Embed Size (px)
Citation preview

Floral Development of the Early-Branching Papilionoid Legume Amburana cearensis(Leguminosae) Reveals Rare and Novel CharactersAuthor(s): Viviane Gonçalves Leite, Simone Padua Teixeira, Vidal Freitas Mansano, GerhardPrennerSource: International Journal of Plant Sciences, Vol. 176, No. 1 (January 2015), pp. 94-106Published by: The University of Chicago PressStable URL: http://www.jstor.org/stable/10.1086/678468 .
Accessed: 21/01/2015 03:50
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The University of Chicago Press is collaborating with JSTOR to digitize, preserve and extend access toInternational Journal of Plant Sciences.
http://www.jstor.org
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions

FLORAL DEVELOPMENT OF THE EARLY-BRANCHING PAPILIONOID LEGUME AMBURANACEARENSIS (LEGUMINOSAE) REVEALS RARE AND NOVEL CHARACTERS
Viviane Gonçalves Leite,*,† Simone Padua Teixeira,† Vidal Freitas Mansano,‡ and Gerhard Prenner1,§
*Departamento de Biologia, Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, Universidade de São Paulo (USP), Avenida Bandeirantes,3900, Ribeirão Preto, SP 14040-901, Brazil; †Departamento de Ciências Farmacêuticas, Faculdade de Ciências Farmacêuticas de Ribeirão
Preto, USP, Avenida do Café, s/n., Ribeirão Preto, SP 14040-903, Brazil; ‡Instituto de Pesquisa Jardim Botânico do Rio de Janeiro,Diretoria de Pesquisas (DIPEQ), Rua Pacheco Leão, 915 Jardim Botânico, Rio de Janeiro, RJ 22460-030, Brazil; and §Jodrell
Laboratory, Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3DS, United Kingdom
Editor: Patrick S. Herendeen
Premise of research. We present a detailed analysis of the floral ontogeny ofAmburana cearensis, which is anearly-branching papilionoid legume with an unusual floral morphology. Especially the reduced one-petalledcorolla is noteworthy and stimulates the question about the ontogenetic background of this reduction. Are all fivepetals initiated and later reduced, or are four petals completely lost (i.e., without any primordia evident)? On-togenetic studies of early-branching papilionoids provide important insights for the understanding of relation-ships down to caesalpinioid legumes and up to derived papilionoid lineages.
Methodology. We analyzed inflorescences, flowers, and flower buds with the SEM using standard pro-cedures.
Pivotal results. Sepals and petals are initiated bidirectionally, which is a rare character among papilionoidlegumes. The reduced one-petalled corolla is the result of early suppression of four petal primordia. Stamenformation is in reversed unidirectional order starting from the adaxial side of the flower. This is a rare characteramong Leguminosae. Floral characters that could be important for floral ecology are the rapid growth of theadaxial petal, which participates in bud protection; a long hypanthium with nectariferous stomata in the lowerhalf; two fenestrae at the left and right of the adaxial stamen, through which nectar is accessible; and reducedanthers in the inner stamen whorl (i.e., heteranthery), which might be related to the reduction of four petals.
Conclusions. Our study highlights the importance of ontogenetic studies among the diverse early-branchingpapilionoid lineages. Floral ontogeny inAmburana deviates considerably from the unidirectional pathway that isa common pattern among more derived papilionoid lineages. Organ reduction is based on the abortion of ini-tiated primordia. Our study contributes to a better understanding of early-branching papilionoids and theirsystematic relationships.
Keywords: Amburana, Amburaneae (Papilionoideae), evolution, Fabaceae, floral ontogeny, floral morphology,Leguminosae.
Introduction
Amburana is an early-branching papilionoid genus that con-sists of about three woody species found in non-AmazonianBrazil, Peru, Bolívia, Paraguay, andNArgentina (Ireland 2005;E. Selene, unpublished data). The genus is characterized by im-paripinnate leaves with alternate leaflets, racemose inflores-cences composed of flowers with a long and tubular hypan-thium, a corolla that consists of a single petal, free stamens,and a one-seeded fruit with a distal seed surrounded by a pa-leaceous endocarp (Spegazzini 1921; Queiroz 2009).
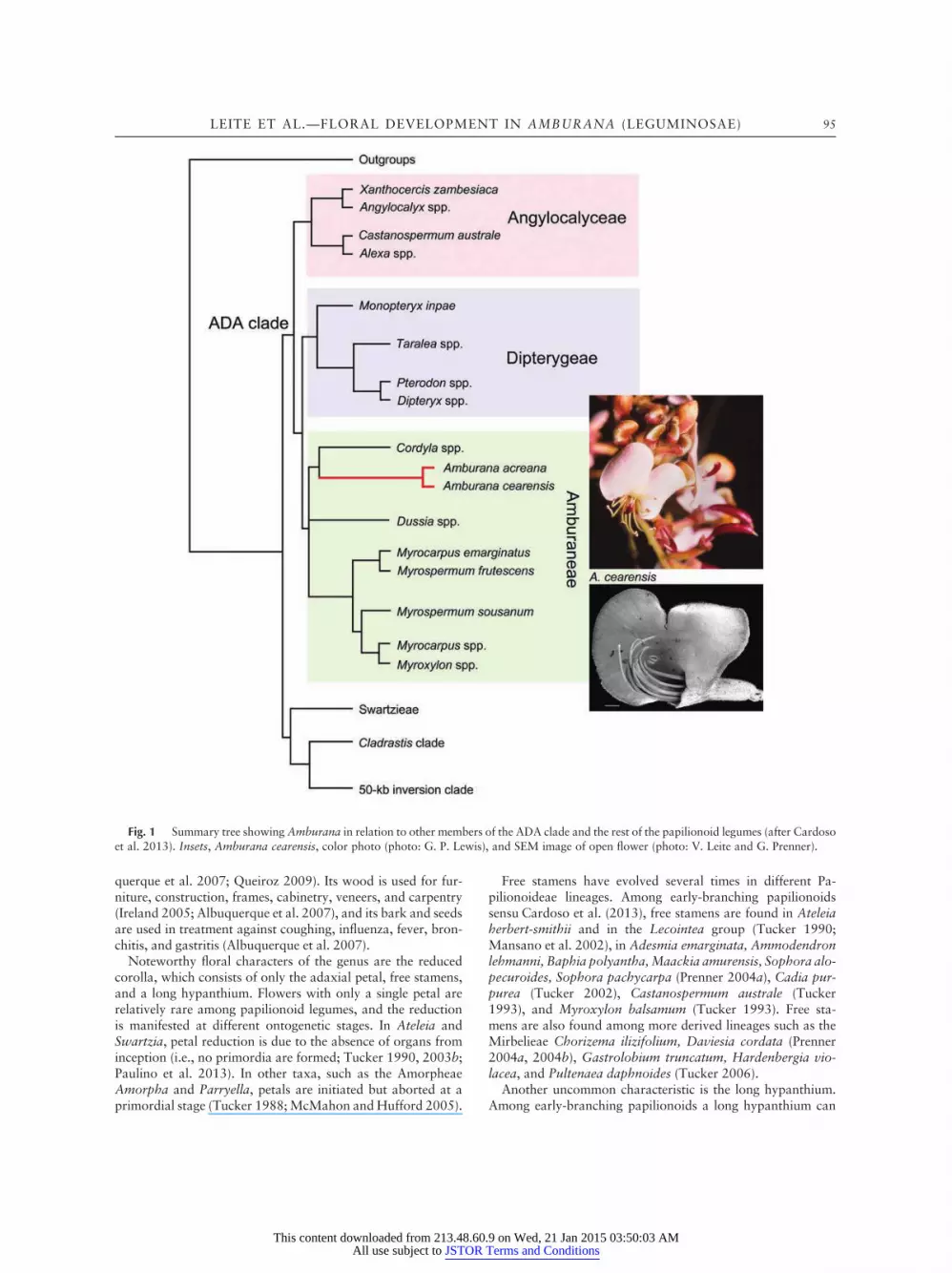
Amburana is part of the ADA clade, which includes threedistinct subclades, Angylocalyceae, Dipterygeae, and Ambu-raneae (fig. 1; Cardoso et al. 2012, 2013). The Amburaneaeclade is morphologically heterogeneous and sister to the Dip-terygeae clade. It consists of the genera Amburana, Cordyla,Dussia, Myrocarpus, Myrospermum, Myroxylon, and Mild-braediodendron, which are grouped in three subclades (fig. 1;see also Cardoso et al. 2012, 2013). One subclade is formedby the genus Dussia; another subclade consists of Amburanaand Cordyla; and the third subclade includes the remaininggenera Myrocarpus, Myrospermum, and Myroxylon. The re-lationships among the three subclades and the systematic posi-tion of the genus Mildbraediodendron are currently unclear.
Amburana cearensis (Allemão) A.C. Sm. is the most wide-spread and economically important species of the genus. Itis popularly known as imburana-de-cheiro or cumaru (Albu-
1 Author for correspondence; e-mail: [email protected].
Manuscript received May 2014; revised manuscript received July 2014; elec-tronically published December 11, 2014.
Int. J. Plant Sci. 176(1):94–106. 2015.q 2014 by The University of Chicago. All rights reserved.1058-5893/2015/17601-0008$15.00 DOI: 10.1086/678468
94
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions
querque et al. 2007; Queiroz 2009). Its wood is used for fur-niture, construction, frames, cabinetry, veneers, and carpentry(Ireland 2005; Albuquerque et al. 2007), and its bark and seedsare used in treatment against coughing, influenza, fever, bron-chitis, and gastritis (Albuquerque et al. 2007).
Noteworthy floral characters of the genus are the reducedcorolla, which consists of only the adaxial petal, free stamens,and a long hypanthium. Flowers with only a single petal arerelatively rare among papilionoid legumes, and the reductionis manifested at different ontogenetic stages. In Ateleia andSwartzia, petal reduction is due to the absence of organs frominception (i.e., no primordia are formed; Tucker 1990, 2003b;Paulino et al. 2013). In other taxa, such as the AmorpheaeAmorpha and Parryella, petals are initiated but aborted at aprimordial stage (Tucker 1988;McMahon and Hufford 2005).
Free stamens have evolved several times in different Pa-pilionoideae lineages. Among early-branching papilionoidssensu Cardoso et al. (2013), free stamens are found in Ateleiaherbert-smithii and in the Lecointea group (Tucker 1990;Mansano et al. 2002), in Adesmia emarginata, Ammodendronlehmanni, Baphia polyantha, Maackia amurensis, Sophora alo-pecuroides, Sophora pachycarpa (Prenner 2004a), Cadia pur-purea (Tucker 2002), Castanospermum australe (Tucker1993), and Myroxylon balsamum (Tucker 1993). Free sta-mens are also found among more derived lineages such as theMirbelieae Chorizema ilizifolium, Daviesia cordata (Prenner2004a, 2004b), Gastrolobium truncatum, Hardenbergia vio-lacea, and Pultenaea daphnoides (Tucker 2006).Another uncommon characteristic is the long hypanthium.
Among early-branching papilionoids a long hypanthium can
Fig. 1 Summary tree showing Amburana in relation to other members of the ADA clade and the rest of the papilionoid legumes (after Cardosoet al. 2013). Insets, Amburana cearensis, color photo (photo: G. P. Lewis), and SEM image of open flower (photo: V. Leite and G. Prenner).
LEITE ET AL.—FLORAL DEVELOPMENT IN AMBURANA (LEGUMINOSAE) 95
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions
be found only in Angylocalyx (G. Prenner, personal observa-tion), which belongs to the Angylocalyceae (sister to Diptery-geae 1 Amburaneae; see fig. 1). A short hypanthium is foundin Myroxylon (Tucker 1993) and in Dipterygeae (Leite et al.2014), which is sister to the Amburaneae clade (Cardoso et al.2013).
Based on a detailed ontogenetic study of the flowers of A.cearensis, we aim to answer the following questions: (1) Whatis the ontogenetic basis for the single-petalled corolla in Am-burana, and do Amburana, Swartzia, and Ateleia share onto-genic pathways leading to this feature? (2) Does the reductionin petal number have an impact on organ formation in otherwhorls? (3) Are there similarities in floral ontogeny amongAmburaneae and between Amburaneae and its sister tribeDipterygeae? Answering these questions will help in under-standing floral evolution in the early-branching papilionoidlegumes and their systematic relationships.
Material and Methods
Floral buds at different stages of development and flowerswere collected in June 2011 in Bahia and Rio de Janeiro, Brazil.Voucher specimens were deposited in RB under the followingregister numbers: Seleme, E.P.n. 460; Nunes, N.L. 19.
For SEM, samples were fixed in FAA 50 (Johansen 1940),dissected in 70% alcohol under a stereomicroscope, critical-point dried using an Autosamdri-815B critical-point dryer,mounted on aluminum stubs with clear nail polish, and coatedwith platinum in aQuorumQ150T sputter-coater. SEM imageswere taken with a Hitachi S-4700-II cold field emission SEM.Images were processed using Adobe Photoshop CS5.
The terminology used to describe the floral ontogeny fol-lowed Tucker (1987, 1997, 2003b), Klitgaard (1999), andPrenner (2004a, 2004c, 2013b). The adaxial side of the flower,which is the closest to the inflorescence axis, is at the top ofeach image; the abaxial side of the flower is the lower one,which is opposite the inflorescence axis and closest to the bract.
Results
Flower Morphology
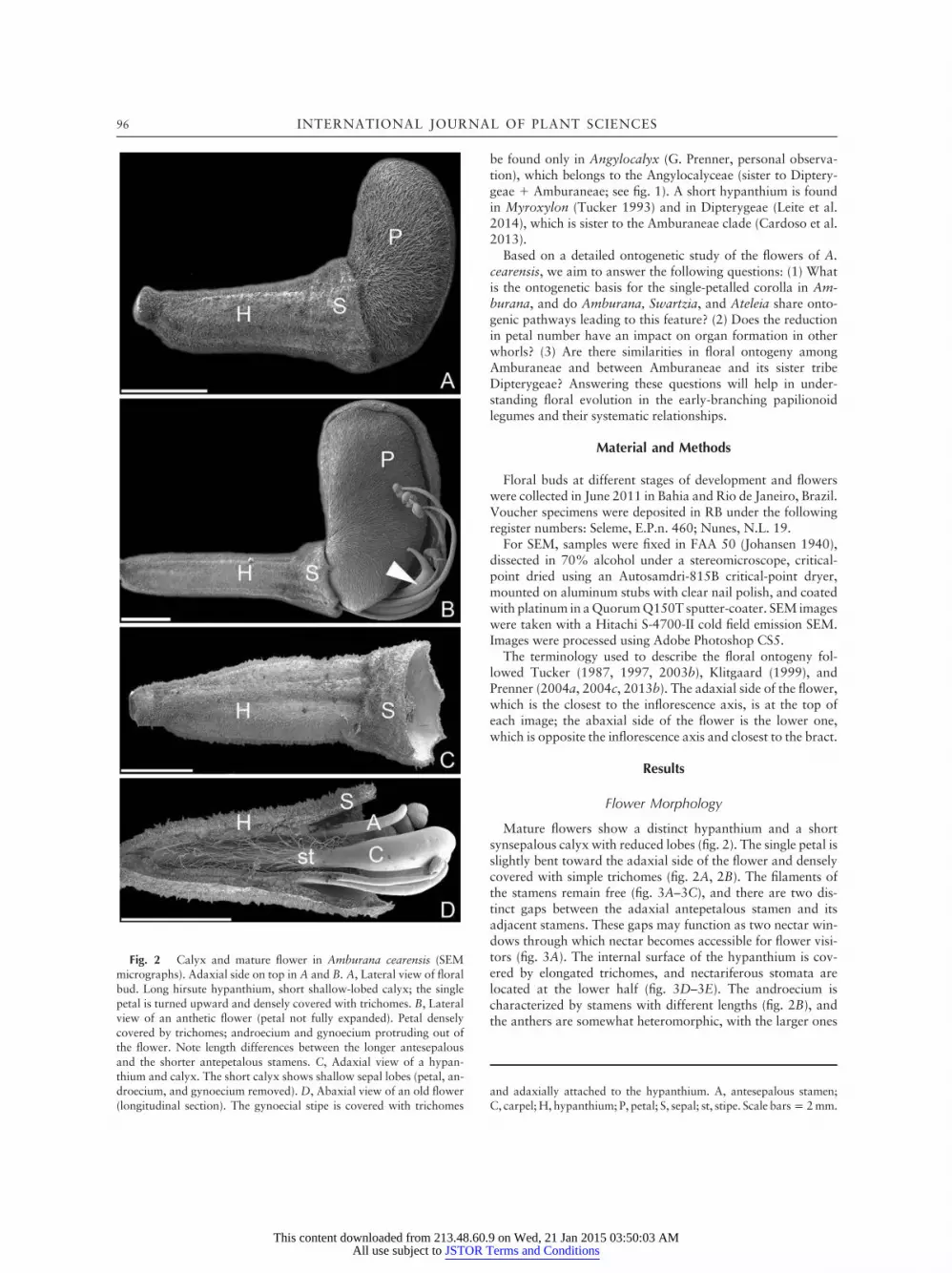
Mature flowers show a distinct hypanthium and a shortsynsepalous calyx with reduced lobes (fig. 2). The single petal isslightly bent toward the adaxial side of the flower and denselycovered with simple trichomes (fig. 2A, 2B). The filaments ofthe stamens remain free (fig. 3A–3C), and there are two dis-tinct gaps between the adaxial antepetalous stamen and itsadjacent stamens. These gaps may function as two nectar win-dows through which nectar becomes accessible for flower visi-tors (fig. 3A). The internal surface of the hypanthium is cov-ered by elongated trichomes, and nectariferous stomata arelocated at the lower half (fig. 3D–3E). The androecium ischaracterized by stamens with different lengths (fig. 2B), andthe anthers are somewhat heteromorphic, with the larger ones
Fig. 2 Calyx and mature flower in Amburana cearensis (SEMmicrographs). Adaxial side on top in A and B. A, Lateral view of floralbud. Long hirsute hypanthium, short shallow-lobed calyx; the singlepetal is turned upward and densely covered with trichomes. B, Lateralview of an anthetic flower (petal not fully expanded). Petal denselycovered by trichomes; androecium and gynoecium protruding out ofthe flower. Note length differences between the longer antesepalousand the shorter antepetalous stamens. C, Adaxial view of a hypan-thium and calyx. The short calyx shows shallow sepal lobes (petal, an-droecium, and gynoecium removed). D, Abaxial view of an old flower(longitudinal section). The gynoecial stipe is covered with trichomes
and adaxially attached to the hypanthium. A, antesepalous stamen;C, carpel; H, hypanthium; P, petal; S, sepal; st, stipe. Scale barsp 2mm.
96 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions

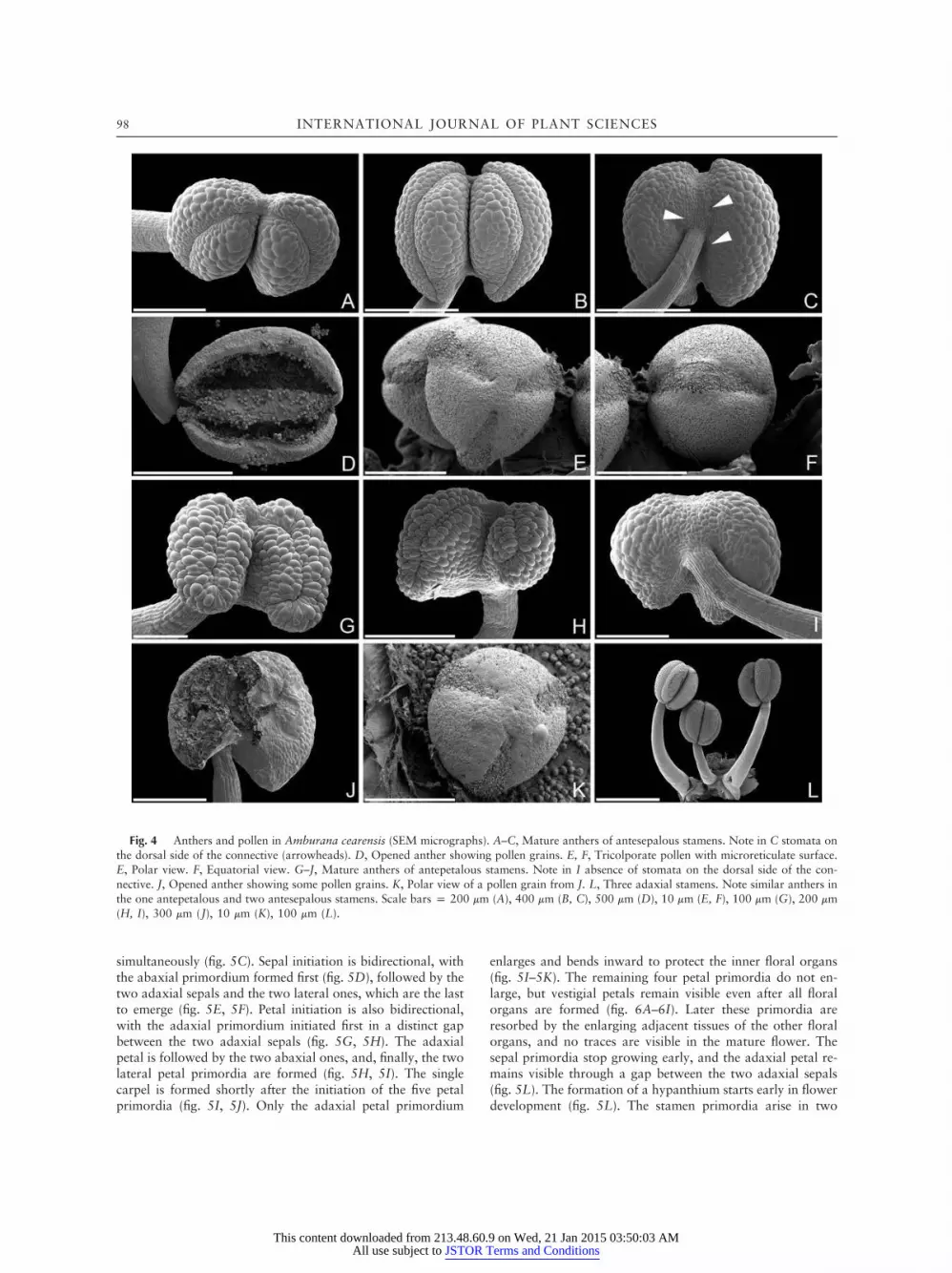
in the antesepalous whorl (fig. 4A–4D). These anthers havestomata on their connective (fig. 4C). All anthers are appen-diculate, dithecous, and longitudinally dehiscent (fig. 4B, 4D).Anthers in the antepetalous whorl are smaller and lack stomataon the connective (fig. 4G–4J). Pollen grains from the two an-ther types do not show any significant difference; they aretricolporate with a microreticulate surface (fig. 4E, 4F, 4K).However, pollen quantity seems to be highly reduced in thereduced anther type (fig. 4D, 4J). The gynoecium is mono-
carpellate; the carpel stipe is coveredwith simple trichomes, andit is adaxially attached to the hypanthium (figs. 2D, 3A, 7D, 8).
Floral Ontogeny
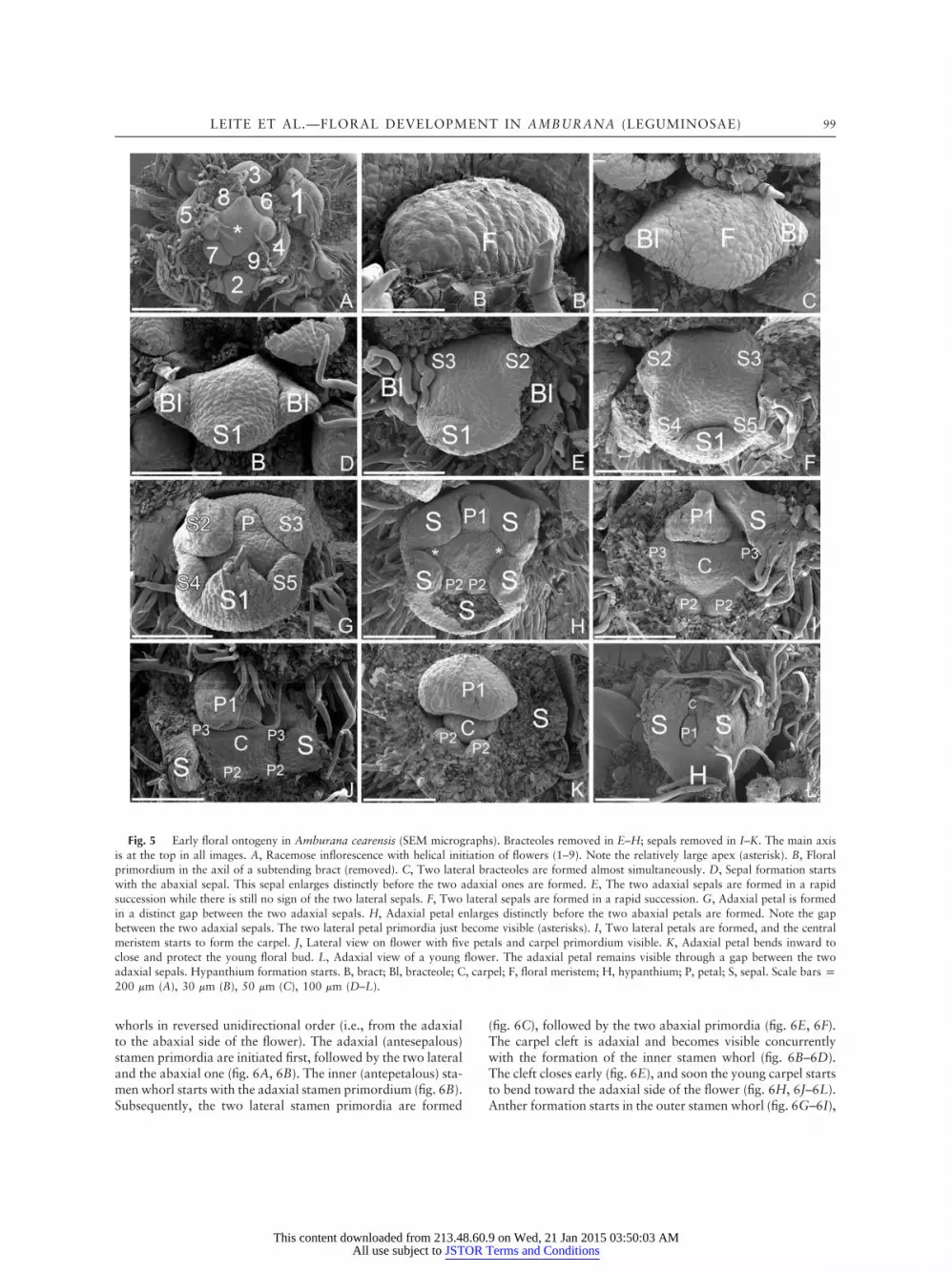
On the inflorescence axis, the flowers are initiated acrope-tally and in helical order (fig. 5A). The transversely elongatedfloral primordium is subtended by a bract (fig. 5A, 5B) andpreceded by two lateral bracteoles, which are formed almost
Fig. 3 Hypanthium of mature flower in Amburana cearensis (SEM micrographs). A, Adaxial view of mature flower (longisection). Tri-chomes on carpel stipe and hypanthium wall. Note the two gaps between the adaxial antepetalous stamen and its adjacent stamens (asterisks).B, C, Abaxial (B) and lateral (C) view of hypanthium in mature flower (longisection). Hypanthium with trichomes and stomata in its lower half.Stamen filaments are free and without distinct gaps. D, E, Detail of B and C, showing nectar stomata from the lower half of the hypanthium.A, outer whorl stamen; a, inner whorl stamen; C, carpel; H, hypanthium; S, sepal. Scale bars p 1 mm (A–C), 20 mm (D, E).
LEITE ET AL.—FLORAL DEVELOPMENT IN AMBURANA (LEGUMINOSAE) 97
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions
simultaneously (fig. 5C). Sepal initiation is bidirectional, withthe abaxial primordium formed first (fig. 5D), followed by thetwo adaxial sepals and the two lateral ones, which are the lastto emerge (fig. 5E, 5F). Petal initiation is also bidirectional,with the adaxial primordium initiated first in a distinct gapbetween the two adaxial sepals (fig. 5G, 5H). The adaxialpetal is followed by the two abaxial ones, and, finally, the twolateral petal primordia are formed (fig. 5H, 5I). The singlecarpel is formed shortly after the initiation of the five petalprimordia (fig. 5I, 5J). Only the adaxial petal primordium
enlarges and bends inward to protect the inner floral organs(fig. 5I–5K). The remaining four petal primordia do not en-large, but vestigial petals remain visible even after all floralorgans are formed (fig. 6A–6I). Later these primordia areresorbed by the enlarging adjacent tissues of the other floralorgans, and no traces are visible in the mature flower. Thesepal primordia stop growing early, and the adaxial petal re-mains visible through a gap between the two adaxial sepals(fig. 5L). The formation of a hypanthium starts early in flowerdevelopment (fig. 5L). The stamen primordia arise in two
Fig. 4 Anthers and pollen in Amburana cearensis (SEM micrographs). A–C, Mature anthers of antesepalous stamens. Note in C stomata onthe dorsal side of the connective (arrowheads). D, Opened anther showing pollen grains. E, F, Tricolporate pollen with microreticulate surface.E, Polar view. F, Equatorial view. G–J, Mature anthers of antepetalous stamens. Note in I absence of stomata on the dorsal side of the con-nective. J, Opened anther showing some pollen grains. K, Polar view of a pollen grain from J. L, Three adaxial stamens. Note similar anthers inthe one antepetalous and two antesepalous stamens. Scale bars p 200 mm (A), 400 mm (B, C), 500 mm (D), 10 mm (E, F), 100 mm (G), 200 mm(H, I), 300 mm (J), 10 mm (K), 100 mm (L).
98 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions
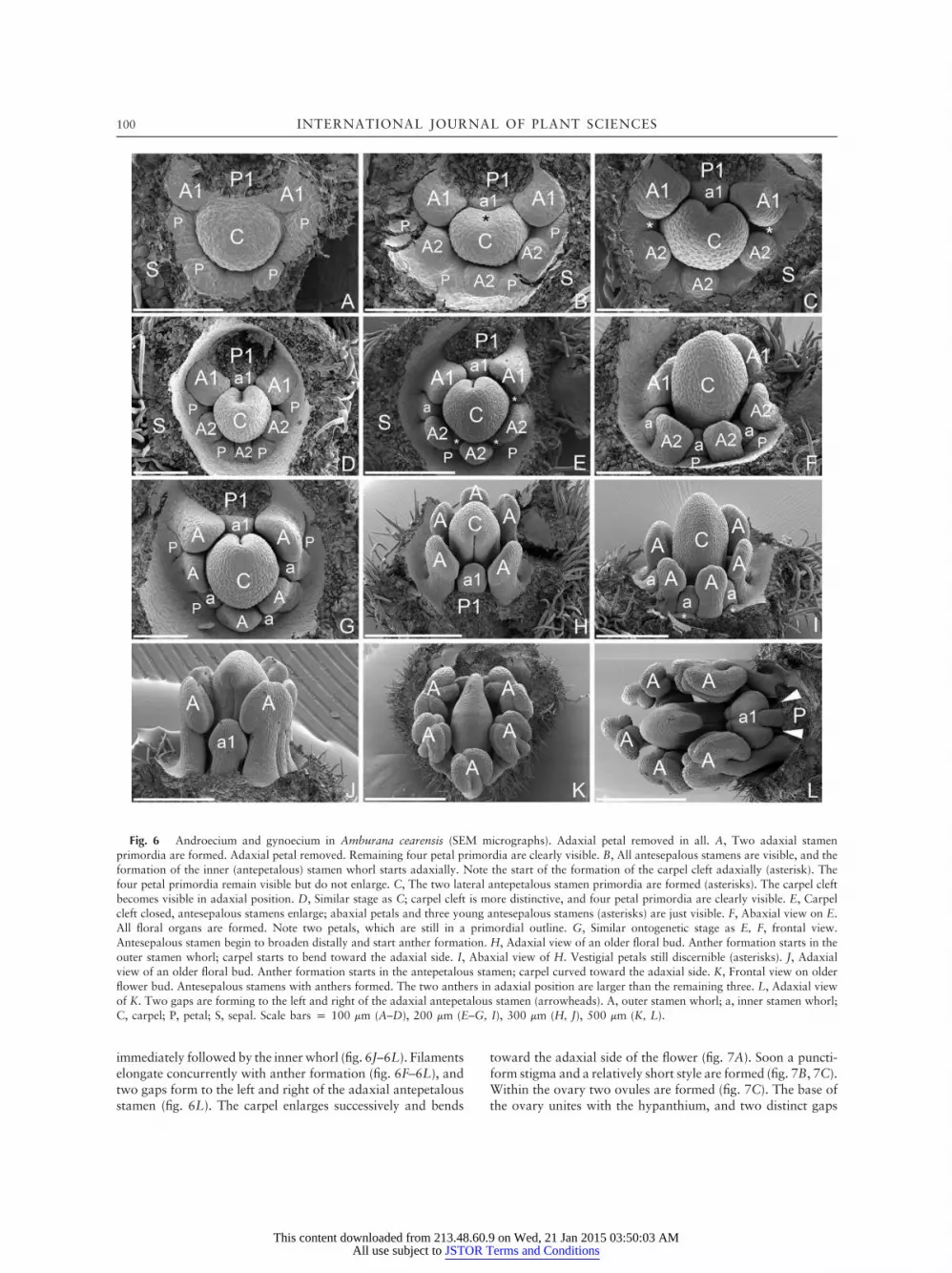
whorls in reversed unidirectional order (i.e., from the adaxialto the abaxial side of the flower). The adaxial (antesepalous)stamen primordia are initiated first, followed by the two lateraland the abaxial one (fig. 6A, 6B). The inner (antepetalous) sta-men whorl starts with the adaxial stamen primordium (fig. 6B).Subsequently, the two lateral stamen primordia are formed
(fig. 6C), followed by the two abaxial primordia (fig. 6E, 6F).The carpel cleft is adaxial and becomes visible concurrentlywith the formation of the inner stamen whorl (fig. 6B–6D).The cleft closes early (fig. 6E), and soon the young carpel startsto bend toward the adaxial side of the flower (fig. 6H, 6J–6L).Anther formation starts in the outer stamen whorl (fig. 6G–6I),
Fig. 5 Early floral ontogeny in Amburana cearensis (SEM micrographs). Bracteoles removed in E–H; sepals removed in I–K. The main axisis at the top in all images. A, Racemose inflorescence with helical initiation of flowers (1–9). Note the relatively large apex (asterisk). B, Floralprimordium in the axil of a subtending bract (removed). C, Two lateral bracteoles are formed almost simultaneously. D, Sepal formation startswith the abaxial sepal. This sepal enlarges distinctly before the two adaxial ones are formed. E, The two adaxial sepals are formed in a rapidsuccession while there is still no sign of the two lateral sepals. F, Two lateral sepals are formed in a rapid succession. G, Adaxial petal is formedin a distinct gap between the two adaxial sepals. H, Adaxial petal enlarges distinctly before the two abaxial petals are formed. Note the gapbetween the two adaxial sepals. The two lateral petal primordia just become visible (asterisks). I, Two lateral petals are formed, and the centralmeristem starts to form the carpel. J, Lateral view on flower with five petals and carpel primordium visible. K, Adaxial petal bends inward toclose and protect the young floral bud. L, Adaxial view of a young flower. The adaxial petal remains visible through a gap between the twoadaxial sepals. Hypanthium formation starts. B, bract; Bl, bracteole; C, carpel; F, floral meristem; H, hypanthium; P, petal; S, sepal. Scale bars p200 mm (A), 30 mm (B), 50 mm (C), 100 mm (D–L).
LEITE ET AL.—FLORAL DEVELOPMENT IN AMBURANA (LEGUMINOSAE) 99
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions
immediately followed by the inner whorl (fig. 6J–6L). Filamentselongate concurrently with anther formation (fig. 6F–6L), andtwo gaps form to the left and right of the adaxial antepetalousstamen (fig. 6L). The carpel enlarges successively and bends
toward the adaxial side of the flower (fig. 7A). Soon a puncti-form stigma and a relatively short style are formed (fig. 7B, 7C).Within the ovary two ovules are formed (fig. 7C). The base ofthe ovary unites with the hypanthium, and two distinct gaps
Fig. 6 Androecium and gynoecium in Amburana cearensis (SEM micrographs). Adaxial petal removed in all. A, Two adaxial stamenprimordia are formed. Adaxial petal removed. Remaining four petal primordia are clearly visible. B, All antesepalous stamens are visible, and theformation of the inner (antepetalous) stamen whorl starts adaxially. Note the start of the formation of the carpel cleft adaxially (asterisk). Thefour petal primordia remain visible but do not enlarge. C, The two lateral antepetalous stamen primordia are formed (asterisks). The carpel cleftbecomes visible in adaxial position. D, Similar stage as C; carpel cleft is more distinctive, and four petal primordia are clearly visible. E, Carpelcleft closed, antesepalous stamens enlarge; abaxial petals and three young antesepalous stamens (asterisks) are just visible. F, Abaxial view on E.All floral organs are formed. Note two petals, which are still in a primordial outline. G, Similar ontogenetic stage as E, F, frontal view.Antesepalous stamen begin to broaden distally and start anther formation. H, Adaxial view of an older floral bud. Anther formation starts in theouter stamen whorl; carpel starts to bend toward the adaxial side. I, Abaxial view of H. Vestigial petals still discernible (asterisks). J, Adaxialview of an older floral bud. Anther formation starts in the antepetalous stamen; carpel curved toward the adaxial side. K, Frontal view on olderflower bud. Antesepalous stamens with anthers formed. The two anthers in adaxial position are larger than the remaining three. L, Adaxial viewof K. Two gaps are forming to the left and right of the adaxial antepetalous stamen (arrowheads). A, outer stamen whorl; a, inner stamen whorl;C, carpel; P, petal; S, sepal. Scale bars p 100 mm (A–D), 200 mm (E–G, I), 300 mm (H, J), 500 mm (K, L).
100 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions
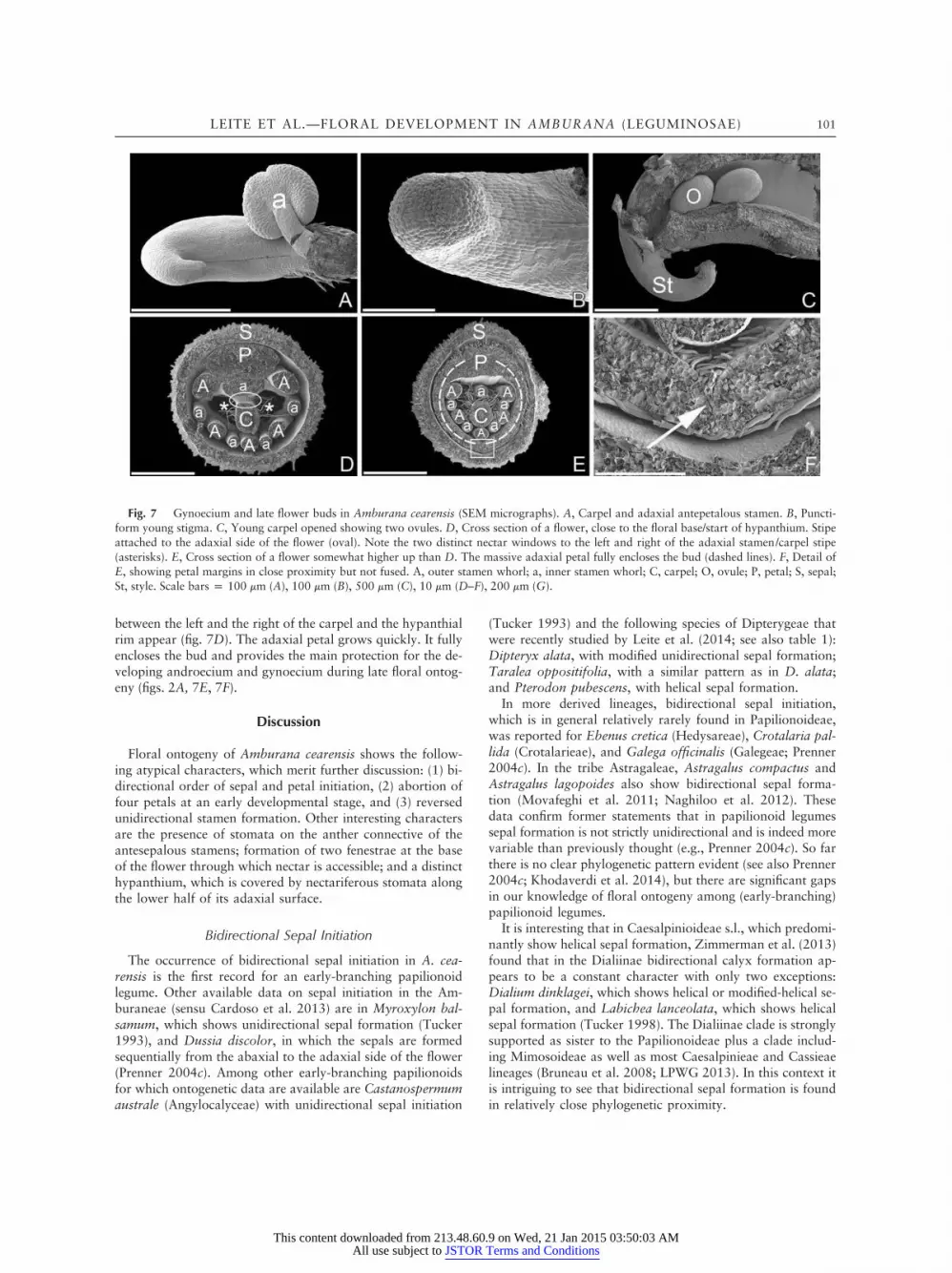
between the left and the right of the carpel and the hypanthialrim appear (fig. 7D). The adaxial petal grows quickly. It fullyencloses the bud and provides the main protection for the de-veloping androecium and gynoecium during late floral ontog-eny (figs. 2A, 7E, 7F).
Discussion
Floral ontogeny of Amburana cearensis shows the follow-ing atypical characters, which merit further discussion: (1) bi-directional order of sepal and petal initiation, (2) abortion offour petals at an early developmental stage, and (3) reversedunidirectional stamen formation. Other interesting charactersare the presence of stomata on the anther connective of theantesepalous stamens; formation of two fenestrae at the baseof the flower through which nectar is accessible; and a distincthypanthium, which is covered by nectariferous stomata alongthe lower half of its adaxial surface.
Bidirectional Sepal Initiation
The occurrence of bidirectional sepal initiation in A. cea-rensis is the first record for an early-branching papilionoidlegume. Other available data on sepal initiation in the Am-buraneae (sensu Cardoso et al. 2013) are in Myroxylon bal-samum, which shows unidirectional sepal formation (Tucker1993), and Dussia discolor, in which the sepals are formedsequentially from the abaxial to the adaxial side of the flower(Prenner 2004c). Among other early-branching papilionoidsfor which ontogenetic data are available are Castanospermumaustrale (Angylocalyceae) with unidirectional sepal initiation
(Tucker 1993) and the following species of Dipterygeae thatwere recently studied by Leite et al. (2014; see also table 1):Dipteryx alata, with modified unidirectional sepal formation;Taralea oppositifolia, with a similar pattern as in D. alata;and Pterodon pubescens, with helical sepal formation.In more derived lineages, bidirectional sepal initiation,
which is in general relatively rarely found in Papilionoideae,was reported for Ebenus cretica (Hedysareae), Crotalaria pal-lida (Crotalarieae), and Galega officinalis (Galegeae; Prenner2004c). In the tribe Astragaleae, Astragalus compactus andAstragalus lagopoides also show bidirectional sepal forma-tion (Movafeghi et al. 2011; Naghiloo et al. 2012). Thesedata confirm former statements that in papilionoid legumessepal formation is not strictly unidirectional and is indeed morevariable than previously thought (e.g., Prenner 2004c). So farthere is no clear phylogenetic pattern evident (see also Prenner2004c; Khodaverdi et al. 2014), but there are significant gapsin our knowledge of floral ontogeny among (early-branching)papilionoid legumes.It is interesting that in Caesalpinioideae s.l., which predomi-
nantly show helical sepal formation, Zimmerman et al. (2013)found that in the Dialiinae bidirectional calyx formation ap-pears to be a constant character with only two exceptions:Dialium dinklagei, which shows helical or modified-helical se-pal formation, and Labichea lanceolata, which shows helicalsepal formation (Tucker 1998). The Dialiinae clade is stronglysupported as sister to the Papilionoideae plus a clade includ-ing Mimosoideae as well as most Caesalpinieae and Cassieaelineages (Bruneau et al. 2008; LPWG 2013). In this context itis intriguing to see that bidirectional sepal formation is foundin relatively close phylogenetic proximity.
Fig. 7 Gynoecium and late flower buds in Amburana cearensis (SEM micrographs). A, Carpel and adaxial antepetalous stamen. B, Puncti-form young stigma. C, Young carpel opened showing two ovules. D, Cross section of a flower, close to the floral base/start of hypanthium. Stipeattached to the adaxial side of the flower (oval). Note the two distinct nectar windows to the left and right of the adaxial stamen/carpel stipe(asterisks). E, Cross section of a flower somewhat higher up than D. The massive adaxial petal fully encloses the bud (dashed lines). F, Detail ofE, showing petal margins in close proximity but not fused. A, outer stamen whorl; a, inner stamen whorl; C, carpel; O, ovule; P, petal; S, sepal;St, style. Scale bars p 100 mm (A), 100 mm (B), 500 mm (C), 10 mm (D–F), 200 mm (G).
LEITE ET AL.—FLORAL DEVELOPMENT IN AMBURANA (LEGUMINOSAE) 101
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions

The Corolla: Bidirectional Initiation andEarly Abortion of Primordia
The peculiar one-petalled corolla of A. cearensis differsstrongly from the typical papilionoid corolla, which consists ofan adaxial vexillum (or standard petal), two lateral wing petals,and two abaxial keel petals. In A. cearensis, the adaxial vexil-lum is the only remaining petal, and no petal vestiges can befound in mature flowers. Our study shows that this reductionis the result of the abortion of four petal primordia during earlyto middevelopmental stages. Petal primordia do not enlargeand are resorbed early in development so that no vestiges of thefour primordia can be detected in the mature flower.
Reduction in petal number is rare within papilionoid le-gumes. A corolla is absent in the Amburaneae genera Cordylaand Mildbraediodendron (Cowan 1981), which are sister toAmburana (Cardoso et al. 2013). No ontogenetic studies areavailable for these taxa. More taxa with a reduced petal num-ber are found in Swartzieae (sensu Cardoso et al. 2013), whereonly a single petal or no petal is formed in Swartzia (Tucker2003b; Paulino et al. 2013), Bobgunnia (Tucker 2003b), and
Ateleia (Tucker 1990; see table 2). As in Amburana, in thesetaxa the adaxial petal is the only petal that is found in themature flower, but in contrast with Amburana, in these taxajust one petal primordium is present and the other primordiado not initiate (they can be classified as lost organs sensu Tucker1988). Bocoa, Cyathostegia, Candolleodendron, and Trischi-dium also have only one petal (Cowan 1981; Polhill 1981;Ireland 2007), but no ontogenetic data are available for thesetaxa.
Taxa with a reduced number of petals are also found in theAmorpheae, which is part of the Dalbergioids s.l. and belongsto a large clade of Papilionoideae with the 50-kb inversion (seeCardoso et al. 2013). In Amorpha, only the adaxial petal de-velops, while the remaining four petals are aborted at a pri-mordial stage (Tucker 1988; McMahon and Hufford 2005).In Parryella filifolia, which lacks petals entirely, McMahonand Hufford (2005) found little evidence for any petal pri-mordium (i.e., maybe a complete loss of petals). Mature flow-ers of Errazurizia rotundata, which is sister to P. filifolia, alsodo not show petals (McMahon and Hufford 2005). A de-tailed ontogenetic analysis of this species is lacking.
Compared with the other two subfamilies, it is intriguingthat petal reduction is more frequent among caesalpinioid le-gumes. Most recently, Bruneau et al. (2014) showed that co-rolla merosity changed 15 times in their sampling of Detarieae,and Fougére-Danezan et al. (2009) noted that in the resin-producingDetarieae a complete loss of petals occurred six timesin parallel.
Considering that papilionoid legumes evolved from caesal-pinioid ancestors, we speculate that to a certain extent thepronounced experimental phase (postulated by Prenner andKlitgaard 2008) that occurs among Caesalpinioideae s.l. alsoapplies to early-branching papilionoid legumes such as Am-buraneae. A pattern such as petal reduction also occurs in morederived taxa within Papilionoideae such as the Amorpheae. Butexceptions from the pattern of flowers consisting of 21 organsin five whorls become increasingly rare among the most derivedNPAAA clade, where we frequently find variation in petalmorphology but not in petal merosity. Exceptions include petalabortion in the subterranean flowers of the Phaseoleae Amphi-carpaea edgeworthii (Zhang et al. 2006) or petal reduction insome Erythrina species (Tucker 1987). However, in these taxa,organ suppression takes place at a much later ontogenetic stageand could be regarded as an ecological adaptation. Due to thesubterranean flowering in A. edgeworthii, petals are no longerneeded to attract pollinators, and in Erythrina petal reductioncan be correlated with certain pollination syndromes such asbird or bat pollination (see Arroyo 1981).
Another noteworthy pattern in the petal whorl of A. cea-rensis is the bidirectional initiation, starting with the adaxialpetal, which is followed by the two abaxial and, finally, thetwo lateral ones, formed last. Here, we speculate that theremight be a correlation between petal reduction and this ini-tiation pattern. The first petal formed is the only petal thatwill enlarge and become visible in the mature flower, whilethe others are early reduced or resorbed during ontogeny.
To our knowledge, there is only one other record of bidirec-tional petal formation in Papilionoideae, which was recentlydescribed in some species of Hedysareae (Khodaverdi et al.2014). However, in these taxa, a complete pentamerous co-rolla is formed, and no correlation with organ reduction or
Fig. 8 Amburana cearensis. Floral diagram and floral formula(the latter format following Prenner et al. 2010). The inflorescenceaxis is on top (crossed circle). The flower is subtended by a bract andpreceded by two lateral bracteoles (all in black), followed by fivefused sepals (green), five petals where just one of them develops intothe adaxial standard petal and four are aborted in the primordialstage (red), 10 free stamens (yellow) in two whorls of five, and anunicarpellate gynoecium in the floral center (blue). The stipe of thecarpel is fused to the adaxial part of the hypanthium (not shown inthe diagram and formula). Two ovules are attached to the ovary by amarginal placentation (Vm). A, androecium; C, corolla; G, gynoecium;K, calyx; m, marginal placentation; V, ovules.
102 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions

abortion is evident. Khodaverdi et al. (2014) highlighted thatbidirectional organ formation has evolved several times in-dependently among papilionoid legumes. Moço and Mariath(2009) proposed a bidirectional petal formation in Adesmiamuricata. However, their images do not show this charac-ter very clearly, and a simultaneous formation seems equallypossible.
Is There a Relation of Petal Reduction and Heteranthery?
The heteromorphic stamens are another puzzling characterof A. cearensis. One explanation for this feature could be thatthe inner, shorter stamen whorl, which has reduced anthers,might functionally replace wing and keel petals of a papil-ionaceous flower. In typical papilionaceous flowers, keel andwing petals have important functions in the pollination pro-cess. They act as a landing platform for floral visitors, andthey enclose the androecium and gynoecium. In the one-petalled corolla of A. cearensis, some of these functions maybe transferred to the inner stamen whorl; floral visitors coulduse the stamens to hold onto the flower while they inserttheir proboscis to the left and right of the flower, where thetwo fenestrae allow them access to the nectar exuded fromnectariferous stomata in the lower half of the hypanthium.Studies of floral biology are necessary to verify this hypothesis.
Tucker (1996) highlights the occurrence of androecial het-eromorphy in a wider range of legume flowers. Among Papil-ionoideae she highlights Swartzia macrosperma and Swartziasimplex, which show three larger stamens and about 60–70smaller ones (see also Tucker 2003b). Paulino et al. (2013)showed that in Swartzia dipetala pollen from larger and smalleranthers differed in reserve content. The authors propose that thelarger anthers produce pollen for reproduction, whereas thesmaller ones produce pollen for feeding. In other papilionoidlegumes with regular flag blossoms, heteromorphic anthers areimportant for the presentation of pollen in the so-called pumpmechanism in which the smaller anthers act as pistons movingpollen out of the keel (see Arroyo 1981).
The pattern of two nectar windows to the left and right ofthe adaxial antepetalous stamen, as found in A. cearensis, is arelatively common character and was also reported in othertribes such as Adesmieae, Carmichaelieae, Cicereae, Desmo-dieae, Galegeae, Hedysareae, Indigofereae, Liparieae, Loteae,Millettieae, Mirbelieae, Phaseoleae, Podalyrieae, Psoraleae,Robinieae, Sophoreae, and Vicieae (Prenner 2004a, 2013a).The position of the nectariferous stomata on the inner faceof the hypanthium, which was also described in the tribeDipterygeae (Leite et al. 2014), seems to be a characteristic ofsome lineages of Papilionoideae, such as the tribe Vicieae(Prenner 2003b), two reports in Dalbergieae (Klitgaard 1999),one report in Loteae (Prenner 2003a), and 13 reports forGenisteae (Vogel 1997).
Reversed Unidirectional Order of Stamen Initiation
Both stamen whorls are initiated in reversed unidirectionalorder starting adaxially and proceeding in abaxial direction.This is the reversed pattern of the frequently found unidi-rectional pattern that starts abaxially and proceeds towardthe adaxial side of the flower (see Gemmeke 1982; Tucker2003a; Prenner 2004b, 2004c). So far, reversed unidirectionalstamen formation was rarely documented among papilionoidlegumes. Prenner (2004b) found it in the outer stamen whorlof Daviesia cordata (Mirbelieae), and Moço and Mariath(2009) showed it in the sepal and stamens of several Adesmiaspecies (Adesmieae). It is interesting that in all these taxa thestamens are mainly free. Further investigations are necessaryto see whether there is a connection between filament unionand the stamen initiation pattern. Is there a different under-lying organization pattern among united and free androecia?Among mimosoid legumes, reversed unidirectional organ
formation was reported in the sepal whorl of Inga grandis,Inga hispida (Paulino 2012), Lysiloma vogelianum (Gemmeke1982), and Calliandra angustifolia (Prenner 2004d). In Cae-salpinioideae s.l. reversed unidirectional organ formation wasreported in Sindora klaineana (for both stamen whorls) and in
Table 1
Floral Ontogeny and Morphology in Amburana cearensis and the Dipterygeae Clade
Amburana cearensisa Dipteryx alatab Pterodon pubescensb Taralea oppositifoliab
Order of bracteole initiation Simultaneous Asynchronous Simultaneous SimultaneousOrder of sepal initiation Bidirectional Modified unidirectional Helical Sequential or modified
sequentialOrder of petal initiation Bidirectional Simultaneous Simultaneous SimultaneousNo. petals initiated 5 5 5 5No. petals in the matureflower 1 5 5 5
Order of antesepalousstamens Reversed unidirectional Modified unidirectional Modified unidirectional Modified unidirectional
Stigma shape Punctiform Truncate Capitate PunctiformStyle curvature Curved Curved Curved ErectStipe length Long Long Long ShortOvary surface Glabrous Glabrous Glabrous PiloseOccurrence of antherappendix Absent Present (discrete) Present (conspicuous) Absent
Type of hypanthium Long Short Short ShortNectary structure Nectary modified stomata Nectary modified stomata Nectary modified stomata Nectary modified stomata
a This study.b Leite et al. (2014).
LEITE ET AL.—FLORAL DEVELOPMENT IN AMBURANA (LEGUMINOSAE) 103
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions

Julbernardia pellegriniana (in petals and both stamen whorls;Tucker 2003a).
Outlook
Our study gives some insights into the floral morphologyand evolution among early-branching papilionoid legumes.Amburana cearensis shares some ontogenetic characters withits sister clade Dipterygeae, such as number of petals initiatedand the presence of modified stomata on the hypanthial nec-tary (table 1). Other characters, such as organ reduction andbidirectional and reversed unidirectional organ formation, arefound only rarely among papilionoid legumes, and most ofthese rare characters seem to be more common among early-branching lineages. Therefore, the data acquired lead to im-portant questions about the conditions in other taxa that areso far unstudied; studies of these taxa will help us to betterunderstand the rather complex developmental patterns foundin the calyx whorl of early-branching papilionoids. An in-depth understanding of these species-poor early-branchingpapilionoids is vital for the understanding of legumes as awhole, because they represent a link both down to the para-phyletic subfamily Caesalpinioideae and up toward more de-rived papilionoid lineages, which harbor some of the mostsuccessful plant genera such as Astragalus (2300–2500 spe-cies), Indigofera (ca. 700 species), and Crotalaria (ca. 690 spe-cies; Lewis et al. 2005). Among early-branching papilionoid
legumes some rarely found ontogenetic patterns point to-ward an extended experimental phase during which a broaderrange of ontogenetic and morphological pathways were re-alized. It is noticeable that these taxa consist of mainly mod-erate species number. The breadth of morphological and on-togenetic variability is comparable with that found in differentlineages that used to be grouped in the nonmonophyletic sub-family Caesalpinioideae. A canalization of ontogenetic path-ways led to a more uniform unidirectional organ formationand may have been an important milestone in the formationof the successful flag blossom found in the majority of themore derived and species-rich papilionoid taxa.
Acknowledgments
This work was supported by São Paulo Research Founda-tion (FAPESP; process no. 2012/15644-8), Conselho Nacionalde Desenvolvimento Científico e Tecnológico (CNPq; processnos. 309987/2012-1 and 302204/2012-1), and Coordenaçãode Aperfeiçoamento de Pessoal de Nível Superior (CAPES).We are grateful to E. Seleme and N. L. Nunes for technicalassistance. Gwilym Lewis (RBG, Kew) kindly provided thecolor image of Amburana cearensis in figure 1. Paula Rudall(RBG, Kew) read through the final version of the manuscriptand provided valuable input. We thank Domingos Cardosoand an anonymous reviewer for their helpful comments andconstructive criticism.
Table 2
Abortion or Absence of Petals in Species of ADA and Swartzioid Clades
Tribe/species or genera No. petals initiatedNo. petals in themature flower Petal abortion or absence Reference
Angylocalyceae:Alexa sp. 5 5 . . . Polhill 1981Angylocalyx sp. 5 5 . . . Polhill 1981Castanospermum australe 5 5 . . . Tucker 1993Uleanthus sp. 5 5 . . . Polhill 1981Xanthocercis sp. 5 5 . . . Polhill 1981
Dipterygeae:Dipteryx alata 5 5 . . . Leite et al. 2014Pterodon pubescens 5 5 . . . Leite et al. 2014Taralea oppositifolia 5 5 . . . Leite et al. 2014
Amburaneae:Amburana acreana ? 1 4 V. Leite, personal observationAmburana cearensis 5 1 4 (abortion) This studyCordyla sp. ? 0 ? Cowan 1981Dussia sp. 5 5 . . . Polhill 1981Mildbraediodendron sp. ? 0 ? Cowan 1981Myrocarpus sp. 5 5 . . . Polhill 1981Myrospermum sp. 5 5 . . . Polhill 1981Myroxylum balsamum 5 5 . . . Tucker 1993
Swartzieae:Ateleia herbertsmith 1 1 4 (absence) Tucker 1990Bobgunniamadagascariensis 1 1 4 (absence) Tucker 2003bSwartzia apetala 0 0 5 (absence) Tucker 2003bSwartzia aureosericea 1 1 4 (absence) Tucker 2003bSwartzia dipetala 1 or 2 1 or 2 4 or 3 (absence) Paulino et al. 2013Swartzia laurifolia 1 1 4 (absence) Tucker 2003bSwartzia sericea 0 or 1 0 or 1 4 or 5 (absence) Tucker 2003bSwartzia simplex 1 1 4 (absence) Tucker 2003b
Note. Tribes are designated according to Cardoso et al. (2012, 2013). An ellipsis indicates no occurrence.
104 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions

Literature Cited
Albuquerque UP, JM Monteiro, MA Ramos, ELCA Amorim 2007Medicinal and magic plants from a public market in northeasternBrazil. J Ethnopharmacol 110:76–91.
Arroyo MTK 1981 Breeding systems and pollination biology inLeguminosae. Pages 723–769 in RM Polhill, PH Raven, eds. Ad-vances in legume systematics. Pt 2. Royal Botanic Gardens, Kew.
Bruneau A, BB Klitgaard, G Prenner, M Fougére-Danezan, SCTucker 2014 Floral evolution in the Detarieae (Leguminosae):phylogenetic evidence for labile floral development in an early-diverging legume lineage. Int J Plant Sci 175:392–417.
Bruneau A, M Mercure, GP Lewis, PS Herendeen 2008 Phylogeneticpatterns and diversification in the caesalpinioid legumes. Botany86:697–718.
Cardoso D, RT Pennington, LP Queiroz, JS Boatwright, BE VanWyk, MF Wojciechowski, M Lavin 2013 Reconstructing thedeep-branching relationships of papilionoid legumes. S Afr J Bot89:58–75.
Cardoso D, LP Queiroz, RT Pennington, HC Lima, E Fonty, MFWojciechowski, M Lavin 2012 Revisiting the phylogeny of pa-pilionoid legumes: new insights from comprehensively sampledearly-branching lineages. Am J Bot 99:1–23.
Cowan RS 1981 Swartzieae DC (1825). Pages 209–212 in RMPolhill, PH Raven, eds. Advances in legume systematics. Pt 1.Royal Botanic Gardens, Kew.
Fougére-Danezan M, PS Herendeen, S Maumont, A Bruneau 2009Morphological evolution in the variable resin-producing Detarieae(Leguminosae): do morphological characters retain a phylogeneticsignal? Ann Bot 105:311–325.
Gemmeke V 1982 Entwicklungsgeschichtliche Untersuchungen andMimosaceen-Blüten. Bot Jahrb Syst 103:185–210.
Ireland HE 2005 Swartzieae. Pages 215–225 in G Lewis, B Schrire, BMackinder, M Lock, eds. Legumes of the world. Royal BotanicGardens, Kew.
——— 2007 Taxonomic changes in the South American genus Bocoa(Leguminosae-Swartzieae): reinstatement of the name Trischidiumand a synopsis of both genera. Kew Bull 62:333–349.
Johansen DA 1940 Plant microtechnique. McGraw-Hill, New York.Khodaverdi M, A Movafeghi, MR Dadpour, S Naghiloo, M Ranj-bar, G Prenner 2014 Comparative study of floral development inOnobrychis melanotricha,Hedysarum varium and Alhagi persarum(Leguminosae: Papilionoideae: Hedysareae). Flora 209:23–33.
Klitgaard BB 1999 Floral ontogeny in tribe Dalbergieae (Legumi-nosae: Papilionoideae):Dalbergia brasiliensis,Machaerium villosumsl., Platymiscium floribundum and Pterocarpus rotundifolius. PlantSyst Evol 219:1–25.
Leite VG, VFMansano, SP Teixeira 2014 Floral ontogeny in Diptery-geae (Fabaceae) reveals new insights into one of the earliest branch-ing tribes in papilionoid legumes. Bot J Linn Soc 174:529–550.
Legume Phylogeny Working Group (LPWG) 2013 Legume phylog-eny and classification in the 21st century: progress, prospects andlessons for other species-rich clades. Taxon 62:217–248.
Lewis G, B Schrire, B Mackinder, M Lock 2005 Legumes of theworld. Royal Botanic Gardens, Kew.
Mansano VF, SC Tucker, AMGA Tozzi 2002 Floral ontogeny ofLecointea, Zollernia, Exostiles and Harleyodendron (Leguminosae:Papilionoideae: Swartzieae sl.). Am J Bot 89:1553–1569.
McMahon MM, L Hufford 2005 Evolution and development inthe amorphoid clade (Amorpheae: Papilionoideae: Leguminosae):petal loss and dedifferentiation. Int J Plant Sci 166:383–396.
Moço MCC, JEA Mariath 2009 Comparative floral ontogeny inAdesmia (Leguminosae: Papilionoideae: Dalbergieae). Aust J Bot57:65–75.
Movafeghi A, S Naghiloo, MR Dadpour 2011 Inflorescence andfloral development in Astragalus lagopoides Lam. (Leguminosae:Papilionoideae: Galegeae). Flora 206:219–226.
Naghiloo S, MR Dadpour, A Movafeghi 2012 Floral ontogeny inAstragalus compactus (Leguminosae: Papilionoideae: Galegeae):variable occurrence of bracteoles and variable patterns of sepalinitiation. Planta 235:793–805.
Paulino JV 2012 Ontogenia e morfologia de flores pluricarpeladasde espécies de Leguminosae. PhD diss. Universidade de Sao Paulo.
Paulino JV, VF Mansano, SP Teixeira 2013 Elucidating the unusualfloral features of Swartzia dipetala (Fabaceae). Bot J Linn Soc 173:303–320.
Polhill RM 1981 Papilionoideae. Pages 191–208 in RM Polhill, PHRaven, eds. Advances in legume systematics. Pt 1. Royal BotanicGardens, Kew.
Prenner G 2003a A developmental analysis of the inflorescence andthe flower of Lotus corniculatus (Fabaceae-Loteae). Mitt NaturwissVerein Steiermark 133:99–107.
——— 2003b Floral ontogeny in Lathyrus latifolius (Fabaceae-Vicieae). Phyton 43:392–400.
——— 2004a The asymmetric androecium in Papilionoideae (Legu-minosae): definition, occurrence, and possible systematic value. Int JPlant Sci 165:499–510.
——— 2004b Floral development in Daviesia cordata (Legumi-nosae: Papilionoideae: Mirbelieae) and its systematic implications.Aust J Bot 52:285–291.
——— 2004c New aspects in floral development of Papilionoideae:initiated but suppressed bracteoles and variable initiation of sepals.Ann Bot 93:537–545.
——— 2004d Floral ontogeny in Calliandra angustifolia (Legumi-nosae: Mimosoideae: Ingeae) and its systematic implications. Int JPlant Sci 165:417–426.
——— 2013a Flower development in Abrus precatorius (Legumi-nosae: Papilionoideae: Abreae) and a review of androecial char-acters in Papilionoideae. S Afr J Bot 89:210–218.
——— 2013b Papilionoid inflorescences revisited (Leguminosae:Papilionoideae). Ann Bot 112:1567–1576.
Prenner G, RM Bateman, PJ Rudall 2010 Floral formulae updatedfor routine inclusion in formal taxonomic descriptions. Taxon59:241–250.
Prenner G, BB Klitgaard 2008 Towards unlocking the deep nodes ofthe Leguminosae: floral development and morphology of the en-igmatic Duparquetia orchidacea (Leguminosae: Caesalpinioideae).Am J Bot 95:1349–1365.
Queiroz LP 2009 Leguminosas da caatinga. Universidade Estadualde Feira de Santana/Royal Botanic Gardens, Kew.
Spegazzini C 1921 Plantas nuevas o interesantes. An Soc Cient Ar-gent 91–92:77–123.
Tucker SC 1987 Floral initiation and development in legumes.Pages 183–239 in CH Stirton, ed. Advances in legume systematics.Pt 3. Royal Botanic Gardens, Kew.
——— 1988 Loss versus suppression of floral organs. Pages 69–82 inP Leins, SC Tucker, PK Endress, eds. Aspects of floral development.Springer, Berlin.
——— 1990 Loss of floral organs in Ateleia (Leguminosae: Papil-ionoideae). Am J Bot 77:750–761.
——— 1993 Floral ontogeny in Sophoreae (Leguminosae: Papil-ionoideae). I. Myroxylon (Myroxylon group) and Castanospermum(Angylocalyx group). Am J Bot 80:65–75.
——— 1996 Stamen structure and development in legumes, withemphasis on poricidal stamens of caesalpinioid tribe Cassieae.Pages 236–254 in WG D’Arcy, RC Keating, eds. Cambridge Uni-versity Press, Cambridge.
——— 1997 Floral evolution, development, and convergence: thehierarchical-significance hypothesis. Int J Plant Sci 156:143–161.
——— 1998 Floral ontogeny in legume genera Petalostylis, Labichea,and Dialium (Caesalpinioideae: Cassieae): a series in floral reduc-tion. Am J Bot 85:184–208.
LEITE ET AL.—FLORAL DEVELOPMENT IN AMBURANA (LEGUMINOSAE) 105
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions

——— 2002 Floral ontogeny in Sophoreae (Leguminosae: Papil-ionoideae). III. Radial symmetry and random petal aestivation inCadia purpurea. Am J Bot 89:748–757.
——— 2003a Comparative floral ontogeny in Detarieae (Legumi-nosae: Caesalpinioideae). III. Adaxially initiated whorls in Julber-nardia and Sindora. Int J Plant Sci 164:275–286.
——— 2003b Floral ontogeny in Swartzia (Leguminosae: Papil-ionoideae: Swartzieae): distribution and role of the ring meristem.Am J Bot 90:1271–1292.
——— 2006 Floral ontogeny of Hardenbergia violacea (Fabaceae:Faboideae: Phaseoleae) and taxa of tribes Bossiaeeae andMirbelieae,
with emphasis on presence of pseudoraceme inflorescences. Aust SystBot 19:193–210.
Vogel S 1997 Remarkable nectaries: structure, ecology, organo-phyletic perspectives. I. Substitutive nectaries. Flora 192:305–333.
Zhang Y, J Yang, G-Y Rao 2006 Comparative study on the aerialand subterranean flower development in Amphicarpaea edgeworthiiBenth. (Leguminosae: Papilionoideae), an amphicarpic species. Int JPlant Sci 167:943–949.
Zimmerman E, G Prenner, A Bruneau 2013 Floral ontogeny inDialiinae (Caesalpinioideae: Cassieae), a study in organ loss andinstability. S Afr J Bot 89:188–209.
106 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 213.48.60.9 on Wed, 21 Jan 2015 03:50:03 AMAll use subject to JSTOR Terms and Conditions
















![Foster 2008 Legume Sm Plot[1]](https://img.dokumen.tips/doc/110x75/6321245b80403fa2920c9a0b/foster-2008-legume-sm-plot1.jpg)












