Embed Size (px)
Citation preview

(CANCER RESEARCH 52, 3760-3767. July I. 1992]
Oncogene Complementation in Fetal Brain Transplants1
Otmar D. Wiestler,2'3 Adriano Aguzzi,4 Markus Schneemann, Robert Eibl, Andreas von Deimling, and Paul Kleihues
Institute ofNeuropathology, Department of Pathology, University of Zurich, CH-8091 Zurich, Switzerland
ABSTRACT
Using a neural transplantation model and retrovirus-mediated genetransfer, we have introduced the oncogenes v-Ha-ras and \-myr into thedeveloping rat brain. Upon insertion of a construct encoding i-l la-ra.vand the Escherichia colt /3-galactosidase marker gene, the retroviralvector was found to be expressed in neurons, astrocytes, and endothelialcells of the graft. After latency periods of several months, fascicularneoplasms with expression of S-100 protein were observed in 50% of thetransplants. The foreign genes were shown to be highly expressed in thetumors and in intact donor cells, by 5-bromo-4-chloro-3-indolyl-/?-D-galactopyranoside histochemistry, indicating that an activated I la-raioncogene has the potential to initiate neoplastic transformation of glialcells. Introduction of the \-myc oncogene into 15 grafts resulted in onlya single primitive neuroectodermal tumor. However, simultaneous expression of the v-Ha-ras and \-myc genes yielded highly malignant, poh donai
neoplasms in all recipient animals, as early as 13 days after transplantation, from which cell lines could be easily derived. In addition, neoplastictransformation was also observed in vitro following introduction of rasand mir into embryonic forebrain cultures and into newborn cerebellarcultures. These data indicate a powerful complementary transformingeffect of ras and myc on neural progenitors in vivoand in vitro. Coexpres-sion of ras and myc may, therefore, provide a highly efficient tool fortransforming neural precursor cells in distinct segments of the centralnervous system at different stages of development.
INTRODUCTION
It is commonly accepted that neoplastic transformation ofprimary cells proceeds as a multistep process which involvesthe combined activation and/or inactivation of several genes. Aparadigm model for cooperation of oncogenes was establishedby Land et al. (l, 2), using primary rat embryo fibroblasts.Simultaneous introduction of activated ras and myc genes intothese cells results in malignant transformation, whereas eachoncogene alone does not induce full transformation. Accordingto their functional activity in this system, oncogenes have beenallocated to two cooperation groups (reviewed in Refs. 3-5).Members of the first group include genes encoding membrane-associated gene products such as ras, src, and the middle Tantigen of polyoma virus. Upon introduction into embryo fibroblasts, these genes cause morphological features of transformation but are unable to immortalize. Oncogenes of the mycfamily, adenovirus EIA, polyoma large T antigen, and papil-loma virus E7 induce immortalization without a significantchange in cell morphology.
Recently, cooperation of ras and myc genes has also beendemonstrated in vivo. Using retrovirus-mediated gene transferin a prostatic gland reconstitution model, Thompson et al. (6)were able to induce carcinomas with a combination of v-Ha-rasand v-myc, while the single oncogenes caused hyperplasia and
Received 11/27/91: accepted 4/17/92.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported by the Swiss National Science Foundation and the
Cancer League of the Kanton of Zürich.: Present address: Institute ofNeuropathology. University of Bonn, Sigmund-
Freud-Strasse 25 D-5300, Bonn 1. Germany.' To whom requests for reprints should be addressed.4 Present address: Research Institute of Molecular Pathology, Dr. Bohr Gasse
7. A-10.10 Vienna, Austria.
dysplastic development of prostatic cells but failed to elicitprostatic neoplasms. Also, cross-breeding of transgenic miceexpressing high levels of ras and myc in the mammary glandwas shown to enhance mammary tumorigenesis dramatically(7, 8). Clynes et al. (9) reported a high incidence of plasmocy-tomas in Pristane-pretreated mice following infection with aretroviral vector encoding both ras and myc oncogenes, whereaseach oncogene alone did not induce neoplasms.
We have developed a retrovirus-mediated gene transfer modelwhich allows us to introduce and express foreign genes inneuroectodermal and mesodermal cells of fetal brain transplants (10-12). Introduction of viral oncogenes with an intrinsicor associated protein tyrosine kinase activity into neural transplants resulted in cell type-specific tumor induction in the
grafts. While the polyoma middle T gene rapidly and selectivelyinterfered with the development of endothelial cells, expressionof \-src produced glial and mesenchymal neoplasms, followingmultistep kinetics. We have now generated neural transplantscarrying replication-defective retroviral vectors encoding v-Ha-ras, \-myc, or a combination of both oncogenes. The resultsdemonstrate a powerful cooperative transforming effect of rasand myc in the brain and indicate that gene transfer into CNS5
transplants provides a unique opportunity for studying oncogene cooperation in the nervous system.
MATERIALS AND METHODS
Retroviral Vectors. Replication-defective retroviral vectors encodingthe v-Ha-ras gene, the \-gag/myc gene of the MC29 avian myelocyto-matosis virus, or both oncogenes together were generously provided byHartmut Land (Imperial Cancer Research Fund Laboratories, London).Fig. 1 illustrates the structure of the retroviruses. All constructs arederived from the Moloney leukemia virus and have been describedelsewhere (6). A retrovirus that encodes only the neomycin phospho-transferase gene (neo) was used for control transplants. Packaging celllines which produce the appropriate virus were grown in Iscove'smodified Dulbecco's medium containing 10% newborn calf serum.
Supernatants were collected when cells were >80% confluent and werefiltered. Titer and transforming activity were tested in a focus-formingassay on NIH-3T3 cells.
Preparation of Fetal CNS Donor Cells. Timed pregnant Fischer F344rats were anesthetized at day 13.5 to 15.5 of gestation, the embryoswere removed, and the entire fetal brain was carefully dissected using astereomicroscope. Special care was taken to avoid contamination withleptomeninges. Approximately 10 fetal brains were then enzymaticallydissociated (0.25% trypsin, 0.1% DNase in phosphate-buffered saline;
10 min at room temperature) and gently triturated, in order to obtaina single-cell suspension. For retroviral infection, the cells were incubated with tissue culture supernatant of the respective packaging cellline, in the presence of polybrene (3 iig/ml), for 4 h at 37°Cand 5%
CO?, at a multiplicity of infection of 1-2. After adsorption of the virus,the cells were washed with Hanks' balanced salt solution, to remove
free retroviral particles. Cells were then pelleted and immediately usedfor transplantation. Small aliquots of these pellets were resuspendedand kept in culture, to assess cell viability and to test potential in vitroeffects on neural cells of the respective oncogene. Organotypic cultures
'The abbreviations used are: CNS, central nervous system; GFAP. glialfibrillary acidic protein; NSE, neuron-specific enolase; PCR, polymerase chainreaction: X-gal, 5-bromo-4-chloro-.Vindolyl-/i-D-galactopyranoside; BrdU,bromodeoxvuridine.
3760
on June 27, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ONCOGENE COMPLEMENTATION IN BRAIN TRANSPLANTS
LTK
neo SV(X)
DoK v-myc
ras&lacZ
ras&myc
neoK SV40 LTR
LTR v-gag/myc SV40 neo1* LTU
I
LTR_ ras lacZ LTR
LTR ^ ras ^ v-ga%/myc LTR
Fig. I. Structure of the retroviral vectors used for oncogene transfer into braintransplants. In neo SV(X), expression of the neo gene is driven by the Moloneyvirus long terminal repeat. In DoK v-myc, v-gag/myc is controlled by the longterminal repeat and neo is controlled by an internal SV40 early promoter, ras/myc and ras/lacZ. are based on the :ip vector, in which coexpression of the twogenes relies on a partial mRNA splice mechanism. The 0.7-kilobase ras complementary DNA was cloned between the endogenous splice donor (Sd) and spliceacceptor (Sa) signals of the Moloney virus. While the primary transcript leads totranslation of ras, a part of the mRNA is spliced and confers expression of thesecond gene. While ras/myc encodes v-Ha-ras and a viral form of myc, the zipras/lacZ construct encodes v-Ha-rai and an E. coli /1-galactosidase marker gene,which allows identification of target cells by histochemistry with an appropriatesubstrate. The Psi-2 packaging cell line was used to generate replication-defectiveretroviruses.
of postnatal day 2 rat cerebellum were prepared using a protocoldescribed by Trenkner and Sidman (13).
Stereotaxic Transplantation. Adult male F344 rats (150-200 g bodyweight) were used as recipients, and a stereotaxic injection system(Narishige, Tokyo, Japan) was used for transplantation. Host animalswere anesthetized with pentobarbital (20 mg/kg body weight) andreceived a single injection of 5 ^1of cell suspension, containing approximately IO6cells, into the center of the left caudoputamen (coordinates:
bregma +0.8 mm, left +2 mm).Microscopic and Immunocytochemical Analysis of the Grafts, l'osi
operatively, the host animals were closely monitored for signs ofneurological impairment. Animals with severe neurological symptomswere anesthetized with diethyl ether and sequentially perfused withphosphate-buffered saline containing 5 lU/ml heparin and with 4%buffered paraformaldehyde (pH 7.6). Frontal brain slices were embedded in paraffin and 5-^m sections were prepared following standardprocedures. Sections from all animals were stained with hematoxylin-eosin and with luxol fast blue. Selected sections were also stained withelastica-van Gieson and reticulin stains. For immunocytochemical re
actions, polyclonal antibodies to GFAP (Dakopatts, Copenhagen. Denmark), NSE (Dakopatts), factor Vlll-related antigen (Dakopatts), mye-lin basic protein (Dakopatts), and S 100 protein (Dakopatts) and monoclonal antibodies to synaptophysin (SY-38, Boehringer, Mannheim,FRG), neurofilament protein (Beckton Dickinson), and vimentin (Dakopatts) were used. Polyclonal antibodies were visualized with a commercial avidin-biotin-peroxidase kit (Dakopatts), and monoclonal antibodies with a peroxidase-antiperoxidase reaction. For cell proliferation studies, selected animals received injections of the nucleosidebromodeoxyuridine (20 mg/kg every 8 h; 48-h survival) and wereperfused with 70"¿ethanol. Brain slices were then dehydrated and
embedded in paraffin without prior paraformaldehyde fixation. Proliferating cells were detected immunocytochemically as described (14, 15),using a monoclonal antibody to Hull (Dakopatts).
Generation of Tumor-derived Cell Lines. Fresh ras/myc-induced malignant tumors were mechanically dissociated and cultured in Iscove'smodified Dulbecco's medium containing 10% fetal calf serum. Rapidly
proliferating cultures were obtained from selected tumors and kept invitro for several passages. One of these cultures (TZ 363) was found tobe clonal, by Southern blot analysis, and was used for retransplantation
and assayed for oncogene expression.Proviral Integration and Oncogene Expression. For detection of pro-
viral DNA and oncogene transcripts, genomic DNA and total RNAwere extracted from tumor cells and subjected to Southern and Northern analyses following standard protocols. DNA probes for the ras andmyc oncogenes were a generous gift of JürgenMuser (Department ofMicrobiology, University of Basel). NIH-3T3 cells transformed withthe ras/myc retroviral vector were used as a positive control.
Escherichia coli /i-galactosidase activity was visualized by X-gal his
tochemistry, as described (6. 16). Briefly, animals were perfused sequentially with 4% buffered paraformaldehyde, permeabilization buffercontaining MgCl2, sodium deoxycholate. and Nonidet P-40. and finallywith a substrate solution containing 1 mg/ml X-gal (Boehringer). Thebrain was removed and cut in slices, and the reaction was allowed toproceed for up to 72 h. The tissue was then processed for paraffinhistology. In some cases, tissues were fixed in Delaunay's solution and
frozen sections were prepared, which yielded equivalent staining results.Exon 1 of v-Ha-ras and a junction fragment of the v-gag/myc
sequence were amplified from paraffin sections of tumor tissue, usingthe PCR. Briefly, tumor-bearing transplants were excised from dewaxed5-fim sections, treated overnight with proteinase K, and subjected tothe PCR reaction, following a protocol provided by the manufacturer(Cetus Corporation). Amplification products were directly sequencedby a modified Sanger method (17). NIH-3T3 cells transformed byinfection with the ras/myc retrovirus and normal Fischer rat brainserved as positive and negative controls, respectively.
RESULTS
Development and Microscopic Appearance of Control Transplants. Donor cell suspensions were prepared from fetal ratbrains at 13.5 days of gestational age, as described (12). Theywere mock-infected or incubated with the neo SV(X) retrovirusencoding the neomycin resistance (neo) gene (Fig. 1) and weregrafted intracerebrally into syngeneic host animals. Twomonths after stereotaxic implantation, the grafts showed thecharacteristic histológica! appearance of fetal brain transplants,with an irregular distribution of neurons and glial cells andformation of pseudocortical patterns (Fig. 2, A and B). Immu-nohistochemically, all major neuroectodermal and non-neuralcell types of the CNS were identified in the grafts, usingantibodies to NSE, synaptophysin, neurofilament protein,GFAP, and myelin basic protein (Fig. 2). No evidence ofdevelopmental defects or tumorigenesis was found in thesecontrol transplants. Since retroviruses are believed to integrateonly into proliferating cells, we determined the proliferationpotential of the transplanted fetal CNS cells by labeling therecipient animals with BrdU and counting labeled cells onhistológica! sections stained with an antibody to BrdU. Asignificant fraction of donor cells was shown to be proliferating7 days after implantation (Fig. 2C).
Identification of Target Cells for Retroviral Infection. Theretroviral vector ras/lacZ, encoding both the v-Ha-rai oncogeneand the E. coli 0-galactosidase (/acZ) gene, was used to identify
target cells prone to infection and capable of expressing thetransferred genes in E13.5 CNS grafts. By double-labeling tissuesections with a chromogenic /3-galactosidase substrate (X-gal)and with antibodies to synaptophysin or to GFAP, we coulddemonstrate expression of the retroviral construct in all majorcell types of the graft, including neurons, astrocytes, and endo-thelial cells (Fig. 2D). The experiments established that it ispossible to study the effects of oncogenes in neuroectodermaland non-neuroectodermal cells by using this gene transfermodel.
3761
on June 27, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ONCOGENE COMPLEMENTATION IN BRAIN TRANSPLANTS
Fig. 2. Morphology of neural transplants. A, transverse section through the basal ganglia of a rat with a control transplant, stained with hematoxylin-eosin/luxolfast blue. The chessboard-like pattern of the caudoputamen is interrupted on the left side by a large graft (dotted line) which, 3 months after stereotaxic implantation,has smoothly integrated into the surrounding host brain. B, photomicrograph of a control transplant, stained immunohistochemically with an antibody tosynaptophysin. which demonstrates synaptic vesicles in the graft, x 400. Among other cell types, several fully differentiated neurons are visible, most of which tendto organize into patterns reminiscent of cortical architectures. C, cell proliferation in the graft. To label proliferating cells in the graft, the recipient animal receivedthree injections of the nucleoside analogue bromodeoxyuridine 48 h before death. Immunohistochemical reaction with an antibody to bromodeoxyuridine, x 400. D,retrovirus-mediated gene transfer. Target cells for the ras/lacZ retroviral vector have been visualized by X-gal histochemistry and an immunocytochemical reactionwith an antiserum to GFAP. x 400. A high number of astrocytes and several neurons (arrow) are labeled in this field.
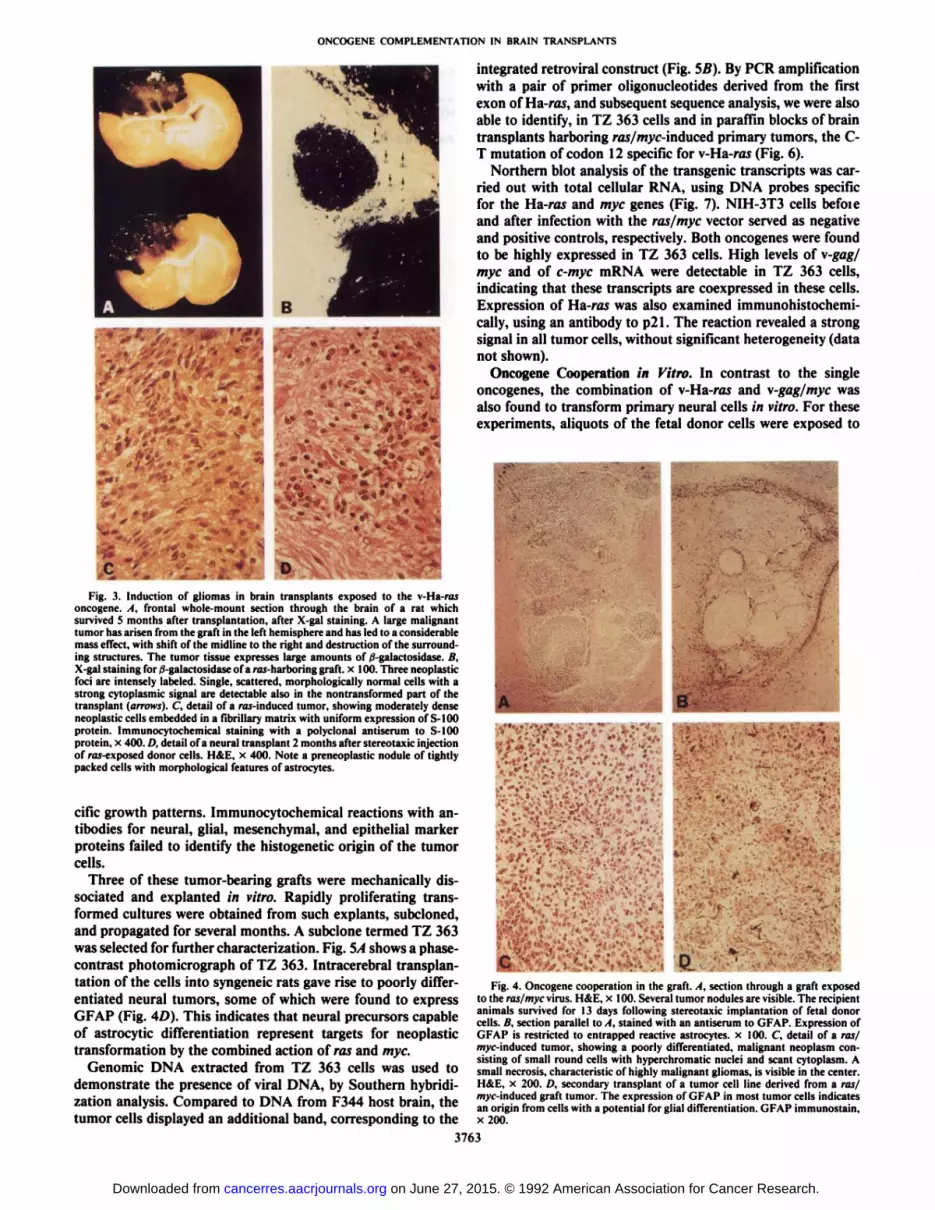
Tumor Induction in Transplants Exposed to the v-l Ia-m.vGene. A total of 10 transplants were generated with the ras/lacZ retro virus and fetal donor cells of embryonic days 13.5and 14.5. The recipient animals were observed for periods ofup to 12 months. Of eight animals with a vital graft, fourdeveloped large neoplasms in the transplant within 3-6 monthsafter implantation (Fig. 3/1). The remaining four grafts weremorphologically normal 12 months after transplantation. His-tologically, the tumors were characterized by a spindle cellmorphology and a fascicular growth pattern (Fig. 3, C and D).Extensive immunohistochemical analyses revealed uniformexpression of the neuroectodermal calcium-binding protein S-100 in these neoplasms but failed to demonstrate GFAP, neuronal marker proteins (NSE, neurofilament protein, and synaptophysin), or vimentin (Fig. 3C). Therefore, we have tentatively diagnosed the tumors as immature gliomas which havenot advanced to the stage of expression of GFAP; however, analternative histogenesis cannot be ruled out.
Expression of the retroviral vector was studied by X-galhistochemistry. It was shown previously that NIH-3T3 cellsinfected with the ras/lacZ retrovirus reliably coexpress bothgenes (6). A strong X-gal stain was detectable in tumors and inearly neoplastic foci of the transplants (Fig. 3, A and B). Inaddition, phenotypically normal cells within the residual graftwere labeled intensely, indicating that these cells tolerate theactivated oncogene without necessarily advancing to malignanttransformation. The remaining four transplanted animals didnot develop lesions.
Aliquots of the donor cell populations were established asorganotypic cultures and propagated in vitro. During an observation period of several months, no significant differences werenoted, compared to control cultures; none of the cultures developed transformed foci.
Transplants with the v-gag/myc Retroviral Vector. E13.5 donor cell populations exposed to the \-gag/myc retroviral vector
were transplanted into 13 recipient animals. Twelve of therecipients were followed for periods of 12-18 months withoutsigns of neurological impairment. Neuropathological analysisrevealed phenotypically normal CNS grafts, without evidencefor abnormal development or tumor formation. One recipientanimal developed a progressive hemiparesis 49 days after transplantation. A large tumor mass was found at the implantationsite in the left hemisphere. Microscopically, this neoplasmexhibited features of a primitive neuroectodermal tumor, withextensive areas of differentiation containing hypertrophie neurons and neoplastic glial cells (data not shown). The retroviralvector was identified in paraffin sections of the tumor by PCRamplification of a gag/myc fusion fragment, using oligonucle-otide primers to a 3'-gag/5'-myc primer of the virus (data not
shown). Suitable tumor tissue for expression analysis was notavailable. Aliquots of the infected donor cell populations wereestablished as organotypic cultures and were grown in selectionmedium containing G418 sulfate (0.5 mg/ml), starting 7 daysafter primary seeding. G418-resistant colonies derived fromthis experiment did not develop features of malignanttransformation.
Oncogene Cooperation in Fetal Brain Transplants. Donor cellswere prepared at days 13.5, 14.5, and 15.5 of embryonic development, incubated with the retroviral vector ras/myc, whichencodes both v-Ha-ras and v-gag/myc oncogenes, and stereo-taxically implanted into the brains of 13 adult recipients. Twoof the recipients did not carry vital grafts. The remaining 11host animals presented with severe neurological impairmentafter latency periods of 13-60 days. Numerous tumor noduleswere found in these transplants. A representative example of agraft with multiple neoplastic foci is shown in Fig. 4. Thisanimal survived for only 13 days following transplantation. Adetailed histopathological evaluation of these neoplasms revealed rapidly proliferating, highly malignant tumors, withoutmorphological evidence for differentiation or formation of spe-
3762
on June 27, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
ONCOGENE COMPLEMENTATION IN BRAIN TRANSPLANTS
integrated retroviral construct (Fig. 55). By PCR amplificationwith a pair of primer oligonucleotides derived from the firstexon of Ha-ras, and subsequent sequence analysis, we were alsoable to identify, in TZ 363 cells and in paraffin blocks of braintransplants harboring ras/myc-induced primary tumors, the C-T mutation of codon 12 specific for v-Ha-ras (Fig. 6).
Northern blot analysis of the transgenic transcripts was carried out with total cellular RNA, using DNA probes specificfor the Ha-ras and myc genes (Fig. 7). NIH-3T3 cells befoieand after infection with the ras/myc vector served as negativeand positive controls, respectively. Both oncogenes were foundto be highly expressed in TZ 363 cells. High levels of \-gagfmyc and of c-myc mRNA were detectable in TZ 363 cells,indicating that these transcripts are coexpressed in these cells.Expression of Ha-ras was also examined immunohistochemi-
cally, using an antibody to p21. The reaction revealed a strongsignal in all tumor cells, without significant heterogeneity (datanot shown).
Oncogene Cooperation in Vitro. In contrast to the singleoncogenes, the combination of v-Ha-ras and \-gag/myc wasalso found to transform primary neural cells in vitro. For theseexperiments, aliquots of the fetal donor cells were exposed to
Fig. 3. Induction of gliomas in brain transplants exposed to the v-Ha-rasoncogene. A. frontal whole-mount section through the brain of a rat whichsurvived 5 months after transplantation, after X-gal staining. A large malignanttumor has arisen from the graft in the left hemisphere and has led to a considerablemass effect, with shift of the midline to the right and destruction of the surrounding structures. The tumor tissue expresses large amounts of J-galactosidase. B.X-gal staining for fj-galactosidase of a ras-harboring graft, x 100. Three neoplasticfoci are intensely labeled. Single, scattered, morphologically normal cells with astrong cytoplasmic signal are detectable also in the nontransformed part of thetransplant (arrows). C, detail of a raj-induced tumor, showing moderately denseneoplastic cells embedded in a fibrillary matrix with uniform expression of S-100protein. Immunocytochemical staining with a polyclonal antiserum to S-IOOprotein, x 400. D, detail of a neural transplant 2 months after stereotaxic injectionof ras-exposed donor cells. H&E, x 400. Note a preneoplastic nodule of tightlypacked cells with morphological features of astrocytes.
cific growth patterns. Immunocytochemical reactions with antibodies for neural, glial, mesenchymal, and epithelial markerproteins failed to identify the histogenetic origin of the tumorcells.
Three of these tumor-bearing grafts were mechanically dissociated and explanted in vitro. Rapidly proliferating transformed cultures were obtained from such expiants, subcloned,and propagated for several months. A subclone termed TZ 363was selected for further characterization. Fig. 5A shows a phase-contrast photomicrograph of TZ 363. Intracerebral transplantation of the cells into syngeneic rats gave rise to poorly differentiated neural tumors, some of which were found to expressGFAP (Fig. 4D). This indicates that neural precursors capableof astrocytic differentiation represent targets for neoplastictransformation by the combined action of ras and myc.
Genomic DNA extracted from TZ 363 cells was used todemonstrate the presence of viral DNA, by Southern hybridization analysis. Compared to DNA from F344 host brain, thetumor cells displayed an additional band, corresponding to the
3763
S
•¿�•¿�•¿�.-.•-
Fig. 4. Oncogene cooperation in the graft. A, section through a graft exposedto the ras/myc virus. H&E, x 100. Several tumor nodules are visible. The recipientanimals survived for 13 days following stereotaxic implantation of fetal donorcells. B, section parallel to A. stained with an antiserum to GFAP. Expression ofGFAP is restricted to entrapped reactive astrocytes. x 100. C, detail of a ras/m>'f-induced tumor, showing a poorly differentiated, malignant neoplasm consisting of small round cells with hyperchromatic nuclei and scant cytoplasm. Asmall necrosis, characteristic of highly malignant gliomas, is visible in the center.H&E, x 200. D, secondary transplant of a tumor cell line derived from a ras/m>'c-induced graft tumor. The expression of GFAP in most tumor cells indicatesan origin from cells with a potential for glial differentiation. GFAP immunostain,X200.
on June 27, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ONCOGENE COMPLEMENTATION IN BRAIN TRANSPLANTS
Fig. 5. Characterization of the cell line TZ 363,derived from a ras/m>'c-induced graft tumor. A,phase-contrast photomicrograph, showing clustersof undifferentiated tumor cells. These cells arehighly refractile and readily form foci in vitro. B,genomic Southern blot analysis of the proviralintegration site. A band corresponding to a singleintegration is detectable in the tumor cell line TZ363 (arrows), demonstrating its clonal origin. Acomplementary DNA fragment of Ha-ras was usedas hybridization probe.
coCOCO
Ç'CO
co
C"co
CO .û
S 3N CO
-23.1 kb
Bgl
A C G T AC GT
Codon 12
TZ363 Cerebellum
132 bpgag-myc
B
E3
Fig. 6. PCR-mediated amplification of theintegrated ras/myc provirus. The first exon ofHa-ras and a gag/myc fusion fragment wereamplified from paraffin sections of normal andtumor tissue. A, sequence analysis of the firstexon of Ha-ras amplified from a ras/myc-m-duced graft tumor (TZ 363) demonstrates thediagnostic C-T point mutation in codon 12characteristic of v-Ha-ra.s. Normal rat cerebellum shows the wild-type sequence. B, detectionof a 132-base pair gag/myc fusion fragmentspecific for the v-gag/myc gene of the ras/mycretrovirus. This fragment is detectable in theras/m>T-induced graft tumor, TZ 363, and ina tumor derived from rai/m}>c-transformedperinatal cerebellar cultures, but not in normalcerebellar tissue.
the retroviral vector and then propagated in culture. Within10-14 days, multiple transformed foci appeared in these cultures and started to replace the organotypic brain reaggregates(Fig. 5/4). Such foci were never observed in control dishesexposed to a retroviral vector encoding only neo.
The newborn rodent cerebellum harbors an expanding population of proliferating neuronal precursor cells in the externalgranular cell layer. In order to examine oncogene cooperationin this tissue, organotypic cultures obtained from postnatal day2 cerebellum were infected with the ras/myc retrovirus andpropagated in vitro. Numerous transformed foci developed inthese cultures within 2-4 weeks. The neoplastic colonies didnot exhibit cytological or immunocytochemical features of neu
ronal or glial maturation. Upon intracerebral implantation intoF344 rats, aggressive neoplasms, with morphological featuresindistinguishable from those of the neoplasms induced in fore-brain grafts, developed within 2-3 weeks. Transcripts of both
oncogenes were readily detectable in total RNA prepared fromthese transplantation tumors and from in vitro transformedcells (CBP2; Fig. 4). It was also possible to demonstrate v-Ha-ras and v-gag/myc genomic sequences by PCR amplification
(data not shown). These findings indicate that both embryonicforebrain and perinatal cerebellum are highly susceptible toneoplastic transformation by the combined action of ras andmyc.
3764
on June 27, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ONCOGENE COMPLEMENTATION IN BRAIN TRANSPLANTS
t
t
t
3T3
3T3 H-ras/myc
TZ 363
CBP2
3T3
3T3 H-ras/myc
TZ 363
CBP2
Fig. 7. Northern analysis of oncogene expression. Twelve n%of total cellularRNA were extracted from parental NIH-3T3 cells (3T3). from NIH-3T3 cellstransformed by the ras/myc retroviral vector (3T3 H-ras/myc). from the TZ 363cell line derived from a ros/myr-induced transplant tumor, or from a postnatalrat cerebellum culture transformed by exposure to the ras/myc retroviral vector(CBP2). Left, expression of Ha-ras mRNA. Right, expression of \-gag/myc (upperhand) and c-myc mRNA (lower hand).
DISCUSSION
The present work was initiated by experiments which demonstrated cell type-specific transformation in brain transplantsexposed to replication-defective retroviruses encoding eitherpolyoma middle T or v-src (10-12). These studies also revealedthat all major cell types in the brain can be stably targeted withretroviral constructs and can express the transferred genes.Using retrovirus-mediated gene transfer into CNS transplants,we have now examined the effects of the v-Ha-ras and v-gag/myc oncogenes on the developing and adult nervous system invivo.
Recipient animals with transplants harboring an activated v-Ha-ras gene developed solid tumors in the graft at an incidenceof approximately 50%. Based on the histopathological featuresand immunocytochemical reactivity pattern, with no expressionof the examined markers except for S-100 protein, we tentatively classified these neoplasms as immature gliomas of astro-cytic origin (18). Because S-100 protein is not a specific markerfor glial cells, however, an alternative histogenesis cannot beexcluded. The tumors induced by v-Ha-ras exhibited a uniformmorphological appearance. This suggests that astroglial precursors are susceptible to ras-initiated transformation. Other celltypes in the graft were not visibly affected, although X-galhistochemistry demonstrated expression of the retroviral construct in neurons, glia, and endothelial cells. Similar lesionshave not been previously described, to our knowledge, in animals exposed to an activated ras gene. Transduction of activatedHa-ras into midgestation embryos resulted in a high incidenceof skin lesions in chimeric mice, without evidence for involvement of the nervous system (19, 20). Corallini et al. (21) didnot observe brain tumors following inoculation, into the brainof newborn hamsters, of a plasmid encoding activated c-Ha-ras.
Special attention was given to the neuronal compartment inthe graft, since many observations point to a role for ras genes
in neuronal differentiation pathways (22-24). Introduction ofactivated ras into PC 12 pheochromocytoma cells induces features of neuronal differentiation (25), and nerve growth factor-induced neurogenesis in PCI 2 cells can be abolished by microinjection of antibodies to p21"". Introduction of p21â„¢'in primary
neurons may obviate the requirement for neurotrophic factors(26). In Schwann cell cultures, an activated ras gene can inhibitcell proliferation (27). These observations indicate that rasproteins act in a highly cell type-specific fashion. A carefulhistopathological and immunocytochemical analysis of thetransplants failed to identify phenotypic changes in X-gal-labeled neurons, suggesting that expression of activated p21""
is tolerated in CNS neurons without causing morphologicalsigns of abnormal neurogenesis.
The latency period of several months after transplantationand the detection of /3-galactosidase activity in phenotypicallynormal neuro-glial cells within the grafts suggest that v-Ha-rasinitiates an early stage of neoplastic transformation, whichrequires additional genetic alterations for progression into macroscopic tumors. Complementing spontaneous mutations havealso been proposed in mammary carcinomas and skin tumorsof transgenic mice, with targeted expression of an activated rasgene in the respective tissue (8).
In contrast to oncogenes of the ras family, which so far havenot been associated with human brain tumors, myc genes appearto be frequently involved in the pathogenesis of neuroectoder-mal tumors. Prominent examples include amplification of theN-/n>'c gene in neuroblastomas, an event which correlates witha poor clinical outcome for the individual patient, and c-mycamplification in glioblastoma multiforme and medulloblastomacell lines (28, 29). Recently, we have demonstrated overexpres-sion of N-myc in neuroblastomas of transgenic mice expressingthe polyoma middle T antigen (30). Thus, myc was a particularly interesting candidate for introduction into CNS transplants. Intense efforts with a high titer retroviral vector failedto induce neoplastic transformation in the graft, with the exception of a primitive neuroectodermal tumor in a single recipient. The integrated provirus could be demonstrated in thistumor by PCR analysis. Since additional tumors have not beenobserved in the same and subsequent experiments, we cannotexclude the possibility that positional effects of the retroviralintegration, perhaps in combination with myc expression, mightbe responsible for tumorigenesis in this particular graft (31,32). Evidence for insertional mutagenesis was not found in anyof the neoplasms induced by ras or by ras/myc in ourexperiments.
No evidence of grossly impaired neurogenesis was seen intransplants harboring the v-gag/myc gene, suggesting that mycoverexpression is generally tolerated by transplanted CNS neurons. However, preliminary observations suggest that fetal CNScells pretreated with alkylating carcinogenic agents becomehighly susceptible to tumorigenesis when infected with the v-gag/myc retrovirus.*1 A similar observation was recently madein transgenic mice containing the pim-1 oncogene (33).
Fetal brain transplants harboring both v-Ha-ras and \-mycdemonstrated a dramatic cooperative transforming effect ofthese oncogenes in transforming neural cells in vivo. All recipient animals with vital transplants displayed multiple tumorsin the graft within 2-6 weeks of stereotaxic implantation. Thesepoorly differentiated, malignant tumors most likely originatedfrom glial precursor cells in the fetal CNS preparation, asindicated by the finding of GFAP immunoreactivity in retrans-
1O. D. Wiestier. O. Brüstle.and K. Plate, manuscript in preparation.
3765
on June 27, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ONCOGENE COMPLEMENTATION IN BRAIN TRANSPLANTS
planted cell lines derived from these neoplasms. Since integration and expression of zip retroviral vectors occur in a varietyof cell types in the graft, the predominant induction of malignant neuroectodermal tumors indicates that these progenitorcells are highly susceptible to the transforming potential of thev-Ha-ras and \-gag/myc oncogene combination. The observation of rapid induction of multiple tumors in the grafts stronglysuggests that the combination of raÃand myc may be sufficientto transform primary neural cells in vivo. Although we are notable to show clonality of single neoplasms in the graft, thenodular appearance of the tumors suggests that the singlenodules are monoclonal.
In contrast to other transforming genes which we have introduced into brain transplants, the coexpression of ras and mycalso elicits transformed foci in organotypic CNS cultures invitro. The short latency period required for focus formation invitro would, again, be compatible with a single-step transfor
mation event. Upon cytological and immunocytochemicalanalysis, it was shown that these colonies were derived fromimmature neural cells, without evidence for neuronal or glialdifferentiation. Stereotaxic implantation into the brain of syn-
geneic recipients resulted in malignant neoplasms indistinguishable from ras/myc-tnduced primary tumors. This in vitrotransformation potential was not restricted to the embryonicbrain, since cultures of postnatal rat cerebellum also gave riseto rapidly proliferating and tumorigenic transformed coloniesafter introduction of the ras/myc retroviral vector. We, therefore, conclude that neural precursor cells are a major target cellpopulation for ras/myc oncogene cooperation. Coexpression ofras and myc provides a rapid and efficient tool to transformthese cells in distinct segments of the CNS and at differentstages of neurogenesis.
Following injection of the ras/myc retroviral construct intomouse embryos, Compere et al. (19) observed a high incidenceof malignant neoplasms in the brain and in other tissues, witha median latency period of only 3 weeks. Based on their histo-pathological appearance, the intracranial tumors were classifiedas vascular meningea! neoplasms; gliomas were not observed.The authors proposed that additional genetic lesions may berequired to complete the transformation event initiated by raÃand myc; introduction of either ras or myc alone did not inducea pathological phenotype in the nervous system. An alternativeapproach for transfer of oncogenes into the brain was chosenby Corallini et al. (21 ). Plasmids encoding an activated ras gene,the BK virus early region including BK large T antigen, or acombination of both were injected into the brain of newbornhamsters. Inoculation of the construct harboring both transforming genes led to the development of intracranial sarcomaswithin 2-6 weeks, whereas the single genes did not inducetumor formation. Neuroectodermal tumors were not observed.Although these findings provide evidence for in vivo cooperation in the brain between an activated raÃgene and the BK viruslarge T antigen, the differences in the histogenetic origin ofintracranial tumors following introduction of complementingoncogenes into either neural transplants, midgestation mouseembryos, or newborn hamster brains suggest that distinct targetcells are preferentially transformed at different developmentalstages.
Retrovirus-mediated oncogene transfer into fetal brain transplants provides a novel approach to study neoplastic transformation pathways in the nervous system. In contrast to trans-genie animals, in which all cells contain exogenous DNA, targetcells for the retroviral vector develop in an environment of
unmodified normal cells, thus mimicking important features ofsporadic tumorigenesis in the brain. Using this transplantationmodel, it will be possible to evaluate the transforming potentialof alternative oncogene combinations in the CNS. Candidategenes include the N-myc oncogene and the epidermal growthfactor receptor, which have recently been implicated in thepathogenesis of human brain tumors (28, 29). Finally, genetransfer into neural transplants can also be applied to the studyof the molecular pathogenesis of other neurological disorders(34).
ACKNOWLEDGMENTS
We thank Hartmut Land for generously providing us with theretroviral vectors and for many stimulating discussions and E. F.Wagner, A. Grigoriadis, and J. Manfredi for critical reading of themanuscript. The excellent technical assistance of Marianne KönigandBrigitta Tomlinson is gratefully acknowledged.
REFERENCES
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
Land, H.. Parada, L. F., and Weinberg, R. A. Tumorigenic conversion ofprimary embryo fibroblasts requires at least two cooperating oncogenes.Nature (Lond.). 304: 596-602. 1983.Land, H.. Chen. A. C., Morganstern, J. P., Parada, L. F., and Weinberg, R.A. Behavior of myc and ras oncogenes in transformation of rat embryofibroblasts. Mol. Cell. Biol., 6: 1917-1925, 1986.Weinberg. R. A. The action of oncogenes in the cytoplasm and nucleus.Science (Washington DC), 230: 770-776. 1985.Weinberg, R. A. Oncogenes. antioncogenes. and the molecular bases ofmultistep carcinogenesis. Cancer Res.. 49: 3713-3721. 1989.Hunter. T. Cooperation between oncogenes. Cell. 64: 249-270. 1991.Thompson. T. C.. Southgate. J., Kitchener, G., and Land. H. Multistagecarcinogenesis induced by ras and myc oncogenes in a reconstituted organ.Cell. 56:917-930. 1989.Sinn, E., Muller, W., Pattengale, P., Tepler, I., Wallace, R., and Leder, P.Coexpression of MMTV/v-Ha-ras and MMTV/c-m>r genes in transgenicmice: synergistic action of oncogenes in vivo. Cell, 49:465-475, 1987.Pattengale, P. K.. Stewart, T. A., Leder, A., Sinn, E.. Muller, W., Tepler, I.,Schmidt, E., and Leder, P. Animal models of human disease. Pathology andmolecular biology of spontaneous neoplasms occuring in transgenic micecarrying and expressing activated cellular oncogenes. Am. J. Pathol., 135:39-61. 1989.Clynes. R.. Wax. J.. Stanton. L. E.. Smith-Gill. S.. Potter, M., and Marcu,K. B. Rapid induction of IgM-secreting murine plasmocytomas by Pristaneand an immunoglobulin heavy-chain promoter/enhancer-driven c-mj'c/v-Ha-ras retrovirus. Proc. Nati. Acad. Sci. USA, 85: 6067-6071, 1988.Kleihues. P.. Aguzzi, A., and Wiestier, O. D. Cellular and molecular aspectsof neurocarcinogenesis. Toxicol. Pathol.. 18: 193-203, 1990.Aguzzi, A.. Kleihues, P., Heckl, K., and Wiestier. O. D. Cell type-specifictumor induction in neural transplants by retrovirus-mediated oncogene transfer. Oncogene, 6: 113-118. 1991.Wiestier, O. D., Brüstle.O., Eibl, R. H.. Radner. H.. Aguzzi. A., and Kleihues,P. Retrovirus-mediated oncogene transfer into neural transplants. BrainPathol.. 2: 62-74, 1992.Trenkner, E.. and Sidman, R. L. Histogenesis of mouse cerebellum inmicrowell cultures. J. Cell Biol., 75: 915-940, 1978.Williams, R. L., Risau. W., Zerwes, H. G., Drexler, H., Aguzzi, A., andWagner. E. F. Endothelioma cells expressing the polyoma middle T oncogeneinduce hemangiomas by host cell recruitment. Cell, 57: 1053-1063. 1989.Gratzner, H. G. Monoclonal antibody to 5-bromo- and 5-iododeoxyuridine:a new reagent for detection of DNA replication. Science (Washington DC),218: 474-476, 1982.Cepko. C. Lineage analysis and immortalization of neural cells via retrovirusvectors. In: A. A. Boulton, G. B. Baker, and A. T. Campagnoni (eds.),Neuromethods. Vol. 16, Molecular Neurobiological Techniques, pp. 177-219. Clifton. NJ: The Humana Press. Inc., 1989.Wang, Y.. You, M.. Reynolds. S. H.. Stoner. G. D., and Anderson, M. W.Mutational activation of the cellular Harvey ras oncogene in rat esophagealpapillomas induced by methylbenzylnitrosamine. Cancer Res., 50: 1591-1595. 1990.Kleihues, P., Kiessling, M., and Janzer, R. C. Morphological markers inneuro-oncology. Curr. Top. Pathol., 77: 307-338, 1987.Compere, S. J., Baldacci, P., Sharpe, A. H.. Thompson, T.. Land, H., andJaenisch. L. The ras and myc oncogenes coooperate in tumor induction inmany tissues when introduced into midgestation mouse embryos by retroviralvectors. Proc. Nati. Acad. Sci. USA, 86: 2224-2228. 1989.Bailleul, B., Surani, M. A., White. S., Barton, S. C.. Brown, K., Blessing,M.. Jorcano. J.. and Balmain. A. Skin hyperkeratosis and papilloma forma-
3766
on June 27, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ONCOGENE COMPLEMENTATION IN BRAIN TRANSPLANTS
tion in transgenic mice expressing a ras oncogene from a suprabasal keratinpromoter. Cell, 62: 697-708, 1990.
21. Corallini, A., Pagnani, M., Caputo. A., Negrini, M., Altavilla, G., Catozzi,L., and Barbanti-Brodano. G. Cooperation in oncogenesis between BK virusearly region gene and the activated human c-Harvey ras oncogene. J. Gen.Virol.. 69:2671-2679, 1988.
22. Noda, M., Ko, M., Ogura. A., Liu. D-g., Amano, T., Takano, T., and Ikawa,Y. Sarcoma viruses carrying ras oncogenes induce differentiation-associatedproperties in a neuronal cell line. Nature (Lond.), 318: 73-75, 1985.
23. Bar-Sagi, D., and Feramisco, J. R. Microinjection of the raÃoncogene proteininto PC 12 cells induces morphological differentiation. Cell, 42: 841-848,1985.
24. Furth, M. E., Aldrich, T. H., and Cordon-Cardo, C. Expression of raj protooncogene proteins in normal human tissues. Oncogene, /: 59-69. 1987.
25. Hagag, N., Halegoua, S., and Viola, M. Inhibition of growth factor-induceddifferentiation of PCI 2 cells by microinjection of antibody to raÃp21. Nature(Lond.), 319: 680-682, 1986.
26. Borasio, G. D., John, J., Wittinghofer, A.. Barde. Y-A., Sendtner. M., andHeumann, R. Ras p21 protein promotes survival and fiber outgrowth ofcultured embryonic neurons. Neuron. 2: 1087-1096. 1989.
27. Ridley, A. J., Paterson, H. F., Noble, M., and Land, H. raj-mediated cell
cycle arrest is altered by nuclear oncogenes to induce Schwann cell transformation. EMBO J., 7: 1635-1645, 1988.
28. Schwab, M. Amplification of the MYCN oncogene and deletion of putativetumour suppressor gene in human neuroblastomas. Brain Pathol.. /: 41-46,1990.
29. Bigner, S. H., and Vogelstein, B. Cytogenetics and molecular genetics ofmalignant gliomas and medulloblastomas. Brain Pathol., /: 12-18, 1990.
30. Aguzzi, A., Wagner, E. F., Williams, R. L., and Courtneidge, S. A. Sympathetic hyperplasia and neuroblastomas in transgenic mice expressing polyomamiddle T antigen. New Biologist, 2: 533-543. 1990.
31. Woychik. R. P., Stewart. T. A., Davis, L. G., D'Eustachio, P., and Leder, P.
An inherited limb deformity created by insertional mutagenesis in a transgenic mouse. Nature (Lond"), 318: 36-40, 1985.
32. Schnieke, A., Harbers, K., and Jaenisch, R. Embryonic lethal mutation inmice induced by retrovirus insertion into the alpha\(l) collagen gene. Nature(Lond.), 304: 315-320, 1983.
33. Breuer, M., Slebos. R.. Verbeek. S.. van Lohuizen, M.. Wientjens, E., andBerns. A. Very high frequency of lymphoma induction by a chemical carcinogen in pim-\ transgenic mice. Nature (Lond.). 340: 61-63. 1989.
34. Gage, F. H., and Fisher, L. J. Intracerebral grafting: a tool for the neuro-biologist. Neuron, 6: 1-12. 1991.
3767
on June 27, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1992;52:3760-3767. Cancer Res Otmar D. Wiestler, Adriano Aguzzi, Markus Schneemann, et al. Oncogene Complementation in Fetal Brain Transplants
Updated version
http://cancerres.aacrjournals.org/content/52/13/3760
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on June 27, 2015. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from





























