Embed Size (px)
Citation preview

185
American Journal of Botany 88(2): 185–195. 2001.
FLORAL DEVELOPMENT IN APHANDRA (ARECACEAE)1
ANDERS S. BARFOD AND NATALIE W. UHL2
Department of Systematic Botany, Aarhus University, Nordlandsvej 68, DK-8240 Risskov, Denmark; and L. H. Bailey Hortorium,467 Mann Library, Cornell University, Ithaca, New York 14853 USA
The organogenesis of staminate flower clusters and flowers and some observations on the corresponding pistillate structures ofAphandra natalia are described and compared with those of the other two genera in the Phytelephantoideae (Arecaceae). In Aphandra,staminate flowers are borne in monopodial clusters of mostly four (1–6) flowers. Each flower is surrounded by two pairs of suboppositebracteoles and has two rather indistinctly four-parted whorls of perianth parts. Stamen primordia arise on a shallow apical dome andthen centrifugally down the sides of a long, angled, and laterally flattened receptacle. Immediately before the staminate bud opens,the floral receptacle below the androecium rapidly elongates, becoming funnel-shaped, with the bracteoles and a perianth sheath adnateto it forming a pseudopedicel. Epidermal and subepidermal layers of these pseudopedicels split at anthesis and release a great numberof raphide idioblasts that resemble the pollen grains in shape and size. It is hypothesized that the idioblasts deter pollen feeding orovidepositing insects. The phylogenetic implications of these findings are important within the Phytelephantoideae and among palmsin general.
Key words: Arecaceae; Aphandra natalia; floral development; monotocotyledons; palms; Phytelephantoideae; pollination; pseu-dopedicel.
Aphandra natalia (Balslev and Henderson) Barfod is a pin-nate leaved, single-stemmed palm found in Amazonian Ec-uador and Peru near the foothills of the Andes. The genusbelongs to the subfamily Phytelephantoideae, which includesonly three small genera and constitutes a morphologically iso-lated group of dioecious genera within the Palmae (Uhl andDransfield, 1987). Developmental studies have shown that thephytelephantoid genera have the only monopodial flower clus-ters in the family, a four-parted perianth otherwise known onlyin only one species of Chelyocarpus (Coryphoideae), and cen-trifugal stamen inception. Partial centrifugal stamen develop-ment is known to occur elsewhere only in the genus Eugeis-sona (Calamoideae) (Uhl and Moore, 1977; Uhl and Drans-field, 1984).
Balslev and Henderson (1987) originally referred Aphandranatalia to Ammandra based on the prominent submarginalveins on the pinnae and the pedicellate condition of the sta-minate flower clusters. Monographic work on the subfamilyPhytelephantoideae has shown that it is a distinct genus (Bar-fod, Henderson, and Balslev, 1987; Barfod, 1991) (Table 1)and that the structure of the floral pedicel is critical. Devel-opmental studies of the inflorescence and flowers of Aphandraare important for eludication of the pedicel and for comparisonwith developmental patterns previously described for the othergenera of Phytelephantoideae (Uhl and Moore, 1977; Uhl andDransfield, 1984). In this study we address three issues in par-ticular: ontogeny of the staminate flower cluster, stamen in-ception, and the structure of the pedicel of the staminate flow-er. Some observations are also presented on the developmentof the pistillate flower clusters and flowers.
MATERIALS AND METHODS
Plant material—Aphandra is exploited for fibers extracted from the leafbases (Borgtoft Pedersen and Balslev, 1990); developmental material is dif-
1 Manuscript received 21 December, 1999; revision accepted 27 April 2000.The authors thank Anni Sloth for assistance with laboratory work, SEM,
and electronic processing of the photographs; and the reviewers. Support isacknowledged from NSF grant BSR-8806042 to NWU.
2 Author for correspondence (e-mail: [email protected]).
ficult to obtain since large valuable trees must be felled to get a series. Inflo-rescence stages were obtained from three individuals growing in semi-culti-vated populations near Logrono, Province of Morona-Santiago in Ecuador, thetype locality of Aphandra natalia. The young inflorescences (Figs. 1–4) (Ta-ble 2) were fixed in FAA (formalin-acetic acid-alcohol) in the field and after2 d, transferred to storage in nine parts 70% glycerine alcohol to one part10% glycerine. Prophylls and peduncular bracts (PD bracts) were cut open tofacilitate infiltration.
Scanning electron microscopy—The various pistillate and staminate de-velopmental stages were critical point dried in a Biorad CPD-750 Apparatus,mounted on stubs with double-adhesive tape, and coated with platinum in anEdwards S150B Sputter Coater. They were studied with a JEOL 840 ScanningMicroscope at 20 kV.
Clearings—Material for study of vasculature was cleared using a 5% so-lution of NaOH and left overnight in an oven at 508C. After 24 h the NaOHwas changed and we repeated the procedure until clearing was satisfactory.The material was then rinsed in distilled water and treated with commercialbleach to remove any cloudiness. Subsequently preparations were stored inglycerine alcohol. The material was stained in a basic fuchsin solution (1 gof fuchsin and 6 g of solid KOH in 100 mL water) for photographing.
RESULTS
Inflorescence structure—In wild populations of Aphandranatalia the distribution of staminate and pistillate individualsis almost even. Flowering occurs throughout the year, peakingslightly in February–March (Barfod, 1991). Inflorescencesform in acropetal sequence in the axils of their subtendingleaves. All leaves produced during periods of flowering sub-tend a young inflorescence bud. The pistillate and staminatebuds have a similar overall appearance while they are stillincluded in the sheaths of the subtending leaves, but they de-velop differently after the peduncular bract (PD bract) breaksthrough the prophyll (Figs. 1–7). In the staminate bud, the PDbract continues to grow and before it splits will attain morethan twice the size of the prophyll. At anthesis the entire in-florescence is exposed and pendant. The pistillate bud, in con-trast, elongates only a little after the splitting of the prophyll,the PD bract reaching only about three-quarters the length of

186 [Vol. 88AMERICAN JOURNAL OF BOTANY
TABLE 1. Diagnostic table.
Ammandra decasperma Aphrandra natalia Phytelephas macrocarpa Phytelephas aequatorialis
Pistillate inflorescenceNo. of flowers per inflorescence 6–10 30–50 5–25 15–25No. of sepaloid bracts 0? 4–6 4–6 4–6No. of perianth segments 7–10 7–9 5–8 6–9No. of carpels 6–9 6–8 4–5 5–6
Staminate inflorescenceLength of rachis 30–60 100–170 30–60 90–150Axis of flower cluster at anthes-
isapresent indistinguishable indistinguishable indistinguishable
No. of flowers per clusterb 6–9 4 (25) 4 (25) 4 (25)Pedicel pseudopedicel?c pseudopedicelc indistinguishable pedicelDistribution of idioblasts concentrated in
blisterseven even even
Idioblast released at anthesis yes yes no noNo. of sepaloid bracts 0? 4 4 4No. of perianth segments 8? 8 8 8Shape of male receptacle at an-
thesiselongated, recurved elongated, dome
shapedflat to dome shaped widened, flat
Stamen inception centrifugal? centrifugal, sequential centrifugal, sequential centrifugal, in 2phases
No. of stamens 300–500 400–650 150–300 500–700Length of filament at anthesis 0.1–0.2 2.5–3 3–9 6–10Length of anthers at anthesis 0.5 3–3.5 4–6.5 3–5Pistillode 1/2 present absent absent absentSize and shape of pollen grain 45–55, rounded 60–70, elliptic 55–65, elliptic to
elongate80, elongate
Pollen grain aperture monoporate monosulcate monosulcate monosulcate
a The individual pseudopedicels or pedicels of the flowers of Aphandra natalia and Phytelephas aequatorialis are fused, thus forming a pseudo-axis.
b Aberrant number in parentheses.c Pseudopedicel formed by fusion and elongation of the perianth and the receptacle (see text).
the prophyll. The rachis of the pistillate inflorescence is muchshorter, and the flowers remain closely bunched. At anthesis,they are partly contained in the PD bract, which splits longi-tudinally along the abaxial side to expose only the tips of longstrap-shaped perianth parts and stigma branches (Fig. 19C).
Structure of the staminate flower cluster—The staminateinflorescence of Aphandra natalia breaks through the PD bractin the evening and finishes development overnight. Elongationproceeds at up to 10 cm/h until the rachis has attained a lengthof 100–170 cm at anthesis the following day. The staminateflowers are gathered in 200–300 spirally arranged and denselyinserted clusters (Figs. 6, 19A). The individual clusters havemostly four (1–6) flowers. Lower, stalklike parts of the flowersare united to varying degrees and taper proximally to a com-mon attachment (Figs. 8, 19A). The basal clusters have fewerflowers, usually two or three, and a number of solitary flowersare always present at the apex of the inflorescence. Each clus-ter is subtended by a bract that differs in shape and size fromdeltoid and is ;3 cm long at the base of the rachis to a narrow,few millimetre wide rim near the apex.
The flower cluster usually consists of two pairs of subop-posite flowers (Fig. 8). Rarely, one or two more flowers de-velop distal to these. The lower pair of flowers are in lateralpositions and the upper two flowers are smaller and in anteriorand posterior postions. The receptacle of the anterior flower isnarrower than those of the lateral ones (Fig. 8), and the pos-terior flower never attains the size of the others. All flowersare sessile (Figs. 6, 7) until late in ontogeny when a curvedstalk develops, and the cluster bends over and ultimately be-
comes adpressed to the rachis with the posterior flower partlyhidden (Fig. 19A).
Organogenesis of staminate flower clusters and flowers—In the earliest developmental stages available to us, the flowercluster sites are covered by their subtending bracts. Removalof the bract exposes a lentil-shaped apical dome in each flowercluster (Fig. 16). The two bracts that subtend the lateral flow-ers (Fig. 16: A, B) are clearly discernible (ba and bb), whereasthe subtending bracts of the posterior and anterior flowers,respectively, are not visible in distal view. The apices of thefour flowers are flattened to slightly dome-shaped (Fig. 17: A,B, and D). In distal outline, they appear three- to four-lobedbecause of primordial bulges (Fig. 18, 19D). On the lateralflowers, a pair of subopposite bracteoles develops first, slightlydisplaced relative to the plane of symmetry (Figs. 16, 19: 1b,2b, 1a, 2a). The posterior member of this pair is usually visiblebefore the anterior one. On the posterior flower of the cluster,the first-formed pair of subopposite bracteoles can be distin-guished in lateral positions (Fig. 16: 1c, 2c). The anterior floralprimordium is partly hidden under the basal part of the bractthat subtends the entire flower cluster.
Each floral apex produces an additional pair of suboppositebracteoles, perpendicular to the first formed ones (Fig. 19D).In Fig. 15 (3a, 3b), a bracteole is formed at a latero-posteriorposition, and the opposite one in a latero-anterior position (4a,4b). The former appears first and attains a larger size than anyof the remaining bracts. Later in ontogeny, the two pairs ofsubopposite bracteoles are adnate to the receptacle as it be-comes elongate. Their presence in the mature flower is ob-
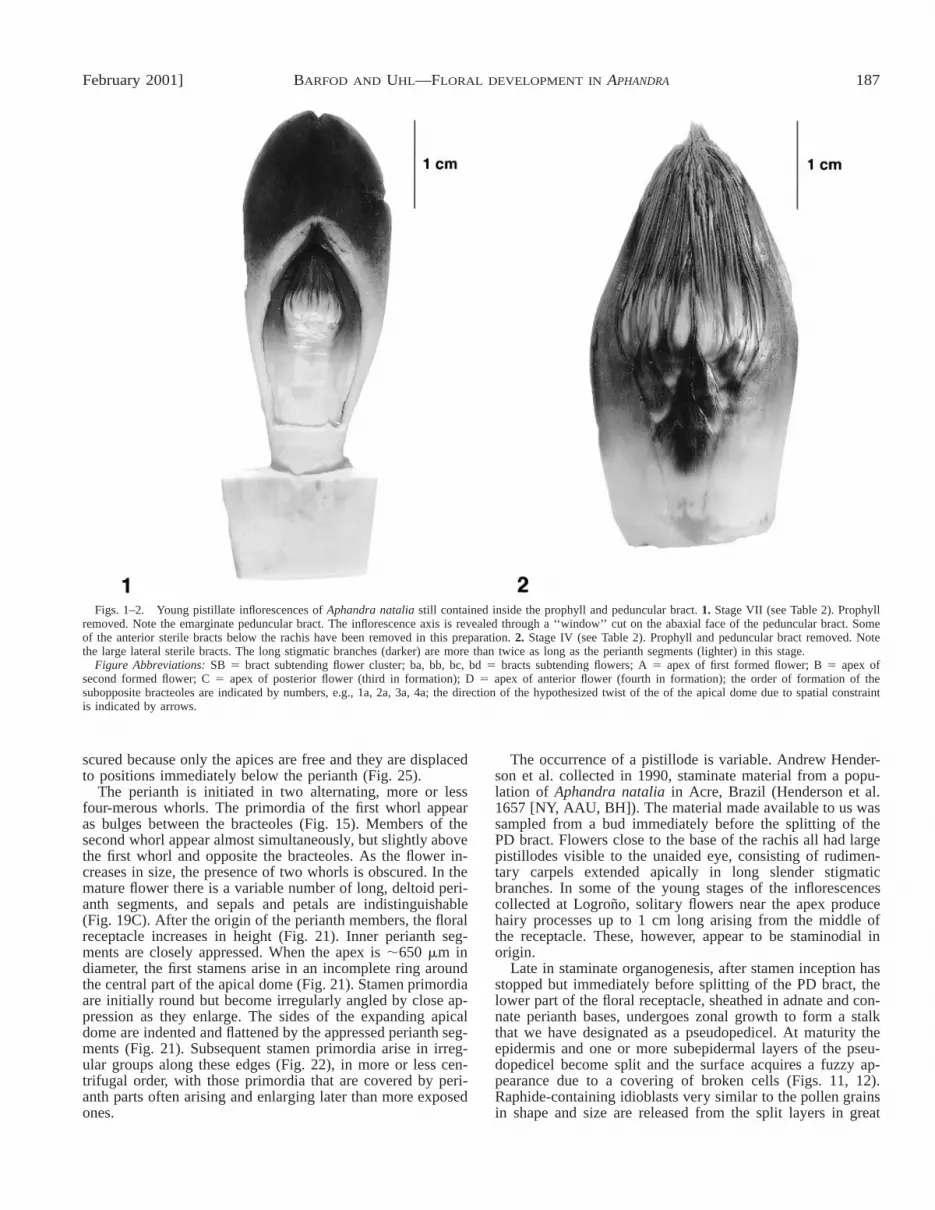
February 2001] 187BARFOD AND UHL—FLORAL DEVELOPMENT IN APHANDRA
Figs. 1–2. Young pistillate inflorescences of Aphandra natalia still contained inside the prophyll and peduncular bract. 1. Stage VII (see Table 2). Prophyllremoved. Note the emarginate peduncular bract. The inflorescence axis is revealed through a ‘‘window’’ cut on the abaxial face of the peduncular bract. Someof the anterior sterile bracts below the rachis have been removed in this preparation. 2. Stage IV (see Table 2). Prophyll and peduncular bract removed. Notethe large lateral sterile bracts. The long stigmatic branches (darker) are more than twice as long as the perianth segments (lighter) in this stage.
Figure Abbreviations: SB 5 bract subtending flower cluster; ba, bb, bc, bd 5 bracts subtending flowers; A 5 apex of first formed flower; B 5 apex ofsecond formed flower; C 5 apex of posterior flower (third in formation); D 5 apex of anterior flower (fourth in formation); the order of formation of thesubopposite bracteoles are indicated by numbers, e.g., 1a, 2a, 3a, 4a; the direction of the hypothesized twist of the of the apical dome due to spatial constraintis indicated by arrows.
scured because only the apices are free and they are displacedto positions immediately below the perianth (Fig. 25).
The perianth is initiated in two alternating, more or lessfour-merous whorls. The primordia of the first whorl appearas bulges between the bracteoles (Fig. 15). Members of thesecond whorl appear almost simultaneously, but slightly abovethe first whorl and opposite the bracteoles. As the flower in-creases in size, the presence of two whorls is obscured. In themature flower there is a variable number of long, deltoid peri-anth segments, and sepals and petals are indistinguishable(Fig. 19C). After the origin of the perianth members, the floralreceptacle increases in height (Fig. 21). Inner perianth seg-ments are closely appressed. When the apex is ;650 mm indiameter, the first stamens arise in an incomplete ring aroundthe central part of the apical dome (Fig. 21). Stamen primordiaare initially round but become irregularly angled by close ap-pression as they enlarge. The sides of the expanding apicaldome are indented and flattened by the appressed perianth seg-ments (Fig. 21). Subsequent stamen primordia arise in irreg-ular groups along these edges (Fig. 22), in more or less cen-trifugal order, with those primordia that are covered by peri-anth parts often arising and enlarging later than more exposedones.
The occurrence of a pistillode is variable. Andrew Hender-son et al. collected in 1990, staminate material from a popu-lation of Aphandra natalia in Acre, Brazil (Henderson et al.1657 [NY, AAU, BH]). The material made available to us wassampled from a bud immediately before the splitting of thePD bract. Flowers close to the base of the rachis all had largepistillodes visible to the unaided eye, consisting of rudimen-tary carpels extended apically in long slender stigmaticbranches. In some of the young stages of the inflorescencescollected at Logrono, solitary flowers near the apex producehairy processes up to 1 cm long arising from the middle ofthe receptacle. These, however, appear to be staminodial inorigin.
Late in staminate organogenesis, after stamen inception hasstopped but immediately before splitting of the PD bract, thelower part of the floral receptacle, sheathed in adnate and con-nate perianth bases, undergoes zonal growth to form a stalkthat we have designated as a pseudopedicel. At maturity theepidermis and one or more subepidermal layers of the pseu-dopedicel become split and the surface acquires a fuzzy ap-pearance due to a covering of broken cells (Figs. 11, 12).Raphide-containing idioblasts very similar to the pollen grainsin shape and size are released from the split layers in great

188 [Vol. 88AMERICAN JOURNAL OF BOTANY
TABLE 2. Measurements of developmental stages of the pistillate andstaminate inflorescences of Aphandra natalia.
Length of pro-phyll (cm)
Length of PDa
bract (cm)Distance between
(mm)Length of
rachis (mm)
Pistillate seriesStage IXStage VIIIStage VIIStage VIStage VStage IVStage IIIStage II
5.512252931353539
33455.56.38.5
10.5
58
101519202025
2556779
10Staminate series IStage VIIIStage VIIStage VIStage VStage IVStage IIIStage IIStage I
36.5
2332363636cut
1.84.5
148
10121524
1018355550505280
10203040456080
155Staminate series II
4042
6162
92105
360370
a PD 5 peduncular bract.
Figs. 3–4. Developmental stages of staminate and pistillate inflorescences of Aphandra natalia enclosed by the prophyll and the peduncular bract. TheRoman numerals indicate the consecutive order in which the buds were removed from the crown. They are referred to in Table 2 and throughout the text. 3.Staminate series (from left to right, stages I–VII). Besides the stages shown here two additional stages were collected for study: one immediately before splittingof the PD bract, and one immediately before anthesis. 4. Pistillate series (from left to right, stages I–IX).
numbers. They remain trapped in the epidermal shreds on thesurface where they mix with pollen grains at anthesis (Figs.12, 13).
When the inflorescence breaks through the peduncular bract,400–650 stamens have been produced. A narrow zone of thefloral receptacle above the perianth and below the stamen-bearing part remains naked (Figs. 9, 12).
Vascular anatomy—The large number of collateral bundlessupplying each flower varies with the size and developmentalstage. As the floral apex expands longitudinally and in diam-eter the bundles curve towards the periphery and anastomosefrequently (Fig. 10).
Pistillate inflorescence and flowers: structure anddevelopment—Stages of pistillate organs were incomplete butsome observations were possible. The PD bract of the pistillate
inflorescence bud differs from its staminate homologue byhaving an emarginate apex (Figs. 1, 5). The rachis of the in-florescence resembles the staminate but bears only ;20 soli-tary, spirally arranged, pistillate flowers, each in the axil of abract. It is noteworthy that two subopposite pairs of bracteolesare present below each flower. In fully developed flowers, thebracteoles can be distinguished from the perianth segments bybeing smaller and less fleshy. In Fig. 1, the youngest stageavailable, the subtending bract, four bracteoles, and the peri-anth of the pistillate flowers are already elongate. The centralwhitish bulges in individual flowers are the multicarpellate gy-noecia. The flowers in stage number VII of our collection havefree carpel primordia that are conduplicate. The carpel pri-mordia are present before the initiation of the staminodes,which first appear as a single series of primordia surroundingthe gynoecium, a second whorl eventually forming on the out-side.
The development of the pistillate perianth is irregular. Ourcollections did not comprise the earliest stages of perianth in-ception, but the sizes and insertion of the segments suggestthat more than one whorl is involved. Whereas the first-formedsegments are uniform in shape and in size, additional segmentsusually form on the abaxial side of the flower. This phenom-enon, possibly related to asymmetrical expansion of the apex,was also observed in Phytelephas macrocarpa R. and P. (Bar-fod, 1991).
Early in organogenesis, the apices of the carpels elongate.The resulting stigmatic branches become longer than the peri-anth segments as can be determined from the material shownin Figs. 1 and 2. The stylar portion of the gynoecium elongatescontinuously until anthesis when the stigmatic branches aresurrounded by the apices of the long perianth segments. Atthis time the style length ranges between 20 and 25 cm andthe stigmatic branches are 4–5 cm long (Fig. 19C).
DISCUSSION
This study confirms that in staminate flowers of Aphandra,two pairs of sepaloid bracteoles develop before the perianthon each floral apex (Barfod, 1991), that the floral apex expandsin width and height during stamen development, and that sta-men development is for the most part centrifugal (Uhl andMoore, 1977). The stalk of the staminate flower was deter-

February 2001] 189BARFOD AND UHL—FLORAL DEVELOPMENT IN APHANDRA
Figs. 5–9. Developmental stages of the staminate inflorescence of Aphandra natalia. 5. Stage VI. The prophyll and peduncular bract have both been splitopen along their flattened rims. Note that the prophyll is much larger than the peduncular bract. The latter is pointed and not notched like its pistillate homologuein Fig. 1. 6. Stage III. Note the sessile flower clusters. At this stage the pseudopedicels have not formed. The two large lateral sterile bracts at the base arehomologous to similar bracts found in the pistillate inflorescence (Fig. 2). 7. Stage VI. Rachis showing the two sterile lateral bracts at the base and the blacktips of the bracts subtending the flower clusters. 8. Monopodial cluster of four flowers. The stamens have been removed to reveal the uneven size and shapeof the receptacles. 9. Adaxial side of a fully developed lateral flower of a cluster. Note the irregular borderline between the perianth fused to the receptacle andthe naked peripheral zone of the apical dome. Raphide idioblasts are visible on the pseudopedicel stalk as tiny luminescent spots.

190 [Vol. 88AMERICAN JOURNAL OF BOTANY
Figs. 10–13. Preparations of the staminate flowers of Aphandra natalia at anthesis. 10. Section of cleared flower stained to show the vascular bundles. 11.SEM preparation showing the borderline between the naked, glabrous peripheral zone of the apical dome (upper half of picture) and split superficial layers ofthe pseudopedicel covered by masses of pollen grains and raphide idioblasts. 12. Detail of Fig. 11 showing the surface of pseudopedicel. Note the mixture ofpollen grains and raphide idioblasts almost indistinguishable in shape and size. 13. LM preparation of the pollen grains and raphide idioblasts scraped from thesurface of the psudopedicel.

February 2001] 191BARFOD AND UHL—FLORAL DEVELOPMENT IN APHANDRA
Figs. 14–18. Staminate inflorescence of Aphandra natalia. Development of the flower cluster. 14. Stage VII. Part of an inflorescence rachis with flowerclusters in different stages. Numbers indicate the ontogenetic sequence from 1 (youngest cluster) to 12 (oldest cluster). 15. Stage VII. Cluster number 10 inFig. 14. The perianth primordia (asterisks) are visible. 16. Stage VIII. 17. Stage VII. Cluster number 1 in Fig. 14. Latero-distal view. 18. Stage VII. Detailshowing cluster number 1 in Fig 14. Distal view.

192 [Vol. 88AMERICAN JOURNAL OF BOTANY
Fig. 19. (A) Part of a mature staminate inflorescence showing four-merousflower clusters; each flower with numerous stamens. (B) A young staminateflower dissected from an unopened bud; note the sterile bracts and perianthsegments surrounding the closely appressed stamen primordia, and the largeproximal bract which subtends the flower cluster. (C) A pistillate flower atanthesis showing strap-shaped sepaloid bracteoles at the base, the staminodes,long perianth segments, and sinuous stigmatic branches. (D) Diagram of sta-minate flower cluster of Aphandra natalia to show the approximate configu-ration of bracts and bracteoles. Note the slightly displaced position of thebracteoles of the lateral flowers, probably due to spatial constraint in thedeveloping floral apices.
mined to be structurally unique and designated as a pseudo-pedicel, which consists of an enlarged, more or less funnel-shaped floral receptacle with an adnate perianth tube. Thesmall tips of the perianth members are free distally. The epi-dermis and one or more underlying layers of the pseudopedicelcontain large raphide idioblasts, resembling the pollen grainsin shape. The raphides are freed by disintegration of the epi-dermal cells and become mixed with pollen grains on the sur-face of the pseudopedicel at anthesis. Comparison with othergenera of the Phytelephantoideae and further consideration ofthese findings are given below.
Staminate flower cluster—Uhl and Dransfield (1984) sug-gested that in Phytelephas aequatorialis (syn. Palandra ae-quatorialis), the four bract-like appendages surrounding theperianth in the pistillate flowers are homologous to the fourbracts that subtend the flowers of the staminate cluster andtermed them ‘‘bracteoles.’’ Aphandra differs from P. aequa-torialis in that four bract-like appendages are also present be-low the perianth on each staminate flower. Barfod (1991) re-ferred to these as sepaloid bracts. The four first-formed brac-teoles on each staminate floral axis and the four bracteoles, orsepaloid bracts, of the pistillate flower are homologous. Thus,the homology is between the pistillate flower and a singleflower of the staminate cluster. The extra bracteoles might sug-gest a more branched staminate flower cluster in Aphandra.
Stamen inception—In multistaminate palms, floral recep-tacles expand to accommodate more stamens, the shape of thereceptacle and pattern of origin of primordia varying in dif-ferent groups (Uhl and Moore, 1980). The three genera ofphytelephantoid palms are distinguished by bizarre differencesin receptacle shape during stamen initiation. Floral develop-ment of two species of Phytelephas has been studied. In Phy-telephas aequatorialis the staminate flowers are pedicellatefrom early in organogenesis; the pedicels are true stalks, notpseudopedicels as in Aphandra. When the first stamen pri-mordia originate, the receptacle in P. aequatorialis is ;800mm in diameter and relatively flat. Stamen inception occursrapidly in two phases. During a first phase, the entire apexbecomes covered by primordia except for a marginal meriste-matic area. Although the primordia surrounding the center arelarger, Uhl and Moore (1977) did not observe any apices witha single ring of primordia. In the second phase, the stamenprimordia develop in centrifugal order in radial rows along themeristematic periphery and thereby increase the size of theapex. At this stage, a pistillode was observed in some flowers.Five to seven hundred stamens are usually produced in eachstaminate flower.
In Phytelephas macrocarpa, the staminate clusters and flow-ers are sessile, the flowers remaining sessile throughout de-velopment. At the time of inception of the first stamens, thereceptacle is ;550 mm wide and slightly curved with a raisedapex. Two types of meristem are active, either simultanouslyor in succession. One produces a limited number of stamenprimordia at early stages of stamen inception, apparently in acentripetal pattern (Uhl and Moore, 1977). The remaining sta-mens are produced in a centrifugal sequence by the peripheralmeristem, which is active throughout organogenesis. Pistillo-des were not observed. The number of stamens formed in eachstaminate flower varies from 150 to 300.
In species of Phytelephas and in Aphandra the receptaclesare shallow domes in early ontogenetic stages. The receptacleexpands in diameter but only slightly in height in both speciesof Phytelephas. In Aphandra natalia, however, the receptaclealso elongates, becoming somewhat funnel-shaped, with sta-men primordia originating centrifugally along the angled andflattened sides, often partly under the perianth segments. Atanthesis a naked zone remains between the level of insertionof perianth segments and the stamen-bearing part of the re-ceptacle. Our evidence is not conclusive as to whether thereis a short centripetal phase previous to centrifugal stamen in-ception in Aphandra. In some apices studied one to six pri-mordia may develop later in the center of the flower as sug-gested by the presence of inner bulges. They do not differ from

February 2001] 193BARFOD AND UHL—FLORAL DEVELOPMENT IN APHANDRA
Figs. 20–25. Staminate inflorescence. Stamen inception and development. 20. Stage VII. Floral apex of a lateral member of a flower cluster. The perianthsegments are irregular in size and shape and indistinguishable from the bracteoles. The apex is covered by stamen primordia except for a naked center. Notethat some of the inner primordia seem to develop later than the adjacent outer ones. 21. Stage VII. Floral apex with the perianth segments removed. Note theimpressions left by these in the apex and stamen primordia arising in irregular groups along their edges. 22. Stage VII. Floral apex with some of the perianthsegments removed, and stamen primordia differing in size and shape. Some stamen primordia are in the process of splitting as indicated by white arrows. 23.Stage VII. Close-up showing the naked center of the floral apex. 24. Stage VI. Distal view of the rachis showing details of adjacent four-flowered clusters. Notethe irregular perianth segments. 25. Stage IV. A flower cluster in lateral view (anterior flower to the left, lateral flower to the right). The pseudopedicel of thelateral flower is partly developed at this stage; bracteoles are indicated by white arrows.

194 [Vol. 88AMERICAN JOURNAL OF BOTANY
the surrounding stamen primordia in shape, only in size. Thebulges observed could represent the carpel primordia, but thatremains to be demonstrated. A similar developmental patternwas considered partial centrifugal development in Phytelephasmacrocarpa (Uhl and Moore, 1977, fig. 18).
In the third genus, Ammandra, the receptacle at anthesis isirregularly chunky and becomes expanded basally to cover theperianth apices on the abaxial side of the flower. The stamenfilaments are much shorter, and the anthers are smaller than inthe other phytelephantoid genera (Barfod, 1991). Button-shaped pistillodes, inserted in shallow depressions and usuallycaducous at anthesis, are almost universally present. Anatom-ical sections of the fully developed flowers show that most ofthe major trunk bundles extend to about three-fourths theheight of the flower and then curve toward the periphery andextend downward (Uhl and Moore, 1977), suggesting centrif-ugal initiation. No other observations of stamen inception havebeen made. Up to 1200 stamens have been recorded in A.dasyneura (Burret) Barfod (Barfod, 1991), the highest numberin the family.
The pseudopedicel—Floral stalks or pedicels vary amongthe phytelephantoid genera. Both sessile and pedicellate flow-ers occur in Phytelephas. The stalked condition of the stami-nate flower cluster was used to separate the genus Palandrafrom Phytelephas (Cook, 1927), but the character was foundvariable among the species of Phytelephas, and Palandra hassince been placed in synonomy (Barfod, 1991). It is notewor-thy that in Phytelephas tumacana Cook, the clusters in themiddle and proximal part of the inflorescence are composedof four (rarely five) flowers that are sessile to subsessile,whereas a number of solitary flowers are always present at theapex with pedicels up to 7 mm long (Barfod, 1991). In otherspecies of Phytelephas, as in P. macrocarpa, all flowers aresessile. Here also Ammandra differs in having stalked stami-nate flowers borne in clusters of up to nine on a 1.5–3 cmlong branch. The floral stalk appears to be a short branch, butthis requires confirmation by developmental and anatomicalstudies.
In Aphandra the pseudopedicel is a special structure, anelongate floral receptacle with an adnate and connate perianthtube. Several characters appear associated with the pedicellatecondition. The rapid expansion of the pseudopedicel may helpgenerate the pressure that is needed for the flower-bearing tis-sues to break through the PD bract. Secondly, such rapid in-crease in size may contribute to the heating of the bud aboveambient temperature during pollination. Thirdly, the epidermisdisintegrates releasing raphide idioblasts at the same time aspollen grains are shed, and the idioblasts and pollen intermin-gle, covering the surface of the pseudopedicel. In Ammandraraphide idioblasts are also found in great numbers, not scat-tered in subepidermal layers as in Aphandra, but inside blister-like structures that occur scattered on the pedicel and the re-ceptacle as well. The blisters rupture at anthesis to release theidioblasts. As in Aphandra the idioblasts resemble the roundedpollen grains in size and shape. Such idioblasts may help todeter pollen feeding and perforation of pedicels and recepta-cles for ovipositing.
Pistillate flower clusters and flowers—Except for the num-ber of flowers in the inflorescence (see Table 1), structure anddevelopment of the pistillate flower clusters and flowers inAphrandra are like those found for Phytelephas aequatorialis
where similar carpel primordia become fused laterally later indevelopment, the ventral sutures remaining open throughout(Uhl and Dransfield, 1984). The presence of two pairs of se-paloid bracts below the perianth has been noted. Two indistinctfour-parted perianth whorls are followed by a ring of condu-plicate carpel primordia. In P. aequatorialis carpels becamelaterally connate, but ventral sutures remain open and the sin-gle ovule of each carpel is initiated directly on the floral axis,in a position that might be considered axillary to the carpel.At maturity the gynoecium of all phytelephantoid genera hasa central, cone-shaped receptacle, histologically distinguishedby large parenchyma cells and scattered tannins. Similar gyn-oecial structure is found elsewhere in the family only in thegenera of the Calamoideae, where the receptacle is differentin shape and histology.
Staminodes in Aphandra originate in two whorls, the firstwhorl next to the carpel primorida and a second outside thefirst. Uhl and Moore (1977) demonstrated the same kind ofcentrifugal staminodial inception in the pistillate flowers ofPhytelephas aequatorialis.
Pollination—Several studies have been undertaken on thepollination of phytelephantoid palms. Barfod, Henderson, andBalslev (1987) and Barfod (1991) studied Phytelephas macro-carpa ssp. tenuicaulis Barfod in Amazonian Ecuador and con-cluded that several insect groups mediate pollen transfer. It isnoteworthy that beetles of the families Staphylinidae, Nituli-dae, and Curculionidae were all ovipositing in the staminateinflorescence. At the time of anthesis, they had perforated thereceptacle completely. Bernal and Ervik (1996) described thefloral biology and pollination of Phytelephas seemannii Cookin Colombia. Pollination is mostly carried out by species ofpollen-eating and predating staphylinids. One particular spe-cies of the genus Amazoncharis reproduces in the male inflo-rescences by constructing egg chambers in the fleshy recep-tacles of the flowers. This reproductive behavior resemblesthat of beetles in the closely related subtribe Gyrophaenina thatfeed on the spores and reproduce in fleshy mushrooms.
Aphandra natalia is pollinated mainly by Baridinae (Cole-optera) that feed on pollen and, in contrast to the situation inother phytelephantoid species, prefer the pistillate rather thanstaminate inflorescence for oviposition (Ervik, 1992). Thepseudopedicel may play a key role in the interaction with thevisiting insects. The raphide idioblasts that are released fromthe hypodermal layers in great numbers could be detrimentalto the larvae. Their similarity in size and shape to the pollengrains are striking (Figs. 12, 13). A simple bio-assay could bedesigned to demonstrate whether the pollinating insects areable to distinguish between the two.
Ervik, Tollsten, and Knudsen (in press) have analyzed thefloral scent of Ammandra decasperma, A. dasyneura, Aphan-dra natalia, Phytelephas aequatorialis, P. macrocarpa ssp.tenuicaulis and P. seemannii. Their results show that the majorconstituents of the floral scents of the three genera are of com-pletely different biochemical origin. The floral scent of Aphan-dra is unusual in being dominated by a pyrazine. The presenceof this compound could explain the absence in Aphandra na-talia of Derelomini and Mystrops that are common visitors inall Phytelephas species studied as well as in many other palmand cyclanth species (Henderson, 1986; Gottsberger, 1991; Er-iksson, 1994).
Phylogenetic implications—Cladistic analyses based on

February 2001] 195BARFOD AND UHL—FLORAL DEVELOPMENT IN APHANDRA
morphology and restriction site fragments found the Phytele-phantoideae monophyletic (Barfod, 1991; Uhl et al., 1995).The group is strongly supported by a number of synapomor-phies, although some of the characters are found elsewherethroughout the family, such as the multicarpellate condition(some species of Attalea), seeds attached to an extension ofthe receptacle (Eugeissona), floral buds open (e.g., Ceroxylon),and the breaking up of the fruit mesocarp into corky processes(e.g., Manicaria, Pelagodoxa, Sommiera, and Johanneste-ijsmannia).
Barfod (1991) used morphological data to infer relation-ships among genera of the Phytelephantoideae. An outgroupwas assembled representing the major evolutionary lineageswithin the palms according to Uhl and Dransfield (1987). Flo-ral characters were emphasized by weighting procedures. Theresult was ambiguous due to lack of a likely sister group, butthe cladogram favored had Aphandra and Phytelephas as aclade with Ammandra as sister group. One problem of weight-ing in favor of floral characters is that these are often the resultof coevolutionary relationships and therefore do not necces-sarily reflect phylogeny. The findings of this study further cor-robate an Aphandra–Phytelephas clade. The early, sessile on-togenetical stages of the staminate flowers of Aphandra na-talia are very similar to similar stages in Phytelephas macro-carpa. This similarity applies to number of flowers per cluster,stamen inception, length of the flower-bearing axis, and shapeand vasculature of the receptacle. Subsequent development,however, is very different.
This study further supports the circumscription of Aphandrabased on several characters of the staminate flower, in partic-ular the presence of four sepaloid bracteoles, the shape of thefloral receptacle, and a structurally unique pseudopedicel. Sta-men number, and the morphology and number of pistillateflowers per inflorescence, are also different (Table 1).
The relationship of the phytelephantoid genera to otherpalms is not yet clear. Some trees (Uhl et al., 1995) based onmorphological and molecular characters resolved the Ceroxy-leae as a sister group. A chloroplast DNA (cpDNA) restrictionfragment analysis that compares representatives of the threephytelephantoid genera with a joint outgroup of 12 taxa iscurrently underway to further seek a likely sister group. Sev-eral of the gene trees generated using simple parsimony andNypa as a functional outgroup have Ceroxylon as sister group.Tribe Ceroxyleae in the ceroxyloid palms has a Gondwanicdistribution pattern with representatives in the Juan FernandesIslands, northern South America, Madagascar, and Australia.In general the group is less specialized than the Phytelephan-toideae, but further research is necessary to establish this re-lationship. The Phytelephantoideae is of primary importancein understanding evolutionary trends within the palm family.The superficial resemblance of inflorescences and flowers withcertain cyclanthaceous groups such as Cardulovica is note-worthy. The phytelephantoid palms share with members of thisgenus, strong floral dimorphy, monopodial flower clusters,pseudopedicels, a multistaminate condition, four-merous floralwhorls, spicate inflorescences, and lack of vessels in the stem(Dahlgren, Clifford, and Yeo, 1985). There are, however, nu-merous differences, and comparative analysis of restrictionfragments (Davis, 1995) as well as DNA sequences (Chase et
al., 1995) has clearly demonstrated that these groups are onlyremotely related to each other. The respective pollination syn-dromes of the cyclanthoid genus Cardulovica and the phyt-elephantoid genera are very similar. Both groups are beetlepollinated, and they share the following features: nocturnalflowering, color, scent and temperature elevation (Barfod andHenderson, 1987; Barfod, 1991; Gottsberger, 1991; Eriksson,1994). Their similarities appear to represent a striking exampleof convergent evolution in response to uniform selection pres-sures in similar habitats.
LITERATURE CITED
BALSLEV, H., AND A. HENDERSON. 1987. A new Ammandra (Palmae) fromEcuador. Systematic Botany 12: 501–504.
BARFOD, A. S. 1991. A monographic study of the subfamily Phytelephan-toideae. Opera Botanica 104: 1–76.
———, A. HENDERSON, AND H. BALSLEV. 1987. A note on the pollinationof Phytelephas microcarpa (Palmae). Biotropica 19: 191–192.
BERNAL, R., AND F. ERVIK. 1996. Floral biology of the dioecious palm Phy-telephas seemannii in Colombia: an adaptation to staphylinid beetles.Biotropica 28: 682–696.
BORGTOFT PEDERSEN, H., AND H. BALSLEV. 1990. Ecuadorean palms foragroforestry. AAU reports 23: 1–122.
CHASE, M. W., D. W. STEVENSON, P. WILKIN, AND P. J. RUDALL. 1995.Monocot Systematics: a combined analysis. In P. J. Rudall et al. [eds.],Monocotyledons: systematics and evolution, 685–730. Royal BotanicGardens, Kew, Richmond, Surrey, UK.
COOK, O. F. 1927. New genera and species of Ivory Palms from Colombia,Ecuador and Peru. Journal of the Washington Academy of Sciences 17:218–230.
DAHLGREN, R., H. T. CLIFFORD, AND P. F. YEO. 1985. The families of themonocotyledons: structure, evolution, and taxonomy. Springer Verlag,Berlin, Germany.
DAVIS, J. I. 1995. A phylogenetic structure for the monocotyledons, as in-ferred from chloroplast dna restriction site variation and a comparisonof measures of clade support. Systematic Botany 20: 503–527.
ERIKSSON, R. 1994. The remarkable weevil pollination of the neotropicalCardulovicoideae (Cyclanthaceae). Plant Systematics and Evolution 189:75–81.
ERVIK, F. 1992. Notes on the phenology and pollination of the dioeciouspalms Mauritia flexuosa (Calamoideae) and Aphandra natalia (Phytele-phantoideae) in Ecuador. In W. Barthlott et al. [eds.], Animal-plant in-teractions in tropical environments, 7–12. Zoologisches Forschungsinsti-tut und Museum Alexander Koenig, Bonn, Germany.
ERVIK, F., L. TOLLSTEN, AND J. T. KNUDSEN. In press. Floral scent chemistryand pollination ecology in phytelephantoid palms (Arecaceae). Plant Sys-tematics and Evolution.
GOTTSBERGER, G. 1991. Pollination of some species of the Carludovicoideae,and remarks on the origin and evolution of the Cyclanthaceae. Botanis-che Jahrbucher fur Systematik 113: 221–235.
HENDERSON, A. 1986. A review of pollination studies in the Palmae. Botan-ical Review 52: 221–259.
UHL, N. W., AND J. DRANSFIELD. 1984. Development of the inflorescence,androecium, and gynoecium with reference to palms. In R. A. White andW. C. Dickison [eds.], Contemporary problems in plant anatomy, 397–449. Academic Press, New York, New York, USA.
———, AND H. E. MOORE, JR. 1977. Centrifugal stamen initiation in phy-telephantoid palms. American Journal of Botany 64: 1152–1161.
———, AND ———. 1980. Androecial development in six polyandrous gen-era representing five major groups of palms. Annals of Botany, London45: 57–75.
———, AND ———. 1987. Genera Palmarum. L.H. Bailey Hortorium andInternational Palm Society. Allen Press, Lawrence, Kansas, USA.
———, J. I. DAVIS, M. A. LUCKOW, K. S. HANSEN, AND J. J. DOYLE. 1995.Phylogenetic relationships among palms: cladistic analyses of morpho-logical and chloroplast DNA restriction site variation. In P. J. Rudall etal. [eds.], Monocotyledons: systematics and evolution, 623–661. RoyalBotanic Gardens, Kew, Richmond, Surrey, UK.





























