicates that thisgene lineage probably underwent two ancient duplication
events during asterid evolution In both instances the twocopies were retained in some extant species lineageswhereas in other taxa one copy appears to have been lostor further duplicated An initial ancient duplication probablyoccurred early in the evolution of the core asterids becausethe Bayesian analysis supports two paralogous cladesa clade containing Gentianales Solanales and Asteralessequences (BPP 95) and one with Solanales and Lamialessequences (BPP 100) The latter is also strongly supportedbyMPbootstrapping (86) and aLRT testing (10099) Givenour current knowledge of asterid evolution (Albach et al2001 Bremer et al 2002) this suggests an ancient duplica-tion event before the origin of the two major core asteridclades (euasterids I and euasterids II) (Bayesian analysis)or at least before the origin of the euasterid II clade compris-ing Solanales Lamiales and Gentianales (MP analysis andaLRT testing) This duplication event was then followed byindependent losses in each paralogous lineageAlthoughourresults allow us to infer such a scenario no Garryales Dip-sacales or Apiales PI representatives are currently availablethat would further support such a hypothesis In addition thePI sequences of Aquifoliales an order situated at the base ofthe euasterid II clade occupy a nonsupported position at thebase of basal asterid PI sequences Together this suggeststhat an even denser sampling of asterid sequences would bedesirable to determine the exact moment of the PI duplica-tion in the core asterid lineage with even more confidence
The ML tree further suggests a second ancient dupli-cation event consistent with an origin early in the radiationof the order Ericales The PI sequences from Ericales rep-resentatives that are strongly supported by Bayesian support(BPP 96) and aLRT support (9999) are placed into two sep-arate clades A first clade is supported by a BPP value of 90and a v2-value of 92 The weakly supported second clade(BPP 67 aLRT 7088) consists of two strongly supportedclades (BPP 96 aLRT 9499 and BPP 90 aLRT 8999)where one (BPP 96) consists of the proposed basal lineagesof the Ericales the balsaminoıd clade and Lecythidaceae(Geuten et al 2004 Schonenberger et al 2005) The pres-ence of three strongly supported clades within the Ericalessuggests that the duplication in Ericales could have occurredeither at the base of the Ericales (as the ML phylogenyshows) or after the divergence of the basal lineages (balsami-noids and Lecythidaceae)Galax urceolata (Diapensiaceae)Hdiptera (Styracaceae)Saurauia zahlbruckneri (Actinidia-ceae) S pseudocamellia and C japonica (Theaceae) andSymplocos chinensis (Symplocaceae) all have multiple PIcopies belonging to two strongly supported clades due to thisancient duplication event Again this ancestral duplication isfollowed by different gene fateswith independent losses andduplications in each paralogous lineage Although furthersampling could lead to alternative explanations for the pro-posed multiple loss events there is some support for the hy-pothesis of independent gene loss in eachparalogous lineageSouthern blot analyses of Marcgravia umbellata (Geutenet al 2006) and Diospyros digyna (data not shown) both in-dicate that there is likely only one PI gene present in each ofthese species (fig 1 underlined) This observation supportsthe hypothesis of multiple PI gene losses within the Ericales
Because we wanted to explore the effect of omittingtaxa with single-copy genes of which the paralogous was
2632 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
putatively lost after duplication we constructed a secondmatrix with only duplicated PI genes due to the lsquolsquoancientrsquorsquoduplication from G urceolata (Diapensiaceae) H diptera(Styracaceae) S zahlbruckneri (Actinidiaceae) S pseudo-camellia and C japonica (Theaceae) and S chinensis(Symplocaceae) The MP analysis resulted in just one mostparsimonious tree that is identical to the tree with the high-est likelihood As in the previous analysis the same twoparalogous clades were identified In this case both BPPvalues (100 and 97) and ML bootstrap values (57 and53) were higher in this case supporting a duplication eventpresumably at least before the divergence of these familiesand after the divergence of the primuloid clade in Ericalesphylogeny (Schonenberger et al 2006)
In addition to these two ancient duplication events weidentified several other more recent duplication events inasterid evolution From I aquifolium (Aquifoliaceae) weamplified two PI copies but more extensive sampling ofAquifoliales representatives should reveal the timing of thisduplication more accurately In Cornales duplicate genesare found in both Davidia involucrata and Nyssa sylvatica(Nyssaceae) suggesting that the duplication that resulted inthese copies occurred in the lineage leading to NyssaceaeIn the clade of Ericales representatives there are seven spe-cies with more than one PI copy which appear to be morerecent than the previously described duplication event atthe base of the Ericales order For example we isolatedtwo PI homologues from Impatiens glandulifera (IgGLO1and IgGLO2 Balsaminaceae) A previously characterizedPI gene from Impatiens hawkeri (IhGLO Geuten et al2006) groups with IgGLO2 but not with the PI gene fromM umbellata (MuGLO Marcgraviaceae) suggesting thatthis duplication occurred at the base of the BalsaminaceaeSimilarly we isolated twoPI genes fromG brasiliensis (Le-cythidaceae) and the phylogeny indicates that one copy ismore related to the PI gene from another Lecythidaceae rep-resentativeNapoleonaea vogeliiBecauseGustavia is basalin the phylogeny of Lecythidaceae (Mori et al 2007) the du-plication leading to paralogous copies inGustavia probablyoccurred at the origin of the Lecythidaceae or earlier
We also identified two PI copies in Erica hiemalis(Ericaceae) and Actinidia chinensis (Actinidiaceae) In bothS pseudocamellia (Theaceae) and S zahlbruckneri (Acti-nidiaceae) a third PI copy is present presumably as a resultof a more recent duplication in one of their paralogs As thesequence similarity of the paralogs in these four species ishigh (91ndash98) it is hard to decide whether these genes areat the same genetic locus or at different loci We suggestthese genes to be paralogous copies with a very recent or-igin and not shared by any other species in our samplingFinally two PI genes were identified in P denticulata (Pri-mulaceae) Although sequence similarity among thesegenes is significantly lower (82) this duplication is notshared by any other PI genes in our analyses indicating thatthis duplication is specific to the Primulaceae
Duplications in the AP3 and TM6 Lineages Are Few andNot Correlated with PI Duplications
We also carried out phylogenetic analyses of asteridAP3 sequences using a number of newly identified AP3
sequences as well as previously characterized genes andEST sequences from Genbank (table 1) The phylogenywas inferred from a nucleotide alignment of genes from85 taxa which was analyzed using both MP and Bayesianmethods (fig 2) In the Bayesian analysis AlAP3 was usedto root the AP3-like tree Similar to previous analyses(Kramer and Irish 1999 Kramer et al 2006) our Bayesiananalysis recovered two highly supported clades A firstclade contains the euAP3 homologues (BPP 98) whereasa second clade contains the TM6 homologues (BPP 100)(fig 2) The duplication event that gave rise to these two eu-dicot AP3 gene clades is placed near the base of the core eu-dicots and likely occurred coincident with the origin ofTrochodendraceae and Buxaceae (Kramer et al 2006)Within core asterids our phylogeny gave no indicationsofmajor duplication events but onlyfiveof eight core asteridorderswere sampled Similarly in basal asterids duplicationevents have seemingly not played an important role duringAP3 evolution because the observed gene phylogeny did notconflict with the species phylogeny (Schonenberger et al2006) In several cases thoughparalogousAP3genesofveryrecent origin were identified in four species one from Eri-cales (I hawkeri Geuten et al 2006) and three from core as-terids (Hieracium piloselloides Mimulus cultstatus andSalvia coccinea) (Aagaard et al 2005) As for thePI phylog-eny the bootstrap values in the MP analyses were low andthese analyses resulted in a poorly resolved tree
After the basal eudicot and rosid representativesbranch off in the euAP3 clade (BPP 98) the Cornales rep-resentatives are sister to two large clades The first cladecontains the newly sampled Ericales representatives(BPP 81) whereas the second contains all core asterid se-quences (BPP 54) The unexpected association of Brassica-ceae representatives (AP3 BjAP3) with the euAP3 genesfrom Asterales was also present in previous analyses andprobably illustrates the need for even denser sampling(de Martino et al 2006 Kramer et al 2006) Within theEricales euAP3 clade (BPP 81) most relationships arepoorly resolved However a strongly supported primuloid(BPP 100) ericoid (BPP 91) and balsaminoid clade (BPP100) can be observed (Schonenberger et al 2005) Withinthe TM6-clade Ericales representatives are supported witha BPP of 99 but overall support within this clade is lowHowever we did not recover any paralogous copies of TM6genes in the species sampled suggesting that duplicationevents in this clade either do not occur readily or are se-lected against We also observed an unexpected associationof Asterales TM6 representatives with rosids (BPP 71) asreported in previous analyses (de Martino et al 2006Kramer et al 2006) The TM6 genes within the Solanalesare strongly supported as a clade (BPP 100)
RT-PCR Reveals Divergence of Expression PatternsAfter Duplication
To investigate whether gene duplicates have divergedin expression patterns or have seemingly remained underthe same regulatory control we used RT-PCR to monitorgene expression levels in mature dissected floral organs(fig 3) We limited our survey to Ericales species thatcontain representatives of PI genes arising from both the
PISTILLATAmdashDuplications in Asterids 2633
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
ancient and recent duplications We find that the expressionpattern of PI paralogs has diverged considerably and fairlyrapidly The PI protein is known to require an AP3-like in-teraction partner for its in vivo function in a number of coreeudicots (Jack et al 1992 Trobner et al 1992 Goto andMeyerowitz 1994) We therefore also examined the expres-sion patterns of TM6 and euAP3 genes in the same species
We compared Ericales expression patterns with the as-terid speciesPetunia hybrida (Solanaceae) BothPI genesin Petunia show a very similar expression pattern (Vanden-bussche et al 2004) Expression of these twoPI homologues(FBP1 and PMADS2) is mainly confined to the second andthirdwhorls (Angenent et al 1992Van derKrol et al 1993)Expression of PhDEF the AP3 homologue was mainly
FIG 2mdashEvolutionary reconstruction of APETALA3-like gene phylogeny in asterids as inferred from Bayesian analysis Bayesian posteriorprobabilities are indicated above branches nonparametric bootstrap support values are shown below branches Gray circle5 inferred duplication event
2634 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
confined to petals and stamens with low expression in firstand fourth whorl organs PhTM6 has a different expressionprofilewithstrongexpressioninstamensandcarpelswhereasexpression in sepals and petals is low (Vandenbussche et al2004) Similar expression patterns to Petunia of PI AP3 and
TM6 were detected in S lycopersicon another core asteridSolanaceae species (de Martino et al 2006)
Multiple copies of PI have been retained in the ge-nomes of S pseudocamellia C japonica and H dipteraafter the major duplication event at the base of the Ericales
FIG 3mdashRT-PCR expression data for B-class genes from selected Ericales representatives Sep 5 sepals pet 5 petals pet up 5 upper part ofpetals fus 5 fused part of petals and stamens sta 5 stamens sta up 5 upper part of stamens sta fus 5 fused part of stamens gyn 5 gynoecium andflo 5 flower
PISTILLATAmdashDuplications in Asterids 2635
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Both copies in each of these species appear to have divergedin their expression patterns after duplication (fig 3AndashC)Although HdGLO2 is expressed in the petals and stamensof H diptera the other copy (HdGLO1) is mainly ex-pressed in stamens and the gynoecium and absent in petalsIn comparison the TM6 and euAP3 lineage genes show un-expected expression patterns with strong expression ofHdTM6 in all disected organs but HdDEF expression beingmainly restricted to petals and carpels InC japonica one PIparalog (CjGLO2) is expressed throughout sepals petalsand stamens whereas expression of the second paralog(CjGLO1) appeared to be restricted to the nonfused partsof petals stamens and carpels Although both CjDEF andCjTM6 expression are detected in all organs strong expres-sion ofCjDEF is seenmainly in the sepals and petalsCjTM6expression is strongest in all stamen parts and in carpelsS pseudocamellia also displays differences in the expressionof its two PI copies One paralog (SpGLO2) is expressed inpetals stamens carpels and expression of the second paral-og (SpGLO1) is more restricted to the fused part of petals andstamens and upper part of petals The AP3 homologueSpDEF is strongly expressed in all stamen parts and carpelswhereas strong expression of SpTM6 is detected in sepals allstamen parts and carpels
In summary in these three cases there is one PI copywith a broad expression pattern in the flower (even in sepalsand carpels) whereas expression of the other PI copy ismore restricted to specific parts of petals and stamensThese expression patterns differ from those of PI lineagegenes in core asterid species (Vandenbussche et al2004 de Martino et al 2006) where PI expression (alsoin a duplicated configuration) is mainly restricted to the sec-ond and third whorl organs petals and stamens and nostrong expression in the first and fourth whorl organs is de-tected Interestingly the divergence in expression patternsin these species does not follow the gene phylogeny Theparalogous copies with a general expression pattern are notgrouped in one clade of PI genes in Ericales but rather arefound in two clades This suggests that the PI expressiondomains were not fixed immediately after duplication in-dicating that subfunctionalization in spatial expression doesnot explain retention of the duplicates Alternatively ifregulatory subfunctionalization was the main reason forretention evidence for this has been obscured throughtime Differences in expression between AP3 and TM6in C japonica seem to be very similar to members ofthe same lineages in the core asterid species Expressionof AP3-like genes in H diptera and S pseudocamelliahowever differs from those of Camellia with strongAP3 expression in the carpels of Stewartia and Halesiaand weak expression of AP3 in stamens of Halesia In eachof these species expression of one or both AP3-like genes isdetected in the sepals
Sarracenia drummondii (fig 3D) Styrax japonicus(fig 3E) C persicum (fig 3F) and Phlox paniculata(fig 3G) maintained probably only one of the paralogousPI copies in their genome after the ancient Ericales dupli-cation Moreover Sarracenia and Styrax maintained a dif-ferent copy (fig 1 BPP 68) than did Cyclamen and Phlox(fig 1 BPP89) We investigated the expression patterns ofPI genes in these species to monitor the possible divergence
between the two paralogous PI clades after duplication Ineach of these four species PI is expressed in petals and sta-mens similar to the expression patterns of PI in core aster-ids in previous studies (Vandenbussche et al 2004 deMartino et al 2006) In both Sarracenia (SdGLO) andPhlox (PpGLO) there is some expression detected in thecarpels whereas SjGLO (Styrax) is also expressed in sepalsNo clear divergence in expression of PI from the two dif-ferent clades (fig 1 BPP68 and BPP89) is detected in thesecond and third whorl organs The expression patterns ofAP3 and TM6 lineage genes are different between these fourspecies Although in S drummondii both AP3 and TM6 areexpressed in all four floral whorls the expression in Styraxis very similar to that of TM6 and AP3 in core asterid spe-cies with in addition to expression in petals and stamensexpression of TM6 in carpels and AP3 in sepals In Cycla-men TM6 expression is absent in the upper part of the sta-mens and strong expression of AP3 is detected in thecarpels In Phlox however TM6 expression is absent fromthe carpels
The multiple PI copies in G brasiliensis (Lecythida-ceae) and P acaulis (Primulaceae) are the result of a recentduplication probably at the base of these two families(fig 1) We looked at the expression pattern of each of theseduplicated genes and of the AP3-like genes in these speciesIn G brasiliensis GbGLO2 is expressed in petals and sta-mens whereas GbGLO1 is mainly expressed in the stami-nal parts and less in the petals (fig 3H) Expression of AP3and TM6 seems to be lsquolsquoinversersquorsquo compared with the ex-pected asterid expression with AP3 expression mainly re-stricted to the stamens and strong TM6 expression in thepetals No AP3-like expression is detected in the carpelsIn P acaulis there are differences between the expressionpatterns of both PI copies (fig 3I) PaGLO2 is expressed inall organs tested whereas the expression of PaGLO1 ismainly restricted to the sepals and petals Both AP3 andTM6 are strongly expressed in sepals and as inC persicumTM6-expression is weak in the staminal parts Even afterthese recent duplication events divergence in the expres-sion domain of the two PI copies is detected
In contrast to the duplicated PI genes of Petunia(Vandenbussche et al 2004) the expression domains ofPI genes have changed considerably after duplication inEricales For both ancient and several recent duplicationevents divergence in expression was detected indicatingthat the regulation of the duplicate copies has divergedIn the studied species where only one PI copy was isolatedit appears that an lsquolsquoancestralrsquorsquo expression pattern of PI isreestablished However a second copy could have beenmissed during amplification Also within the AP3-like lin-eage some derived expression patterns are detected
In Situ Hybridization Demonstrates ExpressionDivergence in Early Flower Development
To analyze the expression patterns of the PI genes afterduplication in early developmental stages we examined thespatio-temporal expression patterns of PI in selected Eri-cales species by in situ hybridization (fig 4) We selectedexamples of each type of fate after duplication retention
2636 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
of both ancient duplicates (Stewartia) retention of only oneof each PI paralog (C persicum and S rubra) and retentionof recent duplicates (P acaulis)
In S rostrata (fig 4AndashF) we could detect expressiondivergence between each of the PI copies at early stages offloral development At the stage when petal primordia areevident but stamen primordia are apparent only as smallmeristematic outgrowths SpGLO1 expression is detectedin petal and stamen primordia (fig 4A) whereas SpGLO2transcripts are also detected in the tip of the sepals (fig 4D)As the flower develops SpGLO2 expression is low butexpands to include developing carpels (fig 4E) and expres-sion is maintained in the carpels in later stages (fig 4F) Incontrast SpGLO1 expression is maintained at high levelsonly in petals and stamens through later stages (fig 4Band C) and does not expand into sepals and carpels Theseearly patterns of S rostrata PI gene expression are consis-tent with the later patterns of expression as detected byRT-PCR (fig 3I)
Expression patterns of PI homologues were furtherinvestigated in C persicum (fig 4GndashI) and S rubra(fig 4JndashK) CpGLO expression first appears in the youngpetal primordia (fig 4G) followed by expression in the sta-men primordia as they emerge (fig 4H) Expression inthese organs is maintained in later stages (fig 4I) In con-trast to the expression patterns in mature organs as docu-mented by RT-PCR weak expression in the developingsepals (fig 4H) and developing carpels (fig 4I) was de-tected For S rubra we were not able to obtain the earliestdevelopmental stages For later stages of floral develop-ment we observed strong expression of PI in petals andstamens (fig 4J) Weak expression of SdGLO was also de-tected in the developing carpels (fig 4K) which is consis-tent with the RT-PCR data (fig 3) These observationstogether with the RT-PCR data (fig 3) support the ideathat species with presumably only one PI copy in their ge-nome show expression that is more confined to petals andstamens
Finally we investigated the expression of recent PIduplicates in P acaulis (fig 4LndashQ) At early stages bothPaGLO1 (fig 4L) and PaGLO2 (fig 4O) are expressedacross the meristematic dome in sepal primordia commonpetalndashstamen primordia and carpel primordia Expressionof PaGLO2 is maintained in all organs at older stages(fig 4R and Q) whereas PaGLO1 expression is restrictedto the stamens (fig 4M and N)
The in situ expression data (fig 4) in comparison tothe RT-PCR data (fig 3) indicate that early patterns of ex-pression of PI genes are often developmentally regulatedwith different patterns of expression being observed early incomparison to later in flower development Nonethelessboth sets of data indicate that in species possessing dupli-cate PI genes divergence in expression patterns betweenthe two copies is a common theme
Positive Selection and Relaxed Purifying Selection ActsAfter the Duplication Event in Ericales in the K- and C-Cerminal Domain
The mRNA expression data indicate that duplicate PIparalogs have acquired different expression patterns sug-gesting independent evolution of their transcriptional reg-ulatory control It was also demonstrated that adaptiveevolution in the coding regions of MADS-box transcriptionfactors has played a role in the diversification of thisgene family (Martinez-Castilla and Alvarez-Buylla2003) Within class B genes Hernandez-Hernandez et al(2007) demonstrated the role of positive selection after crit-ical duplication events like the AP3-PI duplication and theeuAP3ndashTM6 duplication To investigate whether parallelchanges occurred in the coding regions of the genes exam-ined in this study we investigated the nonsynonymous ver-sus synonymous amino acid replacement rates forsignificant increases before and after duplication in thespecific paralogous clades and along the coding sequenceThe results of likelihood ratio tests are shown in table 2 and
FIG 4mdashIn situ hybridization data for PI genes in various developmental stages of Stewartia rostrata (AndashC SpGLO1 DndashF SpGLO2) Cyclamenpersicum (GndashI CpGLO) Sarracenia rubra (JndashK SdGLO) Primula acaulis (LndashN PaGLO1 OndashQ PaGLO2) lsquolsquoprsquorsquo indicates petalprimordia lsquolsquosrsquorsquoindicates stamen primordia
PISTILLATAmdashDuplications in Asterids 2637
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
specific sites are marked in the alignment presented infigure 5 Parameter estimates log-likelihood scores andBayes Empirical Bayes identified sites are listed in supple-mentary table S2 Supplementary Material online
Comparison of the likelihood scores calculated usingbranch models R1 and R2 for matrix A indicates a signifi-cantly increased evolutionary rate following the ancient du-plication event in Ericales The dNdS estimate from thetwo-ratio branch model after duplication (x 5 0285) istwice compared with the value before duplication (x 50113) illustrating the relaxed selective regime resultingfrom the duplication event However the same test appliedon matrix B did not reveal this change in selective pressureOne possible explanation is that by adding the single-copylineages that evolved after duplication the observed patternin matrix A for the duplicates is obscured In addition wefound no evidence that the two paralogous gene cladescould be evolving at different rates because the three-and four-ratio models did not result in a significantly in-creased log-likelihood as calculated for either of the datamatrices
To formally test for the presence of positive selectionon sequence sites we used the site models proposed byNielsen and Yang (1998) and Yang et al (2000) Analysisof matrix A shows that both M2a and M8 two models thatallow for positive selection fit the data significantly betterthan the corresponding neutral models (M1a and M7) Sub-stitution patterns in about 5 of the sites for the M2amodel and about 9 of the sites for the M8 model appearto result from adaptive evolution Here also analysis of ma-trix B suggests positively selected sites M8 fits the databetter than the neutral model M7 with an estimation of
10 sites under positive selection Bayes empirical Bayes(Yang et al 2005) identification of the sites under positiveselection in matrix A finds seven sites to be positively se-lected under model M8 two of these sites have a posteriorprobability higher than 99 Within matrix B only twopositively selected sites were detected which were alsoidentified in matrix A both with posterior probability high-er than 91 Interestingly all the identified sites are locatedwithin the K2-subdomain the interhelical region betweenK2 and K3 the K3-subdomain and the C-terminal domain(fig 5) These functional domains have been shown to beimportant for proteinndashprotein interactions among Arabi-dopsis MADS-domain proteins (Ma et al 1991 Pnueliet al 1991 Shore and Sharrocks 1995 Riechmann et al1996 Yang et al 2003) The strength of the AP3ndashPI inter-action is mediated primarily by K1 and K2 whereas thePIndashSEP interaction is mediated by K2 the amino-terminalportion of K3 and possibly the C-domain (Yang et al 2003Yang and Jack 2004) The C-terminal region normallyshows much lower sequence conservation but containsshort highly conserved motifs (Lamb and Irish 2003) Sev-eral studies have suggested that this domain is involvedin the formation of higher order protein complexes(Egea-Cortines et al 1999 Honma and Goto 2001) yetthe specific function of the class B C-terminal domainsis unclear (Lamb and Irish 2003 Piwarzyk et al 2007)None of the sites identified by Yang et al (2003) to be re-sponsible for the direct interaction between APETALA3and PISTILLATA are positively selected for in our dataset However site 49 contains a hydrophobic amino acidthat when mutated disrupts the alpha-helical secondarystructure (Yang et al 2003) In addition sites 49 and site60 in figure 5 were identified by Yang and Jack (2004) to becritical for PIndashSEP interaction Site 49 is situated withinthe K2 subdomain which is required for both AP3ndashPIand PIndashSEP interaction whereas site 60 which is locatedat the amino-terminal end of K3 affects when mutated thestrength of the PIndashSEP interaction
To identify evolutionary pressures acting on specificsites along certain branches and in clades we calculatedlikelihoods using the branch-site models First we specifiedthe two branches immediately following the duplication asforeground branches in matrix A Branch-site models thatallow for positive selection in these foreground brancheswere not better at explaining the data than the nested sitemodels M1a and the null model A and indicate no evidencefor a burst of positive selection in the branches immediatelyfollowing the duplication event (Bielawski and Yang 2003)(data not shown) Second we treated all branches in eitherclade of paralogs as foreground branches When these twoclades are treated separately the model allowing for
Table 2Likelihood Ratio Tests Comparing Branch-Models (R1ndashR4)Site Models (M1ndashM8) and Branch-Site Models (Model AndashModel A Null)
Matrix Models Compared2(ln L1 ln L0)
Matrix A One ratio (R1) versus two ratio (R2) 18174Two ratio (R2) versus three ratio (R3) 0435Three ratio (R3) versus four ratio (R4) 1298Neutral (M1a) versus selection (M2a) 20566Beta (M7) versus beta amp w 1 (M8) 2683Model A versus M1a (Test 1) 874Model A versus model anull (Test 2) 252
Matrix B One ratio (R1) versus two ratio (R2) 2348Two ratio (R2) versus three ratio (R3) 001Three ratio (R3) versus four ratio (R4) 01Neutral (M1a) versus selection (M2a) 367Beta (M7) versus beta amp w 1 (M8) 7054
Significant at the 95 level significant at the 99 level
Table 3B Class MADS-Box Protein Interactions as Determined by Yeast Two-Hybrid Assays in Camellia japonica
CjDEF-BD CjTM6-BD CjGLO1-BD CjGLO2-BD BD
CjDEF-AD thorn mdash mdash mdash mdashCjTM6-AD mdash mdash thorn mdash mdashCjGLO1-AD thorn thorn mdash mdash mdashCjGLO2-AD thorn mdash mdash mdash mdashAD mdash mdash mdash mdash mdash
2638 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
positive selection is not significantly better in explaining thedata than the model M1a and null model A (data notshown) When the two clades are treated together as fore-ground branches model A is significantly better than thenested site model M1a (test 1) suggesting that positive se-lection is acting in both clades that arose after duplicationBayes empirical Bayes identification resulted in seven po-sitions though all with posterior probability below 90The improved test 2 from Zhang et al (2005) resulted ina value 252 just beneath the critical value 271 at the5 level but far above the critical value at the 10 level164 These two tests support the hypothesis that positiveselection acts after the duplication event in both paralogousgene clades
Together the identification of changes in selectivepressure in time and along the coding sequence indicatethat upon duplication PI paralogs underwent an increasednonsynonymous substitution rate and that this occurred
specifically in the domains shown to be necessary forproteinndashprotein interactions
Diversification in the Interaction Behavior of B-ClassGenes After the Ancient Duplication Event in BasalAsterids
As sites in the interaction domains of the PI genesfrom basal asterids were detected to be positively selectedafter duplication we further investigated the possible func-tional divergence of the duplicated PI proteins by yeast-2-hybrid analyses We tested the interactions between thefour class B genes in C japonica Camellia japonica con-tains two AP3-like genes one belonging to the euAP3-typeand one from the paleoAP3-type and two PI-like genesfrom an ancient duplication event at the base of the EricalesThis situation is similar to the B-class configuration in thecore asterid species Petunia hybrida (Vandenbussche
FIG 5mdashAlignment of the K-domain and C-terminal domain of the PI duplicates from matrix A used for the analysis in PAML Differentsubdomains and the characteristic PI motif are indicated (Kramer et al 1998) Codon positions identified to be under positive selection are marked withgray boxes
PISTILLATAmdashDuplications in Asterids 2639
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

However the extent to which this dynamic pattern ofclass B gene evolution has played a role in core eudicot flo-ral diversification is not well understood Core eudicotflowers in general are organized in a regular manner witha set number of organ types a clear distinction between se-pals and petals and fused carpels Nonetheless significantmorphological variation can be observed within this groupand in particular the asterids display a very wide range offloral diversity in shape form symmetry and color of thefloral organs The asterids contain some 80000 extantspecies and comprise a total of about one-third of all flower-ing plant species (Stevens 2001 onwards) During early as-terid diversification two extant groups branched off theorders Cornales and Ericales which together we denoteas the lsquolsquobasal asteridsrsquorsquo (Albach et al 2001 Bremer et al2002) These early diverging clades have retained severalfloral characters that are present in both rosids and asteridsincluding a high variation in the degree of sympetaly sta-men number adnation of stamens to corolla and ovulemorphology and anatomy (Stevens 2001 onwards Kubitzki2003) Moreover this group is of special interest becausesome floral morphological features that are constant in coreasterids are variable in basal asterids
Although a number of functional and phylogeneticanalyses have been carried out with several members ofthe asterid clade (eg Petunia Vandenbussche et al 2004Rijpkema et al 2006 Solanum de Martino et al 2006Antirrhinum Sommer et al 1990 Trobner et al 1992 Ger-bera Yu et al 1999 and Lamiales Aagaard et al 20052006) the evolutionary history of class B genes in asteridsis not yet well characterized In particular a well-describedphylogenetic framework for the PI lineage is still lackingFurthermore there has been virtually no sampling of thehighly variable basal asterid families except for Hydrangea(Cornales) (Kramer and Irish 2000)
We reconstructed the evolutionary history ofPISTILLATA- APETALA3- and TM6-like genes in asteridsby isolating new sequences from basal asterids and Aqui-foliales and combining these with previously characterizedclass B genes and expressed sequence tag (EST) data Ouranalyses suggest that during the evolution of both basaland core asterids old and recent PI gene duplications haveoccurred and that these are not correlated with duplicationsin the AP3 lineage Furthermore we have examined thepossibility of functional divergence of duplicate genes inbasal asterids by carrying out expression studies includingreverse transcription polymerase chain reaction (RTndashPCR)and in situ hybridization data as well as by assessing se-quence divergence and changes in interaction behaviorthrough yeast-2-hybrid analysis These data together withprevious analyses of class B gene functional capabilities incore asterids suggest that PI gene duplications have oc-curred multiple times during the evolution of the asteridsand may be associated with the recruitment of these novelduplicates to new floral functions in this clade
Material and MethodsSpecies Sampling
Table 1 lists the species used in this study Our ratio-nale was to sample densely across the orders Ericales and
Cornales because the largest variation in floral morphologyof asterids is found here For these orders species from 19 of23Ericales families and four of sevenCornales familieswerestudied The unsampled families were mainly monospecificor low in taxon number From seven larger families morethan one species was sampled In addition we added Ilexaquifolium from the core asterid order Aquifoliales Thenewly generated sequences are indicated in bold in table 1To supplement our sampling with species from core asteridswe performedGenbank Blast searches in both the nucleotideand EST databases In this way sequences from 27 otherasterid species were added of which the EST sequencesare marked with an asterisk in table 1 Naming of sequencesexplicitly follows the priority rule in which newmembers ofa gene lineage are named after the first discoveredmember ofthis cladeDEFICIENS (Sommer et al 1990) andGLOBOSA(Trobner et al 1992) fromAntirrhinummajus and TM6 fromSolanum lycopersicon (Pnueli et al 1991)
Cloning of Class B MADS-Box Genes
Floral buds were frozen in liquid nitrogen and stored at80 C Total RNA was isolated from these using the In-visorb Spin Plant RNA kit (Invitek Berlin Germany) Fromthis total RNA mRNA was isolated using the NucleotrapmRNA kit (Macherey-Nagel Duren DE) and reverse tran-scribed into cDNA using avian myeloblastosis virus (AMV)reverse transcriptase(PromegaMadisonWI)andthe includedoligo-dT primer Our initial strategy was applying 3-rapidamplification of CDNA by PCR (Frohmann et al 1988) usingprimers (MADSBFCT 5-GGGGTACCAAYMGICAR-GTIACITAYTCIAAGMGIMG-3 RQVT 5-CGRCARG-TGACSTTCTSCAARCG-3 oligodT 5-CCGGATCCT-CTAGAGCGGCCGC(T)17-3) and polymerase chain reac-tion (PCR)-programs from the literature (Kramer et al 1998Winter et al 1999) After a provisional alignment of basal as-terid PISTILLATA-sequences the primer from Kramer et al(1998) was modified (5-GGGGTACCAAYMGGCARGT-GACGTAYTCGAAGMGGMG-3) Based on the samealignment new forward and reverse primers (5-GYTYTGG-GATGCTAARCATGAG-3 3-AGMNAGNACRTYTA-AWCCRAC-5) were designed to amplify a second copy ofPI in Galax urceolata Using this forward primer in com-bination with the oligodT as reverse primer PI homologuesfromMaesaargenteaandAcanthogiliagloriosawere isolatedA second copy of Primula denticulata was isolated usinga primer based on a short PI sequence of Primula vulgarisin Genbank (DQ381443) After creating an alignment ofAP3-like genes a reverse primer on the C-terminus (3-YCCRTYRTCVACYARCCCRTARTGWGGATC) was de-signed and combined with the MADSBFCT forward primer(Krameretal1998)toamplifyAP3-likegenesmorespecificallyAll PCR amplifications were carried out using Taq DNAPolymerase (Invitrogen Carlsbad CA) PCR products weregel-purified with the Nucleospin extract 2 kit (Macherey-Nagel) and cloned into the pGEM-T vector (Promega) Aftertransformation between 50 and 100white cloneswere checkedforinsertsinaPCRreactionusingthesameprimersandprogramPlasmidDNA for selected cloneswas extractedwith theNucle-ospinPlasmidkit (Macherey-Nagel) Inaneffort to sampleclassB members exhaustively for each species between 20 and 40
2628 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Table 1List of Class B Homologues Included in This Study
Order Family Species PI AP3 TM6
Cornales Cornaceae Alangium platinifolium ApGLO ApDEF ApTM6Nyssaceae Nyssa sylvatica NsGLO1 NsGLO2 NsDEF NsTM6
Davidia involucrata DiGLO1 DiGLO2 DiTM6Loasaceae Loasa vulcanica LvGLO LvTM6Hydrangeaceae Hydrangea macrophylla HmPI HmAP3 HmTM6
Philadelphus pubescens PhpTM6Ericales Balsaminaceae Impatiens hawkeri IhGLO IhDEF1
IhDEF2Impatiens glandulifera IgGLO1 IgGLO2
Marcgraviaceae Marcgravia umbellata MuGLO MuDEFLecythidaceae Gustavia brasiliensis GbGLO1 GbGLO2 GbTM6
Napoleonaea vogelii NvGLO NvDEFPolemoniaceae Phlox paniculata PpGLO PpDEF PpTM6
Acanthogilia gloriosa AgGLOIpomopsis aggregata IaGLO
Sapotaceae Synsepalum dulcificum SydGLO SydDEFDiapensiaceae Galax urceolata GuGLO1 GuGLO2 GuDEF GuTM6Styracaceae Halesia diptera HdGLO1 HdGLO2 HdDEF
Styrax japonicus SjGLO SjDEF SjTM6Roridulaceae Roridula gorgonias RgGLOActinidiaceae Saurauia zahlbruckneri SzGLO1 SzGLO2
SzGLO3SzTM6
Actinidia chinensis AcGLO1 AcGLO2Clethraceae Clethra tomentosa CtGLO CtDEF CtTM6Theaceae Stewartia pseudocamellia SpGLO1 SpGLO2
SpGLO3SpDEF
Camellia japonica CjGLO1 CjGLO2 CjDEF CjTM6Symplocaceae Symplocos chinensis ScGLO1 ScGLO2 ScTM6Sarraceniaceae Sarracenia drummondii SdGLO SdDEF SdTM6Ericaceae Enkianthus campanulatus EcGLO
Erica hiemalis EhGLO1 EhGLO2 EhDEFEbenaceae Diospyros digyna DdGLO DdDEFPrimulaceae Primula denticulata PdGLO1 PdGLO2 PdDEFMyrsinaceae Cyclamen persicum CpGLO CpDEF
Hymenandra walichii HwGLO HwDEF HwTM6Theophrastaceae Jacquinia auriantaca JaGLO JaDEF JaTM6
Clavija latifolia ClGLO ClDEFMaesaceae Maesa argentea MaGLO MaDEF
Aquifoliales Aquifoliaceae Ilex aquifolium IaGLO1 IaGLO2 IxaAP3 IxaTM6Asterales Asteraceae Gerbera hybrida GGLO1 GDEF2 GDEF1
Lactuca sativa LsGLOTaraxacum officinale ToGLOCichorium intybus CiGLOSenecio vulgaris SvGLOHelianthus annuus HaPI HaAP3 HaTM6Centaurea solstitialis CsGLOHieracium piloselloides HpDEF1
HpDEF2Gentianales Rubiaceae Hedyotis terminalis HtGLO
Apocynaceae Catharanthus roseus CrGLOGentianaceae Eustoma grandiflorum EgGLO1 EgDEF
Solanales Solanaceae Petunia hybrida FBP1 PMADS2 PhDEF PhTM6Solanum lycopersicon SlGLO1 SlGLO2 LeAP3Ipomoea nil InGLONicotiana tabacum NtGLOCestrum elegans CeDEF CeTM6Nicotiana benthamiana NtTM6Solanum pseudolulo SopDEF SpTM6Brunfelsia uniflora BuDEF BuTM6
Lamiales Scrophulariaceae Mimulus guttatus MgGLOMimulus ringens MrDEFA
MrDEFBVerbenaceae Verbena hybrida VhGLOPlantaginaceae Antirrhinum majus GLO DEF
Misopates orontium MoGLO MoDEFChelone glabra CgDEF
Oleaceae Syringa vulgaris SvPI SvAP3Lamiaceae Salvia coccinea ScDEFA
ScDEFB
PISTILLATAmdashDuplications in Asterids 2629
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
plasmid inserts were sequenced using T7 and SP6 universalprimers using the BigDye Terminator 11 kit (Applied Bio-systems Foster City CA) on an Applied Biosystems 310 se-quencer or the plasmids were sent for sequencing (MacroGenInc Seoul South Korea) New sequences were deposited inGenbank (Accession numbers GQ141105ndashGQ141187)
Identification and Sequence Analysis
Obtained nucleotide sequences were translated intothe corresponding amino acid sequence and manuallyaligned using MacClade 4 (Maddison RP and MaddisonWP 2003) together with previously cloned and character-ized class B sequences from Genbank (table 1) After align-ment nucleotide sequences were analyzed with PAUP4b10 (Swofford 2002) MrBayes 312 (Huelsenbeck andRonquist 2001) and PHYML (Guindon and Gascuel2003) (PI matrix only) Paup 4b10 (Swofford 2002)was used for parsimony bootstrap analysis A MaximumParsimony (MP) heuristic search was conducted using1000 random addition sequences with tree bisection andreconnection branch swapping and saving of multiple par-simonous trees (MulTrees on) Branch support was calcu-lated by nonparametric bootstrap analysis on 1000pseudoreplicate data sets (Felsenstein 1985) Parametersfor the Bayesian analysis and maximum likelihood (ML)analysis were estimated using Modeltest 306 (Posadaand Crandall 2001) Modeltest selected the GTR (generaltime reversible) thorn I thorn G substitution model using theAkaike Information criterion MrBayes was run for 5 mil-lion generations where for every 100 generations one treewas saved For both matrices the search reached the sta-tionary phase around 50000 generations This numberwas considered the lsquolsquoburn-in periodrsquorsquo and was excludedwhen the consensus phylogeny was constructed PHYML(Guindon and Gascuel 2003) was used for ML inference ofthe PImatrix Confidence in the clades was estimated by theapproximate likelihood ratio test (aLRT) method from Ani-simova and Gascuel (2006) and traditional bootstrap anal-
ysis with 100 replicates For the aLRT tests we used boththe v2 statistic and the more conservative shimodeiva-Hasqawa (SH)-like test as branch support measures To fur-ther look into the supposed ancient PI duplication duringthe evolution of the Ericales clade we additionally con-structed a matrix with only the duplicated PI genes resultingfrom this duplication Cornales PI genes were used as out-groups as the reconstructed PI phylogeny gives us noclear answers on the exact timing of duplication A similaranalysis strategy was used but Modeltest selected theHasegawandashKishinondashYano (HKY) thorn G substitution modelfor the Bayesian and ML analyses
Semiquantitative RT-PCR
RNA was extracted from each organ type separatelyusing the Invisorb Spin Plant RNA kit (Invitek) and eachRNA sample was DNase treated using TURBO DNA-free(Ambion Austin TX) Total RNA was reverse transcribedinto cDNA using AMV reverse transcriptase (Promega) andthe included oligo-dT primer and random primers Gene-specific primers are listed in supplementary table S1 Sup-plementary Material online Expression of AP3 in Gustaviabrasiliensis and of TM6 in Cyclamen persicum Primulaacaulis Stewartia pseudocamellia andHalesia diptera wereexaminedwith primers fromAP3-like sequences from closelyrelated species Ribosomal 18S RNA was used as a control
In Situ Hybridization
Antisense probes were in vitro transcribed using T7RNApolymerase (NewEnglandBiolabs IpswichAustralia)in thepresence of digoxigenin-labeled uracil-5-triphosphate(Roche Basel Switzerland) from PCR-amplified templatesthat included a T7 promoter Tissues were fixed in 4 para-formaldehydeparaffinembeddedandsectionedat8lmSec-tions were mounted on Probe-On-Plus slides (FisherScientific Pittsburgh PA) Prehybridization hybridizationand detection essentially followed Carr and Irish (1997)
Table 1Continued
Order Family Species PI AP3 TM6
Ranunculales Ranunculaceae Aquilegia vulgaris AqvPITrautvetteria carolinensis TrcAP3
Vitales Vitaceae Vitis vinifera VvDEF VvTM6Saxifragales Grossulariacaea Ribes sanguineum
Saxifragaceae Saxifraga careyana SxcAP3 SxcTM6Malpighiales Salicaceae Populus deltoides
Populus tomentosa PtTM6Myrtales Myrtaceae Melaleuca quinquenerviaRosales Rosaceae Malus domestica MdTM6
Rosa rugosa RrDEF RrTM6Cucurbitales Cucurbitaceae Cucumis sativusFagales Betulaceae Betula pendula
Juglandaceae Juglans regia JrDEFTrochodendrales Trochodendraceae Trochodendron aralioides TraPI1 TroAP3Magnoliales Annonaceae Asimina longifolia AlAP3Dilleniales Dilleniaceae Dillenia indica DiiAP3 DiiTM6Asparagales Orchidaceae Phalaenopsis hybrid PhahAP3
Brassicaceae Arabidopsis thaliana AP3Brassica juncea BjAP3
Newly generated asterid sequences are marked in bold EST data are highlighted with an asterisk
2630 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Hybridizations in Sarracenia rubra and Stewartia rostratawere carried out using heterologous probes of the closely re-lated speciesSarracenia drummondii andS pseudocamellia
Codon Substitution Models
Omega (x or dNdS) is a measure of the amount of nat-ural selection acting on the protein To test whether positiveselection acted after the duplication event at the base of theEricales the previously obtained Bayesian topology of PIgenes was pruned in TreeEdit (indicated in fig 1) and thematching data sets were analyzed using PAML version314 (Yang 1997) We used two data matrices to investigate
selective pressure a matrix in which only the duplicatedgenes that originated after the duplication event at the baseof the Ericales are present (hereafter matrix A) and a secondmatrix in which the paralogous copies from species whohave lost either one of both paralogs are incorporated inmatrix A (hereafter matrix B) The log-likelihood of the se-quence alignments was calculated using different codonsubstitution models and likelihood ratio tests were usedto detect significant differences in fit between nested mod-els Branch models (Yang 1998 Yang and Nielsen 1998)were used to test for episodic adaptive evolution after du-plication either in all lineages combined or in paralogouslineages separately To test for the presence of positive
FIG 1mdashEvolutionary reconstruction of PI gene phylogeny in asterids as inferred from ML and Bayesian analysis Bayesian posterior probabilitiesand nonparametric bootstrap support values are indicated above branches SH-like and X2-like supports are shown below branches Gray circle 5
inferred duplication event Data sets used for assessments of selection pressures are indicated by stars black star 5 sequences included in matrix Awhite star 5 extra sequences added to matrix A to form Matrix B Marcgravia umbellata and Diospyros digyna PI sequences are underlined
PISTILLATAmdashDuplications in Asterids 2631
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
selection on sequence sites site models were applied thathold x constant among all branches but allow x to varyamong codons Site models include M1a M2a M7 andM8 (Nielsen and Yang 1998 Wong et al 2004 Yang et al2005) Positive selection is inferred when M2a and M8 arerespectively significantly better than M1a and M7 Branch-sitemodels allow thex ratio to vary amongboth sites and lin-eages (Zhang et al 2005) and can be used to detect positivelyselected sites in specific lineages within the tree We usedthe modified branch-site model A and compared this modelwith the nested site model M1a (Test 1) and the null modelA (Test 2) to test for positive selection (Zhang et al 2005)
Yeast Two-Hybrid Assays
The intervening- keratin-like and C-terminal domaindomains of the two PI (CjGLO1 and CjGLO2) the AP3(CjDEF) and TM6 genes (CjTM6) from Camellia japonicawere fusedwith theGAL4activationdomain in thepGAD424vector and the GAL4 DNA-binding domain in the pGBT9vector (ClontechMountainView CA) Each possible vectorcombination was transformed in yeast and two colonies pertransformation were used for B-galactosidase liquid assaysusing the protocol available at httpwwwfhcrcorgsciencelabsgottschlingyeast20ProtocolsBgalhtml
Results
In total 83 new B-lineage sequences were amplifiedfrom asterid species Forty-seven new PI and 36 new AP3-like sequences were identified from 31 and 25 asterid speciesrespectively (table 1) All but one of these species belong tothe basal asterid orders Cornales and Ericales In addition PIhomologues were amplified from I aquifolium (Aquifoliales)
Two Ancient and Frequent Recent Duplications HaveOccurred in the Asterid PI Lineage
For the phylogenetic analyses of the PI lineage wecombined the newly identified PI genes with previouslyidentified asterid PI sequences and EST data from Genbankto generate one data set Basal eudicot sequences were usedas outgroups for this data set and AqvPI (Aquilegia Ranun-culales) was used to root the PI tree A nucleotide alignmentof 74 taxa was analyzed using MP ML and Bayesian anal-yses The results are summarized in figure 1
The Bayesian topology which has relatively high pos-terior probabilities for a majority of nodes did not conflictwith the most likely tree from the ML analysis The MPanalysis however did not resolve the relationships betweenthe major clades and high bootstrap values were only ob-tained for nodes at lower taxonomical levels Similar to thebootstrap values from the MP analysis bootstrap supportfor the ML tree is low and we chose to only indicate thebootstrap values from the MP analysis on the phylogenyHowever to provide an additional measure of support ina likelihood framework we show SH-like and v2-like aLRTtest values These provide additional support for the Bayes-ian inferred relationships
The inferred PI phylogeny (fig 1) indicates that thisgene lineage probably underwent two ancient duplication
events during asterid evolution In both instances the twocopies were retained in some extant species lineageswhereas in other taxa one copy appears to have been lostor further duplicated An initial ancient duplication probablyoccurred early in the evolution of the core asterids becausethe Bayesian analysis supports two paralogous cladesa clade containing Gentianales Solanales and Asteralessequences (BPP 95) and one with Solanales and Lamialessequences (BPP 100) The latter is also strongly supportedbyMPbootstrapping (86) and aLRT testing (10099) Givenour current knowledge of asterid evolution (Albach et al2001 Bremer et al 2002) this suggests an ancient duplica-tion event before the origin of the two major core asteridclades (euasterids I and euasterids II) (Bayesian analysis)or at least before the origin of the euasterid II clade compris-ing Solanales Lamiales and Gentianales (MP analysis andaLRT testing) This duplication event was then followed byindependent losses in each paralogous lineageAlthoughourresults allow us to infer such a scenario no Garryales Dip-sacales or Apiales PI representatives are currently availablethat would further support such a hypothesis In addition thePI sequences of Aquifoliales an order situated at the base ofthe euasterid II clade occupy a nonsupported position at thebase of basal asterid PI sequences Together this suggeststhat an even denser sampling of asterid sequences would bedesirable to determine the exact moment of the PI duplica-tion in the core asterid lineage with even more confidence
The ML tree further suggests a second ancient dupli-cation event consistent with an origin early in the radiationof the order Ericales The PI sequences from Ericales rep-resentatives that are strongly supported by Bayesian support(BPP 96) and aLRT support (9999) are placed into two sep-arate clades A first clade is supported by a BPP value of 90and a v2-value of 92 The weakly supported second clade(BPP 67 aLRT 7088) consists of two strongly supportedclades (BPP 96 aLRT 9499 and BPP 90 aLRT 8999)where one (BPP 96) consists of the proposed basal lineagesof the Ericales the balsaminoıd clade and Lecythidaceae(Geuten et al 2004 Schonenberger et al 2005) The pres-ence of three strongly supported clades within the Ericalessuggests that the duplication in Ericales could have occurredeither at the base of the Ericales (as the ML phylogenyshows) or after the divergence of the basal lineages (balsami-noids and Lecythidaceae)Galax urceolata (Diapensiaceae)Hdiptera (Styracaceae)Saurauia zahlbruckneri (Actinidia-ceae) S pseudocamellia and C japonica (Theaceae) andSymplocos chinensis (Symplocaceae) all have multiple PIcopies belonging to two strongly supported clades due to thisancient duplication event Again this ancestral duplication isfollowed by different gene fateswith independent losses andduplications in each paralogous lineage Although furthersampling could lead to alternative explanations for the pro-posed multiple loss events there is some support for the hy-pothesis of independent gene loss in eachparalogous lineageSouthern blot analyses of Marcgravia umbellata (Geutenet al 2006) and Diospyros digyna (data not shown) both in-dicate that there is likely only one PI gene present in each ofthese species (fig 1 underlined) This observation supportsthe hypothesis of multiple PI gene losses within the Ericales
Because we wanted to explore the effect of omittingtaxa with single-copy genes of which the paralogous was
2632 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
putatively lost after duplication we constructed a secondmatrix with only duplicated PI genes due to the lsquolsquoancientrsquorsquoduplication from G urceolata (Diapensiaceae) H diptera(Styracaceae) S zahlbruckneri (Actinidiaceae) S pseudo-camellia and C japonica (Theaceae) and S chinensis(Symplocaceae) The MP analysis resulted in just one mostparsimonious tree that is identical to the tree with the high-est likelihood As in the previous analysis the same twoparalogous clades were identified In this case both BPPvalues (100 and 97) and ML bootstrap values (57 and53) were higher in this case supporting a duplication eventpresumably at least before the divergence of these familiesand after the divergence of the primuloid clade in Ericalesphylogeny (Schonenberger et al 2006)
In addition to these two ancient duplication events weidentified several other more recent duplication events inasterid evolution From I aquifolium (Aquifoliaceae) weamplified two PI copies but more extensive sampling ofAquifoliales representatives should reveal the timing of thisduplication more accurately In Cornales duplicate genesare found in both Davidia involucrata and Nyssa sylvatica(Nyssaceae) suggesting that the duplication that resulted inthese copies occurred in the lineage leading to NyssaceaeIn the clade of Ericales representatives there are seven spe-cies with more than one PI copy which appear to be morerecent than the previously described duplication event atthe base of the Ericales order For example we isolatedtwo PI homologues from Impatiens glandulifera (IgGLO1and IgGLO2 Balsaminaceae) A previously characterizedPI gene from Impatiens hawkeri (IhGLO Geuten et al2006) groups with IgGLO2 but not with the PI gene fromM umbellata (MuGLO Marcgraviaceae) suggesting thatthis duplication occurred at the base of the BalsaminaceaeSimilarly we isolated twoPI genes fromG brasiliensis (Le-cythidaceae) and the phylogeny indicates that one copy ismore related to the PI gene from another Lecythidaceae rep-resentativeNapoleonaea vogeliiBecauseGustavia is basalin the phylogeny of Lecythidaceae (Mori et al 2007) the du-plication leading to paralogous copies inGustavia probablyoccurred at the origin of the Lecythidaceae or earlier
We also identified two PI copies in Erica hiemalis(Ericaceae) and Actinidia chinensis (Actinidiaceae) In bothS pseudocamellia (Theaceae) and S zahlbruckneri (Acti-nidiaceae) a third PI copy is present presumably as a resultof a more recent duplication in one of their paralogs As thesequence similarity of the paralogs in these four species ishigh (91ndash98) it is hard to decide whether these genes areat the same genetic locus or at different loci We suggestthese genes to be paralogous copies with a very recent or-igin and not shared by any other species in our samplingFinally two PI genes were identified in P denticulata (Pri-mulaceae) Although sequence similarity among thesegenes is significantly lower (82) this duplication is notshared by any other PI genes in our analyses indicating thatthis duplication is specific to the Primulaceae
Duplications in the AP3 and TM6 Lineages Are Few andNot Correlated with PI Duplications
We also carried out phylogenetic analyses of asteridAP3 sequences using a number of newly identified AP3
sequences as well as previously characterized genes andEST sequences from Genbank (table 1) The phylogenywas inferred from a nucleotide alignment of genes from85 taxa which was analyzed using both MP and Bayesianmethods (fig 2) In the Bayesian analysis AlAP3 was usedto root the AP3-like tree Similar to previous analyses(Kramer and Irish 1999 Kramer et al 2006) our Bayesiananalysis recovered two highly supported clades A firstclade contains the euAP3 homologues (BPP 98) whereasa second clade contains the TM6 homologues (BPP 100)(fig 2) The duplication event that gave rise to these two eu-dicot AP3 gene clades is placed near the base of the core eu-dicots and likely occurred coincident with the origin ofTrochodendraceae and Buxaceae (Kramer et al 2006)Within core asterids our phylogeny gave no indicationsofmajor duplication events but onlyfiveof eight core asteridorderswere sampled Similarly in basal asterids duplicationevents have seemingly not played an important role duringAP3 evolution because the observed gene phylogeny did notconflict with the species phylogeny (Schonenberger et al2006) In several cases thoughparalogousAP3genesofveryrecent origin were identified in four species one from Eri-cales (I hawkeri Geuten et al 2006) and three from core as-terids (Hieracium piloselloides Mimulus cultstatus andSalvia coccinea) (Aagaard et al 2005) As for thePI phylog-eny the bootstrap values in the MP analyses were low andthese analyses resulted in a poorly resolved tree
After the basal eudicot and rosid representativesbranch off in the euAP3 clade (BPP 98) the Cornales rep-resentatives are sister to two large clades The first cladecontains the newly sampled Ericales representatives(BPP 81) whereas the second contains all core asterid se-quences (BPP 54) The unexpected association of Brassica-ceae representatives (AP3 BjAP3) with the euAP3 genesfrom Asterales was also present in previous analyses andprobably illustrates the need for even denser sampling(de Martino et al 2006 Kramer et al 2006) Within theEricales euAP3 clade (BPP 81) most relationships arepoorly resolved However a strongly supported primuloid(BPP 100) ericoid (BPP 91) and balsaminoid clade (BPP100) can be observed (Schonenberger et al 2005) Withinthe TM6-clade Ericales representatives are supported witha BPP of 99 but overall support within this clade is lowHowever we did not recover any paralogous copies of TM6genes in the species sampled suggesting that duplicationevents in this clade either do not occur readily or are se-lected against We also observed an unexpected associationof Asterales TM6 representatives with rosids (BPP 71) asreported in previous analyses (de Martino et al 2006Kramer et al 2006) The TM6 genes within the Solanalesare strongly supported as a clade (BPP 100)
RT-PCR Reveals Divergence of Expression PatternsAfter Duplication
To investigate whether gene duplicates have divergedin expression patterns or have seemingly remained underthe same regulatory control we used RT-PCR to monitorgene expression levels in mature dissected floral organs(fig 3) We limited our survey to Ericales species thatcontain representatives of PI genes arising from both the
PISTILLATAmdashDuplications in Asterids 2633
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
ancient and recent duplications We find that the expressionpattern of PI paralogs has diverged considerably and fairlyrapidly The PI protein is known to require an AP3-like in-teraction partner for its in vivo function in a number of coreeudicots (Jack et al 1992 Trobner et al 1992 Goto andMeyerowitz 1994) We therefore also examined the expres-sion patterns of TM6 and euAP3 genes in the same species
We compared Ericales expression patterns with the as-terid speciesPetunia hybrida (Solanaceae) BothPI genesin Petunia show a very similar expression pattern (Vanden-bussche et al 2004) Expression of these twoPI homologues(FBP1 and PMADS2) is mainly confined to the second andthirdwhorls (Angenent et al 1992Van derKrol et al 1993)Expression of PhDEF the AP3 homologue was mainly
FIG 2mdashEvolutionary reconstruction of APETALA3-like gene phylogeny in asterids as inferred from Bayesian analysis Bayesian posteriorprobabilities are indicated above branches nonparametric bootstrap support values are shown below branches Gray circle5 inferred duplication event
2634 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
confined to petals and stamens with low expression in firstand fourth whorl organs PhTM6 has a different expressionprofilewithstrongexpressioninstamensandcarpelswhereasexpression in sepals and petals is low (Vandenbussche et al2004) Similar expression patterns to Petunia of PI AP3 and
TM6 were detected in S lycopersicon another core asteridSolanaceae species (de Martino et al 2006)
Multiple copies of PI have been retained in the ge-nomes of S pseudocamellia C japonica and H dipteraafter the major duplication event at the base of the Ericales
FIG 3mdashRT-PCR expression data for B-class genes from selected Ericales representatives Sep 5 sepals pet 5 petals pet up 5 upper part ofpetals fus 5 fused part of petals and stamens sta 5 stamens sta up 5 upper part of stamens sta fus 5 fused part of stamens gyn 5 gynoecium andflo 5 flower
PISTILLATAmdashDuplications in Asterids 2635
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Both copies in each of these species appear to have divergedin their expression patterns after duplication (fig 3AndashC)Although HdGLO2 is expressed in the petals and stamensof H diptera the other copy (HdGLO1) is mainly ex-pressed in stamens and the gynoecium and absent in petalsIn comparison the TM6 and euAP3 lineage genes show un-expected expression patterns with strong expression ofHdTM6 in all disected organs but HdDEF expression beingmainly restricted to petals and carpels InC japonica one PIparalog (CjGLO2) is expressed throughout sepals petalsand stamens whereas expression of the second paralog(CjGLO1) appeared to be restricted to the nonfused partsof petals stamens and carpels Although both CjDEF andCjTM6 expression are detected in all organs strong expres-sion ofCjDEF is seenmainly in the sepals and petalsCjTM6expression is strongest in all stamen parts and in carpelsS pseudocamellia also displays differences in the expressionof its two PI copies One paralog (SpGLO2) is expressed inpetals stamens carpels and expression of the second paral-og (SpGLO1) is more restricted to the fused part of petals andstamens and upper part of petals The AP3 homologueSpDEF is strongly expressed in all stamen parts and carpelswhereas strong expression of SpTM6 is detected in sepals allstamen parts and carpels
In summary in these three cases there is one PI copywith a broad expression pattern in the flower (even in sepalsand carpels) whereas expression of the other PI copy ismore restricted to specific parts of petals and stamensThese expression patterns differ from those of PI lineagegenes in core asterid species (Vandenbussche et al2004 de Martino et al 2006) where PI expression (alsoin a duplicated configuration) is mainly restricted to the sec-ond and third whorl organs petals and stamens and nostrong expression in the first and fourth whorl organs is de-tected Interestingly the divergence in expression patternsin these species does not follow the gene phylogeny Theparalogous copies with a general expression pattern are notgrouped in one clade of PI genes in Ericales but rather arefound in two clades This suggests that the PI expressiondomains were not fixed immediately after duplication in-dicating that subfunctionalization in spatial expression doesnot explain retention of the duplicates Alternatively ifregulatory subfunctionalization was the main reason forretention evidence for this has been obscured throughtime Differences in expression between AP3 and TM6in C japonica seem to be very similar to members ofthe same lineages in the core asterid species Expressionof AP3-like genes in H diptera and S pseudocamelliahowever differs from those of Camellia with strongAP3 expression in the carpels of Stewartia and Halesiaand weak expression of AP3 in stamens of Halesia In eachof these species expression of one or both AP3-like genes isdetected in the sepals
Sarracenia drummondii (fig 3D) Styrax japonicus(fig 3E) C persicum (fig 3F) and Phlox paniculata(fig 3G) maintained probably only one of the paralogousPI copies in their genome after the ancient Ericales dupli-cation Moreover Sarracenia and Styrax maintained a dif-ferent copy (fig 1 BPP 68) than did Cyclamen and Phlox(fig 1 BPP89) We investigated the expression patterns ofPI genes in these species to monitor the possible divergence
between the two paralogous PI clades after duplication Ineach of these four species PI is expressed in petals and sta-mens similar to the expression patterns of PI in core aster-ids in previous studies (Vandenbussche et al 2004 deMartino et al 2006) In both Sarracenia (SdGLO) andPhlox (PpGLO) there is some expression detected in thecarpels whereas SjGLO (Styrax) is also expressed in sepalsNo clear divergence in expression of PI from the two dif-ferent clades (fig 1 BPP68 and BPP89) is detected in thesecond and third whorl organs The expression patterns ofAP3 and TM6 lineage genes are different between these fourspecies Although in S drummondii both AP3 and TM6 areexpressed in all four floral whorls the expression in Styraxis very similar to that of TM6 and AP3 in core asterid spe-cies with in addition to expression in petals and stamensexpression of TM6 in carpels and AP3 in sepals In Cycla-men TM6 expression is absent in the upper part of the sta-mens and strong expression of AP3 is detected in thecarpels In Phlox however TM6 expression is absent fromthe carpels
The multiple PI copies in G brasiliensis (Lecythida-ceae) and P acaulis (Primulaceae) are the result of a recentduplication probably at the base of these two families(fig 1) We looked at the expression pattern of each of theseduplicated genes and of the AP3-like genes in these speciesIn G brasiliensis GbGLO2 is expressed in petals and sta-mens whereas GbGLO1 is mainly expressed in the stami-nal parts and less in the petals (fig 3H) Expression of AP3and TM6 seems to be lsquolsquoinversersquorsquo compared with the ex-pected asterid expression with AP3 expression mainly re-stricted to the stamens and strong TM6 expression in thepetals No AP3-like expression is detected in the carpelsIn P acaulis there are differences between the expressionpatterns of both PI copies (fig 3I) PaGLO2 is expressed inall organs tested whereas the expression of PaGLO1 ismainly restricted to the sepals and petals Both AP3 andTM6 are strongly expressed in sepals and as inC persicumTM6-expression is weak in the staminal parts Even afterthese recent duplication events divergence in the expres-sion domain of the two PI copies is detected
In contrast to the duplicated PI genes of Petunia(Vandenbussche et al 2004) the expression domains ofPI genes have changed considerably after duplication inEricales For both ancient and several recent duplicationevents divergence in expression was detected indicatingthat the regulation of the duplicate copies has divergedIn the studied species where only one PI copy was isolatedit appears that an lsquolsquoancestralrsquorsquo expression pattern of PI isreestablished However a second copy could have beenmissed during amplification Also within the AP3-like lin-eage some derived expression patterns are detected
In Situ Hybridization Demonstrates ExpressionDivergence in Early Flower Development
To analyze the expression patterns of the PI genes afterduplication in early developmental stages we examined thespatio-temporal expression patterns of PI in selected Eri-cales species by in situ hybridization (fig 4) We selectedexamples of each type of fate after duplication retention
2636 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
of both ancient duplicates (Stewartia) retention of only oneof each PI paralog (C persicum and S rubra) and retentionof recent duplicates (P acaulis)
In S rostrata (fig 4AndashF) we could detect expressiondivergence between each of the PI copies at early stages offloral development At the stage when petal primordia areevident but stamen primordia are apparent only as smallmeristematic outgrowths SpGLO1 expression is detectedin petal and stamen primordia (fig 4A) whereas SpGLO2transcripts are also detected in the tip of the sepals (fig 4D)As the flower develops SpGLO2 expression is low butexpands to include developing carpels (fig 4E) and expres-sion is maintained in the carpels in later stages (fig 4F) Incontrast SpGLO1 expression is maintained at high levelsonly in petals and stamens through later stages (fig 4Band C) and does not expand into sepals and carpels Theseearly patterns of S rostrata PI gene expression are consis-tent with the later patterns of expression as detected byRT-PCR (fig 3I)
Expression patterns of PI homologues were furtherinvestigated in C persicum (fig 4GndashI) and S rubra(fig 4JndashK) CpGLO expression first appears in the youngpetal primordia (fig 4G) followed by expression in the sta-men primordia as they emerge (fig 4H) Expression inthese organs is maintained in later stages (fig 4I) In con-trast to the expression patterns in mature organs as docu-mented by RT-PCR weak expression in the developingsepals (fig 4H) and developing carpels (fig 4I) was de-tected For S rubra we were not able to obtain the earliestdevelopmental stages For later stages of floral develop-ment we observed strong expression of PI in petals andstamens (fig 4J) Weak expression of SdGLO was also de-tected in the developing carpels (fig 4K) which is consis-tent with the RT-PCR data (fig 3) These observationstogether with the RT-PCR data (fig 3) support the ideathat species with presumably only one PI copy in their ge-nome show expression that is more confined to petals andstamens
Finally we investigated the expression of recent PIduplicates in P acaulis (fig 4LndashQ) At early stages bothPaGLO1 (fig 4L) and PaGLO2 (fig 4O) are expressedacross the meristematic dome in sepal primordia commonpetalndashstamen primordia and carpel primordia Expressionof PaGLO2 is maintained in all organs at older stages(fig 4R and Q) whereas PaGLO1 expression is restrictedto the stamens (fig 4M and N)
The in situ expression data (fig 4) in comparison tothe RT-PCR data (fig 3) indicate that early patterns of ex-pression of PI genes are often developmentally regulatedwith different patterns of expression being observed early incomparison to later in flower development Nonethelessboth sets of data indicate that in species possessing dupli-cate PI genes divergence in expression patterns betweenthe two copies is a common theme
Positive Selection and Relaxed Purifying Selection ActsAfter the Duplication Event in Ericales in the K- and C-Cerminal Domain
The mRNA expression data indicate that duplicate PIparalogs have acquired different expression patterns sug-gesting independent evolution of their transcriptional reg-ulatory control It was also demonstrated that adaptiveevolution in the coding regions of MADS-box transcriptionfactors has played a role in the diversification of thisgene family (Martinez-Castilla and Alvarez-Buylla2003) Within class B genes Hernandez-Hernandez et al(2007) demonstrated the role of positive selection after crit-ical duplication events like the AP3-PI duplication and theeuAP3ndashTM6 duplication To investigate whether parallelchanges occurred in the coding regions of the genes exam-ined in this study we investigated the nonsynonymous ver-sus synonymous amino acid replacement rates forsignificant increases before and after duplication in thespecific paralogous clades and along the coding sequenceThe results of likelihood ratio tests are shown in table 2 and
FIG 4mdashIn situ hybridization data for PI genes in various developmental stages of Stewartia rostrata (AndashC SpGLO1 DndashF SpGLO2) Cyclamenpersicum (GndashI CpGLO) Sarracenia rubra (JndashK SdGLO) Primula acaulis (LndashN PaGLO1 OndashQ PaGLO2) lsquolsquoprsquorsquo indicates petalprimordia lsquolsquosrsquorsquoindicates stamen primordia
PISTILLATAmdashDuplications in Asterids 2637
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
specific sites are marked in the alignment presented infigure 5 Parameter estimates log-likelihood scores andBayes Empirical Bayes identified sites are listed in supple-mentary table S2 Supplementary Material online
Comparison of the likelihood scores calculated usingbranch models R1 and R2 for matrix A indicates a signifi-cantly increased evolutionary rate following the ancient du-plication event in Ericales The dNdS estimate from thetwo-ratio branch model after duplication (x 5 0285) istwice compared with the value before duplication (x 50113) illustrating the relaxed selective regime resultingfrom the duplication event However the same test appliedon matrix B did not reveal this change in selective pressureOne possible explanation is that by adding the single-copylineages that evolved after duplication the observed patternin matrix A for the duplicates is obscured In addition wefound no evidence that the two paralogous gene cladescould be evolving at different rates because the three-and four-ratio models did not result in a significantly in-creased log-likelihood as calculated for either of the datamatrices
To formally test for the presence of positive selectionon sequence sites we used the site models proposed byNielsen and Yang (1998) and Yang et al (2000) Analysisof matrix A shows that both M2a and M8 two models thatallow for positive selection fit the data significantly betterthan the corresponding neutral models (M1a and M7) Sub-stitution patterns in about 5 of the sites for the M2amodel and about 9 of the sites for the M8 model appearto result from adaptive evolution Here also analysis of ma-trix B suggests positively selected sites M8 fits the databetter than the neutral model M7 with an estimation of
10 sites under positive selection Bayes empirical Bayes(Yang et al 2005) identification of the sites under positiveselection in matrix A finds seven sites to be positively se-lected under model M8 two of these sites have a posteriorprobability higher than 99 Within matrix B only twopositively selected sites were detected which were alsoidentified in matrix A both with posterior probability high-er than 91 Interestingly all the identified sites are locatedwithin the K2-subdomain the interhelical region betweenK2 and K3 the K3-subdomain and the C-terminal domain(fig 5) These functional domains have been shown to beimportant for proteinndashprotein interactions among Arabi-dopsis MADS-domain proteins (Ma et al 1991 Pnueliet al 1991 Shore and Sharrocks 1995 Riechmann et al1996 Yang et al 2003) The strength of the AP3ndashPI inter-action is mediated primarily by K1 and K2 whereas thePIndashSEP interaction is mediated by K2 the amino-terminalportion of K3 and possibly the C-domain (Yang et al 2003Yang and Jack 2004) The C-terminal region normallyshows much lower sequence conservation but containsshort highly conserved motifs (Lamb and Irish 2003) Sev-eral studies have suggested that this domain is involvedin the formation of higher order protein complexes(Egea-Cortines et al 1999 Honma and Goto 2001) yetthe specific function of the class B C-terminal domainsis unclear (Lamb and Irish 2003 Piwarzyk et al 2007)None of the sites identified by Yang et al (2003) to be re-sponsible for the direct interaction between APETALA3and PISTILLATA are positively selected for in our dataset However site 49 contains a hydrophobic amino acidthat when mutated disrupts the alpha-helical secondarystructure (Yang et al 2003) In addition sites 49 and site60 in figure 5 were identified by Yang and Jack (2004) to becritical for PIndashSEP interaction Site 49 is situated withinthe K2 subdomain which is required for both AP3ndashPIand PIndashSEP interaction whereas site 60 which is locatedat the amino-terminal end of K3 affects when mutated thestrength of the PIndashSEP interaction
To identify evolutionary pressures acting on specificsites along certain branches and in clades we calculatedlikelihoods using the branch-site models First we specifiedthe two branches immediately following the duplication asforeground branches in matrix A Branch-site models thatallow for positive selection in these foreground brancheswere not better at explaining the data than the nested sitemodels M1a and the null model A and indicate no evidencefor a burst of positive selection in the branches immediatelyfollowing the duplication event (Bielawski and Yang 2003)(data not shown) Second we treated all branches in eitherclade of paralogs as foreground branches When these twoclades are treated separately the model allowing for
Table 2Likelihood Ratio Tests Comparing Branch-Models (R1ndashR4)Site Models (M1ndashM8) and Branch-Site Models (Model AndashModel A Null)
Matrix Models Compared2(ln L1 ln L0)
Matrix A One ratio (R1) versus two ratio (R2) 18174Two ratio (R2) versus three ratio (R3) 0435Three ratio (R3) versus four ratio (R4) 1298Neutral (M1a) versus selection (M2a) 20566Beta (M7) versus beta amp w 1 (M8) 2683Model A versus M1a (Test 1) 874Model A versus model anull (Test 2) 252
Matrix B One ratio (R1) versus two ratio (R2) 2348Two ratio (R2) versus three ratio (R3) 001Three ratio (R3) versus four ratio (R4) 01Neutral (M1a) versus selection (M2a) 367Beta (M7) versus beta amp w 1 (M8) 7054
Significant at the 95 level significant at the 99 level
Table 3B Class MADS-Box Protein Interactions as Determined by Yeast Two-Hybrid Assays in Camellia japonica
CjDEF-BD CjTM6-BD CjGLO1-BD CjGLO2-BD BD
CjDEF-AD thorn mdash mdash mdash mdashCjTM6-AD mdash mdash thorn mdash mdashCjGLO1-AD thorn thorn mdash mdash mdashCjGLO2-AD thorn mdash mdash mdash mdashAD mdash mdash mdash mdash mdash
2638 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
positive selection is not significantly better in explaining thedata than the model M1a and null model A (data notshown) When the two clades are treated together as fore-ground branches model A is significantly better than thenested site model M1a (test 1) suggesting that positive se-lection is acting in both clades that arose after duplicationBayes empirical Bayes identification resulted in seven po-sitions though all with posterior probability below 90The improved test 2 from Zhang et al (2005) resulted ina value 252 just beneath the critical value 271 at the5 level but far above the critical value at the 10 level164 These two tests support the hypothesis that positiveselection acts after the duplication event in both paralogousgene clades
Together the identification of changes in selectivepressure in time and along the coding sequence indicatethat upon duplication PI paralogs underwent an increasednonsynonymous substitution rate and that this occurred
specifically in the domains shown to be necessary forproteinndashprotein interactions
Diversification in the Interaction Behavior of B-ClassGenes After the Ancient Duplication Event in BasalAsterids
As sites in the interaction domains of the PI genesfrom basal asterids were detected to be positively selectedafter duplication we further investigated the possible func-tional divergence of the duplicated PI proteins by yeast-2-hybrid analyses We tested the interactions between thefour class B genes in C japonica Camellia japonica con-tains two AP3-like genes one belonging to the euAP3-typeand one from the paleoAP3-type and two PI-like genesfrom an ancient duplication event at the base of the EricalesThis situation is similar to the B-class configuration in thecore asterid species Petunia hybrida (Vandenbussche
FIG 5mdashAlignment of the K-domain and C-terminal domain of the PI duplicates from matrix A used for the analysis in PAML Differentsubdomains and the characteristic PI motif are indicated (Kramer et al 1998) Codon positions identified to be under positive selection are marked withgray boxes
PISTILLATAmdashDuplications in Asterids 2639
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

Table 1List of Class B Homologues Included in This Study
Order Family Species PI AP3 TM6
Cornales Cornaceae Alangium platinifolium ApGLO ApDEF ApTM6Nyssaceae Nyssa sylvatica NsGLO1 NsGLO2 NsDEF NsTM6
Davidia involucrata DiGLO1 DiGLO2 DiTM6Loasaceae Loasa vulcanica LvGLO LvTM6Hydrangeaceae Hydrangea macrophylla HmPI HmAP3 HmTM6
Philadelphus pubescens PhpTM6Ericales Balsaminaceae Impatiens hawkeri IhGLO IhDEF1
IhDEF2Impatiens glandulifera IgGLO1 IgGLO2
Marcgraviaceae Marcgravia umbellata MuGLO MuDEFLecythidaceae Gustavia brasiliensis GbGLO1 GbGLO2 GbTM6
Napoleonaea vogelii NvGLO NvDEFPolemoniaceae Phlox paniculata PpGLO PpDEF PpTM6
Acanthogilia gloriosa AgGLOIpomopsis aggregata IaGLO
Sapotaceae Synsepalum dulcificum SydGLO SydDEFDiapensiaceae Galax urceolata GuGLO1 GuGLO2 GuDEF GuTM6Styracaceae Halesia diptera HdGLO1 HdGLO2 HdDEF
Styrax japonicus SjGLO SjDEF SjTM6Roridulaceae Roridula gorgonias RgGLOActinidiaceae Saurauia zahlbruckneri SzGLO1 SzGLO2
SzGLO3SzTM6
Actinidia chinensis AcGLO1 AcGLO2Clethraceae Clethra tomentosa CtGLO CtDEF CtTM6Theaceae Stewartia pseudocamellia SpGLO1 SpGLO2
SpGLO3SpDEF
Camellia japonica CjGLO1 CjGLO2 CjDEF CjTM6Symplocaceae Symplocos chinensis ScGLO1 ScGLO2 ScTM6Sarraceniaceae Sarracenia drummondii SdGLO SdDEF SdTM6Ericaceae Enkianthus campanulatus EcGLO
Erica hiemalis EhGLO1 EhGLO2 EhDEFEbenaceae Diospyros digyna DdGLO DdDEFPrimulaceae Primula denticulata PdGLO1 PdGLO2 PdDEFMyrsinaceae Cyclamen persicum CpGLO CpDEF
Hymenandra walichii HwGLO HwDEF HwTM6Theophrastaceae Jacquinia auriantaca JaGLO JaDEF JaTM6
Clavija latifolia ClGLO ClDEFMaesaceae Maesa argentea MaGLO MaDEF
Aquifoliales Aquifoliaceae Ilex aquifolium IaGLO1 IaGLO2 IxaAP3 IxaTM6Asterales Asteraceae Gerbera hybrida GGLO1 GDEF2 GDEF1
Lactuca sativa LsGLOTaraxacum officinale ToGLOCichorium intybus CiGLOSenecio vulgaris SvGLOHelianthus annuus HaPI HaAP3 HaTM6Centaurea solstitialis CsGLOHieracium piloselloides HpDEF1
HpDEF2Gentianales Rubiaceae Hedyotis terminalis HtGLO
Apocynaceae Catharanthus roseus CrGLOGentianaceae Eustoma grandiflorum EgGLO1 EgDEF
Solanales Solanaceae Petunia hybrida FBP1 PMADS2 PhDEF PhTM6Solanum lycopersicon SlGLO1 SlGLO2 LeAP3Ipomoea nil InGLONicotiana tabacum NtGLOCestrum elegans CeDEF CeTM6Nicotiana benthamiana NtTM6Solanum pseudolulo SopDEF SpTM6Brunfelsia uniflora BuDEF BuTM6
Lamiales Scrophulariaceae Mimulus guttatus MgGLOMimulus ringens MrDEFA
MrDEFBVerbenaceae Verbena hybrida VhGLOPlantaginaceae Antirrhinum majus GLO DEF
Misopates orontium MoGLO MoDEFChelone glabra CgDEF
Oleaceae Syringa vulgaris SvPI SvAP3Lamiaceae Salvia coccinea ScDEFA
ScDEFB
PISTILLATAmdashDuplications in Asterids 2629
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
plasmid inserts were sequenced using T7 and SP6 universalprimers using the BigDye Terminator 11 kit (Applied Bio-systems Foster City CA) on an Applied Biosystems 310 se-quencer or the plasmids were sent for sequencing (MacroGenInc Seoul South Korea) New sequences were deposited inGenbank (Accession numbers GQ141105ndashGQ141187)
Identification and Sequence Analysis
Obtained nucleotide sequences were translated intothe corresponding amino acid sequence and manuallyaligned using MacClade 4 (Maddison RP and MaddisonWP 2003) together with previously cloned and character-ized class B sequences from Genbank (table 1) After align-ment nucleotide sequences were analyzed with PAUP4b10 (Swofford 2002) MrBayes 312 (Huelsenbeck andRonquist 2001) and PHYML (Guindon and Gascuel2003) (PI matrix only) Paup 4b10 (Swofford 2002)was used for parsimony bootstrap analysis A MaximumParsimony (MP) heuristic search was conducted using1000 random addition sequences with tree bisection andreconnection branch swapping and saving of multiple par-simonous trees (MulTrees on) Branch support was calcu-lated by nonparametric bootstrap analysis on 1000pseudoreplicate data sets (Felsenstein 1985) Parametersfor the Bayesian analysis and maximum likelihood (ML)analysis were estimated using Modeltest 306 (Posadaand Crandall 2001) Modeltest selected the GTR (generaltime reversible) thorn I thorn G substitution model using theAkaike Information criterion MrBayes was run for 5 mil-lion generations where for every 100 generations one treewas saved For both matrices the search reached the sta-tionary phase around 50000 generations This numberwas considered the lsquolsquoburn-in periodrsquorsquo and was excludedwhen the consensus phylogeny was constructed PHYML(Guindon and Gascuel 2003) was used for ML inference ofthe PImatrix Confidence in the clades was estimated by theapproximate likelihood ratio test (aLRT) method from Ani-simova and Gascuel (2006) and traditional bootstrap anal-
ysis with 100 replicates For the aLRT tests we used boththe v2 statistic and the more conservative shimodeiva-Hasqawa (SH)-like test as branch support measures To fur-ther look into the supposed ancient PI duplication duringthe evolution of the Ericales clade we additionally con-structed a matrix with only the duplicated PI genes resultingfrom this duplication Cornales PI genes were used as out-groups as the reconstructed PI phylogeny gives us noclear answers on the exact timing of duplication A similaranalysis strategy was used but Modeltest selected theHasegawandashKishinondashYano (HKY) thorn G substitution modelfor the Bayesian and ML analyses
Semiquantitative RT-PCR
RNA was extracted from each organ type separatelyusing the Invisorb Spin Plant RNA kit (Invitek) and eachRNA sample was DNase treated using TURBO DNA-free(Ambion Austin TX) Total RNA was reverse transcribedinto cDNA using AMV reverse transcriptase (Promega) andthe included oligo-dT primer and random primers Gene-specific primers are listed in supplementary table S1 Sup-plementary Material online Expression of AP3 in Gustaviabrasiliensis and of TM6 in Cyclamen persicum Primulaacaulis Stewartia pseudocamellia andHalesia diptera wereexaminedwith primers fromAP3-like sequences from closelyrelated species Ribosomal 18S RNA was used as a control
In Situ Hybridization
Antisense probes were in vitro transcribed using T7RNApolymerase (NewEnglandBiolabs IpswichAustralia)in thepresence of digoxigenin-labeled uracil-5-triphosphate(Roche Basel Switzerland) from PCR-amplified templatesthat included a T7 promoter Tissues were fixed in 4 para-formaldehydeparaffinembeddedandsectionedat8lmSec-tions were mounted on Probe-On-Plus slides (FisherScientific Pittsburgh PA) Prehybridization hybridizationand detection essentially followed Carr and Irish (1997)
Table 1Continued
Order Family Species PI AP3 TM6
Ranunculales Ranunculaceae Aquilegia vulgaris AqvPITrautvetteria carolinensis TrcAP3
Vitales Vitaceae Vitis vinifera VvDEF VvTM6Saxifragales Grossulariacaea Ribes sanguineum
Saxifragaceae Saxifraga careyana SxcAP3 SxcTM6Malpighiales Salicaceae Populus deltoides
Populus tomentosa PtTM6Myrtales Myrtaceae Melaleuca quinquenerviaRosales Rosaceae Malus domestica MdTM6
Rosa rugosa RrDEF RrTM6Cucurbitales Cucurbitaceae Cucumis sativusFagales Betulaceae Betula pendula
Juglandaceae Juglans regia JrDEFTrochodendrales Trochodendraceae Trochodendron aralioides TraPI1 TroAP3Magnoliales Annonaceae Asimina longifolia AlAP3Dilleniales Dilleniaceae Dillenia indica DiiAP3 DiiTM6Asparagales Orchidaceae Phalaenopsis hybrid PhahAP3
Brassicaceae Arabidopsis thaliana AP3Brassica juncea BjAP3
Newly generated asterid sequences are marked in bold EST data are highlighted with an asterisk
2630 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Hybridizations in Sarracenia rubra and Stewartia rostratawere carried out using heterologous probes of the closely re-lated speciesSarracenia drummondii andS pseudocamellia
Codon Substitution Models
Omega (x or dNdS) is a measure of the amount of nat-ural selection acting on the protein To test whether positiveselection acted after the duplication event at the base of theEricales the previously obtained Bayesian topology of PIgenes was pruned in TreeEdit (indicated in fig 1) and thematching data sets were analyzed using PAML version314 (Yang 1997) We used two data matrices to investigate
selective pressure a matrix in which only the duplicatedgenes that originated after the duplication event at the baseof the Ericales are present (hereafter matrix A) and a secondmatrix in which the paralogous copies from species whohave lost either one of both paralogs are incorporated inmatrix A (hereafter matrix B) The log-likelihood of the se-quence alignments was calculated using different codonsubstitution models and likelihood ratio tests were usedto detect significant differences in fit between nested mod-els Branch models (Yang 1998 Yang and Nielsen 1998)were used to test for episodic adaptive evolution after du-plication either in all lineages combined or in paralogouslineages separately To test for the presence of positive
FIG 1mdashEvolutionary reconstruction of PI gene phylogeny in asterids as inferred from ML and Bayesian analysis Bayesian posterior probabilitiesand nonparametric bootstrap support values are indicated above branches SH-like and X2-like supports are shown below branches Gray circle 5
inferred duplication event Data sets used for assessments of selection pressures are indicated by stars black star 5 sequences included in matrix Awhite star 5 extra sequences added to matrix A to form Matrix B Marcgravia umbellata and Diospyros digyna PI sequences are underlined
PISTILLATAmdashDuplications in Asterids 2631
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
selection on sequence sites site models were applied thathold x constant among all branches but allow x to varyamong codons Site models include M1a M2a M7 andM8 (Nielsen and Yang 1998 Wong et al 2004 Yang et al2005) Positive selection is inferred when M2a and M8 arerespectively significantly better than M1a and M7 Branch-sitemodels allow thex ratio to vary amongboth sites and lin-eages (Zhang et al 2005) and can be used to detect positivelyselected sites in specific lineages within the tree We usedthe modified branch-site model A and compared this modelwith the nested site model M1a (Test 1) and the null modelA (Test 2) to test for positive selection (Zhang et al 2005)
Yeast Two-Hybrid Assays
The intervening- keratin-like and C-terminal domaindomains of the two PI (CjGLO1 and CjGLO2) the AP3(CjDEF) and TM6 genes (CjTM6) from Camellia japonicawere fusedwith theGAL4activationdomain in thepGAD424vector and the GAL4 DNA-binding domain in the pGBT9vector (ClontechMountainView CA) Each possible vectorcombination was transformed in yeast and two colonies pertransformation were used for B-galactosidase liquid assaysusing the protocol available at httpwwwfhcrcorgsciencelabsgottschlingyeast20ProtocolsBgalhtml
Results
In total 83 new B-lineage sequences were amplifiedfrom asterid species Forty-seven new PI and 36 new AP3-like sequences were identified from 31 and 25 asterid speciesrespectively (table 1) All but one of these species belong tothe basal asterid orders Cornales and Ericales In addition PIhomologues were amplified from I aquifolium (Aquifoliales)
Two Ancient and Frequent Recent Duplications HaveOccurred in the Asterid PI Lineage
For the phylogenetic analyses of the PI lineage wecombined the newly identified PI genes with previouslyidentified asterid PI sequences and EST data from Genbankto generate one data set Basal eudicot sequences were usedas outgroups for this data set and AqvPI (Aquilegia Ranun-culales) was used to root the PI tree A nucleotide alignmentof 74 taxa was analyzed using MP ML and Bayesian anal-yses The results are summarized in figure 1
The Bayesian topology which has relatively high pos-terior probabilities for a majority of nodes did not conflictwith the most likely tree from the ML analysis The MPanalysis however did not resolve the relationships betweenthe major clades and high bootstrap values were only ob-tained for nodes at lower taxonomical levels Similar to thebootstrap values from the MP analysis bootstrap supportfor the ML tree is low and we chose to only indicate thebootstrap values from the MP analysis on the phylogenyHowever to provide an additional measure of support ina likelihood framework we show SH-like and v2-like aLRTtest values These provide additional support for the Bayes-ian inferred relationships
The inferred PI phylogeny (fig 1) indicates that thisgene lineage probably underwent two ancient duplication
events during asterid evolution In both instances the twocopies were retained in some extant species lineageswhereas in other taxa one copy appears to have been lostor further duplicated An initial ancient duplication probablyoccurred early in the evolution of the core asterids becausethe Bayesian analysis supports two paralogous cladesa clade containing Gentianales Solanales and Asteralessequences (BPP 95) and one with Solanales and Lamialessequences (BPP 100) The latter is also strongly supportedbyMPbootstrapping (86) and aLRT testing (10099) Givenour current knowledge of asterid evolution (Albach et al2001 Bremer et al 2002) this suggests an ancient duplica-tion event before the origin of the two major core asteridclades (euasterids I and euasterids II) (Bayesian analysis)or at least before the origin of the euasterid II clade compris-ing Solanales Lamiales and Gentianales (MP analysis andaLRT testing) This duplication event was then followed byindependent losses in each paralogous lineageAlthoughourresults allow us to infer such a scenario no Garryales Dip-sacales or Apiales PI representatives are currently availablethat would further support such a hypothesis In addition thePI sequences of Aquifoliales an order situated at the base ofthe euasterid II clade occupy a nonsupported position at thebase of basal asterid PI sequences Together this suggeststhat an even denser sampling of asterid sequences would bedesirable to determine the exact moment of the PI duplica-tion in the core asterid lineage with even more confidence
The ML tree further suggests a second ancient dupli-cation event consistent with an origin early in the radiationof the order Ericales The PI sequences from Ericales rep-resentatives that are strongly supported by Bayesian support(BPP 96) and aLRT support (9999) are placed into two sep-arate clades A first clade is supported by a BPP value of 90and a v2-value of 92 The weakly supported second clade(BPP 67 aLRT 7088) consists of two strongly supportedclades (BPP 96 aLRT 9499 and BPP 90 aLRT 8999)where one (BPP 96) consists of the proposed basal lineagesof the Ericales the balsaminoıd clade and Lecythidaceae(Geuten et al 2004 Schonenberger et al 2005) The pres-ence of three strongly supported clades within the Ericalessuggests that the duplication in Ericales could have occurredeither at the base of the Ericales (as the ML phylogenyshows) or after the divergence of the basal lineages (balsami-noids and Lecythidaceae)Galax urceolata (Diapensiaceae)Hdiptera (Styracaceae)Saurauia zahlbruckneri (Actinidia-ceae) S pseudocamellia and C japonica (Theaceae) andSymplocos chinensis (Symplocaceae) all have multiple PIcopies belonging to two strongly supported clades due to thisancient duplication event Again this ancestral duplication isfollowed by different gene fateswith independent losses andduplications in each paralogous lineage Although furthersampling could lead to alternative explanations for the pro-posed multiple loss events there is some support for the hy-pothesis of independent gene loss in eachparalogous lineageSouthern blot analyses of Marcgravia umbellata (Geutenet al 2006) and Diospyros digyna (data not shown) both in-dicate that there is likely only one PI gene present in each ofthese species (fig 1 underlined) This observation supportsthe hypothesis of multiple PI gene losses within the Ericales
Because we wanted to explore the effect of omittingtaxa with single-copy genes of which the paralogous was
2632 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
putatively lost after duplication we constructed a secondmatrix with only duplicated PI genes due to the lsquolsquoancientrsquorsquoduplication from G urceolata (Diapensiaceae) H diptera(Styracaceae) S zahlbruckneri (Actinidiaceae) S pseudo-camellia and C japonica (Theaceae) and S chinensis(Symplocaceae) The MP analysis resulted in just one mostparsimonious tree that is identical to the tree with the high-est likelihood As in the previous analysis the same twoparalogous clades were identified In this case both BPPvalues (100 and 97) and ML bootstrap values (57 and53) were higher in this case supporting a duplication eventpresumably at least before the divergence of these familiesand after the divergence of the primuloid clade in Ericalesphylogeny (Schonenberger et al 2006)
In addition to these two ancient duplication events weidentified several other more recent duplication events inasterid evolution From I aquifolium (Aquifoliaceae) weamplified two PI copies but more extensive sampling ofAquifoliales representatives should reveal the timing of thisduplication more accurately In Cornales duplicate genesare found in both Davidia involucrata and Nyssa sylvatica(Nyssaceae) suggesting that the duplication that resulted inthese copies occurred in the lineage leading to NyssaceaeIn the clade of Ericales representatives there are seven spe-cies with more than one PI copy which appear to be morerecent than the previously described duplication event atthe base of the Ericales order For example we isolatedtwo PI homologues from Impatiens glandulifera (IgGLO1and IgGLO2 Balsaminaceae) A previously characterizedPI gene from Impatiens hawkeri (IhGLO Geuten et al2006) groups with IgGLO2 but not with the PI gene fromM umbellata (MuGLO Marcgraviaceae) suggesting thatthis duplication occurred at the base of the BalsaminaceaeSimilarly we isolated twoPI genes fromG brasiliensis (Le-cythidaceae) and the phylogeny indicates that one copy ismore related to the PI gene from another Lecythidaceae rep-resentativeNapoleonaea vogeliiBecauseGustavia is basalin the phylogeny of Lecythidaceae (Mori et al 2007) the du-plication leading to paralogous copies inGustavia probablyoccurred at the origin of the Lecythidaceae or earlier
We also identified two PI copies in Erica hiemalis(Ericaceae) and Actinidia chinensis (Actinidiaceae) In bothS pseudocamellia (Theaceae) and S zahlbruckneri (Acti-nidiaceae) a third PI copy is present presumably as a resultof a more recent duplication in one of their paralogs As thesequence similarity of the paralogs in these four species ishigh (91ndash98) it is hard to decide whether these genes areat the same genetic locus or at different loci We suggestthese genes to be paralogous copies with a very recent or-igin and not shared by any other species in our samplingFinally two PI genes were identified in P denticulata (Pri-mulaceae) Although sequence similarity among thesegenes is significantly lower (82) this duplication is notshared by any other PI genes in our analyses indicating thatthis duplication is specific to the Primulaceae
Duplications in the AP3 and TM6 Lineages Are Few andNot Correlated with PI Duplications
We also carried out phylogenetic analyses of asteridAP3 sequences using a number of newly identified AP3
sequences as well as previously characterized genes andEST sequences from Genbank (table 1) The phylogenywas inferred from a nucleotide alignment of genes from85 taxa which was analyzed using both MP and Bayesianmethods (fig 2) In the Bayesian analysis AlAP3 was usedto root the AP3-like tree Similar to previous analyses(Kramer and Irish 1999 Kramer et al 2006) our Bayesiananalysis recovered two highly supported clades A firstclade contains the euAP3 homologues (BPP 98) whereasa second clade contains the TM6 homologues (BPP 100)(fig 2) The duplication event that gave rise to these two eu-dicot AP3 gene clades is placed near the base of the core eu-dicots and likely occurred coincident with the origin ofTrochodendraceae and Buxaceae (Kramer et al 2006)Within core asterids our phylogeny gave no indicationsofmajor duplication events but onlyfiveof eight core asteridorderswere sampled Similarly in basal asterids duplicationevents have seemingly not played an important role duringAP3 evolution because the observed gene phylogeny did notconflict with the species phylogeny (Schonenberger et al2006) In several cases thoughparalogousAP3genesofveryrecent origin were identified in four species one from Eri-cales (I hawkeri Geuten et al 2006) and three from core as-terids (Hieracium piloselloides Mimulus cultstatus andSalvia coccinea) (Aagaard et al 2005) As for thePI phylog-eny the bootstrap values in the MP analyses were low andthese analyses resulted in a poorly resolved tree
After the basal eudicot and rosid representativesbranch off in the euAP3 clade (BPP 98) the Cornales rep-resentatives are sister to two large clades The first cladecontains the newly sampled Ericales representatives(BPP 81) whereas the second contains all core asterid se-quences (BPP 54) The unexpected association of Brassica-ceae representatives (AP3 BjAP3) with the euAP3 genesfrom Asterales was also present in previous analyses andprobably illustrates the need for even denser sampling(de Martino et al 2006 Kramer et al 2006) Within theEricales euAP3 clade (BPP 81) most relationships arepoorly resolved However a strongly supported primuloid(BPP 100) ericoid (BPP 91) and balsaminoid clade (BPP100) can be observed (Schonenberger et al 2005) Withinthe TM6-clade Ericales representatives are supported witha BPP of 99 but overall support within this clade is lowHowever we did not recover any paralogous copies of TM6genes in the species sampled suggesting that duplicationevents in this clade either do not occur readily or are se-lected against We also observed an unexpected associationof Asterales TM6 representatives with rosids (BPP 71) asreported in previous analyses (de Martino et al 2006Kramer et al 2006) The TM6 genes within the Solanalesare strongly supported as a clade (BPP 100)
RT-PCR Reveals Divergence of Expression PatternsAfter Duplication
To investigate whether gene duplicates have divergedin expression patterns or have seemingly remained underthe same regulatory control we used RT-PCR to monitorgene expression levels in mature dissected floral organs(fig 3) We limited our survey to Ericales species thatcontain representatives of PI genes arising from both the
PISTILLATAmdashDuplications in Asterids 2633
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
ancient and recent duplications We find that the expressionpattern of PI paralogs has diverged considerably and fairlyrapidly The PI protein is known to require an AP3-like in-teraction partner for its in vivo function in a number of coreeudicots (Jack et al 1992 Trobner et al 1992 Goto andMeyerowitz 1994) We therefore also examined the expres-sion patterns of TM6 and euAP3 genes in the same species
We compared Ericales expression patterns with the as-terid speciesPetunia hybrida (Solanaceae) BothPI genesin Petunia show a very similar expression pattern (Vanden-bussche et al 2004) Expression of these twoPI homologues(FBP1 and PMADS2) is mainly confined to the second andthirdwhorls (Angenent et al 1992Van derKrol et al 1993)Expression of PhDEF the AP3 homologue was mainly
FIG 2mdashEvolutionary reconstruction of APETALA3-like gene phylogeny in asterids as inferred from Bayesian analysis Bayesian posteriorprobabilities are indicated above branches nonparametric bootstrap support values are shown below branches Gray circle5 inferred duplication event
2634 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
confined to petals and stamens with low expression in firstand fourth whorl organs PhTM6 has a different expressionprofilewithstrongexpressioninstamensandcarpelswhereasexpression in sepals and petals is low (Vandenbussche et al2004) Similar expression patterns to Petunia of PI AP3 and
TM6 were detected in S lycopersicon another core asteridSolanaceae species (de Martino et al 2006)
Multiple copies of PI have been retained in the ge-nomes of S pseudocamellia C japonica and H dipteraafter the major duplication event at the base of the Ericales
FIG 3mdashRT-PCR expression data for B-class genes from selected Ericales representatives Sep 5 sepals pet 5 petals pet up 5 upper part ofpetals fus 5 fused part of petals and stamens sta 5 stamens sta up 5 upper part of stamens sta fus 5 fused part of stamens gyn 5 gynoecium andflo 5 flower
PISTILLATAmdashDuplications in Asterids 2635
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Both copies in each of these species appear to have divergedin their expression patterns after duplication (fig 3AndashC)Although HdGLO2 is expressed in the petals and stamensof H diptera the other copy (HdGLO1) is mainly ex-pressed in stamens and the gynoecium and absent in petalsIn comparison the TM6 and euAP3 lineage genes show un-expected expression patterns with strong expression ofHdTM6 in all disected organs but HdDEF expression beingmainly restricted to petals and carpels InC japonica one PIparalog (CjGLO2) is expressed throughout sepals petalsand stamens whereas expression of the second paralog(CjGLO1) appeared to be restricted to the nonfused partsof petals stamens and carpels Although both CjDEF andCjTM6 expression are detected in all organs strong expres-sion ofCjDEF is seenmainly in the sepals and petalsCjTM6expression is strongest in all stamen parts and in carpelsS pseudocamellia also displays differences in the expressionof its two PI copies One paralog (SpGLO2) is expressed inpetals stamens carpels and expression of the second paral-og (SpGLO1) is more restricted to the fused part of petals andstamens and upper part of petals The AP3 homologueSpDEF is strongly expressed in all stamen parts and carpelswhereas strong expression of SpTM6 is detected in sepals allstamen parts and carpels
In summary in these three cases there is one PI copywith a broad expression pattern in the flower (even in sepalsand carpels) whereas expression of the other PI copy ismore restricted to specific parts of petals and stamensThese expression patterns differ from those of PI lineagegenes in core asterid species (Vandenbussche et al2004 de Martino et al 2006) where PI expression (alsoin a duplicated configuration) is mainly restricted to the sec-ond and third whorl organs petals and stamens and nostrong expression in the first and fourth whorl organs is de-tected Interestingly the divergence in expression patternsin these species does not follow the gene phylogeny Theparalogous copies with a general expression pattern are notgrouped in one clade of PI genes in Ericales but rather arefound in two clades This suggests that the PI expressiondomains were not fixed immediately after duplication in-dicating that subfunctionalization in spatial expression doesnot explain retention of the duplicates Alternatively ifregulatory subfunctionalization was the main reason forretention evidence for this has been obscured throughtime Differences in expression between AP3 and TM6in C japonica seem to be very similar to members ofthe same lineages in the core asterid species Expressionof AP3-like genes in H diptera and S pseudocamelliahowever differs from those of Camellia with strongAP3 expression in the carpels of Stewartia and Halesiaand weak expression of AP3 in stamens of Halesia In eachof these species expression of one or both AP3-like genes isdetected in the sepals
Sarracenia drummondii (fig 3D) Styrax japonicus(fig 3E) C persicum (fig 3F) and Phlox paniculata(fig 3G) maintained probably only one of the paralogousPI copies in their genome after the ancient Ericales dupli-cation Moreover Sarracenia and Styrax maintained a dif-ferent copy (fig 1 BPP 68) than did Cyclamen and Phlox(fig 1 BPP89) We investigated the expression patterns ofPI genes in these species to monitor the possible divergence
between the two paralogous PI clades after duplication Ineach of these four species PI is expressed in petals and sta-mens similar to the expression patterns of PI in core aster-ids in previous studies (Vandenbussche et al 2004 deMartino et al 2006) In both Sarracenia (SdGLO) andPhlox (PpGLO) there is some expression detected in thecarpels whereas SjGLO (Styrax) is also expressed in sepalsNo clear divergence in expression of PI from the two dif-ferent clades (fig 1 BPP68 and BPP89) is detected in thesecond and third whorl organs The expression patterns ofAP3 and TM6 lineage genes are different between these fourspecies Although in S drummondii both AP3 and TM6 areexpressed in all four floral whorls the expression in Styraxis very similar to that of TM6 and AP3 in core asterid spe-cies with in addition to expression in petals and stamensexpression of TM6 in carpels and AP3 in sepals In Cycla-men TM6 expression is absent in the upper part of the sta-mens and strong expression of AP3 is detected in thecarpels In Phlox however TM6 expression is absent fromthe carpels
The multiple PI copies in G brasiliensis (Lecythida-ceae) and P acaulis (Primulaceae) are the result of a recentduplication probably at the base of these two families(fig 1) We looked at the expression pattern of each of theseduplicated genes and of the AP3-like genes in these speciesIn G brasiliensis GbGLO2 is expressed in petals and sta-mens whereas GbGLO1 is mainly expressed in the stami-nal parts and less in the petals (fig 3H) Expression of AP3and TM6 seems to be lsquolsquoinversersquorsquo compared with the ex-pected asterid expression with AP3 expression mainly re-stricted to the stamens and strong TM6 expression in thepetals No AP3-like expression is detected in the carpelsIn P acaulis there are differences between the expressionpatterns of both PI copies (fig 3I) PaGLO2 is expressed inall organs tested whereas the expression of PaGLO1 ismainly restricted to the sepals and petals Both AP3 andTM6 are strongly expressed in sepals and as inC persicumTM6-expression is weak in the staminal parts Even afterthese recent duplication events divergence in the expres-sion domain of the two PI copies is detected
In contrast to the duplicated PI genes of Petunia(Vandenbussche et al 2004) the expression domains ofPI genes have changed considerably after duplication inEricales For both ancient and several recent duplicationevents divergence in expression was detected indicatingthat the regulation of the duplicate copies has divergedIn the studied species where only one PI copy was isolatedit appears that an lsquolsquoancestralrsquorsquo expression pattern of PI isreestablished However a second copy could have beenmissed during amplification Also within the AP3-like lin-eage some derived expression patterns are detected
In Situ Hybridization Demonstrates ExpressionDivergence in Early Flower Development
To analyze the expression patterns of the PI genes afterduplication in early developmental stages we examined thespatio-temporal expression patterns of PI in selected Eri-cales species by in situ hybridization (fig 4) We selectedexamples of each type of fate after duplication retention
2636 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
of both ancient duplicates (Stewartia) retention of only oneof each PI paralog (C persicum and S rubra) and retentionof recent duplicates (P acaulis)
In S rostrata (fig 4AndashF) we could detect expressiondivergence between each of the PI copies at early stages offloral development At the stage when petal primordia areevident but stamen primordia are apparent only as smallmeristematic outgrowths SpGLO1 expression is detectedin petal and stamen primordia (fig 4A) whereas SpGLO2transcripts are also detected in the tip of the sepals (fig 4D)As the flower develops SpGLO2 expression is low butexpands to include developing carpels (fig 4E) and expres-sion is maintained in the carpels in later stages (fig 4F) Incontrast SpGLO1 expression is maintained at high levelsonly in petals and stamens through later stages (fig 4Band C) and does not expand into sepals and carpels Theseearly patterns of S rostrata PI gene expression are consis-tent with the later patterns of expression as detected byRT-PCR (fig 3I)
Expression patterns of PI homologues were furtherinvestigated in C persicum (fig 4GndashI) and S rubra(fig 4JndashK) CpGLO expression first appears in the youngpetal primordia (fig 4G) followed by expression in the sta-men primordia as they emerge (fig 4H) Expression inthese organs is maintained in later stages (fig 4I) In con-trast to the expression patterns in mature organs as docu-mented by RT-PCR weak expression in the developingsepals (fig 4H) and developing carpels (fig 4I) was de-tected For S rubra we were not able to obtain the earliestdevelopmental stages For later stages of floral develop-ment we observed strong expression of PI in petals andstamens (fig 4J) Weak expression of SdGLO was also de-tected in the developing carpels (fig 4K) which is consis-tent with the RT-PCR data (fig 3) These observationstogether with the RT-PCR data (fig 3) support the ideathat species with presumably only one PI copy in their ge-nome show expression that is more confined to petals andstamens
Finally we investigated the expression of recent PIduplicates in P acaulis (fig 4LndashQ) At early stages bothPaGLO1 (fig 4L) and PaGLO2 (fig 4O) are expressedacross the meristematic dome in sepal primordia commonpetalndashstamen primordia and carpel primordia Expressionof PaGLO2 is maintained in all organs at older stages(fig 4R and Q) whereas PaGLO1 expression is restrictedto the stamens (fig 4M and N)
The in situ expression data (fig 4) in comparison tothe RT-PCR data (fig 3) indicate that early patterns of ex-pression of PI genes are often developmentally regulatedwith different patterns of expression being observed early incomparison to later in flower development Nonethelessboth sets of data indicate that in species possessing dupli-cate PI genes divergence in expression patterns betweenthe two copies is a common theme
Positive Selection and Relaxed Purifying Selection ActsAfter the Duplication Event in Ericales in the K- and C-Cerminal Domain
The mRNA expression data indicate that duplicate PIparalogs have acquired different expression patterns sug-gesting independent evolution of their transcriptional reg-ulatory control It was also demonstrated that adaptiveevolution in the coding regions of MADS-box transcriptionfactors has played a role in the diversification of thisgene family (Martinez-Castilla and Alvarez-Buylla2003) Within class B genes Hernandez-Hernandez et al(2007) demonstrated the role of positive selection after crit-ical duplication events like the AP3-PI duplication and theeuAP3ndashTM6 duplication To investigate whether parallelchanges occurred in the coding regions of the genes exam-ined in this study we investigated the nonsynonymous ver-sus synonymous amino acid replacement rates forsignificant increases before and after duplication in thespecific paralogous clades and along the coding sequenceThe results of likelihood ratio tests are shown in table 2 and
FIG 4mdashIn situ hybridization data for PI genes in various developmental stages of Stewartia rostrata (AndashC SpGLO1 DndashF SpGLO2) Cyclamenpersicum (GndashI CpGLO) Sarracenia rubra (JndashK SdGLO) Primula acaulis (LndashN PaGLO1 OndashQ PaGLO2) lsquolsquoprsquorsquo indicates petalprimordia lsquolsquosrsquorsquoindicates stamen primordia
PISTILLATAmdashDuplications in Asterids 2637
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
specific sites are marked in the alignment presented infigure 5 Parameter estimates log-likelihood scores andBayes Empirical Bayes identified sites are listed in supple-mentary table S2 Supplementary Material online
Comparison of the likelihood scores calculated usingbranch models R1 and R2 for matrix A indicates a signifi-cantly increased evolutionary rate following the ancient du-plication event in Ericales The dNdS estimate from thetwo-ratio branch model after duplication (x 5 0285) istwice compared with the value before duplication (x 50113) illustrating the relaxed selective regime resultingfrom the duplication event However the same test appliedon matrix B did not reveal this change in selective pressureOne possible explanation is that by adding the single-copylineages that evolved after duplication the observed patternin matrix A for the duplicates is obscured In addition wefound no evidence that the two paralogous gene cladescould be evolving at different rates because the three-and four-ratio models did not result in a significantly in-creased log-likelihood as calculated for either of the datamatrices
To formally test for the presence of positive selectionon sequence sites we used the site models proposed byNielsen and Yang (1998) and Yang et al (2000) Analysisof matrix A shows that both M2a and M8 two models thatallow for positive selection fit the data significantly betterthan the corresponding neutral models (M1a and M7) Sub-stitution patterns in about 5 of the sites for the M2amodel and about 9 of the sites for the M8 model appearto result from adaptive evolution Here also analysis of ma-trix B suggests positively selected sites M8 fits the databetter than the neutral model M7 with an estimation of
10 sites under positive selection Bayes empirical Bayes(Yang et al 2005) identification of the sites under positiveselection in matrix A finds seven sites to be positively se-lected under model M8 two of these sites have a posteriorprobability higher than 99 Within matrix B only twopositively selected sites were detected which were alsoidentified in matrix A both with posterior probability high-er than 91 Interestingly all the identified sites are locatedwithin the K2-subdomain the interhelical region betweenK2 and K3 the K3-subdomain and the C-terminal domain(fig 5) These functional domains have been shown to beimportant for proteinndashprotein interactions among Arabi-dopsis MADS-domain proteins (Ma et al 1991 Pnueliet al 1991 Shore and Sharrocks 1995 Riechmann et al1996 Yang et al 2003) The strength of the AP3ndashPI inter-action is mediated primarily by K1 and K2 whereas thePIndashSEP interaction is mediated by K2 the amino-terminalportion of K3 and possibly the C-domain (Yang et al 2003Yang and Jack 2004) The C-terminal region normallyshows much lower sequence conservation but containsshort highly conserved motifs (Lamb and Irish 2003) Sev-eral studies have suggested that this domain is involvedin the formation of higher order protein complexes(Egea-Cortines et al 1999 Honma and Goto 2001) yetthe specific function of the class B C-terminal domainsis unclear (Lamb and Irish 2003 Piwarzyk et al 2007)None of the sites identified by Yang et al (2003) to be re-sponsible for the direct interaction between APETALA3and PISTILLATA are positively selected for in our dataset However site 49 contains a hydrophobic amino acidthat when mutated disrupts the alpha-helical secondarystructure (Yang et al 2003) In addition sites 49 and site60 in figure 5 were identified by Yang and Jack (2004) to becritical for PIndashSEP interaction Site 49 is situated withinthe K2 subdomain which is required for both AP3ndashPIand PIndashSEP interaction whereas site 60 which is locatedat the amino-terminal end of K3 affects when mutated thestrength of the PIndashSEP interaction
To identify evolutionary pressures acting on specificsites along certain branches and in clades we calculatedlikelihoods using the branch-site models First we specifiedthe two branches immediately following the duplication asforeground branches in matrix A Branch-site models thatallow for positive selection in these foreground brancheswere not better at explaining the data than the nested sitemodels M1a and the null model A and indicate no evidencefor a burst of positive selection in the branches immediatelyfollowing the duplication event (Bielawski and Yang 2003)(data not shown) Second we treated all branches in eitherclade of paralogs as foreground branches When these twoclades are treated separately the model allowing for
Table 2Likelihood Ratio Tests Comparing Branch-Models (R1ndashR4)Site Models (M1ndashM8) and Branch-Site Models (Model AndashModel A Null)
Matrix Models Compared2(ln L1 ln L0)
Matrix A One ratio (R1) versus two ratio (R2) 18174Two ratio (R2) versus three ratio (R3) 0435Three ratio (R3) versus four ratio (R4) 1298Neutral (M1a) versus selection (M2a) 20566Beta (M7) versus beta amp w 1 (M8) 2683Model A versus M1a (Test 1) 874Model A versus model anull (Test 2) 252
Matrix B One ratio (R1) versus two ratio (R2) 2348Two ratio (R2) versus three ratio (R3) 001Three ratio (R3) versus four ratio (R4) 01Neutral (M1a) versus selection (M2a) 367Beta (M7) versus beta amp w 1 (M8) 7054
Significant at the 95 level significant at the 99 level
Table 3B Class MADS-Box Protein Interactions as Determined by Yeast Two-Hybrid Assays in Camellia japonica
CjDEF-BD CjTM6-BD CjGLO1-BD CjGLO2-BD BD
CjDEF-AD thorn mdash mdash mdash mdashCjTM6-AD mdash mdash thorn mdash mdashCjGLO1-AD thorn thorn mdash mdash mdashCjGLO2-AD thorn mdash mdash mdash mdashAD mdash mdash mdash mdash mdash
2638 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
positive selection is not significantly better in explaining thedata than the model M1a and null model A (data notshown) When the two clades are treated together as fore-ground branches model A is significantly better than thenested site model M1a (test 1) suggesting that positive se-lection is acting in both clades that arose after duplicationBayes empirical Bayes identification resulted in seven po-sitions though all with posterior probability below 90The improved test 2 from Zhang et al (2005) resulted ina value 252 just beneath the critical value 271 at the5 level but far above the critical value at the 10 level164 These two tests support the hypothesis that positiveselection acts after the duplication event in both paralogousgene clades
Together the identification of changes in selectivepressure in time and along the coding sequence indicatethat upon duplication PI paralogs underwent an increasednonsynonymous substitution rate and that this occurred
specifically in the domains shown to be necessary forproteinndashprotein interactions
Diversification in the Interaction Behavior of B-ClassGenes After the Ancient Duplication Event in BasalAsterids
As sites in the interaction domains of the PI genesfrom basal asterids were detected to be positively selectedafter duplication we further investigated the possible func-tional divergence of the duplicated PI proteins by yeast-2-hybrid analyses We tested the interactions between thefour class B genes in C japonica Camellia japonica con-tains two AP3-like genes one belonging to the euAP3-typeand one from the paleoAP3-type and two PI-like genesfrom an ancient duplication event at the base of the EricalesThis situation is similar to the B-class configuration in thecore asterid species Petunia hybrida (Vandenbussche
FIG 5mdashAlignment of the K-domain and C-terminal domain of the PI duplicates from matrix A used for the analysis in PAML Differentsubdomains and the characteristic PI motif are indicated (Kramer et al 1998) Codon positions identified to be under positive selection are marked withgray boxes
PISTILLATAmdashDuplications in Asterids 2639
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

plasmid inserts were sequenced using T7 and SP6 universalprimers using the BigDye Terminator 11 kit (Applied Bio-systems Foster City CA) on an Applied Biosystems 310 se-quencer or the plasmids were sent for sequencing (MacroGenInc Seoul South Korea) New sequences were deposited inGenbank (Accession numbers GQ141105ndashGQ141187)
Identification and Sequence Analysis
Obtained nucleotide sequences were translated intothe corresponding amino acid sequence and manuallyaligned using MacClade 4 (Maddison RP and MaddisonWP 2003) together with previously cloned and character-ized class B sequences from Genbank (table 1) After align-ment nucleotide sequences were analyzed with PAUP4b10 (Swofford 2002) MrBayes 312 (Huelsenbeck andRonquist 2001) and PHYML (Guindon and Gascuel2003) (PI matrix only) Paup 4b10 (Swofford 2002)was used for parsimony bootstrap analysis A MaximumParsimony (MP) heuristic search was conducted using1000 random addition sequences with tree bisection andreconnection branch swapping and saving of multiple par-simonous trees (MulTrees on) Branch support was calcu-lated by nonparametric bootstrap analysis on 1000pseudoreplicate data sets (Felsenstein 1985) Parametersfor the Bayesian analysis and maximum likelihood (ML)analysis were estimated using Modeltest 306 (Posadaand Crandall 2001) Modeltest selected the GTR (generaltime reversible) thorn I thorn G substitution model using theAkaike Information criterion MrBayes was run for 5 mil-lion generations where for every 100 generations one treewas saved For both matrices the search reached the sta-tionary phase around 50000 generations This numberwas considered the lsquolsquoburn-in periodrsquorsquo and was excludedwhen the consensus phylogeny was constructed PHYML(Guindon and Gascuel 2003) was used for ML inference ofthe PImatrix Confidence in the clades was estimated by theapproximate likelihood ratio test (aLRT) method from Ani-simova and Gascuel (2006) and traditional bootstrap anal-
ysis with 100 replicates For the aLRT tests we used boththe v2 statistic and the more conservative shimodeiva-Hasqawa (SH)-like test as branch support measures To fur-ther look into the supposed ancient PI duplication duringthe evolution of the Ericales clade we additionally con-structed a matrix with only the duplicated PI genes resultingfrom this duplication Cornales PI genes were used as out-groups as the reconstructed PI phylogeny gives us noclear answers on the exact timing of duplication A similaranalysis strategy was used but Modeltest selected theHasegawandashKishinondashYano (HKY) thorn G substitution modelfor the Bayesian and ML analyses
Semiquantitative RT-PCR
RNA was extracted from each organ type separatelyusing the Invisorb Spin Plant RNA kit (Invitek) and eachRNA sample was DNase treated using TURBO DNA-free(Ambion Austin TX) Total RNA was reverse transcribedinto cDNA using AMV reverse transcriptase (Promega) andthe included oligo-dT primer and random primers Gene-specific primers are listed in supplementary table S1 Sup-plementary Material online Expression of AP3 in Gustaviabrasiliensis and of TM6 in Cyclamen persicum Primulaacaulis Stewartia pseudocamellia andHalesia diptera wereexaminedwith primers fromAP3-like sequences from closelyrelated species Ribosomal 18S RNA was used as a control
In Situ Hybridization
Antisense probes were in vitro transcribed using T7RNApolymerase (NewEnglandBiolabs IpswichAustralia)in thepresence of digoxigenin-labeled uracil-5-triphosphate(Roche Basel Switzerland) from PCR-amplified templatesthat included a T7 promoter Tissues were fixed in 4 para-formaldehydeparaffinembeddedandsectionedat8lmSec-tions were mounted on Probe-On-Plus slides (FisherScientific Pittsburgh PA) Prehybridization hybridizationand detection essentially followed Carr and Irish (1997)
Table 1Continued
Order Family Species PI AP3 TM6
Ranunculales Ranunculaceae Aquilegia vulgaris AqvPITrautvetteria carolinensis TrcAP3
Vitales Vitaceae Vitis vinifera VvDEF VvTM6Saxifragales Grossulariacaea Ribes sanguineum
Saxifragaceae Saxifraga careyana SxcAP3 SxcTM6Malpighiales Salicaceae Populus deltoides
Populus tomentosa PtTM6Myrtales Myrtaceae Melaleuca quinquenerviaRosales Rosaceae Malus domestica MdTM6
Rosa rugosa RrDEF RrTM6Cucurbitales Cucurbitaceae Cucumis sativusFagales Betulaceae Betula pendula
Juglandaceae Juglans regia JrDEFTrochodendrales Trochodendraceae Trochodendron aralioides TraPI1 TroAP3Magnoliales Annonaceae Asimina longifolia AlAP3Dilleniales Dilleniaceae Dillenia indica DiiAP3 DiiTM6Asparagales Orchidaceae Phalaenopsis hybrid PhahAP3
Brassicaceae Arabidopsis thaliana AP3Brassica juncea BjAP3
Newly generated asterid sequences are marked in bold EST data are highlighted with an asterisk
2630 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Hybridizations in Sarracenia rubra and Stewartia rostratawere carried out using heterologous probes of the closely re-lated speciesSarracenia drummondii andS pseudocamellia
Codon Substitution Models
Omega (x or dNdS) is a measure of the amount of nat-ural selection acting on the protein To test whether positiveselection acted after the duplication event at the base of theEricales the previously obtained Bayesian topology of PIgenes was pruned in TreeEdit (indicated in fig 1) and thematching data sets were analyzed using PAML version314 (Yang 1997) We used two data matrices to investigate
selective pressure a matrix in which only the duplicatedgenes that originated after the duplication event at the baseof the Ericales are present (hereafter matrix A) and a secondmatrix in which the paralogous copies from species whohave lost either one of both paralogs are incorporated inmatrix A (hereafter matrix B) The log-likelihood of the se-quence alignments was calculated using different codonsubstitution models and likelihood ratio tests were usedto detect significant differences in fit between nested mod-els Branch models (Yang 1998 Yang and Nielsen 1998)were used to test for episodic adaptive evolution after du-plication either in all lineages combined or in paralogouslineages separately To test for the presence of positive
FIG 1mdashEvolutionary reconstruction of PI gene phylogeny in asterids as inferred from ML and Bayesian analysis Bayesian posterior probabilitiesand nonparametric bootstrap support values are indicated above branches SH-like and X2-like supports are shown below branches Gray circle 5
inferred duplication event Data sets used for assessments of selection pressures are indicated by stars black star 5 sequences included in matrix Awhite star 5 extra sequences added to matrix A to form Matrix B Marcgravia umbellata and Diospyros digyna PI sequences are underlined
PISTILLATAmdashDuplications in Asterids 2631
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
selection on sequence sites site models were applied thathold x constant among all branches but allow x to varyamong codons Site models include M1a M2a M7 andM8 (Nielsen and Yang 1998 Wong et al 2004 Yang et al2005) Positive selection is inferred when M2a and M8 arerespectively significantly better than M1a and M7 Branch-sitemodels allow thex ratio to vary amongboth sites and lin-eages (Zhang et al 2005) and can be used to detect positivelyselected sites in specific lineages within the tree We usedthe modified branch-site model A and compared this modelwith the nested site model M1a (Test 1) and the null modelA (Test 2) to test for positive selection (Zhang et al 2005)
Yeast Two-Hybrid Assays
The intervening- keratin-like and C-terminal domaindomains of the two PI (CjGLO1 and CjGLO2) the AP3(CjDEF) and TM6 genes (CjTM6) from Camellia japonicawere fusedwith theGAL4activationdomain in thepGAD424vector and the GAL4 DNA-binding domain in the pGBT9vector (ClontechMountainView CA) Each possible vectorcombination was transformed in yeast and two colonies pertransformation were used for B-galactosidase liquid assaysusing the protocol available at httpwwwfhcrcorgsciencelabsgottschlingyeast20ProtocolsBgalhtml
Results
In total 83 new B-lineage sequences were amplifiedfrom asterid species Forty-seven new PI and 36 new AP3-like sequences were identified from 31 and 25 asterid speciesrespectively (table 1) All but one of these species belong tothe basal asterid orders Cornales and Ericales In addition PIhomologues were amplified from I aquifolium (Aquifoliales)
Two Ancient and Frequent Recent Duplications HaveOccurred in the Asterid PI Lineage
For the phylogenetic analyses of the PI lineage wecombined the newly identified PI genes with previouslyidentified asterid PI sequences and EST data from Genbankto generate one data set Basal eudicot sequences were usedas outgroups for this data set and AqvPI (Aquilegia Ranun-culales) was used to root the PI tree A nucleotide alignmentof 74 taxa was analyzed using MP ML and Bayesian anal-yses The results are summarized in figure 1
The Bayesian topology which has relatively high pos-terior probabilities for a majority of nodes did not conflictwith the most likely tree from the ML analysis The MPanalysis however did not resolve the relationships betweenthe major clades and high bootstrap values were only ob-tained for nodes at lower taxonomical levels Similar to thebootstrap values from the MP analysis bootstrap supportfor the ML tree is low and we chose to only indicate thebootstrap values from the MP analysis on the phylogenyHowever to provide an additional measure of support ina likelihood framework we show SH-like and v2-like aLRTtest values These provide additional support for the Bayes-ian inferred relationships
The inferred PI phylogeny (fig 1) indicates that thisgene lineage probably underwent two ancient duplication
events during asterid evolution In both instances the twocopies were retained in some extant species lineageswhereas in other taxa one copy appears to have been lostor further duplicated An initial ancient duplication probablyoccurred early in the evolution of the core asterids becausethe Bayesian analysis supports two paralogous cladesa clade containing Gentianales Solanales and Asteralessequences (BPP 95) and one with Solanales and Lamialessequences (BPP 100) The latter is also strongly supportedbyMPbootstrapping (86) and aLRT testing (10099) Givenour current knowledge of asterid evolution (Albach et al2001 Bremer et al 2002) this suggests an ancient duplica-tion event before the origin of the two major core asteridclades (euasterids I and euasterids II) (Bayesian analysis)or at least before the origin of the euasterid II clade compris-ing Solanales Lamiales and Gentianales (MP analysis andaLRT testing) This duplication event was then followed byindependent losses in each paralogous lineageAlthoughourresults allow us to infer such a scenario no Garryales Dip-sacales or Apiales PI representatives are currently availablethat would further support such a hypothesis In addition thePI sequences of Aquifoliales an order situated at the base ofthe euasterid II clade occupy a nonsupported position at thebase of basal asterid PI sequences Together this suggeststhat an even denser sampling of asterid sequences would bedesirable to determine the exact moment of the PI duplica-tion in the core asterid lineage with even more confidence
The ML tree further suggests a second ancient dupli-cation event consistent with an origin early in the radiationof the order Ericales The PI sequences from Ericales rep-resentatives that are strongly supported by Bayesian support(BPP 96) and aLRT support (9999) are placed into two sep-arate clades A first clade is supported by a BPP value of 90and a v2-value of 92 The weakly supported second clade(BPP 67 aLRT 7088) consists of two strongly supportedclades (BPP 96 aLRT 9499 and BPP 90 aLRT 8999)where one (BPP 96) consists of the proposed basal lineagesof the Ericales the balsaminoıd clade and Lecythidaceae(Geuten et al 2004 Schonenberger et al 2005) The pres-ence of three strongly supported clades within the Ericalessuggests that the duplication in Ericales could have occurredeither at the base of the Ericales (as the ML phylogenyshows) or after the divergence of the basal lineages (balsami-noids and Lecythidaceae)Galax urceolata (Diapensiaceae)Hdiptera (Styracaceae)Saurauia zahlbruckneri (Actinidia-ceae) S pseudocamellia and C japonica (Theaceae) andSymplocos chinensis (Symplocaceae) all have multiple PIcopies belonging to two strongly supported clades due to thisancient duplication event Again this ancestral duplication isfollowed by different gene fateswith independent losses andduplications in each paralogous lineage Although furthersampling could lead to alternative explanations for the pro-posed multiple loss events there is some support for the hy-pothesis of independent gene loss in eachparalogous lineageSouthern blot analyses of Marcgravia umbellata (Geutenet al 2006) and Diospyros digyna (data not shown) both in-dicate that there is likely only one PI gene present in each ofthese species (fig 1 underlined) This observation supportsthe hypothesis of multiple PI gene losses within the Ericales
Because we wanted to explore the effect of omittingtaxa with single-copy genes of which the paralogous was
2632 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
putatively lost after duplication we constructed a secondmatrix with only duplicated PI genes due to the lsquolsquoancientrsquorsquoduplication from G urceolata (Diapensiaceae) H diptera(Styracaceae) S zahlbruckneri (Actinidiaceae) S pseudo-camellia and C japonica (Theaceae) and S chinensis(Symplocaceae) The MP analysis resulted in just one mostparsimonious tree that is identical to the tree with the high-est likelihood As in the previous analysis the same twoparalogous clades were identified In this case both BPPvalues (100 and 97) and ML bootstrap values (57 and53) were higher in this case supporting a duplication eventpresumably at least before the divergence of these familiesand after the divergence of the primuloid clade in Ericalesphylogeny (Schonenberger et al 2006)
In addition to these two ancient duplication events weidentified several other more recent duplication events inasterid evolution From I aquifolium (Aquifoliaceae) weamplified two PI copies but more extensive sampling ofAquifoliales representatives should reveal the timing of thisduplication more accurately In Cornales duplicate genesare found in both Davidia involucrata and Nyssa sylvatica(Nyssaceae) suggesting that the duplication that resulted inthese copies occurred in the lineage leading to NyssaceaeIn the clade of Ericales representatives there are seven spe-cies with more than one PI copy which appear to be morerecent than the previously described duplication event atthe base of the Ericales order For example we isolatedtwo PI homologues from Impatiens glandulifera (IgGLO1and IgGLO2 Balsaminaceae) A previously characterizedPI gene from Impatiens hawkeri (IhGLO Geuten et al2006) groups with IgGLO2 but not with the PI gene fromM umbellata (MuGLO Marcgraviaceae) suggesting thatthis duplication occurred at the base of the BalsaminaceaeSimilarly we isolated twoPI genes fromG brasiliensis (Le-cythidaceae) and the phylogeny indicates that one copy ismore related to the PI gene from another Lecythidaceae rep-resentativeNapoleonaea vogeliiBecauseGustavia is basalin the phylogeny of Lecythidaceae (Mori et al 2007) the du-plication leading to paralogous copies inGustavia probablyoccurred at the origin of the Lecythidaceae or earlier
We also identified two PI copies in Erica hiemalis(Ericaceae) and Actinidia chinensis (Actinidiaceae) In bothS pseudocamellia (Theaceae) and S zahlbruckneri (Acti-nidiaceae) a third PI copy is present presumably as a resultof a more recent duplication in one of their paralogs As thesequence similarity of the paralogs in these four species ishigh (91ndash98) it is hard to decide whether these genes areat the same genetic locus or at different loci We suggestthese genes to be paralogous copies with a very recent or-igin and not shared by any other species in our samplingFinally two PI genes were identified in P denticulata (Pri-mulaceae) Although sequence similarity among thesegenes is significantly lower (82) this duplication is notshared by any other PI genes in our analyses indicating thatthis duplication is specific to the Primulaceae
Duplications in the AP3 and TM6 Lineages Are Few andNot Correlated with PI Duplications
We also carried out phylogenetic analyses of asteridAP3 sequences using a number of newly identified AP3
sequences as well as previously characterized genes andEST sequences from Genbank (table 1) The phylogenywas inferred from a nucleotide alignment of genes from85 taxa which was analyzed using both MP and Bayesianmethods (fig 2) In the Bayesian analysis AlAP3 was usedto root the AP3-like tree Similar to previous analyses(Kramer and Irish 1999 Kramer et al 2006) our Bayesiananalysis recovered two highly supported clades A firstclade contains the euAP3 homologues (BPP 98) whereasa second clade contains the TM6 homologues (BPP 100)(fig 2) The duplication event that gave rise to these two eu-dicot AP3 gene clades is placed near the base of the core eu-dicots and likely occurred coincident with the origin ofTrochodendraceae and Buxaceae (Kramer et al 2006)Within core asterids our phylogeny gave no indicationsofmajor duplication events but onlyfiveof eight core asteridorderswere sampled Similarly in basal asterids duplicationevents have seemingly not played an important role duringAP3 evolution because the observed gene phylogeny did notconflict with the species phylogeny (Schonenberger et al2006) In several cases thoughparalogousAP3genesofveryrecent origin were identified in four species one from Eri-cales (I hawkeri Geuten et al 2006) and three from core as-terids (Hieracium piloselloides Mimulus cultstatus andSalvia coccinea) (Aagaard et al 2005) As for thePI phylog-eny the bootstrap values in the MP analyses were low andthese analyses resulted in a poorly resolved tree
After the basal eudicot and rosid representativesbranch off in the euAP3 clade (BPP 98) the Cornales rep-resentatives are sister to two large clades The first cladecontains the newly sampled Ericales representatives(BPP 81) whereas the second contains all core asterid se-quences (BPP 54) The unexpected association of Brassica-ceae representatives (AP3 BjAP3) with the euAP3 genesfrom Asterales was also present in previous analyses andprobably illustrates the need for even denser sampling(de Martino et al 2006 Kramer et al 2006) Within theEricales euAP3 clade (BPP 81) most relationships arepoorly resolved However a strongly supported primuloid(BPP 100) ericoid (BPP 91) and balsaminoid clade (BPP100) can be observed (Schonenberger et al 2005) Withinthe TM6-clade Ericales representatives are supported witha BPP of 99 but overall support within this clade is lowHowever we did not recover any paralogous copies of TM6genes in the species sampled suggesting that duplicationevents in this clade either do not occur readily or are se-lected against We also observed an unexpected associationof Asterales TM6 representatives with rosids (BPP 71) asreported in previous analyses (de Martino et al 2006Kramer et al 2006) The TM6 genes within the Solanalesare strongly supported as a clade (BPP 100)
RT-PCR Reveals Divergence of Expression PatternsAfter Duplication
To investigate whether gene duplicates have divergedin expression patterns or have seemingly remained underthe same regulatory control we used RT-PCR to monitorgene expression levels in mature dissected floral organs(fig 3) We limited our survey to Ericales species thatcontain representatives of PI genes arising from both the
PISTILLATAmdashDuplications in Asterids 2633
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
ancient and recent duplications We find that the expressionpattern of PI paralogs has diverged considerably and fairlyrapidly The PI protein is known to require an AP3-like in-teraction partner for its in vivo function in a number of coreeudicots (Jack et al 1992 Trobner et al 1992 Goto andMeyerowitz 1994) We therefore also examined the expres-sion patterns of TM6 and euAP3 genes in the same species
We compared Ericales expression patterns with the as-terid speciesPetunia hybrida (Solanaceae) BothPI genesin Petunia show a very similar expression pattern (Vanden-bussche et al 2004) Expression of these twoPI homologues(FBP1 and PMADS2) is mainly confined to the second andthirdwhorls (Angenent et al 1992Van derKrol et al 1993)Expression of PhDEF the AP3 homologue was mainly
FIG 2mdashEvolutionary reconstruction of APETALA3-like gene phylogeny in asterids as inferred from Bayesian analysis Bayesian posteriorprobabilities are indicated above branches nonparametric bootstrap support values are shown below branches Gray circle5 inferred duplication event
2634 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
confined to petals and stamens with low expression in firstand fourth whorl organs PhTM6 has a different expressionprofilewithstrongexpressioninstamensandcarpelswhereasexpression in sepals and petals is low (Vandenbussche et al2004) Similar expression patterns to Petunia of PI AP3 and
TM6 were detected in S lycopersicon another core asteridSolanaceae species (de Martino et al 2006)
Multiple copies of PI have been retained in the ge-nomes of S pseudocamellia C japonica and H dipteraafter the major duplication event at the base of the Ericales
FIG 3mdashRT-PCR expression data for B-class genes from selected Ericales representatives Sep 5 sepals pet 5 petals pet up 5 upper part ofpetals fus 5 fused part of petals and stamens sta 5 stamens sta up 5 upper part of stamens sta fus 5 fused part of stamens gyn 5 gynoecium andflo 5 flower
PISTILLATAmdashDuplications in Asterids 2635
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Both copies in each of these species appear to have divergedin their expression patterns after duplication (fig 3AndashC)Although HdGLO2 is expressed in the petals and stamensof H diptera the other copy (HdGLO1) is mainly ex-pressed in stamens and the gynoecium and absent in petalsIn comparison the TM6 and euAP3 lineage genes show un-expected expression patterns with strong expression ofHdTM6 in all disected organs but HdDEF expression beingmainly restricted to petals and carpels InC japonica one PIparalog (CjGLO2) is expressed throughout sepals petalsand stamens whereas expression of the second paralog(CjGLO1) appeared to be restricted to the nonfused partsof petals stamens and carpels Although both CjDEF andCjTM6 expression are detected in all organs strong expres-sion ofCjDEF is seenmainly in the sepals and petalsCjTM6expression is strongest in all stamen parts and in carpelsS pseudocamellia also displays differences in the expressionof its two PI copies One paralog (SpGLO2) is expressed inpetals stamens carpels and expression of the second paral-og (SpGLO1) is more restricted to the fused part of petals andstamens and upper part of petals The AP3 homologueSpDEF is strongly expressed in all stamen parts and carpelswhereas strong expression of SpTM6 is detected in sepals allstamen parts and carpels
In summary in these three cases there is one PI copywith a broad expression pattern in the flower (even in sepalsand carpels) whereas expression of the other PI copy ismore restricted to specific parts of petals and stamensThese expression patterns differ from those of PI lineagegenes in core asterid species (Vandenbussche et al2004 de Martino et al 2006) where PI expression (alsoin a duplicated configuration) is mainly restricted to the sec-ond and third whorl organs petals and stamens and nostrong expression in the first and fourth whorl organs is de-tected Interestingly the divergence in expression patternsin these species does not follow the gene phylogeny Theparalogous copies with a general expression pattern are notgrouped in one clade of PI genes in Ericales but rather arefound in two clades This suggests that the PI expressiondomains were not fixed immediately after duplication in-dicating that subfunctionalization in spatial expression doesnot explain retention of the duplicates Alternatively ifregulatory subfunctionalization was the main reason forretention evidence for this has been obscured throughtime Differences in expression between AP3 and TM6in C japonica seem to be very similar to members ofthe same lineages in the core asterid species Expressionof AP3-like genes in H diptera and S pseudocamelliahowever differs from those of Camellia with strongAP3 expression in the carpels of Stewartia and Halesiaand weak expression of AP3 in stamens of Halesia In eachof these species expression of one or both AP3-like genes isdetected in the sepals
Sarracenia drummondii (fig 3D) Styrax japonicus(fig 3E) C persicum (fig 3F) and Phlox paniculata(fig 3G) maintained probably only one of the paralogousPI copies in their genome after the ancient Ericales dupli-cation Moreover Sarracenia and Styrax maintained a dif-ferent copy (fig 1 BPP 68) than did Cyclamen and Phlox(fig 1 BPP89) We investigated the expression patterns ofPI genes in these species to monitor the possible divergence
between the two paralogous PI clades after duplication Ineach of these four species PI is expressed in petals and sta-mens similar to the expression patterns of PI in core aster-ids in previous studies (Vandenbussche et al 2004 deMartino et al 2006) In both Sarracenia (SdGLO) andPhlox (PpGLO) there is some expression detected in thecarpels whereas SjGLO (Styrax) is also expressed in sepalsNo clear divergence in expression of PI from the two dif-ferent clades (fig 1 BPP68 and BPP89) is detected in thesecond and third whorl organs The expression patterns ofAP3 and TM6 lineage genes are different between these fourspecies Although in S drummondii both AP3 and TM6 areexpressed in all four floral whorls the expression in Styraxis very similar to that of TM6 and AP3 in core asterid spe-cies with in addition to expression in petals and stamensexpression of TM6 in carpels and AP3 in sepals In Cycla-men TM6 expression is absent in the upper part of the sta-mens and strong expression of AP3 is detected in thecarpels In Phlox however TM6 expression is absent fromthe carpels
The multiple PI copies in G brasiliensis (Lecythida-ceae) and P acaulis (Primulaceae) are the result of a recentduplication probably at the base of these two families(fig 1) We looked at the expression pattern of each of theseduplicated genes and of the AP3-like genes in these speciesIn G brasiliensis GbGLO2 is expressed in petals and sta-mens whereas GbGLO1 is mainly expressed in the stami-nal parts and less in the petals (fig 3H) Expression of AP3and TM6 seems to be lsquolsquoinversersquorsquo compared with the ex-pected asterid expression with AP3 expression mainly re-stricted to the stamens and strong TM6 expression in thepetals No AP3-like expression is detected in the carpelsIn P acaulis there are differences between the expressionpatterns of both PI copies (fig 3I) PaGLO2 is expressed inall organs tested whereas the expression of PaGLO1 ismainly restricted to the sepals and petals Both AP3 andTM6 are strongly expressed in sepals and as inC persicumTM6-expression is weak in the staminal parts Even afterthese recent duplication events divergence in the expres-sion domain of the two PI copies is detected
In contrast to the duplicated PI genes of Petunia(Vandenbussche et al 2004) the expression domains ofPI genes have changed considerably after duplication inEricales For both ancient and several recent duplicationevents divergence in expression was detected indicatingthat the regulation of the duplicate copies has divergedIn the studied species where only one PI copy was isolatedit appears that an lsquolsquoancestralrsquorsquo expression pattern of PI isreestablished However a second copy could have beenmissed during amplification Also within the AP3-like lin-eage some derived expression patterns are detected
In Situ Hybridization Demonstrates ExpressionDivergence in Early Flower Development
To analyze the expression patterns of the PI genes afterduplication in early developmental stages we examined thespatio-temporal expression patterns of PI in selected Eri-cales species by in situ hybridization (fig 4) We selectedexamples of each type of fate after duplication retention
2636 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
of both ancient duplicates (Stewartia) retention of only oneof each PI paralog (C persicum and S rubra) and retentionof recent duplicates (P acaulis)
In S rostrata (fig 4AndashF) we could detect expressiondivergence between each of the PI copies at early stages offloral development At the stage when petal primordia areevident but stamen primordia are apparent only as smallmeristematic outgrowths SpGLO1 expression is detectedin petal and stamen primordia (fig 4A) whereas SpGLO2transcripts are also detected in the tip of the sepals (fig 4D)As the flower develops SpGLO2 expression is low butexpands to include developing carpels (fig 4E) and expres-sion is maintained in the carpels in later stages (fig 4F) Incontrast SpGLO1 expression is maintained at high levelsonly in petals and stamens through later stages (fig 4Band C) and does not expand into sepals and carpels Theseearly patterns of S rostrata PI gene expression are consis-tent with the later patterns of expression as detected byRT-PCR (fig 3I)
Expression patterns of PI homologues were furtherinvestigated in C persicum (fig 4GndashI) and S rubra(fig 4JndashK) CpGLO expression first appears in the youngpetal primordia (fig 4G) followed by expression in the sta-men primordia as they emerge (fig 4H) Expression inthese organs is maintained in later stages (fig 4I) In con-trast to the expression patterns in mature organs as docu-mented by RT-PCR weak expression in the developingsepals (fig 4H) and developing carpels (fig 4I) was de-tected For S rubra we were not able to obtain the earliestdevelopmental stages For later stages of floral develop-ment we observed strong expression of PI in petals andstamens (fig 4J) Weak expression of SdGLO was also de-tected in the developing carpels (fig 4K) which is consis-tent with the RT-PCR data (fig 3) These observationstogether with the RT-PCR data (fig 3) support the ideathat species with presumably only one PI copy in their ge-nome show expression that is more confined to petals andstamens
Finally we investigated the expression of recent PIduplicates in P acaulis (fig 4LndashQ) At early stages bothPaGLO1 (fig 4L) and PaGLO2 (fig 4O) are expressedacross the meristematic dome in sepal primordia commonpetalndashstamen primordia and carpel primordia Expressionof PaGLO2 is maintained in all organs at older stages(fig 4R and Q) whereas PaGLO1 expression is restrictedto the stamens (fig 4M and N)
The in situ expression data (fig 4) in comparison tothe RT-PCR data (fig 3) indicate that early patterns of ex-pression of PI genes are often developmentally regulatedwith different patterns of expression being observed early incomparison to later in flower development Nonethelessboth sets of data indicate that in species possessing dupli-cate PI genes divergence in expression patterns betweenthe two copies is a common theme
Positive Selection and Relaxed Purifying Selection ActsAfter the Duplication Event in Ericales in the K- and C-Cerminal Domain
The mRNA expression data indicate that duplicate PIparalogs have acquired different expression patterns sug-gesting independent evolution of their transcriptional reg-ulatory control It was also demonstrated that adaptiveevolution in the coding regions of MADS-box transcriptionfactors has played a role in the diversification of thisgene family (Martinez-Castilla and Alvarez-Buylla2003) Within class B genes Hernandez-Hernandez et al(2007) demonstrated the role of positive selection after crit-ical duplication events like the AP3-PI duplication and theeuAP3ndashTM6 duplication To investigate whether parallelchanges occurred in the coding regions of the genes exam-ined in this study we investigated the nonsynonymous ver-sus synonymous amino acid replacement rates forsignificant increases before and after duplication in thespecific paralogous clades and along the coding sequenceThe results of likelihood ratio tests are shown in table 2 and
FIG 4mdashIn situ hybridization data for PI genes in various developmental stages of Stewartia rostrata (AndashC SpGLO1 DndashF SpGLO2) Cyclamenpersicum (GndashI CpGLO) Sarracenia rubra (JndashK SdGLO) Primula acaulis (LndashN PaGLO1 OndashQ PaGLO2) lsquolsquoprsquorsquo indicates petalprimordia lsquolsquosrsquorsquoindicates stamen primordia
PISTILLATAmdashDuplications in Asterids 2637
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
specific sites are marked in the alignment presented infigure 5 Parameter estimates log-likelihood scores andBayes Empirical Bayes identified sites are listed in supple-mentary table S2 Supplementary Material online
Comparison of the likelihood scores calculated usingbranch models R1 and R2 for matrix A indicates a signifi-cantly increased evolutionary rate following the ancient du-plication event in Ericales The dNdS estimate from thetwo-ratio branch model after duplication (x 5 0285) istwice compared with the value before duplication (x 50113) illustrating the relaxed selective regime resultingfrom the duplication event However the same test appliedon matrix B did not reveal this change in selective pressureOne possible explanation is that by adding the single-copylineages that evolved after duplication the observed patternin matrix A for the duplicates is obscured In addition wefound no evidence that the two paralogous gene cladescould be evolving at different rates because the three-and four-ratio models did not result in a significantly in-creased log-likelihood as calculated for either of the datamatrices
To formally test for the presence of positive selectionon sequence sites we used the site models proposed byNielsen and Yang (1998) and Yang et al (2000) Analysisof matrix A shows that both M2a and M8 two models thatallow for positive selection fit the data significantly betterthan the corresponding neutral models (M1a and M7) Sub-stitution patterns in about 5 of the sites for the M2amodel and about 9 of the sites for the M8 model appearto result from adaptive evolution Here also analysis of ma-trix B suggests positively selected sites M8 fits the databetter than the neutral model M7 with an estimation of
10 sites under positive selection Bayes empirical Bayes(Yang et al 2005) identification of the sites under positiveselection in matrix A finds seven sites to be positively se-lected under model M8 two of these sites have a posteriorprobability higher than 99 Within matrix B only twopositively selected sites were detected which were alsoidentified in matrix A both with posterior probability high-er than 91 Interestingly all the identified sites are locatedwithin the K2-subdomain the interhelical region betweenK2 and K3 the K3-subdomain and the C-terminal domain(fig 5) These functional domains have been shown to beimportant for proteinndashprotein interactions among Arabi-dopsis MADS-domain proteins (Ma et al 1991 Pnueliet al 1991 Shore and Sharrocks 1995 Riechmann et al1996 Yang et al 2003) The strength of the AP3ndashPI inter-action is mediated primarily by K1 and K2 whereas thePIndashSEP interaction is mediated by K2 the amino-terminalportion of K3 and possibly the C-domain (Yang et al 2003Yang and Jack 2004) The C-terminal region normallyshows much lower sequence conservation but containsshort highly conserved motifs (Lamb and Irish 2003) Sev-eral studies have suggested that this domain is involvedin the formation of higher order protein complexes(Egea-Cortines et al 1999 Honma and Goto 2001) yetthe specific function of the class B C-terminal domainsis unclear (Lamb and Irish 2003 Piwarzyk et al 2007)None of the sites identified by Yang et al (2003) to be re-sponsible for the direct interaction between APETALA3and PISTILLATA are positively selected for in our dataset However site 49 contains a hydrophobic amino acidthat when mutated disrupts the alpha-helical secondarystructure (Yang et al 2003) In addition sites 49 and site60 in figure 5 were identified by Yang and Jack (2004) to becritical for PIndashSEP interaction Site 49 is situated withinthe K2 subdomain which is required for both AP3ndashPIand PIndashSEP interaction whereas site 60 which is locatedat the amino-terminal end of K3 affects when mutated thestrength of the PIndashSEP interaction
To identify evolutionary pressures acting on specificsites along certain branches and in clades we calculatedlikelihoods using the branch-site models First we specifiedthe two branches immediately following the duplication asforeground branches in matrix A Branch-site models thatallow for positive selection in these foreground brancheswere not better at explaining the data than the nested sitemodels M1a and the null model A and indicate no evidencefor a burst of positive selection in the branches immediatelyfollowing the duplication event (Bielawski and Yang 2003)(data not shown) Second we treated all branches in eitherclade of paralogs as foreground branches When these twoclades are treated separately the model allowing for
Table 2Likelihood Ratio Tests Comparing Branch-Models (R1ndashR4)Site Models (M1ndashM8) and Branch-Site Models (Model AndashModel A Null)
Matrix Models Compared2(ln L1 ln L0)
Matrix A One ratio (R1) versus two ratio (R2) 18174Two ratio (R2) versus three ratio (R3) 0435Three ratio (R3) versus four ratio (R4) 1298Neutral (M1a) versus selection (M2a) 20566Beta (M7) versus beta amp w 1 (M8) 2683Model A versus M1a (Test 1) 874Model A versus model anull (Test 2) 252
Matrix B One ratio (R1) versus two ratio (R2) 2348Two ratio (R2) versus three ratio (R3) 001Three ratio (R3) versus four ratio (R4) 01Neutral (M1a) versus selection (M2a) 367Beta (M7) versus beta amp w 1 (M8) 7054
Significant at the 95 level significant at the 99 level
Table 3B Class MADS-Box Protein Interactions as Determined by Yeast Two-Hybrid Assays in Camellia japonica
CjDEF-BD CjTM6-BD CjGLO1-BD CjGLO2-BD BD
CjDEF-AD thorn mdash mdash mdash mdashCjTM6-AD mdash mdash thorn mdash mdashCjGLO1-AD thorn thorn mdash mdash mdashCjGLO2-AD thorn mdash mdash mdash mdashAD mdash mdash mdash mdash mdash
2638 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
positive selection is not significantly better in explaining thedata than the model M1a and null model A (data notshown) When the two clades are treated together as fore-ground branches model A is significantly better than thenested site model M1a (test 1) suggesting that positive se-lection is acting in both clades that arose after duplicationBayes empirical Bayes identification resulted in seven po-sitions though all with posterior probability below 90The improved test 2 from Zhang et al (2005) resulted ina value 252 just beneath the critical value 271 at the5 level but far above the critical value at the 10 level164 These two tests support the hypothesis that positiveselection acts after the duplication event in both paralogousgene clades
Together the identification of changes in selectivepressure in time and along the coding sequence indicatethat upon duplication PI paralogs underwent an increasednonsynonymous substitution rate and that this occurred
specifically in the domains shown to be necessary forproteinndashprotein interactions
Diversification in the Interaction Behavior of B-ClassGenes After the Ancient Duplication Event in BasalAsterids
As sites in the interaction domains of the PI genesfrom basal asterids were detected to be positively selectedafter duplication we further investigated the possible func-tional divergence of the duplicated PI proteins by yeast-2-hybrid analyses We tested the interactions between thefour class B genes in C japonica Camellia japonica con-tains two AP3-like genes one belonging to the euAP3-typeand one from the paleoAP3-type and two PI-like genesfrom an ancient duplication event at the base of the EricalesThis situation is similar to the B-class configuration in thecore asterid species Petunia hybrida (Vandenbussche
FIG 5mdashAlignment of the K-domain and C-terminal domain of the PI duplicates from matrix A used for the analysis in PAML Differentsubdomains and the characteristic PI motif are indicated (Kramer et al 1998) Codon positions identified to be under positive selection are marked withgray boxes
PISTILLATAmdashDuplications in Asterids 2639
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

Hybridizations in Sarracenia rubra and Stewartia rostratawere carried out using heterologous probes of the closely re-lated speciesSarracenia drummondii andS pseudocamellia
Codon Substitution Models
Omega (x or dNdS) is a measure of the amount of nat-ural selection acting on the protein To test whether positiveselection acted after the duplication event at the base of theEricales the previously obtained Bayesian topology of PIgenes was pruned in TreeEdit (indicated in fig 1) and thematching data sets were analyzed using PAML version314 (Yang 1997) We used two data matrices to investigate
selective pressure a matrix in which only the duplicatedgenes that originated after the duplication event at the baseof the Ericales are present (hereafter matrix A) and a secondmatrix in which the paralogous copies from species whohave lost either one of both paralogs are incorporated inmatrix A (hereafter matrix B) The log-likelihood of the se-quence alignments was calculated using different codonsubstitution models and likelihood ratio tests were usedto detect significant differences in fit between nested mod-els Branch models (Yang 1998 Yang and Nielsen 1998)were used to test for episodic adaptive evolution after du-plication either in all lineages combined or in paralogouslineages separately To test for the presence of positive
FIG 1mdashEvolutionary reconstruction of PI gene phylogeny in asterids as inferred from ML and Bayesian analysis Bayesian posterior probabilitiesand nonparametric bootstrap support values are indicated above branches SH-like and X2-like supports are shown below branches Gray circle 5
inferred duplication event Data sets used for assessments of selection pressures are indicated by stars black star 5 sequences included in matrix Awhite star 5 extra sequences added to matrix A to form Matrix B Marcgravia umbellata and Diospyros digyna PI sequences are underlined
PISTILLATAmdashDuplications in Asterids 2631
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
selection on sequence sites site models were applied thathold x constant among all branches but allow x to varyamong codons Site models include M1a M2a M7 andM8 (Nielsen and Yang 1998 Wong et al 2004 Yang et al2005) Positive selection is inferred when M2a and M8 arerespectively significantly better than M1a and M7 Branch-sitemodels allow thex ratio to vary amongboth sites and lin-eages (Zhang et al 2005) and can be used to detect positivelyselected sites in specific lineages within the tree We usedthe modified branch-site model A and compared this modelwith the nested site model M1a (Test 1) and the null modelA (Test 2) to test for positive selection (Zhang et al 2005)
Yeast Two-Hybrid Assays
The intervening- keratin-like and C-terminal domaindomains of the two PI (CjGLO1 and CjGLO2) the AP3(CjDEF) and TM6 genes (CjTM6) from Camellia japonicawere fusedwith theGAL4activationdomain in thepGAD424vector and the GAL4 DNA-binding domain in the pGBT9vector (ClontechMountainView CA) Each possible vectorcombination was transformed in yeast and two colonies pertransformation were used for B-galactosidase liquid assaysusing the protocol available at httpwwwfhcrcorgsciencelabsgottschlingyeast20ProtocolsBgalhtml
Results
In total 83 new B-lineage sequences were amplifiedfrom asterid species Forty-seven new PI and 36 new AP3-like sequences were identified from 31 and 25 asterid speciesrespectively (table 1) All but one of these species belong tothe basal asterid orders Cornales and Ericales In addition PIhomologues were amplified from I aquifolium (Aquifoliales)
Two Ancient and Frequent Recent Duplications HaveOccurred in the Asterid PI Lineage
For the phylogenetic analyses of the PI lineage wecombined the newly identified PI genes with previouslyidentified asterid PI sequences and EST data from Genbankto generate one data set Basal eudicot sequences were usedas outgroups for this data set and AqvPI (Aquilegia Ranun-culales) was used to root the PI tree A nucleotide alignmentof 74 taxa was analyzed using MP ML and Bayesian anal-yses The results are summarized in figure 1
The Bayesian topology which has relatively high pos-terior probabilities for a majority of nodes did not conflictwith the most likely tree from the ML analysis The MPanalysis however did not resolve the relationships betweenthe major clades and high bootstrap values were only ob-tained for nodes at lower taxonomical levels Similar to thebootstrap values from the MP analysis bootstrap supportfor the ML tree is low and we chose to only indicate thebootstrap values from the MP analysis on the phylogenyHowever to provide an additional measure of support ina likelihood framework we show SH-like and v2-like aLRTtest values These provide additional support for the Bayes-ian inferred relationships
The inferred PI phylogeny (fig 1) indicates that thisgene lineage probably underwent two ancient duplication
events during asterid evolution In both instances the twocopies were retained in some extant species lineageswhereas in other taxa one copy appears to have been lostor further duplicated An initial ancient duplication probablyoccurred early in the evolution of the core asterids becausethe Bayesian analysis supports two paralogous cladesa clade containing Gentianales Solanales and Asteralessequences (BPP 95) and one with Solanales and Lamialessequences (BPP 100) The latter is also strongly supportedbyMPbootstrapping (86) and aLRT testing (10099) Givenour current knowledge of asterid evolution (Albach et al2001 Bremer et al 2002) this suggests an ancient duplica-tion event before the origin of the two major core asteridclades (euasterids I and euasterids II) (Bayesian analysis)or at least before the origin of the euasterid II clade compris-ing Solanales Lamiales and Gentianales (MP analysis andaLRT testing) This duplication event was then followed byindependent losses in each paralogous lineageAlthoughourresults allow us to infer such a scenario no Garryales Dip-sacales or Apiales PI representatives are currently availablethat would further support such a hypothesis In addition thePI sequences of Aquifoliales an order situated at the base ofthe euasterid II clade occupy a nonsupported position at thebase of basal asterid PI sequences Together this suggeststhat an even denser sampling of asterid sequences would bedesirable to determine the exact moment of the PI duplica-tion in the core asterid lineage with even more confidence
The ML tree further suggests a second ancient dupli-cation event consistent with an origin early in the radiationof the order Ericales The PI sequences from Ericales rep-resentatives that are strongly supported by Bayesian support(BPP 96) and aLRT support (9999) are placed into two sep-arate clades A first clade is supported by a BPP value of 90and a v2-value of 92 The weakly supported second clade(BPP 67 aLRT 7088) consists of two strongly supportedclades (BPP 96 aLRT 9499 and BPP 90 aLRT 8999)where one (BPP 96) consists of the proposed basal lineagesof the Ericales the balsaminoıd clade and Lecythidaceae(Geuten et al 2004 Schonenberger et al 2005) The pres-ence of three strongly supported clades within the Ericalessuggests that the duplication in Ericales could have occurredeither at the base of the Ericales (as the ML phylogenyshows) or after the divergence of the basal lineages (balsami-noids and Lecythidaceae)Galax urceolata (Diapensiaceae)Hdiptera (Styracaceae)Saurauia zahlbruckneri (Actinidia-ceae) S pseudocamellia and C japonica (Theaceae) andSymplocos chinensis (Symplocaceae) all have multiple PIcopies belonging to two strongly supported clades due to thisancient duplication event Again this ancestral duplication isfollowed by different gene fateswith independent losses andduplications in each paralogous lineage Although furthersampling could lead to alternative explanations for the pro-posed multiple loss events there is some support for the hy-pothesis of independent gene loss in eachparalogous lineageSouthern blot analyses of Marcgravia umbellata (Geutenet al 2006) and Diospyros digyna (data not shown) both in-dicate that there is likely only one PI gene present in each ofthese species (fig 1 underlined) This observation supportsthe hypothesis of multiple PI gene losses within the Ericales
Because we wanted to explore the effect of omittingtaxa with single-copy genes of which the paralogous was
2632 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
putatively lost after duplication we constructed a secondmatrix with only duplicated PI genes due to the lsquolsquoancientrsquorsquoduplication from G urceolata (Diapensiaceae) H diptera(Styracaceae) S zahlbruckneri (Actinidiaceae) S pseudo-camellia and C japonica (Theaceae) and S chinensis(Symplocaceae) The MP analysis resulted in just one mostparsimonious tree that is identical to the tree with the high-est likelihood As in the previous analysis the same twoparalogous clades were identified In this case both BPPvalues (100 and 97) and ML bootstrap values (57 and53) were higher in this case supporting a duplication eventpresumably at least before the divergence of these familiesand after the divergence of the primuloid clade in Ericalesphylogeny (Schonenberger et al 2006)
In addition to these two ancient duplication events weidentified several other more recent duplication events inasterid evolution From I aquifolium (Aquifoliaceae) weamplified two PI copies but more extensive sampling ofAquifoliales representatives should reveal the timing of thisduplication more accurately In Cornales duplicate genesare found in both Davidia involucrata and Nyssa sylvatica(Nyssaceae) suggesting that the duplication that resulted inthese copies occurred in the lineage leading to NyssaceaeIn the clade of Ericales representatives there are seven spe-cies with more than one PI copy which appear to be morerecent than the previously described duplication event atthe base of the Ericales order For example we isolatedtwo PI homologues from Impatiens glandulifera (IgGLO1and IgGLO2 Balsaminaceae) A previously characterizedPI gene from Impatiens hawkeri (IhGLO Geuten et al2006) groups with IgGLO2 but not with the PI gene fromM umbellata (MuGLO Marcgraviaceae) suggesting thatthis duplication occurred at the base of the BalsaminaceaeSimilarly we isolated twoPI genes fromG brasiliensis (Le-cythidaceae) and the phylogeny indicates that one copy ismore related to the PI gene from another Lecythidaceae rep-resentativeNapoleonaea vogeliiBecauseGustavia is basalin the phylogeny of Lecythidaceae (Mori et al 2007) the du-plication leading to paralogous copies inGustavia probablyoccurred at the origin of the Lecythidaceae or earlier
We also identified two PI copies in Erica hiemalis(Ericaceae) and Actinidia chinensis (Actinidiaceae) In bothS pseudocamellia (Theaceae) and S zahlbruckneri (Acti-nidiaceae) a third PI copy is present presumably as a resultof a more recent duplication in one of their paralogs As thesequence similarity of the paralogs in these four species ishigh (91ndash98) it is hard to decide whether these genes areat the same genetic locus or at different loci We suggestthese genes to be paralogous copies with a very recent or-igin and not shared by any other species in our samplingFinally two PI genes were identified in P denticulata (Pri-mulaceae) Although sequence similarity among thesegenes is significantly lower (82) this duplication is notshared by any other PI genes in our analyses indicating thatthis duplication is specific to the Primulaceae
Duplications in the AP3 and TM6 Lineages Are Few andNot Correlated with PI Duplications
We also carried out phylogenetic analyses of asteridAP3 sequences using a number of newly identified AP3
sequences as well as previously characterized genes andEST sequences from Genbank (table 1) The phylogenywas inferred from a nucleotide alignment of genes from85 taxa which was analyzed using both MP and Bayesianmethods (fig 2) In the Bayesian analysis AlAP3 was usedto root the AP3-like tree Similar to previous analyses(Kramer and Irish 1999 Kramer et al 2006) our Bayesiananalysis recovered two highly supported clades A firstclade contains the euAP3 homologues (BPP 98) whereasa second clade contains the TM6 homologues (BPP 100)(fig 2) The duplication event that gave rise to these two eu-dicot AP3 gene clades is placed near the base of the core eu-dicots and likely occurred coincident with the origin ofTrochodendraceae and Buxaceae (Kramer et al 2006)Within core asterids our phylogeny gave no indicationsofmajor duplication events but onlyfiveof eight core asteridorderswere sampled Similarly in basal asterids duplicationevents have seemingly not played an important role duringAP3 evolution because the observed gene phylogeny did notconflict with the species phylogeny (Schonenberger et al2006) In several cases thoughparalogousAP3genesofveryrecent origin were identified in four species one from Eri-cales (I hawkeri Geuten et al 2006) and three from core as-terids (Hieracium piloselloides Mimulus cultstatus andSalvia coccinea) (Aagaard et al 2005) As for thePI phylog-eny the bootstrap values in the MP analyses were low andthese analyses resulted in a poorly resolved tree
After the basal eudicot and rosid representativesbranch off in the euAP3 clade (BPP 98) the Cornales rep-resentatives are sister to two large clades The first cladecontains the newly sampled Ericales representatives(BPP 81) whereas the second contains all core asterid se-quences (BPP 54) The unexpected association of Brassica-ceae representatives (AP3 BjAP3) with the euAP3 genesfrom Asterales was also present in previous analyses andprobably illustrates the need for even denser sampling(de Martino et al 2006 Kramer et al 2006) Within theEricales euAP3 clade (BPP 81) most relationships arepoorly resolved However a strongly supported primuloid(BPP 100) ericoid (BPP 91) and balsaminoid clade (BPP100) can be observed (Schonenberger et al 2005) Withinthe TM6-clade Ericales representatives are supported witha BPP of 99 but overall support within this clade is lowHowever we did not recover any paralogous copies of TM6genes in the species sampled suggesting that duplicationevents in this clade either do not occur readily or are se-lected against We also observed an unexpected associationof Asterales TM6 representatives with rosids (BPP 71) asreported in previous analyses (de Martino et al 2006Kramer et al 2006) The TM6 genes within the Solanalesare strongly supported as a clade (BPP 100)
RT-PCR Reveals Divergence of Expression PatternsAfter Duplication
To investigate whether gene duplicates have divergedin expression patterns or have seemingly remained underthe same regulatory control we used RT-PCR to monitorgene expression levels in mature dissected floral organs(fig 3) We limited our survey to Ericales species thatcontain representatives of PI genes arising from both the
PISTILLATAmdashDuplications in Asterids 2633
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
ancient and recent duplications We find that the expressionpattern of PI paralogs has diverged considerably and fairlyrapidly The PI protein is known to require an AP3-like in-teraction partner for its in vivo function in a number of coreeudicots (Jack et al 1992 Trobner et al 1992 Goto andMeyerowitz 1994) We therefore also examined the expres-sion patterns of TM6 and euAP3 genes in the same species
We compared Ericales expression patterns with the as-terid speciesPetunia hybrida (Solanaceae) BothPI genesin Petunia show a very similar expression pattern (Vanden-bussche et al 2004) Expression of these twoPI homologues(FBP1 and PMADS2) is mainly confined to the second andthirdwhorls (Angenent et al 1992Van derKrol et al 1993)Expression of PhDEF the AP3 homologue was mainly
FIG 2mdashEvolutionary reconstruction of APETALA3-like gene phylogeny in asterids as inferred from Bayesian analysis Bayesian posteriorprobabilities are indicated above branches nonparametric bootstrap support values are shown below branches Gray circle5 inferred duplication event
2634 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
confined to petals and stamens with low expression in firstand fourth whorl organs PhTM6 has a different expressionprofilewithstrongexpressioninstamensandcarpelswhereasexpression in sepals and petals is low (Vandenbussche et al2004) Similar expression patterns to Petunia of PI AP3 and
TM6 were detected in S lycopersicon another core asteridSolanaceae species (de Martino et al 2006)
Multiple copies of PI have been retained in the ge-nomes of S pseudocamellia C japonica and H dipteraafter the major duplication event at the base of the Ericales
FIG 3mdashRT-PCR expression data for B-class genes from selected Ericales representatives Sep 5 sepals pet 5 petals pet up 5 upper part ofpetals fus 5 fused part of petals and stamens sta 5 stamens sta up 5 upper part of stamens sta fus 5 fused part of stamens gyn 5 gynoecium andflo 5 flower
PISTILLATAmdashDuplications in Asterids 2635
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Both copies in each of these species appear to have divergedin their expression patterns after duplication (fig 3AndashC)Although HdGLO2 is expressed in the petals and stamensof H diptera the other copy (HdGLO1) is mainly ex-pressed in stamens and the gynoecium and absent in petalsIn comparison the TM6 and euAP3 lineage genes show un-expected expression patterns with strong expression ofHdTM6 in all disected organs but HdDEF expression beingmainly restricted to petals and carpels InC japonica one PIparalog (CjGLO2) is expressed throughout sepals petalsand stamens whereas expression of the second paralog(CjGLO1) appeared to be restricted to the nonfused partsof petals stamens and carpels Although both CjDEF andCjTM6 expression are detected in all organs strong expres-sion ofCjDEF is seenmainly in the sepals and petalsCjTM6expression is strongest in all stamen parts and in carpelsS pseudocamellia also displays differences in the expressionof its two PI copies One paralog (SpGLO2) is expressed inpetals stamens carpels and expression of the second paral-og (SpGLO1) is more restricted to the fused part of petals andstamens and upper part of petals The AP3 homologueSpDEF is strongly expressed in all stamen parts and carpelswhereas strong expression of SpTM6 is detected in sepals allstamen parts and carpels
In summary in these three cases there is one PI copywith a broad expression pattern in the flower (even in sepalsand carpels) whereas expression of the other PI copy ismore restricted to specific parts of petals and stamensThese expression patterns differ from those of PI lineagegenes in core asterid species (Vandenbussche et al2004 de Martino et al 2006) where PI expression (alsoin a duplicated configuration) is mainly restricted to the sec-ond and third whorl organs petals and stamens and nostrong expression in the first and fourth whorl organs is de-tected Interestingly the divergence in expression patternsin these species does not follow the gene phylogeny Theparalogous copies with a general expression pattern are notgrouped in one clade of PI genes in Ericales but rather arefound in two clades This suggests that the PI expressiondomains were not fixed immediately after duplication in-dicating that subfunctionalization in spatial expression doesnot explain retention of the duplicates Alternatively ifregulatory subfunctionalization was the main reason forretention evidence for this has been obscured throughtime Differences in expression between AP3 and TM6in C japonica seem to be very similar to members ofthe same lineages in the core asterid species Expressionof AP3-like genes in H diptera and S pseudocamelliahowever differs from those of Camellia with strongAP3 expression in the carpels of Stewartia and Halesiaand weak expression of AP3 in stamens of Halesia In eachof these species expression of one or both AP3-like genes isdetected in the sepals
Sarracenia drummondii (fig 3D) Styrax japonicus(fig 3E) C persicum (fig 3F) and Phlox paniculata(fig 3G) maintained probably only one of the paralogousPI copies in their genome after the ancient Ericales dupli-cation Moreover Sarracenia and Styrax maintained a dif-ferent copy (fig 1 BPP 68) than did Cyclamen and Phlox(fig 1 BPP89) We investigated the expression patterns ofPI genes in these species to monitor the possible divergence
between the two paralogous PI clades after duplication Ineach of these four species PI is expressed in petals and sta-mens similar to the expression patterns of PI in core aster-ids in previous studies (Vandenbussche et al 2004 deMartino et al 2006) In both Sarracenia (SdGLO) andPhlox (PpGLO) there is some expression detected in thecarpels whereas SjGLO (Styrax) is also expressed in sepalsNo clear divergence in expression of PI from the two dif-ferent clades (fig 1 BPP68 and BPP89) is detected in thesecond and third whorl organs The expression patterns ofAP3 and TM6 lineage genes are different between these fourspecies Although in S drummondii both AP3 and TM6 areexpressed in all four floral whorls the expression in Styraxis very similar to that of TM6 and AP3 in core asterid spe-cies with in addition to expression in petals and stamensexpression of TM6 in carpels and AP3 in sepals In Cycla-men TM6 expression is absent in the upper part of the sta-mens and strong expression of AP3 is detected in thecarpels In Phlox however TM6 expression is absent fromthe carpels
The multiple PI copies in G brasiliensis (Lecythida-ceae) and P acaulis (Primulaceae) are the result of a recentduplication probably at the base of these two families(fig 1) We looked at the expression pattern of each of theseduplicated genes and of the AP3-like genes in these speciesIn G brasiliensis GbGLO2 is expressed in petals and sta-mens whereas GbGLO1 is mainly expressed in the stami-nal parts and less in the petals (fig 3H) Expression of AP3and TM6 seems to be lsquolsquoinversersquorsquo compared with the ex-pected asterid expression with AP3 expression mainly re-stricted to the stamens and strong TM6 expression in thepetals No AP3-like expression is detected in the carpelsIn P acaulis there are differences between the expressionpatterns of both PI copies (fig 3I) PaGLO2 is expressed inall organs tested whereas the expression of PaGLO1 ismainly restricted to the sepals and petals Both AP3 andTM6 are strongly expressed in sepals and as inC persicumTM6-expression is weak in the staminal parts Even afterthese recent duplication events divergence in the expres-sion domain of the two PI copies is detected
In contrast to the duplicated PI genes of Petunia(Vandenbussche et al 2004) the expression domains ofPI genes have changed considerably after duplication inEricales For both ancient and several recent duplicationevents divergence in expression was detected indicatingthat the regulation of the duplicate copies has divergedIn the studied species where only one PI copy was isolatedit appears that an lsquolsquoancestralrsquorsquo expression pattern of PI isreestablished However a second copy could have beenmissed during amplification Also within the AP3-like lin-eage some derived expression patterns are detected
In Situ Hybridization Demonstrates ExpressionDivergence in Early Flower Development
To analyze the expression patterns of the PI genes afterduplication in early developmental stages we examined thespatio-temporal expression patterns of PI in selected Eri-cales species by in situ hybridization (fig 4) We selectedexamples of each type of fate after duplication retention
2636 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
of both ancient duplicates (Stewartia) retention of only oneof each PI paralog (C persicum and S rubra) and retentionof recent duplicates (P acaulis)
In S rostrata (fig 4AndashF) we could detect expressiondivergence between each of the PI copies at early stages offloral development At the stage when petal primordia areevident but stamen primordia are apparent only as smallmeristematic outgrowths SpGLO1 expression is detectedin petal and stamen primordia (fig 4A) whereas SpGLO2transcripts are also detected in the tip of the sepals (fig 4D)As the flower develops SpGLO2 expression is low butexpands to include developing carpels (fig 4E) and expres-sion is maintained in the carpels in later stages (fig 4F) Incontrast SpGLO1 expression is maintained at high levelsonly in petals and stamens through later stages (fig 4Band C) and does not expand into sepals and carpels Theseearly patterns of S rostrata PI gene expression are consis-tent with the later patterns of expression as detected byRT-PCR (fig 3I)
Expression patterns of PI homologues were furtherinvestigated in C persicum (fig 4GndashI) and S rubra(fig 4JndashK) CpGLO expression first appears in the youngpetal primordia (fig 4G) followed by expression in the sta-men primordia as they emerge (fig 4H) Expression inthese organs is maintained in later stages (fig 4I) In con-trast to the expression patterns in mature organs as docu-mented by RT-PCR weak expression in the developingsepals (fig 4H) and developing carpels (fig 4I) was de-tected For S rubra we were not able to obtain the earliestdevelopmental stages For later stages of floral develop-ment we observed strong expression of PI in petals andstamens (fig 4J) Weak expression of SdGLO was also de-tected in the developing carpels (fig 4K) which is consis-tent with the RT-PCR data (fig 3) These observationstogether with the RT-PCR data (fig 3) support the ideathat species with presumably only one PI copy in their ge-nome show expression that is more confined to petals andstamens
Finally we investigated the expression of recent PIduplicates in P acaulis (fig 4LndashQ) At early stages bothPaGLO1 (fig 4L) and PaGLO2 (fig 4O) are expressedacross the meristematic dome in sepal primordia commonpetalndashstamen primordia and carpel primordia Expressionof PaGLO2 is maintained in all organs at older stages(fig 4R and Q) whereas PaGLO1 expression is restrictedto the stamens (fig 4M and N)
The in situ expression data (fig 4) in comparison tothe RT-PCR data (fig 3) indicate that early patterns of ex-pression of PI genes are often developmentally regulatedwith different patterns of expression being observed early incomparison to later in flower development Nonethelessboth sets of data indicate that in species possessing dupli-cate PI genes divergence in expression patterns betweenthe two copies is a common theme
Positive Selection and Relaxed Purifying Selection ActsAfter the Duplication Event in Ericales in the K- and C-Cerminal Domain
The mRNA expression data indicate that duplicate PIparalogs have acquired different expression patterns sug-gesting independent evolution of their transcriptional reg-ulatory control It was also demonstrated that adaptiveevolution in the coding regions of MADS-box transcriptionfactors has played a role in the diversification of thisgene family (Martinez-Castilla and Alvarez-Buylla2003) Within class B genes Hernandez-Hernandez et al(2007) demonstrated the role of positive selection after crit-ical duplication events like the AP3-PI duplication and theeuAP3ndashTM6 duplication To investigate whether parallelchanges occurred in the coding regions of the genes exam-ined in this study we investigated the nonsynonymous ver-sus synonymous amino acid replacement rates forsignificant increases before and after duplication in thespecific paralogous clades and along the coding sequenceThe results of likelihood ratio tests are shown in table 2 and
FIG 4mdashIn situ hybridization data for PI genes in various developmental stages of Stewartia rostrata (AndashC SpGLO1 DndashF SpGLO2) Cyclamenpersicum (GndashI CpGLO) Sarracenia rubra (JndashK SdGLO) Primula acaulis (LndashN PaGLO1 OndashQ PaGLO2) lsquolsquoprsquorsquo indicates petalprimordia lsquolsquosrsquorsquoindicates stamen primordia
PISTILLATAmdashDuplications in Asterids 2637
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
specific sites are marked in the alignment presented infigure 5 Parameter estimates log-likelihood scores andBayes Empirical Bayes identified sites are listed in supple-mentary table S2 Supplementary Material online
Comparison of the likelihood scores calculated usingbranch models R1 and R2 for matrix A indicates a signifi-cantly increased evolutionary rate following the ancient du-plication event in Ericales The dNdS estimate from thetwo-ratio branch model after duplication (x 5 0285) istwice compared with the value before duplication (x 50113) illustrating the relaxed selective regime resultingfrom the duplication event However the same test appliedon matrix B did not reveal this change in selective pressureOne possible explanation is that by adding the single-copylineages that evolved after duplication the observed patternin matrix A for the duplicates is obscured In addition wefound no evidence that the two paralogous gene cladescould be evolving at different rates because the three-and four-ratio models did not result in a significantly in-creased log-likelihood as calculated for either of the datamatrices
To formally test for the presence of positive selectionon sequence sites we used the site models proposed byNielsen and Yang (1998) and Yang et al (2000) Analysisof matrix A shows that both M2a and M8 two models thatallow for positive selection fit the data significantly betterthan the corresponding neutral models (M1a and M7) Sub-stitution patterns in about 5 of the sites for the M2amodel and about 9 of the sites for the M8 model appearto result from adaptive evolution Here also analysis of ma-trix B suggests positively selected sites M8 fits the databetter than the neutral model M7 with an estimation of
10 sites under positive selection Bayes empirical Bayes(Yang et al 2005) identification of the sites under positiveselection in matrix A finds seven sites to be positively se-lected under model M8 two of these sites have a posteriorprobability higher than 99 Within matrix B only twopositively selected sites were detected which were alsoidentified in matrix A both with posterior probability high-er than 91 Interestingly all the identified sites are locatedwithin the K2-subdomain the interhelical region betweenK2 and K3 the K3-subdomain and the C-terminal domain(fig 5) These functional domains have been shown to beimportant for proteinndashprotein interactions among Arabi-dopsis MADS-domain proteins (Ma et al 1991 Pnueliet al 1991 Shore and Sharrocks 1995 Riechmann et al1996 Yang et al 2003) The strength of the AP3ndashPI inter-action is mediated primarily by K1 and K2 whereas thePIndashSEP interaction is mediated by K2 the amino-terminalportion of K3 and possibly the C-domain (Yang et al 2003Yang and Jack 2004) The C-terminal region normallyshows much lower sequence conservation but containsshort highly conserved motifs (Lamb and Irish 2003) Sev-eral studies have suggested that this domain is involvedin the formation of higher order protein complexes(Egea-Cortines et al 1999 Honma and Goto 2001) yetthe specific function of the class B C-terminal domainsis unclear (Lamb and Irish 2003 Piwarzyk et al 2007)None of the sites identified by Yang et al (2003) to be re-sponsible for the direct interaction between APETALA3and PISTILLATA are positively selected for in our dataset However site 49 contains a hydrophobic amino acidthat when mutated disrupts the alpha-helical secondarystructure (Yang et al 2003) In addition sites 49 and site60 in figure 5 were identified by Yang and Jack (2004) to becritical for PIndashSEP interaction Site 49 is situated withinthe K2 subdomain which is required for both AP3ndashPIand PIndashSEP interaction whereas site 60 which is locatedat the amino-terminal end of K3 affects when mutated thestrength of the PIndashSEP interaction
To identify evolutionary pressures acting on specificsites along certain branches and in clades we calculatedlikelihoods using the branch-site models First we specifiedthe two branches immediately following the duplication asforeground branches in matrix A Branch-site models thatallow for positive selection in these foreground brancheswere not better at explaining the data than the nested sitemodels M1a and the null model A and indicate no evidencefor a burst of positive selection in the branches immediatelyfollowing the duplication event (Bielawski and Yang 2003)(data not shown) Second we treated all branches in eitherclade of paralogs as foreground branches When these twoclades are treated separately the model allowing for
Table 2Likelihood Ratio Tests Comparing Branch-Models (R1ndashR4)Site Models (M1ndashM8) and Branch-Site Models (Model AndashModel A Null)
Matrix Models Compared2(ln L1 ln L0)
Matrix A One ratio (R1) versus two ratio (R2) 18174Two ratio (R2) versus three ratio (R3) 0435Three ratio (R3) versus four ratio (R4) 1298Neutral (M1a) versus selection (M2a) 20566Beta (M7) versus beta amp w 1 (M8) 2683Model A versus M1a (Test 1) 874Model A versus model anull (Test 2) 252
Matrix B One ratio (R1) versus two ratio (R2) 2348Two ratio (R2) versus three ratio (R3) 001Three ratio (R3) versus four ratio (R4) 01Neutral (M1a) versus selection (M2a) 367Beta (M7) versus beta amp w 1 (M8) 7054
Significant at the 95 level significant at the 99 level
Table 3B Class MADS-Box Protein Interactions as Determined by Yeast Two-Hybrid Assays in Camellia japonica
CjDEF-BD CjTM6-BD CjGLO1-BD CjGLO2-BD BD
CjDEF-AD thorn mdash mdash mdash mdashCjTM6-AD mdash mdash thorn mdash mdashCjGLO1-AD thorn thorn mdash mdash mdashCjGLO2-AD thorn mdash mdash mdash mdashAD mdash mdash mdash mdash mdash
2638 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
positive selection is not significantly better in explaining thedata than the model M1a and null model A (data notshown) When the two clades are treated together as fore-ground branches model A is significantly better than thenested site model M1a (test 1) suggesting that positive se-lection is acting in both clades that arose after duplicationBayes empirical Bayes identification resulted in seven po-sitions though all with posterior probability below 90The improved test 2 from Zhang et al (2005) resulted ina value 252 just beneath the critical value 271 at the5 level but far above the critical value at the 10 level164 These two tests support the hypothesis that positiveselection acts after the duplication event in both paralogousgene clades
Together the identification of changes in selectivepressure in time and along the coding sequence indicatethat upon duplication PI paralogs underwent an increasednonsynonymous substitution rate and that this occurred
specifically in the domains shown to be necessary forproteinndashprotein interactions
Diversification in the Interaction Behavior of B-ClassGenes After the Ancient Duplication Event in BasalAsterids
As sites in the interaction domains of the PI genesfrom basal asterids were detected to be positively selectedafter duplication we further investigated the possible func-tional divergence of the duplicated PI proteins by yeast-2-hybrid analyses We tested the interactions between thefour class B genes in C japonica Camellia japonica con-tains two AP3-like genes one belonging to the euAP3-typeand one from the paleoAP3-type and two PI-like genesfrom an ancient duplication event at the base of the EricalesThis situation is similar to the B-class configuration in thecore asterid species Petunia hybrida (Vandenbussche
FIG 5mdashAlignment of the K-domain and C-terminal domain of the PI duplicates from matrix A used for the analysis in PAML Differentsubdomains and the characteristic PI motif are indicated (Kramer et al 1998) Codon positions identified to be under positive selection are marked withgray boxes
PISTILLATAmdashDuplications in Asterids 2639
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

selection on sequence sites site models were applied thathold x constant among all branches but allow x to varyamong codons Site models include M1a M2a M7 andM8 (Nielsen and Yang 1998 Wong et al 2004 Yang et al2005) Positive selection is inferred when M2a and M8 arerespectively significantly better than M1a and M7 Branch-sitemodels allow thex ratio to vary amongboth sites and lin-eages (Zhang et al 2005) and can be used to detect positivelyselected sites in specific lineages within the tree We usedthe modified branch-site model A and compared this modelwith the nested site model M1a (Test 1) and the null modelA (Test 2) to test for positive selection (Zhang et al 2005)
Yeast Two-Hybrid Assays
The intervening- keratin-like and C-terminal domaindomains of the two PI (CjGLO1 and CjGLO2) the AP3(CjDEF) and TM6 genes (CjTM6) from Camellia japonicawere fusedwith theGAL4activationdomain in thepGAD424vector and the GAL4 DNA-binding domain in the pGBT9vector (ClontechMountainView CA) Each possible vectorcombination was transformed in yeast and two colonies pertransformation were used for B-galactosidase liquid assaysusing the protocol available at httpwwwfhcrcorgsciencelabsgottschlingyeast20ProtocolsBgalhtml
Results
In total 83 new B-lineage sequences were amplifiedfrom asterid species Forty-seven new PI and 36 new AP3-like sequences were identified from 31 and 25 asterid speciesrespectively (table 1) All but one of these species belong tothe basal asterid orders Cornales and Ericales In addition PIhomologues were amplified from I aquifolium (Aquifoliales)
Two Ancient and Frequent Recent Duplications HaveOccurred in the Asterid PI Lineage
For the phylogenetic analyses of the PI lineage wecombined the newly identified PI genes with previouslyidentified asterid PI sequences and EST data from Genbankto generate one data set Basal eudicot sequences were usedas outgroups for this data set and AqvPI (Aquilegia Ranun-culales) was used to root the PI tree A nucleotide alignmentof 74 taxa was analyzed using MP ML and Bayesian anal-yses The results are summarized in figure 1
The Bayesian topology which has relatively high pos-terior probabilities for a majority of nodes did not conflictwith the most likely tree from the ML analysis The MPanalysis however did not resolve the relationships betweenthe major clades and high bootstrap values were only ob-tained for nodes at lower taxonomical levels Similar to thebootstrap values from the MP analysis bootstrap supportfor the ML tree is low and we chose to only indicate thebootstrap values from the MP analysis on the phylogenyHowever to provide an additional measure of support ina likelihood framework we show SH-like and v2-like aLRTtest values These provide additional support for the Bayes-ian inferred relationships
The inferred PI phylogeny (fig 1) indicates that thisgene lineage probably underwent two ancient duplication
events during asterid evolution In both instances the twocopies were retained in some extant species lineageswhereas in other taxa one copy appears to have been lostor further duplicated An initial ancient duplication probablyoccurred early in the evolution of the core asterids becausethe Bayesian analysis supports two paralogous cladesa clade containing Gentianales Solanales and Asteralessequences (BPP 95) and one with Solanales and Lamialessequences (BPP 100) The latter is also strongly supportedbyMPbootstrapping (86) and aLRT testing (10099) Givenour current knowledge of asterid evolution (Albach et al2001 Bremer et al 2002) this suggests an ancient duplica-tion event before the origin of the two major core asteridclades (euasterids I and euasterids II) (Bayesian analysis)or at least before the origin of the euasterid II clade compris-ing Solanales Lamiales and Gentianales (MP analysis andaLRT testing) This duplication event was then followed byindependent losses in each paralogous lineageAlthoughourresults allow us to infer such a scenario no Garryales Dip-sacales or Apiales PI representatives are currently availablethat would further support such a hypothesis In addition thePI sequences of Aquifoliales an order situated at the base ofthe euasterid II clade occupy a nonsupported position at thebase of basal asterid PI sequences Together this suggeststhat an even denser sampling of asterid sequences would bedesirable to determine the exact moment of the PI duplica-tion in the core asterid lineage with even more confidence
The ML tree further suggests a second ancient dupli-cation event consistent with an origin early in the radiationof the order Ericales The PI sequences from Ericales rep-resentatives that are strongly supported by Bayesian support(BPP 96) and aLRT support (9999) are placed into two sep-arate clades A first clade is supported by a BPP value of 90and a v2-value of 92 The weakly supported second clade(BPP 67 aLRT 7088) consists of two strongly supportedclades (BPP 96 aLRT 9499 and BPP 90 aLRT 8999)where one (BPP 96) consists of the proposed basal lineagesof the Ericales the balsaminoıd clade and Lecythidaceae(Geuten et al 2004 Schonenberger et al 2005) The pres-ence of three strongly supported clades within the Ericalessuggests that the duplication in Ericales could have occurredeither at the base of the Ericales (as the ML phylogenyshows) or after the divergence of the basal lineages (balsami-noids and Lecythidaceae)Galax urceolata (Diapensiaceae)Hdiptera (Styracaceae)Saurauia zahlbruckneri (Actinidia-ceae) S pseudocamellia and C japonica (Theaceae) andSymplocos chinensis (Symplocaceae) all have multiple PIcopies belonging to two strongly supported clades due to thisancient duplication event Again this ancestral duplication isfollowed by different gene fateswith independent losses andduplications in each paralogous lineage Although furthersampling could lead to alternative explanations for the pro-posed multiple loss events there is some support for the hy-pothesis of independent gene loss in eachparalogous lineageSouthern blot analyses of Marcgravia umbellata (Geutenet al 2006) and Diospyros digyna (data not shown) both in-dicate that there is likely only one PI gene present in each ofthese species (fig 1 underlined) This observation supportsthe hypothesis of multiple PI gene losses within the Ericales
Because we wanted to explore the effect of omittingtaxa with single-copy genes of which the paralogous was
2632 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
putatively lost after duplication we constructed a secondmatrix with only duplicated PI genes due to the lsquolsquoancientrsquorsquoduplication from G urceolata (Diapensiaceae) H diptera(Styracaceae) S zahlbruckneri (Actinidiaceae) S pseudo-camellia and C japonica (Theaceae) and S chinensis(Symplocaceae) The MP analysis resulted in just one mostparsimonious tree that is identical to the tree with the high-est likelihood As in the previous analysis the same twoparalogous clades were identified In this case both BPPvalues (100 and 97) and ML bootstrap values (57 and53) were higher in this case supporting a duplication eventpresumably at least before the divergence of these familiesand after the divergence of the primuloid clade in Ericalesphylogeny (Schonenberger et al 2006)
In addition to these two ancient duplication events weidentified several other more recent duplication events inasterid evolution From I aquifolium (Aquifoliaceae) weamplified two PI copies but more extensive sampling ofAquifoliales representatives should reveal the timing of thisduplication more accurately In Cornales duplicate genesare found in both Davidia involucrata and Nyssa sylvatica(Nyssaceae) suggesting that the duplication that resulted inthese copies occurred in the lineage leading to NyssaceaeIn the clade of Ericales representatives there are seven spe-cies with more than one PI copy which appear to be morerecent than the previously described duplication event atthe base of the Ericales order For example we isolatedtwo PI homologues from Impatiens glandulifera (IgGLO1and IgGLO2 Balsaminaceae) A previously characterizedPI gene from Impatiens hawkeri (IhGLO Geuten et al2006) groups with IgGLO2 but not with the PI gene fromM umbellata (MuGLO Marcgraviaceae) suggesting thatthis duplication occurred at the base of the BalsaminaceaeSimilarly we isolated twoPI genes fromG brasiliensis (Le-cythidaceae) and the phylogeny indicates that one copy ismore related to the PI gene from another Lecythidaceae rep-resentativeNapoleonaea vogeliiBecauseGustavia is basalin the phylogeny of Lecythidaceae (Mori et al 2007) the du-plication leading to paralogous copies inGustavia probablyoccurred at the origin of the Lecythidaceae or earlier
We also identified two PI copies in Erica hiemalis(Ericaceae) and Actinidia chinensis (Actinidiaceae) In bothS pseudocamellia (Theaceae) and S zahlbruckneri (Acti-nidiaceae) a third PI copy is present presumably as a resultof a more recent duplication in one of their paralogs As thesequence similarity of the paralogs in these four species ishigh (91ndash98) it is hard to decide whether these genes areat the same genetic locus or at different loci We suggestthese genes to be paralogous copies with a very recent or-igin and not shared by any other species in our samplingFinally two PI genes were identified in P denticulata (Pri-mulaceae) Although sequence similarity among thesegenes is significantly lower (82) this duplication is notshared by any other PI genes in our analyses indicating thatthis duplication is specific to the Primulaceae
Duplications in the AP3 and TM6 Lineages Are Few andNot Correlated with PI Duplications
We also carried out phylogenetic analyses of asteridAP3 sequences using a number of newly identified AP3
sequences as well as previously characterized genes andEST sequences from Genbank (table 1) The phylogenywas inferred from a nucleotide alignment of genes from85 taxa which was analyzed using both MP and Bayesianmethods (fig 2) In the Bayesian analysis AlAP3 was usedto root the AP3-like tree Similar to previous analyses(Kramer and Irish 1999 Kramer et al 2006) our Bayesiananalysis recovered two highly supported clades A firstclade contains the euAP3 homologues (BPP 98) whereasa second clade contains the TM6 homologues (BPP 100)(fig 2) The duplication event that gave rise to these two eu-dicot AP3 gene clades is placed near the base of the core eu-dicots and likely occurred coincident with the origin ofTrochodendraceae and Buxaceae (Kramer et al 2006)Within core asterids our phylogeny gave no indicationsofmajor duplication events but onlyfiveof eight core asteridorderswere sampled Similarly in basal asterids duplicationevents have seemingly not played an important role duringAP3 evolution because the observed gene phylogeny did notconflict with the species phylogeny (Schonenberger et al2006) In several cases thoughparalogousAP3genesofveryrecent origin were identified in four species one from Eri-cales (I hawkeri Geuten et al 2006) and three from core as-terids (Hieracium piloselloides Mimulus cultstatus andSalvia coccinea) (Aagaard et al 2005) As for thePI phylog-eny the bootstrap values in the MP analyses were low andthese analyses resulted in a poorly resolved tree
After the basal eudicot and rosid representativesbranch off in the euAP3 clade (BPP 98) the Cornales rep-resentatives are sister to two large clades The first cladecontains the newly sampled Ericales representatives(BPP 81) whereas the second contains all core asterid se-quences (BPP 54) The unexpected association of Brassica-ceae representatives (AP3 BjAP3) with the euAP3 genesfrom Asterales was also present in previous analyses andprobably illustrates the need for even denser sampling(de Martino et al 2006 Kramer et al 2006) Within theEricales euAP3 clade (BPP 81) most relationships arepoorly resolved However a strongly supported primuloid(BPP 100) ericoid (BPP 91) and balsaminoid clade (BPP100) can be observed (Schonenberger et al 2005) Withinthe TM6-clade Ericales representatives are supported witha BPP of 99 but overall support within this clade is lowHowever we did not recover any paralogous copies of TM6genes in the species sampled suggesting that duplicationevents in this clade either do not occur readily or are se-lected against We also observed an unexpected associationof Asterales TM6 representatives with rosids (BPP 71) asreported in previous analyses (de Martino et al 2006Kramer et al 2006) The TM6 genes within the Solanalesare strongly supported as a clade (BPP 100)
RT-PCR Reveals Divergence of Expression PatternsAfter Duplication
To investigate whether gene duplicates have divergedin expression patterns or have seemingly remained underthe same regulatory control we used RT-PCR to monitorgene expression levels in mature dissected floral organs(fig 3) We limited our survey to Ericales species thatcontain representatives of PI genes arising from both the
PISTILLATAmdashDuplications in Asterids 2633
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
ancient and recent duplications We find that the expressionpattern of PI paralogs has diverged considerably and fairlyrapidly The PI protein is known to require an AP3-like in-teraction partner for its in vivo function in a number of coreeudicots (Jack et al 1992 Trobner et al 1992 Goto andMeyerowitz 1994) We therefore also examined the expres-sion patterns of TM6 and euAP3 genes in the same species
We compared Ericales expression patterns with the as-terid speciesPetunia hybrida (Solanaceae) BothPI genesin Petunia show a very similar expression pattern (Vanden-bussche et al 2004) Expression of these twoPI homologues(FBP1 and PMADS2) is mainly confined to the second andthirdwhorls (Angenent et al 1992Van derKrol et al 1993)Expression of PhDEF the AP3 homologue was mainly
FIG 2mdashEvolutionary reconstruction of APETALA3-like gene phylogeny in asterids as inferred from Bayesian analysis Bayesian posteriorprobabilities are indicated above branches nonparametric bootstrap support values are shown below branches Gray circle5 inferred duplication event
2634 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
confined to petals and stamens with low expression in firstand fourth whorl organs PhTM6 has a different expressionprofilewithstrongexpressioninstamensandcarpelswhereasexpression in sepals and petals is low (Vandenbussche et al2004) Similar expression patterns to Petunia of PI AP3 and
TM6 were detected in S lycopersicon another core asteridSolanaceae species (de Martino et al 2006)
Multiple copies of PI have been retained in the ge-nomes of S pseudocamellia C japonica and H dipteraafter the major duplication event at the base of the Ericales
FIG 3mdashRT-PCR expression data for B-class genes from selected Ericales representatives Sep 5 sepals pet 5 petals pet up 5 upper part ofpetals fus 5 fused part of petals and stamens sta 5 stamens sta up 5 upper part of stamens sta fus 5 fused part of stamens gyn 5 gynoecium andflo 5 flower
PISTILLATAmdashDuplications in Asterids 2635
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Both copies in each of these species appear to have divergedin their expression patterns after duplication (fig 3AndashC)Although HdGLO2 is expressed in the petals and stamensof H diptera the other copy (HdGLO1) is mainly ex-pressed in stamens and the gynoecium and absent in petalsIn comparison the TM6 and euAP3 lineage genes show un-expected expression patterns with strong expression ofHdTM6 in all disected organs but HdDEF expression beingmainly restricted to petals and carpels InC japonica one PIparalog (CjGLO2) is expressed throughout sepals petalsand stamens whereas expression of the second paralog(CjGLO1) appeared to be restricted to the nonfused partsof petals stamens and carpels Although both CjDEF andCjTM6 expression are detected in all organs strong expres-sion ofCjDEF is seenmainly in the sepals and petalsCjTM6expression is strongest in all stamen parts and in carpelsS pseudocamellia also displays differences in the expressionof its two PI copies One paralog (SpGLO2) is expressed inpetals stamens carpels and expression of the second paral-og (SpGLO1) is more restricted to the fused part of petals andstamens and upper part of petals The AP3 homologueSpDEF is strongly expressed in all stamen parts and carpelswhereas strong expression of SpTM6 is detected in sepals allstamen parts and carpels
In summary in these three cases there is one PI copywith a broad expression pattern in the flower (even in sepalsand carpels) whereas expression of the other PI copy ismore restricted to specific parts of petals and stamensThese expression patterns differ from those of PI lineagegenes in core asterid species (Vandenbussche et al2004 de Martino et al 2006) where PI expression (alsoin a duplicated configuration) is mainly restricted to the sec-ond and third whorl organs petals and stamens and nostrong expression in the first and fourth whorl organs is de-tected Interestingly the divergence in expression patternsin these species does not follow the gene phylogeny Theparalogous copies with a general expression pattern are notgrouped in one clade of PI genes in Ericales but rather arefound in two clades This suggests that the PI expressiondomains were not fixed immediately after duplication in-dicating that subfunctionalization in spatial expression doesnot explain retention of the duplicates Alternatively ifregulatory subfunctionalization was the main reason forretention evidence for this has been obscured throughtime Differences in expression between AP3 and TM6in C japonica seem to be very similar to members ofthe same lineages in the core asterid species Expressionof AP3-like genes in H diptera and S pseudocamelliahowever differs from those of Camellia with strongAP3 expression in the carpels of Stewartia and Halesiaand weak expression of AP3 in stamens of Halesia In eachof these species expression of one or both AP3-like genes isdetected in the sepals
Sarracenia drummondii (fig 3D) Styrax japonicus(fig 3E) C persicum (fig 3F) and Phlox paniculata(fig 3G) maintained probably only one of the paralogousPI copies in their genome after the ancient Ericales dupli-cation Moreover Sarracenia and Styrax maintained a dif-ferent copy (fig 1 BPP 68) than did Cyclamen and Phlox(fig 1 BPP89) We investigated the expression patterns ofPI genes in these species to monitor the possible divergence
between the two paralogous PI clades after duplication Ineach of these four species PI is expressed in petals and sta-mens similar to the expression patterns of PI in core aster-ids in previous studies (Vandenbussche et al 2004 deMartino et al 2006) In both Sarracenia (SdGLO) andPhlox (PpGLO) there is some expression detected in thecarpels whereas SjGLO (Styrax) is also expressed in sepalsNo clear divergence in expression of PI from the two dif-ferent clades (fig 1 BPP68 and BPP89) is detected in thesecond and third whorl organs The expression patterns ofAP3 and TM6 lineage genes are different between these fourspecies Although in S drummondii both AP3 and TM6 areexpressed in all four floral whorls the expression in Styraxis very similar to that of TM6 and AP3 in core asterid spe-cies with in addition to expression in petals and stamensexpression of TM6 in carpels and AP3 in sepals In Cycla-men TM6 expression is absent in the upper part of the sta-mens and strong expression of AP3 is detected in thecarpels In Phlox however TM6 expression is absent fromthe carpels
The multiple PI copies in G brasiliensis (Lecythida-ceae) and P acaulis (Primulaceae) are the result of a recentduplication probably at the base of these two families(fig 1) We looked at the expression pattern of each of theseduplicated genes and of the AP3-like genes in these speciesIn G brasiliensis GbGLO2 is expressed in petals and sta-mens whereas GbGLO1 is mainly expressed in the stami-nal parts and less in the petals (fig 3H) Expression of AP3and TM6 seems to be lsquolsquoinversersquorsquo compared with the ex-pected asterid expression with AP3 expression mainly re-stricted to the stamens and strong TM6 expression in thepetals No AP3-like expression is detected in the carpelsIn P acaulis there are differences between the expressionpatterns of both PI copies (fig 3I) PaGLO2 is expressed inall organs tested whereas the expression of PaGLO1 ismainly restricted to the sepals and petals Both AP3 andTM6 are strongly expressed in sepals and as inC persicumTM6-expression is weak in the staminal parts Even afterthese recent duplication events divergence in the expres-sion domain of the two PI copies is detected
In contrast to the duplicated PI genes of Petunia(Vandenbussche et al 2004) the expression domains ofPI genes have changed considerably after duplication inEricales For both ancient and several recent duplicationevents divergence in expression was detected indicatingthat the regulation of the duplicate copies has divergedIn the studied species where only one PI copy was isolatedit appears that an lsquolsquoancestralrsquorsquo expression pattern of PI isreestablished However a second copy could have beenmissed during amplification Also within the AP3-like lin-eage some derived expression patterns are detected
In Situ Hybridization Demonstrates ExpressionDivergence in Early Flower Development
To analyze the expression patterns of the PI genes afterduplication in early developmental stages we examined thespatio-temporal expression patterns of PI in selected Eri-cales species by in situ hybridization (fig 4) We selectedexamples of each type of fate after duplication retention
2636 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
of both ancient duplicates (Stewartia) retention of only oneof each PI paralog (C persicum and S rubra) and retentionof recent duplicates (P acaulis)
In S rostrata (fig 4AndashF) we could detect expressiondivergence between each of the PI copies at early stages offloral development At the stage when petal primordia areevident but stamen primordia are apparent only as smallmeristematic outgrowths SpGLO1 expression is detectedin petal and stamen primordia (fig 4A) whereas SpGLO2transcripts are also detected in the tip of the sepals (fig 4D)As the flower develops SpGLO2 expression is low butexpands to include developing carpels (fig 4E) and expres-sion is maintained in the carpels in later stages (fig 4F) Incontrast SpGLO1 expression is maintained at high levelsonly in petals and stamens through later stages (fig 4Band C) and does not expand into sepals and carpels Theseearly patterns of S rostrata PI gene expression are consis-tent with the later patterns of expression as detected byRT-PCR (fig 3I)
Expression patterns of PI homologues were furtherinvestigated in C persicum (fig 4GndashI) and S rubra(fig 4JndashK) CpGLO expression first appears in the youngpetal primordia (fig 4G) followed by expression in the sta-men primordia as they emerge (fig 4H) Expression inthese organs is maintained in later stages (fig 4I) In con-trast to the expression patterns in mature organs as docu-mented by RT-PCR weak expression in the developingsepals (fig 4H) and developing carpels (fig 4I) was de-tected For S rubra we were not able to obtain the earliestdevelopmental stages For later stages of floral develop-ment we observed strong expression of PI in petals andstamens (fig 4J) Weak expression of SdGLO was also de-tected in the developing carpels (fig 4K) which is consis-tent with the RT-PCR data (fig 3) These observationstogether with the RT-PCR data (fig 3) support the ideathat species with presumably only one PI copy in their ge-nome show expression that is more confined to petals andstamens
Finally we investigated the expression of recent PIduplicates in P acaulis (fig 4LndashQ) At early stages bothPaGLO1 (fig 4L) and PaGLO2 (fig 4O) are expressedacross the meristematic dome in sepal primordia commonpetalndashstamen primordia and carpel primordia Expressionof PaGLO2 is maintained in all organs at older stages(fig 4R and Q) whereas PaGLO1 expression is restrictedto the stamens (fig 4M and N)
The in situ expression data (fig 4) in comparison tothe RT-PCR data (fig 3) indicate that early patterns of ex-pression of PI genes are often developmentally regulatedwith different patterns of expression being observed early incomparison to later in flower development Nonethelessboth sets of data indicate that in species possessing dupli-cate PI genes divergence in expression patterns betweenthe two copies is a common theme
Positive Selection and Relaxed Purifying Selection ActsAfter the Duplication Event in Ericales in the K- and C-Cerminal Domain
The mRNA expression data indicate that duplicate PIparalogs have acquired different expression patterns sug-gesting independent evolution of their transcriptional reg-ulatory control It was also demonstrated that adaptiveevolution in the coding regions of MADS-box transcriptionfactors has played a role in the diversification of thisgene family (Martinez-Castilla and Alvarez-Buylla2003) Within class B genes Hernandez-Hernandez et al(2007) demonstrated the role of positive selection after crit-ical duplication events like the AP3-PI duplication and theeuAP3ndashTM6 duplication To investigate whether parallelchanges occurred in the coding regions of the genes exam-ined in this study we investigated the nonsynonymous ver-sus synonymous amino acid replacement rates forsignificant increases before and after duplication in thespecific paralogous clades and along the coding sequenceThe results of likelihood ratio tests are shown in table 2 and
FIG 4mdashIn situ hybridization data for PI genes in various developmental stages of Stewartia rostrata (AndashC SpGLO1 DndashF SpGLO2) Cyclamenpersicum (GndashI CpGLO) Sarracenia rubra (JndashK SdGLO) Primula acaulis (LndashN PaGLO1 OndashQ PaGLO2) lsquolsquoprsquorsquo indicates petalprimordia lsquolsquosrsquorsquoindicates stamen primordia
PISTILLATAmdashDuplications in Asterids 2637
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
specific sites are marked in the alignment presented infigure 5 Parameter estimates log-likelihood scores andBayes Empirical Bayes identified sites are listed in supple-mentary table S2 Supplementary Material online
Comparison of the likelihood scores calculated usingbranch models R1 and R2 for matrix A indicates a signifi-cantly increased evolutionary rate following the ancient du-plication event in Ericales The dNdS estimate from thetwo-ratio branch model after duplication (x 5 0285) istwice compared with the value before duplication (x 50113) illustrating the relaxed selective regime resultingfrom the duplication event However the same test appliedon matrix B did not reveal this change in selective pressureOne possible explanation is that by adding the single-copylineages that evolved after duplication the observed patternin matrix A for the duplicates is obscured In addition wefound no evidence that the two paralogous gene cladescould be evolving at different rates because the three-and four-ratio models did not result in a significantly in-creased log-likelihood as calculated for either of the datamatrices
To formally test for the presence of positive selectionon sequence sites we used the site models proposed byNielsen and Yang (1998) and Yang et al (2000) Analysisof matrix A shows that both M2a and M8 two models thatallow for positive selection fit the data significantly betterthan the corresponding neutral models (M1a and M7) Sub-stitution patterns in about 5 of the sites for the M2amodel and about 9 of the sites for the M8 model appearto result from adaptive evolution Here also analysis of ma-trix B suggests positively selected sites M8 fits the databetter than the neutral model M7 with an estimation of
10 sites under positive selection Bayes empirical Bayes(Yang et al 2005) identification of the sites under positiveselection in matrix A finds seven sites to be positively se-lected under model M8 two of these sites have a posteriorprobability higher than 99 Within matrix B only twopositively selected sites were detected which were alsoidentified in matrix A both with posterior probability high-er than 91 Interestingly all the identified sites are locatedwithin the K2-subdomain the interhelical region betweenK2 and K3 the K3-subdomain and the C-terminal domain(fig 5) These functional domains have been shown to beimportant for proteinndashprotein interactions among Arabi-dopsis MADS-domain proteins (Ma et al 1991 Pnueliet al 1991 Shore and Sharrocks 1995 Riechmann et al1996 Yang et al 2003) The strength of the AP3ndashPI inter-action is mediated primarily by K1 and K2 whereas thePIndashSEP interaction is mediated by K2 the amino-terminalportion of K3 and possibly the C-domain (Yang et al 2003Yang and Jack 2004) The C-terminal region normallyshows much lower sequence conservation but containsshort highly conserved motifs (Lamb and Irish 2003) Sev-eral studies have suggested that this domain is involvedin the formation of higher order protein complexes(Egea-Cortines et al 1999 Honma and Goto 2001) yetthe specific function of the class B C-terminal domainsis unclear (Lamb and Irish 2003 Piwarzyk et al 2007)None of the sites identified by Yang et al (2003) to be re-sponsible for the direct interaction between APETALA3and PISTILLATA are positively selected for in our dataset However site 49 contains a hydrophobic amino acidthat when mutated disrupts the alpha-helical secondarystructure (Yang et al 2003) In addition sites 49 and site60 in figure 5 were identified by Yang and Jack (2004) to becritical for PIndashSEP interaction Site 49 is situated withinthe K2 subdomain which is required for both AP3ndashPIand PIndashSEP interaction whereas site 60 which is locatedat the amino-terminal end of K3 affects when mutated thestrength of the PIndashSEP interaction
To identify evolutionary pressures acting on specificsites along certain branches and in clades we calculatedlikelihoods using the branch-site models First we specifiedthe two branches immediately following the duplication asforeground branches in matrix A Branch-site models thatallow for positive selection in these foreground brancheswere not better at explaining the data than the nested sitemodels M1a and the null model A and indicate no evidencefor a burst of positive selection in the branches immediatelyfollowing the duplication event (Bielawski and Yang 2003)(data not shown) Second we treated all branches in eitherclade of paralogs as foreground branches When these twoclades are treated separately the model allowing for
Table 2Likelihood Ratio Tests Comparing Branch-Models (R1ndashR4)Site Models (M1ndashM8) and Branch-Site Models (Model AndashModel A Null)
Matrix Models Compared2(ln L1 ln L0)
Matrix A One ratio (R1) versus two ratio (R2) 18174Two ratio (R2) versus three ratio (R3) 0435Three ratio (R3) versus four ratio (R4) 1298Neutral (M1a) versus selection (M2a) 20566Beta (M7) versus beta amp w 1 (M8) 2683Model A versus M1a (Test 1) 874Model A versus model anull (Test 2) 252
Matrix B One ratio (R1) versus two ratio (R2) 2348Two ratio (R2) versus three ratio (R3) 001Three ratio (R3) versus four ratio (R4) 01Neutral (M1a) versus selection (M2a) 367Beta (M7) versus beta amp w 1 (M8) 7054
Significant at the 95 level significant at the 99 level
Table 3B Class MADS-Box Protein Interactions as Determined by Yeast Two-Hybrid Assays in Camellia japonica
CjDEF-BD CjTM6-BD CjGLO1-BD CjGLO2-BD BD
CjDEF-AD thorn mdash mdash mdash mdashCjTM6-AD mdash mdash thorn mdash mdashCjGLO1-AD thorn thorn mdash mdash mdashCjGLO2-AD thorn mdash mdash mdash mdashAD mdash mdash mdash mdash mdash
2638 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
positive selection is not significantly better in explaining thedata than the model M1a and null model A (data notshown) When the two clades are treated together as fore-ground branches model A is significantly better than thenested site model M1a (test 1) suggesting that positive se-lection is acting in both clades that arose after duplicationBayes empirical Bayes identification resulted in seven po-sitions though all with posterior probability below 90The improved test 2 from Zhang et al (2005) resulted ina value 252 just beneath the critical value 271 at the5 level but far above the critical value at the 10 level164 These two tests support the hypothesis that positiveselection acts after the duplication event in both paralogousgene clades
Together the identification of changes in selectivepressure in time and along the coding sequence indicatethat upon duplication PI paralogs underwent an increasednonsynonymous substitution rate and that this occurred
specifically in the domains shown to be necessary forproteinndashprotein interactions
Diversification in the Interaction Behavior of B-ClassGenes After the Ancient Duplication Event in BasalAsterids
As sites in the interaction domains of the PI genesfrom basal asterids were detected to be positively selectedafter duplication we further investigated the possible func-tional divergence of the duplicated PI proteins by yeast-2-hybrid analyses We tested the interactions between thefour class B genes in C japonica Camellia japonica con-tains two AP3-like genes one belonging to the euAP3-typeand one from the paleoAP3-type and two PI-like genesfrom an ancient duplication event at the base of the EricalesThis situation is similar to the B-class configuration in thecore asterid species Petunia hybrida (Vandenbussche
FIG 5mdashAlignment of the K-domain and C-terminal domain of the PI duplicates from matrix A used for the analysis in PAML Differentsubdomains and the characteristic PI motif are indicated (Kramer et al 1998) Codon positions identified to be under positive selection are marked withgray boxes
PISTILLATAmdashDuplications in Asterids 2639
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

putatively lost after duplication we constructed a secondmatrix with only duplicated PI genes due to the lsquolsquoancientrsquorsquoduplication from G urceolata (Diapensiaceae) H diptera(Styracaceae) S zahlbruckneri (Actinidiaceae) S pseudo-camellia and C japonica (Theaceae) and S chinensis(Symplocaceae) The MP analysis resulted in just one mostparsimonious tree that is identical to the tree with the high-est likelihood As in the previous analysis the same twoparalogous clades were identified In this case both BPPvalues (100 and 97) and ML bootstrap values (57 and53) were higher in this case supporting a duplication eventpresumably at least before the divergence of these familiesand after the divergence of the primuloid clade in Ericalesphylogeny (Schonenberger et al 2006)
In addition to these two ancient duplication events weidentified several other more recent duplication events inasterid evolution From I aquifolium (Aquifoliaceae) weamplified two PI copies but more extensive sampling ofAquifoliales representatives should reveal the timing of thisduplication more accurately In Cornales duplicate genesare found in both Davidia involucrata and Nyssa sylvatica(Nyssaceae) suggesting that the duplication that resulted inthese copies occurred in the lineage leading to NyssaceaeIn the clade of Ericales representatives there are seven spe-cies with more than one PI copy which appear to be morerecent than the previously described duplication event atthe base of the Ericales order For example we isolatedtwo PI homologues from Impatiens glandulifera (IgGLO1and IgGLO2 Balsaminaceae) A previously characterizedPI gene from Impatiens hawkeri (IhGLO Geuten et al2006) groups with IgGLO2 but not with the PI gene fromM umbellata (MuGLO Marcgraviaceae) suggesting thatthis duplication occurred at the base of the BalsaminaceaeSimilarly we isolated twoPI genes fromG brasiliensis (Le-cythidaceae) and the phylogeny indicates that one copy ismore related to the PI gene from another Lecythidaceae rep-resentativeNapoleonaea vogeliiBecauseGustavia is basalin the phylogeny of Lecythidaceae (Mori et al 2007) the du-plication leading to paralogous copies inGustavia probablyoccurred at the origin of the Lecythidaceae or earlier
We also identified two PI copies in Erica hiemalis(Ericaceae) and Actinidia chinensis (Actinidiaceae) In bothS pseudocamellia (Theaceae) and S zahlbruckneri (Acti-nidiaceae) a third PI copy is present presumably as a resultof a more recent duplication in one of their paralogs As thesequence similarity of the paralogs in these four species ishigh (91ndash98) it is hard to decide whether these genes areat the same genetic locus or at different loci We suggestthese genes to be paralogous copies with a very recent or-igin and not shared by any other species in our samplingFinally two PI genes were identified in P denticulata (Pri-mulaceae) Although sequence similarity among thesegenes is significantly lower (82) this duplication is notshared by any other PI genes in our analyses indicating thatthis duplication is specific to the Primulaceae
Duplications in the AP3 and TM6 Lineages Are Few andNot Correlated with PI Duplications
We also carried out phylogenetic analyses of asteridAP3 sequences using a number of newly identified AP3
sequences as well as previously characterized genes andEST sequences from Genbank (table 1) The phylogenywas inferred from a nucleotide alignment of genes from85 taxa which was analyzed using both MP and Bayesianmethods (fig 2) In the Bayesian analysis AlAP3 was usedto root the AP3-like tree Similar to previous analyses(Kramer and Irish 1999 Kramer et al 2006) our Bayesiananalysis recovered two highly supported clades A firstclade contains the euAP3 homologues (BPP 98) whereasa second clade contains the TM6 homologues (BPP 100)(fig 2) The duplication event that gave rise to these two eu-dicot AP3 gene clades is placed near the base of the core eu-dicots and likely occurred coincident with the origin ofTrochodendraceae and Buxaceae (Kramer et al 2006)Within core asterids our phylogeny gave no indicationsofmajor duplication events but onlyfiveof eight core asteridorderswere sampled Similarly in basal asterids duplicationevents have seemingly not played an important role duringAP3 evolution because the observed gene phylogeny did notconflict with the species phylogeny (Schonenberger et al2006) In several cases thoughparalogousAP3genesofveryrecent origin were identified in four species one from Eri-cales (I hawkeri Geuten et al 2006) and three from core as-terids (Hieracium piloselloides Mimulus cultstatus andSalvia coccinea) (Aagaard et al 2005) As for thePI phylog-eny the bootstrap values in the MP analyses were low andthese analyses resulted in a poorly resolved tree
After the basal eudicot and rosid representativesbranch off in the euAP3 clade (BPP 98) the Cornales rep-resentatives are sister to two large clades The first cladecontains the newly sampled Ericales representatives(BPP 81) whereas the second contains all core asterid se-quences (BPP 54) The unexpected association of Brassica-ceae representatives (AP3 BjAP3) with the euAP3 genesfrom Asterales was also present in previous analyses andprobably illustrates the need for even denser sampling(de Martino et al 2006 Kramer et al 2006) Within theEricales euAP3 clade (BPP 81) most relationships arepoorly resolved However a strongly supported primuloid(BPP 100) ericoid (BPP 91) and balsaminoid clade (BPP100) can be observed (Schonenberger et al 2005) Withinthe TM6-clade Ericales representatives are supported witha BPP of 99 but overall support within this clade is lowHowever we did not recover any paralogous copies of TM6genes in the species sampled suggesting that duplicationevents in this clade either do not occur readily or are se-lected against We also observed an unexpected associationof Asterales TM6 representatives with rosids (BPP 71) asreported in previous analyses (de Martino et al 2006Kramer et al 2006) The TM6 genes within the Solanalesare strongly supported as a clade (BPP 100)
RT-PCR Reveals Divergence of Expression PatternsAfter Duplication
To investigate whether gene duplicates have divergedin expression patterns or have seemingly remained underthe same regulatory control we used RT-PCR to monitorgene expression levels in mature dissected floral organs(fig 3) We limited our survey to Ericales species thatcontain representatives of PI genes arising from both the
PISTILLATAmdashDuplications in Asterids 2633
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
ancient and recent duplications We find that the expressionpattern of PI paralogs has diverged considerably and fairlyrapidly The PI protein is known to require an AP3-like in-teraction partner for its in vivo function in a number of coreeudicots (Jack et al 1992 Trobner et al 1992 Goto andMeyerowitz 1994) We therefore also examined the expres-sion patterns of TM6 and euAP3 genes in the same species
We compared Ericales expression patterns with the as-terid speciesPetunia hybrida (Solanaceae) BothPI genesin Petunia show a very similar expression pattern (Vanden-bussche et al 2004) Expression of these twoPI homologues(FBP1 and PMADS2) is mainly confined to the second andthirdwhorls (Angenent et al 1992Van derKrol et al 1993)Expression of PhDEF the AP3 homologue was mainly
FIG 2mdashEvolutionary reconstruction of APETALA3-like gene phylogeny in asterids as inferred from Bayesian analysis Bayesian posteriorprobabilities are indicated above branches nonparametric bootstrap support values are shown below branches Gray circle5 inferred duplication event
2634 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
confined to petals and stamens with low expression in firstand fourth whorl organs PhTM6 has a different expressionprofilewithstrongexpressioninstamensandcarpelswhereasexpression in sepals and petals is low (Vandenbussche et al2004) Similar expression patterns to Petunia of PI AP3 and
TM6 were detected in S lycopersicon another core asteridSolanaceae species (de Martino et al 2006)
Multiple copies of PI have been retained in the ge-nomes of S pseudocamellia C japonica and H dipteraafter the major duplication event at the base of the Ericales
FIG 3mdashRT-PCR expression data for B-class genes from selected Ericales representatives Sep 5 sepals pet 5 petals pet up 5 upper part ofpetals fus 5 fused part of petals and stamens sta 5 stamens sta up 5 upper part of stamens sta fus 5 fused part of stamens gyn 5 gynoecium andflo 5 flower
PISTILLATAmdashDuplications in Asterids 2635
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Both copies in each of these species appear to have divergedin their expression patterns after duplication (fig 3AndashC)Although HdGLO2 is expressed in the petals and stamensof H diptera the other copy (HdGLO1) is mainly ex-pressed in stamens and the gynoecium and absent in petalsIn comparison the TM6 and euAP3 lineage genes show un-expected expression patterns with strong expression ofHdTM6 in all disected organs but HdDEF expression beingmainly restricted to petals and carpels InC japonica one PIparalog (CjGLO2) is expressed throughout sepals petalsand stamens whereas expression of the second paralog(CjGLO1) appeared to be restricted to the nonfused partsof petals stamens and carpels Although both CjDEF andCjTM6 expression are detected in all organs strong expres-sion ofCjDEF is seenmainly in the sepals and petalsCjTM6expression is strongest in all stamen parts and in carpelsS pseudocamellia also displays differences in the expressionof its two PI copies One paralog (SpGLO2) is expressed inpetals stamens carpels and expression of the second paral-og (SpGLO1) is more restricted to the fused part of petals andstamens and upper part of petals The AP3 homologueSpDEF is strongly expressed in all stamen parts and carpelswhereas strong expression of SpTM6 is detected in sepals allstamen parts and carpels
In summary in these three cases there is one PI copywith a broad expression pattern in the flower (even in sepalsand carpels) whereas expression of the other PI copy ismore restricted to specific parts of petals and stamensThese expression patterns differ from those of PI lineagegenes in core asterid species (Vandenbussche et al2004 de Martino et al 2006) where PI expression (alsoin a duplicated configuration) is mainly restricted to the sec-ond and third whorl organs petals and stamens and nostrong expression in the first and fourth whorl organs is de-tected Interestingly the divergence in expression patternsin these species does not follow the gene phylogeny Theparalogous copies with a general expression pattern are notgrouped in one clade of PI genes in Ericales but rather arefound in two clades This suggests that the PI expressiondomains were not fixed immediately after duplication in-dicating that subfunctionalization in spatial expression doesnot explain retention of the duplicates Alternatively ifregulatory subfunctionalization was the main reason forretention evidence for this has been obscured throughtime Differences in expression between AP3 and TM6in C japonica seem to be very similar to members ofthe same lineages in the core asterid species Expressionof AP3-like genes in H diptera and S pseudocamelliahowever differs from those of Camellia with strongAP3 expression in the carpels of Stewartia and Halesiaand weak expression of AP3 in stamens of Halesia In eachof these species expression of one or both AP3-like genes isdetected in the sepals
Sarracenia drummondii (fig 3D) Styrax japonicus(fig 3E) C persicum (fig 3F) and Phlox paniculata(fig 3G) maintained probably only one of the paralogousPI copies in their genome after the ancient Ericales dupli-cation Moreover Sarracenia and Styrax maintained a dif-ferent copy (fig 1 BPP 68) than did Cyclamen and Phlox(fig 1 BPP89) We investigated the expression patterns ofPI genes in these species to monitor the possible divergence
between the two paralogous PI clades after duplication Ineach of these four species PI is expressed in petals and sta-mens similar to the expression patterns of PI in core aster-ids in previous studies (Vandenbussche et al 2004 deMartino et al 2006) In both Sarracenia (SdGLO) andPhlox (PpGLO) there is some expression detected in thecarpels whereas SjGLO (Styrax) is also expressed in sepalsNo clear divergence in expression of PI from the two dif-ferent clades (fig 1 BPP68 and BPP89) is detected in thesecond and third whorl organs The expression patterns ofAP3 and TM6 lineage genes are different between these fourspecies Although in S drummondii both AP3 and TM6 areexpressed in all four floral whorls the expression in Styraxis very similar to that of TM6 and AP3 in core asterid spe-cies with in addition to expression in petals and stamensexpression of TM6 in carpels and AP3 in sepals In Cycla-men TM6 expression is absent in the upper part of the sta-mens and strong expression of AP3 is detected in thecarpels In Phlox however TM6 expression is absent fromthe carpels
The multiple PI copies in G brasiliensis (Lecythida-ceae) and P acaulis (Primulaceae) are the result of a recentduplication probably at the base of these two families(fig 1) We looked at the expression pattern of each of theseduplicated genes and of the AP3-like genes in these speciesIn G brasiliensis GbGLO2 is expressed in petals and sta-mens whereas GbGLO1 is mainly expressed in the stami-nal parts and less in the petals (fig 3H) Expression of AP3and TM6 seems to be lsquolsquoinversersquorsquo compared with the ex-pected asterid expression with AP3 expression mainly re-stricted to the stamens and strong TM6 expression in thepetals No AP3-like expression is detected in the carpelsIn P acaulis there are differences between the expressionpatterns of both PI copies (fig 3I) PaGLO2 is expressed inall organs tested whereas the expression of PaGLO1 ismainly restricted to the sepals and petals Both AP3 andTM6 are strongly expressed in sepals and as inC persicumTM6-expression is weak in the staminal parts Even afterthese recent duplication events divergence in the expres-sion domain of the two PI copies is detected
In contrast to the duplicated PI genes of Petunia(Vandenbussche et al 2004) the expression domains ofPI genes have changed considerably after duplication inEricales For both ancient and several recent duplicationevents divergence in expression was detected indicatingthat the regulation of the duplicate copies has divergedIn the studied species where only one PI copy was isolatedit appears that an lsquolsquoancestralrsquorsquo expression pattern of PI isreestablished However a second copy could have beenmissed during amplification Also within the AP3-like lin-eage some derived expression patterns are detected
In Situ Hybridization Demonstrates ExpressionDivergence in Early Flower Development
To analyze the expression patterns of the PI genes afterduplication in early developmental stages we examined thespatio-temporal expression patterns of PI in selected Eri-cales species by in situ hybridization (fig 4) We selectedexamples of each type of fate after duplication retention
2636 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
of both ancient duplicates (Stewartia) retention of only oneof each PI paralog (C persicum and S rubra) and retentionof recent duplicates (P acaulis)
In S rostrata (fig 4AndashF) we could detect expressiondivergence between each of the PI copies at early stages offloral development At the stage when petal primordia areevident but stamen primordia are apparent only as smallmeristematic outgrowths SpGLO1 expression is detectedin petal and stamen primordia (fig 4A) whereas SpGLO2transcripts are also detected in the tip of the sepals (fig 4D)As the flower develops SpGLO2 expression is low butexpands to include developing carpels (fig 4E) and expres-sion is maintained in the carpels in later stages (fig 4F) Incontrast SpGLO1 expression is maintained at high levelsonly in petals and stamens through later stages (fig 4Band C) and does not expand into sepals and carpels Theseearly patterns of S rostrata PI gene expression are consis-tent with the later patterns of expression as detected byRT-PCR (fig 3I)
Expression patterns of PI homologues were furtherinvestigated in C persicum (fig 4GndashI) and S rubra(fig 4JndashK) CpGLO expression first appears in the youngpetal primordia (fig 4G) followed by expression in the sta-men primordia as they emerge (fig 4H) Expression inthese organs is maintained in later stages (fig 4I) In con-trast to the expression patterns in mature organs as docu-mented by RT-PCR weak expression in the developingsepals (fig 4H) and developing carpels (fig 4I) was de-tected For S rubra we were not able to obtain the earliestdevelopmental stages For later stages of floral develop-ment we observed strong expression of PI in petals andstamens (fig 4J) Weak expression of SdGLO was also de-tected in the developing carpels (fig 4K) which is consis-tent with the RT-PCR data (fig 3) These observationstogether with the RT-PCR data (fig 3) support the ideathat species with presumably only one PI copy in their ge-nome show expression that is more confined to petals andstamens
Finally we investigated the expression of recent PIduplicates in P acaulis (fig 4LndashQ) At early stages bothPaGLO1 (fig 4L) and PaGLO2 (fig 4O) are expressedacross the meristematic dome in sepal primordia commonpetalndashstamen primordia and carpel primordia Expressionof PaGLO2 is maintained in all organs at older stages(fig 4R and Q) whereas PaGLO1 expression is restrictedto the stamens (fig 4M and N)
The in situ expression data (fig 4) in comparison tothe RT-PCR data (fig 3) indicate that early patterns of ex-pression of PI genes are often developmentally regulatedwith different patterns of expression being observed early incomparison to later in flower development Nonethelessboth sets of data indicate that in species possessing dupli-cate PI genes divergence in expression patterns betweenthe two copies is a common theme
Positive Selection and Relaxed Purifying Selection ActsAfter the Duplication Event in Ericales in the K- and C-Cerminal Domain
The mRNA expression data indicate that duplicate PIparalogs have acquired different expression patterns sug-gesting independent evolution of their transcriptional reg-ulatory control It was also demonstrated that adaptiveevolution in the coding regions of MADS-box transcriptionfactors has played a role in the diversification of thisgene family (Martinez-Castilla and Alvarez-Buylla2003) Within class B genes Hernandez-Hernandez et al(2007) demonstrated the role of positive selection after crit-ical duplication events like the AP3-PI duplication and theeuAP3ndashTM6 duplication To investigate whether parallelchanges occurred in the coding regions of the genes exam-ined in this study we investigated the nonsynonymous ver-sus synonymous amino acid replacement rates forsignificant increases before and after duplication in thespecific paralogous clades and along the coding sequenceThe results of likelihood ratio tests are shown in table 2 and
FIG 4mdashIn situ hybridization data for PI genes in various developmental stages of Stewartia rostrata (AndashC SpGLO1 DndashF SpGLO2) Cyclamenpersicum (GndashI CpGLO) Sarracenia rubra (JndashK SdGLO) Primula acaulis (LndashN PaGLO1 OndashQ PaGLO2) lsquolsquoprsquorsquo indicates petalprimordia lsquolsquosrsquorsquoindicates stamen primordia
PISTILLATAmdashDuplications in Asterids 2637
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
specific sites are marked in the alignment presented infigure 5 Parameter estimates log-likelihood scores andBayes Empirical Bayes identified sites are listed in supple-mentary table S2 Supplementary Material online
Comparison of the likelihood scores calculated usingbranch models R1 and R2 for matrix A indicates a signifi-cantly increased evolutionary rate following the ancient du-plication event in Ericales The dNdS estimate from thetwo-ratio branch model after duplication (x 5 0285) istwice compared with the value before duplication (x 50113) illustrating the relaxed selective regime resultingfrom the duplication event However the same test appliedon matrix B did not reveal this change in selective pressureOne possible explanation is that by adding the single-copylineages that evolved after duplication the observed patternin matrix A for the duplicates is obscured In addition wefound no evidence that the two paralogous gene cladescould be evolving at different rates because the three-and four-ratio models did not result in a significantly in-creased log-likelihood as calculated for either of the datamatrices
To formally test for the presence of positive selectionon sequence sites we used the site models proposed byNielsen and Yang (1998) and Yang et al (2000) Analysisof matrix A shows that both M2a and M8 two models thatallow for positive selection fit the data significantly betterthan the corresponding neutral models (M1a and M7) Sub-stitution patterns in about 5 of the sites for the M2amodel and about 9 of the sites for the M8 model appearto result from adaptive evolution Here also analysis of ma-trix B suggests positively selected sites M8 fits the databetter than the neutral model M7 with an estimation of
10 sites under positive selection Bayes empirical Bayes(Yang et al 2005) identification of the sites under positiveselection in matrix A finds seven sites to be positively se-lected under model M8 two of these sites have a posteriorprobability higher than 99 Within matrix B only twopositively selected sites were detected which were alsoidentified in matrix A both with posterior probability high-er than 91 Interestingly all the identified sites are locatedwithin the K2-subdomain the interhelical region betweenK2 and K3 the K3-subdomain and the C-terminal domain(fig 5) These functional domains have been shown to beimportant for proteinndashprotein interactions among Arabi-dopsis MADS-domain proteins (Ma et al 1991 Pnueliet al 1991 Shore and Sharrocks 1995 Riechmann et al1996 Yang et al 2003) The strength of the AP3ndashPI inter-action is mediated primarily by K1 and K2 whereas thePIndashSEP interaction is mediated by K2 the amino-terminalportion of K3 and possibly the C-domain (Yang et al 2003Yang and Jack 2004) The C-terminal region normallyshows much lower sequence conservation but containsshort highly conserved motifs (Lamb and Irish 2003) Sev-eral studies have suggested that this domain is involvedin the formation of higher order protein complexes(Egea-Cortines et al 1999 Honma and Goto 2001) yetthe specific function of the class B C-terminal domainsis unclear (Lamb and Irish 2003 Piwarzyk et al 2007)None of the sites identified by Yang et al (2003) to be re-sponsible for the direct interaction between APETALA3and PISTILLATA are positively selected for in our dataset However site 49 contains a hydrophobic amino acidthat when mutated disrupts the alpha-helical secondarystructure (Yang et al 2003) In addition sites 49 and site60 in figure 5 were identified by Yang and Jack (2004) to becritical for PIndashSEP interaction Site 49 is situated withinthe K2 subdomain which is required for both AP3ndashPIand PIndashSEP interaction whereas site 60 which is locatedat the amino-terminal end of K3 affects when mutated thestrength of the PIndashSEP interaction
To identify evolutionary pressures acting on specificsites along certain branches and in clades we calculatedlikelihoods using the branch-site models First we specifiedthe two branches immediately following the duplication asforeground branches in matrix A Branch-site models thatallow for positive selection in these foreground brancheswere not better at explaining the data than the nested sitemodels M1a and the null model A and indicate no evidencefor a burst of positive selection in the branches immediatelyfollowing the duplication event (Bielawski and Yang 2003)(data not shown) Second we treated all branches in eitherclade of paralogs as foreground branches When these twoclades are treated separately the model allowing for
Table 2Likelihood Ratio Tests Comparing Branch-Models (R1ndashR4)Site Models (M1ndashM8) and Branch-Site Models (Model AndashModel A Null)
Matrix Models Compared2(ln L1 ln L0)
Matrix A One ratio (R1) versus two ratio (R2) 18174Two ratio (R2) versus three ratio (R3) 0435Three ratio (R3) versus four ratio (R4) 1298Neutral (M1a) versus selection (M2a) 20566Beta (M7) versus beta amp w 1 (M8) 2683Model A versus M1a (Test 1) 874Model A versus model anull (Test 2) 252
Matrix B One ratio (R1) versus two ratio (R2) 2348Two ratio (R2) versus three ratio (R3) 001Three ratio (R3) versus four ratio (R4) 01Neutral (M1a) versus selection (M2a) 367Beta (M7) versus beta amp w 1 (M8) 7054
Significant at the 95 level significant at the 99 level
Table 3B Class MADS-Box Protein Interactions as Determined by Yeast Two-Hybrid Assays in Camellia japonica
CjDEF-BD CjTM6-BD CjGLO1-BD CjGLO2-BD BD
CjDEF-AD thorn mdash mdash mdash mdashCjTM6-AD mdash mdash thorn mdash mdashCjGLO1-AD thorn thorn mdash mdash mdashCjGLO2-AD thorn mdash mdash mdash mdashAD mdash mdash mdash mdash mdash
2638 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
positive selection is not significantly better in explaining thedata than the model M1a and null model A (data notshown) When the two clades are treated together as fore-ground branches model A is significantly better than thenested site model M1a (test 1) suggesting that positive se-lection is acting in both clades that arose after duplicationBayes empirical Bayes identification resulted in seven po-sitions though all with posterior probability below 90The improved test 2 from Zhang et al (2005) resulted ina value 252 just beneath the critical value 271 at the5 level but far above the critical value at the 10 level164 These two tests support the hypothesis that positiveselection acts after the duplication event in both paralogousgene clades
Together the identification of changes in selectivepressure in time and along the coding sequence indicatethat upon duplication PI paralogs underwent an increasednonsynonymous substitution rate and that this occurred
specifically in the domains shown to be necessary forproteinndashprotein interactions
Diversification in the Interaction Behavior of B-ClassGenes After the Ancient Duplication Event in BasalAsterids
As sites in the interaction domains of the PI genesfrom basal asterids were detected to be positively selectedafter duplication we further investigated the possible func-tional divergence of the duplicated PI proteins by yeast-2-hybrid analyses We tested the interactions between thefour class B genes in C japonica Camellia japonica con-tains two AP3-like genes one belonging to the euAP3-typeand one from the paleoAP3-type and two PI-like genesfrom an ancient duplication event at the base of the EricalesThis situation is similar to the B-class configuration in thecore asterid species Petunia hybrida (Vandenbussche
FIG 5mdashAlignment of the K-domain and C-terminal domain of the PI duplicates from matrix A used for the analysis in PAML Differentsubdomains and the characteristic PI motif are indicated (Kramer et al 1998) Codon positions identified to be under positive selection are marked withgray boxes
PISTILLATAmdashDuplications in Asterids 2639
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

ancient and recent duplications We find that the expressionpattern of PI paralogs has diverged considerably and fairlyrapidly The PI protein is known to require an AP3-like in-teraction partner for its in vivo function in a number of coreeudicots (Jack et al 1992 Trobner et al 1992 Goto andMeyerowitz 1994) We therefore also examined the expres-sion patterns of TM6 and euAP3 genes in the same species
We compared Ericales expression patterns with the as-terid speciesPetunia hybrida (Solanaceae) BothPI genesin Petunia show a very similar expression pattern (Vanden-bussche et al 2004) Expression of these twoPI homologues(FBP1 and PMADS2) is mainly confined to the second andthirdwhorls (Angenent et al 1992Van derKrol et al 1993)Expression of PhDEF the AP3 homologue was mainly
FIG 2mdashEvolutionary reconstruction of APETALA3-like gene phylogeny in asterids as inferred from Bayesian analysis Bayesian posteriorprobabilities are indicated above branches nonparametric bootstrap support values are shown below branches Gray circle5 inferred duplication event
2634 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
confined to petals and stamens with low expression in firstand fourth whorl organs PhTM6 has a different expressionprofilewithstrongexpressioninstamensandcarpelswhereasexpression in sepals and petals is low (Vandenbussche et al2004) Similar expression patterns to Petunia of PI AP3 and
TM6 were detected in S lycopersicon another core asteridSolanaceae species (de Martino et al 2006)
Multiple copies of PI have been retained in the ge-nomes of S pseudocamellia C japonica and H dipteraafter the major duplication event at the base of the Ericales
FIG 3mdashRT-PCR expression data for B-class genes from selected Ericales representatives Sep 5 sepals pet 5 petals pet up 5 upper part ofpetals fus 5 fused part of petals and stamens sta 5 stamens sta up 5 upper part of stamens sta fus 5 fused part of stamens gyn 5 gynoecium andflo 5 flower
PISTILLATAmdashDuplications in Asterids 2635
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Both copies in each of these species appear to have divergedin their expression patterns after duplication (fig 3AndashC)Although HdGLO2 is expressed in the petals and stamensof H diptera the other copy (HdGLO1) is mainly ex-pressed in stamens and the gynoecium and absent in petalsIn comparison the TM6 and euAP3 lineage genes show un-expected expression patterns with strong expression ofHdTM6 in all disected organs but HdDEF expression beingmainly restricted to petals and carpels InC japonica one PIparalog (CjGLO2) is expressed throughout sepals petalsand stamens whereas expression of the second paralog(CjGLO1) appeared to be restricted to the nonfused partsof petals stamens and carpels Although both CjDEF andCjTM6 expression are detected in all organs strong expres-sion ofCjDEF is seenmainly in the sepals and petalsCjTM6expression is strongest in all stamen parts and in carpelsS pseudocamellia also displays differences in the expressionof its two PI copies One paralog (SpGLO2) is expressed inpetals stamens carpels and expression of the second paral-og (SpGLO1) is more restricted to the fused part of petals andstamens and upper part of petals The AP3 homologueSpDEF is strongly expressed in all stamen parts and carpelswhereas strong expression of SpTM6 is detected in sepals allstamen parts and carpels
In summary in these three cases there is one PI copywith a broad expression pattern in the flower (even in sepalsand carpels) whereas expression of the other PI copy ismore restricted to specific parts of petals and stamensThese expression patterns differ from those of PI lineagegenes in core asterid species (Vandenbussche et al2004 de Martino et al 2006) where PI expression (alsoin a duplicated configuration) is mainly restricted to the sec-ond and third whorl organs petals and stamens and nostrong expression in the first and fourth whorl organs is de-tected Interestingly the divergence in expression patternsin these species does not follow the gene phylogeny Theparalogous copies with a general expression pattern are notgrouped in one clade of PI genes in Ericales but rather arefound in two clades This suggests that the PI expressiondomains were not fixed immediately after duplication in-dicating that subfunctionalization in spatial expression doesnot explain retention of the duplicates Alternatively ifregulatory subfunctionalization was the main reason forretention evidence for this has been obscured throughtime Differences in expression between AP3 and TM6in C japonica seem to be very similar to members ofthe same lineages in the core asterid species Expressionof AP3-like genes in H diptera and S pseudocamelliahowever differs from those of Camellia with strongAP3 expression in the carpels of Stewartia and Halesiaand weak expression of AP3 in stamens of Halesia In eachof these species expression of one or both AP3-like genes isdetected in the sepals
Sarracenia drummondii (fig 3D) Styrax japonicus(fig 3E) C persicum (fig 3F) and Phlox paniculata(fig 3G) maintained probably only one of the paralogousPI copies in their genome after the ancient Ericales dupli-cation Moreover Sarracenia and Styrax maintained a dif-ferent copy (fig 1 BPP 68) than did Cyclamen and Phlox(fig 1 BPP89) We investigated the expression patterns ofPI genes in these species to monitor the possible divergence
between the two paralogous PI clades after duplication Ineach of these four species PI is expressed in petals and sta-mens similar to the expression patterns of PI in core aster-ids in previous studies (Vandenbussche et al 2004 deMartino et al 2006) In both Sarracenia (SdGLO) andPhlox (PpGLO) there is some expression detected in thecarpels whereas SjGLO (Styrax) is also expressed in sepalsNo clear divergence in expression of PI from the two dif-ferent clades (fig 1 BPP68 and BPP89) is detected in thesecond and third whorl organs The expression patterns ofAP3 and TM6 lineage genes are different between these fourspecies Although in S drummondii both AP3 and TM6 areexpressed in all four floral whorls the expression in Styraxis very similar to that of TM6 and AP3 in core asterid spe-cies with in addition to expression in petals and stamensexpression of TM6 in carpels and AP3 in sepals In Cycla-men TM6 expression is absent in the upper part of the sta-mens and strong expression of AP3 is detected in thecarpels In Phlox however TM6 expression is absent fromthe carpels
The multiple PI copies in G brasiliensis (Lecythida-ceae) and P acaulis (Primulaceae) are the result of a recentduplication probably at the base of these two families(fig 1) We looked at the expression pattern of each of theseduplicated genes and of the AP3-like genes in these speciesIn G brasiliensis GbGLO2 is expressed in petals and sta-mens whereas GbGLO1 is mainly expressed in the stami-nal parts and less in the petals (fig 3H) Expression of AP3and TM6 seems to be lsquolsquoinversersquorsquo compared with the ex-pected asterid expression with AP3 expression mainly re-stricted to the stamens and strong TM6 expression in thepetals No AP3-like expression is detected in the carpelsIn P acaulis there are differences between the expressionpatterns of both PI copies (fig 3I) PaGLO2 is expressed inall organs tested whereas the expression of PaGLO1 ismainly restricted to the sepals and petals Both AP3 andTM6 are strongly expressed in sepals and as inC persicumTM6-expression is weak in the staminal parts Even afterthese recent duplication events divergence in the expres-sion domain of the two PI copies is detected
In contrast to the duplicated PI genes of Petunia(Vandenbussche et al 2004) the expression domains ofPI genes have changed considerably after duplication inEricales For both ancient and several recent duplicationevents divergence in expression was detected indicatingthat the regulation of the duplicate copies has divergedIn the studied species where only one PI copy was isolatedit appears that an lsquolsquoancestralrsquorsquo expression pattern of PI isreestablished However a second copy could have beenmissed during amplification Also within the AP3-like lin-eage some derived expression patterns are detected
In Situ Hybridization Demonstrates ExpressionDivergence in Early Flower Development
To analyze the expression patterns of the PI genes afterduplication in early developmental stages we examined thespatio-temporal expression patterns of PI in selected Eri-cales species by in situ hybridization (fig 4) We selectedexamples of each type of fate after duplication retention
2636 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
of both ancient duplicates (Stewartia) retention of only oneof each PI paralog (C persicum and S rubra) and retentionof recent duplicates (P acaulis)
In S rostrata (fig 4AndashF) we could detect expressiondivergence between each of the PI copies at early stages offloral development At the stage when petal primordia areevident but stamen primordia are apparent only as smallmeristematic outgrowths SpGLO1 expression is detectedin petal and stamen primordia (fig 4A) whereas SpGLO2transcripts are also detected in the tip of the sepals (fig 4D)As the flower develops SpGLO2 expression is low butexpands to include developing carpels (fig 4E) and expres-sion is maintained in the carpels in later stages (fig 4F) Incontrast SpGLO1 expression is maintained at high levelsonly in petals and stamens through later stages (fig 4Band C) and does not expand into sepals and carpels Theseearly patterns of S rostrata PI gene expression are consis-tent with the later patterns of expression as detected byRT-PCR (fig 3I)
Expression patterns of PI homologues were furtherinvestigated in C persicum (fig 4GndashI) and S rubra(fig 4JndashK) CpGLO expression first appears in the youngpetal primordia (fig 4G) followed by expression in the sta-men primordia as they emerge (fig 4H) Expression inthese organs is maintained in later stages (fig 4I) In con-trast to the expression patterns in mature organs as docu-mented by RT-PCR weak expression in the developingsepals (fig 4H) and developing carpels (fig 4I) was de-tected For S rubra we were not able to obtain the earliestdevelopmental stages For later stages of floral develop-ment we observed strong expression of PI in petals andstamens (fig 4J) Weak expression of SdGLO was also de-tected in the developing carpels (fig 4K) which is consis-tent with the RT-PCR data (fig 3) These observationstogether with the RT-PCR data (fig 3) support the ideathat species with presumably only one PI copy in their ge-nome show expression that is more confined to petals andstamens
Finally we investigated the expression of recent PIduplicates in P acaulis (fig 4LndashQ) At early stages bothPaGLO1 (fig 4L) and PaGLO2 (fig 4O) are expressedacross the meristematic dome in sepal primordia commonpetalndashstamen primordia and carpel primordia Expressionof PaGLO2 is maintained in all organs at older stages(fig 4R and Q) whereas PaGLO1 expression is restrictedto the stamens (fig 4M and N)
The in situ expression data (fig 4) in comparison tothe RT-PCR data (fig 3) indicate that early patterns of ex-pression of PI genes are often developmentally regulatedwith different patterns of expression being observed early incomparison to later in flower development Nonethelessboth sets of data indicate that in species possessing dupli-cate PI genes divergence in expression patterns betweenthe two copies is a common theme
Positive Selection and Relaxed Purifying Selection ActsAfter the Duplication Event in Ericales in the K- and C-Cerminal Domain
The mRNA expression data indicate that duplicate PIparalogs have acquired different expression patterns sug-gesting independent evolution of their transcriptional reg-ulatory control It was also demonstrated that adaptiveevolution in the coding regions of MADS-box transcriptionfactors has played a role in the diversification of thisgene family (Martinez-Castilla and Alvarez-Buylla2003) Within class B genes Hernandez-Hernandez et al(2007) demonstrated the role of positive selection after crit-ical duplication events like the AP3-PI duplication and theeuAP3ndashTM6 duplication To investigate whether parallelchanges occurred in the coding regions of the genes exam-ined in this study we investigated the nonsynonymous ver-sus synonymous amino acid replacement rates forsignificant increases before and after duplication in thespecific paralogous clades and along the coding sequenceThe results of likelihood ratio tests are shown in table 2 and
FIG 4mdashIn situ hybridization data for PI genes in various developmental stages of Stewartia rostrata (AndashC SpGLO1 DndashF SpGLO2) Cyclamenpersicum (GndashI CpGLO) Sarracenia rubra (JndashK SdGLO) Primula acaulis (LndashN PaGLO1 OndashQ PaGLO2) lsquolsquoprsquorsquo indicates petalprimordia lsquolsquosrsquorsquoindicates stamen primordia
PISTILLATAmdashDuplications in Asterids 2637
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
specific sites are marked in the alignment presented infigure 5 Parameter estimates log-likelihood scores andBayes Empirical Bayes identified sites are listed in supple-mentary table S2 Supplementary Material online
Comparison of the likelihood scores calculated usingbranch models R1 and R2 for matrix A indicates a signifi-cantly increased evolutionary rate following the ancient du-plication event in Ericales The dNdS estimate from thetwo-ratio branch model after duplication (x 5 0285) istwice compared with the value before duplication (x 50113) illustrating the relaxed selective regime resultingfrom the duplication event However the same test appliedon matrix B did not reveal this change in selective pressureOne possible explanation is that by adding the single-copylineages that evolved after duplication the observed patternin matrix A for the duplicates is obscured In addition wefound no evidence that the two paralogous gene cladescould be evolving at different rates because the three-and four-ratio models did not result in a significantly in-creased log-likelihood as calculated for either of the datamatrices
To formally test for the presence of positive selectionon sequence sites we used the site models proposed byNielsen and Yang (1998) and Yang et al (2000) Analysisof matrix A shows that both M2a and M8 two models thatallow for positive selection fit the data significantly betterthan the corresponding neutral models (M1a and M7) Sub-stitution patterns in about 5 of the sites for the M2amodel and about 9 of the sites for the M8 model appearto result from adaptive evolution Here also analysis of ma-trix B suggests positively selected sites M8 fits the databetter than the neutral model M7 with an estimation of
10 sites under positive selection Bayes empirical Bayes(Yang et al 2005) identification of the sites under positiveselection in matrix A finds seven sites to be positively se-lected under model M8 two of these sites have a posteriorprobability higher than 99 Within matrix B only twopositively selected sites were detected which were alsoidentified in matrix A both with posterior probability high-er than 91 Interestingly all the identified sites are locatedwithin the K2-subdomain the interhelical region betweenK2 and K3 the K3-subdomain and the C-terminal domain(fig 5) These functional domains have been shown to beimportant for proteinndashprotein interactions among Arabi-dopsis MADS-domain proteins (Ma et al 1991 Pnueliet al 1991 Shore and Sharrocks 1995 Riechmann et al1996 Yang et al 2003) The strength of the AP3ndashPI inter-action is mediated primarily by K1 and K2 whereas thePIndashSEP interaction is mediated by K2 the amino-terminalportion of K3 and possibly the C-domain (Yang et al 2003Yang and Jack 2004) The C-terminal region normallyshows much lower sequence conservation but containsshort highly conserved motifs (Lamb and Irish 2003) Sev-eral studies have suggested that this domain is involvedin the formation of higher order protein complexes(Egea-Cortines et al 1999 Honma and Goto 2001) yetthe specific function of the class B C-terminal domainsis unclear (Lamb and Irish 2003 Piwarzyk et al 2007)None of the sites identified by Yang et al (2003) to be re-sponsible for the direct interaction between APETALA3and PISTILLATA are positively selected for in our dataset However site 49 contains a hydrophobic amino acidthat when mutated disrupts the alpha-helical secondarystructure (Yang et al 2003) In addition sites 49 and site60 in figure 5 were identified by Yang and Jack (2004) to becritical for PIndashSEP interaction Site 49 is situated withinthe K2 subdomain which is required for both AP3ndashPIand PIndashSEP interaction whereas site 60 which is locatedat the amino-terminal end of K3 affects when mutated thestrength of the PIndashSEP interaction
To identify evolutionary pressures acting on specificsites along certain branches and in clades we calculatedlikelihoods using the branch-site models First we specifiedthe two branches immediately following the duplication asforeground branches in matrix A Branch-site models thatallow for positive selection in these foreground brancheswere not better at explaining the data than the nested sitemodels M1a and the null model A and indicate no evidencefor a burst of positive selection in the branches immediatelyfollowing the duplication event (Bielawski and Yang 2003)(data not shown) Second we treated all branches in eitherclade of paralogs as foreground branches When these twoclades are treated separately the model allowing for
Table 2Likelihood Ratio Tests Comparing Branch-Models (R1ndashR4)Site Models (M1ndashM8) and Branch-Site Models (Model AndashModel A Null)
Matrix Models Compared2(ln L1 ln L0)
Matrix A One ratio (R1) versus two ratio (R2) 18174Two ratio (R2) versus three ratio (R3) 0435Three ratio (R3) versus four ratio (R4) 1298Neutral (M1a) versus selection (M2a) 20566Beta (M7) versus beta amp w 1 (M8) 2683Model A versus M1a (Test 1) 874Model A versus model anull (Test 2) 252
Matrix B One ratio (R1) versus two ratio (R2) 2348Two ratio (R2) versus three ratio (R3) 001Three ratio (R3) versus four ratio (R4) 01Neutral (M1a) versus selection (M2a) 367Beta (M7) versus beta amp w 1 (M8) 7054
Significant at the 95 level significant at the 99 level
Table 3B Class MADS-Box Protein Interactions as Determined by Yeast Two-Hybrid Assays in Camellia japonica
CjDEF-BD CjTM6-BD CjGLO1-BD CjGLO2-BD BD
CjDEF-AD thorn mdash mdash mdash mdashCjTM6-AD mdash mdash thorn mdash mdashCjGLO1-AD thorn thorn mdash mdash mdashCjGLO2-AD thorn mdash mdash mdash mdashAD mdash mdash mdash mdash mdash
2638 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
positive selection is not significantly better in explaining thedata than the model M1a and null model A (data notshown) When the two clades are treated together as fore-ground branches model A is significantly better than thenested site model M1a (test 1) suggesting that positive se-lection is acting in both clades that arose after duplicationBayes empirical Bayes identification resulted in seven po-sitions though all with posterior probability below 90The improved test 2 from Zhang et al (2005) resulted ina value 252 just beneath the critical value 271 at the5 level but far above the critical value at the 10 level164 These two tests support the hypothesis that positiveselection acts after the duplication event in both paralogousgene clades
Together the identification of changes in selectivepressure in time and along the coding sequence indicatethat upon duplication PI paralogs underwent an increasednonsynonymous substitution rate and that this occurred
specifically in the domains shown to be necessary forproteinndashprotein interactions
Diversification in the Interaction Behavior of B-ClassGenes After the Ancient Duplication Event in BasalAsterids
As sites in the interaction domains of the PI genesfrom basal asterids were detected to be positively selectedafter duplication we further investigated the possible func-tional divergence of the duplicated PI proteins by yeast-2-hybrid analyses We tested the interactions between thefour class B genes in C japonica Camellia japonica con-tains two AP3-like genes one belonging to the euAP3-typeand one from the paleoAP3-type and two PI-like genesfrom an ancient duplication event at the base of the EricalesThis situation is similar to the B-class configuration in thecore asterid species Petunia hybrida (Vandenbussche
FIG 5mdashAlignment of the K-domain and C-terminal domain of the PI duplicates from matrix A used for the analysis in PAML Differentsubdomains and the characteristic PI motif are indicated (Kramer et al 1998) Codon positions identified to be under positive selection are marked withgray boxes
PISTILLATAmdashDuplications in Asterids 2639
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
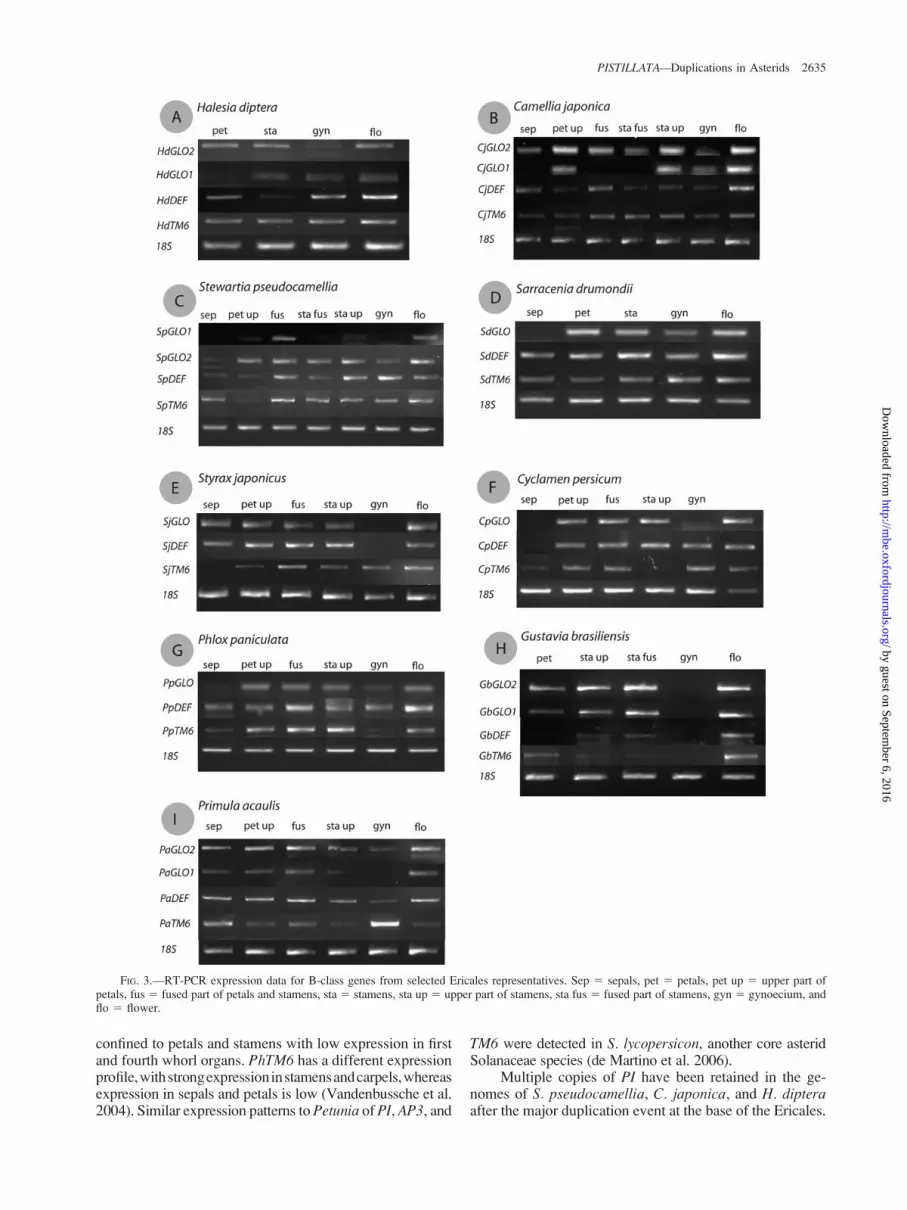
confined to petals and stamens with low expression in firstand fourth whorl organs PhTM6 has a different expressionprofilewithstrongexpressioninstamensandcarpelswhereasexpression in sepals and petals is low (Vandenbussche et al2004) Similar expression patterns to Petunia of PI AP3 and
TM6 were detected in S lycopersicon another core asteridSolanaceae species (de Martino et al 2006)
Multiple copies of PI have been retained in the ge-nomes of S pseudocamellia C japonica and H dipteraafter the major duplication event at the base of the Ericales
FIG 3mdashRT-PCR expression data for B-class genes from selected Ericales representatives Sep 5 sepals pet 5 petals pet up 5 upper part ofpetals fus 5 fused part of petals and stamens sta 5 stamens sta up 5 upper part of stamens sta fus 5 fused part of stamens gyn 5 gynoecium andflo 5 flower
PISTILLATAmdashDuplications in Asterids 2635
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Both copies in each of these species appear to have divergedin their expression patterns after duplication (fig 3AndashC)Although HdGLO2 is expressed in the petals and stamensof H diptera the other copy (HdGLO1) is mainly ex-pressed in stamens and the gynoecium and absent in petalsIn comparison the TM6 and euAP3 lineage genes show un-expected expression patterns with strong expression ofHdTM6 in all disected organs but HdDEF expression beingmainly restricted to petals and carpels InC japonica one PIparalog (CjGLO2) is expressed throughout sepals petalsand stamens whereas expression of the second paralog(CjGLO1) appeared to be restricted to the nonfused partsof petals stamens and carpels Although both CjDEF andCjTM6 expression are detected in all organs strong expres-sion ofCjDEF is seenmainly in the sepals and petalsCjTM6expression is strongest in all stamen parts and in carpelsS pseudocamellia also displays differences in the expressionof its two PI copies One paralog (SpGLO2) is expressed inpetals stamens carpels and expression of the second paral-og (SpGLO1) is more restricted to the fused part of petals andstamens and upper part of petals The AP3 homologueSpDEF is strongly expressed in all stamen parts and carpelswhereas strong expression of SpTM6 is detected in sepals allstamen parts and carpels
In summary in these three cases there is one PI copywith a broad expression pattern in the flower (even in sepalsand carpels) whereas expression of the other PI copy ismore restricted to specific parts of petals and stamensThese expression patterns differ from those of PI lineagegenes in core asterid species (Vandenbussche et al2004 de Martino et al 2006) where PI expression (alsoin a duplicated configuration) is mainly restricted to the sec-ond and third whorl organs petals and stamens and nostrong expression in the first and fourth whorl organs is de-tected Interestingly the divergence in expression patternsin these species does not follow the gene phylogeny Theparalogous copies with a general expression pattern are notgrouped in one clade of PI genes in Ericales but rather arefound in two clades This suggests that the PI expressiondomains were not fixed immediately after duplication in-dicating that subfunctionalization in spatial expression doesnot explain retention of the duplicates Alternatively ifregulatory subfunctionalization was the main reason forretention evidence for this has been obscured throughtime Differences in expression between AP3 and TM6in C japonica seem to be very similar to members ofthe same lineages in the core asterid species Expressionof AP3-like genes in H diptera and S pseudocamelliahowever differs from those of Camellia with strongAP3 expression in the carpels of Stewartia and Halesiaand weak expression of AP3 in stamens of Halesia In eachof these species expression of one or both AP3-like genes isdetected in the sepals
Sarracenia drummondii (fig 3D) Styrax japonicus(fig 3E) C persicum (fig 3F) and Phlox paniculata(fig 3G) maintained probably only one of the paralogousPI copies in their genome after the ancient Ericales dupli-cation Moreover Sarracenia and Styrax maintained a dif-ferent copy (fig 1 BPP 68) than did Cyclamen and Phlox(fig 1 BPP89) We investigated the expression patterns ofPI genes in these species to monitor the possible divergence
between the two paralogous PI clades after duplication Ineach of these four species PI is expressed in petals and sta-mens similar to the expression patterns of PI in core aster-ids in previous studies (Vandenbussche et al 2004 deMartino et al 2006) In both Sarracenia (SdGLO) andPhlox (PpGLO) there is some expression detected in thecarpels whereas SjGLO (Styrax) is also expressed in sepalsNo clear divergence in expression of PI from the two dif-ferent clades (fig 1 BPP68 and BPP89) is detected in thesecond and third whorl organs The expression patterns ofAP3 and TM6 lineage genes are different between these fourspecies Although in S drummondii both AP3 and TM6 areexpressed in all four floral whorls the expression in Styraxis very similar to that of TM6 and AP3 in core asterid spe-cies with in addition to expression in petals and stamensexpression of TM6 in carpels and AP3 in sepals In Cycla-men TM6 expression is absent in the upper part of the sta-mens and strong expression of AP3 is detected in thecarpels In Phlox however TM6 expression is absent fromthe carpels
The multiple PI copies in G brasiliensis (Lecythida-ceae) and P acaulis (Primulaceae) are the result of a recentduplication probably at the base of these two families(fig 1) We looked at the expression pattern of each of theseduplicated genes and of the AP3-like genes in these speciesIn G brasiliensis GbGLO2 is expressed in petals and sta-mens whereas GbGLO1 is mainly expressed in the stami-nal parts and less in the petals (fig 3H) Expression of AP3and TM6 seems to be lsquolsquoinversersquorsquo compared with the ex-pected asterid expression with AP3 expression mainly re-stricted to the stamens and strong TM6 expression in thepetals No AP3-like expression is detected in the carpelsIn P acaulis there are differences between the expressionpatterns of both PI copies (fig 3I) PaGLO2 is expressed inall organs tested whereas the expression of PaGLO1 ismainly restricted to the sepals and petals Both AP3 andTM6 are strongly expressed in sepals and as inC persicumTM6-expression is weak in the staminal parts Even afterthese recent duplication events divergence in the expres-sion domain of the two PI copies is detected
In contrast to the duplicated PI genes of Petunia(Vandenbussche et al 2004) the expression domains ofPI genes have changed considerably after duplication inEricales For both ancient and several recent duplicationevents divergence in expression was detected indicatingthat the regulation of the duplicate copies has divergedIn the studied species where only one PI copy was isolatedit appears that an lsquolsquoancestralrsquorsquo expression pattern of PI isreestablished However a second copy could have beenmissed during amplification Also within the AP3-like lin-eage some derived expression patterns are detected
In Situ Hybridization Demonstrates ExpressionDivergence in Early Flower Development
To analyze the expression patterns of the PI genes afterduplication in early developmental stages we examined thespatio-temporal expression patterns of PI in selected Eri-cales species by in situ hybridization (fig 4) We selectedexamples of each type of fate after duplication retention
2636 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
of both ancient duplicates (Stewartia) retention of only oneof each PI paralog (C persicum and S rubra) and retentionof recent duplicates (P acaulis)
In S rostrata (fig 4AndashF) we could detect expressiondivergence between each of the PI copies at early stages offloral development At the stage when petal primordia areevident but stamen primordia are apparent only as smallmeristematic outgrowths SpGLO1 expression is detectedin petal and stamen primordia (fig 4A) whereas SpGLO2transcripts are also detected in the tip of the sepals (fig 4D)As the flower develops SpGLO2 expression is low butexpands to include developing carpels (fig 4E) and expres-sion is maintained in the carpels in later stages (fig 4F) Incontrast SpGLO1 expression is maintained at high levelsonly in petals and stamens through later stages (fig 4Band C) and does not expand into sepals and carpels Theseearly patterns of S rostrata PI gene expression are consis-tent with the later patterns of expression as detected byRT-PCR (fig 3I)
Expression patterns of PI homologues were furtherinvestigated in C persicum (fig 4GndashI) and S rubra(fig 4JndashK) CpGLO expression first appears in the youngpetal primordia (fig 4G) followed by expression in the sta-men primordia as they emerge (fig 4H) Expression inthese organs is maintained in later stages (fig 4I) In con-trast to the expression patterns in mature organs as docu-mented by RT-PCR weak expression in the developingsepals (fig 4H) and developing carpels (fig 4I) was de-tected For S rubra we were not able to obtain the earliestdevelopmental stages For later stages of floral develop-ment we observed strong expression of PI in petals andstamens (fig 4J) Weak expression of SdGLO was also de-tected in the developing carpels (fig 4K) which is consis-tent with the RT-PCR data (fig 3) These observationstogether with the RT-PCR data (fig 3) support the ideathat species with presumably only one PI copy in their ge-nome show expression that is more confined to petals andstamens
Finally we investigated the expression of recent PIduplicates in P acaulis (fig 4LndashQ) At early stages bothPaGLO1 (fig 4L) and PaGLO2 (fig 4O) are expressedacross the meristematic dome in sepal primordia commonpetalndashstamen primordia and carpel primordia Expressionof PaGLO2 is maintained in all organs at older stages(fig 4R and Q) whereas PaGLO1 expression is restrictedto the stamens (fig 4M and N)
The in situ expression data (fig 4) in comparison tothe RT-PCR data (fig 3) indicate that early patterns of ex-pression of PI genes are often developmentally regulatedwith different patterns of expression being observed early incomparison to later in flower development Nonethelessboth sets of data indicate that in species possessing dupli-cate PI genes divergence in expression patterns betweenthe two copies is a common theme
Positive Selection and Relaxed Purifying Selection ActsAfter the Duplication Event in Ericales in the K- and C-Cerminal Domain
The mRNA expression data indicate that duplicate PIparalogs have acquired different expression patterns sug-gesting independent evolution of their transcriptional reg-ulatory control It was also demonstrated that adaptiveevolution in the coding regions of MADS-box transcriptionfactors has played a role in the diversification of thisgene family (Martinez-Castilla and Alvarez-Buylla2003) Within class B genes Hernandez-Hernandez et al(2007) demonstrated the role of positive selection after crit-ical duplication events like the AP3-PI duplication and theeuAP3ndashTM6 duplication To investigate whether parallelchanges occurred in the coding regions of the genes exam-ined in this study we investigated the nonsynonymous ver-sus synonymous amino acid replacement rates forsignificant increases before and after duplication in thespecific paralogous clades and along the coding sequenceThe results of likelihood ratio tests are shown in table 2 and
FIG 4mdashIn situ hybridization data for PI genes in various developmental stages of Stewartia rostrata (AndashC SpGLO1 DndashF SpGLO2) Cyclamenpersicum (GndashI CpGLO) Sarracenia rubra (JndashK SdGLO) Primula acaulis (LndashN PaGLO1 OndashQ PaGLO2) lsquolsquoprsquorsquo indicates petalprimordia lsquolsquosrsquorsquoindicates stamen primordia
PISTILLATAmdashDuplications in Asterids 2637
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
specific sites are marked in the alignment presented infigure 5 Parameter estimates log-likelihood scores andBayes Empirical Bayes identified sites are listed in supple-mentary table S2 Supplementary Material online
Comparison of the likelihood scores calculated usingbranch models R1 and R2 for matrix A indicates a signifi-cantly increased evolutionary rate following the ancient du-plication event in Ericales The dNdS estimate from thetwo-ratio branch model after duplication (x 5 0285) istwice compared with the value before duplication (x 50113) illustrating the relaxed selective regime resultingfrom the duplication event However the same test appliedon matrix B did not reveal this change in selective pressureOne possible explanation is that by adding the single-copylineages that evolved after duplication the observed patternin matrix A for the duplicates is obscured In addition wefound no evidence that the two paralogous gene cladescould be evolving at different rates because the three-and four-ratio models did not result in a significantly in-creased log-likelihood as calculated for either of the datamatrices
To formally test for the presence of positive selectionon sequence sites we used the site models proposed byNielsen and Yang (1998) and Yang et al (2000) Analysisof matrix A shows that both M2a and M8 two models thatallow for positive selection fit the data significantly betterthan the corresponding neutral models (M1a and M7) Sub-stitution patterns in about 5 of the sites for the M2amodel and about 9 of the sites for the M8 model appearto result from adaptive evolution Here also analysis of ma-trix B suggests positively selected sites M8 fits the databetter than the neutral model M7 with an estimation of
10 sites under positive selection Bayes empirical Bayes(Yang et al 2005) identification of the sites under positiveselection in matrix A finds seven sites to be positively se-lected under model M8 two of these sites have a posteriorprobability higher than 99 Within matrix B only twopositively selected sites were detected which were alsoidentified in matrix A both with posterior probability high-er than 91 Interestingly all the identified sites are locatedwithin the K2-subdomain the interhelical region betweenK2 and K3 the K3-subdomain and the C-terminal domain(fig 5) These functional domains have been shown to beimportant for proteinndashprotein interactions among Arabi-dopsis MADS-domain proteins (Ma et al 1991 Pnueliet al 1991 Shore and Sharrocks 1995 Riechmann et al1996 Yang et al 2003) The strength of the AP3ndashPI inter-action is mediated primarily by K1 and K2 whereas thePIndashSEP interaction is mediated by K2 the amino-terminalportion of K3 and possibly the C-domain (Yang et al 2003Yang and Jack 2004) The C-terminal region normallyshows much lower sequence conservation but containsshort highly conserved motifs (Lamb and Irish 2003) Sev-eral studies have suggested that this domain is involvedin the formation of higher order protein complexes(Egea-Cortines et al 1999 Honma and Goto 2001) yetthe specific function of the class B C-terminal domainsis unclear (Lamb and Irish 2003 Piwarzyk et al 2007)None of the sites identified by Yang et al (2003) to be re-sponsible for the direct interaction between APETALA3and PISTILLATA are positively selected for in our dataset However site 49 contains a hydrophobic amino acidthat when mutated disrupts the alpha-helical secondarystructure (Yang et al 2003) In addition sites 49 and site60 in figure 5 were identified by Yang and Jack (2004) to becritical for PIndashSEP interaction Site 49 is situated withinthe K2 subdomain which is required for both AP3ndashPIand PIndashSEP interaction whereas site 60 which is locatedat the amino-terminal end of K3 affects when mutated thestrength of the PIndashSEP interaction
To identify evolutionary pressures acting on specificsites along certain branches and in clades we calculatedlikelihoods using the branch-site models First we specifiedthe two branches immediately following the duplication asforeground branches in matrix A Branch-site models thatallow for positive selection in these foreground brancheswere not better at explaining the data than the nested sitemodels M1a and the null model A and indicate no evidencefor a burst of positive selection in the branches immediatelyfollowing the duplication event (Bielawski and Yang 2003)(data not shown) Second we treated all branches in eitherclade of paralogs as foreground branches When these twoclades are treated separately the model allowing for
Table 2Likelihood Ratio Tests Comparing Branch-Models (R1ndashR4)Site Models (M1ndashM8) and Branch-Site Models (Model AndashModel A Null)
Matrix Models Compared2(ln L1 ln L0)
Matrix A One ratio (R1) versus two ratio (R2) 18174Two ratio (R2) versus three ratio (R3) 0435Three ratio (R3) versus four ratio (R4) 1298Neutral (M1a) versus selection (M2a) 20566Beta (M7) versus beta amp w 1 (M8) 2683Model A versus M1a (Test 1) 874Model A versus model anull (Test 2) 252
Matrix B One ratio (R1) versus two ratio (R2) 2348Two ratio (R2) versus three ratio (R3) 001Three ratio (R3) versus four ratio (R4) 01Neutral (M1a) versus selection (M2a) 367Beta (M7) versus beta amp w 1 (M8) 7054
Significant at the 95 level significant at the 99 level
Table 3B Class MADS-Box Protein Interactions as Determined by Yeast Two-Hybrid Assays in Camellia japonica
CjDEF-BD CjTM6-BD CjGLO1-BD CjGLO2-BD BD
CjDEF-AD thorn mdash mdash mdash mdashCjTM6-AD mdash mdash thorn mdash mdashCjGLO1-AD thorn thorn mdash mdash mdashCjGLO2-AD thorn mdash mdash mdash mdashAD mdash mdash mdash mdash mdash
2638 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
positive selection is not significantly better in explaining thedata than the model M1a and null model A (data notshown) When the two clades are treated together as fore-ground branches model A is significantly better than thenested site model M1a (test 1) suggesting that positive se-lection is acting in both clades that arose after duplicationBayes empirical Bayes identification resulted in seven po-sitions though all with posterior probability below 90The improved test 2 from Zhang et al (2005) resulted ina value 252 just beneath the critical value 271 at the5 level but far above the critical value at the 10 level164 These two tests support the hypothesis that positiveselection acts after the duplication event in both paralogousgene clades
Together the identification of changes in selectivepressure in time and along the coding sequence indicatethat upon duplication PI paralogs underwent an increasednonsynonymous substitution rate and that this occurred
specifically in the domains shown to be necessary forproteinndashprotein interactions
Diversification in the Interaction Behavior of B-ClassGenes After the Ancient Duplication Event in BasalAsterids
As sites in the interaction domains of the PI genesfrom basal asterids were detected to be positively selectedafter duplication we further investigated the possible func-tional divergence of the duplicated PI proteins by yeast-2-hybrid analyses We tested the interactions between thefour class B genes in C japonica Camellia japonica con-tains two AP3-like genes one belonging to the euAP3-typeand one from the paleoAP3-type and two PI-like genesfrom an ancient duplication event at the base of the EricalesThis situation is similar to the B-class configuration in thecore asterid species Petunia hybrida (Vandenbussche
FIG 5mdashAlignment of the K-domain and C-terminal domain of the PI duplicates from matrix A used for the analysis in PAML Differentsubdomains and the characteristic PI motif are indicated (Kramer et al 1998) Codon positions identified to be under positive selection are marked withgray boxes
PISTILLATAmdashDuplications in Asterids 2639
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

Both copies in each of these species appear to have divergedin their expression patterns after duplication (fig 3AndashC)Although HdGLO2 is expressed in the petals and stamensof H diptera the other copy (HdGLO1) is mainly ex-pressed in stamens and the gynoecium and absent in petalsIn comparison the TM6 and euAP3 lineage genes show un-expected expression patterns with strong expression ofHdTM6 in all disected organs but HdDEF expression beingmainly restricted to petals and carpels InC japonica one PIparalog (CjGLO2) is expressed throughout sepals petalsand stamens whereas expression of the second paralog(CjGLO1) appeared to be restricted to the nonfused partsof petals stamens and carpels Although both CjDEF andCjTM6 expression are detected in all organs strong expres-sion ofCjDEF is seenmainly in the sepals and petalsCjTM6expression is strongest in all stamen parts and in carpelsS pseudocamellia also displays differences in the expressionof its two PI copies One paralog (SpGLO2) is expressed inpetals stamens carpels and expression of the second paral-og (SpGLO1) is more restricted to the fused part of petals andstamens and upper part of petals The AP3 homologueSpDEF is strongly expressed in all stamen parts and carpelswhereas strong expression of SpTM6 is detected in sepals allstamen parts and carpels
In summary in these three cases there is one PI copywith a broad expression pattern in the flower (even in sepalsand carpels) whereas expression of the other PI copy ismore restricted to specific parts of petals and stamensThese expression patterns differ from those of PI lineagegenes in core asterid species (Vandenbussche et al2004 de Martino et al 2006) where PI expression (alsoin a duplicated configuration) is mainly restricted to the sec-ond and third whorl organs petals and stamens and nostrong expression in the first and fourth whorl organs is de-tected Interestingly the divergence in expression patternsin these species does not follow the gene phylogeny Theparalogous copies with a general expression pattern are notgrouped in one clade of PI genes in Ericales but rather arefound in two clades This suggests that the PI expressiondomains were not fixed immediately after duplication in-dicating that subfunctionalization in spatial expression doesnot explain retention of the duplicates Alternatively ifregulatory subfunctionalization was the main reason forretention evidence for this has been obscured throughtime Differences in expression between AP3 and TM6in C japonica seem to be very similar to members ofthe same lineages in the core asterid species Expressionof AP3-like genes in H diptera and S pseudocamelliahowever differs from those of Camellia with strongAP3 expression in the carpels of Stewartia and Halesiaand weak expression of AP3 in stamens of Halesia In eachof these species expression of one or both AP3-like genes isdetected in the sepals
Sarracenia drummondii (fig 3D) Styrax japonicus(fig 3E) C persicum (fig 3F) and Phlox paniculata(fig 3G) maintained probably only one of the paralogousPI copies in their genome after the ancient Ericales dupli-cation Moreover Sarracenia and Styrax maintained a dif-ferent copy (fig 1 BPP 68) than did Cyclamen and Phlox(fig 1 BPP89) We investigated the expression patterns ofPI genes in these species to monitor the possible divergence
between the two paralogous PI clades after duplication Ineach of these four species PI is expressed in petals and sta-mens similar to the expression patterns of PI in core aster-ids in previous studies (Vandenbussche et al 2004 deMartino et al 2006) In both Sarracenia (SdGLO) andPhlox (PpGLO) there is some expression detected in thecarpels whereas SjGLO (Styrax) is also expressed in sepalsNo clear divergence in expression of PI from the two dif-ferent clades (fig 1 BPP68 and BPP89) is detected in thesecond and third whorl organs The expression patterns ofAP3 and TM6 lineage genes are different between these fourspecies Although in S drummondii both AP3 and TM6 areexpressed in all four floral whorls the expression in Styraxis very similar to that of TM6 and AP3 in core asterid spe-cies with in addition to expression in petals and stamensexpression of TM6 in carpels and AP3 in sepals In Cycla-men TM6 expression is absent in the upper part of the sta-mens and strong expression of AP3 is detected in thecarpels In Phlox however TM6 expression is absent fromthe carpels
The multiple PI copies in G brasiliensis (Lecythida-ceae) and P acaulis (Primulaceae) are the result of a recentduplication probably at the base of these two families(fig 1) We looked at the expression pattern of each of theseduplicated genes and of the AP3-like genes in these speciesIn G brasiliensis GbGLO2 is expressed in petals and sta-mens whereas GbGLO1 is mainly expressed in the stami-nal parts and less in the petals (fig 3H) Expression of AP3and TM6 seems to be lsquolsquoinversersquorsquo compared with the ex-pected asterid expression with AP3 expression mainly re-stricted to the stamens and strong TM6 expression in thepetals No AP3-like expression is detected in the carpelsIn P acaulis there are differences between the expressionpatterns of both PI copies (fig 3I) PaGLO2 is expressed inall organs tested whereas the expression of PaGLO1 ismainly restricted to the sepals and petals Both AP3 andTM6 are strongly expressed in sepals and as inC persicumTM6-expression is weak in the staminal parts Even afterthese recent duplication events divergence in the expres-sion domain of the two PI copies is detected
In contrast to the duplicated PI genes of Petunia(Vandenbussche et al 2004) the expression domains ofPI genes have changed considerably after duplication inEricales For both ancient and several recent duplicationevents divergence in expression was detected indicatingthat the regulation of the duplicate copies has divergedIn the studied species where only one PI copy was isolatedit appears that an lsquolsquoancestralrsquorsquo expression pattern of PI isreestablished However a second copy could have beenmissed during amplification Also within the AP3-like lin-eage some derived expression patterns are detected
In Situ Hybridization Demonstrates ExpressionDivergence in Early Flower Development
To analyze the expression patterns of the PI genes afterduplication in early developmental stages we examined thespatio-temporal expression patterns of PI in selected Eri-cales species by in situ hybridization (fig 4) We selectedexamples of each type of fate after duplication retention
2636 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
of both ancient duplicates (Stewartia) retention of only oneof each PI paralog (C persicum and S rubra) and retentionof recent duplicates (P acaulis)
In S rostrata (fig 4AndashF) we could detect expressiondivergence between each of the PI copies at early stages offloral development At the stage when petal primordia areevident but stamen primordia are apparent only as smallmeristematic outgrowths SpGLO1 expression is detectedin petal and stamen primordia (fig 4A) whereas SpGLO2transcripts are also detected in the tip of the sepals (fig 4D)As the flower develops SpGLO2 expression is low butexpands to include developing carpels (fig 4E) and expres-sion is maintained in the carpels in later stages (fig 4F) Incontrast SpGLO1 expression is maintained at high levelsonly in petals and stamens through later stages (fig 4Band C) and does not expand into sepals and carpels Theseearly patterns of S rostrata PI gene expression are consis-tent with the later patterns of expression as detected byRT-PCR (fig 3I)
Expression patterns of PI homologues were furtherinvestigated in C persicum (fig 4GndashI) and S rubra(fig 4JndashK) CpGLO expression first appears in the youngpetal primordia (fig 4G) followed by expression in the sta-men primordia as they emerge (fig 4H) Expression inthese organs is maintained in later stages (fig 4I) In con-trast to the expression patterns in mature organs as docu-mented by RT-PCR weak expression in the developingsepals (fig 4H) and developing carpels (fig 4I) was de-tected For S rubra we were not able to obtain the earliestdevelopmental stages For later stages of floral develop-ment we observed strong expression of PI in petals andstamens (fig 4J) Weak expression of SdGLO was also de-tected in the developing carpels (fig 4K) which is consis-tent with the RT-PCR data (fig 3) These observationstogether with the RT-PCR data (fig 3) support the ideathat species with presumably only one PI copy in their ge-nome show expression that is more confined to petals andstamens
Finally we investigated the expression of recent PIduplicates in P acaulis (fig 4LndashQ) At early stages bothPaGLO1 (fig 4L) and PaGLO2 (fig 4O) are expressedacross the meristematic dome in sepal primordia commonpetalndashstamen primordia and carpel primordia Expressionof PaGLO2 is maintained in all organs at older stages(fig 4R and Q) whereas PaGLO1 expression is restrictedto the stamens (fig 4M and N)
The in situ expression data (fig 4) in comparison tothe RT-PCR data (fig 3) indicate that early patterns of ex-pression of PI genes are often developmentally regulatedwith different patterns of expression being observed early incomparison to later in flower development Nonethelessboth sets of data indicate that in species possessing dupli-cate PI genes divergence in expression patterns betweenthe two copies is a common theme
Positive Selection and Relaxed Purifying Selection ActsAfter the Duplication Event in Ericales in the K- and C-Cerminal Domain
The mRNA expression data indicate that duplicate PIparalogs have acquired different expression patterns sug-gesting independent evolution of their transcriptional reg-ulatory control It was also demonstrated that adaptiveevolution in the coding regions of MADS-box transcriptionfactors has played a role in the diversification of thisgene family (Martinez-Castilla and Alvarez-Buylla2003) Within class B genes Hernandez-Hernandez et al(2007) demonstrated the role of positive selection after crit-ical duplication events like the AP3-PI duplication and theeuAP3ndashTM6 duplication To investigate whether parallelchanges occurred in the coding regions of the genes exam-ined in this study we investigated the nonsynonymous ver-sus synonymous amino acid replacement rates forsignificant increases before and after duplication in thespecific paralogous clades and along the coding sequenceThe results of likelihood ratio tests are shown in table 2 and
FIG 4mdashIn situ hybridization data for PI genes in various developmental stages of Stewartia rostrata (AndashC SpGLO1 DndashF SpGLO2) Cyclamenpersicum (GndashI CpGLO) Sarracenia rubra (JndashK SdGLO) Primula acaulis (LndashN PaGLO1 OndashQ PaGLO2) lsquolsquoprsquorsquo indicates petalprimordia lsquolsquosrsquorsquoindicates stamen primordia
PISTILLATAmdashDuplications in Asterids 2637
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
specific sites are marked in the alignment presented infigure 5 Parameter estimates log-likelihood scores andBayes Empirical Bayes identified sites are listed in supple-mentary table S2 Supplementary Material online
Comparison of the likelihood scores calculated usingbranch models R1 and R2 for matrix A indicates a signifi-cantly increased evolutionary rate following the ancient du-plication event in Ericales The dNdS estimate from thetwo-ratio branch model after duplication (x 5 0285) istwice compared with the value before duplication (x 50113) illustrating the relaxed selective regime resultingfrom the duplication event However the same test appliedon matrix B did not reveal this change in selective pressureOne possible explanation is that by adding the single-copylineages that evolved after duplication the observed patternin matrix A for the duplicates is obscured In addition wefound no evidence that the two paralogous gene cladescould be evolving at different rates because the three-and four-ratio models did not result in a significantly in-creased log-likelihood as calculated for either of the datamatrices
To formally test for the presence of positive selectionon sequence sites we used the site models proposed byNielsen and Yang (1998) and Yang et al (2000) Analysisof matrix A shows that both M2a and M8 two models thatallow for positive selection fit the data significantly betterthan the corresponding neutral models (M1a and M7) Sub-stitution patterns in about 5 of the sites for the M2amodel and about 9 of the sites for the M8 model appearto result from adaptive evolution Here also analysis of ma-trix B suggests positively selected sites M8 fits the databetter than the neutral model M7 with an estimation of
10 sites under positive selection Bayes empirical Bayes(Yang et al 2005) identification of the sites under positiveselection in matrix A finds seven sites to be positively se-lected under model M8 two of these sites have a posteriorprobability higher than 99 Within matrix B only twopositively selected sites were detected which were alsoidentified in matrix A both with posterior probability high-er than 91 Interestingly all the identified sites are locatedwithin the K2-subdomain the interhelical region betweenK2 and K3 the K3-subdomain and the C-terminal domain(fig 5) These functional domains have been shown to beimportant for proteinndashprotein interactions among Arabi-dopsis MADS-domain proteins (Ma et al 1991 Pnueliet al 1991 Shore and Sharrocks 1995 Riechmann et al1996 Yang et al 2003) The strength of the AP3ndashPI inter-action is mediated primarily by K1 and K2 whereas thePIndashSEP interaction is mediated by K2 the amino-terminalportion of K3 and possibly the C-domain (Yang et al 2003Yang and Jack 2004) The C-terminal region normallyshows much lower sequence conservation but containsshort highly conserved motifs (Lamb and Irish 2003) Sev-eral studies have suggested that this domain is involvedin the formation of higher order protein complexes(Egea-Cortines et al 1999 Honma and Goto 2001) yetthe specific function of the class B C-terminal domainsis unclear (Lamb and Irish 2003 Piwarzyk et al 2007)None of the sites identified by Yang et al (2003) to be re-sponsible for the direct interaction between APETALA3and PISTILLATA are positively selected for in our dataset However site 49 contains a hydrophobic amino acidthat when mutated disrupts the alpha-helical secondarystructure (Yang et al 2003) In addition sites 49 and site60 in figure 5 were identified by Yang and Jack (2004) to becritical for PIndashSEP interaction Site 49 is situated withinthe K2 subdomain which is required for both AP3ndashPIand PIndashSEP interaction whereas site 60 which is locatedat the amino-terminal end of K3 affects when mutated thestrength of the PIndashSEP interaction
To identify evolutionary pressures acting on specificsites along certain branches and in clades we calculatedlikelihoods using the branch-site models First we specifiedthe two branches immediately following the duplication asforeground branches in matrix A Branch-site models thatallow for positive selection in these foreground brancheswere not better at explaining the data than the nested sitemodels M1a and the null model A and indicate no evidencefor a burst of positive selection in the branches immediatelyfollowing the duplication event (Bielawski and Yang 2003)(data not shown) Second we treated all branches in eitherclade of paralogs as foreground branches When these twoclades are treated separately the model allowing for
Table 2Likelihood Ratio Tests Comparing Branch-Models (R1ndashR4)Site Models (M1ndashM8) and Branch-Site Models (Model AndashModel A Null)
Matrix Models Compared2(ln L1 ln L0)
Matrix A One ratio (R1) versus two ratio (R2) 18174Two ratio (R2) versus three ratio (R3) 0435Three ratio (R3) versus four ratio (R4) 1298Neutral (M1a) versus selection (M2a) 20566Beta (M7) versus beta amp w 1 (M8) 2683Model A versus M1a (Test 1) 874Model A versus model anull (Test 2) 252
Matrix B One ratio (R1) versus two ratio (R2) 2348Two ratio (R2) versus three ratio (R3) 001Three ratio (R3) versus four ratio (R4) 01Neutral (M1a) versus selection (M2a) 367Beta (M7) versus beta amp w 1 (M8) 7054
Significant at the 95 level significant at the 99 level
Table 3B Class MADS-Box Protein Interactions as Determined by Yeast Two-Hybrid Assays in Camellia japonica
CjDEF-BD CjTM6-BD CjGLO1-BD CjGLO2-BD BD
CjDEF-AD thorn mdash mdash mdash mdashCjTM6-AD mdash mdash thorn mdash mdashCjGLO1-AD thorn thorn mdash mdash mdashCjGLO2-AD thorn mdash mdash mdash mdashAD mdash mdash mdash mdash mdash
2638 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
positive selection is not significantly better in explaining thedata than the model M1a and null model A (data notshown) When the two clades are treated together as fore-ground branches model A is significantly better than thenested site model M1a (test 1) suggesting that positive se-lection is acting in both clades that arose after duplicationBayes empirical Bayes identification resulted in seven po-sitions though all with posterior probability below 90The improved test 2 from Zhang et al (2005) resulted ina value 252 just beneath the critical value 271 at the5 level but far above the critical value at the 10 level164 These two tests support the hypothesis that positiveselection acts after the duplication event in both paralogousgene clades
Together the identification of changes in selectivepressure in time and along the coding sequence indicatethat upon duplication PI paralogs underwent an increasednonsynonymous substitution rate and that this occurred
specifically in the domains shown to be necessary forproteinndashprotein interactions
Diversification in the Interaction Behavior of B-ClassGenes After the Ancient Duplication Event in BasalAsterids
As sites in the interaction domains of the PI genesfrom basal asterids were detected to be positively selectedafter duplication we further investigated the possible func-tional divergence of the duplicated PI proteins by yeast-2-hybrid analyses We tested the interactions between thefour class B genes in C japonica Camellia japonica con-tains two AP3-like genes one belonging to the euAP3-typeand one from the paleoAP3-type and two PI-like genesfrom an ancient duplication event at the base of the EricalesThis situation is similar to the B-class configuration in thecore asterid species Petunia hybrida (Vandenbussche
FIG 5mdashAlignment of the K-domain and C-terminal domain of the PI duplicates from matrix A used for the analysis in PAML Differentsubdomains and the characteristic PI motif are indicated (Kramer et al 1998) Codon positions identified to be under positive selection are marked withgray boxes
PISTILLATAmdashDuplications in Asterids 2639
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

of both ancient duplicates (Stewartia) retention of only oneof each PI paralog (C persicum and S rubra) and retentionof recent duplicates (P acaulis)
In S rostrata (fig 4AndashF) we could detect expressiondivergence between each of the PI copies at early stages offloral development At the stage when petal primordia areevident but stamen primordia are apparent only as smallmeristematic outgrowths SpGLO1 expression is detectedin petal and stamen primordia (fig 4A) whereas SpGLO2transcripts are also detected in the tip of the sepals (fig 4D)As the flower develops SpGLO2 expression is low butexpands to include developing carpels (fig 4E) and expres-sion is maintained in the carpels in later stages (fig 4F) Incontrast SpGLO1 expression is maintained at high levelsonly in petals and stamens through later stages (fig 4Band C) and does not expand into sepals and carpels Theseearly patterns of S rostrata PI gene expression are consis-tent with the later patterns of expression as detected byRT-PCR (fig 3I)
Expression patterns of PI homologues were furtherinvestigated in C persicum (fig 4GndashI) and S rubra(fig 4JndashK) CpGLO expression first appears in the youngpetal primordia (fig 4G) followed by expression in the sta-men primordia as they emerge (fig 4H) Expression inthese organs is maintained in later stages (fig 4I) In con-trast to the expression patterns in mature organs as docu-mented by RT-PCR weak expression in the developingsepals (fig 4H) and developing carpels (fig 4I) was de-tected For S rubra we were not able to obtain the earliestdevelopmental stages For later stages of floral develop-ment we observed strong expression of PI in petals andstamens (fig 4J) Weak expression of SdGLO was also de-tected in the developing carpels (fig 4K) which is consis-tent with the RT-PCR data (fig 3) These observationstogether with the RT-PCR data (fig 3) support the ideathat species with presumably only one PI copy in their ge-nome show expression that is more confined to petals andstamens
Finally we investigated the expression of recent PIduplicates in P acaulis (fig 4LndashQ) At early stages bothPaGLO1 (fig 4L) and PaGLO2 (fig 4O) are expressedacross the meristematic dome in sepal primordia commonpetalndashstamen primordia and carpel primordia Expressionof PaGLO2 is maintained in all organs at older stages(fig 4R and Q) whereas PaGLO1 expression is restrictedto the stamens (fig 4M and N)
The in situ expression data (fig 4) in comparison tothe RT-PCR data (fig 3) indicate that early patterns of ex-pression of PI genes are often developmentally regulatedwith different patterns of expression being observed early incomparison to later in flower development Nonethelessboth sets of data indicate that in species possessing dupli-cate PI genes divergence in expression patterns betweenthe two copies is a common theme
Positive Selection and Relaxed Purifying Selection ActsAfter the Duplication Event in Ericales in the K- and C-Cerminal Domain
The mRNA expression data indicate that duplicate PIparalogs have acquired different expression patterns sug-gesting independent evolution of their transcriptional reg-ulatory control It was also demonstrated that adaptiveevolution in the coding regions of MADS-box transcriptionfactors has played a role in the diversification of thisgene family (Martinez-Castilla and Alvarez-Buylla2003) Within class B genes Hernandez-Hernandez et al(2007) demonstrated the role of positive selection after crit-ical duplication events like the AP3-PI duplication and theeuAP3ndashTM6 duplication To investigate whether parallelchanges occurred in the coding regions of the genes exam-ined in this study we investigated the nonsynonymous ver-sus synonymous amino acid replacement rates forsignificant increases before and after duplication in thespecific paralogous clades and along the coding sequenceThe results of likelihood ratio tests are shown in table 2 and
FIG 4mdashIn situ hybridization data for PI genes in various developmental stages of Stewartia rostrata (AndashC SpGLO1 DndashF SpGLO2) Cyclamenpersicum (GndashI CpGLO) Sarracenia rubra (JndashK SdGLO) Primula acaulis (LndashN PaGLO1 OndashQ PaGLO2) lsquolsquoprsquorsquo indicates petalprimordia lsquolsquosrsquorsquoindicates stamen primordia
PISTILLATAmdashDuplications in Asterids 2637
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
specific sites are marked in the alignment presented infigure 5 Parameter estimates log-likelihood scores andBayes Empirical Bayes identified sites are listed in supple-mentary table S2 Supplementary Material online
Comparison of the likelihood scores calculated usingbranch models R1 and R2 for matrix A indicates a signifi-cantly increased evolutionary rate following the ancient du-plication event in Ericales The dNdS estimate from thetwo-ratio branch model after duplication (x 5 0285) istwice compared with the value before duplication (x 50113) illustrating the relaxed selective regime resultingfrom the duplication event However the same test appliedon matrix B did not reveal this change in selective pressureOne possible explanation is that by adding the single-copylineages that evolved after duplication the observed patternin matrix A for the duplicates is obscured In addition wefound no evidence that the two paralogous gene cladescould be evolving at different rates because the three-and four-ratio models did not result in a significantly in-creased log-likelihood as calculated for either of the datamatrices
To formally test for the presence of positive selectionon sequence sites we used the site models proposed byNielsen and Yang (1998) and Yang et al (2000) Analysisof matrix A shows that both M2a and M8 two models thatallow for positive selection fit the data significantly betterthan the corresponding neutral models (M1a and M7) Sub-stitution patterns in about 5 of the sites for the M2amodel and about 9 of the sites for the M8 model appearto result from adaptive evolution Here also analysis of ma-trix B suggests positively selected sites M8 fits the databetter than the neutral model M7 with an estimation of
10 sites under positive selection Bayes empirical Bayes(Yang et al 2005) identification of the sites under positiveselection in matrix A finds seven sites to be positively se-lected under model M8 two of these sites have a posteriorprobability higher than 99 Within matrix B only twopositively selected sites were detected which were alsoidentified in matrix A both with posterior probability high-er than 91 Interestingly all the identified sites are locatedwithin the K2-subdomain the interhelical region betweenK2 and K3 the K3-subdomain and the C-terminal domain(fig 5) These functional domains have been shown to beimportant for proteinndashprotein interactions among Arabi-dopsis MADS-domain proteins (Ma et al 1991 Pnueliet al 1991 Shore and Sharrocks 1995 Riechmann et al1996 Yang et al 2003) The strength of the AP3ndashPI inter-action is mediated primarily by K1 and K2 whereas thePIndashSEP interaction is mediated by K2 the amino-terminalportion of K3 and possibly the C-domain (Yang et al 2003Yang and Jack 2004) The C-terminal region normallyshows much lower sequence conservation but containsshort highly conserved motifs (Lamb and Irish 2003) Sev-eral studies have suggested that this domain is involvedin the formation of higher order protein complexes(Egea-Cortines et al 1999 Honma and Goto 2001) yetthe specific function of the class B C-terminal domainsis unclear (Lamb and Irish 2003 Piwarzyk et al 2007)None of the sites identified by Yang et al (2003) to be re-sponsible for the direct interaction between APETALA3and PISTILLATA are positively selected for in our dataset However site 49 contains a hydrophobic amino acidthat when mutated disrupts the alpha-helical secondarystructure (Yang et al 2003) In addition sites 49 and site60 in figure 5 were identified by Yang and Jack (2004) to becritical for PIndashSEP interaction Site 49 is situated withinthe K2 subdomain which is required for both AP3ndashPIand PIndashSEP interaction whereas site 60 which is locatedat the amino-terminal end of K3 affects when mutated thestrength of the PIndashSEP interaction
To identify evolutionary pressures acting on specificsites along certain branches and in clades we calculatedlikelihoods using the branch-site models First we specifiedthe two branches immediately following the duplication asforeground branches in matrix A Branch-site models thatallow for positive selection in these foreground brancheswere not better at explaining the data than the nested sitemodels M1a and the null model A and indicate no evidencefor a burst of positive selection in the branches immediatelyfollowing the duplication event (Bielawski and Yang 2003)(data not shown) Second we treated all branches in eitherclade of paralogs as foreground branches When these twoclades are treated separately the model allowing for
Table 2Likelihood Ratio Tests Comparing Branch-Models (R1ndashR4)Site Models (M1ndashM8) and Branch-Site Models (Model AndashModel A Null)
Matrix Models Compared2(ln L1 ln L0)
Matrix A One ratio (R1) versus two ratio (R2) 18174Two ratio (R2) versus three ratio (R3) 0435Three ratio (R3) versus four ratio (R4) 1298Neutral (M1a) versus selection (M2a) 20566Beta (M7) versus beta amp w 1 (M8) 2683Model A versus M1a (Test 1) 874Model A versus model anull (Test 2) 252
Matrix B One ratio (R1) versus two ratio (R2) 2348Two ratio (R2) versus three ratio (R3) 001Three ratio (R3) versus four ratio (R4) 01Neutral (M1a) versus selection (M2a) 367Beta (M7) versus beta amp w 1 (M8) 7054
Significant at the 95 level significant at the 99 level
Table 3B Class MADS-Box Protein Interactions as Determined by Yeast Two-Hybrid Assays in Camellia japonica
CjDEF-BD CjTM6-BD CjGLO1-BD CjGLO2-BD BD
CjDEF-AD thorn mdash mdash mdash mdashCjTM6-AD mdash mdash thorn mdash mdashCjGLO1-AD thorn thorn mdash mdash mdashCjGLO2-AD thorn mdash mdash mdash mdashAD mdash mdash mdash mdash mdash
2638 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
positive selection is not significantly better in explaining thedata than the model M1a and null model A (data notshown) When the two clades are treated together as fore-ground branches model A is significantly better than thenested site model M1a (test 1) suggesting that positive se-lection is acting in both clades that arose after duplicationBayes empirical Bayes identification resulted in seven po-sitions though all with posterior probability below 90The improved test 2 from Zhang et al (2005) resulted ina value 252 just beneath the critical value 271 at the5 level but far above the critical value at the 10 level164 These two tests support the hypothesis that positiveselection acts after the duplication event in both paralogousgene clades
Together the identification of changes in selectivepressure in time and along the coding sequence indicatethat upon duplication PI paralogs underwent an increasednonsynonymous substitution rate and that this occurred
specifically in the domains shown to be necessary forproteinndashprotein interactions
Diversification in the Interaction Behavior of B-ClassGenes After the Ancient Duplication Event in BasalAsterids
As sites in the interaction domains of the PI genesfrom basal asterids were detected to be positively selectedafter duplication we further investigated the possible func-tional divergence of the duplicated PI proteins by yeast-2-hybrid analyses We tested the interactions between thefour class B genes in C japonica Camellia japonica con-tains two AP3-like genes one belonging to the euAP3-typeand one from the paleoAP3-type and two PI-like genesfrom an ancient duplication event at the base of the EricalesThis situation is similar to the B-class configuration in thecore asterid species Petunia hybrida (Vandenbussche
FIG 5mdashAlignment of the K-domain and C-terminal domain of the PI duplicates from matrix A used for the analysis in PAML Differentsubdomains and the characteristic PI motif are indicated (Kramer et al 1998) Codon positions identified to be under positive selection are marked withgray boxes
PISTILLATAmdashDuplications in Asterids 2639
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

specific sites are marked in the alignment presented infigure 5 Parameter estimates log-likelihood scores andBayes Empirical Bayes identified sites are listed in supple-mentary table S2 Supplementary Material online
Comparison of the likelihood scores calculated usingbranch models R1 and R2 for matrix A indicates a signifi-cantly increased evolutionary rate following the ancient du-plication event in Ericales The dNdS estimate from thetwo-ratio branch model after duplication (x 5 0285) istwice compared with the value before duplication (x 50113) illustrating the relaxed selective regime resultingfrom the duplication event However the same test appliedon matrix B did not reveal this change in selective pressureOne possible explanation is that by adding the single-copylineages that evolved after duplication the observed patternin matrix A for the duplicates is obscured In addition wefound no evidence that the two paralogous gene cladescould be evolving at different rates because the three-and four-ratio models did not result in a significantly in-creased log-likelihood as calculated for either of the datamatrices
To formally test for the presence of positive selectionon sequence sites we used the site models proposed byNielsen and Yang (1998) and Yang et al (2000) Analysisof matrix A shows that both M2a and M8 two models thatallow for positive selection fit the data significantly betterthan the corresponding neutral models (M1a and M7) Sub-stitution patterns in about 5 of the sites for the M2amodel and about 9 of the sites for the M8 model appearto result from adaptive evolution Here also analysis of ma-trix B suggests positively selected sites M8 fits the databetter than the neutral model M7 with an estimation of
10 sites under positive selection Bayes empirical Bayes(Yang et al 2005) identification of the sites under positiveselection in matrix A finds seven sites to be positively se-lected under model M8 two of these sites have a posteriorprobability higher than 99 Within matrix B only twopositively selected sites were detected which were alsoidentified in matrix A both with posterior probability high-er than 91 Interestingly all the identified sites are locatedwithin the K2-subdomain the interhelical region betweenK2 and K3 the K3-subdomain and the C-terminal domain(fig 5) These functional domains have been shown to beimportant for proteinndashprotein interactions among Arabi-dopsis MADS-domain proteins (Ma et al 1991 Pnueliet al 1991 Shore and Sharrocks 1995 Riechmann et al1996 Yang et al 2003) The strength of the AP3ndashPI inter-action is mediated primarily by K1 and K2 whereas thePIndashSEP interaction is mediated by K2 the amino-terminalportion of K3 and possibly the C-domain (Yang et al 2003Yang and Jack 2004) The C-terminal region normallyshows much lower sequence conservation but containsshort highly conserved motifs (Lamb and Irish 2003) Sev-eral studies have suggested that this domain is involvedin the formation of higher order protein complexes(Egea-Cortines et al 1999 Honma and Goto 2001) yetthe specific function of the class B C-terminal domainsis unclear (Lamb and Irish 2003 Piwarzyk et al 2007)None of the sites identified by Yang et al (2003) to be re-sponsible for the direct interaction between APETALA3and PISTILLATA are positively selected for in our dataset However site 49 contains a hydrophobic amino acidthat when mutated disrupts the alpha-helical secondarystructure (Yang et al 2003) In addition sites 49 and site60 in figure 5 were identified by Yang and Jack (2004) to becritical for PIndashSEP interaction Site 49 is situated withinthe K2 subdomain which is required for both AP3ndashPIand PIndashSEP interaction whereas site 60 which is locatedat the amino-terminal end of K3 affects when mutated thestrength of the PIndashSEP interaction
To identify evolutionary pressures acting on specificsites along certain branches and in clades we calculatedlikelihoods using the branch-site models First we specifiedthe two branches immediately following the duplication asforeground branches in matrix A Branch-site models thatallow for positive selection in these foreground brancheswere not better at explaining the data than the nested sitemodels M1a and the null model A and indicate no evidencefor a burst of positive selection in the branches immediatelyfollowing the duplication event (Bielawski and Yang 2003)(data not shown) Second we treated all branches in eitherclade of paralogs as foreground branches When these twoclades are treated separately the model allowing for
Table 2Likelihood Ratio Tests Comparing Branch-Models (R1ndashR4)Site Models (M1ndashM8) and Branch-Site Models (Model AndashModel A Null)
Matrix Models Compared2(ln L1 ln L0)
Matrix A One ratio (R1) versus two ratio (R2) 18174Two ratio (R2) versus three ratio (R3) 0435Three ratio (R3) versus four ratio (R4) 1298Neutral (M1a) versus selection (M2a) 20566Beta (M7) versus beta amp w 1 (M8) 2683Model A versus M1a (Test 1) 874Model A versus model anull (Test 2) 252
Matrix B One ratio (R1) versus two ratio (R2) 2348Two ratio (R2) versus three ratio (R3) 001Three ratio (R3) versus four ratio (R4) 01Neutral (M1a) versus selection (M2a) 367Beta (M7) versus beta amp w 1 (M8) 7054
Significant at the 95 level significant at the 99 level
Table 3B Class MADS-Box Protein Interactions as Determined by Yeast Two-Hybrid Assays in Camellia japonica
CjDEF-BD CjTM6-BD CjGLO1-BD CjGLO2-BD BD
CjDEF-AD thorn mdash mdash mdash mdashCjTM6-AD mdash mdash thorn mdash mdashCjGLO1-AD thorn thorn mdash mdash mdashCjGLO2-AD thorn mdash mdash mdash mdashAD mdash mdash mdash mdash mdash
2638 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
positive selection is not significantly better in explaining thedata than the model M1a and null model A (data notshown) When the two clades are treated together as fore-ground branches model A is significantly better than thenested site model M1a (test 1) suggesting that positive se-lection is acting in both clades that arose after duplicationBayes empirical Bayes identification resulted in seven po-sitions though all with posterior probability below 90The improved test 2 from Zhang et al (2005) resulted ina value 252 just beneath the critical value 271 at the5 level but far above the critical value at the 10 level164 These two tests support the hypothesis that positiveselection acts after the duplication event in both paralogousgene clades
Together the identification of changes in selectivepressure in time and along the coding sequence indicatethat upon duplication PI paralogs underwent an increasednonsynonymous substitution rate and that this occurred
specifically in the domains shown to be necessary forproteinndashprotein interactions
Diversification in the Interaction Behavior of B-ClassGenes After the Ancient Duplication Event in BasalAsterids
As sites in the interaction domains of the PI genesfrom basal asterids were detected to be positively selectedafter duplication we further investigated the possible func-tional divergence of the duplicated PI proteins by yeast-2-hybrid analyses We tested the interactions between thefour class B genes in C japonica Camellia japonica con-tains two AP3-like genes one belonging to the euAP3-typeand one from the paleoAP3-type and two PI-like genesfrom an ancient duplication event at the base of the EricalesThis situation is similar to the B-class configuration in thecore asterid species Petunia hybrida (Vandenbussche
FIG 5mdashAlignment of the K-domain and C-terminal domain of the PI duplicates from matrix A used for the analysis in PAML Differentsubdomains and the characteristic PI motif are indicated (Kramer et al 1998) Codon positions identified to be under positive selection are marked withgray boxes
PISTILLATAmdashDuplications in Asterids 2639
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

positive selection is not significantly better in explaining thedata than the model M1a and null model A (data notshown) When the two clades are treated together as fore-ground branches model A is significantly better than thenested site model M1a (test 1) suggesting that positive se-lection is acting in both clades that arose after duplicationBayes empirical Bayes identification resulted in seven po-sitions though all with posterior probability below 90The improved test 2 from Zhang et al (2005) resulted ina value 252 just beneath the critical value 271 at the5 level but far above the critical value at the 10 level164 These two tests support the hypothesis that positiveselection acts after the duplication event in both paralogousgene clades
Together the identification of changes in selectivepressure in time and along the coding sequence indicatethat upon duplication PI paralogs underwent an increasednonsynonymous substitution rate and that this occurred
specifically in the domains shown to be necessary forproteinndashprotein interactions
Diversification in the Interaction Behavior of B-ClassGenes After the Ancient Duplication Event in BasalAsterids
As sites in the interaction domains of the PI genesfrom basal asterids were detected to be positively selectedafter duplication we further investigated the possible func-tional divergence of the duplicated PI proteins by yeast-2-hybrid analyses We tested the interactions between thefour class B genes in C japonica Camellia japonica con-tains two AP3-like genes one belonging to the euAP3-typeand one from the paleoAP3-type and two PI-like genesfrom an ancient duplication event at the base of the EricalesThis situation is similar to the B-class configuration in thecore asterid species Petunia hybrida (Vandenbussche
FIG 5mdashAlignment of the K-domain and C-terminal domain of the PI duplicates from matrix A used for the analysis in PAML Differentsubdomains and the characteristic PI motif are indicated (Kramer et al 1998) Codon positions identified to be under positive selection are marked withgray boxes
PISTILLATAmdashDuplications in Asterids 2639
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

et al 2004) and S lycopersicon (Leseberg et al 2008) butdifferent from other model species such as Arabidopsisthaliana in rosids and A majus in core asterids where onlya single pair of class B genes is present
Table 3 reconstructs the interaction specificity be-tween the class B representatives of C japonica CjDEFis able to interact with both GLO proteins (CjGLO1 andCjGLO2) whereas CjTM6 is only able to interact withCjGLO1 This situation is comparable to Petunia butthe two PI copies in both species originate from an inde-pendent duplication event very distant in asterid phylogeny(described above) Intriguingly in Camellia CjDEF is ableto form a homodimer a situation not observed so far in thecore eudicots It has been hypothesized that homodimeriza-tionwas ancestral and that in core eudicots the ability ofAP3gene products to homodimerize was lost (Schwarz-Sommeret al 1992 Riechmann et al 1996Winter et al 2002) How-ever it would be prudent to confirm this homodimeric inter-action using an independent method or to repeat theexperiment with full-length constructs before attributingin vivo relevance to this observation
We find it interesting this diversification in interactionspecificity of PI copies after duplication is analogous to thesituation in Petunia and tomato PhDEF in Petunia is ableto interact with both PhGLO1 and PhGLO2 whereasPhTM6 is only able to interact with PhGLO2 The specificinteraction in Petunia of PhDEF with PhGLO1 is respon-sible for a novel function in the flower development ofPetunia the fusion of the stamen filaments with the petaltube (Vandenbussche et al 2004) The presence of twoGLO genes in both Petunia and Solanum originates fromthe same ancient duplication event at the base of the coreeudicots as already shown in the phylogenetic analysisHowever in S lycopersicon the interaction scheme be-tween the class B genes is different with the TAP3 proteinonly interacting with LePI and TM6 interacting with theother GLO protein TPI (Leseberg et al 2008) suggestingrapid evolution of the interactions between the duplicatedB-class gene products in these two species
DiscussionAsterids Preferentially Adopted PI Duplications to DriveDiversification of B-Function
Although asterids have both AP3 and PI gene lineagesthese species have preferentially retained PI gene duplicatesduring their evolution Both at the base of the Ericales andat the base of its crown group the core asterids a duplica-tion event in the PI lineage occurred and was retained ina significant number of extant species These duplicationsare not correlated with duplications in the sister lineages ofAP3-like genes Similar ancient and evolutionarily retainedPI duplications have not been described in eudicots so farbut occurred in noneudicot groups such as basal angio-sperms where a PI duplication occurred early in the evolu-tion of the magnoliids (Stellari et al 2004) and in thegrasses where a PI duplication occurred before the diver-gence of the lineages with both rice and maize (Munsteret al 2001) Most documented PI duplications howeverhave occurred fairly recently in specific lineages within
families (Chung et al 1995 Kramer et al 1998 Kramerand Irish 1999 2000 Drea et al 2007) As in basal angio-sperms (Stellari et al 2004) PI duplications have continuedto be retained at every phylogenetic level within the asteridclade
As duplications in both the PI lineage and the AP3-likelineage have been documented outside core eudicots it isunclear why asterids would have preferentially undergoneduplications in the PI lineage rather than in the AP3 lineageThis could be the outcome of a random process Alterna-tively the prevalence of PI duplication events could reflecta shift in the regulatory interactions involving this transcrip-tion factor family In Arabidopsis the PI protein interactswith the MADS protein SEPALLATA3 (SEP3) whereasa similar AP3 construct shows only a weak interaction withSEP3 (Yang and Jack 2004) The preference for PI overAP3 duplicate retention could thus reflect their relative im-portance in establishing interactions with the SEP proteinswhich act as bridges for the formation of higher order com-plexes between MADS-domain proteins (Honma and Goto2001 Pelaz et al 2001) Furthermore it has been shown intomato that the duplicate PI products have a different inter-action behavior with the SEP gene products with one copycapable of direct interaction with SEP-like proteins and ableto form higher order complexes (Leseberg et al 2008) Thishypothesis of preferential retention of PI genes in asteridsand consequent changes in interaction specificity can befurther investigated by studying the proteinndashprotein inter-actions among AP3 PI and SEP gene products throughoutthe asterids
Duplicate Lineages in Ericales Do Not Seem to Take onDistinct and Stable Functions but Do Evolve throughSimilar Mechanisms
Our data show that after duplication PI paralogs rap-idly acquire different expression domains both early andlate in development presumably caused by changes inthe cis-regulatory regions of the duplicated genes Thismode of evolution is suggested to be important in the phe-notypic evolution of the flower (Doebley and Lukens1998) However we did not find shared derived expressionpatterns for the distinct paralogous lineages Adaptivechanges in MADS-domain protein sequences also have ap-peared to play an important role suggesting that changeswithin the coding regions of these transcriptional regulatorsalso influence phenotypic evolution of plants (Martinez-Castilla and Alvarez-Buylla 2003) Again although bothparalogs after duplication seem to have acquired substitu-tions in the amino acid domains putatively involved in pro-teinndashprotein interactions these substitutions are notcharacteristic for each clade Together these data suggestthat the duplicate lineages in Ericales have not taken on sta-ble and distinct functions through evolution at least notwhen evaluating their expression patterns and sequenceevolution
Both clades appear to have undergone changes in theirexpression patterns in addition to adaptive evolution in theirprotein-protein interaction domains Although many dupli-cated class B proteins have been shown to have undergone
2640 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

changes in cis-regulatory regions adaptive evolution in theproteinndashprotein interaction domains has also been demon-strated for several examples Both in the case of Papaver(Drea et al 2007) and Petunia (Vandenbussche et al2004) class B duplicates have taken on different proteinndashprotein interaction specificities with their interaction part-ners Also Hernandez-Hernandez et al (2007) identifiedstrong evidence of positive selection after the AP3ndashPIduplication and the euAP3ndashTM6 duplication within theK-domain which is crucial for the proteinndashproteininteractions among MADS-domain proteins (Yang et al2003 Yang and Jack 2004) Within C japonica we werealso able to show a different interaction behavior betweenthe PI duplicates with one copy interacting with DEF andthe other copy interacting with both TM6 and DEF Al-though we do not know if this interaction scheme is con-served in other species with multiple PI copies withinEricales the yeast-2-hybrid result suggests subfunctionali-zation between the two PI copies in Camellia Our datafurther show that after duplication at the base of theEricales there is positive selection on two sites these sitescorrespond to positions that reduce the PIndashSEP interactionwhen mutated in Arabidopsis (Yang and Jack 2004) Thiswould suggest that through elevated amino acid substitu-tion rates the interaction behavior of PI with SEP proteinscould have changed after duplication
From what we know of B-class protein function thusfar these results should not come as a surprise After beingexpressed in a spatially and temporally delimited domainthe protein products interact to bind DNA All three of thesebiochemical processes upstream transcriptional controlproteinndashprotein interaction and DNA binding can in prin-ciple evolve and thus far we have little evidence for any ofthese modes of evolution to be dominant indicating thata challenge ahead is describing at the biochemical levelthe function of these genes in as diverse a sampling of plantsas possible
Functional Plasticity in Asterid PI Genes
Our expression data indicate that after the PI duplica-tion during the evolution of the Ericales expression pat-terns of the duplicate copies can diverge considerablyHowever we have identified multiple cases in the Ericalesin which one duplicate was subsequently lost and these lossevents appear to be correlated with a shift in the expressionpattern of the remaining gene back to a more ancestral con-dition For instance C persicum appears to have lost oneEricales-specific PI duplicate copy However rather thanpresenting a more derived pattern of expression the Cycla-men PI gene displays a more lsquolsquoancestralrsquorsquo expression pat-tern restricted to petals and stamens These conclusionshave to be taken with caution as gene loss was shownfor only two species within the Ericales A similar obser-vation can be made for the duplication event in the PI lin-eage in the core asterid clade The initial functionalcomparison of the B function between Arabidopsis and An-tirrhinum suggested conservation between the two largeclades of core eudicots (Coen andMeyerowitz 1991) How-ever the phylogenetic history of class B genes now indi-
cates that both Arabidopsis and Antirrhinum usea simple but derived B system with the TM6 gene appar-ently absent from their genomes and only a single PI genepresent in the Arabidopsis and Antirrhinum genomes Flex-ibility is thus not only observed when it comes to the originof a duplicate PI transcription factor system which hap-pened several times during asterid evolution but also whenthe duplicated configuration is lost and the prior ancestralcondition is reinstalled
Presence of Duplicated PI Genes Correlates with FusedStamens
Both changes in the upstream transcriptional controland adaptive evolution within the coding region appearto have preserved the PI duplicates in the Ericales suggest-ing that the duplicate genes may have acquired distinct rolesin floral development Because the flower morphology ofEricales is well documented we examined the available in-formation (Fritsch 2003 Nooteboom 2003 Prance andMori 2003 Scott 2003 Stevens et al 2003 Caris and Smets2004 Caris et al 2006) in order to correlate the newly dis-covered PI duplications with derived morphologies in petaland stamen structures in basal asterids For every Ericalesspecies for which we isolated PI loci we looked at the mor-phology of the second and third whorl organs petals andstamens (table 4) All species with two PI loci due to theancient duplication event at the base of the Ericales havethe filaments of their stamens fused in a ring- or tube-likestructure suggesting that the presence of multiple PI genesis associated with this floral trait Within Styracaceae Ha-lesia has two PI copies due to the ancestral duplicationevent whereas only one PI copy was identified in StyraxBoth species have a sympetalous corolla and stamens ad-nate to the corolla tube but only Halesia has fused stamensA similar situation was observed in Actinidiaceae includ-ing A chinensis and S zahlbruckneri Only Saurauia hasretained two PI loci due to the ancient duplication event andActinidia has two copies with low sequence divergence dueto a very recent duplication event Again in this family Sau-rauia has fused stamens whereas the male reproductiveparts in Actinidia are distinct Two other species with fusedstamens I glandulifera and G brasiliensis possess two PIcopies due to a more recent duplication event Howeverthis correlation is not complete because two species fromTheophrastaceae Jacquinia auriantaca and Clavija latifo-lia both possess a staminal tube but only one PI copy wasfound in each species Thus although there are exceptionsspecies with a staminal tube have multiple PI gene copiesdue to both ancient and more recent duplication events Onepossibility is that the same subfunctionalization event hasoccurred several times independently in Ericales and thatthe same pathway responsible for stamen fusion has beenrecruited multiple times as a consequence of a gene dupli-cation Some support for this hypothesis comes from thecore asterid Petunia in which it has been demonstrated thatPI duplications are involved in fusion events within the sec-ond and third whorl organs (Vandenbussche et al 2004)Although speculative the observed correlation in Ericalesis a working hypothesis for further more functional
PISTILLATAmdashDuplications in Asterids 2641
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

research into the roles of PI duplications in basal asteridspecies
Relying on Gene Expression to Understand GeneFunction
Although comparative gene expression data haveproven useful in assessing the developmental role of a geneof interest (Lee et al 2005 Kellogg 2006) some of theobserved gene expression patterns show that speculatingon the function of a gene in establishing a specific morpho-logical trait on the basis of these gene expression patterns isnot always straightforward We show in several species thatcombined expression of a PI and AP3-like gene does notresult in a petaloıd morphology Both expression of PIand an AP3-like gene was detected in the sepals of P acau-lis although they show typical green nonpetaloid sepalsHowever all investigated petaloid and staminoid organsin this study have the combined expression of PI andAP3-like genes thought to be necessary for their organ iden-tity This seems to confirm that there is a strong correlationbetween expression and function of floral MADS-boxgenes although heterotopic expression does not alwayscorrespond to function Thus comparative gene expressionstudies through RT-PCR are useful but such results have tobe interpreted with caution To really capture the functionof a gene in a specific morphological trait functional ap-proaches like Virus Induced Gene Silencing (VIGS) arerequired (eg Hileman et al 2005) The biggest drawback
of this approach until now is the limited number of speciestested for VIGS restricting functional research in nonmodelsystems to a limited number of angiosperm species
Supplementary Material
Supplementary tables S1 and S2 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
We would like to thank Nand Buyl and VivianeLeyman from the National Botanical Garden of BelgiumPaul Uyttebroeck from the Kruidtuin in Leuven AbrahamRammeloo from the Arboretum in Kalmthout KoenKamelbeke from the Arboretum in Wespelaar Marie-Louise Demesmaeker the Berkeley Botanical GardenMark Hachadourian and Amy Litt from the New YorkBotanical Garden Art Vogel from the Hortus Botanicusin Leiden for the collection of fresh flower material Thanksto Toby Kellogg for comments on an early version of themanuscript and Anja Vandeperre for technical assistanceand support An NSF-MORPH travel fellowship wasgranted to Tom Viaene This research was supported bythe Institute for the Promotion of Innovation through Sci-ence and Technology in Flanders (IWT Vlaanderen no35040 to TV) General financial support was provided
Table 4Floral Morphology of the Second and Third Whorl Organs of Every Ericales Species and the Number of Isolated PI Loci
Family Species SympetalyPetalndashStamen
FusionConnateStamens
AncientPI-Duplication
RecentPI-Duplication
Balsaminaceae Impatiens glandulifera thorn mdash thorn xImpatiens hawkeri thorn mdash thorn
Marcgraviaceae Marcgravia umbellata thorn mdash mdashPolemoniaceae Phlox paniculata thorn thorn mdash
Acanthogilia gloriosa thorn thorn mdashIpomopsis aggregata thorn thorn mdash
Lecythidaceae Gustavia brasiliensis mdash mdash thornNapoleoneae vogelii thorn thorn mdash
Sapotaceae Synsepalum dulcificum thorn thorn mdashEbenaceae Diospyros digyna thorn thorn mdashMaesaceae Maesa argentea thorn thorn mdashTheophrastaceae Jacquinia auriantaca thorn thorn thorn
Clavija latifolia thorn thorn thornPrimulaceae Primula denticulata thorn thorn mdash xMyrsinaceae Cyclamen persicum thorn thorn mdash
Hymenandra walichii thorn thorn mdashTheaceae Camellia japonica thorn thorn thorn x
Stewartia pseudocamellia thorn thorn thorn x xSymplocaceae Symplocos chinensis thorn thorn thorn xStyracaceae Styrax japonicus thorn thorn mdash
Halesia diptera thorn thorn thorn xDiapensiaceae Galax urceolata mdash thorn thorn xRoridulaceae Roridula gorgonias mdash mdash mdashActinidiaceae Actinidia chinensis mdash mdash mdash x
Saurauia zahlbruckneri mdash thorn thorn x xSarraceniaceae Sarracenia drumondii mdash mdash mdashClethraceae Clethra tomentosa mdash mdash mdashEricaceae Erica hiemalis thorn mdash mdash x
Enkianthus campanulatus thorn thorn mdash
2642 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

by KU Leuven (OT0535) and the Fund for ScientificResearch-Flanders (Belgium) (FWO G041808)
Literature Cited
Aagaard J Willis J Phillips P 2005 Duplication of floralregulatory genes in Lamiales Am J Bot 921284ndash1293
Aagaard J Willis J Phillips P 2006 Relaxed selection amongduplicate floral regulatory genes in Lamiales J Mol Evol63493ndash503
Albach DC Soltis PS Soltis DE Olmstead RG 2001Phylogenetic analysis of asterids based on sequences of fourgenes Ann Mo Bot Gard 88163ndash212
Angenent GC Busscher M Franken J Mol JN van Tunen AJ1992 Differential expression of two MADS-box genes inwild-type and mutant Petunia flowers Plant Cell 4983ndash993
Anisimova M Gascuel O 2006 Approximate likelihood ratiotest for branches a fast accurate and powerful alternativeSyst Biol 55(4)539ndash552
Becker A Theissen G 2003 The major clades of MADS-boxgenes and their role in the development and evolution offlowering plants Mol Phylogenet Evol 29464ndash489
Bielawski J Yang Z 2003 Maximum likelihood methods fordetecting adaptive evolution after gene duplication JSFG3201ndash212
Blanc G Wolfe KH 2004 Widespread Paleopolyploidy inmodel plant species inferred from age distributions ofduplicate genes Plant Cell 161667ndash1676
Bremer B Bremer K Heidari N Erixon P Olmstead RGAnderberg AA Kallersjo M Barkhordarian E 2002 Phylo-genetics of asterids based on 3 coding and 3 non-codingchloroplast DNA markers and the utility of non-coding DNAat higher taxonomic levels Mol Phylogenet Evol 24274ndash301
Caris PL Geuten KP Janssens SB Smets EF 2006 Floraldevelopment in three species of Impatiens (Balsaminaceae)Am J Bot 931ndash14
Caris PL Smets EF 2004 A floral ontogenetic study on the sistergroup relationship between the genus Samolus (Primulaceae)and the Theophrastaceae Am J Bot 91627ndash643
Carr SM Irish VF 1997 Floral homeotic gene expressiondefines developmental arrest stages in Brassica oleraceaL vars botrytis and italica Planta 201179ndash188
Chung Y-Y Kim S-R Kang H-G Noh Y-S Park M 1995Characterization of two rice MADS box genes homologous toGLOBOSA Plant Sci 10945ndash56
Coen ES Meyerowitz EM 1991 The war of the whorls geneticinteractions controlling flower development Nature 35331ndash37
de Martino G Pan I Emmanuel E Levy A Irish VF 2006Functional analyses of two tomato APETALA3 genesdomonstrate diversification in their roles in regulating floraldevelopment Plant Cell 181833ndash1845
Doebley J Lukens L 1998 Transcriptional regulators and theevolution of plant form Plant cell 101075ndash1082
Drea S Hileman LC de Martino G Irish VF 2007 Functionalanalyses of genetic pathways controlling petal specification inpoppy Development 1344157ndash4166
Egea-Cortines M Saedler H Sommer H 1999 Ternary complexformation between the MADS-box proteins SQUAMOSADEFICIENS and GLOBOSA is involved in the control offloral architecture in Antirrhinum majus EMBO J 185370ndash5379
Felsenstein J 1985 Confidence limits on phylogenies anapproach using the bootstrap Evolution 39783ndash791
Fritsch PW 2003 Styracaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 434ndash442
Frohman MA Dush MK Martin GR 1988 Rapid production offull-length cDNAs from rare transcripts amplification usinga single gene-specific oligonucleotide primer Proc Natl AcadSci USA 858998ndash9002
Geuten K Becker A Kaufmann K Caris P Janssens SViaene T Theissen G Smets E 2006 Petaloidy and petalidentity in the balsaminoid genera Impatiens and MarcgraviaPlant J 47501ndash518
Geuten K Smets E Schols P Yuan Y-M Jannsens S Kupfer PPyck N 2004 Conflicting phylogenies of balsaminoidEricales and the polytomy in Ericales combing data ina Bayesian framework Mol Phyl Evol 31(2)711ndash729
Goto K Meyerowitz EM 1994 Function and regulation of theArabidopsis floral homeotic gene PISTILLATA Genes Dev81548ndash1560
Guindon S Gascuel O 2003 PhyML ndash a simple fast andaccurate algorithm to estimate large phylogenies by maximumlikelihood Syst Biol 52(5)696ndash704
Hernandez-Hernandez T Martinez-Castilla LP Alvarez-Buylla ER 2007 Functional diversification of B MADS-box homeotic regulators of flower development adaptiveevolution in proteinndashprotein interaction domains after majorgene duplication events Mol Biol Evol 24(2)465ndash481
Hileman LC Drea S Martino G Litt A Irish VF 2005 Virus-induced gene silencing is an effective tool for assaying geneexpression in the basal eudicot species Papaver somniferum(opium poppy) Plant J 44334ndash341
Honma T Goto K 2001 Complexes of MADS-box proteins aresufficient to convert leaves into floral organs Nature409525ndash528
Hu J-Y Saedler H 2007 Evolution of the inflated calyxsyndrome in Solanaceae Mol Biol Evol 242443ndash2453
Huelsenbeck JP Ronquist F 2001 MRBAYES Bayesianinference of phylogenetic trees Bioinformatics 17754ndash755
Irish VF 2003 The evolution of floral homeotic gene functionBioessays 25637ndash646
Jack T Brockmann LL Meyerowitz EM 1992 The homeoticgene APETALA3 of Arabidopsis thaliana encodes aMADS box and is expressed in petals and stamens Cell68683ndash697
Jack T Fox GL Meyerowitz EM 1994 Arabidopsis homeoticgene APETALA3 ectopic expression transcriptional andposttranscriptional regulation determine floral organ identityCell 76703ndash716
Kang H-G Jeon J-S Lee S An G 1998 Identification of class Band class C floral organ identity genes from rice plants PlantMol Biol 381021ndash1029
Kellogg EA 2006 Progress and challenges in studies of theevolution of development J Expl Biol 133505ndash3516
Kramer EM Dorit RL Irish VF 1998 Molecular evolution ofgenes controlling petal and stamen development duplicationand divergence within the APETALA 3 and PISTILLATAMADS-box gene lineages Genetics 149765ndash783
Kramer EM Irish VF 1999 Evolution of genetic mechanismscontrolling petal development Nature 399144ndash148
Kramer EM Irish VF 2000 Evolution of the petal and stamendevelopmental programs evidence from comparative studiesof the lower eudicots and basal angiosperms Int J Plant Sci161S29ndashS40
Kramer EM Holappa L Gould B Jaramillo MA Setnikov DSantiago PM 2007 Elaboration of B gene function to includethe identity of novel floral organs in the lower eudicotAquilegia Plant Cell 19750ndash766
Kramer EM Su H-J Wu C-C Hu J-M 2006 A simplifiedexplanation for the frameshift mutation that created a novel C-terminal motif in the APETALA 3 gene lineage BMC EvolBiol 630
PISTILLATAmdashDuplications in Asterids 2643
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

Krizek BA Meyerowitz EM 1996 Mapping the proteinregions responsible for the functional specificities of theArabidopsis MADS protein Proc Natl Acad Sci USA 934063ndash4070
Kubitzki K 2003 The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag
Kyozuka J Kobayashi T Morita M Shimamoto K 2000Spatially and temporally regulated expression of rice MADSbox genes with similarity to Arabidopsis class A B and Cgenes Plant Cell Physiol 41710ndash718
Lamb RS Irish VF 2003 Functional divergence within theAPETALA 3PISTILLATA floral homeotic gene lineages ProcNatl Acad Sci USA 276558ndash6563
Lee J-Y Baum SF Oh S-H Jiang C-Z Chen J-C Bowman JL2005 Recruitment of CRABS CLAW to promote nectarydevelopment within the eudicot clade Development 1325021ndash5032
Leseberg CH Eissler CL Wang X Johns MA Duvall MRMao L 2008 Interaction study of MADS-domain proteins intomato J Exp Biol 82253ndash2265
Liu B Wendel JF 2003 Epigenetic phenomena and theevolution of plant allopolyploids Mol Phylogen Evol 29365ndash379
Ma H Yanofsky MF Meyerowitz EM 1991 AGL1-AGL6 anArabidopsis gene family with similarity to floral homeotic andtranscription factor genes Genes Dev 5484ndash549
Maddison RP Maddison WP 2003 MacClade 4 analysis ofphylogeny and character evolution version 406 Sunderland(MA) Sinauer Associates
Maere S De Bodt S Raes J Casneuf T Van Montagu MKuiper M Van de Peer Y 2005 Modeling gene and genomeduplications in eukaryotes Proc Natl Acad Sci USA 1025454ndash5459
Martinez-Castilla LP Alvarez-Buylla ER 2003 Adaptiveevolution in the Arabidopsis MADS-box gene family inferredfrom its complete resolved phylogeny Proc Natl Acad SciUSA 100(23)13407ndash13412
McGonigle B Bouhidel K Irish VF 1996 Nuclear localizationof the Arabidopsis APETALA3 and PISTILLATA homeoticgene products depends on their simultaneous expressionGenes Dev 101812ndash1821
Mori SA Tsou C-H Wu C-C Cronholm B Anderberg AA2007 Evolution of Lecythidaceae with an emphasis on thecircumscription of neotropical genera information fromcombined ndhF and trnL-F sequence data Am J Bot 94289ndash301
Munster T Wingen LU Faigl W Werth S Saedler HTheissen G 2001 Characterization of three GLOBOSA-likeMADS-box genes from maize evidence for ancient paralogyin one class of floral homeotic B-function genes of grassesGene 2621ndash13
Nielsen R Yang Z 1998 Likelihood models for detectingpositively selected amino acid sites and applications to theHIV-1 envelope gene Genetics 148929ndash936
Nooteboom HP 2003 Symplocaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 444ndash449
Pelaz S Tapia-Lopez R Alvarez-Buylla ER Yanofsky MF2001 Conversion of leaves into petals in Arabidopsis CurrBiol 11182ndash184
Piwarzyk E Yang Y Jack T 2007 Conserved C-terminal motifsof the Arabidopsis proteins APETALA3 and PISTILLATAare dispensable for floral organ identity function Plant Phys1451495ndash1505
Pnueli L Abu-Abeid M Zamir D Nacken W Schwarz-Sommer Z Lifschitz E 1991 The MADS box gene familyin tomato temporal expression during floral development
conserved secondary structures and homology with homeoticgenes from Antirrhinum and Arabiodpsis Plant J 1255ndash266
Posada P Crandall KA 2001 Selecting the best-fit model ofnucleotide substitution Syst Biol 50580ndash601
Prance GT Mori SA 2003 Lecythidaceae In Kubitzki Keditor The families and genera of vascular plants Vol VIBerlin (Germany) Springer-Verlag p 221ndash232
Riechmann JL Krizek BA Meyerowitz EM 1996 Dimerizationspecifity ofArabidopsisdomain homeotic proteinsAPETALA1APETALA 3 PISTILLATA and AGAMOUS Nucleic AcidsRes 243134ndash3141
Rijpkema AS Royaert S Zethof J van der Weerden G Gerats TVandenbussche M 2006 Analysis of the Petunia TM6MADS box gene reveals functional divergence within theDEFAP3 lineage Plant Cell 181819ndash1832
Schonenberger J Anderberg AA Sytsma KJ 2005 MolecularPhylogenetics and patterns of floral evolution in the EricalesInt J Plant Sci 166265ndash288
Schwarz-Sommer Z Hui I Huijser P Flor PJ Hansen RTetens F Lonnig WE Saedler H Sommer H 1992Characterization of the Antirrhinum floral homeotic MADS-box gene deficiens evidence for DNA binding and autor-egulation of its persistent expression throughout flowerdevelopment EMBO J 11251ndash261
Schwarz-Sommer Z Huijser P Nacken W Saedler HSommer H 1990 Genetic control of flower developmentby homeotic genes in Antirrhinum majus Science 250931ndash936
Scott PJ 2003 Diapensiaceae In Kubitzki K editor Thefamilies and genera of vascular plants Vol VI Berlin(Germany) Springer-Verlag p 117ndash121
Shore P Sharrocks AD 1995 The MADS-box family oftranscription factors Eur J Biochem 2291ndash13
Sommer H Beltran JP Huijser P Pape H Lonnig W-ESaedler H Schwarz-Sommer Z 1990 Deficiens a homeoticgene involved in the control of flower morphogenesis inAntirrhinum majus the protein shows homology to transcrip-tion factors EMBO J 9605ndash613
Stellari GM Jaramillo MA Kramer EM 2004 Evolution of theAPETALA3 and PISTILLATA lineages of MADS-boxcontaining genes in the basal angiosperms Mol Biol Evol21506ndash519
Stevens PF 2001 Angiosperm phylogeny website Version 5(May 2004) httpwwwmobotorgMOBOTResearchAPweb
Stevens PF Dressler S Weitzman AL 2003 Theaceae InKubitzki K editor The families and genera of vascular plantsVol VI Berlin (Germany) Springer-Verlag p 463ndash471
Swofford DL 2002 PAUPPhylogenetic analysis using parsi-mony (and other methods) Version 4 Sunderland (MA)Sinauer Associates
Theissen G Kim J Saedler H 1996 Classification andphylogeny of the MADS-box multigene family suggestdefined roles of MADS-box gene subfamilies in themorphological evolution of eukaryotes J Mol Evol 43484ndash516
Theissen G Saedler H 2001 Floral quartets Nature409469ndash471
Trobner W Ramirez L Motte P Hue I Huijser P Lonnig WESaedler H Sommer H Schwarz-Sommer Z 1992 GLOBOSAa homeotic gene which interacts with DEFICIENS in thecontrol of Antirrhinum floral organogenesis EMBO J 114693ndash4704
Vandenbussche M Zethof J Royaert S Weterings K Gerats T2004 The duplicated B-class heterodimer model whorl-specific effects and complex genetic interactions in Petuniahybrida flower development Plant Cell 16741ndash754
2644 Viaene et al
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from
Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from

Van der Krol AR Brunelle A Tsuchimoto S Chua NH 1993Functional analysis of petunia floral homeotic MADS boxgene pMADS1 Genes Dev 71214ndash1228
Winter K-U Becker A Munster T Kim JT Saedler HTheissen G 1999 MADS-box genes reveal that gnetophytesare more closely related to conifers than to flowering plantsProc Natl Acad Sci USA 967342ndash7347
Winter K-U Saedler H Theissen G 2002 On the origin of classB floral homeotic genes functional substitution and dominantinhibition in Arabidopsis by expression of an orthologue fromthe gymnosperm Gnetum Plant J 31457ndash475
Wong WSW Yang Z Goldman N Nielsen R 2004 Accuracyand power of statistical methods for detecting adaptiveevolution in protein coding sequences and for identifyingpositively selected sites Genetics 1681041ndash1051
Yang Y Fanning L Jack T 2003 The K domain mediatesheterodimerization of the Arabidopsis floral organ identityproteins APETALA3 and PISTILLATA Plant J 33177ndash188
Yang Y Jack T 2004 Defining subdomains of the K domainimportant for protein-protein interactions of plant MADSproteins Plant Mol Biol 5545ndash59
Yang Z 1997 PAML a program package for phylogeneticanalysis by maximum likelihood analyses Comput ApplBiosci 13555ndash556
Yang Z 1998 Likelihood ratio tests for detecting positiveselection and application to primate lysozyme evolution MolBiol Evol 15568ndash573
Yang Z Nielsen R 1998 Synonymous and nonsynonymous ratevariation in nuclear genes of mammals J Mol Evol46409ndash418
Yang Z Nielsen R Goldman N Pederson AMK 2000 Codon-substitution models for heterogeneous selection pressure atamino acid sites Genetics 155431ndash449
Yang Z Wong WSW Nielsen R 2005 Bayes empirical Bayesinference of amino acid sites under positive selection MolBiol Evol 221107ndash1118
Yu D Kotilainen M Pollanen E Mehto M Elomaa PHelariutta Y Albert VA Teeri TH 1999 Organ identitygenes and modified patterns of flower development inGerbera hybrida (Asteraceae) Plant J 1751ndash62
Zhang J Nielsen R Yang Z 2005 Evaluation of an improvedbranch-site likelihood method for detecting positive selectionat the molecular level Mol Biol Evol 222472ndash2479
Neelima Sinha Associate Editor
Accepted August 6 2009
PISTILLATAmdashDuplications in Asterids 2645
by guest on September 6 2016
httpmbeoxfordjournalsorg
Dow
nloaded from





























