Embed Size (px)
Citation preview

Secondary immunisation with high-dose heterologouspeptide leads to CD8 T cell populations with reducedfunctional avidity
Sharmal Narayan, Allison Choyce, Germain J. P. Fernando andGraham R. Leggatt
Centre for Immunology and Cancer Research, Princess Alexandra Hospital, Universityof Queensland, Woolloongabba, Brisbane, Australia
Expansion of high- or low-avidity CD8 T cells in vitro inversely correlates with theconcentration of peptide ligand present during culture. In contrast, the selectiveenrichment of high- or low-avidity Tcell populations in vivo using peptide immunisationis not well documented. In our study, a single immunisationwith different doses of wild-type peptide or a variant peptide able to stimulate CTL responses cross-reactive withwild-type peptide failed to shift the average avidity of the responding CD8 T cellpopulation specific to either peptide. However, in contrast to homologous prime-boostimmunisation, heterologous prime-boost immunisation incorporating high doses of thesecond immunogen resulted in peptide-specific CD8 Tcell populations polarized towarda low average functional avidity. These data suggest that sequential exposure tostructurally related viral peptides could impair rather than promote anti-viral immunityby lowering the avidity of the responding CD8 T cell population. This study hasimplications for improving vaccine strategies against viruses and tumours and enhancesour understanding of heterologous immunity during sequential viral infection.
Introduction
Cytotoxic T lymphocytes form an important immunesurveillance system against infection by viruses. Pro-ductive viral infection leads to the synthesis of virallyderived non-self proteins that are enzymatically de-graded into peptides and represent ideal target ligandsfor CTL when presented in conjunction with MHC class Imolecules at the cell surface. Recognition of the largenumber of different peptide/MHC class I ligandsgenerated within the cell requires a correspondingly
large pool of TCR to be maintained within the T cellrepertoire. It is estimated that the number of TCR in themature periphery is insufficient to support a single TCRspecific for every peptide/MHC complex [1]. As aconsequence, degeneracy within TCR recognition iscommon, allowing for a number of different peptide/MHC complexes to be productively recognized by asingle TCR, albeit with varying affinities [1, 2]. This isevident in the numerous reports of Tcell cross-reactivityagainst defined peptide epitopes both in vitro and in vivo[3–5]. Conversely, for any given peptide/MHC complex,there are a number of different T cell clones, each with aunique TCR, capable of binding the peptide/MHCcomplex with varying affinity [1, 6]. Unfortunately,the measurement of membrane-bound TCR affinity atthe immunological synapse between the CTL and targetcell is not easily determined and does not necessarilypredict CTL function. Instead, CTL function is deter-mined by multiple membrane interactions, most im-portantly TCR/peptide/MHC but promoted by adhe-
Correspondence: Dr. Graham Leggatt, Centre for Immunologyand Cancer Research, University of Queensland, 4thFloor,Research Building, Princess Alexandra Hospital, Brisbane, QLD,Australia 4102Fax: +61-7-3240-5946e-mail: [email protected]
Received 10/11/05Revised 17/10/06
Accepted 5/12/06
[DOI 10.1002/eji.200535688]
Key words:Avidity � Cross-
reactivity � CytotoxicT lymphocyte� Heterologous
immunity � Humanpapillomavirus
Abbreviations: HPV: human papillomavirus � KLH: keyholelimpet haemocyanin � SFU: spot-forming units
Sharmal Narayan et al. Eur. J. Immunol. 2007. 37: 406–415406
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

sion/costimulatory molecules and downstream intra-cellular signalling events. Selective recruitment of CTLclones with potentially different TCR has instead beeninvestigated using functional readouts in response totitrated peptide ligand. The concentration of peptidegiving half-maximal effector responses in a CTLpopulation has been termed “functional avidity” andrepresents an important correlate of in vivo protectionagainst viral infection [7]. Alternatively, functionalavidity at the population level has also come to representthe concentration of peptide required to activate 50% ofthe total antigen-specific Tcells within a CD8 population[8, 9]. This latter definition, while not addressing avidityon a per cell basis, is useful in determining the averagefunctional avidity (i.e. the relative proportion of high-and low-avidity clones) in a mixed population ofantigen-specific T cell clones. High-avidity T cell re-sponses have been correlated with in vivo resistance toviral infection, protection against tumours and suscept-ibility to autoimmune diabetes [10–13]. Generation oflow-avidity T cell responses to overexpressed selfantigens in tumours leads to a reduction in tumour sizewhile maintaining tolerance to surrounding tissue [14,15]. These studies suggest that polarization of CTLresponses toward high- and low-avidity phenotypes canhave important biological consequences. To date,approaches to steer the average avidity of a T cellresponse by vaccination have involved the use of DNA/poxvirus prime-boost, peptide-pulsed DC, cytokinessuch as IL-15 and cells transfected with costimulatorymolecules [8, 16–18]. In contrast, the use of peptidevaccination to skew the avidity of a CD8 Tcell response isnot well documented.
In this study, we show that either a high- or a low-avidity phenotype of IFN-c-secreting T cells can beselected by prime-boost immunisation with variantpeptides administered at different doses. Several studieshave now documented dynamic changes in theCD8 T cell repertoire and function after sequentialinfection with heterologous viruses containing cross-reactive T cell epitopes [19, 20]; both beneficial anddetrimental consequences for the host are described [21,22]. A role for changes in T cell population avidity indetermining the outcome of heterologous viral infectionis not well documented but may need further con-sideration on the basis of our findings.
Results
CD8 T cell avidity is unaltered by the dose ofpeptide immunogen
Our previous data demonstrated that, in vitro, highconcentrations of peptide tend to select CTL of relatively
low avidity for expansion, whereas lower concentrationsselect high-avidity Tcells [7]. We therefore hypothesizedthat in vivo immunisation with high peptide doses wouldsimilarly favour generation of lower avidity CD8 cellsand that, conversely, immunisation with low peptidedoses would generate high-avidity CD8 populations.Average functional avidity of a peptide-specificCD8 T cell population has been determined ex vivousing titrated peptide in ELISPOT assays [23, 24]. Toconfirm that the measurement of functional avidity islargely independent of the number of responding cells,we diluted limited numbers (250 to 63 cells) of a humanpapillomavirus (HPV) E7 peptide (GF001)-specific CTLline amongst 1 � 106 C57BL/6 splenocytes in anELISPOT assay. Small numbers of these CTL could bedetected accurately after peptide stimulation in theELISPOT, reflected in the spot-forming units (SFU) atsaturating peptide, with limited variation in thedetermination of functional avidity (bracketed num-bers) (Fig. 1A). Consequently, over the precursorfrequency we tested, functional avidity does not changesubstantially with increased or decreased numbers ofpeptide-specific cells, making the ELISPOT assay anappropriate readout for this study.
Next, mice were immunised with one of two differentdoses of an HPV16E7 peptide (GF001) restricted byH-2Db (Fig. 1C) or, alternatively, with one of five dosesof an E7 variant peptide (GV001) with an alteration inone amino acid compared to GF001 sequence but withimproved binding to H-2Db (Fig. 1B). The E7 variantpeptide (GV001) was first identified based on its abilityto prime CD8 CTL that cross-reacted with wild-type E7peptide and protected against endogenous wild-type E7-bearing tumours in vivo [25]. For each peptide, a singleimmunisation generated CD8 T cell populations wherethe half-maximal concentration of homologous peptidetriggering IFN-c secretion was not significantly alteredby a 10- to 20-fold decrease in immunising peptide dose(Fig. 1). Rather, for each peptide, the lowest immunisingdose of peptide was associated with a reduction in thenumber of IFN-c-secreting cells measured in vitro. Thesedata suggest that during primary activation of peptide-specific CD8 T cells in vivo, varying the immunisingpeptide dose over a 20-fold range does not impact theaverage avidity of the responding T cell population.However in vivo reactivation of previously primedCD8 T cells might more closely mimic the in vitroconditions under which selection for high- and low-avidity populations was previously observed. To testthis, we first primed animals with a fixed dose of peptide(20 lg) and then challenged with a high dose or a lowdose of the same peptide during a secondary immunisa-tion. Even under these conditions, the high or low doseof peptide in the secondary immunisation failed togenerate large differences in the average functional
Eur. J. Immunol. 2007. 37: 406–415 Immunomodulation 407
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

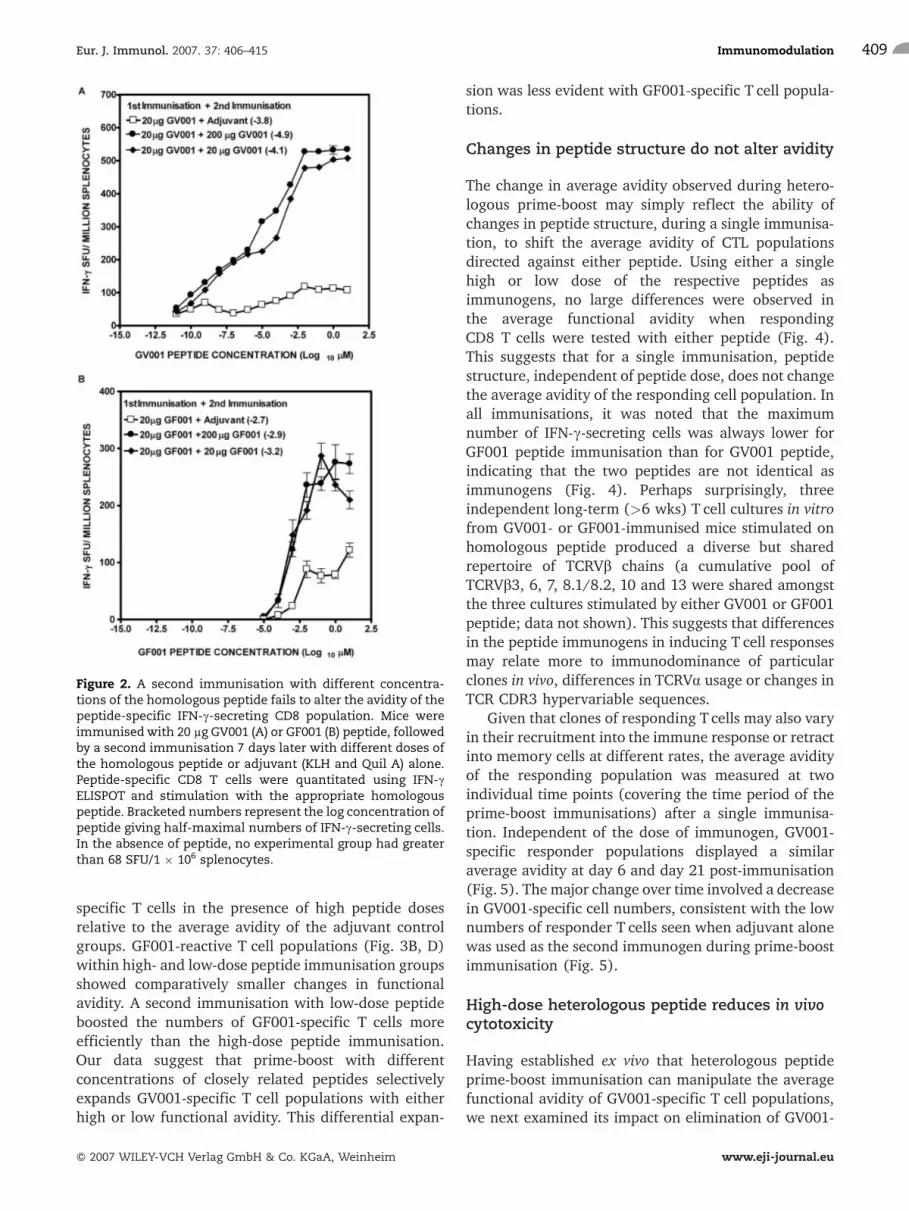
avidity (concentration of peptide leading to IFN-csecretion by 50% of the peptide-specific T cells) of theT cell population specific for either GV001 (Fig. 2A) orGF001 (Fig. 2B). The second peptide immunisation didboost the total number of peptide-specific respondercells relative to the adjuvant control, suggesting an
active role for the second peptide immunisation ineffector cell expansion. Together these data suggest thatthe amount of immunising peptide (over the dose rangetested) does not affect the average functional avidity ofIFN-c-producing CD8 T cells induced in vivo duringprimary or secondary immunisation.
Prime-boost with related peptides changesCD8 T cell avidity
The GV001 and GF001 peptides are able to stimulateCD8 T cells cognate for the other peptide, albeit withdifferent efficiencies [25, 26], suggesting that more thanone clone of cross-reactive T cells exists for this peptidepair and that heterologous peptide stimulation mightselectively expand or delete particular clones, leading toan overall change in functional avidity. We thereforeconsidered whether sequential immunisation withGV001 and GF001 peptides might induce responderT cells with an average avidity different from T cellsprimed and boosted with each peptide alone. Mice wereimmunised with a single dose of either GV001 or GF001peptide, followed by a boost with the alternate peptideat either a high or a low dose. Control mice received asecond immunisation with adjuvant the keyhole limpethaemocyanin (KLH)/Quil A in the absence of peptide.
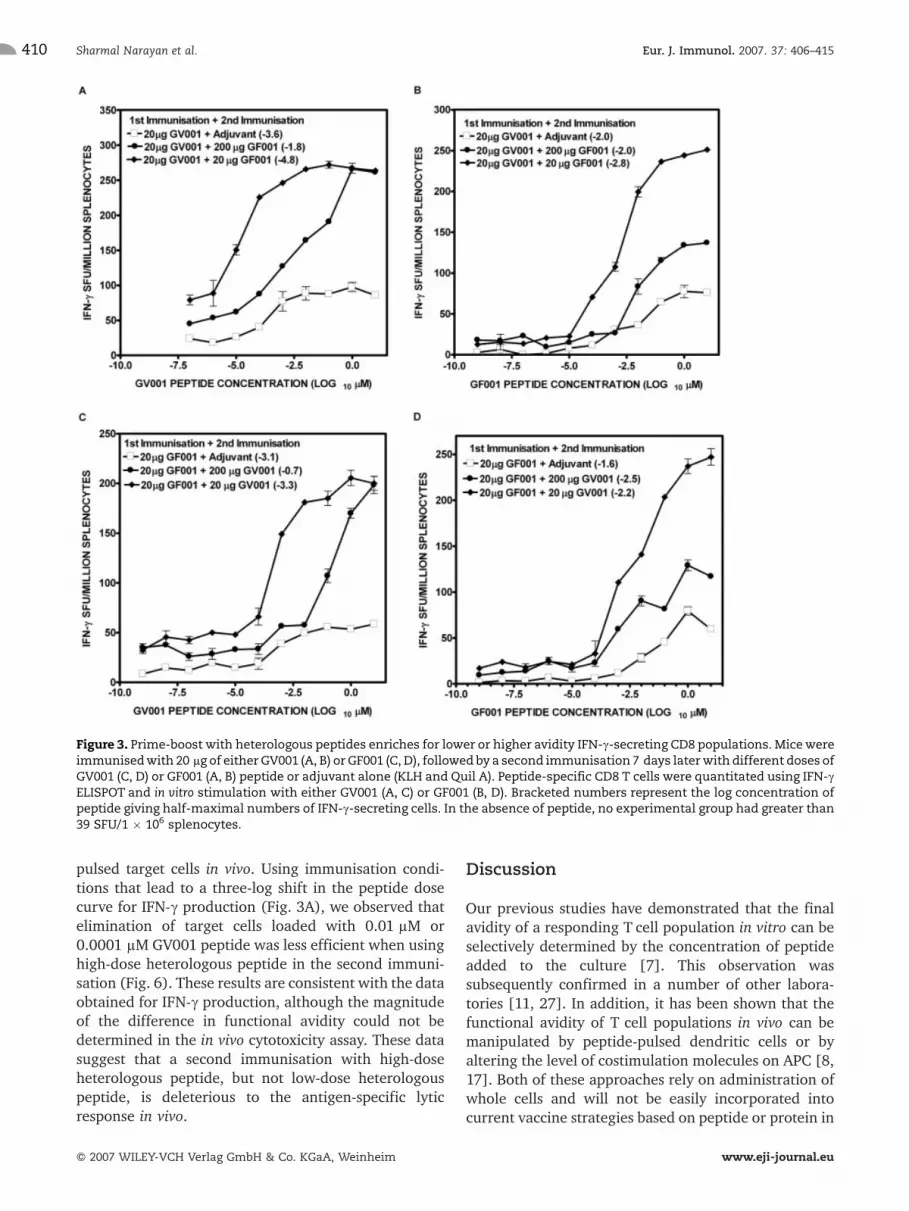
GV001-reactive T cell populations (Fig. 3A, C)showed almost a three-log difference in functionalavidity between the high-dose and low-dose peptideimmunisation groups, independent of the order ofpeptide immunisation. Reimmunisation increased thenumbers of GV001-specific CD8 Tcells in both high- andlow-dose immunisation groups to a similar extent(Fig. 3A, C). The main effect on functional aviditywas the selective expansion of low-avidity GV001-
·
Figure 1.A single immunisationwith different doses of peptideimmunogen does not change the functional avidity of thepeptide-specific CTL population. (A) Cells from an oligoclonallong-termE7-specific CTL linewere spiked into 1 � 106 C57BL/6splenocytes at the indicated percentages, and IFN-c-secretingcells were quantitated using ELISPOT. Spot numbers indicatedon the graph represent the total number of IFN-c-producingcells at saturating peptide as calculated by the curve fit toexperimental data. Experimental determination of E7-specificcells by ELISPOT correlates quite closely with the expectednumber of E7-specific cells plated (i.e. 0.025%= 250 spots vs. 219observed). Bracketed numbers represent the log concentrationof peptide giving half-maximal numbers of IFN-c-secretingcells. (B, C) C57BL/6 mice were immunisedwith different dosesof either GV001 peptide (B) or GF001 peptide (C) mixed withKLH/Quil A. IFN-c-secreting peptide-specific CD8 T cells werequantitated using ELISPOT. Bracketed numbers represent thelog concentration of peptide giving half-maximal numbers ofIFN-c-secreting cells. In the absence of peptide, no experi-mental group had greater than 55 SFU/1 � 106 splenocytes.
Sharmal Narayan et al. Eur. J. Immunol. 2007. 37: 406–415408
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
specific T cells in the presence of high peptide dosesrelative to the average avidity of the adjuvant controlgroups. GF001-reactive T cell populations (Fig. 3B, D)within high- and low-dose peptide immunisation groupsshowed comparatively smaller changes in functionalavidity. A second immunisation with low-dose peptideboosted the numbers of GF001-specific T cells moreefficiently than the high-dose peptide immunisation.Our data suggest that prime-boost with differentconcentrations of closely related peptides selectivelyexpands GV001-specific T cell populations with eitherhigh or low functional avidity. This differential expan-
sion was less evident with GF001-specific T cell popula-tions.
Changes in peptide structure do not alter avidity
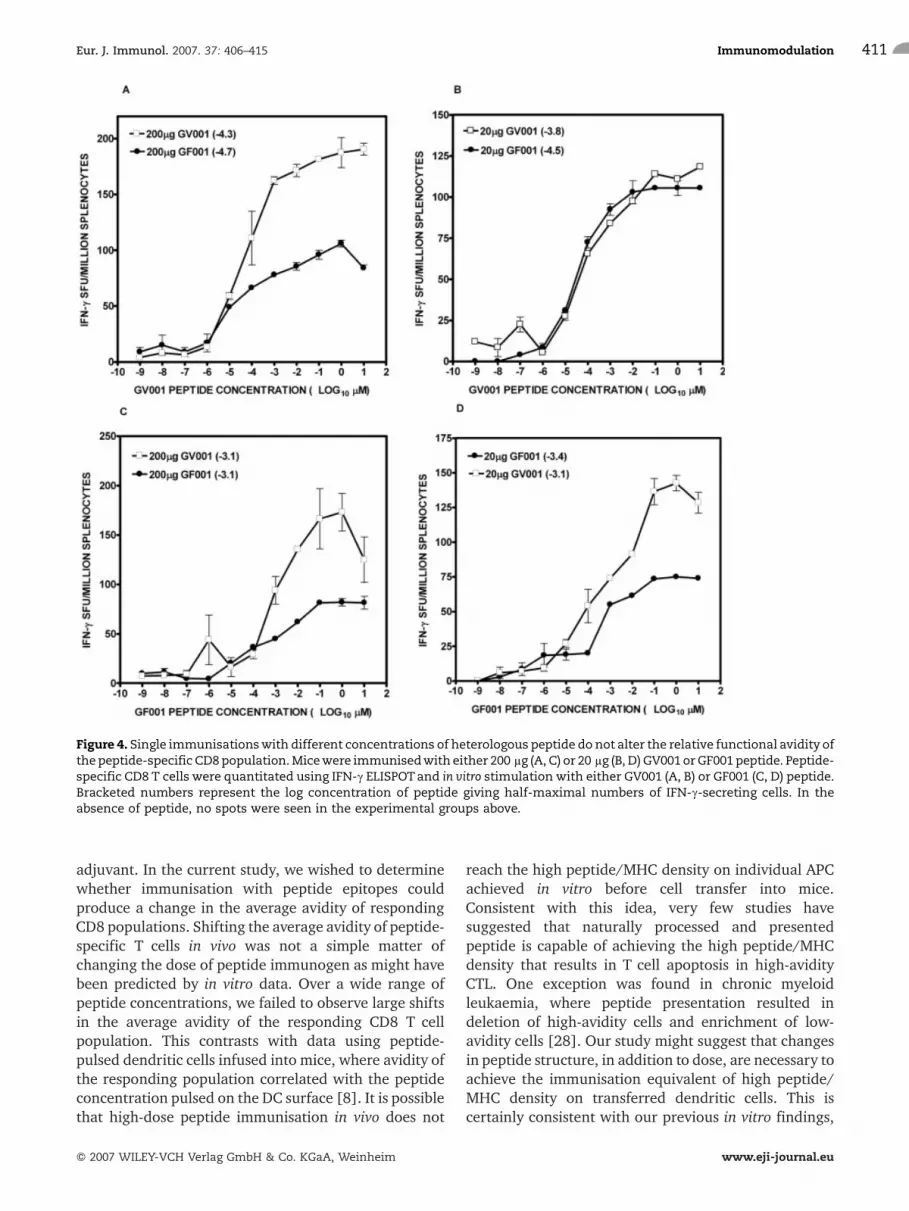
The change in average avidity observed during hetero-logous prime-boost may simply reflect the ability ofchanges in peptide structure, during a single immunisa-tion, to shift the average avidity of CTL populationsdirected against either peptide. Using either a singlehigh or low dose of the respective peptides asimmunogens, no large differences were observed inthe average functional avidity when respondingCD8 T cells were tested with either peptide (Fig. 4).This suggests that for a single immunisation, peptidestructure, independent of peptide dose, does not changethe average avidity of the responding cell population. Inall immunisations, it was noted that the maximumnumber of IFN-c-secreting cells was always lower forGF001 peptide immunisation than for GV001 peptide,indicating that the two peptides are not identical asimmunogens (Fig. 4). Perhaps surprisingly, threeindependent long-term (>6 wks) T cell cultures in vitrofrom GV001- or GF001-immunised mice stimulated onhomologous peptide produced a diverse but sharedrepertoire of TCRVb chains (a cumulative pool ofTCRVb3, 6, 7, 8.1/8.2, 10 and 13 were shared amongstthe three cultures stimulated by either GV001 or GF001peptide; data not shown). This suggests that differencesin the peptide immunogens in inducing T cell responsesmay relate more to immunodominance of particularclones in vivo, differences in TCRVa usage or changes inTCR CDR3 hypervariable sequences.
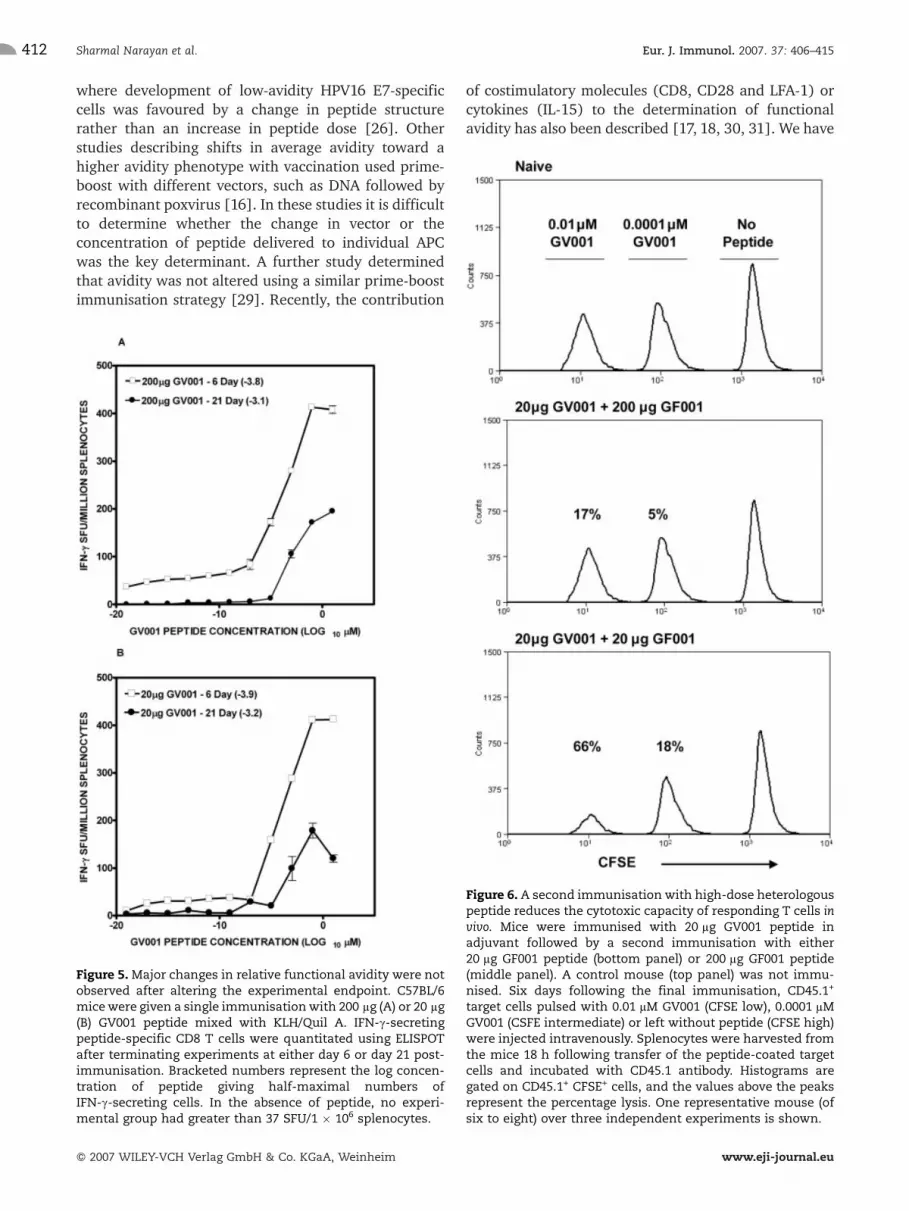
Given that clones of responding T cells may also varyin their recruitment into the immune response or retractinto memory cells at different rates, the average avidityof the responding population was measured at twoindividual time points (covering the time period of theprime-boost immunisations) after a single immunisa-tion. Independent of the dose of immunogen, GV001-specific responder populations displayed a similaraverage avidity at day 6 and day 21 post-immunisation(Fig. 5). The major change over time involved a decreasein GV001-specific cell numbers, consistent with the lownumbers of responder T cells seen when adjuvant alonewas used as the second immunogen during prime-boostimmunisation (Fig. 5).
High-dose heterologous peptide reduces in vivocytotoxicity
Having established ex vivo that heterologous peptideprime-boost immunisation can manipulate the averagefunctional avidity of GV001-specific T cell populations,we next examined its impact on elimination of GV001-
Figure 2. A second immunisation with different concentra-tions of the homologous peptide fails to alter the avidity of thepeptide-specific IFN-c-secreting CD8 population. Mice wereimmunisedwith 20 lg GV001 (A) or GF001 (B) peptide, followedby a second immunisation 7 days later with different doses ofthe homologous peptide or adjuvant (KLH and Quil A) alone.Peptide-specific CD8 T cells were quantitated using IFN-cELISPOT and stimulation with the appropriate homologouspeptide. Bracketed numbers represent the log concentration ofpeptide giving half-maximal numbers of IFN-c-secreting cells.In the absence of peptide, no experimental group had greaterthan 68 SFU/1 � 106 splenocytes.
Eur. J. Immunol. 2007. 37: 406–415 Immunomodulation 409
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
pulsed target cells in vivo. Using immunisation condi-tions that lead to a three-log shift in the peptide dosecurve for IFN-c production (Fig. 3A), we observed thatelimination of target cells loaded with 0.01 lM or0.0001 lM GV001 peptide was less efficient when usinghigh-dose heterologous peptide in the second immuni-sation (Fig. 6). These results are consistent with the dataobtained for IFN-c production, although the magnitudeof the difference in functional avidity could not bedetermined in the in vivo cytotoxicity assay. These datasuggest that a second immunisation with high-doseheterologous peptide, but not low-dose heterologouspeptide, is deleterious to the antigen-specific lyticresponse in vivo.
Discussion
Our previous studies have demonstrated that the finalavidity of a responding T cell population in vitro can beselectively determined by the concentration of peptideadded to the culture [7]. This observation wassubsequently confirmed in a number of other labora-tories [11, 27]. In addition, it has been shown that thefunctional avidity of T cell populations in vivo can bemanipulated by peptide-pulsed dendritic cells or byaltering the level of costimulation molecules on APC [8,17]. Both of these approaches rely on administration ofwhole cells and will not be easily incorporated intocurrent vaccine strategies based on peptide or protein in
Figure 3. Prime-boost with heterologous peptides enriches for lower or higher avidity IFN-c-secreting CD8 populations. Micewereimmunisedwith 20 lg of either GV001 (A, B) or GF001 (C, D), followed by a second immunisation 7 days laterwith different doses ofGV001 (C, D) or GF001 (A, B) peptide or adjuvant alone (KLH and Quil A). Peptide-specific CD8 T cells were quantitated using IFN-cELISPOT and in vitro stimulation with either GV001 (A, C) or GF001 (B, D). Bracketed numbers represent the log concentration ofpeptide giving half-maximal numbers of IFN-c-secreting cells. In the absence of peptide, no experimental group had greater than39 SFU/1 � 106 splenocytes.
Sharmal Narayan et al. Eur. J. Immunol. 2007. 37: 406–415410
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
adjuvant. In the current study, we wished to determinewhether immunisation with peptide epitopes couldproduce a change in the average avidity of respondingCD8 populations. Shifting the average avidity of peptide-specific T cells in vivo was not a simple matter ofchanging the dose of peptide immunogen as might havebeen predicted by in vitro data. Over a wide range ofpeptide concentrations, we failed to observe large shiftsin the average avidity of the responding CD8 T cellpopulation. This contrasts with data using peptide-pulsed dendritic cells infused into mice, where avidity ofthe responding population correlated with the peptideconcentration pulsed on the DC surface [8]. It is possiblethat high-dose peptide immunisation in vivo does not
reach the high peptide/MHC density on individual APCachieved in vitro before cell transfer into mice.Consistent with this idea, very few studies havesuggested that naturally processed and presentedpeptide is capable of achieving the high peptide/MHCdensity that results in T cell apoptosis in high-avidityCTL. One exception was found in chronic myeloidleukaemia, where peptide presentation resulted indeletion of high-avidity cells and enrichment of low-avidity cells [28]. Our study might suggest that changesin peptide structure, in addition to dose, are necessary toachieve the immunisation equivalent of high peptide/MHC density on transferred dendritic cells. This iscertainly consistent with our previous in vitro findings,
Figure 4. Single immunisationswith different concentrations of heterologous peptide do not alter the relative functional avidity ofthepeptide-specific CD8population.Micewere immunisedwith either 200 lg (A, C) or 20 lg (B, D) GV001 or GF001 peptide. Peptide-specific CD8 T cells were quantitated using IFN-c ELISPOTand in vitro stimulation with either GV001 (A, B) or GF001 (C, D) peptide.Bracketed numbers represent the log concentration of peptide giving half-maximal numbers of IFN-c-secreting cells. In theabsence of peptide, no spots were seen in the experimental groups above.
Eur. J. Immunol. 2007. 37: 406–415 Immunomodulation 411
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
where development of low-avidity HPV16 E7-specificcells was favoured by a change in peptide structurerather than an increase in peptide dose [26]. Otherstudies describing shifts in average avidity toward ahigher avidity phenotype with vaccination used prime-boost with different vectors, such as DNA followed byrecombinant poxvirus [16]. In these studies it is difficultto determine whether the change in vector or theconcentration of peptide delivered to individual APCwas the key determinant. A further study determinedthat avidity was not altered using a similar prime-boostimmunisation strategy [29]. Recently, the contribution
of costimulatory molecules (CD8, CD28 and LFA-1) orcytokines (IL-15) to the determination of functionalavidity has also been described [17, 18, 30, 31]. We have
Figure 5. Major changes in relative functional avidity were notobserved after altering the experimental endpoint. C57BL/6micewere given a single immunisationwith 200 lg (A) or 20 lg(B) GV001 peptide mixed with KLH/Quil A. IFN-c-secretingpeptide-specific CD8 T cells were quantitated using ELISPOTafter terminating experiments at either day 6 or day 21 post-immunisation. Bracketed numbers represent the log concen-tration of peptide giving half-maximal numbers ofIFN-c-secreting cells. In the absence of peptide, no experi-mental group had greater than 37 SFU/1 � 106 splenocytes.
Figure 6. A second immunisation with high-dose heterologouspeptide reduces the cytotoxic capacity of responding T cells invivo. Mice were immunised with 20 lg GV001 peptide inadjuvant followed by a second immunisation with either20 lg GF001 peptide (bottom panel) or 200 lg GF001 peptide(middle panel). A control mouse (top panel) was not immu-nised. Six days following the final immunisation, CD45.1+
target cells pulsed with 0.01 lM GV001 (CFSE low), 0.0001 lMGV001 (CSFE intermediate) or left without peptide (CFSE high)were injected intravenously. Splenocytes were harvested fromthe mice 18 h following transfer of the peptide-coated targetcells and incubated with CD45.1 antibody. Histograms aregated on CD45.1+ CFSE+ cells, and the values above the peaksrepresent the percentage lysis. One representative mouse (ofsix to eight) over three independent experiments is shown.
Sharmal Narayan et al. Eur. J. Immunol. 2007. 37: 406–415412
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

not determined whether heterologous prime-boost withpeptide immunogens generates differences in IL-15production or the expression of costimulatory moleculesthat might explain our findings.
Prime-boost with heterologous but closely relatedpeptides has not been widely employed in the study offunctional avidity, despite growing interest in hetero-logous immunity during viral infections [32]. In general,if peptides are identified as improvedMHC binders, thenthey are used in homologous vaccination schemesbefore challenge with infectious agents or tumourscontaining wild-type peptide. Certainly, peptides withimproved binding to MHC have been engineered toinactivate high-avidity CTL during autoimmune disease[33] or to enhance anti-tumour activity [34]. Hetero-logous peptide immunisations have mainly been used inthe context of understanding T cell repertoire degen-eracy through observation of cross-reactive clones invitro. Our study suggests that the boost peptide increaseseffector cell numbers but that certain subsets ofresponding CD8 cells are either enriched or depletedfor IFN-c production according to the dose of the secondimmunogen. We cannot rule out the possibility thatsubsets of responders might also switch their cytokineproduction (e.g. to IL-4) upon encounter with a highdose of heterologous peptide during the secondimmunisation, although the Quil A adjuvant used inour study induces a Th1 environment [35]. However,the in vivo cytotoxicity data were consistent with ex vivoIFN-c production, suggesting that changes in averagefunctional avidity occur for multiple effector functionsin our system. The heterologous peptides do not simplyprime independent populations of T cells with differingavidities, given that single immunisations with eitherpeptide produced CD8 populations with similar averageavidities when assessed on the same peptide readout(Fig. 4A, B). Equally, the shifts in avidity withheterologous immunisation could not be produced bymerely allowing extended harvest times after a singleimmunisation (Fig. 5). This does not conflict withavidity maturation, which may take longer than theshort-term experiments described here [13]. The datainstead favour a model in which the pool of CD8 T cellsprimed during the first immunisation is directlymodified by the peptide used in the second immunisa-tion. We do not believe that CD4 T help is limiting, giventhat KLH was used as a source of CD4 epitopes in boththe primary and secondary immunisations.
It remains to be seen whether a general property ofthe priming or boost peptide, such as MHC affinity, canbe correlated with a skewing of the functional CD8 Tcellrepertoire. This could be determined using large panelsof peptides that induce cross-reactive CD8 T cells butvary in MHC binding affinity. It will also be important totest the robustness of our vaccination strategy using
other peptide systems in which cross-reactive peptideanalogs have been identified. In these systems, high andlow doses of peptide must be determined empirically,given that short peptides will not be equally susceptibleto peptidases in vivo [36]. Predicting the rules by whichthe average avidity of the peptide-specific CD8 popula-tion can be manipulated will be a worthwhile goal giventhe number of disease situations in which the avidity ofthe natural response may be inappropriate or aviditymaturation may fail to occur over time.
Materials and methods
Mice
Female C57BL/6 (H-2b) and B6.SJL (Full name = B6.SJL-PtprcaPop3/BoyJ Arg) mice were obtained from the AnimalResources Centre (Perth, Australia). These mice were main-tained under conventional conditions in specific pathogen-freeholding rooms in the Princess Alexandra Hospital BiologicalResources Facility and used between the ages of 4 and 12 wks.
Peptides
The H-2Db-restricted CTL epitope HPV16 E749–57 (RAHY-NIVTF; GF001) and a variant peptide with a single amino acidchange (RAHYNIVTV; GV001) were purchased from AuspepPty Ltd (Melbourne, Australia) at 83 and 76% purity,respectively. The variant peptide GV001, which contains avaline substitution at the C-terminus, binds more stably to theH-2Db MHC class I molecule [25]. Peptides were dissolved in100% DMSO and stored at –20�C.
Generation of a GF001-specific CTL line
A GF001-specific CTL line was generated and maintained aspreviously described [26]. The CTL line had undergone at leastfour rounds of peptide stimulation in vitro and was used in theELISPOT assay 7 days after the last peptide stimulation.
Immunisations
Single immunisations: Mice were immunised s.c. in the tailwith various doses of GF001 (200 or 20 lg) or GV001 (200,100, 50, 25, 20, 10, 5 or 0.5 lg) peptide in combination with20 lg Quil A (Superfos Biosector, DK-Vedback, Denmark) and20 lg KLH (Sigma). Immunogens were always made up to afinal volume of 50 lL/mouse in PBS. Mice injected with 20 lgQuil A and 20 lg KLH only were used as negative controls.
Double immunisations (Prime-boosting): Mice were im-munised s.c. in the tail with 20 lg GF001 or GV001 inconjunction with 20 lg Quil A and 20 lg KLH. The two groupswere boosted with a second immunisation comprising 200 or20 lg GF001 or GV001 with 20 lg Quil A and 20 lg KLH(homologous or heterologous peptide prime-boosting) 6 or7 days after the primary immunisation.
Each immunisation group consisted of two mice; extractedsplenocytes from each groupwere pooled together for analysis.
Eur. J. Immunol. 2007. 37: 406–415 Immunomodulation 413
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

At least two independent experiments were performed for eachimmunisation regime.
IFN-c ELISPOT assay
Immunised mice were killed 6 days post-immunisation, andspleens were harvested and homogenized in complete RPMImedium (RPMI 1640 supplemented with 10% heat-inactivatedFCS (HI-FCS), 100 U/mL penicillin, 100 lg/mL streptomycin,2 mM glutamine, 5 � 10–5 M 2b-mercaptoethanol, 1 mMsodium pyruvate and 10 mM Hepes buffer) and treated withACK lysing buffer to lyse RBC, then resuspended in completemedia at appropriate cell concentrations. The IFN-c ELISPOTassay is based on slight modifications to a published ELISPOTassay protocol [37, 38]. Nitrocellulose 96-well ELISPOT plates(Millipore Multiscreen-HA) were coated under sterile condi-tions with 75 lL/well anti-mouse IFN-c capture antibody(R4–6A2; PharMingen) at 8 lg/mL in freshly prepared, filter-sterilized 0.1 M NaHCO3 at pH 8.4 (binding buffer). Afterovernight incubation (14–20 h) at 4�C, the capture antibodysolution was discarded and the plates subsequently blocked byincubatingwith 200 lL/well complete RPMImedium for 2 h atroom temperature (23�C). After the blocking, media wasdecanted and the plates washed once with 200 lL RPMImedium without HI-FCS per well. Splenocyte preparationsfrom the immunised mice were then added to the plates at 107
cells/well in a final volume of 100 lL in the presence of 20 U/mL (2 ng/mL) human IL-2 (GibcoBRL). Serial dilutions ofGF001 or GV001 peptides were prepared in complete RPMI,and 100 lL of the various dilutions were also added toappropriate wells. Splenocytes from immunised mice wereincubated in 100 lL complete RPMI alone for negativecontrols. Each sample was tested at least in duplicate. Plateswere incubated at 37�C in 5% CO2 for 16–20 h.
After overnight incubation, the cells were discarded and theplates washed six times with PBST (PBS+ 0.02% Tween 20) toremove residual cells. After washing, 75 lL biotinylated anti-mouse IFN-c detection mAb (XMG1.2; PharMingen) wasadded at a concentration of 1 lg/mL in PBST containing 2%HI-FCS and the plates incubated for 3 h at room temperaturein the dark. To remove unbound antibodies, the plates wereagain washed six times with PBST before addition of 75 lLavidin-horseradish peroxidase solution (Sigma; 1 mg/mLstock solution in PBS diluted 1:400 in PBS containing 1%HI-FCS). Plates were incubated for 1 h at room temperature.After this final incubation step, the plates were first washedthree times with PBST and then washed three times with PBS.3,30-Diaminobenzidine tetra-hydrochloride (DAB) dihydratedeveloping reagent (Sigma) was prepared as recommended bythe manufacturer, and 75 lL of this substrate solution wasadded to each well and kept at room temperature until darkspots appeared (1–3 min). Colour development was stoppedby thoroughly washing the plates in tap water. The plates wereallowed to air-dry overnight. Spots were counted using acomputer-assisted ELISPOT analysis program, and data areexpressed as SFU per 1 � 106 splenocytes.
In vivo cytotoxicity assay
The in vivo cytotoxicity assay has been previously described[39]. Briefly, splenocytes from naive B6.SJL (CD45.1) micewere prepared as a single-cell suspension and depleted of RBCusing ACK lysing buffer. Cells were resuspended in completeRPMI media at 1 � 107 cells/mL and pulsed with either0.01 lM or 0.0001 lM GV001 peptide or left without peptide.Following a 1 h incubation with peptide at 37�C, each group ofsplenocytes was washed three times in complete RPMI mediabefore resuspending in PBS. The 0.01 lM GV001 target cellswere incubated with 0.02 lM CFSE, the 0.0001 lM targetswith 0.2 lM CFSE and the targets without peptide were given2 lM CFSE for 10–15 min at 37�C. Peptide-coated, CFSE-labelled targets were then washed several times beforecounting. Cells from the three groups were combined at aratio of 1:1:1, and mice were injected i.v. with a total of3 � 107 cells in 0.2 mL PBS via the tail vein. Afterapproximately 16 h, splenocytes were harvested, stained forCD45.1 and analysed for CD45.1+ CFSE+ cells using flowcytometry (FACScan, Becton Dickenson). The followingformula was used to calculate % specific lysis: [1 – Ratio innaive (control) mouse / Ratio in immunised mouse] �100,where Ratio = counts in no peptide peak / counts in peptide-pulsed peak.
Analysis of ELISPOT data
The average functional avidity of the populationwas defined asthe log of the peptide concentration that resulted in 50% of themaximal number of IFN-c-producing spots achieved viaELISPOT and was calculated using sigmoidal curve (variableslope) fits to the data (not shown). Graphs were generatedusing the Prism Graphpad program, with the error barsrepresenting the standard deviation of each sample replicate.All graphs are representatives of at least two individualexperiments.
Acknowledgements: This study was supported bygrants from the National Health and Medical ResearchCouncil of Australia, the Lions Medical ResearchFoundation and the Princess Alexandra Hospital Foun-dation.
References
1 Mason, D., A very high level of cross-reactivity is an essential feature of theT-cell receptor. Immunol. Today 1998. 19: 395–404.
2 Wilson, D. B., Wilson, D. H., Schroder, K., Pinilla, C., Blondelle, S.,Houghten, R. A. and Garcia, K. C., Specificity and degeneracy of T cells.Mol. Immunol. 2004. 40: 1047–1055.
3 Burrows, S. R., Silins, S. L., Khanna, R., Burrows, J. M., Rischmueller,M., McCluskey, J. and Moss, D. J., Cross-reactive memory T cells forEpstein-Barr virus augment the alloresponse to common human leukocyteantigens: degenerate recognition of major histocompatibility complex-bound peptide by T cells and its role in alloreactivity. Eur. J. Immunol. 1997.27: 1726–1736.
4 Selin, L. K., Nahill, S. R. and Welsh, R. M., Cross-reactivities in memorycytotoxic T lymphocyte recognition of heterologous viruses. J. Exp. Med.1994. 179: 1933–1943.
Sharmal Narayan et al. Eur. J. Immunol. 2007. 37: 406–415414
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

5 Haanen, J. B., Wolkers, M. C., Kruisbeek, A. M. and Schumacher, T. N.,Selective expansion of cross-reactive CD8(+) memory T cells by viralvariants. J. Exp. Med. 1999. 190: 1319–1328.
6 McHeyzer-Williams,M. G. and Davis, M.M.,Antigen-specific developmentof primary and memory T cells in vivo. Science 1995. 268: 106–111.
7 Alexander-Miller, M. A., Leggatt, G. R. and Berzofsky, J. A., Selectiveexpansion of high- or low-avidity cytotoxic T lymphocytes and efficacy foradoptive immunotherapy. Proc. Natl. Acad. Sci. USA 1996. 93: 4102–4107.
8 Bullock, T. N., Mullins, D. W. and Engelhard, V. H., Antigen densitypresented by dendritic cells in vivo differentially affects the number andavidity of primary, memory, and recall CD8+ Tcells. J. Immunol. 2003. 170:1822–1829.
9 Hofstetter, H. H., Targoni, O. S., Karulin, A. Y., Forsthuber, T. G., Tary-Lehmann, M. and Lehmann, P. V.,Does the frequency and avidity spectrumof the neuroantigen-specific T cells in the blood mirror the autoimmuneprocess in the central nervous system of mice undergoing experimentalallergic encephalomyelitis? J. Immunol. 2005. 174: 4598–4605.
10 Derby, M., Alexander-Miller, M., Tse, R. and Berzofsky, J., High-avidityCTL exploit two complementary mechanisms to provide better protectionagainst viral infection than low-avidity CTL. J. Immunol. 2001. 166:1690–1697.
11 Zeh, H. J. 3rd, Perry-Lalley, D., Dudley, M. E., Rosenberg, S. A. and Yang,J. C., High avidity CTLs for two self-antigens demonstrate superior in vitroand in vivo antitumor efficacy. J. Immunol. 1999. 162: 989–994.
12 Dutoit, V., Rubio-Godoy, V., Dietrich, P. Y., Quiqueres, A. L., Schnuriger,V., Rimoldi, D., Lienard, D. et al., Heterogeneous T-cell response to MAGE-A10(254–262): high avidity-specific cytolytic T lymphocytes show superiorantitumor activity. Cancer Res. 2001. 61: 5850–5856.
13 Amrani, A., Verdaguer, J., Serra, P., Tafuro, S., Tan, R. and Santamaria,P., Progression of autoimmune diabetes driven by avidity maturation of a T-cell population. Nature 2000. 406: 739–742.
14 Lustgarten, J., Dominguez, A. L. and Cuadros, C., The CD8+ T cellrepertoire against Her-2/neu antigens in neu transgenic mice is of lowavidity with antitumor activity. Eur. J. Immunol. 2004. 34: 752–761.
15 Morgan, D. J., Kreuwel, H. T., Fleck, S., Levitsky, H. I., Pardoll, D. M. andSherman, L. A., Activation of low avidity CTL specific for a self epitoperesults in tumor rejection but not autoimmunity. J. Immunol. 1998. 160:643–651.
16 Estcourt, M. J., Ramsay, A. J., Brooks, A., Thomson, S. A., Medveckzy, C.J. and Ramshaw, I. A., Prime-boost immunisation generates a highfrequency, high-avidity CD8(+) cytotoxic T lymphocyte population. Int.Immunol. 2002. 14: 31–37.
17 Oh, S., Hodge, J. W., Ahlers, J. D., Burke, D. S., Schlom, J. and Berzofsky,J. A., Selective induction of high avidity CTL by altering the balance ofsignals from APC. J. Immunol. 2003. 170: 2523–2530.
18 Oh, S., Perera, L. P., Burke, D. S., Waldmann, T. A. and Berzofsky, J. A.,IL-15/IL-15Ralpha-mediated avidity maturation of memory CD8+ T cells.Proc. Natl. Acad. Sci. USA 2004. 101: 15154–15159.
19 Brehm, M. A., Selin, L. K. and Welsh, R. M., CD8 T cell responses to viralinfections in sequence. Cell Microbiol. 2004. 6: 411–421.
20 Selin, L. K. and Welsh, R. M., Plasticity of T cell memory responses toviruses. Immunity 2004. 20: 5–16.
21 Selin, L. K., Lin, M. Y., Kraemer, K. A., Pardoll, D. M., Schneck, J. P.,Varga, S. M., Santolucito, P. A. et al., Attrition of T cell memory: selectiveloss of LCMV epitope-specific memory CD8 T cells following infections withheterologous viruses. Immunity 1999. 11: 733–742.
22 Selin, L. K., Varga, S. M., Wong, I. C. and Welsh, R. M., Protectiveheterologous antiviral immunity and enhanced immunopathogenesismediated by memory Tcell populations. J. Exp. Med. 1998. 188: 1705–1715.
23 Bihl, F., Frahm, N., Di Giammarino, L., Sidney, J., John, M., Yusim, K.,Woodberry, T. et al., Impact of HLA-B alleles, epitope binding affinity,functional avidity, and viral coinfection on the immunodominance of virus-specific CTL responses. J. Immunol. 2006. 176: 4094–4101.
24 Tian, J., Gregori, S., Adorini, L. and Kaufman, D. L., The frequency of highavidity T cells determines the hierarchy of determinant spreading. J.Immunol. 2001. 166: 7144–7150.
25 Vierboom, M. P., Feltkamp, M. C., Neisig, A., Drijfhout, J. W.,ter Schegget, J., Neefjes, J. J., Melief, C. J. and Kast, W. M., Peptidevaccination with an anchor-replaced CTL epitope protects against humanpapillomavirus type 16-induced tumors expressing the wild- type epitope. J.Immunother. 1998. 21: 399–408.
26 Leggatt, G. R., Narayan, S., Fernando, G. J. and Frazer, I. H., Changes topeptide structure, not concentration, contribute to expansion of the lowestavidity cytotoxic T lymphocytes. J. Leukoc. Biol. 2004. 76: 787–795.
27 Gallimore, A., Dumrese, T., Hengartner, H., Zinkernagel, R. M. andRammensee, H. G., Protective immunity does not correlate with thehierarchy of virus- specific cytotoxic T cell responses to naturally processedpeptides. J. Exp. Med. 1998. 187: 1647–1657.
28 Molldrem, J. J., Lee, P. P., Kant, S., Wieder, E., Jiang, W., Lu, S., Wang, C.and Davis, M. M., Chronic myelogenous leukemia shapes host immunity byselective deletion of high-avidity leukemia-specific T cells. J. Clin. Invest.2003. 111: 639–647.
29 Woodberry, T., Gardner, J., Elliott, S. L., Leyrer, S., Purdie, D. M.,Chaplin, P. and Suhrbier, A., Prime boost vaccination strategies: CD8 Tcellnumbers, protection, and Th1 bias. J. Immunol. 2003. 170: 2599–2604.
30 Cawthon, A. G. and Alexander-Miller, M. A.,Optimal colocalization of TCRand CD8 as a novel mechanism for the control of functional avidity. J.Immunol. 2002. 169: 3492–3498.
31 Cawthon, A. G., Kroger, C. J. and Alexander-Miller, M. A., High avidityCD8+ T cells generated from CD28-deficient or wildtype mice exhibit adifferential dependence on lipid raft integrity for activation. Cell Immunol.2004. 227: 148–155.
32 Welsh, R. M. and Selin, L. K., No one is naive: the significance ofheterologous T-cell immunity. Nat. Rev. Immunol. 2002. 2: 417–426.
33 Pearson, C. I., van Ewijk, W. and McDevitt, H. O., Induction of apoptosisand T helper 2 (Th2) responses correlates with peptide affinity for the majorhistocompatibility complex in self-reactive Tcell receptor transgenic mice. J.Exp. Med. 1997. 185: 583–599.
34 Bakker, A. B., van der Burg, S. H., Huijbens, R. J., Drijfhout, J. W., Melief,C. J., Adema, G. J. and Figdor, C. G., Analogues of CTL epitopes withimproved MHC class-I binding capacity elicit anti-melanoma CTL recogniz-ing the wild-type epitope. Int. J. Cancer 1997. 70: 302–309.
35 Fernando, G. J., Stewart, T. J., Tindle, R. W. and Frazer, I. H., Th2-typeCD4+ cells neither enhance nor suppress antitumor CTL activity in a mousetumor model. J. Immunol. 1998. 161: 2421–2427.
36 Widmann, C., Maryanski, J. L., Romero, P. and Corradin, G., Differentialstability of antigenic MHC class I-restricted synthetic peptides. J. Immunol.1991. 147: 3745–3751.
37 Carvalho, L. H., Hafalla, J. C. and Zavala, F., ELISPOT assay to measureantigen-specific murine CD8(+) Tcell responses. J. Immunol. Methods 2001.252: 207–218.
38 Fernando, G. J., Khammanivong, V., Leggatt, G. R., Liu, W. J. and Frazer,I. H., The number of long-lasting functional memory CD8+ Tcells generateddepends on the nature of the initial nonspecific stimulation. Eur. J. Immunol.2002. 32: 1541–1549.
39 Oehen, S., Brduscha-Riem, K., Oxenius, A. and Odermatt, B., A simplemethod for evaluating the rejection of grafted spleen cells by flow cytometryand tracing adoptively transferred cells by light microscopy. J. Immunol.Methods 1997. 207: 33–42.
Eur. J. Immunol. 2007. 37: 406–415 Immunomodulation 415
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu





























