Embed Size (px)
Citation preview

Oecologia (1991) 87:8-18
Oecologia �9 Springer-Verlag 1991
Interactions between freshwater snails and tadpoles: competition and facilitation Christer Briinmark, Simon D. Rundle*, and Ann Erlandsson
Department of Ecology, University of Lund, Helgonav/igen 5, S-223 62 Lund, Sweden
Received December 28, 1990 / Accepted in revised form January 29, 1991
Summary. Freshwater snails and anuran tadpoles have been suggested to have their highest population densities in ponds of intermediate size where abiotic disturbance (e.g. desiccation) is low and large predators absent. Both snails and tadpoles feed on periphytic algae and, thus, there should be a large potential for competitive interac- tions to occur between these two distantly related taxa. In a field experiment we examined the relative strength of competition between two closely related snail species, Lymnaea stagnalis and L. peregra, and between L. stag~ nalis and tadpoles of the common frog, Rana temporaria. Snail growth and egg production and tadpole size at and time to metamorphosis were determined. Effects on the common food source, periphyton, were monitored with the aid of artificial substrates. Periphyton dry weight was dramatically reduced in the presence of snails and/or tadpoles. There were no competitive effects on growth or egg production of the two snail species when they were coexisting. Mortality of L. peregra was high (95 %) after reproduction, but independent of treatment. Growth of L. stagnalis was reduced only at the highest tadpole densities, whereas egg production was reduced both by intraspecific competition and by competition with tad- poles. Differences in egg production were retained after tadpole metamorphosis. Tadpole larval period increased, weight of metamorphosing frogs decreased and growth rate was reduced as a function of increasing tadpole density. However, contrary to expectation, snails had a positive effect on tadpole larval period, weight and growth rate. Further, in experimental containers without snails there was a dense growth of the filamentous green alga Cladophora sp. We suggest that the facilitative ef- fects of snails on tadpoles are due to an "indirect mutual- istic" mechanism, involving competition between food sources of different quality (microalgae and Cladophora sp.) and tadpoles being competitively dominant over snails for the preferred food source (microalgae). In the
* Present address: Catchment Research Group, N,R.A. Laborat- ory, Penyfai House, Furnace, Llanelli, Dyfed SA15 4EL, UK
Offprint requests to: C. Brrnmark
presence of tadpoles snails will be forced to feed on low-quality Cladophora, increasing nutrient turnover rates, which results in enhanced productivity of micro- algae, increasing tadpole food resources. Thus, tadpoles have a negative effect on snails through resource depres- sion, while snails facilitate tadpole growth through an indirect enhancement of food availability.
Key words: Competition - Facilitation Freshwater snails - Tadpoles - Ponds
A large number of experimental field studies have shown the existence and importance ofinterspecific competition (see reviews by Connell 1983; Schoener 1983). However, as pointed out in a recent paper by Morin et al. (1988), there is a strong bias towards studying interactions be- tween closely related species, and remarkably few inves- tigators have considered the effects of interspecific com- petition for food between taxonomically distant species. The importance of interphyletic competition between aquatic herbivores was demonstrated by Morin et al. (1988), who found that the intensity of competition be- tween insect larvae and anuran tadpoles was as strong as when tadpoles of two anuran species were competing. Both the insect larvae and the tadpoles are herbivores, grazing on periphytic algae. Here, we study interactions between tadpoles and another group of pcriphyton grazers, freshwater snails.
According to the predictions of food chain theory (e.g. Hairston et al. 1960; Fretwell 1987) competition should only be an important structuring force at the herbivore level when predators are absent. Lodge et al. (1987), in a conceptual model, argued that competition between freshwater snails is only important in intermediately sized water bodies. In small, temporary ponds snails are regulated by abiotic factors (desiccation, winterkill) but as the size of the pond increases catastrophic events will be less common and populations build up to levels where competition becomes the major structuring force. How-

ever, in larger, permanent ponds predat ion by large predators such as fish and crayfish will reduce snail popu- lations and be the most impor tant structuring force. In parallel, Wilbur (1984) argued that survival o f anuran tadpoles is dependent on desiccation in small, t emporary ponds, whereas in more undisturbed ponds predation will be determining survival and body size at metamor- phosis. Competi t ion between tadpoles is expected to be an impor tant mechanism of communi ty regulation only in ponds that predators have not successfully colonized. Thus, both snails and tadpoles are predicted to have high populat ion densities in small ponds without predators. Because of their ability to effectively reduce periphyton biomass (Dickman 1968; Br6nmark 1989), there should be a large potential for interspecific competi t ion to occur between these two distantly related taxa in small ponds during spring months before tadpoles metamorphose. This period is extremely important to both taxa; popula- tion size of frogs depend on size and abundance of meta- morphosing froglets which, in turn, is affected by com- petitive interactions acting on the tadpole stage (Wilbur 1980), whereas for snails the main reproduction period occurs during these spring months. A large number of laboratory and field experiments have demonstrated den- sity-dependent growth rates in anuran tadpoles (e.g. Wil- bur 1976, 1977; Semlitsch and Caldwell 1982; Travis 1983; Cummins 1989), resulting in differences in size of metamorphos ing frogs and/or length of the larval period. Size and date at metamorphosis are directly related to fitness of the adult amphibians: larger metamorphs can result in larger size at first reproduction (Berven and Gill 1983; Smith 1987; Semlitsch et al. 1988) which, in turn, is related to mating success in males (Howard 1980; Berven 1981) and fecundity of females (Berven 1982; Semlitsch 1985; Gibbons and McCar thy 1986). In snails intra- and interspecific competi t ion has been shown to reduce growth rates and egg product ion (e.g. Eisenberg 1966, 1970; Brown 1979, 1982).
In this study we contrast the relative importance of competi t ion between closely and distantly related taxa. We do this by examining competitive effects between two freshwater snail species, Lymnaea stagnalis and L. pere- gra, and between L. stagnaIis and tadpoles of the com- mon frog, Rana temporaria. Effects of competi t ion on snail growth and reproduction and on tadpole size at and time to metamorphosis were quantified, and, in addition, biomass of the mutual food source, periphytic algae, was monitored.
Materia ls and methods
Experimental design
We used a field experiment to study the effects of interphyletic, interspecific and intraspecific competition on snails and tadpoles. To determine the relative importance of the different kinds of competition we used the experimental design for studying com- petitive asymmetries suggested by Underwood (1986; see also Mor- in and Johnson 1988). The basic design involves five treatments: low density of species A, high density of species A, low density of species B, high density of species B, and species A and B together, each at
Table 1. Experimental design. Density of the freshwater snail Lym- naea stagnalis and a competitor species in the different treatments. The competitor was either L. peregra or tadpoles of the common frog, Rana temporaria
Treatment
1 2 3 4 5
A. Lymnaea stagnalis 5 10 5 - - Competitor - - 5 10 5
B. Lymnaea stagnalis 5 5 5 5 5 Competitor 2 5 10 15 20
the low density (see Table 1). This allowed us to evaluate the relative importance of intra- versus interspecific competition. Further, by comparing the results from treatments with the two snail species to those with snail-tadpole treatments, we could estimate the impor- tance of competition between closely related species as compared to competition between distant taxa. However, with this basic design we could only detect the presence or absence of competition and evaluate possible asymmetries in competitive relations. To investigate how the effect of competition was related to increasing densities of the competing species we added a series of treatments where the density of the snail L. stagnalis was kept constant, while the density of the competitor (L. peregra or tadpoles) gradually increased (response-curve experiment). A final, control treatment contained no animals and was used to estimate periphyton biomass in the absence of grazers. All treatments had five replicates.
The experimental unit consisted of a 25-1 plastic bucket. The 85 buckets used were divided into five groups of 17 and each group was attached to a wooden raft that was placed in a small eutrophic pond 45 km east of Lund, southern Sweden. Fine-mesh (250-gm) win- dows (14 x 10 cm) in the sides of the buckets allowed exchange of water with the surrounding watermass, but prevented colonization by invertebrates. Leaf litter (1.0 1 dried birch leaves) and sand (0.15 1) were added to the bucket to provide some structure. To quantify periphyton we added a strand of plastic flagging anchored at one end with an iron nut and kept vertical with small styrofoam floats. Coarse-mesh (approx. 2 mm) lids precluded colonization by insects and also retained metamorphosing anurans. The buckets were attached to the raft so that there was a small distance (approx. 4 cm) between the water surface and the lid, thus providing an air space for the air breathing pulmonate snails. Rafts were placed in the pond on 20 March, 4 weeks before the start of the experiment, to permit periphyton to colonize.
Study organisms
The two lymneid snail species used in this study commonly co-occur in ponds in southern Sweden (Br6nmark 1985) and many of these ponds serve as breeding grounds for the common frog, Rana tem- poraria (CB, pers.obs.). Lymnaeaperegra reproduces in early spring and has a univoltine life cycle with complete replacement (Young 1975; Dussart 1979; Gaten 1986). In especially productive habitats a second generation may occur in late summer (Russel-Hunter 1961). In southern Sweden Lyrnnaea stagnalis has a biannual life cycle with reproduction after both the first and second overwinter- ing (C.B., pers.obs.; see also Berrie 1965). However, most of the adult snails die after their first reproductive season with less than 10% surviving to breed a second year (Berrie 1965). In early April Rana temporaria egg masses are deposited in ponds and tadpoles hatch after 2-3 weeks. Emergence of metamorphs commence in early June. Both snails and tadpoles feed on periphytic algae (e.g. Calow 1970; Scheerbom and Van Elk 1978; Dickman 1968); in addition, tadpoles can filterfeed on phytoplankton (e.g. Kenny 1969).

10
Newly-laid spawn from a single pair of Rana temporaria in amplexus was taken from a pond close to the study site on 13 April and brought to the laboratory to hatch. On 18-20 April overwinter- ing adult L. stagnalis (24.3 _+ 2.4 ram, mean total length ___ SD) and L. peregra (12.4_+ 1.0 ram) were collected from nearby ponds. All snails of a species were collected from the same pond, thus reducing any effects of genetic inter-population differences in life history traits (Calow 1981). They were individually marked with a plastic number tag (1 x 2 mm) attached to the shell with epoxy resin and their length measured (apex-base shell length; to the nearest 0.5 ram). Marking allowed us to monitor individual growth rate for each treatment. Tadpoles (19.8 rag• 1.1; mean dry weight_+ SD, n = 10) were added to experimental containers on 18 April, L. stagnalis on 20 April and L. peregra on 21 April. By this time a dense algal layer was growing on the inside of the containers and on the plastic flagging. Five snails/tadpoles per bucket (0.2/1) was considered as a low density treatment whereas high density treatments received 10 snails/tadpoles. In the response curve experiment L. stagnalis was kept at a constant density (5 snails/bucket) while L. peregra or tadpoles were added in a gradient from 2 to 20 per bucket (see aIso Table I). The densities used in this experiment are within the range of densities found in natural ponds (CB, personal observation, J. Loman, personal communication). Each raft was considered as one experimental block and treatments were randomized within blocks.
Response variables
Snails. Lymneid snails deposit their eggs in masses on solid sub- strates such as stones or macrophytes. In the containers the snails deposited their eggs on container walls, net windows and on the plastic tape. Starting on 2 May all egg masses were collected weekly from each treatment. Egg masses from each container were placed in pond water in a jar and in the laboratory the number of masses and egg numbers per mass were counted under a dissecting scope. Differences in egg mass shape and egg size (Bondesen 1950) enabled separation of L. stagnalis and L. peregra egg masses. Embryonic development in lymneids take up to 3 weeks (Berrie 1965), thus all egg masses were collected before any eggs had hatched. During the weekly sampling on 22 May we noticed that most L. peregra in- dividuals bad died, so on 24 May this part of the experiment was terminated. All snails were removed and shell lengths were mea- sured on both dead and live snails. Snails from the snail-tadpole experiment were removed and returned to the laboratory when most of the tadpoles had emerged (14 June). After length measure- ments snails were returned to their respective container. Egg mass collection continued after tadpole emergence through to 9 August to monitor whether, during this period, snails could compensate for any reductions in reproduction due to competition with tadpoles. The experiment was ended on 9 August when all snails were collect- ed and measured.
Tadpoles. Tadpole emergence commenced in early June. From this time the pond was visited daily and metamorphs (forelimbs emerged) collected and taken to the laboratory. Froglets were kept in the laboratory until tail resorption was completed when wet weight (towel-dry) was determined to the nearest 0.1 rag. Frogs were then returned to the pond and released.
Periphyton. Periphyton was quantified on 25 May. This was after the end of the L. peregra experiment and, thus, periphyton data from L. peregra treatments were omitted. Plastic strips from the L. stagnalis/tadpole containers were removed and transferred to a jar with tap water, taken to the laboratory and stored in darkness at 4 ~ C until further processed (within 24 h). Periphyton was removed by brushing and suspended in a known volume of water (approxi- mately 0.5 1). From this algal suspension a 50-ml sample was fil- tered onto a precombusted (550 ~ C, 2 h), preweighed (to the nearest 0.1 rag), glass fiber filter (Whatmans GF/C). The filter was then dried
(60 ~ C, 24 h) and weighed. In late June we noticed dense growth of a filamentous green algae, later determined as Cladophora sp., in tadpole treatments without L. stagnalis. To quantify differences between treatments the algae were removed on 10 July with a hand net and returned to the laboratory, dried (60 ~ C, 24 h) and weighed.
The data was log transformed and analysed with a randomized block design ANOVA, each block consisting of a raft of buckets. Differences between treatments were determined a posteriori using a Tukey test. The change in cumulative egg production over time was determined with repeated measures ANOVA with post hoc contrasts (SYSTAT MGLH: Wilkinson 1987).
Results
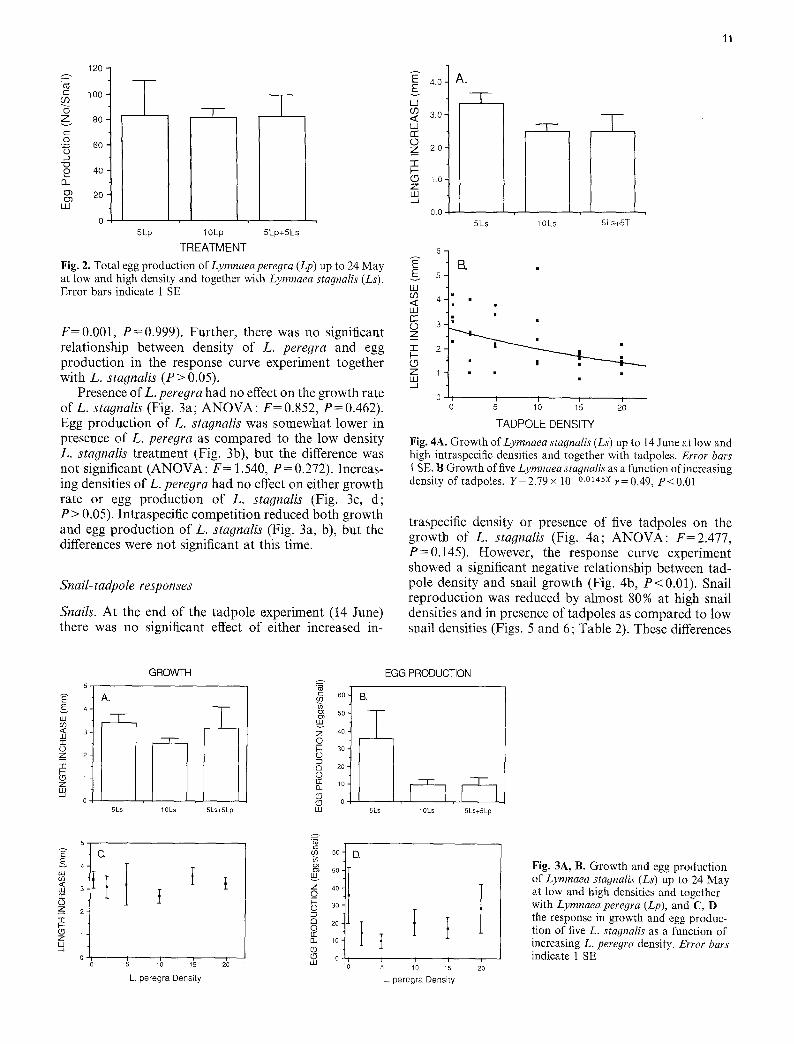
Periphyton responses
Dry weight of periphyton was reduced in the presence of snails and/or tadpoles (Fig. 1; ANOVA: F=13.357, P<0.001). Already at low snail or tadpole densities (5 per container) there was a > 60 % reduction ofperiphy- ton dry weight as compared to controls, and at higher grazing pressures the dry weight decreased to levels close to the detection limit of the method used. However, there was no significant difference between treatments with grazers (Tukey: P>0.05), whereas controls without ei- ther snails or tadpoles had a significantly higher periphy- ton biomass than any of the grazer treatments (Tukey: P<0.05).
Snail-snail responses
Due to massive mortality of L. peregra (95%) in late May, the treatments including this species were ter- minated on 24 May. The growth of Lymnaea peregra over the experimental period was not affected by inter- specific or intraspecific competition. The increase in total length was very small for those individuals that had died by 24 May (mean total length• mm, range = 0-1.0 ram, n=317). Mortality was independent of treatment. The few surviving snails showed a large increase in total length (mean• ram, range= 1.0-5.5 ram, n= 18). L. peregra egg production was not affected by a doubling of intraspecific density or by competition from Lymnaea stagnalis (Fig. 2; ANOVA:
I
oE
k - "1-
(3
h- a
5T ~OT 5Ls lOLs 5+2T 5+5T 5 + l O T 5 + l S T 5 + 2 0 T
T R E A T M E N T
Fig. 1. Periphyton biomass on plastic flagging tape in the different treatments on 25 May. Cont=control (no grazers), T = tadpoles, Ls = Lymnaea stagnalis. Error bars indicate 1 SE
120
c 1 0 0 co -6 Z 80
g �9 ~ 60
0 4 0 -
13.- �9
~ 2 0 -
t_U
0
5Lp 10Lp 5Lp+5Ls
TREATMENT
Fig. 2. Total egg production of Lymnaea peregra (Lp) up to 24 May at low and high density and together with Lymnaea stagnalis (Ls). Error bars indicate 1 SE
F= 0.001, P = 0.999). Further, there was no significant relationship between density of L. peregra and egg production in the response curve experiment together with L. stagnaIis (P>0.05).
Presence ofL. peregra had no effect on the growth rate of L. stagnalis (Fig. 3a; ANOVA: F = 0.852, P = 0.462). Egg production of L. stagnalis was somewhat lower in presence of L. peregra as compared to the low density L. stagnalis treatment (Fig. 3b), but the difference was not significant (ANOVA: F= 1.540, P = 0.272). Increas- ing densities of L. peregra had no effect on either growth rate or egg production of L. stagnalis (Fig. 3c, d; P > 0.05). Intraspecific competition reduced both growth and egg production of L. stagnalis (Fig. 3a, b), but the differences were not significant at this time.
Snail-tadpole responses
Snails. At the end of the tadpole experiment (14 June) there was no significant effect of either increased in-
11
E E v LIJ CO < UJ r r O z 7-
(_9 Z UJ .U
4.0 A .
T 3.0
2.0
1.0
0.0
5Ls 10Ls 5Ls+5T
6
E E 5
LU 09 <: 4 W r r 0 3
i 2
Z 1 LU J
0
B.
I I I I I 5 10 15 20
TADPOLE DENSITY
Fig. 4A. Growth ofLymnaea stagnalis (Ls) up to 14 June at low and high intraspecific densities and together with tadpoles. Error bars 1 SE. B Growth of five Lymnaea stagnalis as a function of increasing density of tadpoles. Y=2.79 x 10 -e.o145x r=0.49, P<0.01
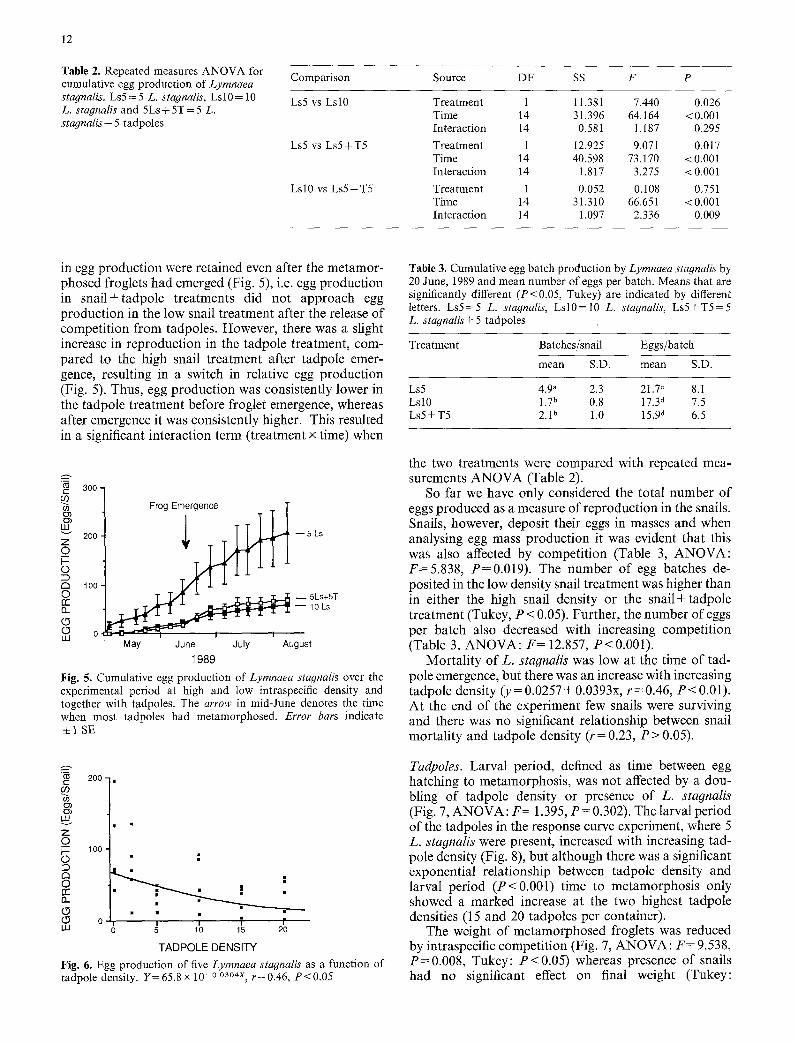
traspecific density or presence of five tadpoles on the growth of L. stagnalis (Fig. 4a; ANOVA: F=2.477, P=0.145). However, the response curve experiment showed a significant negative relationship between tad- pole density and snail growth (Fig. 4b, P<0.01). Snail reproduction was reduced by almost 80% at high snail densities and in presence of tadpoles as compared to low snail densities (Figs. 5 and 6; Table 2). These differences
GROWTH 5-
E 4 -
kU
< 3 kU rr"
0 Z 2
X2
Z
0
& LL/ O9 < LLI n- O z I
(.9 Z
A .
- F -
I
5Ls 10Ls
q--
5Ls+5Lp
Q
' , ' 4 2' L. p e r e g r a D e n s i t y
60
50 uJ Z 40 _o F- 30 O
tm 20 O rr 10 n
0 LU
09
W v Z _o I - O
O rr n r& (.9 LU
60
50
40
3 0
20'
10'
0
EGG PRODUCTION
B,
T
5Ls 10Ls 5Ls~-5Lp
t L j J j 5 10 15 20
L. peregra Density
Fig. 3A, B. Growth and egg production of Lymnaea stagnalis (Ls) up to 24 May at low and high densities and together with Lyrnnaea peregra (Lp), and C, D the response in growth and egg produc- tion of five L. stagnalis as a function of increasing L. peregra density. Error bars indicate 1 SE
12
Table 2. Repeated measures ANOVA for cumulative egg production of Lymnaea stagnalis. L s 5 = 5 L. stagnalis, L s l 0 = 10 L. stagnalis and 5Ls + 5T = 5 L. stagnalis+ 5 tadpoles
Comparison Source D F SS F P
Ls5 vs Lsl0 Treatment 1 11.381 7.440 0.026 Time 14 31.396 64.164 <0.001 Interaction 14 0.581 1.187 0.295
Ls5 vs L s 5 § Treatment 1 12.925 9.071 0.017 Time 14 40.598 73.170 <0.001 Interaction 14 1.817 3.275 <0.001
Lsl0 vs L s 5 § Treatment 1 0.052 0.108 0.751 Time 14 31.310 66.651 <0.001 Interaction 14 1.097 2.336 0.009
in egg production were retained even after the metamor- phosed froglets had emerged (Fig. 5), i.e. egg production in snail+tadpole treatments did not approach egg production in the low snail treatment after the release of competition from tadpoles. However, there was a slight increase in reproduction in the tadpole treatment, com- pared to the high snail treatment after tadpole emer- gence, resulting in a switch in relative egg production (Fig. 5). Thus, egg production was consistently lower in the tadpole treatment before froglet emergence, whereas after emergence it was consistently higher. This resulted in a significant interaction term (treatment x time) when
3O0 c- O9
o3 {3") W v 200 z o
O
~3 100 0 r r O_ (5 (5 W
Frog Emergence _ T I l ~ - - 5 Ls
- - 5Ls+5T - - 10Ls
May June July AugList
1989
Fig. 5. Cumulative egg production of Lymnaea stagnalis over the experimental period at high and low intraspecific density and together with tadpoles. The arrow in mid-June denotes the time when most tadpoles had metamorphosed. Error bars indicate i l SE
2OO c O9 O3 {33
z _o b- 100 O cl �9 o_ (5 (9 0 W
m I
0 g 1'0 1~ 2r0
TADPOLE DENSITY
Fig. 6. Egg production of five Lymnaea stagnalis as a function of tadpole density. Y= 65.8 x 10 -0.o3o4x, r = 0.46, P < 0 . 0 5
Table 3. Cumulative egg batch production by Lymnaea stagnalis by 20 June, 1989 and mean number of eggs per batch. Means that are significantly different ( P < 0.05, Tukey) are indicated by different letters. L s 5 = 5 L. stagnalis, L s l 0 = 1 0 L. stagnalis, L s 5 + T 5 = 5 L. stagnalis + 5 tadpoles
Treatment Batches/snail Eggs/batch
mean S.D. mean S.D.
Ls5 4.9" 2.3 21.7 c 8.1 Lsl0 1.7 b 0.8 17.3 d 7.5 Ls5+T5 2.l 6 1.0 15.9 a 6.5
the two treatments were compared with repeated mea- surements ANOVA (Table 2).
So far we have only considered the total number of eggs produced as a measure of reproduction in the snails. Snails, however, deposit their eggs in masses and when analysing egg mass production it was evident that this was also affected by competition (Table 3, ANOVA: F=5.838, P=0.019). The number of egg batches de- posited in the low density snail treatment was higher than in either the high snail density or the snail+ tadpole treatment (Tukey, P < 0.05). Further, the number of eggs per batch also decreased with increasing competition (Table 3, ANOVA: F = 12.857, P<0.001).
Mortality of L. stagnalis was low at the time of tad- pole emergence, but there was an increase with increasing tadpole density (y = 0.0257 + 0.0393x, r = 0.46, P < 0.01). At the end of the experiment few snails were surviving and there was no significant relationship between snail mortality and tadpole density (r= 0.23, P > 0.05).
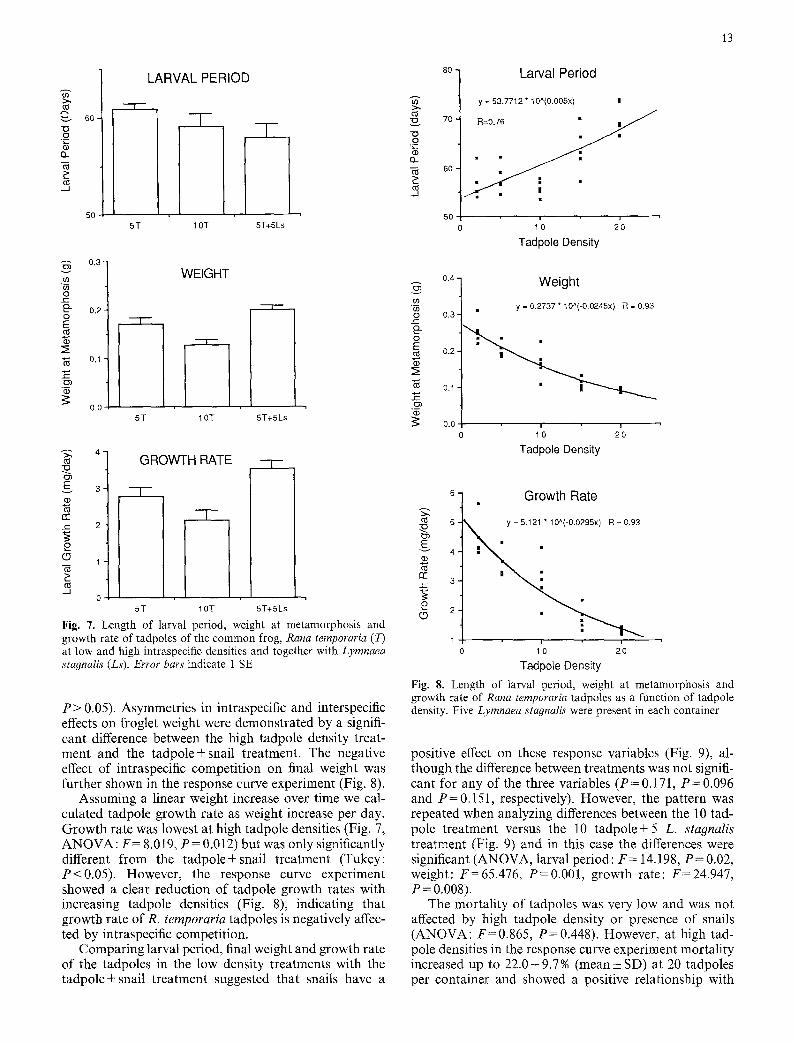
Tadpoles. Larval period, defined as time between egg hatching to metamorphosis, was not affected by a dou- bling of tadpole density or presence of L. stagnalis (Fig. 7, ANOVA: F = 1.395, P = 0.302). The larval period of the tadpoles in the response curve experiment, where 5 L. stagnalis were present, increased with increasing tad- pole density (Fig. 8), but although there was a significant exponential relationship between tadpole density and larval period (P<0.001) time to metamorphosis only showed a marked increase at the two highest tadpole densities (15 and 20 tadpoles per container).
The weight of metamorphosed froglets was reduced by intraspecific competition (Fig. 7, ANOVA: F = 9.538, P = 0.008, Tukey: P < 0.05) whereas presence of snails had no significant effect on final weight (Tukey:
60 "O O Q) n
d
50 5T
LARVAL PERIOD
-T-
1 0T 5T+5Ls
0 . 3 -
(,3
O
O- 0.2- o E c6 .,$
-,~ 0.1 E 03
0.0
- 1 - -
WEIGHT
5T 10Y 5T+5Ls
4
f O') g 3 (3)
rc t- 2 -
O
6J .--I
0 5T 1 0T 5T+5Ls
Fig. 7. Length of larval period, weight at metamorphosis and growth rate of tadpoles of the common frog, Rana temporaria (7) at low and high intraspecific densities and together with Lymnaea stagnaIis (Ls). Error bars indicate 1 SE
GROWTH RATE
T
T
P > 0.05). Asymmetries in intraspecific and interspecific effects on froglet weight were demonstrated by a signifi- cant difference between the high tadpole density treat- ment and the tadpole+ snail treatment. The negative effect of intraspecific competition on final weight was further shown in the response curve experiment (Fig. 8).
Assuming a linear weight increase over time we cal- culated tadpole growth rate as weight increase per day. Growth rate was lowest at high tadpole densities (Fig. 7, ANOVA: F = 8.019, P = 0.012) but was only significantly different from the tadpole+snail treatment (Tukey: P<0.05) . However, the response curve experiment showed a clear reduction of tadpole growth rates with increasing tadpole densities (Fig. 8), indicating that growth rate of R. temporaria tadpoles is negatively affec- ted by intraspecific competition.
Comparing larval period, final weight and growth rate of the tadpoles in the low density treatments with the tadpole+ snail treatment suggested that snails have a
O
13..
t~ _3
q3
r-
O E t~
133
80
70
6 0 ,
50
Larval Period
y = 53.7712 * 10^(0.005x) m
R=0.76 �9
10 20
Tadpole Density
13
0.4] Weight
y = 0.2737 * 10^(-0.0245x) R = 0.93 0,3 �9
0.2
0.1
0.0 n 2~0 0 10
Tadpole Density
e~ 5
~yJ E
v 4
r r 3 t-
O 2
Growth Rate
~ I y = 5.121.* 10^(-0.0295x) R = 0.93
i
1 0 2~0
Tadpole Density Fig. 8. Length of larval period, weight at metamorphosis and growth rate of Rana temporaria tadpoles as a function of tadpole density. Five Lymnaea staonalis were present in each container
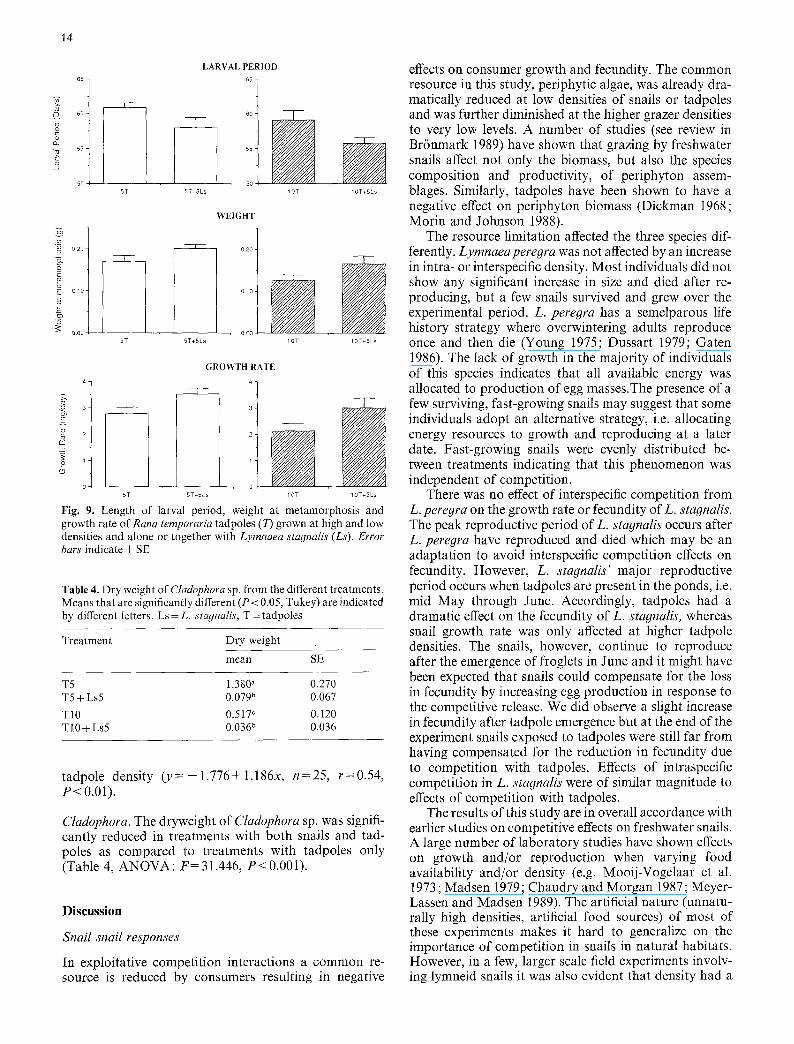
positive effect on these response variables (Fig. 9), al- though the difference between treatments was not signifi- cant for any of the three variables (P=0.171, P=0.096 and P = 0.151, respectively). However, the pattern was repeated when analyzing differences between the 10 tad- pole treatment versus the 10 tadpole+5 L. stagnalis treatment (Fig. 9) and in this case the differences were significant (ANOVA, larval period: F = 14.198, P = 0.02, weight: F = 65.476, P = 0.001, growth rate: F = 24.947, P = 0 . 0 0 8 ) .
The mortality of tadpoles was very low and was not affected by high tadpole density or presence of snails (ANOVA: F=0.865, P=0.448). However, at high tad- pole densities in the response curve experiment mortality increased up to 22.0• 9.7% (mean• SD) at 20 tadpoles per container and showed a positive relationship with
14
v
u~
E
4 -
E
�9
0
5T
-7-
5T+5Ls
LARVAL PERIOD 65
55
5O 10T
W E I G H T
IOT+5Ls
5T 5T+5Ls
0.20
0.10
000 lOT
# IOT+5Ls
GROWTH RATE 4 ,
5Y+5Ls ~ OT IOT+5Ls
Fig. 9. Length of larval period, weight at metamorphosis and growth rate of Rana temporaria tadpoles (7) grown at high and low densities and alone or together with Lymnaea stagnalis (Ls). Error bars indicate 1 SE
Table 4. Dry weight of Cladophora sp. from the different treatments. Means that are significantly different (P < 0.05, Tukey) are indicated by different letters. Ls = L. stagnalis, T = tadpoles
Treatment Dry weight
mean SE
T5 1.380 a 0.270 T5 + Ls5 0.079 b 0.067
T10 0.517 c 0.120 T I 0 + L s 5 0.0368 0.036
tadpole density (y=-1 .776+1 .186x , n=25, r=0.54, P<0.01).
Cladophora. The dryweight of Cladophora sp. was signifi- cantly reduced in treatments with both snails and tad- poles as compared to treatments with tadpoles only (Table 4, ANOVA: F = 31.446, P < 0.001).
Discussion
Snail-snail responses
In exploitative competition interactions a common re- source is reduced by consumers resulting in negative
effects on consumer growth and fecundity. The common resource in this study, periphytic algae, was already dra- matically reduced at low densities of snails or tadpoles and was further diminished at the higher grazer densities to very low levels. A number of studies (see review in Br6nmark 1989) have shown that grazing by freshwater snails affect not only the biomass, but also the species composition and productivity, of periphyton assem- blages. Similarly, tadpoles have been shown to have a negative effect on periphyton biomass (Dickman 1968; Morin and Johnson 1988).
The resource limitation affected the three species dif- ferently. Lymnaeaperegra was not affected by an increase in intra- or interspecific density. Most individuals did not show any significant increase in size and died after re- producing, but a few snails survived and grew over the experimental period. L. peregra has a semelparous life history strategy where overwintering adults reproduce once and then die (Young 1975; Dussart 1979; Gaten 1986). The lack of growth in the majority of individuals of this species indicates that all available energy was allocated to production of egg masses.The presence of a few surviving, fast-growing snails may suggest that some individuals adopt an alternative strategy, i.e. allocating energy resources to growth and reproducing at a later date. Fast-growing snails were evenly distributed be- tween treatments indicating that this phenomenon was independent of competition.
There was no effect of interspecific competition from L. peregra on the growth rate or fecundity of L. stagnalis. The peak reproductive period of L. stagnalis occurs after L. peregra have reproduced and died which may be an adaptation to avoid interspecific competition effects on fecundity. However, L. stagnalis' major reproductive period occurs when tadpoles are present in the ponds, i.e. mid May through June. Accordingly, tadpoles had a dramatic effect on the fecundity of L. stagnalis, whereas snail growth rate was only affected at higher tadpole densities. The snails, however, continue to reproduce after the emergence of froglets in June and it might have been expected that snails could compensate for the loss in fecundity by increasing egg production in response to the competitive release. We did observe a slight increase in fecundity after tadpole emergence but at the end of the experiment snails exposed to tadpoles were still far from having compensated for the reduction in fecundity due to competition with tadpoles. Effects of intraspecific competition in L. stagnalis were of similar magnitude to effects of competition with tadpoles.
The results of this study are in overall accordance with earlier studies on competitive effects on freshwater snails. A large number of laboratory studies have shown effects on growth and/or reproduction when varying food availability and/or density (e.g. Mooij-Vogelaar et al. 1973; Madsen 1979; Chaudry and Morgan 1987; Meyer- Lassen and Madsen 1989). The artificial nature (unnatu- rally high densities, artificial food sources) of most of these experiments makes it hard to generalize on the importance of competition in snails in natural habitats. However, in a few, larger scale field experiments involv- ing lymneid snails it was also evident that density had a

15
negative effect on snail growth and reproduction (Eisen- berg 1966, 1970; Brown 1979, 1982; Brown et al. 1985). Both Eisenberg (1966, 1970) and Brown (1979) found that density dependent effects on fecundity were greatest, whereas adult growth rate was not greatly affected. This agrees with the results of our study. Osenberg (1989), on the other hand, argued that the increased densities of snails found after an increase in resource levels were due to an increase in juvenile survival whereas changes in adult fecundities were small. These studies all argue that increased density has an effect by increasing exploitative competition for food, and additions of high quality food (spinach; Eisenberg 1966, 1970) or fertilization to in- crease algal production (Osenberg 1989) increased growth rate and, especially, fecundity. Further, regional surveys and transplantation experiments have shown that growth and fecundity are correlated with habitat productivity (Russel-Hunter 1961; Brown et al. 1985; Byrne et al. 1989; Lam and Calow 1989).
Observations from laboratory studies have suggested that besides exploitative competition, interference com- petition can be an important factor in interactions be- tween freshwater snails. Madsen (1979) found that the reduced fecundity of BiomphaIaria camerunensis when competing with Helisoma duryi was due to predation on its egg masses by the latter species. We did not find any signs of damaged egg masses in our experiments. Fur- ther, the reduction in fecundity was not only a result of a decrease in the number of egg masses produced, but there was also a reduction in the number of eggs per egg mass, indicating that the fecundity effects were due to a decrease in availability of energy. A similar pattern was found by Eisenberg (1966).
Snail-tadpole responses
Competition for food between distantly related taxa is less well studied than competition between closely related species. Periphytic algae constitute a high quality food source for several groups of aquatic organisms such as snails, crustaceans, insect larvae and tadpoles, and stud- ies on each of these groups have shown that they can reduce periphyton biomass leading to exploitative com- petition (e.g. Br6nmark 1989; Morin and Johnson 1988 ; Hart 1987). As these groups of organisms often coexist in aquatic habitats one would expect that competitive interactions between groups could be an important re- gulating factor. Cuker (1983) and Hawkins and Furnish (1987) showed that presence of snails had a negative effect on the density of periphyton-grazing insect larvae; snails influencing insects through a combination of re- source depression and interference (larval displacement). Morin et al. (1988) studied the effects of competition between toad and tree frog tadpoles and aquatic insect larvae and found that insects reduced mass at metamor- phosis of both tree frogs and toads, probably through exploitative competition for periphyton. The effects of competition between insects and tree Dog tadpoles were as strong as between toad and tree frog tadpoles. Com- petition between tadpoles and snails in this study had a
dramatic negative effect on snail fecundity, but affected snail growth rate to a much lesser degree. The com- petitive relationship was asymmetric, however, as tad- poles were not negatively affected by increased densities of snails. Reviews of interspecific competition have shown that competitive effects are often asymmetric (Lawton and Hassell 1981; Schoener 1983; Connell 1983) and a large number of studies on competition between anuran tadpoles have demonstrated asymmetric effects (see review in Morin and Johnson 1988). For example, Morin and Johnson (1988) studied competition between tadpoles of two North American anurans, Rana sylvatica and Hyla crucifer, and found that Rana had negative effects on all of the response variables measured for Hyla, but showed no adverse effect from the presence of interspecific competitors. The competitive asymmetry was due to exploitative competition, Rana having a greater per capita impact on the biomass of periphyton. Other studies have indicated that chemical substances (growth inhibitors) are responsible for asymmetries in anuran competition (e.g. Steinwascher 1978), but Morin and Johnson (1988) found no evidence for interference via growth inhibitors.
In this study, snails had no negative effect on tadpoles, in fact, on the contrary, the presence of snails facilitated tadpole growth and reduced the length of the larval period. We believe that this is a special case of "indirect mutualism" as described by Vandermeer (1980). He predicted that in a situation where two resources com- pete, a consumer specialized on one of the resources should be positively affected by a consumer specialized on the other resource. Dethier and Duggins (1984) ex- tended this theory to situations with asymmetric com- petition between resources and generalist consumers without food preference, resulting in "indirect commen- salism". In a study of intertidal snails they found that grazing by a large chiton favored two smaller limpet species by removing macroalgal cover. The macroalgae outcompeted microalgae and monopolized the sub- stratum and this decreased available substrate and food resources for the limpets. We propose that a similar mechanism, summarized in Fig. 10, may operate in our system. In this case microalgae (e.g. diatoms, solitary greens) compete with a filamentous green algae, Clado- phora sp., for phosphorus, which is a limiting resource
m
TADPOLE . . . . . . . . "~"SNA 1 L
MICRO-ALGAE ~ ~ CLADOPHORA
\ / NUTRIENTS
Fig. 10. Summary of interactions between snails, tadpoles, algae and nutrients. Arrows between trophic levels denote consumption, whereas broken arrows denote competitive (negative sign) or facili- tative (positive sign) interactions. See text for further explanation

J6
for algal growth during summer in the pond we studied (L.-A. Hansson, pers.comm.). The filamentous algae are competitively dominant over microalgae due to rapid growth early in the growth season (Cheney and Hough 1983) and accumulation of phosphorus in the biomass, reducing its availability in the open water (Confer 1972). Tadpoles are specialist grazers on microalgae (either periphytic or planktonic) that they trap in gill filters and a mucus-entrapment mechanism (Kenny 1969). They can feed on young Cladophora but older filaments that have developed thick cell walls are avoided (Savage 1952). Savage (1952) found that Rana ternporaria tadpoles failed to grow in aquaria with a luxuriant but old stand- ing crop of Cladophora. Lymneid snails, on the other hand, are more generalist, feeding on periphyton, decay- ing macrophytes and detritus (Scheerbom and Van Elk 1978; Reavell 1980). However, they exhibit a clear preference for non-filamentous green algae, whereas fil- amentous greens such as Cladophora are a non-preferred food item (Calow 1970; Calow and Calow 1975; Skoog 1978). Growth and reproduction can be sustained on a diet of Cladophora but is greatly reduced in comparison with when fed with more high-ranked food items (Skoog 1978). This study suggests that R. temporaria tadpoles have a competitive edge in the interaction with snails, which may be due to higher degree of resource depression obtained by high grazing rates (S.D. Rundle, unpublish- ed), continuous feeding (Savage 1952), high food passage rates (Wassersug 1975), and also because of their ability to switch to another high quality food source, planktonic algae (Kenny 1969; Steinwascher 1978). In the presence of tadpoles snails will be forced to feed on Cladophora, a low-quality food item, resulting in a reduction of growth and fecundity. In turn, grazing on Cladophora will increase phosphorus availability, due to depletion of the filamentous algae and excretion by snails (Confer 1972), which will promote productivity of microalgae, especially periphyton (Hansson 1988), increasing tadpole food resources. Thus, tadpoles have a negative com- petitive effect on snails through resource depression, whereas snails have an indirect facilitative effect on tad- poles through an increase of food availability. Wilbur (1987) similarly found that presence of Rana utricularia tadpoles had a strong positive effect on the growth rate of Scaphiopus holbrooki tadpoles. Rana grazing on periphytic algae was suggested to enhance phytoplank- ton production by increasing nutrient concentration. Suspended phytoplankton is preferred by Scaphiopus, whereas periphyton is a low-quality food resource (Wil- bur et al. 1983). Further, Hyla chrysoscelis tadpoles had a higher growth rate and larger metamorph size in young temporary ponds as compared to older ponds where nutrients were tied up in macrophytes and filamentous algae (Wilbur and Alford 1985), which are a lower qual- ity food resource than phytoplankton and periphyton (Seale 1980).
Conclusions
In conclusion, grazing by snails and tadpoles reduced the availability of their common food source, periphytic algae. However, although periphyton was a limiting re-
source the interactions between these three grazers were not simple competitive as predicted. Coexistence of Lym~ naea stagnalis and L. peregra did not result in any nega- tive effects on growth or reproduction. Both species showed little growth during the experiment, i.e. all avail- able energy resources may have been allocated to re- production. The absence of an intra- or interspecific density effect on L. peregra reproduction may indicate that this species used energy reserves, stored before the start of the experiment, for reproduction. The major peak of reproduction in L. stagnaIis occurred after the disappearance of L. peregra, which could be seen as a temporal segregation to avoid interspecific competition during the reproductive period. However, interspecific competition may be of great importance for the juvenile snails during summer when densities of both species are high and most of the growth occurs.
The interactions between snails and tadpoles were more complex involving both competition and facilita- tion, possibly mediated by competitive interactions be- tween different components of their food resource, mi- croalgae and Cladophora. Tadpoles had a clear negative effect on snail reproduction, whereas snails had a positive effect on tadpole growth. This provides us with a par- adox. How can snails and tadpoles coexist in small ponds if the interactions between them are as strong as in- dicated by this study? We predicted that in these small, but permanent, ponds, predation should not be an im- portant regulating factor, because large predators such as fish and crayfish are absent. However, invertebrate predators such as odonate and dytiscid larvae are vora- cious predators on tadpole larvae and are present in these ponds. Several studies have shown that predation by invertebrate predators can dramatically reduce densities of tadpoles (e.g. Smith 1983; Woodward 1983; Crump 1984) and, thus, tadpole densities in ponds with abun- dant invertebrate predators may never reach levels where competition effects on snails are important (cf. Calef 1973; Heyer et al. 1975; Wilbur 1984). Studies involving newts and tadpoles have shown that predation by newts may change the competitive relationships between tad- poles of different species (e.g. Morin 1983; Wilbur et al. 1983; Wilbur 1987; Wilbur and Fauth 1990). Snails on the other hand are not as vulnerable to invertebrate predators (Lodge et al. 1987). For example, although glossiphonid leeches prey upon snails, feeding rates are so low, partly due to anti-predator adaptations in the snails, that leech predation will never limit snail popula- tions (Br6nmark and Malmqvist 1986; Br6nmark, un- published). Rana temporaria uses a broad range of fresh- water habitats as spawning sites, from lakes to small ephemeral ditches (Cooke 1975; Haapanen 1982), but there seems to be a preference for shallow, temporary ponds (Cooke 1975), which may further suggest the im- portance of invertebrate predators. Thus, although this study shows the potential for strong interactions, both competitive and facilitative, predation by invertebrate predators may mediate interaction strength between snails and tadpoles in small, permanent ponds. Acknowledgements. Victor and Oscar Br6nmark enthusiastically helped us with the frog measurements. The manuscript benefitted from comments by K.M. Brown. S.D. Rundle was supported by a

17
postdoctoral fellowship from the Royal Society, Great Britain during this study. Financial support was further received from the Swedish Natural Science Foundation and the Craaford Founda- tion.
References
Berrie A (1965) On the life cycle of Lymnaea stagnalis (L.) in the west of Scotland. Proc Malac Soc Lond 36:283-295
Berven KA (1981) Mate choice in the wood frog, Rana sylvatica. Evolution 35:707 722
Berven KA (1982) The genetical basis of altitudinal variation in the wood frog Rana sylvatica. I. An experimental analysis of life history traits. Evolution 36:962-983
Berven KA, Gill DE (1983) Interpreting geographic variation in life history traits. Am Zool 23:85-97
Bondesen P (1950) A comparative morphological-biological analy- sis of the egg capsules of freshwater pulmonate Gastropods. Natnra Jutlandica 3 : 7-208
Br6nmark C (1985) Freshwater snail diversity: effects of pond area, habitat heterogeneity and isolation. Oecologia 67 : 127-131
Br6nmark C (1989) Interactions between epiphytes, macrophytes and freshwater snails: a review. J Moll Stud 55:299-311
Br6nmark C, Malmqvist B (1986) Interactions between the leech Glossiphonia complanata and its gastropod prey. Oecologia 69 : 268-276
Brown KM (1979) Effects of experimental manipulations on the life history pattern of Lymnaea stagnalis appressa Say (Pulmonata: Lymnaeidae). Hydrobiologia 65:165-176
Brown KM (1982) Resource overlap and competition in pond snails: an experimental analysis. Ecology 63:412-422
Brown KM, DeVries DR, Leathers BK (1985) Causes of life history variation in the freshwater snail Lymnaea elodes. Malacologia 26 : 191-200
Byrne RA, Reynolds JD, McMahon RF (1989) Shell growth, re- production and life cycles of Lymnaea peregra and L. palustris (Pulmonata : Basommatophora) in oligotrophic turloughs (tem- porary lakes) in Ireland. J Zool 217:321-339
Calef GW (1973) Natural mortality of tadpoles in a population of Rana aurora. Ecology 54:741-758
Calow P (1970) Studies on the natural diet of Lymnaea pereger obtusa (Kobelt) and its possible ecological implications. Proc Malacol Soc Lond 39:203-215
Calow P (1981) Adaptational aspects of growth and reproduction in Lymnaea peregra (Gastropoda: Pulmonata) from exposed and sheltered aquatic habitats. Malacologia 21:5-13
Calow P, Calow LJ (1975) Cellulase activity and niche separation in freshwater gastropods. Nature 255:478-480
Chaudry AM, Morgan E (1987) Factors affecting the growth and fecundity of Bulinus tropicus (Krauss) (Gastropoda). J Moll Stud 53:52-61
Cheney C, Hough RA (1983) Factors controlling photosyn- thetic productivity in a population of Cladophora fracta (Chlorophyta) Ecology 64:68-77
Confer JL (1972) Interrelations among plankton, attached algae, and the phosphorus cycle in artificial open systems. Ecol Mo- nogr 42 : 1-23
Connell JH (1983) On the prevalence and relative importance of interspecific competition: evidence from field experiments. Am Nat 122:661-696
Cooke AS (1975) Spawn site selection and colony size of the frog (Rana temporaria) and the toad (Bufo bufo). J Zool 175:29-38
Crump ML (1984) Ontogenetic changes in vulnerability to pre- dation in tadpoles of Hyla pseudopurna. Herpetologica 40:265 271
Cuker BE (1983) Competition and coexistence among the grazing snail Lymnaea, Chironomidae, and microcrustacea in an arctic epilithic lacustrine community. Ecology 64:10-15
Cummins CP (1989) Interaction between the effects of pH and density on growth and development in Rana temporaria L. tadpoles. Funct Ecol 3 : 45-52
Dethier MN, Duggins DO (1984) An "indirect commensalism"
between marine herbivores and the importance of competitive hierarchies. Am Nat 124:205-219
Dickman M (1968) The effect of grazing by tadpoles on the struc- ture of a periphyton community. Ecology 49:1188-1190
Dussart GBJ (1979) Life cycles and distribution of the aquatic gastropod molluscs Bithynia tentaculata (L.), Gyraulus albus (Muller), Planorbisplanorbis (L.) and Lymnaeaperegra (Muller) in relation to water chemistry. Hydrobiologia 67:223-239
Eisenberg RM (1966) The regulation of density in a natural popula- tion of the pond snail, Lymnaea elodes. Ecology 47 : 889-906
Eisenberg RM (1970) The role of food in the regulation of the pond snail, Lymnaea elodes. Ecology 51:680-684
Fretwell SD (1987) Food chain dynamics: the central theory of ecology? Oikos 50:291-301
Gaten E (1986) Life cycle of Lymnaea peregra (Gastropoda: Pul- monata) in the Leicester canal, U.K., with an estimate of annual production. Hydrobiologia 135:45 54
Gibbons MM, McCarthy TK (1986) The reproductive output of frogs Rana temporaria (L.) with particular reference to body size and age. J Zool 209:579-293
Haapanen A (1982) Breeding of the common frog (Rana tem- poraria). Ann Zool Fennici 19:75-79
Hairston N, Smith FE, Slobodkin L (1960) Community structure, population control, and competition. Am Nat 94:421-425
Hansson L-A (1988) Effects of competitive interactions on the biomass development of planktonic and periphytic algae in lakes. Limnol Oceanogr 33 : 121-128
Hart DD (1987) Experimental studies of exploitative competition in a grazing stream insect. Oecologia 73:41M7
Hawkins CP, Furnish JK (1987) Are snails important competitors in stream ecosystems? Oikos 49:209-220
Heyer WR, McDiarmid RW, Weigmann DL (1975) Tadpoles, predation and pond habitats in the tropic. Biotropica 7:100-111
Howard RD (1980) Mating behavior and mating success in wood- frogs, Rana sylvatica. Anim Behav 28:705-716
Kenny JS (1969) Feeding mechanisms in anuran larvae. J Zool 157:225-246
Lam PKS, Calow P (1989) Intraspecific life-history variation in Lymnaea peregra (Gastropoda: Pulmonata). I. Field study. J Anim Ecol 58:571-588
Lawton JH, Hassell MP (1981) Asymmetrical competition in in- sects. Nature 289 : 793-795
Lodge DM, Brown KM, Klosiewski SP, Stein RA, Covich AP, Leathers BK, Br6nmark C (1987) Distribution of freshwater snails: spatial scale and the relative importance of physico- chemical and biotic factors, Am Malac Bull 5:73-84
Madsen H (1979) Further laboratory studies on the interspecific competition between Helisoma duryi (Wetherby) and the inter- mediate hosts of Schistosoma mansoni Sambon: Biomphalaria alexandrina (Ehrenberg) and B. camerunensis (Boettger). Hy- drobiologia 66:181-192
Meyer-Lassen J, Madsen H (1989) The effect of varying relative density and varying food supply on interspecific competition between Helisoma duryi and Bulinus truncatus (Gastropoda, Planorbidae). J Moll Stud 55:89-96
Mooij-Vogelaar JW, Jager JC, Van Der Steen WJ (1973) Effects of density levels, and changes in density levels on reproduction, feeding and growth in the pond snail Lymnaea stagnalis (L.). Proc Ned Akad Wet Ser C 76:245-256
Morin PJ (1983) Predation, competition, and the composition of larval anuran guilds. Ecol Monogr 53:119-138
Morin P J, Johnson EA (1988) Experimental studies of asymmetric competition among anurans. Oikos 53 : 398-407
Morin PJ, Lawler SP, Johnson EA (1988) Competition between aquatic insects and vertebrates : interaction strength and higher order interactions. Ecology 69 : 1401-1409
Osenberg CW (1989) Resource limitation, competition and the influence of life history in a freshwater snail community. Oeeolo- gia 79:512-519
Reavell PE (1980) A study of the diets of some British freshwater gastropods. J Conchol 30:253-271
Russell-Hunter W (1961) Life cycles of four freshwater snails in

18
limited populations in Loch Lomond, with a discussion on intraspecific variation. Proc Zool Soc Lond 137:135-171
Savage RM (1952) Ecological, physiological and anatomical ob- servations on some species of anuran tadpoles. Proc Zool Soc Lond 122: 467-514
Scheerborn JEM, Van Elk R (1978) Field observations on the seasonal variation in the natural diet and the haemolymph- glucose concentration of the pond snail Lymnaea stagnalis (L). Proc K Ned Akad Wet C. Biol Med Sci 81:365-376
Sehoener TW (1983) Field experiments on interspecific competition. Am Nat 122:240-285
Scale DB (1980) Influence of amphibian larvae on primary produc- tion, nutrient flux, and competition in a pond ecosystem. Ecol- ogy 61:1531-1550
Semlitseh RD (1985) Reproductive strategy of a facultative paedo- morphic salamander Ambystoma talpoideum. Oecologia 65:305-313
Semlitsch RD, Caldwell JP (1982) Effects of density on growth, metamorphosis, and survivorship in tadpoles of Scaphiopus holbrooki. Ecology 63:905-911
Semlitsch RD, Scott DE, Pechmann JHK (1988) Time and size at metamorphosis related to adult fitness in Ambystoma tal- poideum. Ecology 69:184-192
Skoog G (1978) Influence of natural food items on growth and egg production in brackish water populations of Lymnaea peregra and Theodoxusfluviatilis (Mollusca). Oikos 31:340-348
Smith DC (1983) Factors controlling tadpole populations of the chorus frog (Pseudacris triseriata) on Isle Royale, Michigan. Ecology 64:501-510
Smith DC (1987) Adult recruitment in chorus frogs: effects of size and date at metamorphosis. Ecology 68:344-350
Steinwascher K (1978) Interference and exploitation competition among tadpoles of Rana utricularia. Ecology 59:1039-1046
Travis J (1983) Variation in growth and survival of Hyla gratiosa larvae in experimental exclosures. Copeia 1983:232-237
Underwood AJ (1986) The analysis of competition by field experi- ments. In: Kikkawa J, Anderson DJ (eds) Community Ecology :
Pattern and Process. Blackwell Scientific Publications, Mel- bourne, pp 240-268
Vandermeer J (1980) Indirect mutualisms: variations on a theme by Stephen Levine. Am Nat 116:441-448
Wassersug R (1975) The adaptive significance of the tadpole stage with comments on the maintenance of complex life cycles in anurans. Amer Zo ol 15 : 405-417
Wilbur HM (1976) Density-dependent aspects of metamorphosis in Ambystoma and Rana syIvatiea. Ecology 57:1289-1296
Wilbur HM (1977) Interactions of food level and population den- sity in Rana sylvatiea. Ecology 58 : 206-209
Wilbur HM (1980) Complex life cycles. Ann Rev Ecol Syst 11 : 67-93
Wilbur HM (1984) Complex life cycles and community organiza- tion in amphibians. In: Price PW, Slobodchikoff CN, Gaud WS (eds) A new ecology: novel approaches to interactive systems. Wiley, New York, pp 195-224
Wilbur HM (1987) Regulation of structure in complex systems: experimental temporary pond communities. Ecology 68 : 1437-1452
Wilbur HM, Alford RA (1985) Priority effects in experimental pond communities: responses of Hyla to Bufo and Rana. Ecology 66:1106-1114
Wilbur HM, Fauth JE (1990) Experimental aquatic food webs: interactions between two predators and two prey. Am Nat 135:176-204
Wilbur HM, Morin PJ, Harris RN (1983) Salamander predation and the structure of experimental communities: anuran re- sponses. Ecology 64:1423-1429
Wilkinson L (1987) SYSTAT: The system for statistics. SYSTAT, Evanston, IL
Woodward BD (1983) Predator-prey interactions and breeding- pond use of temporary-pond species in a desert anuran community. Ecology 64:1549-1555
Young MR (1975) The life cycles of six species of freshwater Mol- luscs in the Worcester-Birmingham canal. Proc Malac Soc Lond 41 : 533-548





























