Embed Size (px)
Citation preview

Journal of Colloid and Interface Science 354 (2011) 181–189
Contents lists available at ScienceDirect
Journal of Colloid and Interface Science
www.elsevier .com/locate / jc is
Colloid-chemical processes in the growth and design of the bio-inorganicaragonite structure in the scleractinian coral Cladocora caespitosa
Ivan Sondi a,⇑, Branka Salopek-Sondi b, Sreco D. Škapin c, Suzana Šegota a, Irena Jurina a, Bojana Vukelic d
a Center for Marine and Environmental Research, Ru -der Boškovic Institute, Zagreb, Croatiab Department of Molecular Biology, Ru -der Boškovic Institute, Zagreb, Croatiac Advanced Materials Department, Jozef Stefan Institute, Ljubljana, Sloveniad Department of Organic Chemistry and Biochemistry, Ru -der Boškovic Institute, Zagreb, Croatia
a r t i c l e i n f o a b s t r a c t
Article history:Received 27 August 2010Accepted 22 October 2010Available online 11 November 2010
Keywords:AggregationAragoniteCoalescenceCoralsHydrolaseMesocrystalsProteinsSubmicrometer fibrous units
0021-9797/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.jcis.2010.10.055
⇑ Corresponding author. Address: Center for MarineRu -der Boškovic Institute, Bijenicka cesta 54, 100004680 242.
E-mail address: [email protected] (I. Sondi).
This study describes the morphological properties and discusses the colloid-chemical mechanisms of theformation of hierarchically structured aragonite fibers in the exoskeleton structure of the Mediterraneanzooxanthellate scleractinian coral Cladocora caespitosa. The study is based on a detailed structural andmorphological examination of the coral exoskeleton and on a preliminary biochemical and molecularidentification of the isolated soluble proteinaceus organic matrix. The biomineral structure was examinedby X-ray diffraction (XRD), field-emission scanning electron microscopy (FESEM) and atomic-forcemicroscopy (AFM), while the isolated protein organic constituents were analyzed by sodium dodecyl sul-fate–polyacrylamide gel electrophoresis (SDS–PAGE) and mass spectrometry (MALDI-TOF-MS). The SDS–PAGE analysis of the soluble protein matrix showed three major protein bands at 15, 41, and 80 kDa.Based on the MALDI-TOF-MS analyses, the identified peptides tend to exhibit an acidic character. Theresults obtained confirm and complement the existing hypotheses relating to the significant role of thesoluble acidic protein matrix and the biologically induced colloid-chemical processes in the phase forma-tion and growth of scleractinian submicrometer fibrous aragonite units. It was also shown that the gen-eral strategy for the morphogenesis of fibrous structured aragonite lies in the nanoscale aggregation andsubsequent coalescence processes that occur simultaneously. The subsequent morphological conversionof the initially formed submicrometer fibrous aragonite units into well-defined, micrometer-sized, pris-matic facets in the skeletal structures of the corals is demonstrated.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction
Aragonite, an anhydrous calcium carbonate polymorph, is acommonly occurring mineral found throughout nature and is theprimary constituent of the bio-inorganic structures of the skele-tons of many mineralizing organisms [1]. Numerous studies reportevidence that these organisms selectively form bio-inorganic solidsof the aragonite structure with unusual structural and morpholog-ical properties [2–6]. In comparison with their abiotic analogues,these solids have surprisingly sophisticated designs, often appear-ing in complex hierarchical shapes that provide superior and mul-tifunctional physical properties, particularly when taking intoaccount that they were formed under ambient conditions [7–9].
Many organisms have developed their own strategies in the for-mation of bio-inorganic structures with desirable properties that
ll rights reserved.
and Environmental Research,Zagreb, Croatia. Fax: +385 1
support their life functions and their adaptation to the environ-ment [10]. The formation of such structures is highly controlled,from the submicrometer size to macroscopic levels, and governedby organic macromolecules, mostly peptides, polypeptides, pro-teins, and polysaccharides [11–15]. These compounds are directlyinvolved in regulating the nucleation, growth and shaping of bio-inorganic solids [12,16,17]. The number of reported studies onthe function, structure and morphology of biominerals has beenincreasing in recent years. These studies have generated consider-able interest in the geo-, bio- and material sciences.
Scleractinian corals are some of the most prolific calcifyingorganisms in nature. An understanding of the structure, morpho-genesis and self-organization of the complex form of biomineralarchitecture provides an insight into the formation of the inorganiccomponents in the skeletons and tissues of many mineralizingorganisms. At the same time, it assists in the development ofnew biomimetical methods capable of generating novel bio-inor-ganic solids with stringent control of their structural and morpho-logical properties that are required for specific needs in materialsscience. The biomineral arrangements in coral skeletons represent

182 I. Sondi et al. / Journal of Colloid and Interface Science 354 (2011) 181–189
an exceptional window of opportunity for understanding many ofthe phenomena of geological, mineralogical, and paleontologicalinterest, particularly in the biological mediation of crystal growth[18–21].
While the primary structural and morphological characteristicsof scleractinians are well documented [18,22,23], the architectureof complex aragonite-based skeletal elements, formed throughbiologically mediated processes, still need more advancedresearch, in particular, taking into account that they have beenshown to be species-dependent [24]. Recently, several importantstudies [22,24–26] reported and discussed fine-scale growth andbiochemical control over the bio-assisted formation of aragonitein scleractinians. These studies follow on from new strategies inthe formation of unusual bio-inorganic structures based on anon-classical crystallization mechanism involving aggregationprocesses [12,27,28], oriented aggregation assembly [29–31], thedevelopment of an oriented architecture of bridged nanocrystalsembedded in organic polymers [32] and the recently developedmesocrystal concept [33,34]. Following these approaches, thisstudy was directed towards a better understanding of theprinciples of the complex biomineralization processes of fibrousaragonite formation in skeletons of a slow-growing colonial zoo-xanthellate scleractinian Mediterranean coral, Cladocora caespitosa.Specifically, it describes the unusual structural and morphologicalcharacteristics of the biomineral structures, revises and comple-ments the insight into the submicrometer scale colloid-chemicalgrowth mechanisms and morphological changes of the sophisticat-edly designed bio-inorganic architecture of fibrous aragonite. Inaddition, this study has accomplished a preliminary biochemicaland molecular identification of the soluble protein componentsof an organic matrix extracted from the exoskeleton of the coralC. caespitosa. It sheds light on the formation and submicrometerscale characteristics of biogenic aragonite in the skeleton of scle-ractinian corals.
2. Materials and methods
2.1. Materials
The C. caespitosa bank in Veliko jezero, on the island of Mljet,was chosen for its significant size of approximately 650 m2 andrepresents one of the largest banks found in the MediterraneanSea [35]. The sample of C. caespitosa (Linnaeus, 1767) was sampledby scuba diving in Veliko Jezero in May, 2008. Corallites up to 1 cmlong and 3–4 mm wide were selected from the colony (Fig. 1),placed in clean plastic box and stored at �80 �C until further use.
Fig. 1. Living colony of coral Cladocora caespitosa showing typical phaceloid form (A) andare pointing at the section of the native calyx surface (a) and the fractured section of co
2.2. Preparation of coral samples
In order to remove the fleshy tissues and debris, the coralliteswere stirred vigorously in 1 M NaOH for 1 h and subsequently in1% NaClO solution for 2 h. The treated sample was washed in dis-tilled water until the corallites were absolutely clean.
2.3. Extraction and analysis of the proteins from the soluble organicmatrix
Approximately 10 g of the cleaned corallites were ground to afine powder and demineralized according to a slightly modifiedprocedure described by Compère and co-workers [36]. Specifically,the powder was demineralized with 300 mL of 0.1 M HCl in thepresence of the Protease Inhibitor Cocktail (Sigma). The pH valuewas monitored during the dissolving process, keeping it betweenpH 3 and 4 by the slow addition of 1 M HCl into the reacting sys-tem. After complete dissolution, a solution containing a solubleprotein organic matrix was placed in dialyzing tubes and dialyzedin 2 L of 10 mM Tris buffer (pH 8) at 4 �C for 4 days with eight buf-fer exchanges. In order to remove the undissolved particles andremaining debris, the obtained dispersion was centrifuged at15,000 rpm for 30 min. The obtained dispersion was freeze-driedand a soluble organic matrix was obtained. The resulting solidswere dissolved into 2–3 mL of the buffer (10 mM Tris, pH 8.0)and additionally desalted by using a PD10 column (Pharmacia). Fi-nally, the concentration of proteins was determined according tothe method described by Bradford [37].
2.4. Characterization of the materials
The morphology of the bio-inorganic structures of the corallitewas examined by field-emission scanning electron microscopy(FESEM, Zeiss Supra 35 VP, Germany). The cleaned corallite sam-ples were carbon coated using a PECS (Gatan, Model 682, Ger-many) in order to ensure good conductivity during the FESEMinvestigation.
The surface topography of the corallite was determined using aMultiMode Probe Microscope with a Nanoscope IIIa controller anda ‘‘J’’ scanner with a vertical engagement (JV) 125 lm (AS-130 Vscanner, Veeco Instruments, Santa Barbara, CA). Imaging was con-ducted in the ‘‘contact mode’’ using a silicon tip (NP, Nom. freq.18 kHz, Nom. spring constant of 0.06 N/m) as well as the ’’tappingmode’’ using a silicon tip (TESP, Veeco, Nom. Freq. 320 kHz, Nom.spring constant of 42 N/m) under ambient conditions in air. Thelinear scanning rate was optimized between 1.0 and 2.0 Hz at a
single corallite from the same colony with top view morphology of calyx (B). Arrowsrallite (b) that was subjected to the microscopy examinations.
I. Sondi et al. / Journal of Colloid and Interface Science 354 (2011) 181–189 183
scan angle of 0�. Atomic-force microscopy (AFM) analyses wereperformed on the surface of the cleaned corallite. Prior to theAFM analysis the surface of the samples was additionally dried ina stream of nitrogen and attached to a metal disc. Any additionaltreatment was avoided due to possible changes taking place be-tween the mineral and organic components.
The mineral composition of the sampled materials was ana-lyzed by X-ray powder diffraction (XRD), using a diffractometerwith Cu Ka radiation and a Sol-X energy-dispersive detector (D4Endeavor, Bruker AXS, Karlsruhe, Germany). The angular range2H was from 10� to 70� with a step size of 0.02� and a collectiontime of 5 s. The obtained XRD diffraction patterns were identifiedaccording to the JCPDS powder-diffraction files. The Fourier Trans-form Infrared (FTIR) spectra were recorded at 20 �C using a Perkin-Elmer spectrometer (Model 2000, USA). The specimens werepressed into small discs using a spectroscopically pure KBr matrix.The spectra were recorded using a KBr beam splitter in the mid IRregion (4000–400 cm�1).
An estimation of the organic matter content in the coral wasdetermined by thermo-gravimetric analysis (TGA) and differentialscanning calorimetry (DSC) on a Netzsch Jupiter 449 simultaneousthermal analysis instrument coupled with a mass spectrometer(MS) (Netzsch QMS 403 C Aëolos quadrupole). The analysis wasperformed in air from 40 to 1000 �C with a heating rate of 10 �C/min using an Al2O3 crucible with a lid. The evolution of H2O andCO2 was monitored by m/z fragments of 18, 17 and m/z fragmentsof 44, 28, respectively.
The proteins were analyzed and separated by sodium dodecylsulfate–polyacrylamide gel electrophoresis (SDS–PAGE) [38] andvisualized by Coomassie brilliant blue and silver staining [39].
The gel bands were then washed with sterile redistilled water,following by 100% methanol, and subjected to a standard protocolof mass-spectrometry identification, which involved the washingof gel pieces, reduction, alkylation, and in-gel tryptic digestion.Proteolytic peptides were extracted from the gel and analyzed byMALDI-TOF mass spectrometry (4800 Plus MALDI TOF/TOF Ana-lyzer, USA).
20 30 40 50
Ar
Ar
Ar
Ar
Ar
Ar ArArAr
Ar
Ar
Ar
inte
nsity
/ a.
u.
2θ/ degree
A
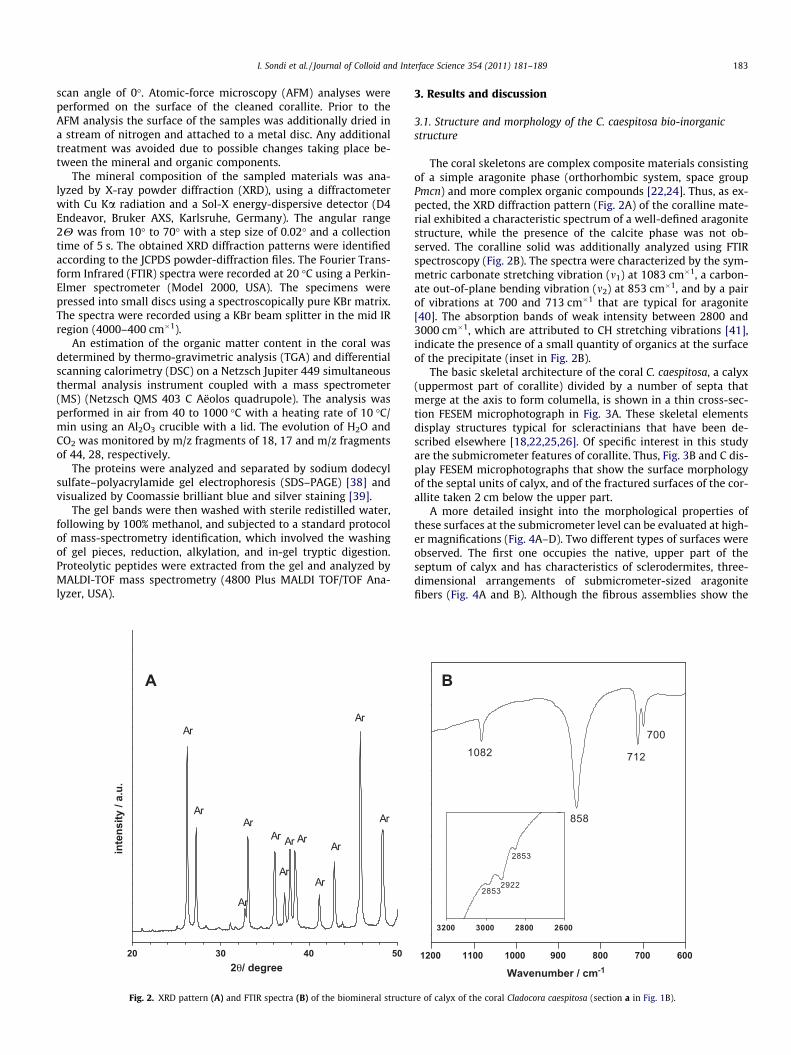
Fig. 2. XRD pattern (A) and FTIR spectra (B) of the biomineral structu
3. Results and discussion
3.1. Structure and morphology of the C. caespitosa bio-inorganicstructure
The coral skeletons are complex composite materials consistingof a simple aragonite phase (orthorhombic system, space groupPmcn) and more complex organic compounds [22,24]. Thus, as ex-pected, the XRD diffraction pattern (Fig. 2A) of the coralline mate-rial exhibited a characteristic spectrum of a well-defined aragonitestructure, while the presence of the calcite phase was not ob-served. The coralline solid was additionally analyzed using FTIRspectroscopy (Fig. 2B). The spectra were characterized by the sym-metric carbonate stretching vibration (m1) at 1083 cm�1, a carbon-ate out-of-plane bending vibration (m2) at 853 cm�1, and by a pairof vibrations at 700 and 713 cm�1 that are typical for aragonite[40]. The absorption bands of weak intensity between 2800 and3000 cm�1, which are attributed to CH stretching vibrations [41],indicate the presence of a small quantity of organics at the surfaceof the precipitate (inset in Fig. 2B).
The basic skeletal architecture of the coral C. caespitosa, a calyx(uppermost part of corallite) divided by a number of septa thatmerge at the axis to form columella, is shown in a thin cross-sec-tion FESEM microphotograph in Fig. 3A. These skeletal elementsdisplay structures typical for scleractinians that have been de-scribed elsewhere [18,22,25,26]. Of specific interest in this studyare the submicrometer features of corallite. Thus, Fig. 3B and C dis-play FESEM microphotographs that show the surface morphologyof the septal units of calyx, and of the fractured surfaces of the cor-allite taken 2 cm below the upper part.
A more detailed insight into the morphological properties ofthese surfaces at the submicrometer level can be evaluated at high-er magnifications (Fig. 4A–D). Two different types of surfaces wereobserved. The first one occupies the native, upper part of theseptum of calyx and has characteristics of sclerodermites, three-dimensional arrangements of submicrometer-sized aragonitefibers (Fig. 4A and B). Although the fibrous assemblies show the
700
712
858
1082
3200 3000 2800 2600
2853
29222853
1200 1100 1000 900 800 700 600
B
Wavenumber / cm-1
re of calyx of the coral Cladocora caespitosa (section a in Fig. 1B).

Fig. 3. FESEM microphotographs of bio-inorganic structures of Cladocora caespitosa:ultra-thin cross-section of the calyx divided by a number of septa (A), three-dimensional morphologies of different levels of corallite: the native surface of theupper part of a septal unit within the calyx (B), (section a in Fig. 1B), and of thefractured septal surface of the lower part of the corallite (C), (section b in Fig. 1B).Inserted high-resolution images in Fig. B and C display the surface morphology ofthe marked sections.
184 I. Sondi et al. / Journal of Colloid and Interface Science 354 (2011) 181–189
crystal structure of aragonite (Fig. 2A) they do not exhibit well-defined crystallographic forms. The apparent size of the fiber unitsis approximately 150 nm in diameter and 2–3 lm in length.According to Constantz [42], the observed narrow and submicrom-eter-sized character of the fibers is characteristics of fast-growingscleractinians. In contrast, the second type of surface that covers
the fractured surface of the lower part of the corallite differs signif-icantly. In comparison with the fibrous structure it is characterizedby micrometer-sized and crystallographically well-defined arago-nite facets that are built up of bundles of thin prismatic needles,all oriented in the same direction (Fig. 4C and D).
Recently, Baronnet and co-workers [43] have shown that high-resolution characterization techniques such as atomic-forcemicroscopy (AFM), even due to difficulties in imaging the porousmorphology, can be very useful for the study of submicrometerbiogenic structures. Accordingly, the investigated structures wereadditionally examined using the high-resolution AFM techniquein order to get a more comprehensive picture of the submicrome-ter-sized hierarchical skeletal architecture. The AFM micrograph ofthe upper part of the septum shows the surface topography of thetopmost and side parts of the elongated aragonite fibers (Fig. 5Aand B). Nearly spherical growths on the AFM image (Fig. 5A) corre-spond to the surface of the upper part of the fibrous units with asize of 100–200 nm that corresponds to the thickness of the fibers.The side AFM image (Fig. 5B) shows that aragonite fibers are builtup of elongated irregular clusters that are incorporated into anirregular bulge structure characterized by high porosity. In con-trast, the surface of the fractured corallite displays crystallograph-ically well-defined forms (Fig. 5C and D).
The described surface morphology is quite similar to otherstudied sclearctinian biomineral structures [19,23,25]. What wasnot previously described and discussed is the fact that the initiallyformed fibrous aragonite in modern corals is subjected to amorphological transformation into micrometer-sized and crystallo-graphically well-defined forms. Notably, significant morphologicalchanges, the transformation of aragonite fibers into a prismatic crys-tal of distinct crystallinity, occurred in the corallite. Recently, Kruzicand Pozar-Domac [35] have determined the skeleton growth rate ofthe coral C. caespitosa, which is estimated to be from 3 to 6 mm peryear. Accordingly, the latest aragonite structure should be 2–3 yearsold, indicating that the morphological transformation of the fibrousstructure into well-defined crystallographic forms occurred in arelatively short period of time. This observation is in accordancewith the previous study, which indicated that the early diageneticchanges in coral skeletons result in morphological changes of sub-micrometer-sized aragonite [19]. These processes are accompaniedby the degradation of organic matrices in exoskeleton structures.
3.2. Growing mechanism of scleractinian revisited
FESEM observations indicate the appearance of two major typesof crystal morphology on the native surface of the septum of calyx,i.e., nanosized spherical crystallites (Fig. 6A) and elongated fibers(Fig. 6D), together with their transitional forms, i.e., elongated coneclusters built up of submicrometer-sized aragonite particles(Fig. 6B and C). This finding, most particularly the structural prop-erties of the transitional morphological forms, offers possibilitiesfor the interpretation of the formation of elongated aragonite fi-bers. The growing patterns of these structures should not be con-sistent with the conventional mechanism, which is primarilybased on diffusion growth. Rather, it supports the recently pro-posed biochemically driven crystallization mechanism, accordingto which scleractinian aragonite is produced through the morecomplex, matrix-mediated growth that includes the simultaneousinvolvement of biotic and abiotic processes [22,24–26]. Recently,Cuif and Dauphin [24] have proposed a ‘‘two-step’’ model ofgrowth for the scleractinian skeletons. This model involves an ini-tial secretion of the organic matrix, of a proteoglycan, and the for-mation of submicrometer-sized granular particles (centers ofcalcification), followed by a ‘‘thickening stage’’. Three-dimensionalarrangements of elongated fibers are formed through an organic

Fig. 4. FESEM microphotographs of fibrous biomineral structures of the upper part of the septum of calyx (A and B) (section a in Fig. 1B) and of bundles of prismatic forms onthe fractured surface of the lower part of corallite (C and D) (section b in Fig. 1B).
Fig. 5. AFM surface topography images presented as a 3D – view of height data obtained from the skeletal nanogranular (A) and fibrous (B) surface regions of the upper partof the septum of calyx (section a in Fig. 1B); 3D view obtained by imaging of a fractured region of the corallite shows the high-density packing of the prismatic aragonitecrystals (C and D) (section b in Fig. 1B).
I. Sondi et al. / Journal of Colloid and Interface Science 354 (2011) 181–189 185
matrix-mediated sequential growth of thick layers consisting ofsubmicrometer crystallites.
Accordingly, the general model for the morphogenesis ofscleractinian could be based on complex organo-mediated

Fig. 6. FESEM microphotographs of different types of morphology on the native surface of a septal unit within the calyx: submicrometer-sized, nearly spherical crystallites(A), transitional forms of elongated cone structures built up of submicrometer-sized particles (B and C), and elongated aragonite fibers (D).
186 I. Sondi et al. / Journal of Colloid and Interface Science 354 (2011) 181–189
precipitation governed by biological activity. It is a highly con-trolled process, from the submicrometer scale to macroscopiclevels, resulting in complex hierarchical architectures and shapes[25,26]. In addition, Holcomb and co-workers [20] in their labora-tory model experiments have shown that the matrix-mediatedgrowth mechanisms of scleractinian corals are accompanied byphysico-chemical conditions driven by cyclic changes in the satura-tion state of the coral’s calcifying fluids. Thus, when the saturationstate is high, granular submicrometer-sized crystals precipitate andform the centers of calcification. As the saturation state decreases,elongated aragonite fiber growth occurs in bundles that radiateout from the centers of calcification. If this hypothesis is true, itwould seem that these two types of crystal morphology, togetherwith their transitional forms, should be present on the same surfaceof the corallite. Indeed, the FESEM microphotographs shown inFig. 6 verify the presence of different morphologies and affirm therole of the saturation state in the formation of aragonite solids.
Following these concepts, the results described herein offer anapproach that complements the recently established growthmechanisms [22,24–26] of scleractinian aragonite. More specifi-cally, this study attempts to explain the formation of a fibrous sub-micrometer-sized aragonite structure through complex colloid-chemical mechanisms that are governed by the biochemical char-acteristics of the proteins involved in these processes.
The formation of hierarchical, submicrometer-sized aragonitefibers occurs in several stages. It is initiated by the organic-medi-ated formation of nearly spherical submicrometer crystallites(Fig. 6A) in localized structures called ‘‘centers of calcification’’.The growth of this structure in association with the organic matrixis significant in understanding the development of biomineral skel-etal elements. Dauphin and co-workers [44] have shown that ara-gonitic nanograins in some scleractinians occur within primaryactive matrix residues of the hydrated proteoglycans. They sug-gested that this organic probably plays a major role in the bio-in-duced formation of aragonite.
Previous laboratory experiments have shown that the initialformation of orthorhombic anhydrous calcium carbonate nano-crystallites, induced by protein activities, is rather common inhighly saturated reacting solutions [45–47]. It was also shown thaton prolonged aging and at a lower saturation state, these solidsgrow into micrometer-sized needle-like clusters through diffusiongrowth [45]. In contrast to these observations, in the C. caespitosaskeleton, the second stage of morphogenesis, in which the initiallyformed nanoparticles convert into fibrous aragonite, is governed byoriented aggregation processes. Indeed, FESEM microphotographsconfirm that the initial formation of nanosized spherulites in cen-ters of calcification (Fig. 6A) is followed by the growth of aggre-gated cone morphologies that are gradually built up of smaller,submicrometer units (Fig. 6B and C). What ensues is the subse-quent morphological conversion and formation of continuous elon-gated fibrous units (Fig. 6D).
This observation implies the significance of the oriented aggre-gation processes in the formation of fibrous structures. This con-cept is much more common in biomineralization processes thanhas been assumed up to now, while the number of reported exam-ples of their significance in biomineral formation has significantlyincreased in recent years [12,45,47,48]. Recent evidence of thenon-classical formation of carbonate solids through orientedaggregation processes [49] and the nanoscale aggregation route[12,47,48], support the mechanism according to which carbonatecolloids can be produced through the aggregation of preformednanosized particles. The obtained submicrometer polycrystallinefibers can be defined as mesocrystals [33,48].
What was not previously well understood is the final stage ofmorphogenesis, in which the initially formed nanoparticlesconvert into fibrous aragonite solids through the organic matrix–nanograin and/or nanograin–nanograin interactions. AFM investi-gations, particularly AFM phase imaging, offers a better insight intothe complex structure and composition of coral aragonite on thenanoscale [24–26]. Thus, the AFM phase-imaging micrograph
0 200 400 600 800 100050
60
70
80
90
100
DSC
/ a.
u.
Wei
ght /
%
Temperature / ºC
Weight loss 2.5 %
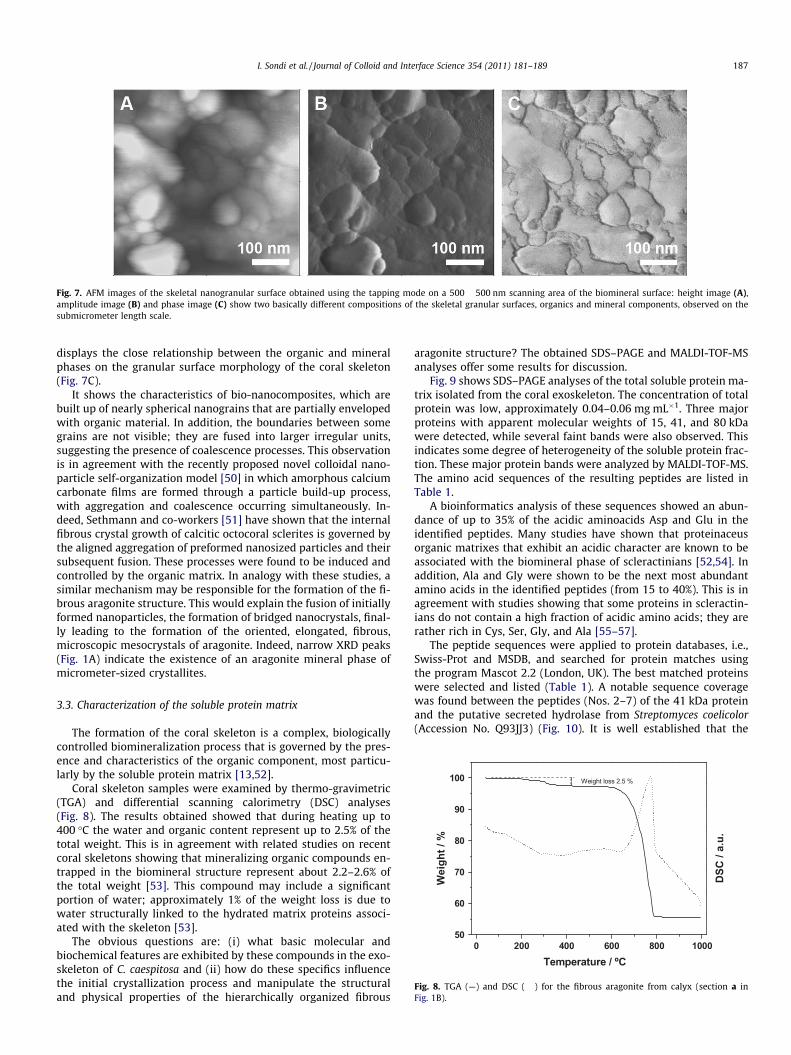
Fig. 8. TGA (—) and DSC (����) for the fibrous aragonite from calyx (section a inFig. 1B).
Fig. 7. AFM images of the skeletal nanogranular surface obtained using the tapping mode on a 500 � 500 nm scanning area of the biomineral surface: height image (A),amplitude image (B) and phase image (C) show two basically different compositions of the skeletal granular surfaces, organics and mineral components, observed on thesubmicrometer length scale.
I. Sondi et al. / Journal of Colloid and Interface Science 354 (2011) 181–189 187
displays the close relationship between the organic and mineralphases on the granular surface morphology of the coral skeleton(Fig. 7C).
It shows the characteristics of bio-nanocomposites, which arebuilt up of nearly spherical nanograins that are partially envelopedwith organic material. In addition, the boundaries between somegrains are not visible; they are fused into larger irregular units,suggesting the presence of coalescence processes. This observationis in agreement with the recently proposed novel colloidal nano-particle self-organization model [50] in which amorphous calciumcarbonate films are formed through a particle build-up process,with aggregation and coalescence occurring simultaneously. In-deed, Sethmann and co-workers [51] have shown that the internalfibrous crystal growth of calcitic octocoral sclerites is governed bythe aligned aggregation of preformed nanosized particles and theirsubsequent fusion. These processes were found to be induced andcontrolled by the organic matrix. In analogy with these studies, asimilar mechanism may be responsible for the formation of the fi-brous aragonite structure. This would explain the fusion of initiallyformed nanoparticles, the formation of bridged nanocrystals, final-ly leading to the formation of the oriented, elongated, fibrous,microscopic mesocrystals of aragonite. Indeed, narrow XRD peaks(Fig. 1A) indicate the existence of an aragonite mineral phase ofmicrometer-sized crystallites.
3.3. Characterization of the soluble protein matrix
The formation of the coral skeleton is a complex, biologicallycontrolled biomineralization process that is governed by the pres-ence and characteristics of the organic component, most particu-larly by the soluble protein matrix [13,52].
Coral skeleton samples were examined by thermo-gravimetric(TGA) and differential scanning calorimetry (DSC) analyses(Fig. 8). The results obtained showed that during heating up to400 �C the water and organic content represent up to 2.5% of thetotal weight. This is in agreement with related studies on recentcoral skeletons showing that mineralizing organic compounds en-trapped in the biomineral structure represent about 2.2–2.6% ofthe total weight [53]. This compound may include a significantportion of water; approximately 1% of the weight loss is due towater structurally linked to the hydrated matrix proteins associ-ated with the skeleton [53].
The obvious questions are: (i) what basic molecular andbiochemical features are exhibited by these compounds in the exo-skeleton of C. caespitosa and (ii) how do these specifics influencethe initial crystallization process and manipulate the structuraland physical properties of the hierarchically organized fibrous
aragonite structure? The obtained SDS–PAGE and MALDI-TOF-MSanalyses offer some results for discussion.
Fig. 9 shows SDS–PAGE analyses of the total soluble protein ma-trix isolated from the coral exoskeleton. The concentration of totalprotein was low, approximately 0.04–0.06 mg mL�1. Three majorproteins with apparent molecular weights of 15, 41, and 80 kDawere detected, while several faint bands were also observed. Thisindicates some degree of heterogeneity of the soluble protein frac-tion. These major protein bands were analyzed by MALDI-TOF-MS.The amino acid sequences of the resulting peptides are listed inTable 1.
A bioinformatics analysis of these sequences showed an abun-dance of up to 35% of the acidic aminoacids Asp and Glu in theidentified peptides. Many studies have shown that proteinaceusorganic matrixes that exhibit an acidic character are known to beassociated with the biomineral phase of scleractinians [52,54]. Inaddition, Ala and Gly were shown to be the next most abundantamino acids in the identified peptides (from 15 to 40%). This is inagreement with studies showing that some proteins in scleractin-ians do not contain a high fraction of acidic amino acids; they arerather rich in Cys, Ser, Gly, and Ala [55–57].
The peptide sequences were applied to protein databases, i.e.,Swiss-Prot and MSDB, and searched for protein matches usingthe program Mascot 2.2 (London, UK). The best matched proteinswere selected and listed (Table 1). A notable sequence coveragewas found between the peptides (Nos. 2–7) of the 41 kDa proteinand the putative secreted hydrolase from Streptomyces coelicolor(Accession No. Q93JJ3) (Fig. 10). It is well established that the

kDa
97
66
45
30
20.1
14.4
Fig. 9. SDS–PAGE analyses of soluble protein matrix: line 1, protein marker; line 2,coral proteins. The major protein bands at 15, 41, and 80 kDa are marked. The gelwas stained by CBB.
188 I. Sondi et al. / Journal of Colloid and Interface Science 354 (2011) 181–189
scleractinian corals calcify extra-cellularly [58]. It can be specu-lated that the secretion of a proteinaceus organic matrix, such asthe secreted acidic hydrolases, creates the crystallization environ-ment. Thus, the initial formation of the nanosized aragonite and itsgrowth into fibrous structures are governed by the specificchemical activity of the secreted protein matrix.
Previous laboratory model experiments have shown that hydro-lyses such as ureases (urea aminohydrolase; EC 3.5.1.5.) signifi-
Table 1Internal amino acid sequences of soluble proteins isolated from the coral Cladocora caespitthe protein databases Swis-Prot.
Protein (kDa) Peptide Nos. Amino acid sequences
80 1 DGASAQYGSDAIAGVVNIILK2 ILDKNLNLNNLIPK3 EAAGVRLKLANSGLPK
41 1 ADLDVIMVAPK2 DFTAAEIAR3 VPTLEQYVHR4 LVVQSFDADSIR5 GASGYAPENTLAAVDR6 TTDVEEVFPDRSPWK7 VAGYDVDGIITNKPDVVR
15 1 DLANGPSVR2 TSADIDAVAIGHDLAR3 NAWTDGHTNFFTVWANR4 QLATNASGSLAVGDPVVVQGR
MHVRAVAVTTTALLGVALTAPLSHARADEAGDDGPTVVTRDGELVVLHDESLARTTDVEEVFPDRSPWKVKDFTAAKLLLELKSPGLYPGIEQQTLKVLANEGWLDRRHVAGRLVAEFADQINPSYGSLSMSYVSAVHAFTGPHGRPLEVLTWT
Fig. 10. Sequence coverage (27.7%) of putative secreted hydrolase with pepti
cantly influence crystal-phase formation and the development ofunusual morphologies of calcium carbonate polymorphs [12]. Spe-cifically, ureases in reacting aqueous solutions of calcium salts in-duce the formation of a nanosized phase, which with a prolongedreaction time aggregates into submicrometer-sized solids [46,47].The initial formation of nanosized particles may be the result ofthe strong interaction between Ca2+ and the acidic amino acidsAsp and Glu of protein macromolecules, forming Ca2+/Asp orCa2+/Glu multi-carboxyl chelate complexes that act as organic tem-plates and induce the initial formation of the nanosized phase[12,16].
A similar mechanism may be responsible for the biologically in-duced formation of submicrometer-sized aragonite in a secretedprotein matrix. This would explain the initial formation on nano-sized particles and their growth into fibrous structures by subse-quent aggregation and coalescence processes.
Peptide No. 2 identified in a protein of 80 kDa (Table 1) showed a5.7% coverage of a sequence with matched carbonic anhydrase fromSulfurilhydrogenibium sp. YO3AOP1 (Accession No. YP_001930770).Several studies have demonstrated the involvement of this enzymein coral calcification, although its role is not entirely understood.Recently, it was shown that carbonic anhydrases are not only struc-tural proteins; they also play an important catalytic role in eliminat-ing the kinetic barrier in the conversion of inorganic carbon at thecalcification sites [59,60]. The latter peptide sequence was alignedwith two complete carbonic anhydrase sequences from the coralStylophora pistillata [61] (GenBank: ACE95141.1 and ACA53457.1).The obtained identity scores of 20 and 28%, respectively, were oneof the non-conserved regions of carbonic anhydrase. There are atleast five classes of carbonic anhydrase described in the literature[62]. Except for several conserved regions, they share very little se-quence or structural similarity. Thus, despite a similar catalyticfunction, they exert different molecular structures that seem to bespecies-dependent.
As a final remark, it needs to be pointed out that the role of theacidic proteinaceus organic matrix in the formation of the aragonitebiomineral structure in scleractinian is far from being completelyunderstood. However, this study revises and complements the in-sight into the nanoscale growth mechanisms and morphologicalchanges of the sophisticatedly designed bio-inorganic architecture
osa exoskeleton, and protein matches found by using the program Mascot 2.2 against
Match to protein (Accession Nos.)
TonB-dependent outer membrane receptor (YP_190844, ZP_00652553)Carbonic anhydrase (YP_001930770)Flagelar M-ring protein FliF (YP_002460594)
Ketol-acid reductoisomerase (AAD32178, NP_947380)Putative secreted hydrolase (Q93JJ3)Putative secreted hydrolase (Q93JJ3)Putative secreted hydrolase (Q93JJ3)Putative secreted hydrolase (Q93JJ3)Putative secreted hydrolase (Q93JJ3)Putative secreted hydrolase (Q93JJ3)
Single stranded DNA binding protein (Q9KYI9)Single stranded DNA binding protein (Q9KYI9)Single stranded DNA binding protein (Q9KYI9)Single stranded DNA binding protein (Q9KYI9)
AHRGASGYAPENTLAAVDRAAELGIRWVENDVQREIARLDAGSWFGSEYAGARVPTLEQYVHRVEHNHQ
VQSFDADSIRTVHDLKPAVKTGFLGTPAVSELPAYVNDADTARRVAGYDVDGIITNKPDVVRDAVD
des 2–7 identified from 41 kDa protein of Cladocora caespitosa (Table 1).

I. Sondi et al. / Journal of Colloid and Interface Science 354 (2011) 181–189 189
of fibrous aragonite in C. caespitosa, and provides a new insight intothe molecular diversity of the proteinaceus organic matrix ofscleractinians.
4. Conclusions
This study complements existing hypotheses on the critical roleof organic compounds and biologically induced colloid-chemicalprocesses at the submicrometer scale in the phase formation,growth and morphological transformation of fibrous aragonite inC. caespitosa.
Molecular and biochemical characterizations of the proteinac-eus organic matrix, isolated from a coral exoskeleton, indicatethe significant presence of acidic hydrolase proteins. The organicmatrix-mediated nucleation and nanograin–nanograin interac-tions, occurring via the aggregation and subsequent coalescenceprocesses, were found to be the main strategy in the formationof hierarchically structured, submicrometer-sized, aragonite fibersin scleractinians.
The results of this study should be considered from the perspec-tive that the molecular and biochemical characteristics of the skel-etal acidic proteinaceus organic matrix and the matrix-mediatedcolloid-chemical processes in the formation of the aragonite struc-ture in different scleractinians may be species-specific properties.
Acknowledgments
This work was supported by the Administration of the Mljet Na-tional Park and by the Ministry of Science, Education and Sports ofthe Republic of Croatia (Grants Nos. 098-0982934-2742, 098-0982913-2829, and 098-1191344-2938, and 098-0982934-2744)and by the Ministry of Higher Education, Science and Technologyof the Republic of Slovenia (Grant P2-0091-0106). The authorsacknowledge the help of Dr. Saša Kazazic with the MALDI-TOF-MS analyses. The comments of the two anonymous reviewers arealso gratefully acknowledged.
References
[1] M.E. Tucker, V.P. Wright, Carbonate Sedimentology, Blackwell ScientificPublications, Oxford, 1990.
[2] G. Falini, S. Albeck, S. Weiner, L. Addadi, Science 271 (1996) 67.[3] H. Tohse, K. Saruwatari, T. Kogure, H. Nagasawa, Y. Takagi, Cryst. Growth Des. 9
(2009) 4897.[4] R.A. Metzler, J.S. Evans, C.E. Killian, D. Zhou, T.H. Churchill, N.P. Appathurai, S.N.
Coppersmith, P.U. Gilbert, J. Am. Chem. Soc. 132 (2010) 6329.[5] A.E. Tanur, N. Gunari, R.M. Sullan, C.J. Kavanagh, G.C. Walker, J. Struct. Biol. 169
(2010) 145.[6] Y. Dauphin, Zoology 109 (2006) 85.[7] S. Mann, Nature 365 (1993) 499.[8] F.C. Meldrum, H. Cölfen, Chem. Rev. 108 (2008) 4332.[9] S. Weiner, J. Struct. Biol. 163 (2008) 229.
[10] A.H. Knoll, Rev. Mineral. Geochem. 54 (2003) 329.[11] J. Aizenberg, S. Albeck, S. Weiner, L. Addadi, J. Cryst. Growth 142 (1994) 156.[12] I. Sondi, B. Salopek-Sondi, Langmuir 21 (2005) 8876.
[13] S. Tambutté, E. Tambutté, D. Zoccola, D. Allemand, in: E. Bäuerlein (Ed.),Handbook of Biomineralization: Biological Aspects and Structure Formation,Wiley-VCH, Weinheim, 2007, p. 243.
[14] Y. Dauphin, Int. J. Biol. Macromol. 28 (2001) 293.[15] H. Nagasawa, Thalassas 20 (2004) 15.[16] H. Tong, W.T. Ma, L.L. Wang, P. Wan, J.M. Hu, L.X. Cao, Biomaterials 25 (2004)
3923.[17] D. Volkmer, M. Fricke, T. Huber, N. Sewald, Chem. Commun. 16 (2004) 1872.[18] J.E. Sorauf, Palaeontology 15 (1972) 88.[19] C. Perrin, D.C. Smith, J. Sediment. Res. 77 (2007) 495.[20] M. Holcomb, A.L. Cohen, R.I. Gabitov, J.L. Hutter, Geochim. Cosmochim. Acta 73
(2009) 4166.[21] J. Stolarski, A. Meibom, R. Przenioslo, M. Mazur, Science 318 (2007) 92.[22] R. Przenioslo, J. Stolarski, M. Mazur, M. Brunelli, J. Struct. Biol. 161 (2008) 74.[23] J. Stolarski, Acta Palaeont. Pol. 48 (2003) 497.[24] J.-P. Cuif, Y. Dauphin, J. Struct. Biol. 150 (2005) 319.[25] J.-P. Cuif, Y. Dauphin, Biogeosci. Discuss. 1 (2004) 625.[26] J.-P. Cuif, Y. Dauphin, A. Meibom, C. Rollion-Bard, E. Salomé, J. Susini, C.T.
Williams, in: W.E.N. Austin, R.H. James (Eds.), Biogeochemical Controls onPalaeoceanographic Environmental Proxies, Geological Society, London, 2008,p. 87.
[27] V. Privman, D.V. Goia, J. Park, E. Matijevic, J. Colloid Interface Sci. 213 (1999)36.
[28] J.K. Bailey, C.J. Brinker, M.L. Mecartney, J. Colloid Interface Sci. 157 (1993) 1.[29] A.P. Alivisatos, Science 289 (2000) 736.[30] J.F. Banfield, S.A. Welch, H. Zhang, T.T. Ebert, R.L. Penn, Science 289 (2000) 751.[31] R.L. Penn, J.F. Banfield, Geochim. Cosmochim. Acta 63 (1999) 1549.[32] Y. Oaki, A. Kotachi, T. Miura, H. Imai, Adv. Mater. 16 (2006) 1633.[33] H. Cölfen, M. Antonietti, Angew. Chem., Int. Ed. 44 (2005) 5576.[34] R.Q. Song, H. Cölfen, Adv. Mater. 22 (2010) 1301.[35] P. Kruzic, A. Pozar-Domac, Period. Biol. 104 (2002) 123.[36] P. Compère, M.F. Jaspar-Versali, G. Goffinet, Biol. Bull. 202 (2002) 61.[37] M.M. Bradford, Anal. Biochem. 72 (1976) 248.[38] U.K. Laemmli, Nature 227 (1970) 680.[39] E. Hempelmann, R. Kaminsky, Electrophoresis 7 (1986) 481.[40] K.-S. Seo, C. Han, J.-H. Wee, J.-K. Park, J.-W. Ahn, J. Cryst. Growth 276 (2005)
680.[41] N.V. Vagenas, A. Gatsouli, C.G. Kontoyannis, Talanta 59 (2003) 831.[42] B.R. Constantz, Palaios 1 (1986) 152.[43] A. Barronet, J.-P. Cuif, Y. Dauphin, B. Farre, J. Nouet, Mineral. Mag. 72 (2008)
617.[44] Y. Dauphin, J.-P. Cuif, P. Massard, Chem. Geol. 231 (2006) 26.[45] I. Sondi, E. Matijevic, Chem. Mater. 15 (2003) 1322.[46] S.D. Škapin, I. Sondi, Cryst. Growth Des. 5 (2005) 1933.[47] I. Sondi, S.D. Škapin, B. Salopek-Sondi, Cryst. Growth Des. 8 (2008) 435.[48] S.D. Škapin, I. Sondi, J. Colloid Interface Sci. 347 (2010) 221.[49] D. Vielzeuf, J. Garrabou, A. Barronet, O. Grauby, C. Marschal, Am. Mineral. 93
(2008) 1799.[50] C. Zhong, C.C. Chu, Langmuir 25 (2009) 3045.[51] I. Sethmann, U. Helbig, G. Worheide, Crystengcomm 9 (2007) 1262.[52] B. Constantz, S. Weiner, J. Exp. Zool. 248 (1988) 253.[53] J.P. Cuif, Y. Dauphin, P. Berthet, J. Jegoudez, Geochem. Geophys. Geosyst. 5
(2004).[54] S. Puverel, E. Tambutté, L. Pereira-Mouriès, D. Zoccola, D. Allemand, S.
Tambutté, Comp. Biochem. Phys., B 141 (2005) 480.[55] S.D. Young, Comp. Biochem. Physiol., B 40 (1971) 113.[56] R.M. Mitterer, Bull. Mar. Sci. 28 (1978) 173.[57] T. Watanabe, I. Fukuda, K. China, Y. Isa, Comp. Biochem. Physiol., B 136 (2003)
767.[58] I.S. Johnston, Int. Rev. Cytol. 67 (1980) 171.[59] S. Tambutté, E. Tambutté, D. Zaccola, N. Caminiti, S. Lotto, A. Moya, D.
Allemand, J. Adkins, Mar. Biol. 151 (2007) 71.[60] A. Moya, S. Tambutté, A. Bertucci, E. Tambutté, S. Lotto, D. Vullo, C.T. Supuran,
D. Allemand, D.J. Zoccola, Biol. Chem. 283 (2008) 25475.[61] M.L. de Boer, D.A. Krupp, V.M. Weis, Biol. Bull. 211 (2006) 18.[62] T.W. Lane, M.A. Saito, G.N. George, I.J. Pickering, R.C. Prince, F.M.M. Morel,
Nature 435 (2005) 42.



























