Embed Size (px)
Citation preview

www.elsevier.com/locate/gloplacha
Global and Planetary Change 40 (2004) 195–200
The Mediterranean coral Cladocora caespitosa: a proxy for
past climate fluctuations?
A. Peiranoa,*, C. Morrib, C.N. Bianchia, J. Aguirrec, F. Antoniolid,G. Calzettae, L. Carobeneb, G. Mastronuzzif, P. Orrug
aENEA, Centro Ricerche Ambiente Marino, PO Box 224, 19100 La Spezia, ItalybDipTeRis, Universita di Genova, Corso Europa 26, 16132 Genoa, Italy
cDep.to Estratigrafıa y Paleontologıa, Universidad de Granada, 18071 Granada, SpaindENEA, Dipartimento Ambiente, Via Anguillarese 301, S.Maria di Galeria, 00060 Rome, Italy
eStudio Eco-X, Via del Corso, 19038 Sarzana, La Spezia, ItalyfDipartimento di Geologia e Geofisica, Via E. Orabona 4, 70125 Bari, Italy
gDiSTer, Universita di Cagliari, Via Trentino 51, 09127 Cagliari, Italy
Received 4 October 2001; accepted 7 May 2003
Abstract
Sclerochronology was applied to recent, Holocene and Pleistocene samples of Cladocora caespitosa. Late Pliocene samples
were recrystallised and thus unsuitable for sclerochronology. Quaternary samples showed a clear, alternating banding pattern as
in the living coral, confirming a marked seasonality of past climate. The computed mean annual growth rates ranged from 2.1 to
6.9 mm year� 1, with highest growth rates during the warmer phase (isotope stage 5e) of the first climate cycle. It is
hypothesised that the largest fossil banks of C. caespitosa grew in a coastal environment with considerable alluvial inputs and
warmer temperatures than today.
D 2003 Elsevier B.V. All rights reserved.
Keywords: Paleoclimatic reconstruction; Cladocora caespitosa; Coral; Mediterranean; Sclerochronology
1. Introduction reefs both in the eastern and western Mediterranean
Cladocora caespitosa (L.) is the main native zoox-
antellate and constructional coral of the Mediterranean
Sea, abundant both in the geological past and in recent
times. Large fossil Cladocora formations were found
as old as Late Pliocene, when this coral formed true
0921-8181/$ - see front matter D 2003 Elsevier B.V. All rights reserved.
doi:10.1016/S0921-8181(03)00110-3
* Corresponding author. Tel.: +39-187-978296; fax: +39-0187-
978273.
E-mail address: [email protected] (A. Peirano).
Sea (Aguirre and Jimenez, 1998; Dornbos and Wil-
son, 1999). Large fossils banks of C. caespitosa are
also known from the Early Pleistocene (Bernasconi et
al., 1997), from the Middle and Late Pleistocene and
from the Holocene (for a review, see Peirano et al.,
1998).
Today, C. caespitosa is a major carbonate producer
among Mediterranean organisms (Peirano et al.,
2001). It occurs on shallow bottoms (4–10 m) form-
ing ‘beds’ of numerous, distinct, hemispherical colo-
nies. From 10 m down to 40 m of water depth, the
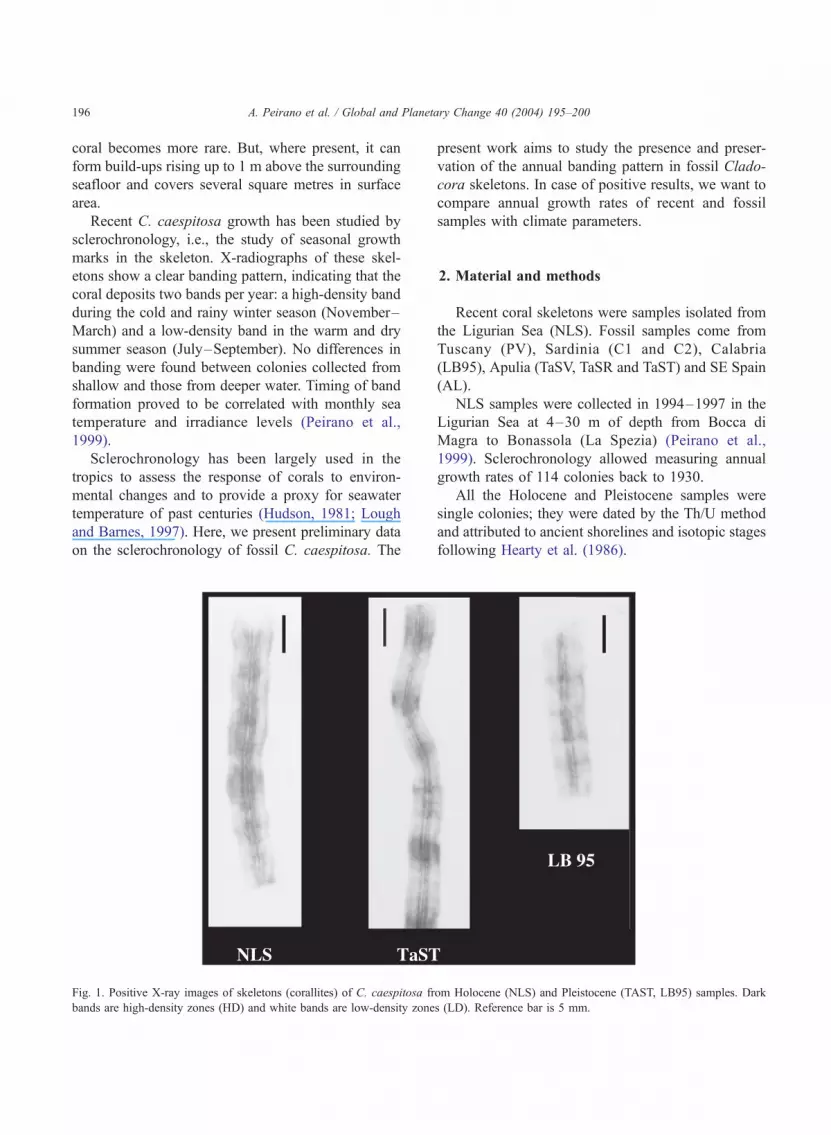
A. Peirano et al. / Global and Planetary Change 40 (2004) 195–200196
coral becomes more rare. But, where present, it can
form build-ups rising up to 1 m above the surrounding
seafloor and covers several square metres in surface
area.
Recent C. caespitosa growth has been studied by
sclerochronology, i.e., the study of seasonal growth
marks in the skeleton. X-radiographs of these skel-
etons show a clear banding pattern, indicating that the
coral deposits two bands per year: a high-density band
during the cold and rainy winter season (November–
March) and a low-density band in the warm and dry
summer season (July–September). No differences in
banding were found between colonies collected from
shallow and those from deeper water. Timing of band
formation proved to be correlated with monthly sea
temperature and irradiance levels (Peirano et al.,
1999).
Sclerochronology has been largely used in the
tropics to assess the response of corals to environ-
mental changes and to provide a proxy for seawater
temperature of past centuries (Hudson, 1981; Lough
and Barnes, 1997). Here, we present preliminary data
on the sclerochronology of fossil C. caespitosa. The
Fig. 1. Positive X-ray images of skeletons (corallites) of C. caespitosa fr
bands are high-density zones (HD) and white bands are low-density zone
present work aims to study the presence and preser-
vation of the annual banding pattern in fossil Clado-
cora skeletons. In case of positive results, we want to
compare annual growth rates of recent and fossil
samples with climate parameters.
2. Material and methods
Recent coral skeletons were samples isolated from
the Ligurian Sea (NLS). Fossil samples come from
Tuscany (PV), Sardinia (C1 and C2), Calabria
(LB95), Apulia (TaSV, TaSR and TaST) and SE Spain
(AL).
NLS samples were collected in 1994–1997 in the
Ligurian Sea at 4–30 m of depth from Bocca di
Magra to Bonassola (La Spezia) (Peirano et al.,
1999). Sclerochronology allowed measuring annual
growth rates of 114 colonies back to 1930.
All the Holocene and Pleistocene samples were
single colonies; they were dated by the Th/U method
and attributed to ancient shorelines and isotopic stages
following Hearty et al. (1986).
om Holocene (NLS) and Pleistocene (TAST, LB95) samples. Dark
s (LD). Reference bar is 5 mm.
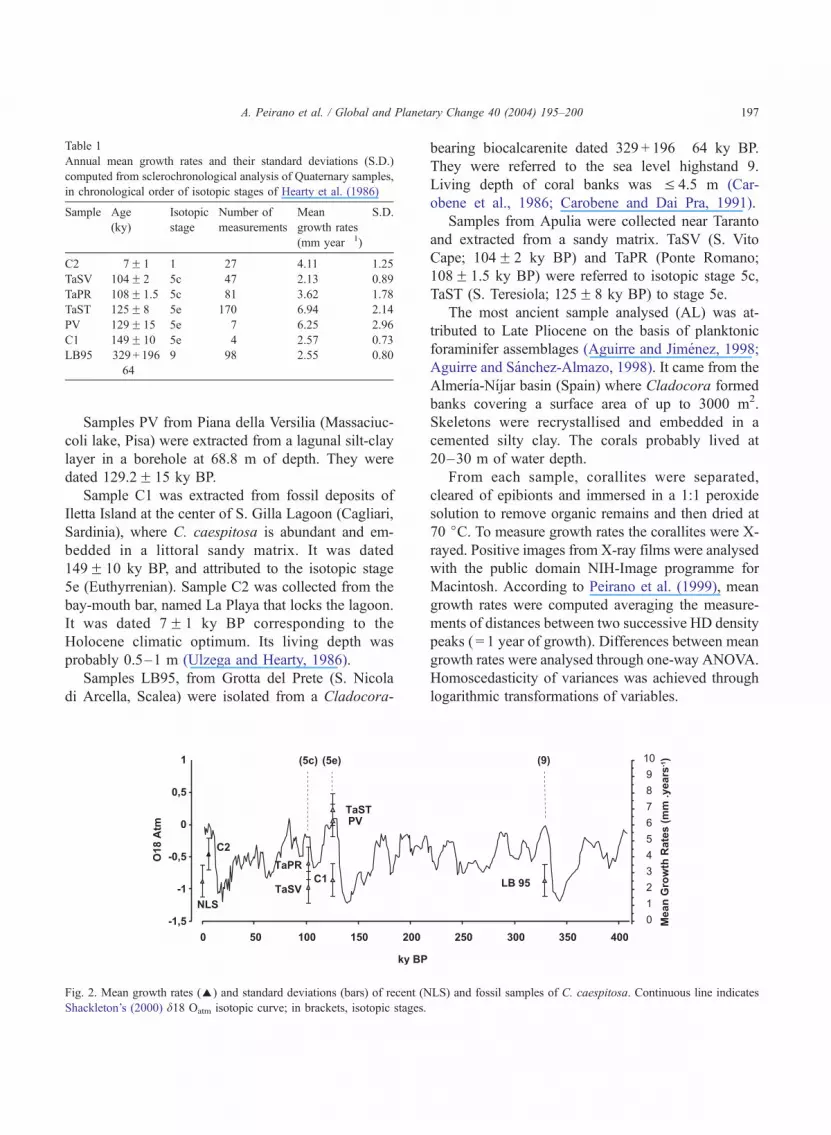
Table 1
Annual mean growth rates and their standard deviations (S.D.)
computed from sclerochronological analysis of Quaternary samples,
in chronological order of isotopic stages of Hearty et al. (1986)
Sample Age
(ky)
Isotopic
stage
Number of
measurements
Mean
growth rates
(mm year� 1)
S.D.
C2 7F 1 1 27 4.11 1.25
TaSV 104F 2 5c 47 2.13 0.89
TaPR 108F 1.5 5c 81 3.62 1.78
TaST 125F 8 5e 170 6.94 2.14
PV 129F 15 5e 7 6.25 2.96
C1 149F 10 5e 4 2.57 0.73
LB95 329 + 196
� 64
9 98 2.55 0.80
A. Peirano et al. / Global and Planetary Change 40 (2004) 195–200 197
Samples PV from Piana della Versilia (Massaciuc-
coli lake, Pisa) were extracted from a lagunal silt-clay
layer in a borehole at 68.8 m of depth. They were
dated 129.2F 15 ky BP.
Sample C1 was extracted from fossil deposits of
Iletta Island at the center of S. Gilla Lagoon (Cagliari,
Sardinia), where C. caespitosa is abundant and em-
bedded in a littoral sandy matrix. It was dated
149F 10 ky BP, and attributed to the isotopic stage
5e (Euthyrrenian). Sample C2 was collected from the
bay-mouth bar, named La Playa that locks the lagoon.
It was dated 7F 1 ky BP corresponding to the
Holocene climatic optimum. Its living depth was
probably 0.5–1 m (Ulzega and Hearty, 1986).
Samples LB95, from Grotta del Prete (S. Nicola
di Arcella, Scalea) were isolated from a Cladocora-
Fig. 2. Mean growth rates (E) and standard deviations (bars) of recent (N
Shackleton’s (2000) d18 Oatm isotopic curve; in brackets, isotopic stages.
bearing biocalcarenite dated 329 + 196� 64 ky BP.
They were referred to the sea level highstand 9.
Living depth of coral banks was V 4.5 m (Car-
obene et al., 1986; Carobene and Dai Pra, 1991).
Samples from Apulia were collected near Taranto
and extracted from a sandy matrix. TaSV (S. Vito
Cape; 104F 2 ky BP) and TaPR (Ponte Romano;
108F 1.5 ky BP) were referred to isotopic stage 5c,
TaST (S. Teresiola; 125F 8 ky BP) to stage 5e.
The most ancient sample analysed (AL) was at-
tributed to Late Pliocene on the basis of planktonic
foraminifer assemblages (Aguirre and Jimenez, 1998;
Aguirre and Sanchez-Almazo, 1998). It came from the
Almerıa-Nıjar basin (Spain) where Cladocora formed
banks covering a surface area of up to 3000 m2.
Skeletons were recrystallised and embedded in a
cemented silty clay. The corals probably lived at
20–30 m of water depth.
From each sample, corallites were separated,
cleared of epibionts and immersed in a 1:1 peroxide
solution to remove organic remains and then dried at
70 jC. To measure growth rates the corallites were X-
rayed. Positive images from X-ray films were analysed
with the public domain NIH-Image programme for
Macintosh. According to Peirano et al. (1999), mean
growth rates were computed averaging the measure-
ments of distances between two successive HD density
peaks ( = 1 year of growth). Differences between mean
growth rates were analysed through one-way ANOVA.
Homoscedasticity of variances was achieved through
logarithmic transformations of variables.
LS) and fossil samples of C. caespitosa. Continuous line indicates

A. Peirano et al. / Global and Planetary Change 40 (2004) 195–200198
3. Results
All the skeletons from the Quaternary showed a
clear banding pattern (Fig. 1). Corallites of the Plio-
cene from Spain (AL), recrystallised, did not show
growth bands. Mean annual growth rates of Quater-
nary samples ranged from 2.1 to 6.9 mm year� 1
(Table 1).
Notwithstanding the low number of fossil corals
analysed and the fact that samples came only from one
colony per site, mean growth rates were in good
accordance with Shackleton’s (2000) isotopic recon-
struction of climate cycles (Fig. 2). In particular,
samples C2, TaPR, TaSV and PV showed a positive
trend of mean growth rates that is consistent with
warm phases of the first climatic cycle. On the
contrary, C1 and LB95 did not fit with previous
samples and exhibited lower growth rates than
expected. Holocene (C2) and recent (NLS) samples
were consistent with the temperature profiles from the
Greenland ice sheet (Dahl-Jensen et al., 1998). An-
nual mean growth rates (2.5F 0.91 mm year� 1) of
recent C. caespitosa (NLS) were significantly differ-
ent from TaPR, TaST, C2 and PV (Table 1). Highest
growth rates were found in TaST (6.94F 2.14 mm
year� 1).
Fig. 3. Interannual variations of mean growth rates of the coral C. caespitos
Mediterranean) since 1866 (lower graph). Curves are the multi-annual tre
To test the possible relationship of past coral
growth with climate, growth rates of recent C. caes-
pitosa collected in the Ligurian Sea were compared to
the 150-year record of air temperature (ISTAT, 1986–
2000), which is strongly and positively correlated
with seawater temperature (Astraldi et al., 1995).
Notwithstanding the variability in the amplitude of
oscillations, due to the differences in the numbers of
corals analysed per year, the growth of C. caespitosa
was in good agreement with climatic data (Fig. 3).
Two synchronous, parallel cycles were recognised: a
sinusoidal long-term cycle of 90 years, and a shorter
cycle of about 7 years. The overall, positive trend is
in agreement with the global warming, and the
warmer climatic phase in the 1930s is consistent with
the temperature profiles from Greenland ice sheet
GRIP (Dahl-Jensen et al., 1998). The five-order
polynomial regression indicated a coral growth in-
crease of 0.7–1.6 mm year� 1 in relation to a rise in
air temperature of 0.7–1 jC. If this relationship is
applied to the fossil coral samples, then the TaST
corals should have grown in a climatic phase charac-
terised by a mean annual air temperature 3.2 jChigher than today. This is in the range of temperatures
reported by Cita et al. (1977) and Zazo (1999) for past
Mediterranean climates.
a (upper graph) compared to air temperature records of Liguria (NW
nds generated by a five-order polynomial regression.

A. Peirano et al. / Global and Planetary Change 40 (2004) 195–200 199
4. Discussion and conclusions
An identical banding pattern in fossil and recent C.
caespitosa indicates that also the past Mediterranean
climate was characterised by a marked seasonality,
with cold winters and warm summers. However, fossil
corals formed larger and higher banks than those of
today, hence the environmental conditions of past
Mediterranean Sea, in terms of both temperature and
water quality should not have been exactly the same.
The descriptions of fossil coral banks, their strati-
graphic sequences and their sedimentological features
often report their presence in sites exposed to fine
sediments and alluvial inputs (Aguirre, 1998). Fornos
et al. (1996) suggested that C. caespitosa banks
developed mainly during maximum flooding condi-
tions at the end of transgressive phases, and massive
input of sediments is supported by Peizhen et al.
(2001) who showed that in the Pliocene–Pleistocene
erosional processes were the highest since 80 Myr.
Such a turbid environment is consistent with the
observation of recent coral ‘beds’ and banks, often
found in coastal waters exposed to important terres-
trial inputs. Here C. caespitosa competes successfully
with soft, frondose algae thanks to its capability of
combining autotrophy and heterotrophy (Peirano et
al., 1998, 1999). In the Pliocene and Early Pleisto-
cene, the presence of extensive ‘beds’ of Ostrea spp.
(Bernasconi et al., 1997; Aguirre and Jimenez, 1998;
Dornbos and Wilson, 1999), forming the basement of
or being intercalated in coral banks, confirms the
assumption of turbid waters, where suspension and
filter feeding organisms were favoured.
Late Pleistocene C. caespitosa banks differed from
older banks in the presence of Strombus bubonius,
which characterised the isotopic stage 5e (our samples
C1, TaST). This gastropod, a Senegalese species, is an
indicator of a warmer climate phase. This coincides
with higher C. caespitosa growth rates (Fig. 1).
In conclusion, we can hypothesise that major fossil
reefs of C. caespitosa grew in coastal waters charac-
terised by a marked seasonality, with alluvial inputs of
fine sediments, higher turbidity and higher tempera-
ture than today. Such results, although preliminary,
suggest that C. caespitosa could be a reference species
for future work on changing Mediterranean climate,
allowing the study of climate cycles at short (annual),
medium (decadal) and long (millennia) terms.
Acknowledgements
Thanks to J. Geister (Bern) and to Kl. Oekentorp
(Muenster) for the critical reading of the manuscript.
This work has been done within the framework of the
research projects SINAPSI and Ambiente Mediterra-
neo (Accordo di programma MURST-CNR ‘‘Ecosis-
temi marini’’).
References
Aguirre, J., 1998. Bioconstrucciones de Saccostrea cuccullata Born,
1778 en el Plioceno superior de Cadiz (SO de Espana): impli-
caciones paleoambientales y paleoclimaticas. Rev. Esp. Paleon-
tol. 13, 27–36.
Aguirre, J., Jimenez, A.P., 1998. Fossil analogues of present-day
Cladocora caespitosa coral banks: Sedimentary setting, dwell-
ing community, and taphonomy (Late Pliocene, W Mediterra-
nean). Coral Reefs 17, 203–213.
Aguirre, J., Sanchez-Almazo, I.M., 1998. Foraminiferal assemb-
lages in upper Pliocene deposits of Almerıa-Nıjar basin (SE
Spain): biostratigraphic and palaeoenvironmental significance.
In: Canaveras, J.C., Garcıa del Cura, M.A., Soria, J. (Eds.),
15th International Sedimentological Congress. Abstract Volume.
Sedimentology at the Dawn of the Third Millennium University
of Alicante, Alicante, p. 120.
Astraldi, M., Bianchi, C.N., Gasparini, G.P., Morri, C., 1995. Cli-
matic fluctuations, current variability and marine species distri-
butions: a case study in the Ligurian sea (north-west mediterra-
nean). Oceanol. Acta 18, 139–149.
Bernasconi, M.P., Corselli, C., Carobene, L., 1997. A bank of the
scleractinian coral Cladocora caespitosa in the Pleistocene of
the Crati valley (Calabria, Southern Italy): growth versus en-
vironmental conditions. Boll. Soc. Paleontol. Ital. 36 (1–2),
53–61.
Carobene, L., Dai Pra, G., 1991. Genesis, chronology and tectonics
of the Quaternary marine terraces of the Tyrrhenian coast of
northern Calabria (Italy). Their correlation with climatic varia-
tions. Quaternario 3/2 (1990), 75–94.
Carobene, L., Dai Pra, G., Gewelt, M., 1986. Niveaux marins du
Pleistocene moyen-superieur de la cote tyrrhenienne de la Cala-
bre (Italie meridionale). Datations 230Tr/234U et tectonique
recente. Z. Geomorphol. N.F., Suppl. 62, 141–158.
Cita, M.B., Vergnaud-Grazzini, C., Robert, C., Chamley, H., Ciar-
anfi, N., D’onofrio, S., 1977. Paleoclimatic record of a long
deep sea core from the Eastern Mediterranean. Quat. Res. 8,
205–235.
Dahl-Jensen, D., Mosegaard, K., Gundestrup, N., Clow, G.D.,
Johnsen, S.J., Hansen, A.W., Balling, N., 1998. Past temperature
directly from the Greeland ice sheet. Science 282, 268–271.
Dornbos, S.Q., Wilson, M.A., 1999. Paleoecology of a Pliocene
coral reef in Cyprus: recovery of a marine comunity from the
Messinian Salinity Crisis. Neues Jarhrb. Geol. Palaontol. Abh.
213/1, 103–118.

A. Peirano et al. / Global and Planetary Change 40 (2004) 195–200200
Fornos, J.J., Baron, A., Pons, G.X., 1996. Arrecifes de corales
hermatıpicos (Cladocora caespitosa) en el relleno holoceno de
la zona de Es Grau (Menorca Mediterraneo Occidental). Geo-
gaceta 20, 303–306.
Hearty, P.J., Miller, G.H., Gifford, H.M., Sterans, C.E., Szabo, B.J.,
1986. Aminostratigraphy of Quaternary shorelines in the Medi-
terranean basins. Geol. Soc. Am. Bull. 97, 850–858.
Hudson, H.J., 1981. Growth rates in Montastrea annularis: a record
of environmental change in Key Largo coral reef marine sanc-
tuary, Florida. Bull. Mar. Sci. 31/2, 444–459.
ISTAT, H.J., 1986. Statistiche Metereologiche. Annuari Istat, Roma.
Lough, J.M., Barnes, D.J., 1997. Several centuries of variation
in skeletal extension, density and calcification in massive
porites colonies from the Great Barrier Reef: a proxy for
seawater temperature and a background of variability against
which to identify unnatural change. J. Exp. Mar. Biol. Ecol.
211, 29–67.
Peirano, A., Morri, C., Mastronuzzi, G., Bianchi, C.N., 1998. The
coral Cladocora caespitosa (Anthozoa, Scleractinia) as a bio-
herm builder in the Mediterranean Sea. Mem. Descr. Carta Geol.
Ital. 52 (1994), 59–74.
Peirano, A., Morri, C., Bianchi, C.N., 1999. Skeleton growth and
density pattern of the zooxanthellate scleractinian Cladocora
caespitosa (L.) from the Ligurian Sea (NW Mediterranean).
Mar. Ecol., Prog. Ser. 185, 195–201.
Peirano, A., Morri, C., Bianchi, C.N., Rodolfo-Metalpa, R., 2001.
Biomass, carbonate standing stock and production of the Med-
iterranean coral Cladocora caespitosa (L.). Facies 44, 75–80.
Peizhen, Z., Molnar, P., Downs, W.R., 2001. Increased sedimenta-
tion rates and grain sizes 2–4 Myr ago due to the influence of
climate change on erosion rates. Nature 410, 891–897.
Shackleton, N.J., 2000. The 100,000-year ice-age cycle identified
and found to lag temperature, carbon dioxide, and orbital eccen-
tricity. Science 289, 1897–1902.
Ulzega, A., Hearty, P.J., 1986. Geomorphology, stratigraphy and
geochronology of Late Quaternary marine deposits in Sardinia.
Z. Geomorphol. N.F., Suppl. 62, 119–129.
Zazo, C., 1999. Interglacial sea level. Quat. Int. 55, 101–113.



























