Embed Size (px)
Citation preview

Identification and Characterization of a SquamateReptilian Amelogenin gene: Iguana iguana
XINPING WANG1, JI-LAO FAN1, YOSHIHIRO ITO1, XIANGHONG LUAN1,AND THOMAS G.H. DIEKWISCH1�1Department of Oral Biology and Allan G. Brodie Laboratory for CraniofacialGenetics, College of Dentistry, University of Illinois at Chicago, Chicago,Illinois 60612
ABSTRACT As the principal components of the developing tooth enamel matrix, amelogeninsplay a significant role in tooth enamel formation and organization. In order to elucidate the structureand function of amelogenins in the evolution of enamel, we have selected the Iguana iguana as asquamate model organism. Here we report the first complete squamate amelogenin sequenceavailable as of yet and document unique features of Iguana amelogenins and enamel. Transmissionelectron microscopy documented randomly oriented Iguana enamel crystals during the elongationphase compared with organized enamel crystal patterns at comparable stages in mammals.Sequencing of PCR amplified products revealed a full-length I. iguana amelogenin cDNA containing877 nucleotides with a 564 nucleotide coding sequence encoding 187 amino acids. The homologies ofthe newly discovered I. iguana amelogenin amino acid sequence with the published mouse, caiman(Palaeosuchus), and snake (Elaphe) amelogenin were 41.3%, 53.5%, and 55.5%, respectively. OnWestern blots one major protein with a molecular weight of 24 kDa, and two minor proteins withmolecular weights of 28 and 13.5 kDa, respectively, were detected based on the cross-reactivity ofantisera against recombinant Rana pipiens amelogenin proteins. Sequence analysis revealed amoderate sequence homology between mammalian and reptilian amelogenin genes. A significantalteration was the deletion of the hydrophilic GSP sequence from exon 3 in the mouse sequenceresulting in a conversion to a hydrophobic region in Iguana. Together, these findings identified anovel amelogenin cDNA sequence in the squamate reptilian I. iguana and functional implications forthe evolution of amelogenins and enamel in squamates. J. Exp. Zool. (Mol. Dev. Evol.)306B:393– 406, 2006. r 2006 Wiley-Liss, Inc.
During vertebrate enamel formation, the ame-logenin gene provides for the majority of enamelmatrix proteins in the developing enamel. Whileamelogenins have been demonstrated to be ex-pressed in tissues outside of the enamel organ aswell (Nebgen et al., ’99; Veis et al., 2000), they areunique among mammalian proteins in that theycomprise over 90% of the enamel matrix proteinsin the developing enamel (Fincham et al., ’99). Inprevious studies, we have provided evidence forthe wide conservation of amelogenins amongdiverse vertebrates (Diekwisch et al., 2002; Satch-ell et al., 2002; Wang et al., 2005), suggesting thatthese unique proteins may play universalroles related to enamel formation in manyvertebrates. Moreover, there have been a numberof studies closely relating enamel microstructurewith the function of amelogenin gene products(Diekwisch et al., ’93; Lyngstadaas et al., ’95;Gibson et al., 2001).
It has been demonstrated that the enamelmicrostructure between mammalian and non-mammalian vertebrates is significantly differentin that mammalian enamel microstructure con-tains prismatic, long, and parallel crystallites,while non-mammalian vertebrate enamel containsprismless, long, and parallel crystallites (Sander,’97; Wood et al., ’99). It has been also suggestedthat the complex microstructure observed in manymammals may require a range of amelogeninproteins to yield a variety of HAP crystallite sizes
Published online 27 February 2006 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/jez.b.21096.
Received 26 October 2005; Accepted 13 December 2005
Grant sponsor: National Science Foundation; Grant number: MCV0242197; Grant sponsor: National Institutes of Health; Grant number:NIDCR DE 13378.�Correspondence to: T.G.H. Diekwisch, Department of Oral Biology
and Allan G. Brodie Laboratory for Craniofacial Genetics, College ofDentistry, University of Illinois at Chicago, 801 South Paulina Street,MC 841 Chicago, IL 60612. E-mail: [email protected]
r 2006 WILEY-LISS, INC.
JOURNAL OF EXPERIMENTAL ZOOLOGY (MOL DEV EVOL) 306B:393–406 (2006)

and orientations, while the simple microstructurefound in non-mammalian enamel may requirefewer and simpler proteins (Mathur and Polly,2000). Because of the close relationship betweenenamel microstructure and amelogenin proteinfunction, it is not unlikely to assume a closerelationship between the evolution of enamelmineral microstructure and the evolution of theamelogenin gene and its protein products.
Complete or partial amelogenin genes have beencloned mostly from mammalian vertebrates(Snead et al., ’85; Shimokawa et al., ’87; Gibsonet al., ’91; Salido et al., ’92; Bonass et al., ’94;Li et al., ’95; Hu et al., ’96a,b; Lyaruu et al., ’98;Toyosawa et al., ’98; Delgado et al., 2005), whileonly few reptilian and amphibian amelogeningenes have been identified so far (Ishiyama et al.,’98; Toyosawa et al., ’98; Wang et al., 2005). Thefunctions of amelogenin have been studied mainlyin mammalian vertebrates, especially in mouse,rat, and bovine (Diekwisch et al., ’93; Lyngstadaaset al., ’95; Gibson et al., 2001). Although muchprogress has been made toward our understandingof mammalian amelogenesis and previous studieshave presented the evidence for the presence ofamelogenins in reptilian teeth (Toyosawa et al.,’98; Ishiyama et al., ’98; Diekwisch et al., 2002),enamel formation and biomineralization in non-mammalian vertebrates still remains largelyunknown. Even after decades of amelogeninresearch, a complete squamate amelogenin se-quence has been curiously missing until thepresent study. Here we are presenting a completenovel squamate reptilian amelogenin gene se-quence in the Green Iguana (Iguana iguana).
MATERIALS AND METHODS
Source and isolation of the genomic DNA
I. iguana were purchased from Carolina Biolo-gical Supply Company (Burlington, NC) andeuthanized according to approved guidelines bythe UIC animal care committee. The genomicDNA was isolated using GenEluteTM MammalianGenomic DNA Miniprep (Sigma-Aldrich Co.,St. Louis, MO) following the manufacturer’sinstruction. The isolated genomic DNA was keptat �801C for future use.
Immunohistochemistryand electron microscopy
Tissues were fixed in 10% buffered formalin andprocessed for paraffin histology. Sections were
either stained using hematoxylin and eosin orprobed for cross-reactivity with amelogenins usinga primary antibody against a mouse recombinantamelogenin antigen generated in E. coli. Immu-noreactions were performed as published pre-viously (Diekwisch et al., 2002). For electronmicroscopy, tissues were fixed in Karnovsky’s,embedded in Epon 812, and cut into 60 nmsections. Thin sections were contrasted in uranylacetate and lead citrate, and observed under aJEOL 1200 electron microscope as describedpreviously (Diekwisch et al., 2002).
RNA isolation
RNA isolation was performed as previouslyreported (Wang et al., 2005). Briefly, I. iguanajaws were removed and immediately frozen inliquid nitrogen and homogenized. The homoge-nized I. iguana teeth tissue in TRI AGENTs
reagent (Sigma) was mixed with 0.2 ml of chloro-form and shaken vigorously for 30 sec. Themixture was centrifuged at 12,000g for 20 min at41C after 10 min incubation at room temperature.The aqueous phase was transferred to a freshtube, mixed with equal volume of isopropanol, andthen centrifuged at 12,000g for 20 min at 41C. Thepellet was washed with 70% ethanol and dissolvedin DEPC-treated H2O. The isolated RNA was keptat �801C for future use.
cDNA synthesis and reverse transcriptase-polymerase chain reaction
Primers were selected based on sequence analy-sis of amelogenin genes from selected speciesincluding human, mouse, snake, caiman, Xenopuslaevis, and Rana pipiens with the aid of Lasergenesoftware (DNASTAR Inc., Madison, WI). Theconsensus sequences among different species werechosen as primers for amplification of I. iguanaamelogenin gene. Reverse transcriptase reactionwas performed using SuperSriptTM II ReverseTranscriptase (Invitrogen, Carlsbad, CA). Briefly,cDNA was synthesized in a volume of 30mlcontaining 25 mM Tris-HCI, pH 8.3, 37.5 mMKCI, 1.5 mM MgCI2, 5 mM DTT, 0.25 mM each ofdATP, dCTP, dGTP, and dTTP, 40 units of RNaseinhibitor, 200 units of M-MLV reverse transcrip-tase, 5 mg of total RNA, and 2.5mM randomprimers. The cDNA synthesis was carried out at42 1C for 60 min. PCR amplification was doneusing Platiums Taq DNA polymerase (Invitro-gen). The reaction was performed in a totalvolume of 50ml containing 20 mM Tris-HCI, pH
X. WANG ET AL.394
J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.b

8.4, 50 mM KCI, 3 mM MgCI2, 0.25 mM each ofdATP, dCTP, dGTP, and dTTP, 1 unit ofPlatiums Taq DNA polymerase, 1 mM of eachprimer, and 2ml of the cDNA synthesized above.The amplification was done under the followingconditions: denaturation at 941C for 4 min; 35cycles of 941C for 40 sec, 56 1C for 1 min, and 721Cfor 1 min; and an additional extension at 721Cfor 10 min.
Cloning and sequencing
The PCR products were analyzed by electro-phoresis using a 1% agarose gel (Invitrogen),purified with QIAquicks Gel Extraction Kit(Qiagen Inc., Valencia, CA), ligated to pGEMs-TEasy vector, and transformed into JM109 compe-tent cells (Promega, Madison, WI). Transformantswere picked up, and cultivated in LB mediumcontaining ampicillin at a final concentration of50mg/ml. Recombinant plasmids were isolatedwith Wizards Plus SV Minipreps DNA Purifica-tion System (Promega), identified by enzymedigestion with EcoRI, and sequenced using theABI 377 sequencer (Northwoods DNA Inc., Becida,MN). Three colonies were selected and sequenced2 times from both orientations with either T7 orSP6 primers.
Western-blot analysis
Western-blot analysis for detection of amelogen-in expressions in I. iguana teeth tissue wasperformed as described previously (Wang andGao, 2003). Briefly, the I. iguana dental tissueswere homogenized, harvested, washed twice withphosphate-buffered saline, pelleted, and resus-pended in 400ml of sodium dodecyl sulfate samplebuffer. The samples were then subjected to 12%SDS-PAGE gel before proteins were transferred tonitrocellulose membrane (Schleicher & Schuell,Keene, NH). The immunoblot was blocked inTBST with 5% non-fat milk, reacted with primaryantibodies against R. pipiens recombinant amelo-genin antigen, then followed by probing with a1:5,000 dilution of second antibody against rabbitimmunoglobulin conjugated with alkaline phos-phatase (Sigma), and revealed by color develop-ment in NBT/BCIP substrate (Stratagene). TheI. iguana tail tissue was processed and used as anamelogenin-negative control.
Northern-blotting analysis
Total RNA was isolated by using TRI AGENTs
according to the instructions of the manufacturer
(Sigma). Northern blot analysis was performed aspreviously reported (Wang et al., 2001). Briefly,20 mg of the total RNA was fractionated on 1%agarose gel containing formaldehyde and trans-ferred to Hybond N1nylon membrane (Amer-sham Pharmacia Biotech, Piscataway, NJ). Theprobe was prepared by labeling I. iguana amelo-genin DNA with [a-32P] dCTP (PerkinElmer LifeScience, Inc. Boston, MA) using RediprimeTM IIkit (Amersham Pharmacia Biotech, Buckingham-shire, England), or labeling oligonucleotides with[g-32P] ATP (PerkinElmer Life Science) using theDNA 50-end labeling system (Promega). Hybridi-zation of the probe to the nylon membrane wasperformed at 681C or 421C for at least 3 hr withthe PerfectHyb Plus Hybridization Buffer follow-ing the manufacturer’s instructions (Sigma). Thespecific hybridization signals on the membranewere captured and analyzed with a MolecularDynamic System (Amersham Bioscience Corp,Piscataway, NJ).
Sequence analysis
The I. iguana amelogenin amino acid sequencewas deduced from the nucleotide sequence ob-tained above, and sequence analyses were per-formed to identify its features. The nucleotidesequence served as a template for searchinghomologous sequences through GenBank(www.ncbi.nlm.nih.gov). Multiple sequence align-ment analyses were performed using the ClustalW method (Thompson et al., ’94). Hydrophilicityplots were generated using the Kyte and Doolittlealgorithm (Kyte and Doolittle, ’82).
RESULTS
Tooth replacement in Iguanidpleurodont dentitions
Similar to many squamates, the I. iguanafeatured a pleurodont dentition with replacementteeth emerging apical from functional teeth(Fig. 1A). Replacement teeth contained newlyformed enamel (Fig. 1A) and were easily accessiblefor tissue preparation and electron microscopy.
The Iguana enamel layer cross-reactedwith antibodies against
mammalian amelogenins
Iguana tooth organ sections were immunor-eacted with antibodies against recombinantM179 mouse amelogenins expressed in E. coli todetermine cross-reactivity of iguana amelogenins.
AMELOGENIN SEQUENCE IN IGUANA IGUANA 395
J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.b
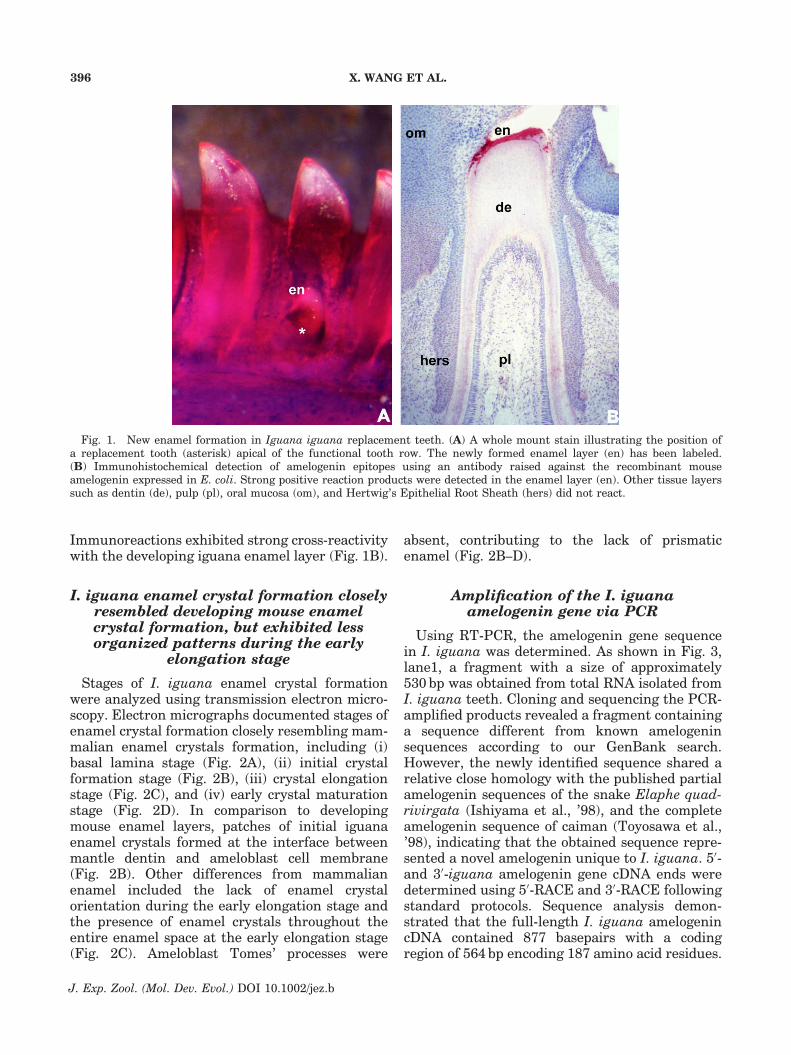
Immunoreactions exhibited strong cross-reactivitywith the developing iguana enamel layer (Fig. 1B).
I. iguana enamel crystal formation closelyresembled developing mouse enamelcrystal formation, but exhibited lessorganized patterns during the early
elongation stage
Stages of I. iguana enamel crystal formationwere analyzed using transmission electron micro-scopy. Electron micrographs documented stages ofenamel crystal formation closely resembling mam-malian enamel crystals formation, including (i)basal lamina stage (Fig. 2A), (ii) initial crystalformation stage (Fig. 2B), (iii) crystal elongationstage (Fig. 2C), and (iv) early crystal maturationstage (Fig. 2D). In comparison to developingmouse enamel layers, patches of initial iguanaenamel crystals formed at the interface betweenmantle dentin and ameloblast cell membrane(Fig. 2B). Other differences from mammalianenamel included the lack of enamel crystalorientation during the early elongation stage andthe presence of enamel crystals throughout theentire enamel space at the early elongation stage(Fig. 2C). Ameloblast Tomes’ processes were
absent, contributing to the lack of prismaticenamel (Fig. 2B–D).
Amplification of the I. iguanaamelogenin gene via PCR
Using RT-PCR, the amelogenin gene sequencein I. iguana was determined. As shown in Fig. 3,lane1, a fragment with a size of approximately530 bp was obtained from total RNA isolated fromI. iguana teeth. Cloning and sequencing the PCR-amplified products revealed a fragment containinga sequence different from known amelogeninsequences according to our GenBank search.However, the newly identified sequence shared arelative close homology with the published partialamelogenin sequences of the snake Elaphe quad-rivirgata (Ishiyama et al., ’98), and the completeamelogenin sequence of caiman (Toyosawa et al.,’98), indicating that the obtained sequence repre-sented a novel amelogenin unique to I. iguana. 50-and 30-iguana amelogenin gene cDNA ends weredetermined using 50-RACE and 30-RACE followingstandard protocols. Sequence analysis demon-strated that the full-length I. iguana amelogenincDNA contained 877 basepairs with a codingregion of 564 bp encoding 187 amino acid residues.
Fig. 1. New enamel formation in Iguana iguana replacement teeth. (A) A whole mount stain illustrating the position ofa replacement tooth (asterisk) apical of the functional tooth row. The newly formed enamel layer (en) has been labeled.(B) Immunohistochemical detection of amelogenin epitopes using an antibody raised against the recombinant mouseamelogenin expressed in E. coli. Strong positive reaction products were detected in the enamel layer (en). Other tissue layerssuch as dentin (de), pulp (pl), oral mucosa (om), and Hertwig’s Epithelial Root Sheath (hers) did not react.
X. WANG ET AL.396
J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.b

The 50-untranslated region was 78 bp upstream ofthe translation start site ATG, the 30-untranslatedregion was 235 bp downstream from the stopcodon, and the AATAAA polyadenylation signalwas located 74 bp upstream of the poly (A) tail(Fig. 4). The deduced I. iguana amelogeninconsisted of 187 amino acid residues with apredicted molecular weight of about 21.5 kDaand a predicted isoelectric point of 7.0. Amino
acid composition analysis established that thenewly sequenced I. iguana amelogenin was en-riched in Proline (37/187), Glutamine (21/187),Methionine (16/187), Histidine (13/187), Glycine(13/187), and Leucine (13/187). Beginningfrom the position of amino acid 141, therewere 11 consecutive triplex (PXX) repeats in theregion corresponding to mammalian amelogeninexon 6.
Fig. 2. Stages of enamel biomineralization in Iguana iguana as visualized via transmission electron microscopy. (A) Positionof the intact ameloblast basal lamina (bl) prior to terminal ameloblast differentiation. The basal lamina was at the interfacebetween ameloblasts (amel) and mineralized mantle dentin (md). (B) Electron micrograph of the onset of enamel crystalformation in the iguana. Initial iguana enamel crystals formed at the interface (if) between ameloblasts (amel) and dentin (de).(C) Initial stage of enamel crystal elongation in the developing iguana enamel. Note that the entire enamel layer (en) containedshort and irregular oriented crystals. (D) Elongating and maturing iguana enamel crystals (en) in adjacency to the ameloblastcells (amel). Note the striking parallel orientation of the maturing enamel crystals (en) closely resembling maturing mammalianenamel crystals at similar stages of development.
AMELOGENIN SEQUENCE IN IGUANA IGUANA 397
J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.b

I. iguana amelogenin gene consistsof six exons with a unique exon 3
To determine the organization of the I. iguanaamelogenin gene, five pairs of I. iguana amelo-genin gene-specific primers were designed andused to define I. iguana amelogenin exon/intronboundaries. The primers for defining exon/intronboundaries are shown in Table 1. After PCRamplification, five fragments with sizes of about2.0, 1.4, 0.7, 2.2, and 1.4 kb were obtained (Fig. 5).Sequencing the PCR-amplified products revealedthe size of each exon and intron as well as theexon/intron boundary sequences as shown inTable 2. The organization of the I. iguanaamelogenin gene is illustrated in Figure 6.I. iguana amelogenin gene sequence analysisrevealed that the full-length I. iguana amelogeninsequence consisted of six exons corresponding tothe mammalian amelogenin exons 1, 2, 3, 5, 6, and7. Similar to other non-mammalian species, nosequence was detected corresponding to mamma-lian amelogenin exon 4. However, exon 3 of theI. iguana amelogenin gene was unique, in that itcontained only 39 bp encoding for 13 amino acidresidues, which was three amino acids less than
the 16 amino acids observed in exon 3 of all otherspecies available so far.
Amelogenin amino acid sequenceis relatively conserved between
mammalian and reptilian
To determine the degree of similarity between asquamate reptilian (I. iguana) and a mammalian(mouse) amelogenin sequence, the I. iguanaamelogenin amino acid sequence was aligned withthose of caiman (Toyosawa et al., ’98), snake(Ishiyama et al., ’98), and mouse (Snead et al., ’85)by the Clustal W Method (Thompson et al., ’94).As shown in Figure 7, several regions were highlyconserved between I. iguana, snake, caiman, andmouse. The N-terminal 61 amino acids of I. iguanaamelogenin shared about 64% homology withmouse amelogenin, and 69% with caiman amelo-genin. Compared with the mouse and caimanamino acid sequence, the I. iguana amelogeninamino acid sequence revealed several deletionsand variations, especially in the center of exon 6.Further analysis indicated that the full-lengthI. iguana amelogenin amino acid sequence shareda moderate homology with partial snake amelo-genin (55.5%), caiman amelogenin (53.5%), andmouse amelogenin (41.3%). Taken together, theseresults established that mammalian and reptilianamelogenin amino acid sequences were moder-ately conserved among each other.
Neighbor-joining analysis placedI. iguana, caiman, and snake in the sameclade and revealed other distinct clades formammalian and amphibian amelogenins
To explore the origin of the I. iguana amelogen-in gene, a phylogenetic analysis of I. iguanaamelogenin in relationship to amelogenins fromselected other species such as cow, pig, horse,human, mouse, rat, guinea pig, caiman, snake,Xenopus, and R. pipiens was performed using theneighbor-joining method (Saitou and Nei, ’87).The sequence analysis resulted in a phylogenetictree with three distinct clades (Fig. 8). The first ofthese three clades was comprised of mammalianamelogenins from cow, pig, horse, human, mouse,rat, and guinea pig; the second clade was formedby the two amphibian amelogenin sequences fromXenopus and R. pipiens; and the third cladecontained the reptilian amelogenins fromI. iguana, snake, and caiman. Based on thephylogenetic analysis, the newly identified I.iguana amelogenin sequence was in the same
Fig. 3. Amplification of the Iguana iguana amelogeningene by RT-PCR. The I. iguana amelogenin gene wasamplified from Iguana tooth organs using RT-PCR withprimers selected in the conserved region. A fragment withthe size of 530 bp was amplified (lane 1). The 1 kb plusTM DNAladder (Invitrogen) was used as a standard to estimate the sizeof the PCR products (lane 2). Lane 3 was the negative control.
X. WANG ET AL.398
J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.b

clade with the caiman and snake amelogenins.Moreover, the amelogenin phylogenetic analysisclearly revealed two branches of the reptilianamelogenins: One branch comprised of Squamataincluding I. iguana and snake and a second branchcomprised of crocodilians such as the caiman.
I. iguana amelogenin shares a highlyconserved hydropathic pattern with
amelogenins from other vertebrate species
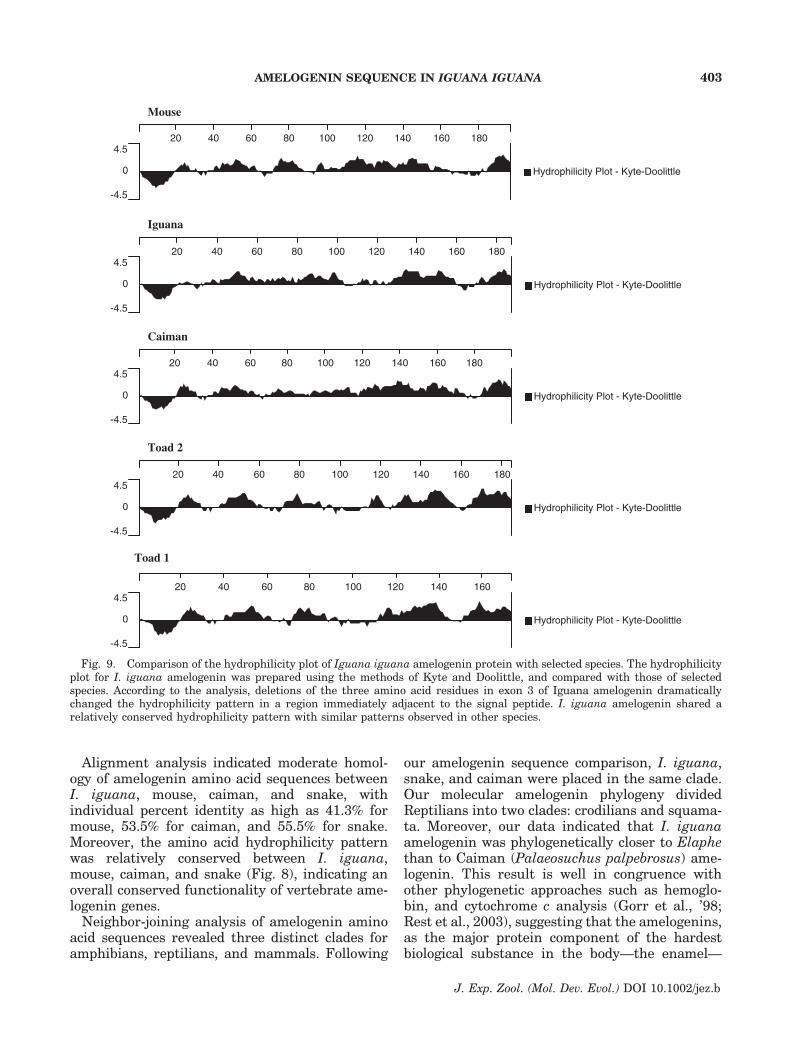
To study the conservation of the amelogeninhydrophilicity pattern in the case of I. iguana,
a hydrophilicity plot was prepared for I. iguanaamelogenin using the methods of Kyte andDoolittle (’82), and compared with amelogeninhydrophilicity plots for other amphibian, reptilian,and mammalian species. As shown in Figure 9, theI. iguana amelogenin gene sequence contained arelatively conserved hydrophilicity pattern thatover considerable stretches resembled the amelo-genin hydrophilicity pattern observed in otherspecies such as caiman, mouse, and Xenopuslaevis. However, there was a significant changein the Iguana amelogenin hydrophilicity patterncompared with those of caiman, mouse, and
ATAAACAACCAAGGCCAC 18
TGGGTCAAAACAGATATCAAGAATCCAAGAAAGCTTCTAAAAGGACCTGCAATGAGGAAG 78
Exon3 ATGGGGGGCTGGACTTTGGTTATGTGCCTCTTATGTACCACATTGGCTATTCCATTGCCA 138 M G G W T L V M C L L C T T L A I P L P 20
CAGCATCCTGGTTTTATCAATTTCAGTTATGAGGTAATGACACCTTTAAAATGGTACCAG 198 Q H P G F I N F S Y E V M T P L K W Y Q 40
AGCCTAATGGGACCCCAGTATCCACGTTATGGTTATGAACCAATGGGAGGATGGATGCGC 258 S L M G P Q Y P R Y G Y E P M G G W M R 60
CATGCTACAGGACCAATGATGCACCAAACTCACATTCAAAACCTTCCTCCAATGCACTCG 318 H A T G P M M H Q T H I Q N L P P M H S 80
CCTATCCATCAGATGCAGCCACAGCACCAAATTCTGAACCCACAAATGCCACAACCAGGG 378 P I H Q M Q P Q H Q I L N P Q M P Q P G 100
CACAACGCATTTGTGCCACTGCCTGGTCAAAATACATTGATGCCGCAGTTTCAACCAGTT 438 H N A F V P L P G Q N T L M P Q F Q P V 120
CATGGAGCAGCACCACCACAAATAGGAGAGCACCCAATGCACCCCCAGCAACATGGAAAT 498 H G A A P P Q I G E H P M H P Q Q H G N 140
CCAATGCAGCCAATCCACCCTCAGCAACCAGCAAATCCAAATCCACCAATGTACCCAGTT 558 P M Q P I H P Q Q P A N P N P P M Y P V 160
CAGCCACTTGCACCAGTGTTCCCAGACACACCTCTTGAATCGTGGCCAGCAGCAGACAAG 618 Q P L A P V F P D T P L E S W P A A D K 180
ACCAAGCAGGAAGAAGTGGACTAAAGGACCCAAGAAAACAGAATAATGCAAAGAATAATT 678 T K Q E E V D 187
TAGGCCTTGTCACTTGCTTTCAGGAACCcATATTTTTACTGCAGTCAGTGATATATATGG 738 TTCCATTAACCCACATTCTTAGCAATTATTTTAGCTAGATGGGAAACAAATATTAAGTAG 798 CAAATAATAAAATATGTTTTGATCAATAAAAAAAAAACCTATAGTGAGTCGTATTAATTC 858 GGATCCGCGAATCCTAAT 876
Exon2
Exon5
Exon6
Exon7
Fig. 4. Full-length Iguana iguana amelogenin cDNA sequence and deduced amino acid residues. 50- and 30-end of iguanaamelogenin cDNA were determined using 50- and 30-RACE. The full-length I. iguana amelogenin cDNA sequence wasdetermined to contain 877 bp, encoding 187 amino acid residues. The 50-untranslated region contains 78 bp upstream of ATGtranslation start site. The 30-untranslated region contains 235 bp. The underlined AATAAA is the polyadenylation signal located74 bp upstream of the poly-(A) tail.
AMELOGENIN SEQUENCE IN IGUANA IGUANA 399
J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.b

Xenopus laevis due to the lack of three amino acids(GSP, mouse AA 24–26, center of exon 3) in aregion immediately following the signal peptide.In addition, there was great variation between the
sequences of mouse, caiman, toad, and iguana inthe region between residues 60 and 130, a regionencoded by the b–c segment of exon 6, which hasbeen considered a ‘‘hotspot’’ in enamel evolution.
Amelogenin expressionin I. iguana tooth organ
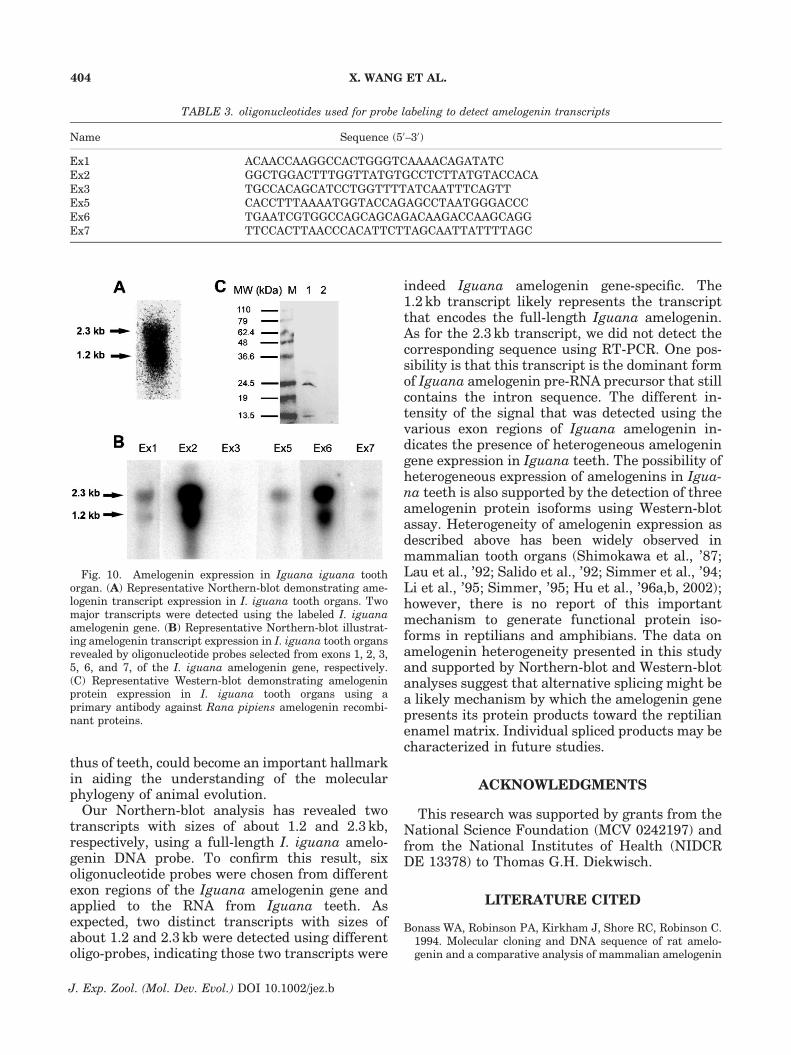
The I. iguana amelogenin gene expression intooth organs was determined via Northern andWestern Blotting. Labeled I. iguana amelogeninprobe was used to hybridize RNA extracts isolatedfrom I. iguana tooth organs resulting in twoobvious transcripts, with the size of 1.2 and2.3 kb, respectively (Fig. 10A). To further confirmthe specificity of these results, oligonucleotideswere generated based on sequence fragments fromeach exon of the I. iguana amelogenin gene(Table 3), labeled with [g-32P] ATP, and used tohybridize the membranes with equal loading ofI. iguana tooth RNA. As shown in Figure 10B,when the oligo-probes (Ex1–Ex7) were applied tothe membranes, two distinct transcripts with sizeof 1.2 and 2.3 kb, respectively, were also revealed.However, the intensity of the signals variedsignificantly using different oligo-probes.
To further verify the amelogenin expressionpattern in tooth organs, a Western-Blot assay wasperformed using an antibody against R. pipiensamelogenin recombinant protein. As shown inFigure 10C, a major protein with a molecularweight of about 24 kDa, and two minor proteinswith molecular weights of about 28 and 13.5 kDawere revealed, respectively. The amelogenin-ne-gative control samples did not react with theantibody against R. pipiens amelogenin recombi-nant proteins. These results confirmed the speci-ficity and heterogeneous expression pattern of theamelogenin gene in I. iguana teeth.
TABLE 1. Oligonucleotide primers used in determination of the exon and intron boundaries
Primer name Sequence (50–30) Orientation
Ex1S CCAAGGCCACTGGGTCAAAACAGATATC SEx2A GTGGTACATAAGAGGCACATAACCAAAGTCC AEx2S GGACTTTGGTTATGTGCCTCTTATGTACCAC SEx3A CTGAAATTGATAAAACCAGGATGCTGTGG AEx3S CCACAGCATCCTGGTTTTATCAATTTCAG SEx5A GGGTCCCATTAGGCTCTGGTACCATTTTAAAGG AEx5S CCTTTAAAATGGTACCAGAGCCTAATGGGACCC SEx6A CATTGGTCCTGTAGCATGGCGCATCCATCC AEx6S TCGTGGCCAGCAGCAGACAAGACCAAGCAGG SEx7A GCAAGTGACAAGGCCTAAATTATTCTTTGC A
S stands for sense; A refers to antisense.
Fig. 5. Amplification of the exon/intron boundary regionsby PCR. Genomic DNA was isolated and used as a template toamplify the exon/intron junctions using primers shown inTable 1. Lanes 1, 2, 3, 5, and 6 illustrating the amplified intronfragments in the junctions between exon1/2, exon2/3, exon3/5,exon5/6, and exon6/7, respectively. Lane 4 is the 1 kb plusTM
DNA ladder (Invitrogen).
X. WANG ET AL.400
J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.b

DISCUSSION
In this study, we have identified and character-ized the first complete squamate amelogenin genesequence using the Green Iguana (I. iguana) as amodel organism. Sequencing PCR-amplified frag-ments revealed a unique amelogenin sequencethat did not match any existing amelogenin genesequence and at the same time displayed closeresemblances with published partial amelogeninsequences from other reptilians. After furtheranalysis, this cDNA turned out to be most closelyrelated to a published partial snake amelogeninsequence (Ishiyama et al., ’98) with a homology of55.5%, and a complete caiman amelogenin se-quence (Toyosawa et al., ’98) with a homology of53.5%. The close homology between I. iguana,Elaphe snake, and caiman amelogenin sequence aswell as its difference from published mammaliansequences reassures our result as the discovery ofa new squamate amelogenin gene.
Amelogenins are the principal enamel matrixproteins and have been demonstrated to play acrucial role in enamel formation (Diekwisch et al.,’93; Gibson et al., 2001; Iijima et al., 2002).Enamel biomineralization in reptilians is ofsignificance to our understanding of vertebrate
hard tissue evolution not only because reptilianshave a unique linkage position between theenormous diversity of tooth minerals in amphi-bians and the relatively homogeneous presenta-tion of mammalian enamel biomineralizationpatterns, but also because of the pivotal positionof reptilian enamel microstructure as a turningpoint from non-prismatic to prismatic enamelcrystal organization. It has been suggested thatenamel microstructure might be correlated withamelogenin sequence components, and that thecomplex enamel microstructure observed in manymammals may require a range of amelogeninproteins to yield a variety of HAP crystallite sizeand orientation, while the simple microstructurefound in non-mammalian enamel may just requirefewer and simpler proteins (Mathur and Polly,2000). Therefore, availability of reptilian amelo-genin sequences could provide important informa-tion to unveil the evolution of vertebrate enamelmicrostructure.
Sequence analysis revealed that the I. iguanaamelogenin consists of six exons corresponding tomammalian amelogenin exons 1, 2, 3, 5, 6, and 7.Similar to findings from amphibian species such asR. pipiens (Wang et al., 2005) and Xenopus laevis(Toyosawa et al., ’98), the Iguana and otherreptilians were lacking sequence elements corre-sponding to amelogenin exon 4 of mammalianvertebrates I. iguana. In addition, exon 4 has onlybeen detected in a few mammalian vertebrates(Simmer et al., ’94; Simmer, ’95; Oida et al., ’96;Yuan et al., ’96; Hu et al., 2002), but is lacking inreptilian and amphibians, suggesting that exon 4might have evolved very recently.
The I. iguana amelogenin is unique in that itsexon 3 contains only 13 amino acids instead of thehighly conserved 16 amino acids reported in allother species available in GenBank so far. Kyteand Doolittle hydrophilicity algorithms reveal that
TABLE 2. Exon– intron sequence organization of the iguana amelogenin gene
Size Sequence of exon–intron junction
Exon Exon (bp) Intron (kb) Splice donor Splice acceptor
1 61 2.0 AAAG gtaaatgtatacttyy.ttacag GAC2 71 1.65 TCCA gtaagtactagttayy.ctacag TTG3 39 0.69 TGAG gtaccattgtttttyy.cccaag GTA5 45 2.2 CCAG gtatgtcctctatayy.taaaag TAT6 420 1.4 AGTG gtaagaatttctgtyy.ttccag GAC7 240
Exon and intron sequences are indicated in upper-case and low-case letters, respectively.
Fig. 6. Iguana amelogenin gene cDNA structure. The full-length Iguana iguana amelogenin cDNA contains six exonscorresponding to mammalian amelogenin exons 1, 2, 3, 5, 6,and 7 (the designation of exon numbers is based on publishedmammalian amelogenin exon numbers, e.g., Gibson et al., ’91;Simmer et al., ’95). Similar as in other non-mammalianspecies, no corresponding sequence elements were detectedresembling exon 4.
AMELOGENIN SEQUENCE IN IGUANA IGUANA 401
J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.b

this lack of three amino acid residues in exon 3 ofthe I. iguana amelogenin dramatically changes thehydrophilicity pattern in a region that immedi-ately follows the signal peptide in the N-terminal
region. We suggest that the absence of these threeamino acids residues within the I. iguana amelo-genin exon 3 is related to the uniqueness ofI. iguana enamel microstructure.
10 20 30 40 50 ------------------+-------------------+-------------------+-------------------+-------------------+- 1 M G G W T L V M C L L C T T L A I P L P Q H P - - - G F I N F S Y E V M T P L K W Y Q S L M G P Q Y Iguana 1 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - * * * * * * * F * * I I * H * * Elaphe(AAD22553) 1 * E * * M * I T * * * G A * F * * * * * P * * H H P * Y V * * * * * * L * * * * * * * * * * R Q P * Caiman(AAC78133) 1 * * T * I * F A * * * G A A F * M * * * P * * G S P * Y * * L * * * * L * * * * * * * * M I R Q P * Mouse(BAA06546)
60 70 80 90 100 ------------------+-------------------+-------------------+-------------------+-------------------+- 51 P R Y G Y E P M G G W M R H A T G P M M H Q T H I Q N L P P M H S P I H Q M Q P Q H - - - - - - Q I Iguana 17 * * * * * * * * * * * * H * N A * * * * * P P * F H G V H A I * P A L * * K Q * * Q - - - - - - L P Elaphe(AAD22553) 51 S S * * * * * * * * * L H Q P M L * - I A * Q * P P - I Q T L T P - - * H Q I * F L - - - - - - S P Caiman (AAC78133) 51 * S * * * * * * * * * L H * Q I I * V L S * Q * P P - S H T L Q P - - * H H L * V V P A Q Q P V A P Mouse(BAA06546)
110 120 130 140 150 ------------------+-------------------+-------------------+-------------------+-------------------+- 95 L N P Q M P Q P G H N A F V P L P G Q - - N T L M P Q F Q P V H G - - - - - - - - A A P P Q I - - - Iguana 61 Q * * * L Q * * * L * * * * * * N * H - - * A * I * H Y * - - * - - - - - - - - - P F Q * * A - - - Elaphe(AAD22553) 91 Q H * L * Q M * * P H Q M M * I * Q * Q P S L Q * * V Q E * * Q P Q A G E H P S P P V Q * * Q - P G Caiman (AAC78133) 98 Q Q * M * * V * * * H S M T * T Q H H Q P * I P P S A Q * * F Q - - - - - - - - Q P F Q * * A I P P Mouse(BAA06546)
160 170 180 190 200 ------------------+-------------------+-------------------+-------------------+-------------------+- 132 - G E H P M H P Q Q H G N P M Q P I H P Q Q P A N P N P P M Y P V Q P L A P V F P D T P L E S W P A Iguana 95 - * * * * I Q S L S - - - * V N * M N * * * * G * A * Q * I * - - - * * P * * I * * * * * * * * * - Elaphe(AAD22553)140 H P N P * * Q P * L P * S * H P * M R * * * * G I * * * * * * * M * * * P * L L * * M * * * P * R P Caiman (AAC78133)140 Q S H Q * * Q * * - - - S * L H * M Q * L A * Q P * L * * L F S M * * * S * I L * E L * * * A * * * Mouse(BAA06546)
210 ------------------+- 181 A D K T K Q E E V D Iguana 137 * * * * * K * * * * Elaphe(AAD22553)190 M * * * * * * * I * Caiman (AAC78133)187 T * * * * R * * * * Mouse(BAA06546)
Fig. 7. Alignment of Iguana iguana amelogenin amino acid sequences with the selected species. I. iguana amelogenin aminoacid sequence was aligned with those of Caiman (AAC78133), Elaphe (AAD22553), and mouse (BAA06546) using the Clustal WMethod with the following parameters: gap penalty, 10.0; gap length penalty, 0.20. I. iguana amelogenin sequence is shownat the top. Identical residues are marked with an asterisk (*). Alignment gaps are indicated by a dash (–). The species namesare shown with a GenBank accession number.
Nucleotide Substitutions (x100)0
64.6
102030405060
Cow(AAA30372)Sus scrofa (AAB41111)Equus caballus (BAA84219)Human(AAL30432)Rat(AAA20491 )Mouse(BAA06546)Guinea pig (CAA09957)IguanaElaphe(AAD22553)Paleosuchus palpebrosus(AAC78133)Xenopus laevis(ACC78134)Xenopus laevis(ACC78135)Frog
Fig. 8. Phylogenetic analysis of Iguana iguana amelogenin in comparison with known amelogenin sequences from selectedvertebrate species. A phylogenetic tree (rooted tree) generated by MegAlign based on the amelogenin protein sequences ofI. iguana, and selected vertebrates were grouped into three distinct clades: (i) one clade for mammalian vertebrates includingcow, goat, pig, horse, human, mouse, rat, hamster, and guinea pig; (ii) a second clade for reptilians containing I. iguana, snake,and caiman; and (iii) a third clade for amphibians including Xenopus and the leopard frog. Note that the newly identifiedI. iguana amelogenin sequence is in the same clade as the already published partial sequence for the Elaphe snake and thecomplete sequence for the caiman (Palaeosuchus palpebrosus). Dotted lines indicate a negative branch length that may resultfrom averaging. Our analysis also demonstrates that the I. iguana amelogenin sequence is much closer to the Elaphe snake thanto the caiman.
X. WANG ET AL.402
J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.b
Alignment analysis indicated moderate homol-ogy of amelogenin amino acid sequences betweenI. iguana, mouse, caiman, and snake, withindividual percent identity as high as 41.3% formouse, 53.5% for caiman, and 55.5% for snake.Moreover, the amino acid hydrophilicity patternwas relatively conserved between I. iguana,mouse, caiman, and snake (Fig. 8), indicating anoverall conserved functionality of vertebrate ame-logenin genes.
Neighbor-joining analysis of amelogenin aminoacid sequences revealed three distinct clades foramphibians, reptilians, and mammals. Following
our amelogenin sequence comparison, I. iguana,snake, and caiman were placed in the same clade.Our molecular amelogenin phylogeny dividedReptilians into two clades: crodilians and squama-ta. Moreover, our data indicated that I. iguanaamelogenin was phylogenetically closer to Elaphethan to Caiman (Palaeosuchus palpebrosus) ame-logenin. This result is well in congruence withother phylogenetic approaches such as hemoglo-bin, and cytochrome c analysis (Gorr et al., ’98;Rest et al., 2003), suggesting that the amelogenins,as the major protein component of the hardestbiological substance in the body—the enamel—
20 40 60 80 100 120 140 160 180
Hydrophilicity Plot - Kyte-Doolittle0
4.5
-4.5
20 40 60 80 100 120 140 160 180
Hydrophilicity Plot - Kyte-Doolittle0
4.5
-4.5
20 40 60 80 100 120 140 160 180
Hydrophilicity Plot - Kyte-Doolittle0
4.5
-4.5
20 40 60 80 100 120 140 160 180
Hydrophilicity Plot - Kyte-Doolittle0
4.5
-4.5
20 40 60 80 100 120 140 160
Hydrophilicity Plot - Kyte-Doolittle0
4.5
-4.5
Toad 2
Toad 1
Mouse
Iguana
Caiman
Fig. 9. Comparison of the hydrophilicity plot of Iguana iguana amelogenin protein with selected species. The hydrophilicityplot for I. iguana amelogenin was prepared using the methods of Kyte and Doolittle, and compared with those of selectedspecies. According to the analysis, deletions of the three amino acid residues in exon 3 of Iguana amelogenin dramaticallychanged the hydrophilicity pattern in a region immediately adjacent to the signal peptide. I. iguana amelogenin shared arelatively conserved hydrophilicity pattern with similar patterns observed in other species.
AMELOGENIN SEQUENCE IN IGUANA IGUANA 403
J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.b
thus of teeth, could become an important hallmarkin aiding the understanding of the molecularphylogeny of animal evolution.
Our Northern-blot analysis has revealed twotranscripts with sizes of about 1.2 and 2.3 kb,respectively, using a full-length I. iguana amelo-genin DNA probe. To confirm this result, sixoligonucleotide probes were chosen from differentexon regions of the Iguana amelogenin gene andapplied to the RNA from Iguana teeth. Asexpected, two distinct transcripts with sizes ofabout 1.2 and 2.3 kb were detected using differentoligo-probes, indicating those two transcripts were
indeed Iguana amelogenin gene-specific. The1.2 kb transcript likely represents the transcriptthat encodes the full-length Iguana amelogenin.As for the 2.3 kb transcript, we did not detect thecorresponding sequence using RT-PCR. One pos-sibility is that this transcript is the dominant formof Iguana amelogenin pre-RNA precursor that stillcontains the intron sequence. The different in-tensity of the signal that was detected using thevarious exon regions of Iguana amelogenin in-dicates the presence of heterogeneous amelogeningene expression in Iguana teeth. The possibility ofheterogeneous expression of amelogenins in Igua-na teeth is also supported by the detection of threeamelogenin protein isoforms using Western-blotassay. Heterogeneity of amelogenin expression asdescribed above has been widely observed inmammalian tooth organs (Shimokawa et al., ’87;Lau et al., ’92; Salido et al., ’92; Simmer et al., ’94;Li et al., ’95; Simmer, ’95; Hu et al., ’96a,b, 2002);however, there is no report of this importantmechanism to generate functional protein iso-forms in reptilians and amphibians. The data onamelogenin heterogeneity presented in this studyand supported by Northern-blot and Western-blotanalyses suggest that alternative splicing might bea likely mechanism by which the amelogenin genepresents its protein products toward the reptilianenamel matrix. Individual spliced products may becharacterized in future studies.
ACKNOWLEDGMENTS
This research was supported by grants from theNational Science Foundation (MCV 0242197) andfrom the National Institutes of Health (NIDCRDE 13378) to Thomas G.H. Diekwisch.
LITERATURE CITED
Bonass WA, Robinson PA, Kirkham J, Shore RC, Robinson C.1994. Molecular cloning and DNA sequence of rat amelo-genin and a comparative analysis of mammalian amelogenin
TABLE 3. oligonucleotides used for probe labeling to detect amelogenin transcripts
Name Sequence (50–30)
Ex1 ACAACCAAGGCCACTGGGTCAAAACAGATATCEx2 GGCTGGACTTTGGTTATGTGCCTCTTATGTACCACAEx3 TGCCACAGCATCCTGGTTTTATCAATTTCAGTTEx5 CACCTTTAAAATGGTACCAGAGCCTAATGGGACCCEx6 TGAATCGTGGCCAGCAGCAGACAAGACCAAGCAGGEx7 TTCCACTTAACCCACATTCTTAGCAATTATTTTAGC
Fig. 10. Amelogenin expression in Iguana iguana toothorgan. (A) Representative Northern-blot demonstrating ame-logenin transcript expression in I. iguana tooth organs. Twomajor transcripts were detected using the labeled I. iguanaamelogenin gene. (B) Representative Northern-blot illustrat-ing amelogenin transcript expression in I. iguana tooth organsrevealed by oligonucleotide probes selected from exons 1, 2, 3,5, 6, and 7, of the I. iguana amelogenin gene, respectively.(C) Representative Western-blot demonstrating amelogeninprotein expression in I. iguana tooth organs using aprimary antibody against Rana pipiens amelogenin recombi-nant proteins.
X. WANG ET AL.404
J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.b

protein sequence divergence. Biochem Biophys Res Com-mun 198:755–763.
Delgado S, Girondot M, Sire JY. 2005. Molecular evolution ofamelogenin in mammals. J Mol Evol 60:12–30.
Diekwisch T, David S, Bringas P Jr, Santos V, Slavkin HC.1993. Antisense inhibition of AMEL translation demon-strates supramolecular controls for enamel HAP crystalgrowth during embryonic mouse molar development. Devel-opment 117:471–482.
Diekwisch TG, Berman BJ, Anderton X, Gurinsky B, OrtegaAJ, Satchell PG, Williams M, Arumugham C, Luan X,McIntosh JE, Yamane A, Carlson DS, Sire JY, Shuler CF.2002. Membranes, minerals, and proteins of developingvertebrate enamel. Microsc Res Tech 59:373–395.
Fincham AG, Moradian-Oldak J, Simmer JP. 1999. Thestructural biology of the developing dental enamel matrix.J Struct Biol 126:270–299.
Gibson C, Golub E, Herold R, Risser M, Ding W, ShimokawaH, Young M, Termine J, Rosenbloom J. 1991. Structure andexpression of the bovine amelogenin gene. Biochemistry 30:1075–1079.
Gibson CW, Yuan ZA, Hall B, Longenecker G, Chen E,Thyagarajan T, Sreenath T, Wright JT, Decker S, Pidding-ton R, Harrison G, Kulkarni AB. 2001. Amelogenin-deficientmice display an amelogenesis imperfecta phenotype. J BiolChem 276:31871–31875.
Gorr TA, Mable BK, Kleinschmidt T. 1998. Phylogeneticanalysis of reptilian hemoglobins: rates, and divergences.J Mol Evol 47:471–485.
Hu CC, Bartlett JD, Zhang CH, Qian Q, Ryu OH, Simmer JP.1996a. Cloning, cDNA sequence, and alternativesplicing of porcine amelogenin mRNAs. J Dent Res 75:1735–1741.
Hu CC, Zhang C, Qian Q, Ryu OH, Moradian-Oldak J,Fincham AG, Simmer JP. 1996b. Cloning, DNA sequence,and alternative splicing of opossum amelogenin mRNAs.J Dent Res 75:1728–1734.
Hu CC, Ryu OH, Yamakoshi Y, Zhang CH, Cao X, Qian Q,Simmer JP. 2002. Pig amelogenin gene expresses a uniqueexon 4. Connect Tissue Res 43:435–440.
Iijima M, Moriwaki Y, Wen HB, Fincham AG, Moradian-OldakJ. 2002. Elongated growth of octacalcium phosphate crystalsin recombinant amelogenin gels under controlled ionic flow.J Dent Res 81:69–73.
Ishiyama M, Mikami M, Shimokawa H, Oida S. 1998.Amelogenin protein in tooth germs of the snake Elaphequadrivirgata, immunohistochemistry, cloning and cDNAsequence. Arch Histol Cytol 61:467–474.
Kyte J, Doolittle RF. 1982. A simple method for displaying thehydropathic character of a protein. J Mol Biol 157:105–132.
Lau EC, Simmer JP, Bringas P Jr, Hsu DD, Hu CC, Zeichner-David M, Thiemann F, Snead ML, Slavkin HC, FinchamAG. 1992. Alternative splicing of the mouse amelogeninprimary RNA transcript contributes to amelogenin hetero-geneity. Biochem Biophys Res Commun 188:1253–1260.
Li R, Li W, DenBesten PK. 1995. Alternative splicing ofamelogenin mRNA from rat incisor ameloblasts. J Dent Res74:1880–1885.
Lyaruu DM, Hu CC, Zhang C, Qian Q, Ryu OH, Moradian-Oldak J, Woltgens JH, Fincham AG, Simmer JP. 1998.Derived protein and cDNA sequences of hamster amelogen-in. Eur J Oral Sci 106 (Suppl. 1):299–307.
Lyngstadaas SP, Risnes S, Sproat BS, Thrane PS, Prydz HP.1995. A synthetic, chemically modified ribozyme eliminates
amelogenin, the major translation product in developingmouse enamel in vivo. EMBO J 14:5224–5229.
Mathur AK, Polly PD. 2000. The evolution of enamelmicrostructure: how important is amelogenin. J MammalEvol 7:23–42.
Nebgen DR, Inoue H, Sabsay B, Wei K, Ho CS, Veis A. 1999.Identification of the chondrogenic-inducing activity frombovine dentin (bCIA) as a low-molecular-mass amelogeninpolypeptide. J Dent Res 78:1484–1494.
Oida S, Miyazaki H, Iimura T, Suzuki M, Sasaki S,Shimokawa H. 1996. Molecular structure of the mouseamelogenin genomic DNA. DNA Seq 6:307–310.
Rest JS, Ast JC, Austin CC, Waddell PJ, Tibbetts EA, Hay JM,Mindell DP. 2003. Molecular systematics of primaryreptilian lineages and the tuatara mitochondrial genome.Mol Phylogenet Evol 29:289–297.
Saitou N, Nei M. 1987. The neighbor-joining method: a newmethod for reconstructing phylogenetic trees. Mol Biol Evol4:406–425.
Salido EC, Yen PH, Koprivnikar K, Yu LC, Shapiro LJ. 1992.The human enamel protein gene amelogenin is expressedfrom both the X and the Y chromosomes. Am J Hum Genet50:303–316.
Sander PM. 1997. Non-mammalian synapsid enamel and theorigin of enamel prisms: the bottom-up perspective. In: vonKoenigswald W, Sander PM, editors. Tooth enamel micro-structure. Rotterdam: Balkema. p 42–62
Satchell PG, Anderton X, Ryu OH, Luan X, Ortega AJ,Opamen R, Berman BJ, Witherspoon DE, Gutmann JL,Yamane A, Zeichner-David M, Simmer JP, Shuler CF,Diekwisch TG. 2002. Conservation and variation in enamelprotein distribution during vertebrate tooth development.J Exp Zool 294:91–106.
Shimokawa H, Ogata Y, Sasaki S, Sobel ME, McQuillan CI,Termine JD, Young MF. 1987. Molecular cloning of bovineamelogenin cDNA. Adv Dent Res 1:293–297.
Simmer JP. 1995. Alternative splicing of amelogenins. Con-nect Tissue Res 32:131–136.
Simmer JP, Hu CC, Lau EC, Sarte P, Slavkin HC, FinchamAG. 1994. Alternative splicing of the mouse amelogeninprimary RNA transcript. Calcif Tissue Int 55:302–310.
Snead ML, Lau EC, Zeichner-David M, Fincham AG, Woo SL,Slavkin HC. 1985. DNA sequence for cloned cDNA formurine amelogenin reveal the amino acid sequence forenamel-specific protein. Biochem Biophys Res Commun 129:812–818.
Thompson JD, Higgins DG, Gibson TJ. 1994. CLUSTAL W:improving the sensitivity of progressive multiple sequencealignment through sequence weighting, position-specific gappenalties and weight matrix choice. Nucl Acids Res 22:4673–4680.
Toyosawa S, O’HUigin C, Figueroa F, Tichy H, Klein J. 1998.Identification and characterization of amelogenin genes inmonotremes, reptiles, and amphibians. Proc Natl Acad SciUSA 95:13056–13061.
Veis A, Tompkins K, Alvares K, Wei K, Wang L, Wang XS,Brownell AG, Jengh SM, Healy KE. 2000. Specific amelo-genin gene splice products have signaling effects on cellsin culture and in implants in vivo. J Biol Chem 275:41263–41272.
Wang XP, Gao SJ. 2003. Auto-activation of the transformingviral interferon regulatory factor encoded by Kaposi’ssarcoma-associated herpesvirus (human herpesvirus-8).J Gen Virol 84:329–336.
AMELOGENIN SEQUENCE IN IGUANA IGUANA 405
J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.b

Wang XP, Zhang YJ, Deng JH, Pan HY, Zhou FC, MontalvoEA, Gao SJ. 2001. Characterization of the promoter regionof the viral interferon regulatory factor encoded by Kaposi’ssarcoma-associated herpesvirus. Oncogene 20:523–530.
Wang X, Ito Y, Luan X, Yamane A, Diekwisch TG. 2005.Amelogenin sequence and enamel biomineralization inRana pipiens. J Exp Zool B Mol Dev Evol 304B:177–186
Wood CB, Dumont ER, Crompton AW. 1999. New studies ofenamel microstructure in mesozoic mammals: a review ofenamel prisms as a mammalian synapomorphy. J MammalEvol 6:177–214.
Yuan ZA, Collier PM, Rosenbloom J, Gibson CW. 1996.Analysis of amelogenin mRNA during bovine tooth deve-lopment. Arch Oral Biol 41:205–213.
X. WANG ET AL.406
J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.b
























