Embed Size (px)
Citation preview

www.elsevier.com/locate/gene
Gene 316 (2003) 157–165
Identification and characterization of the reptilian GnRH-II gene
in the leopard gecko, Eublepharis macularius, and its
evolutionary considerations$
Tadahiro Ikemoto*, Min Kyun Park
Laboratory of Endocrinology, Department of Biological Sciences, Graduate School of Science, The University of Tokyo,
7-3-1 Hongo, Bunkyo, Tokyo 113-0033, Japan
Received 7 March 2003; received in revised form 2 June 2003; accepted 17 June 2003
Received by T. Gojobori
Abstract
To elucidate the molecular phylogeny and evolution of a particular peptide, one must analyze not the limited primary amino acid
sequences of the low molecular weight mature polypeptide, but rather the sequences of the corresponding precursors from various species. Of
all the structural variants of gonadotropin-releasing hormone (GnRH), GnRH-II (chicken GnRH-II, or cGnRH-II) is remarkably conserved
without any sequence substitutions among vertebrates, but its precursor sequences vary considerably. We have identified and characterized
the full-length complementary DNA (cDNA) encoding the GnRH-II precursor and determined its genomic structure, consisting of four exons
and three introns, in a reptilian species, the leopard gecko Eublepharis macularius. This is the first report about the GnRH-II precursor
cDNA/gene from reptiles. The deduced leopard gecko prepro-GnRH-II polypeptide had the highest identities with the corresponding
polypeptides of amphibians. The GnRH-II precursor mRNA was detected in more than half of the tissues and organs examined. This
widespread expression is consistent with the previous findings in several species, though the roles of GnRH outside the hypothalamus–
pituitary–gonadal axis remain largely unknown. Molecular phylogenetic analysis combined with sequence comparison showed that the
leopard gecko is more similar to fishes and amphibians than to eutherian mammals with respect to the GnRH-II precursor sequence. These
results strongly suggest that the divergence of the GnRH-II precursor sequences seen in eutherian mammals may have occurred along with
amniote evolution.
D 2003 Elsevier B.V. All rights reserved.
Keywords: Chicken GnRH-II; Leopard gecko (Eublepharis macularius); Precursor sequence; Genomic structure; Spatial expression pattern; Phylogeny and
evolution
1. Introduction decapeptide, gonadotropin-releasing hormone, which stim-
GnRH plays a pivotal role in the regulation of reproduc-
tion. GnRH was originally identified as a hypothalamic
0378-1119/$ - see front matter D 2003 Elsevier B.V. All rights reserved.
doi:10.1016/S0378-1119(03)00758-3
Abbreviations: bp, base pair(s); cDNA, DNA complementary to RNA;
cGnRH-II, chicken GnRH-II; GAP, GnRH-associated peptide; GnRH,
gonadotropin-releasing hormone; GnRH-II, gene encoding GnRH-II; kb,
kilobase(s) or 1000 bp; NUP, Nested Universal Primer A; ORF, open
reading frame; PCR, polymerase chain reaction; RACE, rapid amplification
of cDNA ends; RT, reverse transcription; TAE, Tris-acetate-EDTA; UPM,
Universal Primer A Mix; UTR, untranslated region(s).$ The nucleotide sequence reported in this paper has been deposited in
the DDBJ/EMBL/GenBank with the accession number AB104485.
* Corresponding author. Tel./fax: +81-3-5841-4439.
E-mail address: [email protected] (T. Ikemoto).
ulates pituitary gonadotropes to synthesize and release
gonadotropins. Since the structure of the decapeptide was
determined from porcine and ovine brains (Matsuo et al.,
1971; Burgus et al., 1972), more than 20 GnRH molecules
or GnRH-like sequences have been identified thus far from
various vertebrates and some invertebrates (Adams et al.,
2003). With a given species, it is generally found that
multiple GnRH isoforms and multiple types of GnRH
receptor are distributed in a wide range of tissues. Thus,
GnRH is thought to have diverse physiological functions in
addition to gonadotropin secretion (Millar, 2003).
Of all the structural variants of GnRH, chicken GnRH-II
(cGnRH-II; first isolated from chicken brain by Miyamoto
et al., 1984) has been found to be universally present in and

T. Ikemoto, M.K. Park / Gene 316 (2003) 157–165158
uniquely conserved among vertebrates without any se-
quence substitutions (Millar and King, 1987). Chicken
GnRH-II appears in all classes of jawed vertebrates, reveal-
ing that it has remained unchanged for at least 500 million
years. This remarkable conservation suggests that it has
been highly constrained throughout evolution, apparently
indicating an important conserved role. This GnRH form
has been renamed GnRH-II (Sealfon et al., 1997).
It is generally accepted that different forms of GnRH
have arisen through gene duplications from a single ances-
tral GnRH form whose origin predates vertebrates (Millar
and King, 1987; Sherwood et al., 1993; Fernald and White,
1999). Indeed, GnRH-like immunoreactivities were reported
in several invertebrates (Tsutsui et al., 1998; Rastogi et al.,
2002), and several GnRH peptides were isolated from two
tunicates and an octopus (Craig et al., 1997; Di Fiore et al.,
2000; Iwakoshi et al., 2002). For analyses of phylogeny and
evolution based on a particular peptide, including GnRH,
the limited primary amino acid sequence of the low molec-
ular weight mature polypeptide is not useful for constructing
molecular phylogenetic trees (Dores et al., 1996). Rather,
precursor sequences from various species are needed to
achieve an understanding of GnRH evolution.
After the discovery and sequencing of the complemen-
tary DNA (cDNA) encoding the GnRH precursor in humans
(Seeburg and Adelman, 1984), a large body of GnRH
precursor sequence data from various species has been
accumulated. Precursor cDNAs for the remarkably con-
served GnRH-II have also been cloned from fishes, amphib-
ians and mammals (for details, see Section 2.7); however,
no sequence data have been reported from reptiles and birds.
Reptiles show some features of both higher and lower
vertebrates. They, together with mammals and birds, are
amniotes. On the other hand, extant reptiles share with
fishes and amphibians the characteristic of being ectother-
mic. Altogether, reptiles are important animals that may be
considered a crucial evolutionary bridge among vertebrates,
and would be absolutely indispensable for clarifying the
evolution of GnRH. The same is also true of GnRH-II,
because its mature peptide sequence remains unchanged but
its precursor sequence is remarkably divergent between
amphibians and mammals (Wang et al., 2001).
These considerations prompted us to search for the
GnRH-II precursor cDNA in reptiles. The existence of
GnRH-II in reptiles was verified by Powell et al. (1986).
Subsequently, this was confirmed by its sequencing in the
American alligator (Lovejoy et al., 1991). Here we describe
the first molecular cloning and characterization of the full-
length GnRH-II precursor cDNA from a reptilian species,
leopard gecko Eublepharis macularius. The physiology and
behavior of this reptile have been vigorously investigated
(Crews et al., 1998). Its ease of breeding and early matura-
tion also make this species a good experimental model. We
also determined the genomic structure of the GnRH-II gene.
In addition, to identify the sites where GnRH-II decapeptide
might be synthesized and to seek clues to its potential
function, the spatial expression pattern of the GnRH-II
precursor was examined by the reverse transcription–poly-
merase chain reaction (RT–PCR)-sequencing method. Fi-
nally, we constructed a molecular phylogeny of the
evolution of GnRH-II, incorporating our new sequence data
from the leopard gecko.
2. Materials and methods
2.1. Animals
Animals were treated according to the guideline of the
Bioscience Committee at the University of Tokyo. The
leopard geckos (E. macularius) were bred over several
generations in groups of one male and at least two females
in the laboratory. Fertile eggs were placed in cases filled
with moist vermiculite at a constant incubation temperature
of 29.0 jC, which produces a female-biased sex ratio
(Crews et al., 1998). One-year-old, fertile male (body
weight 80–100 g) and female (60–80 g) leopard geckos
were maintained under the conditions of 29.0 jC and a long
day photoperiod (14 h light and 10 h dark). The animals
were provided crickets and water ad libitum. Reproductive
fertility was confirmed by egg-laying and at least one
vitellogenic follicle in females in each group. Animals were
anesthetized with sodium pentobarbital (25 Ag/g) and killed
by rapid decapitation, followed by complete bloodletting.
Tissues and organs were immediately dissected and frozen
in liquid nitrogen, and stored at � 80 jC until use.
2.2. RNA preparation and cDNA synthesis
Total RNA was extracted using ISOGEN (NIPPON
GENE, Tokyo, Japan). Male whole brain cDNA used for
rapid amplification of cDNA ends (RACE; see Section 2.3)
was synthesized from 1 Ag of total RNA using a SMART
RACE cDNA Amplification Kit (BD Biosciences Clontech,
Palo Alto, CA), according to the manufacturer’s instructions.
The cDNAs used as templates for RT–PCRwere synthesized
from 3 Ag of denatured total RNA using 5 AM oligo(dT)
primer and 100 units of M-MLV Reverse Transcriptase
(Promega, Madison, WI) in a 20-Al reaction volume with
incubation at 38 jC for 30 min followed by 42 jC for 1.5 h.
2.3. Molecular cloning of the GnRH-II precursor cDNA by
RACE and RT–PCR
RACE was carried out to obtain a partial GnRH-II
precursor cDNA from male whole brain cDNA using four
degenerate primers (Table 1), Universal Primer A Mix
(UPM) and Nested Universal Primer A (NUP) (Clontech).
All of the following PCR amplifications were performed in a
20-Al reaction mixture containing each primer at 1 AM, 0.25
unit of TaKaRa Ex Taq (TaKaRa, Shiga, Japan), each dNTP
at 250 AM and Ex Taq Buffer (TaKaRa). The primary PCRs

Table 1
Oligonucleotide primers used for RACE, RT–PCR and genomic structure determination
Name Nucleotide sequencea Usage and location
01SE 5V-CAYTGGTCNCAYGGNTGGTA-3V for 3V-RACEnest02SE 5V-TGGTCNCAYGGNTGGTAYCC-3V for nested 3V-RACE03AS 5V-GGRTACCANCCRTGNGACCA-3V for 5V-RACEnest04AS 5V-TACCANCCRTGNGACCARTG-3V for nested 5V-RACEGSP-01SE 5V-GAGGCAGAAGAGCCAAGAGGTGAGG-3V for RT–PCR (in the 5V-UTR)GSP-02AS 5V-CAGACGTGAGTGAACACAGCAAGTC-3V for RT–PCR (in the 3V-UTR)EX1-12SE 5V-NNNNNTGGGATTTTCGGAGG-3V for intron A (in exon 1)
EX2-21SE 5V-GCATAATGATCATCGCCACTATCC-3V for intron B (in exon 2)
EX2-22AS 5V-GTGGCGATGATCATTATGCAGAGG-3V for intron A (in exon 2)
EX3-31SE 5V-GCTATGTGACGGGGACGACTGCAC-3V for intron C (in exon 3)
EX3-32AS 5V-GTGCAGTCGTCCCCGTCACATAGC-3V for intron B (in exon 3)
INT2-21AS 5V-GATTCGAACCATCGGTAAAGAAGTAC-3V for intron A (in intron B)
a Abbreviations for degenerate nucleotides: Y, C or T; R, G or A. N represents all four nucleotides.
T. Ikemoto, M.K. Park / Gene 316 (2003) 157–165 159
were performed with 01SE for 3V-RACE or 03AS for 5V-RACE in combination with UPM. The reaction conditions
for the primary PCR were as follows: 94 jC for 5 min, 5
cycles of 94 jC for 40 s, 67 jC for 30 s, 72 jC for 2 min, 7
cycles of 94 jC for 40 s, 65 jC for 30 s, 72 jC for 2 min, 10
cycles of 94 jC for 40 s, 63 jC for 30 s, 72 jC for 2 min, and
72 jC for 7 min. The 40-fold-diluted primary PCR solutions
served as templates for the nested PCRs with nest02SE for
3V-RACE or nest04AS for 5V-RACE in combination with
NUP. The reaction conditions for nested PCR were as
follows: 94 jC for 5 min, 5 cycles of 94 jC for 40 s, 67
jC for 30 s, 72 jC for 2 min, 15 cycles of 94 jC for 40 s, 64.5
jC for 30 s, 72 jC for 2 min, 20 cycles of 94 jC for 40 s, 62
jC for 25 s, 72 jC for 2 min, and 72 jC for 7 min. The
amplified products were separated by electrophoresis in a
1.5% Tris-acetate-EDTA (TAE) agarose gel and visualized
using ethidium bromide staining and a UV transilluminator.
DNA fragments were extracted using a QIAquick Gel
Extraction Kit (QIAGEN K.K., Tokyo, Japan) and directly
sequenced. These analyses were repeated independently
three times to avoid any PCR amplification errors.
After determination of the sequences of the 3V- and 5V-ends of the GnRH-II precursor cDNA, amplification of
cDNA fragments including the entire open reading frame
(ORF) was carried out to determine the complete sequence.
This amplification was performed with gene-specific pri-
mers, GSP-01SE complementary to the 5V-untranslatedregion (5V-UTR) and GSP-02AS complementary to the 3V-UTR (Table 1). The reaction conditions were as follows: 94
jC for 5 min, 30 cycles of 94 jC for 40 s, 65 jC for 30 s,
72 jC for 40 s, and 72 jC for 5 min. The PCR products
were sequenced as described above. This sequence deter-
mination was repeated using cDNAs derived from whole
brain, heart and ovary to rule out any possible errors
introduced during the procedure.
2.4. Comparison of the amino acid sequences of various
GnRH-II precursors
The CLUSTAL X program (version 1.81) (Thompson et
al., 1997) was used with default settings to align the
deduced amino acid sequences of the GnRH-II precursor
of the leopard gecko and other representative species with
each other. The amino acid identities were calculated
between entire ORFs, signal peptides, amidation/proteolytic
processing signals and GnRH-associated peptides (GAPs),
respectively, using the GeneDoc software (version 2.6.002)
(Nicholas and Nicholas, 1997). The amino acid similarity
score, based on the number of similar residues or conser-
vative substitutions, was also calculated between signal
peptides by GeneDoc.
2.5. Genomic structure of the GnRH-II gene
Genomic DNA was extracted from the liver using ISO-
GEN (NIPPON GENE). The GnRH-II gene was cloned by
PCR amplification using several primer pairs (Table 1, Fig.
3A). The reaction conditions were as follows: 94 jC for 5
min, 40 cycles of 94.3 jC for 45 s, 65 jC for 30 s, 72 jC for
2 min, and 72 jC for 7 min. The PCR products were
sequenced as described above. Sequencing procedure was
repeated independently three times.
2.6. Expression analysis
To identify sites where GnRH-II decapeptide might be
synthesized and obtain clues about its potential function, the
spatial expression pattern of the GnRH-II precursor was
examined by the RT–PCR-sequencing method. One micro-
liter of each five-fold-diluted RT product was amplified with
GSP-01SE and GSP-02AS. These primers, specific to the
GnRH-II precursor, were designed to amplify the entire
ORF and span all exon–intron boundaries, so any products
from possible genomic contamination could be eliminated.
For negative controls, PCR amplifications were also con-
ducted using each RNA sample without the RT reaction.
The reaction conditions were as follows: 94 jC for 5 min,
45 cycles of 94 jC for 40 s, 67 jC for 30 s, 72 jC for 40 s,
and 72 jC for 5 min. The PCR products were analyzed by
electrophoresis on a 1.2% TAE agarose gel. Each DNA
fragment was extracted from the gel and sequenced directly
to confirm that it was derived from the GnRH-II precursor

T. Ikemoto, M.K. Park / Gene 316 (2003) 157–165160
mRNA. Some of these fragments required reamplifications.
This procedure was repeated independently at least two
times.
2.7. Molecular phylogenetic analysis
The nucleotide sequences of the entire ORF and the
deduced amino acid sequences of the GnRH-II precursors in
the leopard gecko and all reported species were aligned with
each other using CLUSTAL X with default settings, fol-
lowed by some minor revisions to correct obvious mistakes.
The alignments of both nucleotide and amino acid sequen-
ces were used to generate phylogenetic trees. Phylogenetic
trees were constructed using two methods: the neighbor-
joining method (Saitou and Nei, 1987) with the Tamura 3-
parameter model (Tamura, 1992) for nucleotide sequences
and with default settings for amino acid sequences in the
Mega software (version 2.1) (Kumar et al., 2001), and the
maximum likelihood method with default settings for nu-
cleotide and amino acid sequences in the PHYLIP software
(version 3.6a3) (Felsenstein, 2002). Octopus GnRH (Iwa-
koshi et al., 2002) was used as an outgroup. Bootstrap
values were calculated with 1000 replications to estimate the
robustness of internal branches. Full species names and
GenBank accession numbers of all the reported GnRH-II
precursor sequences are as follows: Anguilla japonica
(Japanese eel), AB026990; Astatotilapia burtoni (Burton’s
mouth-brooder), L27435; Cyprinus carpio (common carp),
AY189961; Carassius auratus (goldfish), (a) U30386, (b)
U40567; Clarias gariepinus (North African catfish),
X78047; Danio rerio (zebrafish), AF511531; Dicentrarchus
labrax (European sea bass), AF224281; Homo sapiens
(human), NM_001501; Macaca mulatta (rhesus monkey),
AF097356; Morone saxatilis (striped sea-bass), AF056313;
Oncorhynchus mykiss (rainbow trout), AF125973; Oreo-
Fig. 1. Nucleotide and deduced amino acid sequence of the cDNA encoding the Gn
5Vto 3V, beginning with the initiator codon (ATG) in the coding region. The ORF i
case letters. Amino acid residues (bottom) are numbered beginning with the first M
decapeptide sequence is depicted in the open box. Sequences for the signal peptide
(Gly-Lys-Arg) is shown by the dotted line. Arrowheads indicate the splice sites. T
sequence has been deposited in the DDBJ/EMBL/GenBank under the accession
chromis niloticus (Nile tilapia), AB101666; Oryzias latipes
(Japanese medaka), AB041330; Rana catesbeiana (bull-
frog), AF186096; Rutilus rutilus (roach), U60668; Sclero-
pages jardinii (Australian bonytongue), AB047326; Sparus
aurata (gilthead seabream), U30325; Suncus murinus
(house shrew), AF107315; Trichosurus vulpecula (silver-
gray brushtail possum), AF193516; Tupaia glis belangeri
(common tree shrew), U63327; Typhlonectes natans (Rio
Cauca caecilian), AF167558; Verasper moseri (barfin floun-
der), AB066359; Octopus vulgaris (common octopus),
AB037165.
3. Results and discussion
3.1. Molecular cloning of the GnRH-II precursor cDNA in
the leopard gecko
The full-length GnRH-II precursor cDNA was isolated
from the leopard gecko whole brain by RACE and RT–PCR
(Fig. 1). The nucleotide sequence was determined by
independent amplifications using different organs (whole
brain, heart and ovary), at least three times each. The 597-
base pair (bp) cDNA consisted of a 5V-UTR of 71 nucleotides,
an ORF of 258 nucleotides encoding a prepro-GnRH-II of 86
amino acid residues, a stop codon (TGA) and a 3V-UTR of 265
nucleotides. The initiation site for translation would be the
ATG at positions + 1 to + 3 (Fig. 1), and was generally in
agreement with the initiation sites of all the other species
analyzed (Fig. 2A). Moreover, the CACCATGG sequence at
positions � 4 to + 4 is the typical consensus sequence for
translation initiation sites in eukaryotic mRNAs (Kozak,
1987). Although there is an additional in-frame ATG at
positions + 40 to + 42, CATAATGA at positions + 36 to
+ 43 is less similar to the Kozak consensus sequence. No
RH-II precursor in the leopard gecko. Nucleotides (top) are numbered from
s indicated by capital letters, and the 5V- and 3V-UTRs are indicated by lower
et residue in the ORF. The asterisk indicates the stop codon. The GnRH-II
and GAP are underlined. The amidation/proteolytic processing (A/P) signal
he polyadenylation signal (AATAAA) is double-underlined. The nucleotide
number AB104485.
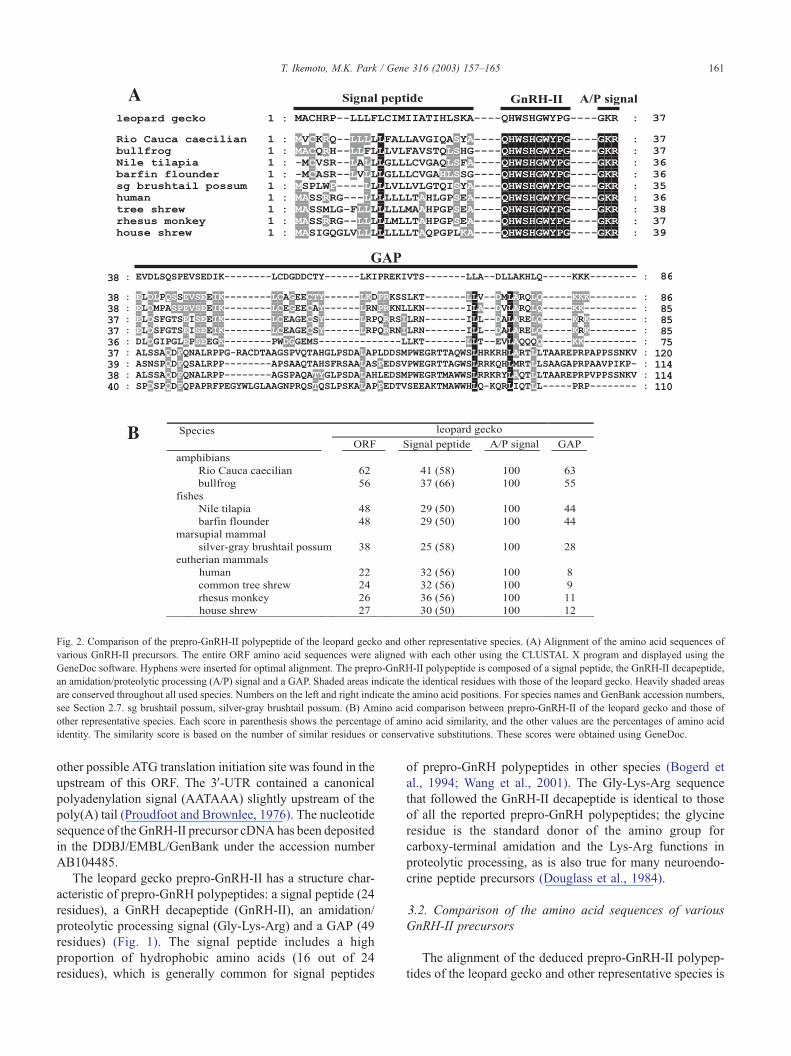
Fig. 2. Comparison of the prepro-GnRH-II polypeptide of the leopard gecko and other representative species. (A) Alignment of the amino acid sequences of
various GnRH-II precursors. The entire ORF amino acid sequences were aligned with each other using the CLUSTAL X program and displayed using the
GeneDoc software. Hyphens were inserted for optimal alignment. The prepro-GnRH-II polypeptide is composed of a signal peptide, the GnRH-II decapeptide,
an amidation/proteolytic processing (A/P) signal and a GAP. Shaded areas indicate the identical residues with those of the leopard gecko. Heavily shaded areas
are conserved throughout all used species. Numbers on the left and right indicate the amino acid positions. For species names and GenBank accession numbers,
see Section 2.7. sg brushtail possum, silver-gray brushtail possum. (B) Amino acid comparison between prepro-GnRH-II of the leopard gecko and those of
other representative species. Each score in parenthesis shows the percentage of amino acid similarity, and the other values are the percentages of amino acid
identity. The similarity score is based on the number of similar residues or conservative substitutions. These scores were obtained using GeneDoc.
T. Ikemoto, M.K. Park / Gene 316 (2003) 157–165 161
other possible ATG translation initiation site was found in the
upstream of this ORF. The 3V-UTR contained a canonical
polyadenylation signal (AATAAA) slightly upstream of the
poly(A) tail (Proudfoot and Brownlee, 1976). The nucleotide
sequence of the GnRH-II precursor cDNA has been deposited
in the DDBJ/EMBL/GenBank under the accession number
AB104485.
The leopard gecko prepro-GnRH-II has a structure char-
acteristic of prepro-GnRH polypeptides: a signal peptide (24
residues), a GnRH decapeptide (GnRH-II), an amidation/
proteolytic processing signal (Gly-Lys-Arg) and a GAP (49
residues) (Fig. 1). The signal peptide includes a high
proportion of hydrophobic amino acids (16 out of 24
residues), which is generally common for signal peptides
of prepro-GnRH polypeptides in other species (Bogerd et
al., 1994; Wang et al., 2001). The Gly-Lys-Arg sequence
that followed the GnRH-II decapeptide is identical to those
of all the reported prepro-GnRH polypeptides; the glycine
residue is the standard donor of the amino group for
carboxy-terminal amidation and the Lys-Arg functions in
proteolytic processing, as is also true for many neuroendo-
crine peptide precursors (Douglass et al., 1984).
3.2. Comparison of the amino acid sequences of various
GnRH-II precursors
The alignment of the deduced prepro-GnRH-II polypep-
tides of the leopard gecko and other representative species is

T. Ikemoto, M.K. Park / Gene 316 (2003) 157–165162
shown in Fig. 2A. For the entire ORF, the leopard gecko
prepro-GnRH-II had the highest identity (62%) with the Rio
Cauca caecilian prepro-GnRH-II (Fig. 2B). The leopard
gecko prepro-GnRH-II also showed relatively high identi-
ties (48–62%) with its counterparts in nonmammalian
species, whereas it exhibited low identities ( < 30%) with
its counterparts in mammalian species except the silver-gray
brushtail possum (38%). This tendency was also observed
for the comparison of the GAP, but the GAP sequences were
remarkably divergent, especially in length, between mam-
malian and nonmammalian species (Fig. 2A). The signal
peptide generally showed low identities ( < 40%), but
exhibited higher similarities (50–66%), which was mainly
due to the common feature of the prepro-GnRH polypeptide
of having a high proportion of hydrophobic amino acids in
the signal peptide.
3.3. Genomic structure of the GnRH-II gene
Sequencing of the PCR-amplified leopard gecko GnRH-
II gene revealed that it was composed of four exons
(designated exons 1, 2, 3 and 4) separated by three introns
(designated introns A, B and C) at similar positions as found
in other GnRH genes (Fig. 3A). The exon–intron bound-
aries were determined by sequence comparison with the
GnRH-II precursor cDNA. The three intron sites are shown
in Fig. 1. All of three 5V- and 3V-splice sites correspond to theconsensus sequences: 5V-GT (donor) and 3V-AG (acceptor)
intron splice sites (Mount, 1982) (Fig. 3B). All three introns
contains a pyrimidine-rich region for mRNA splicing im-
mediately upstream of the 3V-splice sites (Mount, 1982) (Fig.
3B). The three introns are located in the 5V-UTR, in the
Fig. 3. Schematic diagram of the GnRH-II gene in the leopard gecko. (A) Genomi
lines indicate exons and introns, respectively. Arrows indicate the locations of ol
proteolytic processing signal. (B) Sequences of exon– intron boundaries of the Gn
introns are by lower case letters. Exon– intron boundaries are depicted by slashe
contain pyrimidine-rich region (underlined) immediately upstream of the 3V-splice
middle of the GAP, and shortly upstream of the 3V-UTR,respectively. Exon 1 contains a part of the 5V-UTR. Exon 2
encodes the rest of the 5V-UTR, the signal peptide, the
GnRH-II decapeptide, the amidation/proteolytic processing
signal and the N-terminus of the GAP. Exon 3 encodes the
central portion of the GAP. Exon 4 encodes the C-terminus
of the GAP along with the 3V-UTR. It is widely recognized
that the structure of all known vertebrate GnRH genes is
conserved and is composed of four exons and three introns
(Farahmand et al., 2003). The lengths of the introns in all of
the GnRH-II genes reported to date vary between 82
(Japanese medaka intron B) and 845 bp (human intron C)
(White et al., 1998; Okubo et al., 2002). The length of intron
B (91 bp) is relatively short, while those of introns A and C
(about 2 and 4 kilobase (kb), respectively) are much longer
than those found in other species.
3.4. Expression of the GnRH-II precursor mRNA in the
leopard gecko
Single RT–PCR products of the expected size (414 bp)
were obtained from more than half of the tissues and organs
examined (Fig. 4). The authenticity of the RT–PCR prod-
ucts was confirmed by direct sequencing. The possibility of
genomic contamination was eliminated by the observation
of amplifications spanning all exon–intron boundaries. No
products were detected from any negative controls without
the RT reaction. This widespread expression of the GnRH-II
precursor mRNA is consistent with the previous findings in
several species. The presence of the GnRH-II precursor
mRNA in gonads has been reported in the goldfish (Yu et
al., 1998), Burton’s mouth-brooder (White and Fernald,
c structure of the GnRH-II gene in the leopard gecko. Boxes and horizontal
igonucleotide primers to amplify the gene partially. A/P signal, amidation/
RH-II gene in the leopard gecko. Exons are indicated by capital letters, and
s. All of three introns have the consensus 5V-GT and 3V-AG sequences and
sites.

Fig. 4. Expression of the GnRH-II precursor mRNA in the leopard gecko. RT products were PCR-amplified to span all exon– intron boundaries and were
analyzed by electrophoresis on a 1.2% TAE agarose gel with ethidium bromide staining. The authenticity of the RT–PCR products was confirmed by direct
sequencing of each band of the expected size (414 bp; indicated by an arrow). The lane of RT-minus represents the negative control using male whole brain
RNA sample without the RT reaction. M, molecular marker; � , under detection level.
T. Ikemoto, M.K. Park / Gene 316 (2003) 157–165 163
1998), Japanese eel (Okubo et al., 1999), rainbow trout (von
Schalburg et al., 1999), North African catfish (Bogerd et al.,
2002) and humans (Millar, 2003). In the North African
catfish, ubiquitous expression of the GnRH-II precursor
mRNA was observed in all tissues and organs tested
(Bogerd et al., 2002). The hemipenis is the male copulatory
organ, and is one of the major features distinguishing the
Fig. 5. Unrooted neighbor-joining phylogenetic tree of the GnRH-II precursors. The
parameter model for entire ORF nucleotide sequences using the Mega software. Oc
are indicated for all nodes on the tree. The scale bar beneath the tree corresponds
accession numbers, see Section 2.7.
both sexes of the leopard gecko and some other reptiles.
This is the first report to the best of our knowledge on the
expression of the GnRH precursor mRNA in the male
copulatory organ, the hemipenis, and its counterpart in
female, the hemiclitoris.
The roles of GnRH outside the hypothalamus–pituitary–
gonadal axis remain largely unknown. Although the expres-
tree was constructed using the neighbor-joining method with the Tamura 3-
topus GnRH was used as an outgroup. Bootstrap values of 1000 resamplings
to estimated evolutionary distance units. For species names and GenBank

T. Ikemoto, M.K. Park / Gene 316 (2003) 157–165164
sion of the GnRH-II precursor mRNA does not ensure its
translation into a functional GnRH-II decapeptide, the
present results support the notion that GnRH-II may act as
a neurotransmitter and/or a neuromodulator in the brain and
an autocrine and/or a paracrine hormone outside the brain.
Verifying this, however, will require further studies. Under-
standing the roles of the widespread GnRH-II expression
will require detailed knowledge of its specific sites of action
and the distribution of its receptor.
3.5. Molecular phylogenetic analysis
Molecular phylogenetic trees of the GnRH-II precursors
were constructed using the methods of neighbor-joining and
maximum likelihood for nucleotide and amino acid sequen-
ces. Each method yielded essentially the same tree topology.
Fig. 5 shows an unrooted neighbor-joining tree based on the
entire ORF nucleotide sequences. Octopus GnRH used as an
outgroup indicates the putative position of the root. The
robustness of internal branches was estimated by 1000
bootstrap resamplings. As inferred from the results of the
sequence comparison, there were two major clusters: mam-
malian species (except for a marsupial mammal, the silver-
gray brushtail possum), and the other species. The leopard
gecko has a similar precursor sequence with those of fishes
and amphibians, while eutherian mammals have distinct
sequences, especially in the GAP region. Besides, some
mammalian species are thought not to have a GnRH-II
decapeptide (Millar, 2003). Thus the precursor sequence
of the evolutionarily conserved GnRH-II may have become
somewhat divergent during amniote evolution.
3.6. Conclusions
We have identified and characterized the full-length
cDNA encoding the GnRH-II precursor and determined its
genomic structure in the leopard gecko, E. macularius. This
is the first report of such an analysis in reptiles. The deduced
prepro-GnRH-II polypeptide had the highest identities with
the corresponding polypeptides of amphibians.
Expression of the GnRH-II precursor was detected in
more than half of the tissues and organs examined by the
RT–PCR-sequencing method. This widespread expression
is consistent with the previous findings in several species,
though the role of this decapeptide outside the hypothala-
mus–pituitary–gonadal axis remains largely unknown.
Molecular phylogenetic analysis combined with se-
quence comparison showed that the leopard gecko is more
similar to fishes and amphibians than to eutherian mammals
with respect to the GnRH-II precursor sequence. These
results strongly suggest that the divergence of the GnRH-
II precursor sequences seen in eutherian mammals may have
occurred along with amniote evolution. Further character-
ization of the precursor sequence of GnRH, including
GnRH-II, will provide strong insights into the phylogeny
and evolution of GnRH.
Acknowledgements
The authors would like to thank the editor and reviewers
for their helpful comments.
References
Adams, B.A., Tello, J.A., Erchegyi, J., Warby, C., Hong, D.J., Akinsanya,
K.O., Mackie, G.O., Vale, W., Rivier, J.E., Sherwood, N.M., 2003. Six
novel gonadotropin-releasing hormones are encoded as triplets on each
of two genes in the protochordate, Ciona intestinalis. Endocrinology
144, 1907–1919.
Bogerd, J., Zandbergen, T., Andersson, E., Goos, H., 1994. Isolation, char-
acterization and expression of cDNAs encoding the catfish-type and
chicken-II-type gonadotropin-releasing-hormone precursors in the Afri-
can catfish. Eur. J. Biochem. 222, 541–549.
Bogerd, J., Diepenbroek, W.B., Hund, E., van Oosterhout, F., Teves, A.C.,
Leurs, R., Blomenrohr, M., 2002. Two gonadotropin-releasing hormone
receptors in the African catfish: no differences in ligand selectivity, but
differences in tissue distribution. Endocrinology 143, 4673–4682.
Burgus, R., Butcher, M., Amoss, M., Ling, N., Monahan, M., Rivier, J.,
Fellows, R., Blackwell, R., Vale, W., Guillemin, R., 1972. Primary
structure of the ovine hypothalamic luteinizing hormone-releasing fac-
tor (LRF). Proc. Natl. Acad. Sci. U. S. A. 69, 278–282.
Craig, A.G., Fischer, W.H., Park, M., Rivier, J.E., Musselman, B.D., Powell,
J.F., Reska-Skinner, S.M., Prakash, M.O., Mackie, G.O., Sherwood,
N.M., 1997. Sequence of two gonadotropin releasing hormones from
tunicate suggest an important role of conformation in receptor activation.
FEBS Lett. 413, 215–225.
Crews, D., Sakata, J., Rhen, T., 1998. Developmental effects on intersexual
and intrasexual variation in growth and reproduction in a lizard with
temperature-dependent sex determination. Comp. Biochem. Physiol. C,
Pharmacol. Toxicol. Endocrinol. 119, 229–241.
Di Fiore, M.M., Rastogi, R.K., Ceciliani, F., Messi, E., Botte,
V., Botte, L., Pinelli, C., D’Aniello, B., D’Aniello, A., 2000. Mammalian
and chicken I forms of gonadotropin-releasing hormone in the gonads of
a protochordate, Ciona intestinalis. Proc. Natl. Acad. Sci. U. S. A. 97,
2343–2348.
Dores, R.M., Rubin, D.A., Quinn, T.W., 1996. Is it possible to construct
phylogenetic trees using polypeptide hormone sequences? Gen. Comp.
Endocrinol. 103, 1–12.
Douglass, J., Civelli, O., Herbert, E., 1984. Polyprotein gene expression:
generation of diversity of neuroendocrine peptides. Annu. Rev. Bio-
chem. 53, 665–715.
Farahmand, H., Rahman, M.A., Sohm, F., Hwang, G.L., Maclean, N.,
2003. Isolation and expression of tilapia (Oreochromis niloticus) serine
8-type GnRH coding and regulatory sequences. Gene 304, 97–106.
Felsenstein, J., 2002. PHYLIP (Phylogeny Inference Package) version
3.6a3. Distributed by the author. Department of Genome Sciences,
University of Washington, Seattle.
Fernald, R.D., White, R.B., 1999. Gonadotropin-releasing hormone
genes: phylogeny, structure, and functions. Front. Neuroendocrinol.
20, 224–240.
Iwakoshi, E., Takuwa-Kuroda, K., Fujisawa, Y., Hisada, M., Ukena, K.,
Tsutsui, K., Minakata, H., 2002. Isolation and characterization of a
GnRH-like peptide from Octopus vulgaris. Biochem. Biophys. Res.
Commun. 291, 1187–1193.
Kozak, M., 1987. An analysis of 5V-noncoding sequences from 699 verte-
brate messenger RNAs. Nucleic Acids Res. 15, 8125–8148.
Kumar, S., Tamura, K., Jakobsen, I.B., Nei, M., 2001. MEGA2: molecular
evolutionary genetics analysis software. Bioinformatics 17, 1244–1245.
Lovejoy, D.A., Fischer, W.H., Parker, D.B., McRory, J.E., Park, M.,
Lance, V., Swanson, P., Rivier, J.E., Sherwood, N.M., 1991. Primary
structure of two forms of gonadotropin-releasing hormone from brains

T. Ikemoto, M.K. Park / Gene 316 (2003) 157–165 165
of the American alligator (Alligator mississippiensis). Regul. Pept. 33,
105–116.
Matsuo, H., Baba, Y., Nair, R.M., Arimura, A., Schally, A.V., 1971. Structure
of the porcine LH- and FSH-releasing hormone: I. The proposed amino
acid sequence. Biochem. Biophys. Res. Commun. 43, 1334–1339.
Millar, R.P., 2003. GnRH II and type II GnRH receptors. Trends Endocri-
nol. Metab. 14, 35–43.
Millar, R.P., King, J.A., 1987. Structural and functional evolution of gona-
dotropin-releasing hormone. Int. Rev. Cytol. 106, 149–182.
Miyamoto, K., Hasegawa, Y., Nomura, M., Igarashi, M., Kangawa, K.,
Matsuo, H., 1984. Identification of the second gonadotropin-releasing
hormone in chicken hypothalamus: evidence that gonadotropin secre-
tion is probably controlled by two distinct gonadotropin-releasing hor-
mones in avian species. Proc. Natl. Acad. Sci. U. S. A. 81, 3874–3878.
Mount, S.M., 1982. A catalogue of splice junction sequences. Nucleic
Acids Res. 10, 459–472.
Nicholas, K.B., Nicholas Jr., H.B., 1997. GeneDoc: ATool for Editing and
Annotating Multiple Sequence Alignments. Distributed by the author.
Okubo, K., Suetake, H., Aida, K., 1999. Expression of two gonadotropin-
releasing hormone (GnRH) precursor genes in various tissues of the
Japanese eel and evolution of GnRH. Zool. Sci. 16, 471–478.
Okubo, K., Mitani, H., Naruse, K., Kondo, M., Shima, A., Tanaka, M.,
Asakawa, S., Shimizu, N., Yoshiura, Y., Aida, K., 2002. Structural
characterization of GnRH loci in the medaka genome. Gene 293,
181–189.
Powell, R.C., Ciarcia, G., Lance, V., Millar, R.P., King, J.A., 1986. Iden-
tification of diverse molecular forms of GnRH in reptile brain. Peptides
7, 1101–1108.
Proudfoot, N.J., Brownlee, G.G., 1976. 3VNon-coding region sequences in
eukaryotic messenger RNA. Nature 263, 211–214.
Rastogi, R.K., Di Fiore, M.M., D’Aniello, A., Iela, L., Fiorentino, M.,
2002. GnRH in the invertebrates: an overview. Prog. Brain Res. 141,
19–29.
Saitou, N., Nei, M., 1987. The neighbor-joining method: a new method for
reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425.
Sealfon, S.C., Weinstein, H., Millar, R.P., 1997. Molecular mechanisms of
ligand interaction with the gonadotropin-releasing hormone receptor.
Endocr. Rev. 18, 180–205.
Seeburg, P.H., Adelman, J.P., 1984. Characterization of cDNA for precur-
sor of human luteinizing hormone-releasing hormone. Nature 311,
666–668.
Sherwood, N.M., Lovejoy, D.A., Coe, I.R., 1993. Origin of mammalian
gonadotropin-releasing hormones. Endocr. Rev. 14, 241–254.
Tamura, K., 1992. Estimation of the number of nucleotide substitutions
when there are strong transition-transversion and G+C-content biases.
Mol. Biol. Evol. 9, 678–687.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins, D.G.,
1997. The CLUSTAL_X windows interface: flexible strategies for mul-
tiple sequence alignment aided by quality analysis tools. Nucleic Acids
Res. 25, 4876–4882.
Tsutsui, H., Yamamoto, N., Ito, H., Oka, Y., 1998. GnRH-immunoreactive
neuronal system in the presumptive ancestral chordate, Ciona intesti-
nalis (Ascidian). Gen. Comp. Endocrinol. 112, 426–432.
von Schalburg, K.R., Harrower, W.L., Sherwood, N.M., 1999. Regulation
and expression of gonadotropin-releasing hormone in salmon embryo
and gonad. Mol. Cell. Endocrinol. 157, 41–54.
Wang, L., Yoo, M.S., Kang, H.M., Im, W.B., Choi, H.S., Bogerd, J., Kwon,
H.B., 2001. Cloning and characterization of cDNAs encoding the
GnRH1 and GnRH2 precursors from bullfrog (Rana catesbeiana).
J. Exp. Zool. 289, 190–201.
White, R.B., Fernald, R.D., 1998. Genomic structure and expression sites
of three gonadotropin-releasing hormone genes in one species. Gen.
Comp. Endocrinol. 112, 17–25.
White, R.B., Eisen, J.A., Kasten, T.L., Fernald, R.D., 1998. Second gene
for gonadotropin-releasing hormone in humans. Proc. Natl. Acad. Sci.
U. S. A. 95, 305–309.
Yu, K.L., He, M.L., Chik, C.C., Lin, X.W., Chang, J.P., Peter, R.E., 1998.
mRNA expression of gonadotropin-releasing hormones (GnRHs) and
GnRH receptor in goldfish. Gen. Comp. Endocrinol. 112, 303–311.














