Embed Size (px)
Citation preview

Ecology of a corallivorous gastropod, Coralliophila abbreviata,on two scleractinian hosts. II. Feeding, respiration and growth
Received: 13 July 2001 / Accepted: 15 February 2003 / Published online: 28 March 2003� Springer-Verlag 2003
Abstract Coralliophila abbreviata is a corallivorousgastropod that has been observed to cause largefeeding scars on reef-building corals on Floridian andCaribbean reefs. We detected differences in the popu-lation structure (length-frequency distribution and sexratios) of C. abbreviata populations living on two coralhost taxa, Acropora palmata and Montastraea spp., inthe Florida Keys in a previous study. We hypothesizethat diet (host) has a major influence on snail popu-lation structure and, thus, we characterize metabolism,feeding and growth for snails residing on these coraltaxa. Here, we present results of a reciprocal trans-plant experiment demonstrating that the taxon of thehost influences snail growth rates, as indicated bychanges in shell and body tissue weight. Regardless ofthe host from which they were drawn, snails residenton A. palmata grew faster than those resident onMontastraea spp. Thus, diet influences snail populationstructure. However, the tissue of Montastraea spp.provides more N and C per area of tissue than that ofA. palmata. Respiration rates and tissue compositionof snails collected from the two host taxa did notdiffer. Therefore, snails feeding on Montastraea spp.should have to consume less tissue per day to satisfytheir metabolic requirements compared to snails
feeding on A. palmata. Feeding rates for snails onA. palmata were measured in the laboratory over 48 h(1–9 cm2 coral tissue snail)1 day)1) and estimated fromfeeding scars observed in the field (weekly mean rate of1.07 cm2 coral tissue snail)1 day)1). The lack of defi-nition of snail feeding scars on Montastraea spp.required the calculation of coral tissue consumptionrates based on estimated minimum carbon require-ments. Calculated feeding rates for C. abbreviata were0.13–0.88 cm2 coral tissue day)1 snail)1, when feedingon Montastraea spp., and 0.44–3.28 cm2 coral tissueday)1 snail)1, when feeding on A. palmata. The cal-culated range for the latter is consistent with measuredrates. Thus, C. abbreviata exhibits high variationin growth parameters in response to environmentalvariability and/or food source. At mean levels ofsnail density on reefs off Key Largo, Fla., 20% ofA. palmata colonies lose between 1.32 and 9.84 cm2
tissue day)1, while 50% of Montastraea spp. colonieslose between 1.04 and 7.04 cm2 tissue day)1. Togetherwith published coral tissue regeneration rates, theseresults suggest that if sustained, such rates of preda-tion could have a serious effect on the viability of thesecoral populations on Florida’s reefs.
Introduction
Coral populations are in decline both locally on coralreefs within the Floridian reef tract and globally dueto a number of causes, particularly coral bleachingand storm damage (Wilkinson 2000). The FloridaKeys National Marine Sanctuary (FKNMS) ScienceAdvisory Panel has called for increased attention toidentify all causes of coral decline and for the evalu-ation and implementation of management strategies tocombat this decline (http://www.fknms.nos.noaa.gov./research_monitoring/sap2000.html). The impact ofcorallivores is one of the potential threats that, while
Marine Biology (2003) 142: 1093–1101DOI 10.1007/s00227-003-1053-4
I. B. Baums Æ M. W. Miller Æ A. M. Szmant
Communicated by P.W. Sammarco, Chauvin
I. B. Baums (&) Æ A. M. SzmantRosenstiel School of Marine and Atmospheric Science-MarineBiology and Fisheries, University of Miami,4600 Rickenbacker Causeway, Miami, FL 33149, USA
E-mail: [email protected]
M. W. MillerNOAA-Fisheries, Southeast Science Center,75 Virginia Beach Drive, Miami, FL 33149, USA
Present address: A. M. SzmantCenter for Marine Science,University of North Carolina at Wilmington,5600 Marvin K. Moss Lane,Wilmington, NC 28409, USA

recognized, had been poorly studied and quantified inthe Florida Keys. Coralliophila abbreviata is known toprey on a number of coral species, and their poten-tially different effects on various coral host taxa are inneed of evaluation.
Corallivores can have differential effects on preyspecies by either: (1) preferring one prey species (i.e.consuming more tissue) or (2) affecting prey fitness dif-ferently (i.e. different coral species might have differen-tial abilities to cope with the same injury and tissue loss).Coral species differ in their ability to repair lesions bothin the mechanism used and the speed observed (Bak andSteward-Van Es 1980; Meesters et al. 1992), e.g. Acro-pora palmata has faster growth and regeneration rates incomparison to Montastraea spp. (Meesters et al. 1993,1995). To date, information on both Caribbean coralli-vore preference and their prey’s response to predation islimited.
Distinct differences have been reported in the popu-lation structure (length-frequency distribution and sexratios) of the corallivorous snail, C. abbreviata, living ontwo of the most ecologically important coral species inthe Florida Keys: the arborescent, branching A. palmataand the massive M. annularis species complex. Mon-tastraea spp. colonies harbored smaller snails than A.palmata, and the sex ratios of C. abbreviata populationswere skewed toward males on A. palmata, but wereevenly distributed on Montastraea spp. (Baums et al.2003).
The objectives of the current study were: (1) to in-vestigate aspects of the snail predator/coral host dy-namics, such as snail metabolism, that could account forobserved host-specific differences in snail populationcharacteristics and (2) to quantify rates of coral tissueconsumption by C. abbreviata for the two host coraltaxa, A. palmata and Montastraea spp.
To address the first objective, we hypothesized thatdifferences in snail population characteristics could bethe result of differences in snail growth rate. In turn,variation in snail respiration, diet quality, and/or tissuecomposition could provide mechanisms for host-spe-cific growth rates. Differences in snail tissue composi-tion (i.e. C and N content) might indicate differences inconversion efficiency of the two food types. Dietquality of coral tissue was estimated by C and Ncontent per area.
To address the second objective we measured feedingrates on A. palmata in the field and in the laboratory.Multiple attempts to measure snail feeding rates onMontastraea spp. by direct observation failed, due to thesmall, poorly defined feeding scars on this coral.Therefore, we calculated a carbon budget (based on snailand diet parameters measured for objective 1) to esti-mate the amount of coral tissue necessary to sustain anindividual snail feeding at the minimum consumptionrate on Montastraea spp. A similar carbon budget forsnails feeding on A. palmata was calculated for com-parison.
Materials and methods
Snail growth rates
Reciprocal transplant experiment
A reciprocal transplant experiment was performed to test thehypothesis that observed differences in snail size distributionsbetween the two host coral species (Baums et al. 2003) resultedfrom host-specific snail growth rates. Snails between 15 and30 mm in length were collected 15)16 June 1999 at Little Grecianreef (25�07¢N; 80�18¢W) from the two host taxa [n=100 eachfrom Montastraea spp. (including M. annularis sensu strictu, M.faveolata and M. franksi; Weil and Knowlton 1994) and Acroporapalmata]. Shells were cleaned and a small numbered tag(6·3.5 mm, shellfish tag, Floy Tag, Seattle, Wash.), color codedfor its original host, was attached with Krazyglue (Elmers Prod-ucts, Columbus, Ohio).
After recording length and weight (see below), the tagged snailswere returned to Little Grecian reef and placed in a 2·2 factorialdesign (n=50). A total of 50 tagged snails from each host were ran-domly allocated to experimental colonies of their source host species,and the remaining 50 were transplanted to colonies of the alternatecoral host (reciprocal ‘‘feeding’’ host). Experimental colonies wereselected that already harbored snails. These naturally occurring C.abbreviata were removed and replaced with an equal number oftagged snails from each source host (a total of 12–20 snails perMontastraea spp. head, 4–12 snails per A. palmata colony) depend-ing on the size of the colony. At least 50 m separated experimentalMontastraea spp. (n=5) and A. palmata (n=9) colonies.
Snails were retrieved after 3 months, scrubbed, re-measuredand re-weighed. Two SCUBA divers searched on two consecutivedays to increase retrieval efficiency. Shell and body growth werecalculated (see below). Two-way ANOVA was used to detect sig-nificant effects of ‘‘source host’’, ‘‘feeding host’’ and their interac-tion on snail growth. Three growth parameters, change in bodyweight (g), change in shell weight (g) and change in shell length(mm) were analyzed in separate ANOVAs.
Calculation of shell and body weight
Shell and tissue density were found to be constant based on mea-surements of ten sacrificed snails from each host (Baums 2000).This allowed the calculation of shell and body weight of the ex-perimental snails from two non-destructive measurements (Palmer1982; Baums 2000): (1) weight of the whole animal in air (airweight) and (2) weight of the animal immersed in seawater (im-mersed weight).
Mean values for shell density did not differ between snails fromthe two coral hosts, and, thus, data were pooled for a sample size of20, yielding 2.78±0.16 g cm)3 (mean±SD), essentially the densityof pure calcium carbonate. Average immersed weight of body tissuewas zero (±0.01), so body density was set equal to that of seawaterfor all subsequent calculations. The density of seawater was cal-culated from salinity measured with a refractometer and watertemperature (Millero et al. 1980; Fofonoff and Millard 1983). Themass of the shell (MS) can be calculated as:
MS¼Wwim=½ð1� qwÞ=qSÞ� ð1Þwhere Ww im is the immersed weight of the whole animal, qS is shelldensity (2.78 g cm)3 as determined earlier) and qw is the density ofthe seawater.
The mass of the body tissue (MB) alone can be calculated as:
MB¼Mw �MS ð2Þwhere Mw is the mass of the whole animal determined by weighingin air. Calculated shell and body weights were compared withmeasured values for the 20 sacrificed snails. A linear regression of
1094

measured shell and body weight against calculated shell and bodyweight yielded r2 values of 0.998 and 0.956, respectively (P<0.001)(Baums 2000).
Host and snail tissue composition
Subsamples (1 cm diameter) of formalin-fixed, decalcified tissuesfrom eight A. palmata fragments (collected after HurricaneGeorges, August 1998) were dried, ground and analyzed in dupli-cate with a Carbo Erba model 1108 elemental analyzer against anacetanilide standard (10.36% N, 71.09% C). Carbon and nitrogencontent of the tissue was expressed as micrograms atomic (lg-at) Cand lg-at N cm)2 of coral tissue. Due to permitting constraints onthe collection of live coral tissue, values for Montastraea spp. froma previous study (Szmant et al. 1990) utilizing the same method andinstrument were used for comparison.
Snails were collected from both host taxa at Horseshoe(25�08¢N; 80�18¢W), Pickles (24�59¢N; 80�24¢W) and Little Grecianreefs (n=118). After measuring oxygen consumption (see below),soft tissue was removed from the shell, oven dried and analyzed forelemental composition as described above. Average values werereported as lg-at C and lg-at N per snail. N and C content persnail were compared between host groups using non-parametricrank sum tests.
Respirometry
Freshly collected snails were scrubbed to remove all epibionts andtransferred to individual volume-calibrated vials filled with sea-water. At least two vials of seawater without snails were used ineach incubation series of 5–15 snails to control for changes in O2
not associated with snail respiration. Initial oxygen measurementswere taken (Orion Research, model 820 dissolved oxygen meter)from the controls, and all vials were placed on a shaker table.Incubation duration was adjusted between 30 and 90 min, de-pending on the vial and snail size, to yield a drop in oxygen contentof the seawater >0.4 mg O2 l
)1.O2 consumption by snails was corrected for changes in O2 in
control vials (never exceeding ±0.2 mg O2 l)1). After the incu-bation, snail volume was measured (by displacement of seawaterin a graduated cylinder) and subtracted from vial volume to yieldincubation seawater volume. Snail tissues were dried at 65�C toconstant weight, and a subsample of the dried tissue was ashed ina muffle furnace at 475�C for 4 h. Mean (±SD) ash-free dryweight (AFDW) was 89.62±3.2% of the dry tissue weight.Weight-specific snail respiration (mg O2 g)1 AFDW h)1) wascalculated from the decrease in oxygen content normalized byincubation volume, incubation time and snail AFDW biomass.One-way analysis of covariance (ANCOVA) was conducted totest the effect of ‘‘host’’ on oxygen consumption, because O2
consumption changes with body weight. Only snails that weighed>0.03 g and had an oxygen consumption rate >0.03 mg O2 h)1
were included in the analysis (n=58), since measurements weretoo variable below those values.
Snail tissue consumption rates on A. palmata
Laboratory
A. palmata fragments (�10 cm length) were collected fromHorseshoe reef and placed in laboratory aquaria with flow-through seawater. After initial, scaled photographs were taken ofeach coral, one snail was placed on each of the fragments andallowed to feed for 48 h (n=8). Two additional fragments didnot receive snails to control for tissue loss not associated withsnail feeding. Tissue loss (cm2) was quantified by image analyses
of before and after photographs, using Sigma Scan software[Jandel Scientific Software (now SPSS), version 4.0, San Rafael,Calif.].
Field
Photographs of C. abbreviata feeding on A. palmata were taken atsix sites in the northern Florida Keys during field population sur-veys in May 1998 (Baums et al. 2003) with a housed Sony MavicaMVC-7 digital camera. Newly grazed areas are characterized byexposed coral skeleton that is white. After about 5–10 days agreenish tinge appears, indicating early macroalgal colonization.For the purpose of these estimates, a period of 7 days was used asthe average age of white, discernable feeding scars. Twenty-fourimages were judged to have clearly definable scars. The area ofthese scars was determined as the average of three replicate mea-surements with Sigma Scan software. The number of snails presenton each colony and their lengths were recorded during the fieldsurveys. The areas of the feeding scars were regressed against: (1)the number and (2) the cumulative length of snails present. Theregression coefficients provide an estimate of the area of coralgrazed per snail per 7-day period, and the area of coral grazed permillimeter of snail per 7-day period. This equation was then usedtogether with the mean length of A. palmata snails from the fieldsurveys (Baums et al. 2003) to estimate longer-term grazing ratesfor the snail population.
Calculated tissue consumption rates on each coral taxa
The general model for food budgets has been modified when ap-plied to marine gastropods (e.g. Niu et al. 1998) to account for shelland mucus production:
food requirement¼ ðrespirationþbody growthþ shell rowth
þmucus productionÞ=food conversion efficiency
where body growth includes both somatic and gonadal tissues. Theunit used in the following calculations was moles of carbon.
In this study, mean carbon content of A. palmata tissue wasmeasured in summer (see Table 2). The literature values used forMontastraea spp. include both summer and winter measurements(Szmant et al. 1990). Measured snail C content and respirationrates were averaged separately for small (0.033–0.078 g AFDW)and large (0.081–0.367 g AFDW) snails. The mean oxygen con-sumption rates include resting metabolism of female and malesnails from both hosts. Oxygen consumption was converted toCO2 production (CO2 production g)1 AFDW day)1) by assuminga respiratory quotient (RQ) of 0.71 (Schmidt-Nielsen 1997; seealso Table 3), based on the high lipid content of coral tissue(Szmant-Froehlich and Pilson 1980). Snail growth rates weretaken from the transplant experiment (described above) for bothshell and soft tissue components of snails in the large size class oneach coral host. These measurements of body growth pool thegrowth of somatic and gonadal tissues. Growth rates (body andshell) were normalized to 1 g AFDW of snail tissue, assumingthat growth rate is constant over all snail weight classes. Nor-malized growth rates were then calculated for the median sizes ofthe small (0.057 g) and large (0.159 g) size classes. Shell growth isassumed to consist primarily of calcium carbonate, with only 1%organic matrix (Burrows and Hughes 1990) (see Table 3).
Overall food conversion efficiency is a function of feeding ef-ficiency and assimilation efficiency, requiring knowledge of theamount of C lost to feces and excretion. We calculated con-sumption estimates based on food conversion efficiencies of 30%(Ansell 1982) and of 100% to bracket a range of plausible values.It is assumed that food conversion efficiencies are comparable forthe two kinds of coral tissues. The value for mucus productionwas estimated to be 10% of the total amount of carbon needed,regardless of the coral host (but see Niu et al. 1998).
1095

These measured and literature estimates were used to calculatethe area of coral tissue per day of each host taxon required tosupport a single snail and were compared with observed con-sumption rates measured for snails on A. palmata. Since all mea-surements used in this analysis were made during the summer,annual rates may be somewhat lower.
Results
Snail growth rates
In the reciprocal transplant experiment, around 40%of marked snails (Coralliophila abbreviata) were re-captured in three of the four treatment groups (n=17to 20 out of 50). However, only 10% of snails fromAcropora palmata transplanted to Montastraea spp.were recaptured. Hence, the success of recapture wasnot independent of treatment group (G-test, G=213,df=3, P< 0.001). Furthermore, while snails randomlyallocated to each treatment group should have hadsimilar mean snail weights, the A. palmata snailstransplanted to and recovered from Montastraea spp.had significantly lighter initial weights than the re-covered snails from the other three groups (i.e. smallersnails in this treatment were preferentially recovered)(Fig. 1A; one-way ANOVA on ranks, P< 0.05; Tu-key’s post hoc test, P<0.05).
Snails taken originally from both A. palmata andfrom Montastraea spp. had higher tissue growth rateswhen feeding on A. palmata than on Montastraea spp.(Fig. 1B). A 2·2 ANOVA on ranks showed a significanteffect of feeding host (P<0.05), but no effect of sourcehost (P>0.05) and no significant interaction (P>0.05).Mean increases in snail body weight over 3 months forsnails on their host of origin were 0.031 g for A. palmataand )0.002 g for Montastraea spp. (Fig. 1B). Similardifferences between snail groups were obtained whengrowth was measured as change in shell weight or shelllength (Table 1).
Host and snail tissue composition
A. palmata tissue had a higher percentage N than tissueof Montastraea spp. (Table 2), indicating a higher pro-tein content of the former. A. palmata also had a 40%lower elemental C:N ratio than Montastraea spp., indi-cating a lower tissue fat or carbohydrate content for theformer (Table 2). However, Montastraea spp. had more
tissue C and N per square centimeter skeletal area thanA. palmata (Table 2). Snails from A. palmata (n=64)and snails from Montastraea spp. (n=54) had similartissue N (%), C (%) and, hence, C:N ratios (rank sumtest, P>0.5) (Table 2).
Respirometry
Respiration rates did not differ between snails collectedfrom the two coral hosts (one-way ANCOVA, P>0.5,
Fig. 1A, B Coralliophila abbreviata. Reciprocal transplant experi-ment: A initial body weight of retrieved snails in the four treatmentgroups (box plots show median, 5th, 25th, 75th and 95thpercentiles; outliers are indicated by dots) and B change in bodyweight (means±1 SD) during the 3 month experimental growthperiod for the four treatment groups. P-values from two-wayANOVA on rank-transformed data
Table 1 Coralliophila abbreviata. Changes (means±1 SD) in shell weight (g) and maximal length (mm) of snails retrieved after 3 monthsin the reciprocal transplant experiment (NS indicates P>0.05; ** indicates P<0.001)
Snails from Acropora palmata Snails from Montastraea spp. 2·2 ANOVA
A. palmata(n=19)
Montastraea spp.(n=5)
Montastraea spp.(n=17)
A. palmata(n=20)
Originhost
Feedinghost
Interaction
D Shell weight (g) 0.45±0.47 )0.01±0.17 )0.01±0.07 0.60±0.52 NS ** NSD Length (mm) 1.42±1.34 0.26±0.62 0.09±0.43 1.94±1.76 NS ** NS
1096

Fig. 2). Smaller snails (<0.08 g AFDW, n=38) hadhigher weight-specific oxygen consumption(1.20±0.38 mg O2 h)1 g)1 AFDW, mean±1 SD) thanlarger snails (>0.08 g AFDW, n=25; 0.89±0.30 mg O2
h)1 g)1 AFDW).
Snail tissue consumption rates on A. palmata
Laboratory
A. palmata fragments grazed by C. abbreviata in thelaboratory lost a mean of 4 cm2 tissue snail)1 day)1
(range 1–9 cm2 tissue snail)1 day)1). Control coralfragments did not lose tissue over the 48 h duration ofthe experiment. In one case, a grazed fragment contin-ued to slough off tissue after the snail was removed, untilthe entire fragment died (�72 h).
Field
The area of A. palmata scars associated with naturalaggregations of C. abbreviata was correlated withthe number of snails in each aggregation [lesion
area=7.01(no. of snails), r2=0.49, P<0.001, n=24].The goodness-of-fit of the linear regressions betweenlesion area and snail abundance was improved (tor2=0.61) by taking into account the sizes of the snailsfound on each coral (Fig. 3). Using the average snail size(on A. palmata) of 29 mm, this relationship yields anestimate of long-term feeding rates for C. abbreviata of1.07 cm2 A. palmata tissue snail)1 day)1.
Calculated tissue consumption rates
The C requirements for respiration made up the largestcomponent of the carbon budget and were similar forsnails from Montastraea spp. and from A. palmata (Ta-ble 3). The calculated maximum tissue consumption ratewas 3.28 cm2 coral tissue per day (large A. palmata snailsin summer assuming a 30% conversion efficiency; Ansell1982). The higher carbon content and per area biomassof tissue ofMontastraea spp. (see also Szmant et al. 1990)leads to a much smaller amount of coral tissue area re-quired to sustain individual resident snails (0.13–0.88 cm2 tissue snail)1 day)1) compared to A. palmatatissue (0.44–3.28 cm2 tissue snail)1 day)1) (Table 3).
Fig. 2 Coralliophila abbreviata. Oxygen consumption rates of snailscollected from Acropora palmata and Montastraea spp. (AFDWash-free dry weight)
Fig. 3 Coralliophila abbreviata. Field observations of feeding scars.Surface area of lesion (cm2) is plotted against cumulative snaillength (cm). Line and regression equation from least-squares linearregression
Table 2 Tissue carbon and nitrogen content and C:N ratio of twocoral hosts and of Coralliophila abbreviata feeding on the two coralhosts (means±SD). Values for Montastraea spp. tissue are fromSzmant et al. (1990) and include summer and winter collections.
Acropora palmata was collected in summer only. Carbon and ni-trogen contents are expressed for corals as lg-at cm)2 and for snailsas lg-at g)1 ash-free dry weight (AFDW) tissue
A. palmata(n=9)
Montastraea spp.(n=40)
Snails onA. palmata (n=64)
Snails on Montastraea spp.(n=54)
C (% by weight) 55.3±1.7 52.2±2.1 41.3±5.3 41.7±6.6N (% by weight) 10.3±0.5 5.6±0.5 9.4±1.2 9.5±1.8C:N (elemental ratio) 6.3±0.3 10.9±0.5 5.2±0.9 5.2±0.9C (lg-at) 343±91 (cm)2) 983±188 (cm)2) 34,451±4,409 (g)1) 34,721±5,507 (g)1)N (lg-at) 54.7±15.7 (cm)2) 88.2±14.6 (cm)2) 6,719±8,54 (g)1) 6,821±1,266 (g)1)Tissue AFDW (g snail)1) NA NA 0.12±0.12 0.05±0.03
1097
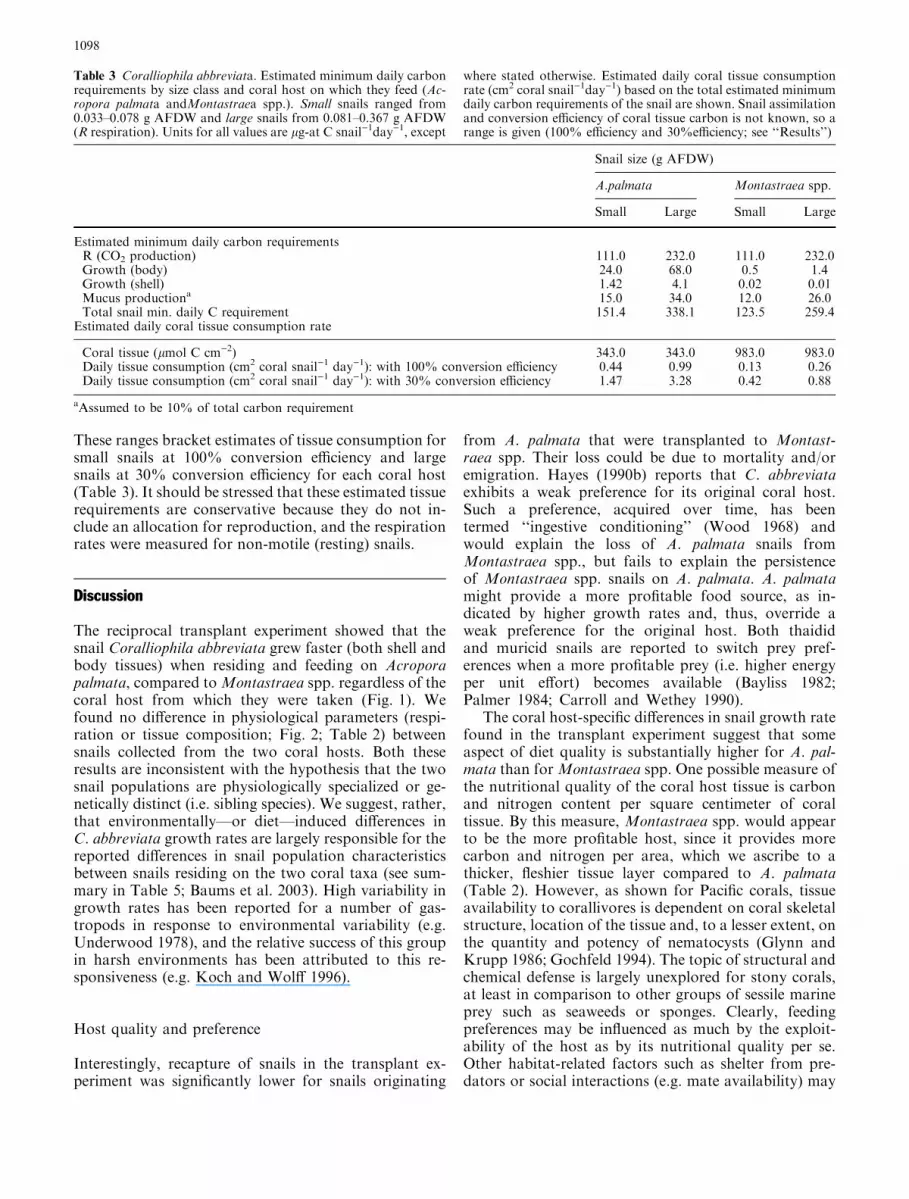
These ranges bracket estimates of tissue consumption forsmall snails at 100% conversion efficiency and largesnails at 30% conversion efficiency for each coral host(Table 3). It should be stressed that these estimated tissuerequirements are conservative because they do not in-clude an allocation for reproduction, and the respirationrates were measured for non-motile (resting) snails.
Discussion
The reciprocal transplant experiment showed that thesnail Coralliophila abbreviata grew faster (both shell andbody tissues) when residing and feeding on Acroporapalmata, compared toMontastraea spp. regardless of thecoral host from which they were taken (Fig. 1). Wefound no difference in physiological parameters (respi-ration or tissue composition; Fig. 2; Table 2) betweensnails collected from the two coral hosts. Both theseresults are inconsistent with the hypothesis that the twosnail populations are physiologically specialized or ge-netically distinct (i.e. sibling species). We suggest, rather,that environmentally—or diet—induced differences inC. abbreviata growth rates are largely responsible for thereported differences in snail population characteristicsbetween snails residing on the two coral taxa (see sum-mary in Table 5; Baums et al. 2003). High variability ingrowth rates has been reported for a number of gas-tropods in response to environmental variability (e.g.Underwood 1978), and the relative success of this groupin harsh environments has been attributed to this re-sponsiveness (e.g. Koch and Wolff 1996).
Host quality and preference
Interestingly, recapture of snails in the transplant ex-periment was significantly lower for snails originating
from A. palmata that were transplanted to Montast-raea spp. Their loss could be due to mortality and/oremigration. Hayes (1990b) reports that C. abbreviataexhibits a weak preference for its original coral host.Such a preference, acquired over time, has beentermed ‘‘ingestive conditioning’’ (Wood 1968) andwould explain the loss of A. palmata snails fromMontastraea spp., but fails to explain the persistenceof Montastraea spp. snails on A. palmata. A. palmatamight provide a more profitable food source, as in-dicated by higher growth rates and, thus, override aweak preference for the original host. Both thaididand muricid snails are reported to switch prey pref-erences when a more profitable prey (i.e. higher energyper unit effort) becomes available (Bayliss 1982;Palmer 1984; Carroll and Wethey 1990).
The coral host-specific differences in snail growth ratefound in the transplant experiment suggest that someaspect of diet quality is substantially higher for A. pal-mata than for Montastraea spp. One possible measure ofthe nutritional quality of the coral host tissue is carbonand nitrogen content per square centimeter of coraltissue. By this measure, Montastraea spp. would appearto be the more profitable host, since it provides morecarbon and nitrogen per area, which we ascribe to athicker, fleshier tissue layer compared to A. palmata(Table 2). However, as shown for Pacific corals, tissueavailability to corallivores is dependent on coral skeletalstructure, location of the tissue and, to a lesser extent, onthe quantity and potency of nematocysts (Glynn andKrupp 1986; Gochfeld 1994). The topic of structural andchemical defense is largely unexplored for stony corals,at least in comparison to other groups of sessile marineprey such as seaweeds or sponges. Clearly, feedingpreferences may be influenced as much by the exploit-ability of the host as by its nutritional quality per se.Other habitat-related factors such as shelter from pre-dators or social interactions (e.g. mate availability) may
Table 3 Coralliophila abbreviata. Estimated minimum daily carbonrequirements by size class and coral host on which they feed (Ac-ropora palmata andMontastraea spp.). Small snails ranged from0.033–0.078 g AFDW and large snails from 0.081–0.367 g AFDW(R respiration). Units for all values are lg-at C snail)1day)1, except
where stated otherwise. Estimated daily coral tissue consumptionrate (cm2 coral snail)1day)1) based on the total estimated minimumdaily carbon requirements of the snail are shown. Snail assimilationand conversion efficiency of coral tissue carbon is not known, so arange is given (100% efficiency and 30%efficiency; see ‘‘Results’’)
Snail size (g AFDW)
A.palmata Montastraea spp.
Small Large Small Large
Estimated minimum daily carbon requirementsR (CO2 production) 111.0 232.0 111.0 232.0Growth (body) 24.0 68.0 0.5 1.4Growth (shell) 1.42 4.1 0.02 0.01Mucus productiona 15.0 34.0 12.0 26.0Total snail min. daily C requirement 151.4 338.1 123.5 259.4
Estimated daily coral tissue consumption rate
Coral tissue (lmol C cm)2) 343.0 343.0 983.0 983.0Daily tissue consumption (cm2 coral snail)1 day)1): with 100% conversion efficiency 0.44 0.99 0.13 0.26Daily tissue consumption (cm2 coral snail)1 day)1): with 30% conversion efficiency 1.47 3.28 0.42 0.88
aAssumed to be 10% of total carbon requirement
1098

also greatly influence host choice by small, relativelyimmobile predators (Lasker et al. 1988).
Snail feeding rates and preferences
Table 4 summarizes feeding rate estimates for snails onA. palmata from this and other studies. Our laboratoryobservations are somewhat higher than available fieldestimates. The correlations between area of tissue dam-age and the cumulative snail length (Fig. 3) or snailabundance (Miller 2001) provide strong evidence thatsnail feeding, rather than some sort of secondary infec-tion, is the direct cause of coral tissue loss.
Repeated attempts to quantify C. abbreviata feedingrates onMontastraea spp. hosts failed (due to the difficultyin distinguishing and measuring the small Montastraeaspp. feeding scars), making direct comparisons of snailimpact on the two coral hosts in terms of area consumedunfeasible. Instead, estimates were derived for snail tissueconsumption rates for both coral hosts from energeticcalculations based on parameters measured in this study(Table 3). The assumptions underlying these calculationsare conservative and should lead to conservative estimatesof snail feeding rates. Indeed, the calculated values forsnails feeding on A. palmata fall within the range ofmeasured field consumption rates (Table 4). This con-gruence of calculated and observed tissue consumptionrates for snails feeding onA. palmata lends confidence thatthe low snail consumption rates (both calculated andobserved) for Montastraea spp. are reasonable.
It does not necessarily follow, however, that Mon-tastraea spp. is the less favored host, since snail size andfitness may be de-coupled. It has been suggested forintertidal gastropods at high density that intraspecificcompetition results in early cessation of somatic growthand continuing investment in reproduction (Underwood1978; Koch and Wolff 1996). Hence, snails living athigher density, such as C. abbreviata on Montastraeaspp. hosts, may have higher fitness despite smaller size.
Alternative feeding modes have been reported forcoralliophillids. Coralliophila violacea, a corallivorepreying on poritid corals in the Red Sea, feeds on hostsecretions at the site of injury that it creates (‘‘prudentsessile feeding’’, Oren et al. 1998). Hayes (1990a) ob-served C. abbreviata with its proboscis inserted into the
polyps of Agaricia tenuifolia and Montastraea annularis,‘‘stealing food’’ from the polyp. We cannot exclude thepossibility that C. abbreviata populations from Floridautilize different feeding strategies depending on the kindof coral host they inhabit, but we think it to be unlikely.Energetic calculations (Table 3) indicate that the higheramount of tissue carbon per skeletal area inMontastraeaspp. might be sufficient to explain differences in the sizeof snail feeding scars between the two coral host taxa.
Effect of corallivory on coral hosts
At measured levels of snail densities on Key Largo, Fla.,reefs, infested colonies of A. palmata were losing be-tween 1.32 and 9.84 cm2 tissue day)1, while infestedcolonies of Montastraea spp. colonies were losing be-tween 1.04 and 7.04 cm2 tissue day)1 due to snail pre-dation. A total of 20% of all surveyed A. palmatacolonies and 50% of all surveyed Montastraea spp.colonies harbored snails (Baums et al. 2003) and weresubject to such predation levels. These calculations werebased on the mean number of snails per infested colony(three and eight snails for A. palmata and Montastraeaspp. respectively; Baums et al. 2003), and calculatedsnail feeding rates for each host taxon (Table 4; presentstudy).
The cost of C. abbreviata predation to its coral hostscan be expressed as the amount of energy that is divertedfrom growth and reproduction to regenerate lost tissue.Demonstrated trade-offs between lesion repair andgrowth rate (e.g. for A. palmata: Bak 1983) and fecun-dity (Montastraea spp.: Szmant-Froehlich 1985; VanVeghel and Bak 1994; Stylophora pistillata: Rinkevichand Loya 1989) indicate that regeneration is a process atleast as important as growth and reproduction (Meesterset al. 1992). Coral species have differential ability torepair lesions according to the mechanism used (higherfor A. palmata than Montastraea spp.: Bak 1983; Me-esters et al. 1992; Van Veghel and Bak 1994) (Table 5).The rate of recovery depends on initial size and shape ofthe lesion (e.g. Meesters et al. 1997; Oren et al. 1998),and corals under stress have lower regeneration poten-tial (Meesters et al. 1992).
Published studies report recovery rates for lesionsinflicted experimentally upon the coral in an area
Table 4 Coralliophila abbreviata. Feeding rates of snails on Montastraea spp. and Acropora palmata, derived from field and laboratoryobservations as well as calculated estimates (see ‘‘Discussion’’) (NA not available)
Snail feeding rates(cm2 coral snail)1 day)1)
Snails on A.palmata
Snails onMontastraea spp.
Time frame ofobservation
Reference
Range Mean Range Mean
Field Up to 6.5 1.9 NA NA 1 month Bruckner et al. (1997) (Puerto Rico)0–4.3 1.3 NA NA 3 month Miller (2001) (Florida Keys)
Field, photos of scars NA 1.1 NA NA �7 days Present study, Fig. 3Laboratory 1.0–9.0 4.0 NA NA 2 days Present studyCalculated 0.4–3.3 NA 0.1–0.9 NA NA Present study, Table 3
1099

surrounded by living tissue. Lesion perimeter is theprimary factor determining the amount of area that willbe regenerated (Meesters et al. 1997). Snail predationoccurs primarily at the edge of living tissue (i.e. less liveperimeter), so published regeneration rates should pro-vide a reasonable estimate of recovery time for snailfeeding scars. Published mean regeneration rates are0.205 cm2 day)1 for A. palmata (Meesters and Bak 1995)and 0.139 cm2 day)1 forM. annularis (Meesters and Bak1993). The apparent difference between tissue re-growthrates of the two coral taxa and the rates of consumptionby C. abbreviata translate into a similar range of re-covery times of 6.4–49.2 and 7.5–50.6 days for the twohosts, respectively, for 1 day of feeding at mean snaildensities (see above). Snail aggregations on Montastraeaspp. consist of smaller, more numerous individuals thanthose on A. palmata, and they may have greater tem-poral persistence (Hayes 1990a); while the feeding scarsare smaller and less noticeable than those on A. palmata,the snails can cause costly damage to both coral taxa(Table 5). In any case, if sustained, the rates of preda-tion described in this study suggest a serious impact onthe viability of the infested colonies of both taxa.
Clearly, field observations are needed of feeding-scarregeneration (rates and metabolic costs), and of subse-quent coral growth and reproductive performance forboth coral taxa. Further, the interaction of snail size anddensity is unknown, and there may be a self-regulatingmechanism for net snail biomass per area. Interspecificdifferences in the ability of corals to defend themselvesagainst corallivores and the costs of doing so are virtu-ally unknown.
The finding of high rates of coral tissue consumptionby C. abbreviata on A. palmata and Montastraea spp. inthe Florida Keys, coupled with the high levels of snail
infestation observed there (Baums et al. 2003) calls forimproved quantification of the relative importance ofcorallivore predation as a threat to the population dy-namics and persistence of these important hermatypiccorals. This need is particularly urgent in the currentcontext of general and global coral population decline(Wilkinson 2000).
Acknowledgements This study was funded by a grant of the Na-tional Undersea Research Center/Univ. of North Carolina, Wil-mington (UNCW 9824) to M.W.M. and A.M.S., and conductedunder Florida Keys National Marine Sanctuary permit no.FKNMS-084-98. This study benefited greatly from the logisticalsupport of the staff at the NURC/UNCW facility, Key Largo. M.Aydin derived the formulas for calculating shell and body weight ofsnails. K. Fite is gratefully acknowledged for analyzing snailfeeding pictures. We are thankful for helpful comments from N.Knowlton and M. Hay on an earlier version of this manuscript.The suggestions of P. Sammarco and three anonymous reviewersimproved the manuscript and are gratefully acknowledged. P.Glynn is thanked for editorial help. G. Hempel provided valuableguidance. Experiments conducted in this study comply with U.S.laws. This study was conducted in partial fulfillment of a Diplomdegree at the University of Bremen, Germany.
References
Ansell AD (1982) Experimental studies of a benthic predator–preyrelationship. II. Energetics of growth and reproduction, andfood-conversion efficiencies, in long-term cultures of the gas-tropod drill Polinices alderi (Forbes) feeding on the bivalveTellina tenuis da Costa. J Exp Mar Biol Ecol 61:1–29
Bak RPM (1976) The growth of coral colonies and the impor-tance of crustose coralline algae and burrowing sponges inrelation with carbonate accumulation. Neth J Sea Res10:285–337
Bak RPM (1983) Neoplasia, regeneration and growth in the reef-building coral Acropora palmata. Mar Biol 77:221–227
Table 5 Coralliohpila abbreviata. Summary of the population andfeeding characteristics of snails residing on Acropora palmata andMontastraea spp. (M male; F female). References: a Baums et al.2003; b Szmant et al. 1990; c Bruckner et al. 1997; d Hayes 1990a;
e Hayes 1990b; f Hayes 1989; g Ott and Lewis 1972; h Wells andLalli 1977; i Miller 2001; j Gladfelter et al. 1978; k Bak 1976; lMeesters et al. 1992)
Characteristics Coral host Fig./Table,present study
Reference
A. palmata Montastraea
Coral host Host density Low–High Low aHost carbon content/area Low High Table 3 bGrowth rate High Low j, kRegeneration potential High Low l
Snail populationinformation
Infestation of corals Low High a, c, dDensity on infested corals Low High c, d, gSnail size Large Small a, d, f, c, hSex ratio M>F M=F a, c, e, hStability of aggregations Low High eMobility of snails High Low e
Snail physiology Snail growth High Low Table 2; Fig. 1Metabolic requirements Equal Equal Fig. 2Tissue composition (C:N) Equal Equal Table 3
Snail impact Area eaten Large Small Table 4 c, iSnail populationregulation
Apparent predationpressure on snailsfrom snail shell scars
High Low Unpubl. data
1100

Bak RPM, Steward-Van Es Y (1980) Regeneration of superficialdamage in the scleractinian corals Agaricia agaricites, F. pur-purea and Porites asteroides. Bull Mar Sci 30:883–887
Baums IB (2000) Population ecology and feeding physiology ofCoralliophila abbreviata, Lamarck (Coralliophilidae), a coral-livorous gastropod in the Florida Keys. Diplomarbeit, Uni-versitat zu Bremen, Bremen
Baums IB, Miller WM, Szmant AM (2003) Ecology of a coralliv-orous gastropod. Coralliophila abbreviata, on two slceractinianhosts I: Population structure of snails and corals. Mar Biol10.1007/s00227-003-1024-9
Bayliss DE (1982) Switching by Lepsiella vinosa (Gastropoda) inSouth Australian mangroves. Oecologia 54:221–226
Bruckner RJ, Bruckner AW, Williams EH Jr (1997) Life historystrategies of Coralliophila abbreviata Lamarck (Gastropoda:Coralliophilidae) on the southeast coast of Puerto Rico. In:Lessios HA, MacIntyre IG (eds) Proc 8th Int Coral Reef Symp,vol 1. Smithsonian Tropical Research Institute, Balboa, Pana-ma, pp 627–632
Burrows MT, Hughes RN (1990) Variation in growth and con-sumption among individuals and populations of the dogwhelksNucella lapillus: a link between foraging behavior and fitness.J Anim Ecol 59:723–742
Carroll ML, Wethey DS (1990) Predator foraging behavior: effectof a novel prey species on prey selection by a marine intertidalgastropod. J Exp Mar Biol Ecol 139:101–117
Fofonoff P, Millard RC Jr (1983) Algorithms for computation offundamental properties of seawater. UNESCO Tech Pap MarSci 44:1–53
Gladfelter EH, Monahan RK, Gladfelter WB (1978) Growth ratesof five reef-building corals in the northeastern Caribbean. BullMar Sci 28:728–734
Glynn PW, Krupp DA (1986) Feeding biology of a Hawaiian seastar corallivore, Culcita novaeguineae Muller & Troschel. J ExpMar Biol Ecol 96:75–96
Gochfeld DJ (1994) Predation-induced defenses in hard corals re-duce susceptibility to future predation. Pac Sci 48:203
Hayes JA (1989) The biology and ecology of Coralliophila abbre-viata Lamarck (Gastropoda: Coralliophilidae): the importanceof corallivores in the ecology of coral reefs. Dissertation, Uni-versity of Texas, Austin
Hayes JA (1990a) Distribution, movement and impact of the cor-allivorous gastropod Coralliophila abbreviata (Lamarck) on aPanamanian patch reef. J Exp Mar Biol Ecol 142:25–42
Hayes JA (1990b) Prey preference in a Caribbean corallivore,Coralliophila abbreviata (Lamarck) (Gastropoda, Coralliophil-idae). Bull Mar Sci 47:557–560
Koch V, Wolff M (1996) The mangrove snail Thais kiosquiformisDuclos: a case of life history adaptation to an extreme envi-ronment. J Shellf Res 15:421–432
Lasker HR, Coffroth MA, Fitzgerald LM (1988) Foraging patternsof Cyphoma gibbosum on octocorals: the roles of host choiceand feeding preference. Biol Bull (Woods Hole) 174:254–266
Meesters EH, Bak RPM (1993) Effects of coral bleaching on tissueregeneration potential and colony survival. Mar Ecol Prog Ser96:189–198
Meesters EH, Bak RPM (1995) Age-related deterioration of aphysical function in the branching coral Acropora palmata. MarEcol Prog Ser 121:203–209
Meesters EH, Bos A, Gast GJ (1992) Effects of sedimentation andlesion position on coral tissue regeneration. In: Richmond RH(ed) Proc 7th Int Coral Reef Symp, vol 2. University of Guam,Mangilao, pp 671–677
Meesters EH, Pauchli W, Bak RPM (1997) Predicting regenerationof physical damage on a reef-building coral by regenerationcapacity and lesion shape. Mar Ecol Prog Ser 146:91–99
Miller MW (2001) Corallivorous snail removal: evaluation of im-pact on Acropora palmata. Coral Reefs 19:293–295
Millero FJ, Chen CT, Bradshaw A, Schleicher K (1980) A new highpressure equation of state for seawater. Deep-Sea Res 27:255–264
Niu C, Nakao S, Goshima S (1998) Energetics of the limpet Lottiakogomogai (Gastropoda: Acmaeidae) in the intertidal rockyshore in southern Hokkaida, Japan. J Exp Mar Biol Ecol224:167–181
Oren U, Brickner I, Loya Y (1998) Prudent sessile feeding by thecorallivore snail Coralliophila violacea on coral energy sinks.Proc R Soc Lond B Biol Sci 265:2051–2055
Ott B, Lewis JB (1972) The importance of the gastropod Coral-liophila abbreviata (Lamarck) and the polychaete Hermodicecarunculata (Pallas) as coral reef predators. Can J Zool50:1651–1656
Palmer RA (1982) Growth in marine gastropods: a non-destructivetechnique for independently measuring shell and body weight.Malacologia 23:63–73
Palmer RA (1984) Prey selection by thaidid gastropods: some ob-servational and experimental field tests of foraging models.Oecologia 62:62–72
Rinkevich B, Loya Y (1989) Reproduction in regenerating coloniesof the coral Sylophora pistillata. In: Spanier E, et al (eds) En-vironmental quality and ecosystem stability, IV, B. Environ-mental quality. Jerusalem, Israel, pp 257–265
Schmidt-Nielsen K (1997) Animal physiology: adaptation and en-vironment, 5th edn. Cambridge University Press, Cambridge
Szmant AM, Ferrer LM, FitzGerald LM (1990) Nitrogen excretionand O:N ratios in reef corals: evidence for conservation of ni-trogen. Mar Biol 104:119–127
Szmant-Froehlich A (1985) The effect of colony size on the re-productive ability of the Caribbean coral Montastrea annularis(Ellis and Solander). In: Gabrie C, et al (eds) Proc 5th Int CoralReef Congr, vol 4. Antenne Museum–EPHE, Moorea, Tahiti,pp 295–300
Szmant-Froehlich A, Pilson MEQ (1980) The effects of feedingfrequency and symbiosis with zooxanthellae on the biochemicalcomposition of Astrangia danaeMilne Edwards & Haime, 1849.J Exp Mar Biol Ecol 48:85–91
Underwood AJ (1978) The ecology of intertidal gastropods. AdvMar Biol 16:111–210
Van Veghel MLJ, Bak RPM (1994) Reproductive characteristics ofthe polymorphic Caribbean reef building coral Montastraeaannularis. III. Reproduction in damaged and regenerating col-onies. Mar Ecol Prog Ser 109:229–233
Weil E, Knowlton N (1994) A multi-character analysis of the Ca-ribbean coral Montastraea annularis (Ellis and Solander, 1786)and its two sibling species, M. faveolata (Ellis and Solander,1786) and M. franksi (Gregory, 1895). Bull Mar Sci 55:151–175
Wells FE, Lalli CM (1977) Reproduction and brood protection inthe Caribbean gastropods Coralliophila abbreviata and C. car-ibaea. J Molluscan Stud 43:79–87
Wilkinson CR (2000) Status of coral reefs of the world. AustralianInstitute of Marine Science, Cape Ferguson
Wood L (1968) Physiological and ecological aspects of prey selec-tion by the marine gastropod Urosalpinx cinerea (Prosobran-chia: Muricidae). Malacologia 6:267–320
1101





























