Embed Size (px)
Citation preview

Gene 227 (1999) 157–167
Cloning of a calcium channel a1 subunit from the reef-building coral,Stylophora pistillata
Didier Zoccola a,1, Eric Tambutte a,2, Francoise Senegas-Balas b, Jean-Francois Michiels c,Jean-Pierre Failla c, Jean Jaubert a, Denis Allemand a,*
a Observatoire Oceanologique Europeen, Centre Scientifique de Monaco, Avenue Saint Martin, MC-98000 Monaco, Monacob Laboratoire d’Histologie, Faculte de Medecine, Avenue de Valombrose, F-06107 Nice, Cedex 2, Francec Laboratoire d’Anatomo-Pathologie, Hopital Pasteur, Avenue de la Voie Romaine, F-06000 Nice, France
Received 26 December 1997; received in revised form 25 November 1998; accepted 5 December 1998
Abstract
While the mechanisms of cellular Ca2+ entry associated with cell activation are well characterized, the pathway of continuousuptake of the large amount of Ca2+ needed in the biomineralization process remains largely unknown. Scleractinian corals areone of the major calcifying groups of organisms. Recent studies have suggested that a voltage-dependent Ca2+ channel is involvedin the transepithelial transport of Ca2+ used for coral calcification. We report here the cloning and sequencing of a cDNA codinga coral a1 subunit Ca2+ channel. This channel is closely related to the L-type family found in vertebrates and invertebrates.Immunohistochemical analysis shows that this channel is present within the calicoblastic ectoderm, the site involved in calciumcarbonate precipitation. These data and previous results provide molecular evidence that voltage-dependent Ca2+ channels areinvolved in calcification. Cnidarians are the most primitive organisms in which a Ca2+ channel has been cloned up to now;evolutionary perspectives on Ca2+ channel diversity are discussed. © 1999 Elsevier Science B.V. All rights reserved.
Keywords: Biomineralization; Cnidaria; Protein evolution; Transepithelial calcium transport
1. Introduction amount of information has been produced in the lastfew years on biomineralization processes, the mechanism
Ca2+ ion is not only a regulatory agent in physiologi- of Ca2+ transport largely remains a biological enigmacal processes but also the primary cation used in bio- (Bawden, 1989; Lundgren and Linde, 1997).mineralized structures in plants, invertebrates and verte- In both intracellular or extracellular biomineralizationbrates (Simkiss and Wilbur, 1989). While small, rapid processes, the primary event is the entry of Ca2+ ionsand transitory Ca2+ uptake through Ca2+ channels at the cell membrane level. On the basis of pharmacolog-appears sufficient to trigger a wide variety of intracellular ical evidence, the involvement of verapamil-sensitiveprocesses, the continuous transport of large amounts of Ca2+ channels has been suggested in a few calcifyingCa2+ is needed to build a skeleton. But although a large marine invertebrates such as echinoids (reviewed in
Dubois and Chen, 1989), crustaceans (Ahearn andZhuang, 1996) and corals (Allemand and Grillo, 1992;* Corresponding author. Tel: +377 9330 1211; Fax: +377 9350 5297;
e-mail: allemand@ unice.fr Marshall, 1996; Tambutte et al., 1996). Dihydro-1 Present address: UMR 6549 CNRS, Faculte de Medecine, Avenue pyridines have been shown to suppress the uptake ofde Valombrose, F-06107 Nice Cedex 2, France. Ca2+ ions during both osteogenesis (Duriez et al., 1993)2 Present address: Service de l’environnement, 3 avenue de Fontvieille,
and dentinogenesis (Lundgren and Linde, 1997) inMC-98000 Monaco, Principality of Monaco.vertebrates. However, only one calcium channel has
Abbreviations: A Ec, aboral ectoderm; A En, aboral endoderm; bp, been cloned up to now in a calcifying cell, the osteoblastbase pairs; Co, coelenteric cavity; DHP, dihydropyridine; O Ec, oral (Meszaros et al., 1996) and this L-type calcium channelectoderm; O En, oral endoderm; ORF, open reading frame; PBS, phos- is more probably involved in calcium signalling ratherphate-buffered saline; PCR, polymerase chain reaction; RACE, rapid
than in the calcium transport process for calcificationamplification of cDNA ends; STPCACH, Stylophora pistillata calciumchanel; Z, zooxanthellae. (Meszaros et al., 1996).
0378-1119/99/$ – see front matter © 1999 Elsevier Science B.V. All rights reserved.PII: S0378-1119 ( 98 ) 00602-7

158 D. Zoccola et al. / Gene 227 (1999) 157–167
Scleractinian corals (Cnidarian, Anthozoa) are one of or fixed on a glass slide using a marine epoxy resin( UW paste, DEVCON Ltd., Ireland). Parent coloniesthe major group of calcifying animals. The rate of
calcification of a tropical coral reef is assumed to be and microcolonies were stored in an aquarium suppliedwith Mediterranean seawater (exchange rate 2%/h),around 10 kg CaCO3/m2 year (Chave et al., 1975),
consequently coral reefs play a major role in the biogeo- heated to 26.0±0.1°C and illuminated with a constantirradiance of 175 mmol photons/m2 s.chemical cycle of calcium carbonate. Aragonitic coral
skeleton is produced externally by the calicoblastic (ecto-dermal ) epithelium (Johnston, 1980). Recent pharmaco- 2.2. RT–PCRlogical studies performed in the scleractinian coralsGalaxea fascicularis (Marshall, 1996) and Stylophora Reverse transcription experiments were performed
using oligo-(dT) or specific primers (see below) with thepistillata (Tambutte et al., 1996) have demonstrated thatcalcification in these animals is inhibited by antagonists RT–PCR kit according to the instructions of the supplier
(Stratagene, La Jolla, CA, USA). PCR experimentsof L-type calcium channels, suggesting that a voltage-dependent calcium channel is involved in the transepithe- were carried out as described in Table 1 using Taq DNA
polymerase (Boehringer Manheim, Germany) on DNAlial transport of Ca2+ used for coral calcification.Despite the fact that knowledge of the molecular biology thermal cycler 480 (Perkin Elmer, Foster City, CA,
USA). Each PCR amplification product was cloned andof voltage-dependent calcium channels in invertebratesis increasing rapidly, only few molecular mechanisms five independent clones from two unrelated amplifica-
tions were sequenced at least, thus reducing sequencehave been elucidated up to now (see Skeer et al., 1996for review). errors resulting from PCR.
Several types of voltage-gated Ca2+ channels (L, T,N, P/Q and R types) have been identified primarily in 2.3. Isolation and characterization of the cDNAexcitable mammalian cells by functional criteria. Cloningof the different subunits (a1, a2, b, c and d) composing An oligo-dT primed cDNA library was constructed
in lambda ZAPA II (Stratagene) using 5 mg poly(A)+these channels revealed the major role played by the a1subunit in voltage dependence as well as in the specificity RNA isolated from Stylophora pistillata colony by the
Chomczynski method (Chomczynski and Sacchi, 1987),of the channel for calcium ions and sensitivity to variouspharmacological agents (see Perez-Reyes and Schneider, followed by one passage over oligo(dT)-cellulose
columns and the Great Lengths@ cDNA Synthesis Kit1994 for review). Two structural subfamilies of a1subunits have emerged from molecular cloning of mam- (Clontech, Palo Alto, CA, USA). It was screened with
a cDNA probe obtained by degenerate oligonucleotidemalian cDNAs: the L-type subfamily, which is pharma-cologically modulated by dihydropyridines (DHP) PCR after reverse transcription of coral mRNA. Primers
(FPA and RPA) (Horne et al., 1993) were synthesized(including a1C, a1D and a1S subunits), and the non-L-type subfamily, which is insensitive to DHP (including based on amino sequences IGMQ(V/M)FG and
VAVIMDNF localized in the highly conserved regionsP/Q type a1A, N-type a1B and R-type a1E subunits).In the present study, we develop a molecular approach in the IV S5 and IV S6 domains of previously cloned
a1 subunits. After three rounds of positive hybridization,to characterize the Ca2+ channel involved in the calcifi-cation process in the scleractinian coral, Stylophora in vivo excision of the recombinant pBluescript SK(−)
phagemid from the lambda ZAPA II clones was per-pistillata. The cloning and sequencing of the complemen-tary DNA encoding a1 subunit of this Ca2+ channel formed. DNA sequencing was carried out on both
strands by the dideoxy chain termination method usingare reported. A comparative analysis between this chan-nel and the others cloned so far in various species is a T7 sequencing kit (Pharmacia, Uppsala, Sweden).
Oligonucleotide sequences were chosen in 5∞ of the firstpresented. An immunohistochemical analysis was usedto localize this channel in coral tissue. clone and used, together with degenerate primers located
in upstream potentially conserved regions, in PCRexperiments (Table 1). For the first round, a specificprimer (RPB1) located at amino acid position2. Materials and methods1170–1176 and a degenerate primer (FPB) correspond-ing to the pore region (P loop) between the IIS5 and2.1. Biological materialIIS6 segments were used. The second round was per-formed with a specific primer (RPB2) located at aminoCloned microcolonies were propagated in the labora-
tory as previously described (Tambutte et al., 1996) acid position 1102–1108 and the same degenerateprimer. By cloning (pTAg vector, R and D Systems)from small fragments of the scleractinian coral
Stylophora pistillata. Briefly, terminal portions of and sequencing the amplified products, we obtained a1378 bp fragment including 1317 bp of a new sequencebranches (6–10 mm long) were cut from parent colonies
and either placed on a nylon net (1 mm×1 mm mesh) located 5∞ of the first one obtained. The same strategy

159D. Zoccola et al. / Gene 227 (1999) 157–167
Table 1Primers and conditions used in PCR analysis
Name Sequence Conditions for assay
FPA 5∞-ATHGGIATGCARRTITTYGG-3∞ FPA/RPA, 3 mM MgCl2RPA 5∞-TAITCRAARTTRTCCATDATIAC-3∞ 35 cycles of 1 min at 94°C,
2 min at 48°C, 1 min at 72°CFPB 5∞-ACNGGNGARGAYTGGAA-3∞ FPB/RPB1, 3 mM MgCl2RPB1 5∞-CGAATATCAGATATTCAAAGGC-3∞ 35 cycles of 1 min at 94°C,
2 min at 48°C, 1 min at 72°CRPB2 5∞-GACCATAAAGAAGGCAATGAT-3∞ FPB/RPB2, 1.5 mM MgCl2
25 cycles of 1 min at 94°C,2 min at 48°C, 1 min at 72°C
FPC 5∞-GYATHACNHTNGARGGNTGGAC-3∞ FPC/RPC1, 3 mM MgCl2RPC1 5∞-AACTGGAAGAGCATGGTAAC-3∞ 35 cycles of 1 min at 94°C,
1 min at 52°C, 2 min at 72°CRPC2 5∞-TCTCCAATGCCACCCCATGC-3∞ FPC/RPC2, 1.5 mM MgCl2
25 cycles of 1 min at 94°C,1 min at 52°C, 2 min at 72°C
RPD1 5∞-CTGTCGCCTTCAATGTCCTCA-3∞ Oligod(T)-anchor/RPD2RPD2 5∞-CGACTTGCTGCTTTTCTCTAA-3∞ Standard conditions, according to the supplier’s
instructionsRPD3 5∞-TGTCGTCTCGCCTTTTCTTTG-3∞ Anchor/RPD3
Standard conditions, according to the supplier’sinstructions
Note: H=A, T or C; I=Inosine; R=A or G; Y=C or T; D=G, A or T; N=A, G, C or T.
was reproduced with specific primers RPC1 and RPC2, Inc.). This antibody is able to inhibit the DHP-sensitivecalcium current in mouse muscle cells (Morton et al.,and a degenerate primer (FPC) corresponding to the P
loop between IS5 and IS6 segments, and an additional 1988). Cross-sections of 3 mm were sequentially incu-bated with primary mouse monoclonal anti-DHP recep-fragment of 1071 bp including 1031 bp of 5∞ sequences
was again obtained. To obtain the NH2-terminus coding tor (24 h, 4°C, 1/20 in phosphate-buffered saline (PBS)pH 7.2, 0.05 M) and a peroxidase conjugated anti-mousesequences, we used Rapid Amplification of cDNA Ends
(RACE) experiments (5∞/3∞ RACE Kit; Boehringer antibody (2 h, room temperature, 1/50 in PBS pH 7.2,0.05 M). The site of antibody binding was visualized byManheim). RPD1 primer located in the 5∞ cloned region
(amino acid position 1279–1299) was used for the incubation in diamino-benzidine (Sigma, St Louis, MO,USA). The slides were counterstained with Harris’ hae-reverse transcription of polyadenylated mRNAs isolated
from coral colonies. The single-strand cDNA obtained matoxylin. Several controls were routinely performed:(a) PBS; or (b) monoclonal mouse anti-bromo deoxyuri-was tailed with a polyA track and PCR was performed
using RPD2 oligonucleotide located at the amino acid dine (DAKO); and (c) staining with only diamino-benzidine.position 1218–1238 and a primer containing an anchor
sequence and a polyT track complementary to the Atail artificially added at the 5∞ extremity of the cDNA.A second nested PCR was performed with RPD3 primer 3. Resultslocated at the amino acid position 1174–1194 and theanchor primer. This technique produced a fragment of 3.1. cDNA cloning1238 bp which was cloned in pTAg vector and sequencedas mentioned above. The first step of cloning consisted of the construction
of a cDNA library corresponding to polyadenylatedmessengers from the coral Stylophora pistillata (see2.4. ImmunohistochemistryMaterials and Methods). Three different types of probeswere found. Databank comparisons (FASTA program)S. pistillata colonies cultured on glass slides were fixed
in 4% paraformaldehyde in 0.6 M KCl (1200 mOsm/l ). indicated that two of them corresponded to N-like(p=2.0 e−24 and 64% homology with channel M94173)They were then decalcified using Rapid Bone
Decalcifying product (Eurobio), dehydrated and embed- and P-like (p=1.7 e−21 and 76% homology with channelM64373) types of calcium channels. A probe corre-ded in paraffin. We used as primary antibody, the
MA3-920 mouse monoclonal antibody raised against sponding to a L-like-type channel was also obtained.This specific probe was then used to pick up a clonerabbit skeletal muscle DHP receptor a1 subunit (ABR,

160 D. Zoccola et al. / Gene 227 (1999) 157–167

161D. Zoccola et al. / Gene 227 (1999) 157–167
Fig
.1.A
min
oac
idal
ignm
ent
ofS
tylo
phor
api
still
ataa
1su
buni
tC
a2+c
hann
el,a
cces
sion
num
ber
U64
465
(ST
PC
AC
H)
wit
hra
bbita
1Csu
buni
t,ac
cess
ion
num
ber
X15
539
(RA
Ba1
C).
Das
hed
lines
repr
esen
tga
psin
trod
uced
byth
eC
LU
STA
Lpr
ogra
mto
opti
miz
eal
ignm
ent.
Iden
tica
lam
ino
acid
resi
dues
are
indi
cate
dby
‘*’
and
amin
oac
idco
nser
vati
vesu
bsti
tuti
ons
by‘.’
.T
hefo
urdo
mai
nsco
ntai
ning
six
puta
tive
mem
bran
esp
anni
ngre
gion
sar
em
arke
dw
ith
solid
lines
and
labe
lled
IS1
toIV
S6.
The
pore
regi
onis
labe
lled
‘Plo
op’.
The
puta
tive
N-g
lyco
syla
tion
site
sar
esh
own
as‘‡
’an
dth
epu
tati
vecA
MP
-dep
ende
ntpr
otei
nki
nase
phos
phor
ylat
ion
site
sas
‘1’.

162 D. Zoccola et al. / Gene 227 (1999) 157–167
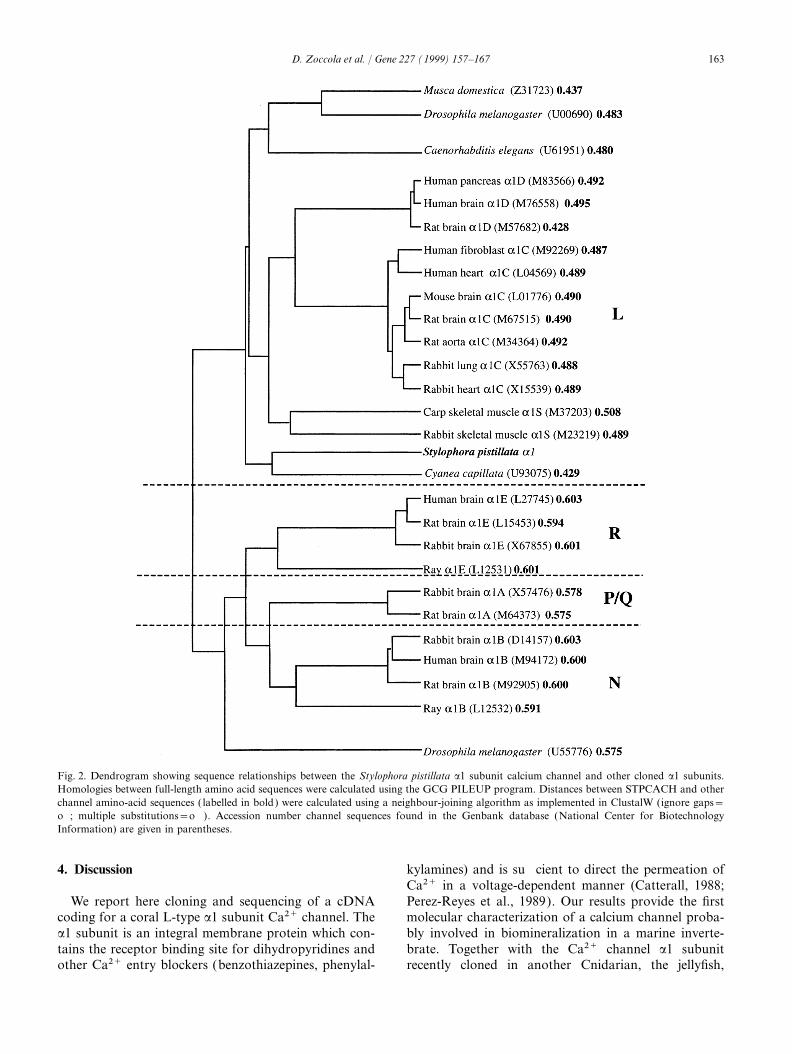
from the specific coral cDNA library. This clone con- et al., 1994), alignment of complete amino acidtained a 4146 bp insert with a potential open reading sequences of the proteins indicated that the best identityframe (ORF) of 768 amino acids and 1842 bp of a non- is obtained with Cyanea a1 subunit (56.6%) and rabbitcoding region corresponding to the most 3∞ of the a1C (52.5%), whereas less identity is obtained withmessenger. From this first clone, 5∞ oligonucleotide Drosophila melanogaster a1 (U55776) and rabbit a1A,sequences were chosen and used, together with degener- a1B and a1E (40%; 41.9%, 42.2% and 42.8%, respec-ate primers located in potentially conserved upstream tively). The PILEUP program was used to generate andregions, in PCR experiments (Table 1). By cloning and plot a dendrogram (Fig. 2) which shows the clusteringsequencing the amplified products, we obtained a new relationships used to obtain the alignments. This dendro-sequence located upstream. The same strategy was gram indicates a separate branching between the L-typerepeated and an additional fragment was again obtained. and the non-L-type subfamily and shows that the coralThe last fragment that we obtained by RACE experi- calcium channel belongs to the L-type subfamily.ments (Frohman et al., 1988) was 1238 bp long and However, the separate branching between the S. pistil-contained at the 5∞-end 110 bp of non-coding sequence lata Ca2+ channel and all the L-type subfamily channelswith stop codons in three frames. From the sequences does not permit us to classify the coral channel amongobtained by the first clone and the PCR products, one of the three subtypes known in mammals. Tocomputer analysis revealed, at 110 pb from the 5∞-end, confirm that the coral subunit belongs to the L-typea methionine in a Kozak’s context ( Kozak, 1984), subfamily, we compared the putative DHP binding sitefollowed by an ORF of 5673 bp which codes for a of the coral subunit with the rabbit ones. The IIIS6 andprotein of 1891 amino acids and a calculated molecular IVS6 domains are important regions of high affinitymass of 213 kDa. binding of DHP agonists and antagonists to L-type
Fig. 1 shows the amino acid sequence of the calcium channels (Grabner et al., 1996; Peterson et al.,Stylophora pistillata Ca2+ channel (STPCACH) as pre- 1997). The sequence aligment of IIIS6 and IVS6dicted from the 7517 bp cloned cDNAs. The structure domains of STPCACH a1 subunit (Fig. 3) shows aof this a1 subunit is typical of a voltage-dependent higher identity with the rabbit L-type subunits than withcalcium channel. It contains four repeated units of the non-L-type subunits (77% vs 42% in the IIIS6homology (amino acid residues 72–351, 469–704, region). Recently, using site-directed mutagenesis850–1115 and 1169–1411). Each repeat has five hydro-
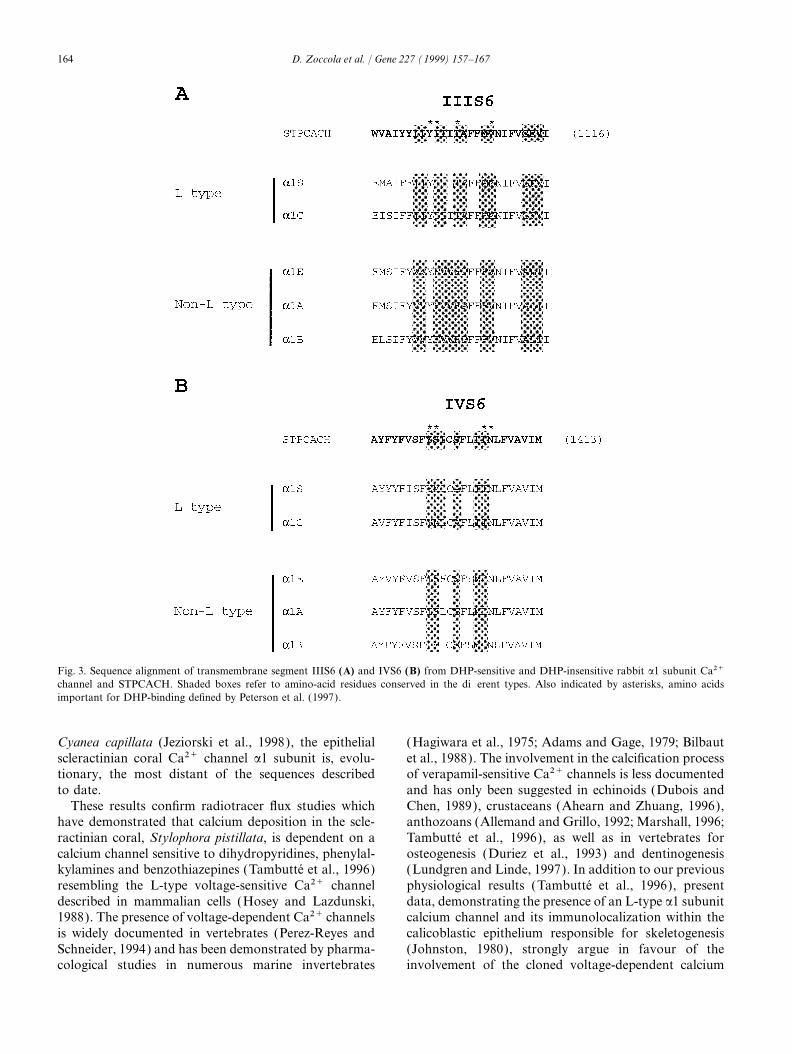
studies, Peterson et al. (1997) have identified amino-phobic segments (S1, S2, S3, S5 and S6) and one
acid residues within transmembrane segments IIIS6 andpositively charged segment (S4) with a conserved ArgIVS6 whose mutation has significant effects on DHPor Lys at every third position. Identities and conservativebinding affinity. In segment IIIS6, Tyr1099, Ile1100,substitutions between the rabbit a1C subunit and the S.Ile1103 are conserved in S. pistillata a1 subunit. However,pistillata Ca2+ channel are 52.5% and 86%, respectively.a Val1108, instead of Met, is present. In segment IVS6,The four repeat domains show a higher identity (I: 69%,the crucial Tyr1397 and Ile1405, Asn1406 are conserved.II: 67%, III: 64%, IV: 60%) than regions such as theHowever, a Ser1398, instead of Met, is present.cytoplasmic COOH-terminus region (49%). In the struc-
tural model given in Fig. 1, two of the twelve potentialN-glycosylation sites (Asn 248 and 874) are located on
3.3. Localization of the coral a1 calcium channelthe external surface. Amino acid analysis reveals severalputative phosphorylation sites for cAMP-dependent
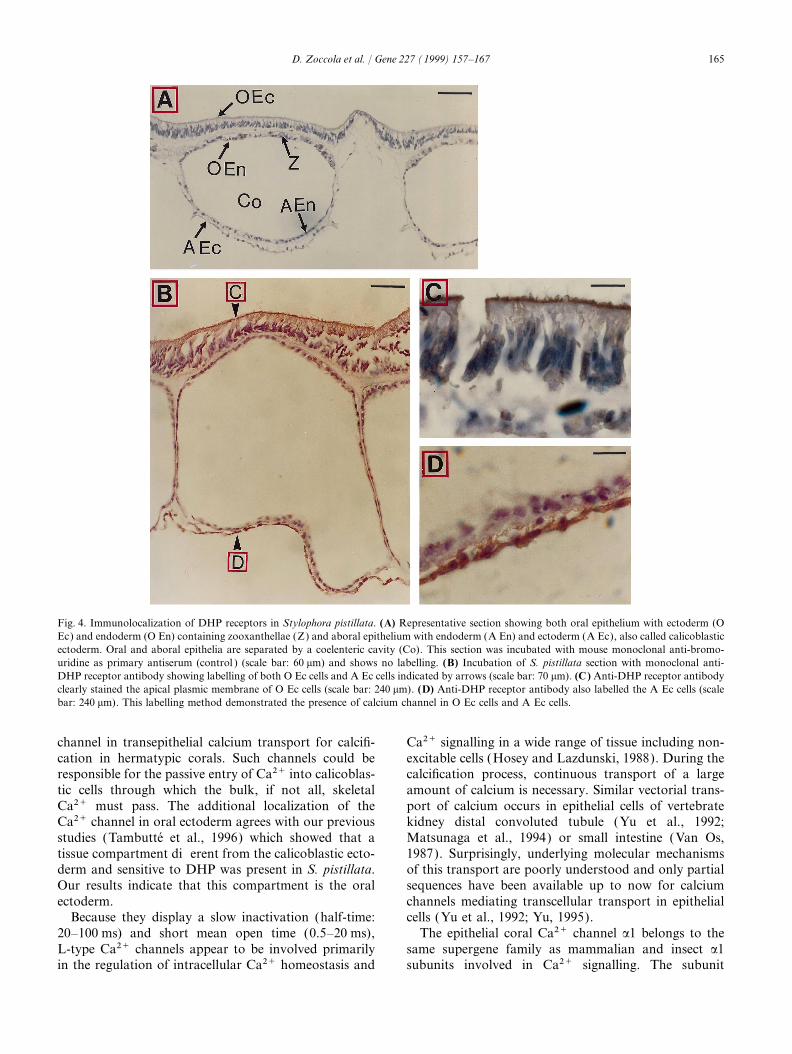
The IIIS6 and IVS6 regions involved in DHP-bindingkinase (four sites: Ser 412, Thr 460, Ser 1676 and Serdetermined in rabbit a1 subunit share 80% homology1677), protein kinase C (17 sites), or tyrosine kinasewith our cloned a1 subunit (Fig. 3). A monoclonal(one site: Tyr 373). Futhermore, primary structure showsmouse antibody, directed against these rabbit regionsthat the I–II cytoplasmic linker contains the conserved(Morton and Froehner, 1987) and able to detect mouseamino-acid sequence, sufficient for the binding of theand human a1 subunits (Morton et al., 1988, 1994),calcium channel b-subunits (Pragnell et al., 1994).was used in an immunocytochemical assay in order tolocate the channel. If the protein encoded by our cloned3.2. a1 subunit encoded by STPCACH cDNA belongs tocDNA is involved in the calcification process, we canthe L-type familypresume that it should be at least located within theepithelial cell layer facing the skeleton, i.e. the calicoblas-In order to classify the Stylophora pistillata Ca2+tic ectoderm (Johnston, 1980). Anti DHP-receptor anti-channel as an L-type or non-L-type, we compared itbody staining shows that the L-type channel is locatedwith previously cloned a1 subunits (Perez-Reyes andas expected within the calicoblastic ectoderm (Fig. 4BSchneider, 1994; Grabner et al., 1994; Wilson et al.,and D) and within the oral ectoderm (Fig. 4B and C ).1994; Zheng et al., 1995; Smith et al., 1996; Jeziorski
et al., 1998). By using clustalW program (Thompson All control tests were negative (Fig. 4A).
163D. Zoccola et al. / Gene 227 (1999) 157–167
Fig. 2. Dendrogram showing sequence relationships between the Stylophora pistillata a1 subunit calcium channel and other cloned a1 subunits.Homologies between full-length amino acid sequences were calculated using the GCG PILEUP program. Distances between STPCACH and otherchannel amino-acid sequences ( labelled in bold) were calculated using a neighbour-joining algorithm as implemented in ClustalW (ignore gaps=off; multiple substitutions=off ). Accession number channel sequences found in the Genbank database (National Center for BiotechnologyInformation) are given in parentheses.
4. Discussion kylamines) and is sufficient to direct the permeation ofCa2+ in a voltage-dependent manner (Catterall, 1988;Perez-Reyes et al., 1989). Our results provide the firstWe report here cloning and sequencing of a cDNA
coding for a coral L-type a1 subunit Ca2+ channel. The molecular characterization of a calcium channel proba-bly involved in biomineralization in a marine inverte-a1 subunit is an integral membrane protein which con-
tains the receptor binding site for dihydropyridines and brate. Together with the Ca2+ channel a1 subunitrecently cloned in another Cnidarian, the jellyfish,other Ca2+ entry blockers (benzothiazepines, phenylal-
164 D. Zoccola et al. / Gene 227 (1999) 157–167
Fig. 3. Sequence alignment of transmembrane segment IIIS6 (A) and IVS6 (B) from DHP-sensitive and DHP-insensitive rabbit a1 subunit Ca2+channel and STPCACH. Shaded boxes refer to amino-acid residues conserved in the different types. Also indicated by asterisks, amino acidsimportant for DHP-binding defined by Peterson et al. (1997).
Cyanea capillata (Jeziorski et al., 1998), the epithelial (Hagiwara et al., 1975; Adams and Gage, 1979; Bilbautet al., 1988). The involvement in the calcification processscleractinian coral Ca2+ channel a1 subunit is, evolu-
tionary, the most distant of the sequences described of verapamil-sensitive Ca2+ channels is less documentedand has only been suggested in echinoids (Dubois andto date.
These results confirm radiotracer flux studies which Chen, 1989), crustaceans (Ahearn and Zhuang, 1996),anthozoans (Allemand and Grillo, 1992; Marshall, 1996;have demonstrated that calcium deposition in the scle-
ractinian coral, Stylophora pistillata, is dependent on a Tambutte et al., 1996), as well as in vertebrates forosteogenesis (Duriez et al., 1993) and dentinogenesiscalcium channel sensitive to dihydropyridines, phenylal-
kylamines and benzothiazepines (Tambutte et al., 1996) (Lundgren and Linde, 1997). In addition to our previousphysiological results (Tambutte et al., 1996), presentresembling the L-type voltage-sensitive Ca2+ channel
described in mammalian cells (Hosey and Lazdunski, data, demonstrating the presence of an L-type a1 subunitcalcium channel and its immunolocalization within the1988). The presence of voltage-dependent Ca2+ channels
is widely documented in vertebrates (Perez-Reyes and calicoblastic epithelium responsible for skeletogenesis(Johnston, 1980), strongly argue in favour of theSchneider, 1994) and has been demonstrated by pharma-
cological studies in numerous marine invertebrates involvement of the cloned voltage-dependent calcium
165D. Zoccola et al. / Gene 227 (1999) 157–167
Fig. 4. Immunolocalization of DHP receptors in Stylophora pistillata. (A) Representative section showing both oral epithelium with ectoderm (OEc) and endoderm (O En) containing zooxanthellae (Z) and aboral epithelium with endoderm (A En) and ectoderm (A Ec), also called calicoblasticectoderm. Oral and aboral epithelia are separated by a coelenteric cavity (Co). This section was incubated with mouse monoclonal anti-bromo-uridine as primary antiserum (control ) (scale bar: 60 mm) and shows no labelling. (B) Incubation of S. pistillata section with monoclonal anti-DHP receptor antibody showing labelling of both O Ec cells and A Ec cells indicated by arrows (scale bar: 70 mm). (C) Anti-DHP receptor antibodyclearly stained the apical plasmic membrane of O Ec cells (scale bar: 240 mm). (D) Anti-DHP receptor antibody also labelled the A Ec cells (scalebar: 240 mm). This labelling method demonstrated the presence of calcium channel in O Ec cells and A Ec cells.
channel in transepithelial calcium transport for calcifi- Ca2+ signalling in a wide range of tissue including non-excitable cells (Hosey and Lazdunski, 1988). During thecation in hermatypic corals. Such channels could be
responsible for the passive entry of Ca2+ into calicoblas- calcification process, continuous transport of a largeamount of calcium is necessary. Similar vectorial trans-tic cells through which the bulk, if not all, skeletal
Ca2+ must pass. The additional localization of the port of calcium occurs in epithelial cells of vertebratekidney distal convoluted tubule (Yu et al., 1992;Ca2+ channel in oral ectoderm agrees with our previous
studies (Tambutte et al., 1996) which showed that a Matsunaga et al., 1994) or small intestine (Van Os,1987). Surprisingly, underlying molecular mechanismstissue compartment different from the calicoblastic ecto-
derm and sensitive to DHP was present in S. pistillata. of this transport are poorly understood and only partialsequences have been available up to now for calciumOur results indicate that this compartment is the oral
ectoderm. channels mediating transcellular transport in epithelialcells (Yu et al., 1992; Yu, 1995).Because they display a slow inactivation (half-time:
20–100 ms) and short mean open time (0.5–20 ms), The epithelial coral Ca2+ channel a1 belongs to thesame supergene family as mammalian and insect a1L-type Ca2+ channels appear to be involved primarily
in the regulation of intracellular Ca2+ homeostasis and subunits involved in Ca2+ signalling. The subunit

166 D. Zoccola et al. / Gene 227 (1999) 157–167
described here shows the same four repeat structures, evolution whose study may provide important andunique insights into the evolution and physiology ofeach containing six transmembrane segments, which is
the characteristic pattern for voltage-dependent Ca2+ Ca2+ channels.and Na+ channels (Catterall, 1988). Concerning theDHP-binding region, sequence aligment shows that thecoral subunit is closer to the L-type family than the Acknowledgementsnon-L-type (Fig. 2). Only two amino-acid discrepancieswere found within the residues defined by Peterson et al. This study was conducted as part of the O.O.E.(1997) as essential for DHP-binding. Despite these 1996–2000 research program. It was supported by thedifferences, it should be noted that the coral Ca2+ Council of Europe (Open Partial Agreement on Majorchannel is sensitive to the DHP antagonist, nifedipine Natural and Technological Disasters). We are indebtedand insensitive to the DHP agonist, Bay K 8644 to Dr V. Paquis and Prof. F. Cuzin for advice, support(Tambutte et al., 1996). and use of technical material. We also wish to thank
Analysis of the S. pistillata Ca2+ channel a1 subunit Prof. J.-P. Cuif, Drs M. Bidet and P. Poujeol for fruitfulprovides some evolutionary perspectives on Ca2+ chan- discussions and Prof. M. Lazdunski and Dr Tina Tentorinel diversity. Only few Ca2+ channels have been cloned for their comments on the manuscript. The nucleotidein invertebrates: one within the phylum Nematoda sequence reported in this paper has been deposited in(Caenorhabditis elegans, Wilson et al., 1994) and two the GenBank with accession numbers U64465,within the phylum Arthropoda (housefly: Grabner et al., AF098943 and AF098944.1996 and Drosophila melanogaster: Zheng et al., 1995;Smith et al., 1996). Together with the recently clonedCa2+ channel a1 subunit in the jellyfish Cyanea capillata References(Jeziorski et al., 1998), the present coral a1 subunit isthe more primitive invertebrate Ca2+ channel studied Adams, D.J., Gage, P.W., 1979. Characteristics of sodium and calciumuntil now. The a1 subunit Ca2+ channel dendrogram conductance changes produced by membrane depolarization in
Aplysia neurons. J. Physiol. Lond. 289, 143–161.(Fig. 2) is consistent with the accepted divergence ofAhearn, G.A., Zhuang, Z.P., 1996. Cellular mechanisms of calciumcnidarians early in metazoan evolution. However, to
transport in crustaceans. Physiol. Zool. 69, 383–402.further substantiate this conclusion, more invertebrateAllemand, D., Grillo, M.-C., 1992. Biocalcification mechanisms in gor-
genes need to be cloned. Based on 18S rRNA data it gonians. 45Ca uptake and deposition by the Mediterranean red coralhas been suggested that cnidarians were not monophy- Corallium rubrum. J. Exp. Zool. 292, 237–246.
Bawden, J.W., 1989. Calcium transport during mineralization. Theletic with higher metazoans (Field et al., 1988). TheAnatomical Record 224, 226–233.high degree of sequence similarity between coral and
Bilbaut, A., Hernandez-Nicaise, M.L., Leech, C.A., Meech, R.W.,vertebrate a1 subunit Ca2+ channel found in the present1988. Membrane currents that govern smooth muscle contraction
study together with the high similarity between cnidarian in a ctenophore. Nature 331, 533–535.and human integrin b1 subunits (Brower et al., 1997) Brower, D.L., Brower, S.M., Hayward, D.C., Ball, E.E., 1997. Molecu-
lar evolution of integrins: gene encoding integrin b subunits from aor nucleoporin p62 (Fischer et al., 1997) suggest thatcoral and a sponge. Proc. Natl. Acad. Sci. USA 94, 9182–9187.cnidarians appear to fall very much in the metazoan
Catterall, W.A., 1988. Structure and function of voltage-sensitive ionmainstream. In this study, we found probes for threechannels. Science 242, 50–61.
types of Ca2+ channels: one member of the L-type Chave, K.E., Smith, S.V., Roy, K.J., 1975. Carbonate production bysubfamily and two members of the non-L-type subfam- coral reefs. Marine Geol. 12, 123–140.
Chomczynski, P., Sacchi, N., 1987. Single-step method of RNA isola-ily. This suggests that L-type and non-L-type subfamiliestion by acid guanidinium thiocyanate–phenol–chloroform extrac-separated before the divergence between diploblastiction. Anal. Biochem. 162, 156–159.and triploblastic eukaryotes, at least 700 millions years
Dubois, P., Chen, C.P., 1989. Calcification in echinoderms. In: Jan-ago. The L-type S. pistillata a1 subunit Ca2+ channel goux, M., Lawrence, J.M., Eds. Echinoderm Studies, Vol. 3, Bal-does not fall into one of the three subtypes (a1C, a1D kema, A.A., Rotterdam, pp. 109–178.
Duriez, J., Flautre, B., Blary, M.C., Hardouin, P., 1993. Effects of theand a1S) known in mammals. This could be due eithercalcium channel blocker nifedipine on epiphyseal growth plate andto the great evolutionary distance between cnidariansbone turnover: a study in rabbit. Calcif. Tissue Int. 52, 120–124.and mammals, or to the existence of an unique form of
Field, K.G., Olsen, G.J., Lane, D.J., Giovannoni, S.J., Ghiselin, M.T.,L-type channel in corals, thus contrasting with that Raff, E.C., Pace, N.R., Raff, R.A., 1988. Molecular phylogeny ofdescribed for example in insects (Pelzer et al., 1989). It the animal kingdom. Science 239, 748–753.
Fischer, R., Cordes, V.C., Franke, W.W., 1997. Sequence analysis ofcan be suggested that the coral a1 subunit Ca2+ channela nuclear pore complex protein in a lower metazoan: nucleoporinhas evolved from a primitive epithelial Ca2+ channel,p62 of the coelenterate Hydra vulgaris. Gene 195, 285–293.which could represent a common ancestor of all Ca2+
Frohman, M.A., Dush, M.K., Martin, G.R., 1988. Rapid productionchannels. Appearance of L-type diversity could have of full-length cDNAs from rare transcripts: amplification using athen evolved late in the Bilateria. In this way, the coral single gene-specific oligonucleotide primer. Proc. Natl. Acad. Sci.
USA 85, 8998–9002.Ca2+ channel may then represent a step in Ca2+ channel

167D. Zoccola et al. / Gene 227 (1999) 157–167
Grabner, M., Bachmann, A., Rosenthal, S., Striessnig, J., Schulz, C., Pelzer, D., 1989. Diversity and novel pharmacological properties ofCa2+ channels in Drosophila brain membranes. EMBO J. 8,Tautz, D., Glossmann, H., 1994. Insect calcium channels. Molecular
cloning of an alpha 1-subunit from housefly (Musca domestica) 2365–2371.Perez-Reyes, E., Kim, H.S., Lacerda, A.E., Horne, W., Wei, X.,muscle. FEBS Lett. 339, 189–194.
Grabner, M., Wang, Z., Hering, S., Striessnig, J., Glossmann, H., 1996. Rampe, D., Campbell, K.P., Brown, A.M., Birnbaumer, L., 1989.Induction of calcium currents by the expression of the a1-subunit ofTransfer of 1,4-dihydropyridine sensitivity from L-type to class A
(BI) calcium channels. Neuron 16, 207–218. the dihydropyridine receptor from skeletal muscle. Nature 340,233–236.Hagiwara, S., Ozawa, S., Sand, O., 1975. Voltage-clamp analysis of
two inward current mechanisms in the egg cell membrane of a star- Perez-Reyes, E., Schneider, T., 1994. Calcium channels: structure,function, and classification. Drug Dev. Res. 33, 295–318.fish. J. Gen. Physiol. 65, 617–644.
Horne, W.A., Ellinor, P.T., Inman, I., Zhou, M., Tsien, R.W., Peterson, B.Z., Johnson, B.D., Hockerman, G.H., Acheson, M.,Scheuer, T., Catterall, W.A., 1997. Analysis of the dihydropyridineSchwarz, T.L., 1993. Molecular diversity of Ca2+ channel alpha 1
subunits from the marine ray Discopyge ommata. Proc. Natl Acad. receptor site of L-type calcium channels by alanine-scanning muta-genesis. J. Biol. Chem. 272, 18752–18758.Sci. USA 90, 3787–3791.
Hosey, M.M., Lazdunski, M., 1988. Calcium channels, molecular Pragnell, M., De Waard, M., Mori, Y., Tanabe, T., Snutch, T.P.,Campbell, K.P., 1994. Calcium channel b-subunit binds to a con-pharmacology, structure and regulation. J. Membr. Biol. 104,
81–105. served motif in the I-II cytoplasmic linker of the a1-subunit. Nature368, 67–70.Jeziorski, M.C., Greenberg, R.M., Clark, K.S., Anderson, P.A., 1998.
Cloning and functional expression of a voltage-gated calcium chan- Simkiss, K., Wilbur, K.M., 1989. Biomineralization: Cell Biology andMineral Deposition. Academic Press, New York.nel alpha1 subunit from jellyfish. J. Biol. Chem. 273, 22792–22799.
Johnston, I.S., 1980. The ultrastructure of skeletogenesis in zooxan- Skeer, J.M., Norman, R.I., Sattelle, D.B., 1996. Invertebrate voltage-dependent calcium channel subtypes. Biol. Rev. 71, 137–154.thellate corals. Int. Rev. Cytol. 67, 171–214.
Kozak, M., 1984. Compilation and analysis of sequences upstream Smith, L.A., Wang, X., Peixoto, A.A., Neumann, E.K., Hall, L.M.,Hall, J.C., 1996. A Drosophila calcium channel alpha1 subunit genefrom the translational start site in eukaryotic mRNAs. Nucleic Acids
Res. 12, 857–871. maps to a genetic locus associated with behavioral and visual defects.J. Neurosci. 16, 7868–7879.Lundgren, T., Linde, A., 1997. Voltage-gated calcium channels and
nonvoltage-gated calcium uptake pathways in the rat incisor odonto- Tambutte, E., Allemand, D., Mueller, E., Jaubert, J., 1996. A compart-mental approach to the mechanism of calcification in hermatypicblast plasma membrane. Calcif. Tissue Int. 60, 79–85.
Marshall, A.T., 1996. Calcification in hermatypic and ahermatypic corals. J. Exp. Biol. 199, 1029–1041.Thompson, J.D., Higgins, D.J., Gibson, T.J., 1994. ClustaW:corals. Science 271, 637–639.
Matsunaga, H., Stanton, B.A., Gesek, F.A., Friedman, P.A., 1994. improving the sensitivity of progressive multiple alignement throughsequence waiting, position-specific gap penalties, and weight matrixEpithelial Ca2+ channels sensitive to dihydropyridines and activated
by hyperpolarizing voltages. Am. J. Physiol. 267, C157–C165. choice. Nucleic Acid Res. 22, 4673–4680.Van Os, C.H., 1987. Transcellular calcium transport in intestinal andMeszaros, J.G., Karin, N.J., Akanbi, K., Farach-Carson, M.C., 1996.
Down regulation of L-type Ca2+ channel transcript levels by renal epithelial cells. Biochim. Biophys. Acta 906, 195–222.Wilson, R., Ainscough, R., Anderson, K. et al., 1994. 2.2 Mb of contig-1,25-dihydroxyvitamin D3. J. Biol. Chem. 271, 32981–32985.
Morton, M.E., Caffrey, J.M., Brown, A.M., Froehner, S.C., 1988. uous nucleotide sequence from chromosome III of C. elegans.Nature 368, 32–38.Monoclonal antibody to the alpha 1-subunit of the dihydropyridine-
binding complex inhibits calcium currents in BC3H1 myocytes. Yu, A.S.L., 1995. Identification and localization of calcium channela1 and b subunit isoforms in the kidney. Kidney Int. 48, 1097–1101.J. Biol. Chem. 263, 613–616.
Morton, M.E., Cassidy, T.N., Froehner, S.C., Gilmour, B.P., Laurens, Yu, A.S.L., Hebert, S.C., Brenner, B.M., Lytton, J., 1992. Molecularcharacterization and nephron distribution of a family of transcriptsR.L., 1994. Alpha 1 and alpha 2 Ca2+ channel subunit expression
in human neuronal and small cell carcinoma cells. FASEB J. 8, encoding the pore-forming subunit of Ca2+ channels in the kidney.Proc. Natl Acad. Sci. USA 89, 10494–10498.884–888.
Morton, M.E., Froehner, S.C., 1987. Monoclonal antibody identifies Zheng, W., Feng, G.P., Ren, D.J., Eberl, D.F., Hannan, F., Dubald,M., Hall, L.M., 1995. Cloning and characterization of a calciuma 200-kDa subunit of the dihydropyridine-sensitive calcium channel.
J. Biol. Chem. 262, 11904–11907. channel subunit from Drosophila melanogaster with similarity to therat brain a1 type D isoform. J. Neurosci. 15, 1132–1143.Pelzer, S., Barhanin, J., Pauron, D., Trautwein, W., Lazdunski, M.,





























