Embed Size (px)
Citation preview

www.elsevier.com/locate/vetpar
Veterinary Parasitology 128 (2005) 195–200
IgG avidity pattern in cattle after ingestion of
Neospora caninum oocysts
C. Bjorkmana,*, L.F.P. Gondimb, K. Naslundc, A.J. Treesd, M.M. McAllisterb
aDepartment of Clinical Sciences, Swedish University of Agricultural Sciences, P.O. Box 7019, SE-75007 Uppsala, SwedenbDepartment of Veterinary Pathobiology, University of Illinois, 2001 South Lincoln Avenue, Urbana Illinois, IL 61802, USA
cDepartment of Parasitology (SWEPAR), National Veterinary Institute and Swedish University of Agricultural Sciences,
SE-75189 Uppsala, SwedendVeterinary Parasitology, Liverpool School of Tropical Medicine and Faculty of Veterinary Science, University of Liverpool,
Pembroke Place, Liverpool L3 5QA, UK
Received 22 June 2004; received in revised form 15 November 2004; accepted 26 November 2004
Abstract
The avidity (functional affinity) of specific antibodies are being used to estimate duration of bovine Neospora caninum
infection. Here, we report for the first time the avidity pattern in cattle orally inoculated with N. caninum oocysts. In all, 16
pregnant cows and 7 calves were administered N. caninum oocysts. In the cows, the avidity increased during the early course of
infection. In all but one, the avidity was �35 during the first 6 weeks after infection and no cow had an avidity value >50 until
week 9. The calves were sampled either week 6 (n = 3) or week 9 (n = 9) after infection, and by then had avidities between 2 and
17. The results are in agreement with results from previous investigations of naturally infected cattle, and calves that were
experimentally infected with tachyzoites. They further validate the ability of the N. caninum iscom avidity ELISA to accurately
assess the duration of infection.
# 2004 Elsevier B.V. All rights reserved.
Keywords: Avidity; Cattle; Neospora caninum; Oocysts; ELISA; Humoral immunity
1. Introduction
The protozoan parasite Neospora caninum is
increasingly recognised as an important pathogen
causing abortion and stillbirth in pregnant cattle
(Dubey, 1999). In many parts of the world, it is
* Corresponding author. Tel.: +46 1867 1778;
fax: +46 1867 3545.
E-mail address: [email protected] (C. Bjorkman).
0304-4017/$ – see front matter # 2004 Elsevier B.V. All rights reserved
doi:10.1016/j.vetpar.2004.11.030
considered one of the most common infectious causes
of bovine abortion. The most commonly recognised
route of infection in cattle is congenital transmission
of the parasite from an infected dam to her fetus (Pare
et al., 1996; Davison et al., 1999b). However, cattle
can also be infected postnatally by ingesting oocysts
shed by a dog or coyote, which are definitive hosts for
the parasite (McAllister et al., 1998; Gondim et al.,
2004d). Infection seems to be life-long, and infected
females can transmit the infection to their offspring
.

C. Bjorkman et al. / Veterinary Parasitology 128 (2005) 195–200196
during consecutive pregnancies (Bjorkman et al.,
1996). Congenital infection can result in abortion or
stillbirth although the majority of infected fetuses are
born clinically healthy but chronically infected.
Congenitally infected heifers can in their turn give
birth to infected calves and the infection can thus,
be kept in a cattle herd for many years without
involvement of a definitive host (Bjorkman et al.,
1996; Anderson et al., 1997). Abortion outbreaks in
cattle herds have been connected with recent infection
in the pregnant heifers and/or cows, whereas endemic
abortion patterns are thought to be consistent with
presence of chronically (persistently) infected dams
(Thurmond et al., 1997; Davison et al., 1999a;
McAllister et al., 2000; Dubey, 2003).
Demonstration of specific antibodies in serum from
an animal is indicative of infection (Bjorkman and
Uggla, 1999; Dubey, 2003). However, as the antibody
levels fluctuate during pregnancy in persistently
infected animals (Stenlund et al., 1999; Guy et al.,
2001), antibody levels or titers cannot be used to
discriminate between acutely and chronically infected
animals. For this purpose, an avidity ELISA was
developed, based on the fact that the first antibodies
synthesised after an antigenic challenge have lower
affinity for the antigen than those produced later on
(Bjorkman et al., 1999). This IgG avidity test utilised
membrane proteins incorporated into iscoms as antigen,
and was the first of its kind to be reported. Since then,
other avidity tests based on tachyzoite extracts (Maley
et al., 2001; Sager et al., 2003) and the N. caninum
membrane protein NcSRS2 (Schares et al., 2002) have
been described. The avidity of N. caninum specific IgG
antibodies increased with time in calves experimentally
infected by subcutaneous or intravenous injections of N.
caninum tachyzoites (Bjorkman et al., 1999). Even
though both the mode of transmission and the parasite
life stage used in these experiments differ from what is
seen under natural conditions, these results suggested
that low IgG avidity levels give an indication whether an
animal has been recently exposed to the infection
(Bjorkman et al., 1999). As far as is known, the only
natural route of postnatal infection in cattle is ingestion
of feed or water contaminated with oocysts shed by a
definitive host. Lactogenic infection with tachyzoites
has been shown experimentally in newborn calves
(Ugglaetal.,1998;Davisonetal.,2001)but isnotknown
to occur naturally.
This investigation was performed in order to further
elucidate the development of IgG avidity during N.
caninum infection by analysing samples from preg-
nant cows and young calves orally inoculated with N.
caninum oocysts. Samples were also analysed from
calves born to the inoculated cows.
2. Materials and methods
2.1. Animals and samples
Serum samples collected from pregnant cows and
from young calves were used in this study.
A) T
hirteen mixed breed beef cows that were derivedfrom a previous study that had—at that time point—
included 19 cows (Gondim et al., 2004b). The cows
were inoculated orally with 1500–115,000 sporu-
lated oocysts of different N. caninum isolates at
different time points during gestation. Blood
samples were collected by venous puncture before
and up to 24 weeks after inoculation. Not all cows
werekeptuntilcalving; therefore,only5calveswere
born to the 13cows. Included in thepresent studyare
the two calves shown to be seropositive before
ingestion of colostrum, one of which was stillborn
and blood sampled at necropsy. The identification
numbers of the cows and calves are the same as in
Gondim et al. (2004b).
B) T
hree 10–12-year-old Hereford cross Friesiancows were at 70 days gestation infected orally with
600 sporulated oocysts of the Nc-Liv isolate and
blood samples were collected up to 30 weeks after
inoculation. This experiment has been described
in detail by Trees et al. (2002).
C) A
lso included were samples from seven 2–14-day-old heifer and bull calves orally given 1400–
29,000 oocysts of Nc-Illinois (Gondim et al.,
2004a), Nc-beef (McAllister et al., 1998; McAll-
ister et al., 2000) or Nc-deer (Gondim et al.,
2004c). Blood samples were collected before and
6–9 weeks after inoculation.
2.2. Antibody analyses
N. caninum antibodies were detected by iscom
ELISA using tachyzoite antigens incorporated into
C. Bjorkman et al. / Veterinary Parasitology 128 (2005) 195–200 197
immunostimulating complexes (iscoms) as antigen
and a monoclonal anti-bovine IgG1 antibody as
conjugate (Bjorkman et al., 1997; Frossling et al.,
2003). All absorbance values were correlated to a
positive control serum with a mean absorbance value
of 1.0, and sera with corrected absorbance �0.20 were
considered positive.
Sera with absorbance values >0.35 in the iscom
ELISA were further tested for IgG avidity by the
method of Bjorkman et al. (1999). Briefly, the sera
were diluted in five-fold serial dilutions from 1:50 and
applied in duplicate microtitre wells. After incubation
with serum, 1 well for each serum was treated with
urea to release low avidity antibodies from the
antigen-antibody complex. Absorbance was measured
in treated and untreated wells after incubation with
conjugate and substrate. The IgG avidity was
calculated using the formula:
IgG avidity ¼ end point titer with urea
end point titer without urea
� �� 100
IgG avidity values could only be calculated who had
endpoint titers �1:50.
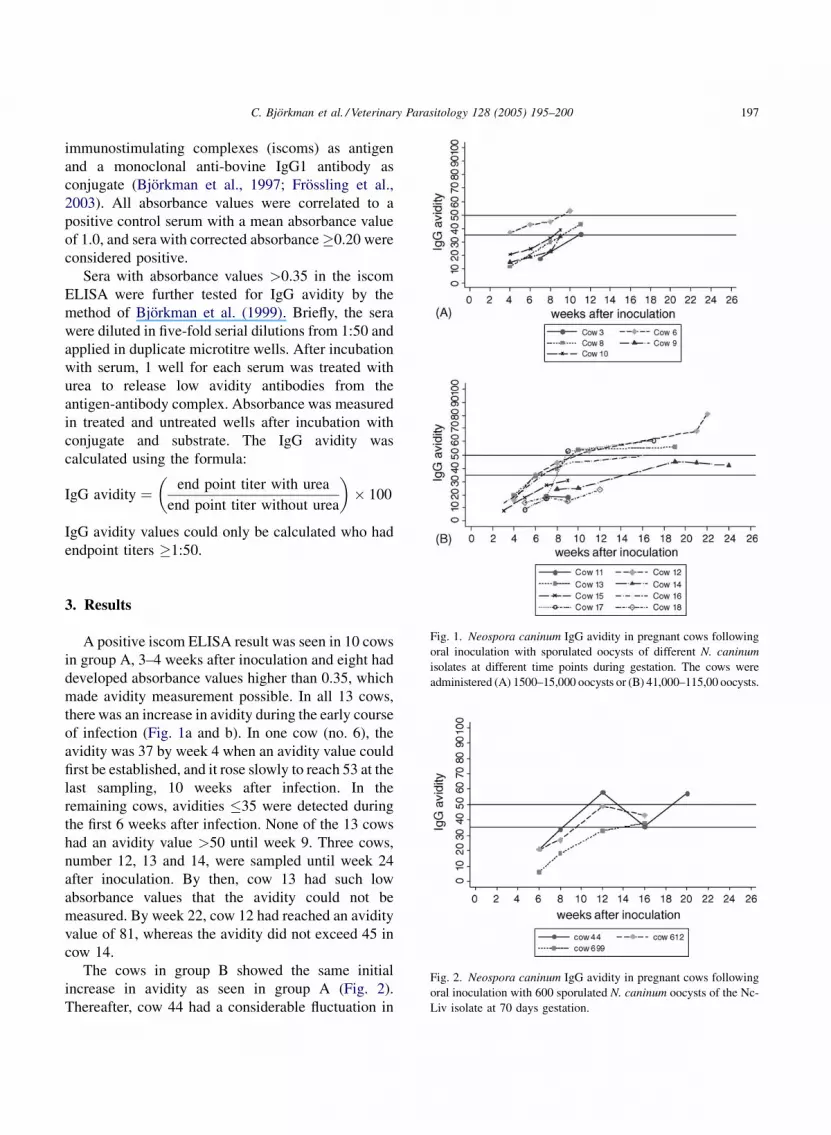
Fig. 1. Neospora caninum IgG avidity in pregnant cows following
oral inoculation with sporulated oocysts of different N. caninum
isolates at different time points during gestation. The cows were
administered (A) 1500–15,000 oocysts or (B) 41,000–115,00 oocysts.
Fig. 2. Neospora caninum IgG avidity in pregnant cows following
oral inoculation with 600 sporulated N. caninum oocysts of the Nc-
Liv isolate at 70 days gestation.
3. Results
A positive iscom ELISA result was seen in 10 cows
in group A, 3–4 weeks after inoculation and eight had
developed absorbance values higher than 0.35, which
made avidity measurement possible. In all 13 cows,
there was an increase in avidity during the early course
of infection (Fig. 1a and b). In one cow (no. 6), the
avidity was 37 by week 4 when an avidity value could
first be established, and it rose slowly to reach 53 at the
last sampling, 10 weeks after infection. In the
remaining cows, avidities �35 were detected during
the first 6 weeks after infection. None of the 13 cows
had an avidity value >50 until week 9. Three cows,
number 12, 13 and 14, were sampled until week 24
after inoculation. By then, cow 13 had such low
absorbance values that the avidity could not be
measured. By week 22, cow 12 had reached an avidity
value of 81, whereas the avidity did not exceed 45 in
cow 14.
The cows in group B showed the same initial
increase in avidity as seen in group A (Fig. 2).
Thereafter, cow 44 had a considerable fluctuation in

C. Bjorkman et al. / Veterinary Parasitology 128 (2005) 195–200198
avidity values between week 8 and 20. Although the
sampling continued until calving 30 weeks post
infection, the antibody levels decreased from week 16
and onwards and no avidity could be estimated later
than 20 weeks after infection in any cow.
Two calves born to group A cows were seropositive
at birth when tested by iscom ELISA with absorbance
values �0.97. One of them was stillborn and had an
avidity of 16. The dam had been inoculated
approximately day 130 in pregnancy. The other
seropositive calf was born to a cow inoculated when
164 days pregnant and had an IgG avidity of 32. Both
dams had avidities >50. The calves born to cows in
group B were all seronegative.
Four of the seven orally infected calves were blood
sampled 9 weeks after inoculation and their avidity
values were 8–17. The remaining three calves were
kept only until week six post inoculation and the
avidity values by then were 2, 3 and 16, respectively.
The iscom ELISA absorbances were 0.83–1.11.
4. Discussion
Here, we report for the first time the avidity pattern
in cattle orally inoculated with N. caninum oocysts. In
the pregnant cows there was an increase in IgG avidity
during the early course of infection similar to that
found in animals that had been injected with
tachyzoites (Bjorkman et al., 1999; Maley et al.,
2001; Schares et al., 2002). There was large individual
variation in avidity pattern, but all calves and most of
the cows had avidity levels <35 during the first 6–7
weeks post infection. The avidity levels in the calves
were considerably lower then the avidities previously
measured in calves infected with tachyzoites using the
same IgG avidity iscom ELISA (Bjorkman et al.,
1999). If this difference in avidity level was a result of
the infection mode, dose or any other circumstances is
not known. They were not, however, associated with
the N. caninum antibody levels. In the earlier study
(Bjorkman et al., 1999), the iscom ELISA absorbance
values of the tachyzoite inoculated calves at week 6
was 0.82–1.00 (not previously reported), and thus, of a
similar magnitude as those reported here.
In cow 44, a temporary drop in avidity, from 58 to
36, occurred between week 12 and 16, while the
ELISA absorbance values increased from 0.60 to 0.80.
A majority of persistently N. caninum infected cows
have consistently high avidity IgG antibodies (Guy
et al., 2001; Bjorkman et al., 2003), although in certain
individuals low avidity antibodies may persist for
many months. Drops in avidity were reported by Sager
et al. (2003) who applied a method based on a tachyzoite
antigen. However, a drop in avidity, as occurred in cow
44, has not previously been observed by the iscom
ELISA test. These sera were re-tested with similar
results. The avidity data from this cow are outliers
compared to other data in this and previous studies.
Indirect evidence that IgG avidity is low in recently
oocyst infected animals has been reported from
investigations performed in naturally infected cattle
herds after putative point source horizontal spread of
the parasite. When the avidity test was applied in a
long-term study in such a herd the mean avidity in the
cows was 30 during the outbreak and increased to 74
during the following 3 years (Bjorkman et al., 2003).
The mode of avidity in the herd during the same period
moved from 21–40 to 61–80. Similar results were
reported by Dijkstra et al. (2002) studying a dairy herd
in which almost 50% of the animals seroconverted
during a period of 6 months between two blood
samplings. However, individual naturally infected
cows can maintain their low avidity in spite of being
seropositive for many years (Bjorkman et al., 2003).
In accordance with previous findings, avidities >50
were only seen in animals that had been infected
longer than 8 weeks. Such overall high avidities have
been detected in herds in which congenital infection is
the principle route of parasite transmission as judged
by high association between seropositivity in dams
and their daughters (Bjorkman et al., 1999; Schares
et al., 2002).
The exact mechanism for the parasite transmission
from a dam to her fetus and what decides the outcome
of pregnancy in an infected cow are not known (Innes
et al., 2002). In the present investigation, two of the
eight cows that were kept throughout pregnancy gave
birth to seropositive calves, one of which was stillborn.
These calves both had high antibody levels and avidity
values below 35, whereas the cows that had been
inoculated more than 17 weeks earlier, had avidities
>50. The lower avidity values in the calves might reflect
that they were only recently infected, i.e. that therewas a
significant time span between when the dams were
infected and when they had transmitted the parasite to

C. Bjorkman et al. / Veterinary Parasitology 128 (2005) 195–200 199
the foetuses. Alternatively, the immature immune
system in the foetus may result in a slower maturation
of the avidity of the IgG antibodies.
Recently, the avidity iscom ELISA has been used to
investigate infection patterns in cattle herds experien-
cing abortionoutbreaks (Jenkins et al., 2000;McAllister
et al., 2000). The aborting cows had lower avidity than
the normally calving cows suggesting a recent point
source exposure to the parasite in these herds. However,
each avidity test must be carefully evaluated before it is
used to establish whether aborting cows are recently or
chronically infected. The reason is that even though the
abortion per se does not seem to affect the avidity
results as measured by the iscom ELISA, a decrease in
avidity at abortion was found by an avidity test based
on tachyzoite extract (Sager et al., 2003). Perhaps this
reflects a difference in binding pattern between
antibodies directed to different epitopes.
The N. caninum iscom avidity ELISA have
previously been applied in naturally N. caninum
infected cattle herds to investigate when the infection
has been acquired and to elucidate the infection
pattern in the herds (Jenkins et al., 2000; McAllister
et al., 2000; Dijkstra et al., 2002; Bjorkman et al.,
2003). Avidity testing provides robust epidemiologi-
cal information when used to analyse groups or herds,
but interpretation of the avidity results for any single
animal is less certain because of individual variation.
The results in the present study validate the ability of
the test to assess the duration of infection following
ingestion of N. caninum oocysts.
Acknowledgements
The authors thank Liying Gao for technical
assistance. The study was financially supported in-
part by the Swedish Research Council for Environ-
ment, Agricultural Sciences and Spatial Planning and
the Swedish Farmers’ Foundation for Agricultural
Research and in-part by United States Department of
Agriculture (USDA-NRICGP 2000–01997).
References
Anderson, M.L., Reynolds, J.P., Rowe, J.D., Sverlow, K.W., Pack-
ham, A.E., Barr, B.C., Conrad, P.A., 1997. Evidence of vertical
transmission of Neospora sp. infection in dairy cattle. J. Am.
Vet. Med. Assoc. 210, 1169–1172.
Bjorkman, C., Johansson, O., Stenlund, S., Holmdahl, J., Uggla, A.,
1996. Neospora species infection in a herd of dairy cattle. J. Am.
Vet. Med. Assoc. 208, 1441–1444.
Bjorkman, C., Holmdahl, O.J.M., Uggla, A., 1997. An indirect
enzyme-linked immunoassay (ELISA) for demonstration of
antibodies to Neospora caninum in serum and milk of cattle.
Vet. Parasitol. 68, 251–260.
Bjorkman, C., Naslund, K., Stenlund, S., Maley, S.W., Buxton, D.,
Uggla, A., 1999. An IgG avidity ELISA to discriminate between
recent and chronic Neospora caninum infection. J. Vet. Diagn.
Invest. 11, 41–44.
Bjorkman, C., Uggla, A., 1999. Serological diagnosis of Neospora
caninum infection. Int. J. Parasitol. 29, 1497–1507.
Bjorkman, C., McAllister, M.M., Frossling, J., Naslund, K., Leung,
F., Uggla, A., 2003. Application of the Neospora caninum IgG
avidity ELISA in assessment of chronic reproductive losses
following an outbreak of neosporosis in a herd of beef cattle.
J. Vet. Diagn. Invest. 15, 3–7.
Davison, H.C., Otter, A., Trees, A.J., 1999a. Significance of Neos-
pora caninum in British dairy cattle determined by estimation of
seroprevalence in normally calving cattle and aborting cattle.
Int. J. Parasitol. 29, 1189–1194.
Davison, H.C., Otter, A., Trees, A.J., 1999b. Estimation of vertical
and horizontal transmission parameters of Neospora caninum
infection in dairy cattle. Int. J. Parasitol. 29, 1683–1689.
Davison, H.C., Guy, C.S., McGarry, J.W., Guy, F., Williams, D.J.L.,
Kelly, D.F., Trees, A.J., 2001. Experimental studies on the
transmission of Neospora caninum between cattle. Res. Vet.
Sci. 70, 163–168.
Dijkstra, T., Barkema, H.W., Bjorkman, C., Wouda, W., 2002. A
high rate of seroconversion for Neospora caninum in a dairy herd
without and obvious increased incidence of abortions. Vet.
Parasitol. 109, 203–211.
Dubey, J.P., 1999. Neosporosis in cattle: biology and economic
impact. J. Am. Vet. Med. Assoc. 214, 1160–1163.
Dubey, J.P., 2003. Neosporosis in cattle. J. Parasitol. 89, S42–
S56.
Frossling, J., Bonnett, B., Lindberg, A., Bjorkman, C., 2003. Vali-
dation of a Neospora caninum iscom ELISA without a gold
standard. Prev. Vet. Med. 57, 141–153.
Gondim, L.F.P., Laski, P., Gao, L., McAllister, M.M., 2004a.
Variation of the internal transcribed spacer 1 sequence within
individual strains and among different strains of Neospora
caninum. J. Parasitol. 90, 119–122.
Gondim, L.F.P., McAllister, M.M., Anderson-Sprecher, R.C., Bjork-
man, C., Lock, T.F., Firkins, L.D., Gao, L., Fisher, W.R.T.,
2004b. Transplacental transmission and abortion in cows admi-
nistered Neospora caninum oocysts. J. Parasitol. 90, 1394–1400.
Gondim, L.F.P., McAllister, M.M., Mateus-Pinilla, N.E., Pitt, W.C.,
Mech, L.D., Nelson, M.E., 2004c. Transmission of Neospora
caninum between wild and domestic animals. J. Parasitol. 90,
1361–1365.
Gondim, L.F.P., McAllister, M.M., Pitt, W.C., Zemlicka, D.E.,
2004d. Coyotes (Canis latrans) are definitive hosts of Neospora
caninum. Int. J. Parasitol. 32, 159–161.

C. Bjorkman et al. / Veterinary Parasitology 128 (2005) 195–200200
Guy, C.S., Williams, D.J.L., Kelly, D.F., McGarry, J.W., Guy, F.,
Bjorkman, C., Smith, R.F., Trees, A.J., 2001. Neospora caninum
in persistently infected, pregnant cows: spontaneous transpla-
cental infection is associated with an acute rise in maternal
antibody. Vet. Rec. 149, 443–449.
Innes, E.A., Andrianarivo, A.G., Bjorkman, C., Williams, D.J.L.,
Conrad, P.A., 2002. Immune responses to Neospora caninum
and prospects for vaccination. Trends Parasitol. 18, 497–
504.
Jenkins, M.C., Caver, J.A., Bjorkman, C., Anderson, T.C., Romand,
S., Vinyard, B., Uggla, A., Thulliez, P., Dubey, J.P., 2000.
Serological investigation of an outbreak of Neospora cani-
num-associated abortion in a dairy herd in southeastern United
States. Vet. Parasitol. 94, 17–26.
Maley, S.W., Buxton, D., Thomson, K.M., Schriefer, C.E.S., Innes,
E.A., 2001. Serological analysis of calves experimentally
infected with Neospora caninum: a 1-year study. Vet. Parasitol.
96, 1–9.
McAllister, M.M., Dubey, J.P., Lindsay, D.S., Jolley, W.R.,
Wills, R.A., McGuire, A.M., 1998. Dogs are definitive
hosts of Neospora caninum. Int. J. Parasitol. 28, 1473–
1478.
McAllister, M.M., Bjorkman, C., Anderson-Sprecher, R., Rogers,
D.G., 2000. Evidence of point-source exposure to Neospora
caninum and protective immunity in a herd of beef cows. J. Am.
Vet. Med. Assoc. 217, 881–887.
Pare, J., Thurmond, M.C., Hietala, S.K., 1996. Congenital Neospora
caninum infection in dairy cattle and associated calfhood mor-
tality. Can. J. Vet. Res. 60, 133–139.
Sager, H., Gloor, M., Bjorkman, C., Kritzner, S., Gottstein, B., 2003.
Assessment of antibody avidity in aborting cattle by a somatic
Neospora caninum tachyzoite antigen IgG avidity ELISA. Vet.
Parasitol. 112, 1–10.
Schares, G., Barwald, A., Staubach, C., Sondgen, P., Rauser, M.,
Schroder, R., Peters, M., Wurm, R., Selhorst, T., Conraths, F.J.,
2002. p38-avidity-ELISA: examination of herds experiencing
epidemic or endemic Neospora caninum-associated bovine
abortion. Vet. Parasitol. 106, 293–305.
Stenlund, S., Kindahl, H., Magnusson, U., Uggla, A., Bjorkman, C.,
1999. Serum antibody profile and reproductive performance
during two consecutive pregnancies of cows naturally infected
with Neospora caninum. Vet. Parasitol. 85, 227–234.
Thurmond, M.C., Hietala, S.H., Blanchard, P.C., 1997. Herd-based
diagnosis of Neospora caninum-induced endemic and epidemic
abortion in cows and evidence for congenital and postnatal
transmission. J. Vet. Diagn. Invest. 9, 44–49.
Trees, A.J., McAllister, M.M., Guy, C.S., McGarry, J.W., Smith,
R.F., Williams, D.J.L., 2002. Neospora caninum: oocyst chal-
lenge of pregnant cows. Vet. Parasitol. 109, 147–154.
Uggla, A., Stenlund, S., Holmdahl, O.J.M., Jakubek, E.-B., Thebo,
P., Kindahl, H., Bjorkman, C., 1998. Oral Neospora caninum
inoculation of neonatal calves. Int. J. Parasitol. 28, 1467–1472.





























