Embed Size (px)
Citation preview

BIODIVERSITY OF CORALLINE ALGAE IN THE NORTHEASTERN ATLANTICINCLUDING CORALLINA CAESPITOSA SP. NOV. (CORALLINOIDEAE, RHODOPHYTA)1
Rachel H. Walker, Juliet Brodie2, Stephen Russell, Linda M. Irvine
Department of Botany, Natural History Museum (BM), Cromwell Road, London, SW7 5BD, UK
and Sotiris Orfanidis
National Agricultural Research Foundation, Fisheries Research Institute, 64007 Nea Peramos, Kavala, Greece
The Corallinoideae (Corallinaceae) is representedin the northeastern Atlantic by Corallina officinalis L.;Corallina elongata J. Ellis et Sol.; Haliptilon squama-tum (L.) H. W. Johans., L. M. Irvine et A. M.Webster; and Jania rubens (L.) J. V. Lamour. Thedelimitation of these geniculate coralline red algaeis based primarily on morphological characters.Molecular analysis based on cox1 and 18S rRNAgene phylogenies supported the division of theCorallinoideae into the tribes Janieae and Corallin-eae. Within the Janieae, a sequence difference of46–48 bp (8.6%–8.9%) between specimens ofH. squamatum and J. rubens in the cox1 phylogenyleads us to conclude that they are congeneric.J. rubens var. rubens and J. rubens var. corniculata (L.)Yendo clustered together in both phylogenies, sug-gesting that for those genes, there was no geneticbasis for the morphological variation. Within theCorallineae, it appears that in some regions, thename C. elongata has been misapplied. C. officinalissamples formed two clusters that differed by45–54 bp (8.4%–10.0%), indicating species-leveldivergence, and morphological differences were suf-ficient to define two species. One of these clusterswas consistent with the morphology of the type spec-imen of C. officinalis (LINN 1293.9). The other spe-cies cluster is therefore described here as Corallinacaespitosa sp. nov. This study has demonstrated thatthere is a clear need for a revision of the genusCorallina to determine the extent of ‘‘pseudo-cryptic’’ diversity in this group of red algae.
Key index words: ‘‘pseudocryptic’’ diversity; 18SrRNA; Corallina caespitosa; Corallina officinalis;coralline algae; cox1; integrative taxonomy;morphology; red algae
Abbreviations: bp, base pairs; CI, consistencyindex; cox1, cytochrome c oxidase subunit I; RC,rescaled consistency index; RI, retention index
The Corallinales is a distinctive and diverse orderof algae, distributed in marine wave-exposed littoraland sublittoral habitats around the world (Johansen1976) and comprising both crustose and geniculate(articulated) forms. Ecologically, they are known toinhabit distinct zones in the littoral and sublittoraland provide niches for many other species (Cham-berlain and Cooke 1984, Kinzie and Buddemeier1996). With 564 currently accepted species, they areconsidered to be the third most species-rich orderwithin the red algae (Brodie and Zuccarello 2007)after the Ceramiales and Gigartinales. Taxonomicand phylogenetic relationships among taxa at alllevels remain largely unresolved (Johansen 1976,1981, Woelkerling 1988, Bailey 1999). Cabioch (1972,1988) included both geniculate and nongeniculatespecies within the subfamily Corallinoideae(Aresch.) Foslie, basing the classification on herinterpretation of their evolutionary relationships.Johansen (1981) and Woelkerling (1988) retainedthe geniculate genera in separate subfamilies forthe convenience of identification. Molecular studiesbased on SSU rRNA gene sequences (Bailey 1999,Bailey et al. 2004), did not fully reflect either classi-fication scheme and indicated that the presence orabsence of genicula was not a phylogenetically infor-mative character at the subfamily level. In this arti-cle, we describe our studies of the geniculatespecies from the northeastern Atlantic belonging tothe subfamily Corallinoideae within the Corallina-ceae.
Two tribes, the Corallineae Aulin and JanieaeH. W. Johans. et P. C. Silva, are recognized within theCorallinoideae. In the northeastern Atlantic, Irvineand Johansen (1994) recognized C. officinalis andC. elongata in the Corallineae, and H. squamatum andJ. rubens, which has two varieties, J. rubens var. rubensand J. rubens var. corniculata, in the Janieae. Kim et al.(2007), who reassessed the generic boundaries in theJanieae on the basis of morphological charactersand SSU rDNA sequence data, inferred that the tribeis monophyletic and proposed that Cheilosporum(Decne.) Zanardini and Haliptilon (Decne.) Lindl.were synonymous with Jania J. V. Lamour. However,given that data from the type of Jania were notincluded in their molecular analysis, we suggest that
1Received 28 March 2008. Accepted 21 August 2008.2Author for correspondence: email [email protected].
J. Phycol. 45, 287–297 (2009)� 2009 Phycological Society of AmericaDOI: 10.1111/j.1529-8817.2008.0637.x
287

further consideration of generic concepts in theJanieae is necessary.
Until the study by Kim et al. (2007), little sequencedata existed for species of the Corallinoideae. A studyby Robba et al. (2006), based on the mitochondrialcox1 gene, suggested that there were two geneticallydistinct species, which had both been morphologi-cally identified as C. officinalis. This finding highlightsthe difficulty of distinguishing species based on mor-phology alone. Current synonyms of C. officinalisinclude: C. calvadosii J. V. Lamour., C. nana Zanardi-ni, and C. compacta P. Crouan et H. Crouan.
The aim of this work was to study phylogeneticrelationships within the Corallinoideae in the north-eastern Atlantic and to determine intra- and inter-specific genetic variation. The study also includedsome material from the Mediterranean and themid-Atlantic to consider diversity within the contextof distributional ranges. An integrative taxonomicapproach included phylogeny reconstructions basedon the cox1 and 18S rRNA genes in addition to acomparative morphological assessment.
MATERIALS AND METHODS
Taxon sampling. In total, 32 specimens were sampled,including individuals of C. officinalis, C. elongata, H. squama-tum, J. rubens var. corniculata, and J. rubens var. rubens(Table S1 in the supplementary material). Fresh materialwas collected from Britain, Iceland, and Greece. Herbariumsamples were selected from BM collections (Table S1).Herbarium abbreviations follow Holmgren et al. (1990). Forsequencing the cox1 gene, 31 coralline specimens wereselected. From separate clades in the cox1 phylogram, 16specimens were then selected for sequencing the 18S rRNAgene.
DNA extraction, PCR amplification, and sequencing. DNA wasextracted from 0.5 cm2 of fresh, silica-gel-preserved, andherbarium material using a modified cetyl trimethyl ammo-nium bromide (CTAB) microextraction protocol (Rogerset al. 1994). Extracted DNA was purified, after isopropanolprecipitation using a GFX PCR DNA purification kit (GEHealthcare Ltd., UK) following the manufacturer’s protocols.The cox1 region was amplified using primers of Saunders(2005), and the 18S rRNA gene region was amplified usingprimers of Saunders and Kraft (1994). Each PCR runcontained 2.5 lL NH4 RXN buffer, 1.5 lL MgCl2, 0.5 lLTaq (all from BIOTAQ DNA Polymerase kit, Bioline Ltd.,UK), 0.5 lL dNTP stock, 1 lL 10 lM forward primer, 1 lL10 lM reverse primer, 17.5 lL H20, and 1 lL of DNAtemplate. The PCR reaction was run on a Techne ThermalCycler model FT Gene 5D (Fisher Scientific Ltd., Loughbor-ough, UK). A standard protocol of PCR (1 cycle at 94�C for2 min, 30 cycles at 94�C for 30 s, 30 cycles at 45�C for 30 s, 30cycles at 72�C for 1 min, 1 cycle at 72�C for 5 min) was usedfor cox1 and 18S rRNA. The successful samples were cleanedusing GFX PCR DNA purification kits, following the manu-facturer’s protocol. Dideoxy cycle sequencing was performed(1 cycle at 96�C for 5 min, 25 cycles at 96�C for 10 s, 25 cyclesat 50�C for 5 s, 25 cycles at 60�C for 4 min) with ABI BigDyeTerminators version 1.1 cycle sequencing kit (AppliedBiosystems, Foster City, CA, USA) in 10 lL volumes usingPCR primers (10 lM) on a Hybaid Omnigene Thermal Cycler(Hybaid Ltd., Cambridge, UK). Excess dye-labeled nucleotidesfrom the sequence reactions were removed by standardethanol-sodium acetate precipitation. Sequence products were
subsequently resuspended and run on a Perkin Elmer (UK)ABI 377 DNA sequencer.
Data analysis. Sequences were assembled and edited inSeqMan (DNAStar Lasergene Navigator package, Version 5.0)and aligned by Clustal W (Thompson et al. 1994) in MegAlign(DNAStar Lasergene Navigator package, Version 5.0, DNAS-TAR, USA). The alignments were edited by eye in MacClade(Maddison and Maddison 2005). Phylogenetic hypotheses wereinferred by maximum-parsimony (MP) and maximum-likeli-hood (ML) optimality criteria. The 38 cox1 sequence data setincluded three outgroup sequences: Chondrus crispus(DQ191341), Mastocarpus stellatus (DQ442899), and Gracilariagracilis (DQ191347). A heuristic parsimony analysis of this dataset was performed in PAUP (Version 4.0b10, Swofford 2002)using tree-bisection-reconnection algorithm with randomsequence addition (100,000 replicates), MULTREES optionin effect, and branch lengths optimized under acceleratedtransformation (ACCTRAN). Characters were unordered andunweighted. This methodology was also carried out for the 18SrRNA sequence data set where gaps were treated as a fifth state.The data set included the outgroup sequences Phymatolithonlaevigatum and P. lenormandii, GenBank accession numbersU60740 and U60741, respectively.
For ML analyses, the nucleotide substitution model was firstdetermined using Modeltest version 3.7 (Posada and Crandall1998). For the cox1 data set, ML analyses employed aTrN + I + G model of sequence evolution with base frequen-cies A = 0.4127, C = 0.1483, G = 0.1370, and T = 0.3020 andthe proportion of invariable sites (I) = 0.4578 with gammadistribution shape parameters 0.7259. For the 18S rRNA dataset, ML analyses employed a TrN + I + G model of sequenceevolution with base frequencies A = 0.2525, C = 0.2024,G = 0.2815, and T = 0.2636 and the proportion of invariablesites (I) = 0.6927 with gamma distribution shape parameters0.7817. ML analyses were carried out using a heuristic search asoutlined above using 1,000 replicates. Bootstrap support wasgenerated in PAUP (Version 4.0b10, Swofford 2002) using1,000 pseudoreplicates (Felsenstein 1985), and Bremer supportvalues (Bremer 1994) were generated in TreeRot using the MPstrict consensus tree (Sorenson 1996).
Morphology. Corallina specimens examined for morphology(excluding the specimen that corresponds to GenBanksequence U60946) are listed in Table S1. External and vege-tative cell morphologies were observed using a hand lens(10·). Reproductive and cell morphology were observed bysquash preparation of the specimens that had been decalcifiedusing 1% hydrochloric acid so that the cells would be visibleunder a microscope (UNH: Olympus BX40 microscope, digitalcamera: Nikon D100), and stained with aniline blue accordingto the methods of Brodie and Guiry (1988). The terminologyused followed that of Irvine and Johansen (1994).
Table 1. Details of parsimony analysis for the two molec-ular markers.
Marker 18S rRNA cox1
Number of taxa 19 38Number of bp included in analysis 1,686 539Number of trees generated byparsimony analysis
8 12
Number of parsimony informativecharacters
125 171
Tree length 184 471CI 0.873 0.659RI 0.932 0.901RC 0.814 0.594
CI, consistency index; RC, rescaled consistency index; RI,retention index.
288 RACHEL H. WALKER ET AL .
Examination of C. officinalis synonym type material. To deter-mine the identity of two genetically distinct species clustersmorphologically identified as C. officinalis, these were com-pared with the type specimen of C. officinalis Linnaeus (1758)(LINN, 1293.9) and with images of the types and descriptionsof the species synonymized by Guiry and Guiry (2008) withC. officinalis, viz. C. calvadosii (PC), C. nana (MCVE), andC. compacta (PC). In addition, the two species were comparedwith images and descriptions of the type specimens of all otherCorallina species currently listed by Guiry and Guiry (2008).
RESULTS
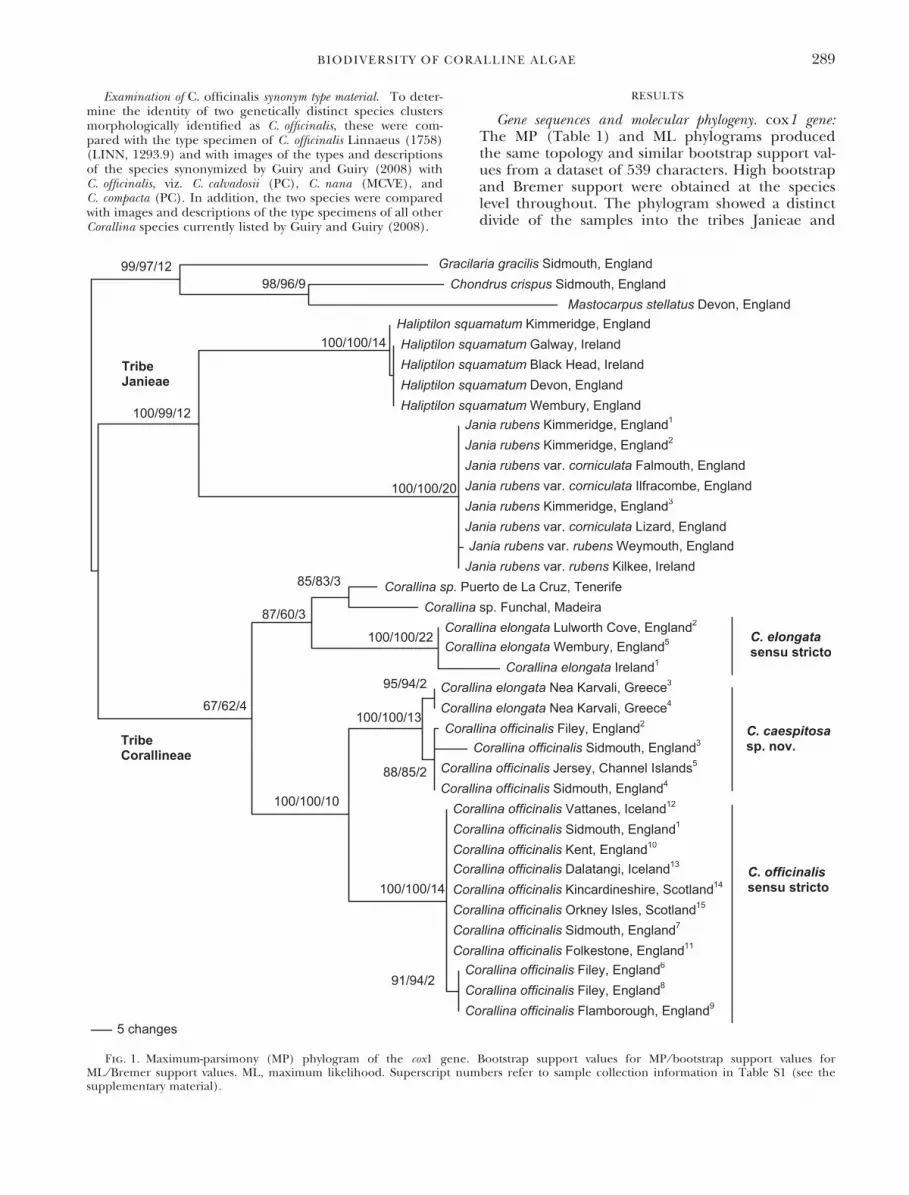
Gene sequences and molecular phylogeny. cox1 gene:The MP (Table 1) and ML phylograms producedthe same topology and similar bootstrap support val-ues from a dataset of 539 characters. High bootstrapand Bremer support were obtained at the specieslevel throughout. The phylogram showed a distinctdivide of the samples into the tribes Janieae and
Fig. 1. Maximum-parsimony (MP) phylogram of the cox1 gene. Bootstrap support values for MP ⁄ bootstrap support values forML ⁄ Bremer support values. ML, maximum likelihood. Superscript numbers refer to sample collection information in Table S1 (see thesupplementary material).
BIODIVERSITY OF CORALLINE ALGAE 289

Corallineae (Fig. 1), differing by 71–100 bp(Table 2). Within Janieae, J. rubens and H. squama-tum sequences were clearly separated, differing by46–48 bp, and intraspecific variation within bothspecies was 0–1 bp. There were no bp differencesbetween J. rubens var. rubens and var. corniculata.Within the Corallineae, samples divided into twodistinct groupings, both of which contained twoclades. In one of these groupings, one cluster (sam-ples from Britain and Iceland) contained specimensidentified as C. officinalis with intraspecific variationof 0–3 bp. The other cluster, containing specimensidentified as either C. officinalis or C. elongata (withsamples from the Mediterranean and Britain andIreland), varied by 0–14 bp and was considered tobe a species distinct from C. officinalis. The differ-ence between these two clusters was 45–54 bp. Theother grouping contained specimens identified asC. elongata (excluding two specimens only identifiedto genus) and contained two distinct clusters, whichvaried by 47–69 bp. Intraspecific variation of onecluster, with samples from the mid-Atlantic, was24 bp, and for the other cluster, with samples fromBritain and Ireland, 0–15 bp.
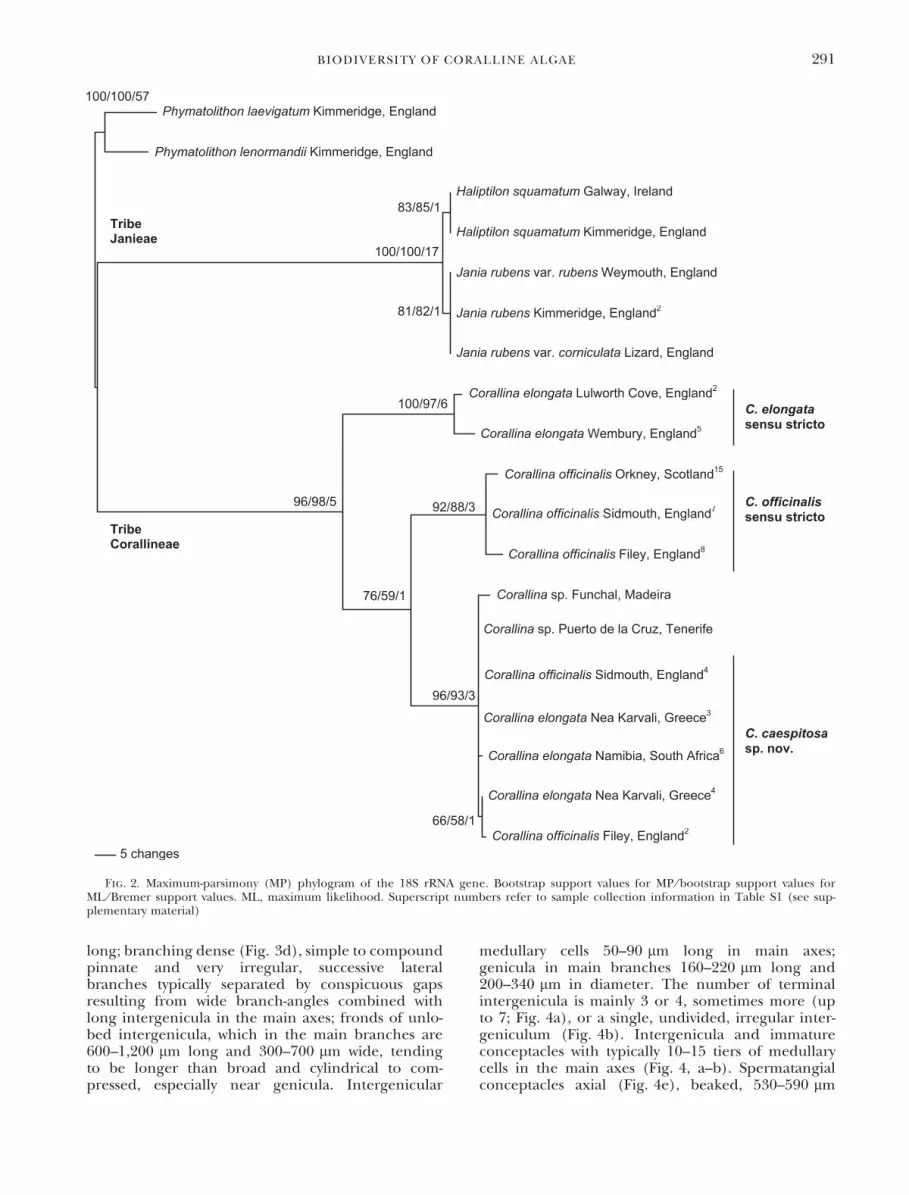
18S rRNA gene: The MP (Table 1) and ML phylo-grams from a data set of 1,686 characters producedthe same topology and similar bootstrap support val-ues. High bootstrap and Bremer support wereobtained for most nodes. The phylogram showed adistinct divide of the samples into the tribes Janieaeand Corallineae (Fig. 2), differing by 45–54 bp(Table 3). Within Janieae, J. rubens and H. squama-tum sequences differed by 4 bp, but there was nointraspecific variation for either species. There wasalso no bp difference between J. rubens var. rubensand var. corniculata. Within the Corallineae, samplesdivided into three clades. One of these clusters withsamples from Britain was consistent with the typespecimen of C. officinalis and had intraspecific varia-tion of 3–5 bp. The cluster containing samples iden-tified as C. elongata from Britain varied by 7 bp.Another cluster, whose morphological characterswere not consistent with those of the type specimenof C. officinalis, contained samples identified eitheras C. elongata or C. officinalis from Tenerife, Britain,
Greece, and South Africa and had intraspecific vari-ation of 0–5 bp.
The major difference between the two gene treeswas the placement of the specimens identified togenus from Madeira and Tenerife, which clusteredwith those identified as C. elongata from Britain andIreland in the cox1 phylogram, but with the clusterof C. elongata and C. officinalis specimens in the 18SrRNA phylogram.
Corallina caespitosa R. H. Walker, J. Brodie etL. M. Irvine sp. nov.
Frondes erectae, usque ad 45 mm altae, rigidae,e crusta basali orte, infra non ramosae, supra ramo-sae ramulis densis, irregularibus simplicibus vel pin-natis compositis; intergenicula plerumque longioraquam lata, cylindrical frondibus basi, compressis,versus apicem frondis cuneiformibus, typice 10–15seriae cellulorum medullosorum instructis; frondisapice intergeniculis 3–4 vel usque ad 7 vel interdumintergeniculo singulo indivisi irregulari instuctis.Conceptacula spermatangialia axialia, rostrata, 530–590 lm in diametro; conceptacula tratrasporica axia-lia, interdum pseudolateralia 440–580 lm in diamet-ro; conceptacula carposporangialia ignota. Data desequentii de holotypus = cox1 Genbank DQ191343et 18S rRNA Genbank FM180101.
Base crustose, erect fronds up to 45 mm, stiff,unbranched stipe, branching dense, irregularly sim-ple to compound pinnate; intergenicula generallylonger than broad, cylindrical at frond base, becom-ing compressed, wedge-shaped toward the frondapex, typically with 10–15 tiers of medullary cells;apex with 3–4 or more (up to 7) intergenicula oroccasionally a single undivided irregular intergenic-ulum. Spermatangial conceptacles axial, beaked,530–590 lm in diameter; tetrasporangial concepta-cles axial, sometimes pseudolateral, 440–580 lm indiameter; carposporangial conceptacles unknown.GenBank accession numbers of the Holotype:DQ191343 [cox1] and FM180101 [18S rRNA].
Holotype: Sidmouth, Devon; coll. Juliet Brodie andLavinia Robba, 23 April 2005, JBLR8 (Fig. 3, a–b)(BM) [BM000804549].
Isotype: JBLR10 (Fig. 3c), (BM) [BM000804550].Type locality: Sidmouth, Devon, UK, in a shallow
rock pool on the exposed rocky shore at Chit Rocks,SY869121.
Etymology: The epithet caespitosa = tufted. Thename refers to dense branching of the fronds.
Seasonality: In Britain and Ireland, specimens havebeen collected in April and from July to September.Spermatangial and tetrasporangial specimens havebeen collected in April, and tetrasporangial speci-mens from July to September.
Habitat: Marine, growing attached to substrata inrock pools from the upper limit of the mid-littoralto the lower limit of the littoral zone.
Description: Thallus with a firmly attached crustosebase, typically >10 mm in diameter bearing tufts ofbranched, stiff, usually erect fronds up to 45 mm
Table 2. Absolute base pair differences for clusters in thecox1 gene phylogram for Corallina, Jania, and Haliptilon.
Corallinaofficinalis
Corallinacaespitosa
Corallinaelongata
(Atlantic)
Corallinaelongata
(Britain)
Janiarubens
Haliptilonsquamatum
C. officinalis 0–3C. caespitosa 45–54 0–14C. elongata(Atlantic)
64–70 62–81 24
C. elongata(Britain)
79–94 77–91 47–69 0–15
J. rubens 90–91 92–99 77–82 88–100 0–1H. squamatum 83 83–84 71–75 87–98 46–48 0–1
290 RACHEL H. WALKER ET AL .
long; branching dense (Fig. 3d), simple to compoundpinnate and very irregular, successive lateralbranches typically separated by conspicuous gapsresulting from wide branch-angles combined withlong intergenicula in the main axes; fronds of unlo-bed intergenicula, which in the main branches are600–1,200 lm long and 300–700 lm wide, tendingto be longer than broad and cylindrical to com-pressed, especially near genicula. Intergenicular
medullary cells 50–90 lm long in main axes;genicula in main branches 160–220 lm long and200–340 lm in diameter. The number of terminalintergenicula is mainly 3 or 4, sometimes more (upto 7; Fig. 4a), or a single, undivided, irregular inter-geniculum (Fig. 4b). Intergenicula and immatureconceptacles with typically 10–15 tiers of medullarycells in the main axes (Fig. 4, a–b). Spermatangialconceptacles axial (Fig. 4e), beaked, 530–590 lm
Fig. 2. Maximum-parsimony (MP) phylogram of the 18S rRNA gene. Bootstrap support values for MP ⁄ bootstrap support values forML ⁄ Bremer support values. ML, maximum likelihood. Superscript numbers refer to sample collection information in Table S1 (see sup-plementary material)
BIODIVERSITY OF CORALLINE ALGAE 291

maximum diameter; spermatangial conceptaclechambers 300–400 lm in diameter, 230–250 lmhigh, with the roofs 390–480 lm thick. Tetrasporan-gial conceptacles (Fig. 4g) mainly axial, sometimespseudolateral (Fig. 4f), protruding markedly, 440–580 lm maximum diameter; axial tetrasporangialconceptacles infrequently bearing branches (<20%)and typically of branches with 1 subtending inter-geniculum; tetrasporangial conceptacle chambers320–460 lm diameter, �380–480 lm high and roofs100–110 lm thick; tetrasporangia �120–160 lmlong and 40–60 lm in diameter after release. Of thefertile material examined, tetrasporangial concepta-cles were more numerous on individuals thanspermatangial conceptacles and contained between15 and 25 tetraporangia. Carposporangial concepta-cles were not observed.
Comparison of C. caespitosa with other northeasternAtlantic Corallina species. Vegetative characters canbe used to distinguish between C. caespitosa sp. nov.,C. officinalis, and C. elongata (Table 4). The crustosebase in C. caespitosa is smaller than in C. officinalisand C. elongata, but bases of individuals may becomefused and therefore indistinguishable. C. caespitosais generally smaller in size with stiff fronds butwith more dense and irregular branching than inC. officinalis or C. elongata (Fig. 3, a–j; 4, a–d). Thegap separation in C. caespitosa between successive lat-eral branches is intermediate. The cylindrical inter-genicula are longer in C. caespitosa and C. officinalisthan in C. elongata. The occurrence of more thanthree terminal intergenicula and single, undividedirregular intergenicula is more frequent in C. caespi-tosa than in C. officinalis (Fig. 4, a–d). The tiers ofmedullary cells are less conspicuous in C. elongataand C. officinalis than in C. caespitosa (Fig. 4, a–d).The main differences in reproductive structuresbetween C. caespitosa and the other two species arethe number of intergenicula supporting theconceptacles and the presence or absence ofpseudolateral conceptacles (Table 4).
Comparison of C. caespitosa with the type specimen ofC. officinalis and currently accepted synonyms. The typespecimen of C. officinalis has intergenicula of‡1 mm in length and conspicuous gaps betweenlateral branches (Fig. 5a), features that distinguishit from C. caespitosa. The syntype fragments of
C. compacta have conspicuous gaps between thelateral branches and do not have single terminalintergenicula (Fig. 5b). Type material of C. nanahas large intergenicula at the base, which becomeincreasingly compressed and reduced in size towardthe tips of the fronds (Fig. 5c). These morphologi-cal characteristics are not observed in C. caespitosa.Lamouroux did not designate a type for C. calvadosii,and the only possible original material lacks local-ity and collector information (Fig. 5, d–e). Thismaterial was identified by Johansen in 1966 asArthrocardia corymbosa (J. V. Lamour.) Decne., whichis a South African endemic. While it superficiallyresembles Corallina, it differs morphologically fromC. caespitosa as the intergenicula are much largerand branching in the main branches is dichoto-mous, as opposed to pinnate. In addition, C. caespi-tosa could be distinguished from the images anddescriptions of the type specimens of all other Coral-lina species currently listed (Guiry and Guiry 2008).
DISCUSSION
Molecular phylogenetics. The cox1 and 18S rRNAgene phylogenies support the tribes Janieae andCorallineae. The very small difference between Janiaand Haliptilon species (4 bp = 0.2%) in our 18SrRNA gene phylogeny agrees with the results ofBailey and Chapman (1998), who also recorded alow sequence divergence between these genera. Thefinding that H. squamatum and J. rubens should beclassified in the same genus, as proposed by Kimet al. (2007), is further supported by the cox1 genephylogeny because the 46–48 bp difference showsspecies-level divergence (Saunders 2005). However,the original description of Haliptilon was based onH. cuvieri (J. V. Lamour.) H. W. Johans. et P. C.Silva, which is now considered to be a synonym ofH. roseum (Lam.) Garbary et H. W. Johans. In thephylogeny of Kim et al. (2007), samples identifiedas H. roseum fall in a separate cluster from samplesof H. squamatum and J. rubens. Therefore, the phylo-genetic position of Haliptilon based on originalmaterial of H. cuvieri and the position of Jania basedon J. rubens need to be established before genericaffinity of Haliptilon and Jania can be resolved.
The 0–1 bp sequence variation in the cox1 geneand no variation in the 18S rRNA gene suggest thatfor the genes studied, there is no genetic basis forthe morphological differences between J. rubens var.rubens and J. rubens var. corniculata. Irvine andJohansen (1994) refrained from consideringJ. rubens var. corniculata as a distinct species becauseboth varieties sometimes share characters that areused to distinguish them. Studies are thereforeneeded to determine the environmental influenceon intergenicula shape and branching pattern, asindividuals of J. rubens with lobed intergenicula andpinnate branching have been identified in theNorth Atlantic (Irvine and Johansen 1994).
Table 3. Absolute base pair differences for clusters inthe 18S rRNA gene phylogram for Corallina, Jania, andHaliptilon.
Corallinaofficinalis
Corallinacaespitosa
Corallinaelongata
(Britain)
Janiarubens
Haliptilonsquamatum
C. officinalis 3–5C. caespitosa 14–19 0–5C. elongata(Britain)
20–22 25–29 7
J. rubens 49–54 45–50 46–50 0H. squamatum 50–54 46–50 47–50 4 0
292 RACHEL H. WALKER ET AL .

Fig. 3. Gross morphology of Corallina spp. Scale bar, 10 mm. (a) Corallina caespitosa (Holotype, Sidmouth, UK, BM000804549,DQ191343 [cox1], FM180101 [18S rRNA]). (b) Corallina caespitosa (Holotype, Sidmouth, UK, BM000804549, DQ191343 [cox1], FM180101[18S rRNA]). (c) C. caespitosa (Isotype, Sidmouth, UK, BM000804550, DQ191342 [cox1]). (d) C. caespitosa fragment (Holotype, DQ191343[cox1], FM180101 [18S rRNA]). (e) C. officinalis (Kent, UK, BM000771429, FM180076 [cox1]). (f) C. officinalis (North Yorkshire, UK,BM000639033, FM180075 [cox1]). (g) C. officinalis fragment (North Yorkshire, UK, BM000639033, FM180075 [cox1]). (h) C. elongata(Devon, UK, BM000804981, FM180069 [cox1], FM180099 [18S rRNA]). (i) C. elongata (Dorset, UK, BM000774725, FM180065 [cox1],FM180095 [18S rRNA]). (j) C. elongata fragment (Dorset, UK, BM000774725, FM180065 [cox1], FM180095 [18S rRNA]).
BIODIVERSITY OF CORALLINE ALGAE 293

The molecular results for both genes are morecomplex for the tribe Corallineae than for Jani-eae. The genetic divergence in the cox1 gene forCorallineae, which is notably greater between thetwo main groupings than the divergence betweenC. officinalis and C. caespitosa in one group andbetween the two clusters of C. elongata in theother group, raises the question as to the parame-ters of these species and when they diverged fromeach other. The 18S rRNA phylogeny shows a dif-ferent relationship between the elements withinthe Corallineae; it does not form two distinctgroupings with clusters as shown in the cox1 tree(Figs. 1 and 2). In contrast, three clusters can beidentified. Of these, C. officinalis sensu stricto andC. elongata from Britain correspond to the similargroups in the cox1 tree, whereas the sequences ofthe unidentified Corallina specimens from Madeiraand Tenerife clustered with the C. caespitosa andC. elongata sequences. The results indicate that thename C. elongata has been misapplied in both theAtlantic and Mediterranean. While the concept ofthe species is not disputed for those samples fromBritain and Ireland, the samples from Greece areC. caespitosa and those from elsewhere in theAtlantic are of an unknown species. The difference
between the sequences of C. elongata from Ireland(GenBank) and those from Dorset and Devon isattributed to differences in the ends of thesequences. We were unable to examine the origi-nal chromatogram. These specimens had beenmorphologically identified as C. elongata. The sameapplies to C. caespitosa from Devon (identified asC. officinalis).
Given that the molecular phylogenies are inter-preted within the context of the genes that theyare based on, Saunders (2005) proposed that forclosely related species, it is essential to assess mito-chondrial data with nuclear markers for evidenceof introgression, hybridization, and incompletespecies boundaries. However, until we have morestudies of red algae using the cox1 marker, we arenot able to explore these possibilities further.Higher divergences, as observed in the sequencesidentified to genus from Tenerife and Madeira inthe cox1 gene, are interpreted as the result ofgeographical isolates and gene pool fragmen-tation (Hebert et al. 2003). These islands aregeographically isolated in the Atlantic, but it isnot known if the populations interbreed withpopulations from elsewhere in the Atlantic andMediterranean.
Fig. 4. Frond tips and internal morphology. (a) Corallina caespitosa, six intergenicula toward end of the frond; scale bar, 0.3 mm. (b)C. caespitosa, single, undivided irregular intergeniculum; scale bar, 0.3 mm. (c) C. officinalis, three terminal intergenicula; scale bar,0.5 mm. (d) C. elongata, three terminal intergenicula; scale bar, 0.4 mm. (e) C. caespitosa spermatangial conceptacle; scale bar, 180 lm. (f)C. caespitosa pseudolateral tetrasporangial conceptacle; scale bar, 220 lm. (g) C. caespitosa tetrasporangial conceptacle; scale bar, 230 lm.
294 RACHEL H. WALKER ET AL .

In this study, the intraspecific and interspecificsequence variation can provide an indication of thebp differences that could be expected (Harvey et al.2002), but as the cox1 and 18S rRNA genes haveshown, the levels of genetic variation betweencurrently recognized species can vary considerably.This variation indicates the importance of consider-ing diversity within the context of distributionalranges. The use of multiple samples will allow theboundaries of biogeographic populations to beinvestigated and the extent to which morphology isgenetically constrained (Pillmann et al. 1997). Thisneed supports current thinking in the field that thelevels of diversity within the marine environmenthave been greatly underestimated (Mathews 2006),even in a well-studied flora such as the seaweeds ofBritain and Ireland (Brodie and Zuccarello 2007).This also highlights that the mechanisms of
speciation and gene flow within the marine environ-ment require further exploration.
Morphology of Corallina. Morphological characterscan be used to distinguish among C. caespitosa,C. officinalis, and C. elongata. The morphology ofC. officinalis and C. elongata from Britain and Irelandagreed with the description in Irvine and Johansen(1994) except for the number of sporangia in eachtetrasporangial conceptacle, which was 15–25 andnot over 30. Characters that are useful for distin-guishing between C. officinalis and C. caespitosasp. nov. are included in the key below:
Key to Corallina species of Britain and Ireland.
(1) Gaps between successive lateral branches absentor inconspicuous; intergenicula in main axestending to be as wide as long, with averagelength <1 mm; conceptacles typically at the end
Table 4. Morphological assessment of Corallina species in Britain and Ireland. Terminology follows that of Irvine andJohansen (1994).
Description
Observations
Corallina caespitosa Corallina officinalis Corallina elongata
Vegetative morphologyCrustose basediameter (mm)
Typically up to 10 Up to 70 Up to 150 or more
Fronds (mm) Stiff, branched, upto 45 long
Stiff, branched, upto 120 long
Limp, branched, upto 200 long
Branching Simple to compoundpinnate, very denseand irregular
Simple to compoundpinnate, dense to sparse,often irregular
Simply to compoundpinnate, usually dense,occasionally irregular
Gaps between successivelateral branches
Typically conspicuous Conspicuous Absent or inconspicuous
Intergenicula size inmain branches (mm)
0.5–1 long, 0.3–0.7 wide,cylindrical to compressed,with concave uppersurfaces
1–2 long, 0.3–1 wide,cylindrical to compressed,with concave upper surfaces
0.5–1.0 long, 0.4–0.8 wide,compressed, with concaveupper surfaces
Number of terminalintergenicula
3 or 4 (up to 7) or1 undivided, irregularintergeniculum
3, occasionally 4 or more;rarely 1 undivided, irregularintergeniculum
3, occasionally 4 or more
Genicula dimensions inmain branches (lm)a
160–220 in length, 200–340in diameter
180–350 in length, 180–350in diameter
140–190 in length,190–240 in diameter
Reproductive morphologya
Spermatangialconceptacles (lm)
Axial, beaked, chambers300–400 diameter, 230–250high, with roofs 390–480 thick
Axial, beaked, chambers250–480 diameter, 100–300high, with roofs 200–600thickb,c
Axial, chambers 185–390diameter, 165–280 high,with roofs 110–415 highthickb
Tetrasporangialconceptacles (lm)
Axial, sometimes pseudolateral,each containing 15–25sporangia; chambers �450diameter, 380 high, withroofs 100 thick
Axial, sometimes pseudolateral,each containing 15–25 sporangia;chambers 330–500 diameter,400–500 high, roofs 60–110 thick
Axial, not pseudolateral,each containing 15–25sporangia; chambers 150–350diameter, 200–320 high,with roofs 80–150 thickb
Tetrasporangia (lm) 100–140 long and 40–60wide after release
100–140 long and 40–60wide after release
95–225 longb
Carposporangialconceptacles (lm)
Not seen Axial, chambers 250–400diameter, 300–320 high,with roofs 150–240 thick,fusion cells 90–200 diameter,6–11 thickb,c
Axial, chambers 250–360in diameter, fusion cells upto 180 in diameter, 6–11thickb
Carposporangia (lm) Not seen 30–80 diameterb,c 40–80 diameterb
aObservable after decalcification using microscope 10·.bAccording to Irvine and Johansen (1994).cThe morphological description of C. officinalis in Irvine and Johansen (1994) probably also encompasses the characteristics ofC. caespitosa sp. nov.
BIODIVERSITY OF CORALLINE ALGAE 295

of lateral branches with >1 subtending inter-geniculum; pseudolateral conceptacles absent.................................................................C. elongataGaps between successive lateral branches typi-cally conspicuous; intergenicula tending to belonger than broad; conceptacles typically at theend of branches with 1 subtending intergenicu-lum; pseudolateral conceptacles sometimes pres-ent, protruding markedly......................................2
(2) Gaps between successive lateral branches con-spicuous; average length of intergenicula inmain axes >1 mm; fronds up to 120 mm long,sparse to dense.....................................C. officinalisGaps between successive lateral branches typicallyconspicuous; average length of intergenicula inmain axes <1 mm; fronds up to 45 mm long, verydense......................................................C. caespitosa
The authors thank Prof. Christine Maggs, Prof. David John,and Prof. Michael Guiry for collecting fresh material;Ms. Jennifer Bryant and Dr. Charlie Jarvis for guidance withherbarium material; and Dr. Norman Robson for assistancewith the Latin translation. We thank the Linnean Society of theBritish Isles for permitting us to use the specimen image of thetype of C. officinalis (1293.3), Dr. Bruno de Reviers (PC) forimages of C. calvadosii and C. compacta, and Dr. Enrico Ratti(MCVE) for images of C. nana. The images of C. calvadosii andC. compacta have also been prepared for inclusion in a forth-coming book on the type specimens of geniculate Corallinaleshoused in PC (Museum National d’Histoire Naturelle, Depart-ement Systematique et Evolution, batiment de Cryptogamie).
Bailey, J. C. 1999. Phylogenetic positions of Lithophyllumincrustans and Titanoderma pustulatum (Corallinaceae, Rhodo-phyta) based on 18S rRNA gene sequence analyses, with arevised classification of the Lithophylloideae. Phycologia38:208–16.
Fig. 5. Gross morphology of Corallina officinalis type specimen and type specimens of currently accepted synonyms. (a) C. officinalis(LINN, 1293.9); scale bar, 17 mm. (b) C. compacta (PC); scale bar, 10 mm. (c) C. nana (MCVE); scale bar, 20 mm. (d) C. calvadosii (PC);scale bar, 10 mm. (e) C. calvadosii (PC); scale bar, 14 mm.
296 RACHEL H. WALKER ET AL .

Bailey, J. C. & Chapman, R. L. 1998. A phylogenetic study of theCorallinales (Rhodophyta) based on nuclear small-subunitrRNA gene sequences. J. Phycol. 34:692–705.
Bailey, J. C., Gabel, J. E. & Freshwater, D. W. 2004. Nuclear 18SrRNA gene sequence analyses indicate that the Mastophoroi-deae (Corallinales, Rhodophyta) is a polyphyletic taxon.Phycologia 43:3–12.
Bremer, B. 1994. Branch support and tree stability. Cladistics10:295–304.
Brodie, J. & Guiry, M. D. 1988. Life history and reproduction ofBotryocladia ardreana sp. nov. (Rhodophyta, Rhodymeniales)from Portugal. Phycologia 27:109–30.
Brodie, J. & Zuccarello, G. C. 2007. Systematics of the species richalgae: red algal classification, phylogeny and speciation. InHodkinson, T. R. [Ed.] Reconstructing the Tree of Life: Taxonomyand Systematics of Species Rich Taxa. CRC ⁄ Taylor & Francis, BocaRaton, Florida, pp. 317–30.
Cabioch, J. 1972. Etude sur les Corallinacees II. La morphogenese,consequences systematiques et phylogenetiques. Cah. Biol.Mar. 13:137–287.
Cabioch, J. 1988. Morphogenesis and generic concepts in corallinealgae – a reappraisal. Helgol. Meeresunters. 42:493–509.
Chamberlain, Y. M. & Cooke, P. J. 1984. Crustose corallines grow-ing in sea-urchin cups at Fanore, Co. Clare. Br. Phycol.J. 19:190–1.
Felsenstein, J. 1985. Confidence limits on phylogenies – anapproach using the bootstrap. Evolution 39:783–91.
Guiry, M. D. & Guiry, M. 2008. AlgaeBase version 4.2. NationalUniversity of Ireland, Galway. http://www.algaebase.org(accessed February 2008).
Harvey, A. S., Woelkerling, W. J. & Millar, A. J. K. 2002. TheSporolithaceae (Corallinales, Rhodophyta) in south-easternAustralia: taxonomy and 18S rRNA phylogeny. Phycologia41:207–27.
Hebert, P. D. N., Cywinska, A., Ball, S. L. & DeWaard, J. R. 2003.Biological identifications through DNA barcodes. Proc. R. Soc.Lond. B Biol. Sci. 270:313–21.
Holmgren, P., Holmgren, N. H. & Bartlett, L. C. 1990. IndexHerbariorum, Part 1. The Herbaria of the World. Koeltz ScientificBooks, Konigstein.
Irvine, L. M. & Johansen, H. W. 1994. Corallinoideae. In Irvine,L. M. & Chamberlain, Y. M. [Eds.] Seaweeds of the British Isles:Vol.1 Rhodophyta Pt. 2B. Corallinales, Hildenbrandiales. HMSO,London, pp. 37–57.
Johansen, H. W. 1976. Current status of generic concepts incoralline algae (Rhodophyta). Phycologia 15:221–44.
Johansen, H. W. 1981. Coralline Algae, a First Synthesis. CRC Press,Boca Raton, Florida, 239 pp.
Kim, J. H., Guiry, M. D., Oak, J. H., Choi, D.-S., Kang, S.-H., Chung,H. & Choi, H.-G. 2007. Phylogenetic relationships within thetribe Janieae (Corallinales, Rhodophyta) based on molecularand morphological data: a reappraisal of Jania. J. Phycol.43:1310–9.
Kinzie, R. A. & Buddemeier, R. W. 1996. Reefs happen. Glob. ChangeBiol. 2:479–94.
Maddison, D. R. & Maddison, W. P. 2005. MacClade. Sinauer Asso-ciates Inc., Sunderland, Massachusetts.
Mathews, L. M. 2006. Cryptic biodiversity and phylogeographicalpatterns in a snapping shrimp species complex. Mol. Ecol.15:4049–63.
Pillmann, A., Woolcott, G. W., Olsen, J. L., Stam, W. T. & King,R. J. 1997. Inter- and intraspecific genetic variation in Caulerpa(Chlorophyta) based on nuclear rDNA ITS sequences. Eur.J. Phycol. 32:379–86.
Posada, D. & Crandall, K. A. 1998. Modeltest: testing the model ofDNA substitution. Bioinformatics 14:817–8.
Robba, L., Russell, S. J., Barker, G. L. & Brodie, J. 2006.Assessing the use of the mitochondrial cox1 marker for usein DNA barcoding of red algae (Rhodophyta). Am. J. Bot.93:1101–8.
Rogers, S. O., Bendich, A. J., Hall, G. E. Jr., Spiker, S., Mac-kenzie, S. A., Price, C. A. & Bisseling, T. 1994. Nucleic acidextraction from plant tissue. In Gelvin, S. B. & Schilperoort,R. A. [Eds.] Plant Molecular Biology Manual. Kluwer, London,pp. A6:1–11.
Saunders, G. W. 2005. Applying DNA barcoding to red macroalgae:a preliminary appraisal holds promise for future applications.Philos. Trans. R. Soc. Lond. B Biol. Sci. 360:1879–88.
Saunders, G. W. & Kraft, G. T. 1994. Small-subunit ribosomal-RNAgene-sequences from representatives of selected families of theGigartinales and Rhodymeniales (Rhodophyta). 1. Evidencefor the Plocamiales ord. nov. Can. J. Bot. 72:1250–63.
Sorenson, M. D. 1996. TreeRot. University of Michigan, Ann Arbor.Swofford, D. 2002. PAUP. Version 4.0b10. Sinauer Associates Inc.,
Sunderland, Massachusetts.Thompson, J. D., Higgins, D. G. & Gibson, T. J. 1994. CLUSTAL W:
improving the sensitivity of progressive multiple sequenceweighting, position-specific gap penalties and weight matrixchoice. Nucleic Acids Res. 22:4673–80.
Woelkerling, W. J. 1988. The Coralline Red Algae: An Analysis of theGenera and Subfamilies of Nongeniculate Corallinaceae. BritishMuseum (Natural History), London, and Oxford UniversityPress, Oxford, 268 pp.
Supplementary Material
The following supplementary material is avail-able for this article:
Table S1. Sample collection information forCorallina, Haliptilon, and Jania. ND, no data.
This material is available as part of the onlinearticle.
Please note: Wiley-Blackwell are not responsi-ble for the content or functionality of any supple-mentary materials supplied by the authors. Anyqueries (other than missing material) shouldbe directed to the corresponding author for thearticle.
BIODIVERSITY OF CORALLINE ALGAE 297





























