Embed Size (px)
Citation preview

Available online at www.sciencedirect.com
www.elsevier.com/locate/ygcen
General and Comparative Endocrinology 155 (2008) 70–78
Short Communication
Expression of sex steroid hormone-related genesin the embryo of the leopard gecko
Daisuke Endo, Yoh-Ichiro Kanaho, Min Kyun Park *
Department of Biological Sciences, Graduate School of Science, The University of Tokyo, 7-3-1 Hongo, Bunkyo, Tokyo 113-0033, Japan
Received 28 November 2006; revised 4 April 2007; accepted 9 April 2007Available online 21 April 2007
Abstract
Sex steroid hormones are known to play a central role in vertebrate sex determination and differentiation. However, the tissues inwhich they are produced or received during development, especially around the period of sex determination of the gonads, have rarelybeen investigated. In this study, we identified the cDNA sequence, including the full-length of the coding region of cholesterol side-chaincleavage enzyme (P450scc), from the leopard gecko; a lizard with temperature-dependent sex determination. Embryonic expression anal-ysis of two steroidogenic enzymes, P450scc and P450 aromatase (P450arom), and four sex steroid hormone receptors, androgen receptor,estrogen receptor a and b, and progesterone receptor, was subsequently conducted. mRNA expression of both steroidogenic enzymeswas observed in the brain and gonads prior to the temperature-sensitive period of sex determination. The mRNAs of the four sex steroidhormone receptors were also detected in the brain and gonads at all stages examined. These results suggest the existence of a gonad-inde-pendent sex steroid hormone signaling system in the developing leopard gecko brain.� 2007 Elsevier Inc. All rights reserved.
Keywords: Reptile; Leopard gecko; Temperature-dependent sex determination; Sex steroid hormone; P450scc
1. Introduction
The first sex determination switch is extremely diverseamong vertebrates. The existence of Sry on the male-spe-cific sex chromosome creates the gonads of embryo testesin mammals (Sinclair et al., 1990; Koopman et al., 1991),while Dmy is the sex-determining gene in medaka, a bonyfish (Matsuda et al., 2002). Conversely, all crocodiles, mostturtles, and some lizards have no sex chromosomes and sexis determined by incubation temperature rather than by agene (Sarre et al., 2004). Sex steroid hormones, however,are known to play a pivotal role in sex determinationand differentiation in all vertebrate classes downstream ofthe first determining switch. The ovaries of adult estrogenreceptor a and b double knockout female mice exhibit fol-licle transdifferentiation to structures resembling seminifer-ous tubules of the testis with Sertoli cells and the expression
0016-6480/$ - see front matter � 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.ygcen.2007.04.010
* Corresponding author. Fax: +81 3 5841 4439.E-mail address: [email protected] (M.K. Park).
of male gonad-specific genes (Couse et al., 1999). Althoughgonadogenesis is essentially resistant to exogenous hor-mones in eutherian mammals, exogenous estrogen caninduce incomplete sex reversal in marsupials and birds.For example, the tammar wallaby, a marsupial species,developed ovary-like gonads when treated with estradiolat day 25 postpartum (Coveney et al., 2001). Similarly,male chick embryos treated with estradiol became femi-nized, although this was not permanent and syntheticinhibitors of the estrogen-synthesizing enzyme, P450 aro-matase (P450arom), could induce permanent female-to-male sex reversal (Smith and Sinclair, 2004). Estrogenor inhibitors of P450arom can complete sex reversal in rep-tiles (Bull et al., 1988; Tousignant and Crews, 1994),amphibians (Chardard and Dournon, 1999), and fish(Kobayashi et al., 2003). The effect of androgen treatmenton sex determination in reptilian species has also beenreported (Wibbels and Crews, 1995; Ganesh et al., 1999).Demonstration of female-specific expression of P450aromin developing gonads also suggests the importance of sex

D. Endo et al. / General and Comparative Endocrinology 155 (2008) 70–78 71
steroid hormones in the molecular mechanisms of sexdetermination or differentiation (Smith and Joss, 1994;Smith et al., 1997; Trant et al., 2001; Akazome et al., 2002).
Sex steroid hormones regulate sexually dimorphic traitsin other tissues as well as the gonads. Testosterone from thegonads gains access to the brain where it is aromatized toestradiol, which then masculinizes the mammalian brain(McCarthy and Konkle, 2005). In addition, hormonalmanipulation affects the formation of the brain structuresrelated to sexual behavior, and can alter traits such asthe lordosis of rodents and the song of songbirds (Cookeet al., 1998).
Recently, it was suggested that the brain could sexuallydifferentiate independently of gonadal steroids. The gynan-dromorphic zebra finch is genetically male in the right halfof the brain and genetically female in the left half. Further-more, the neural song circuit in the right brain has a moremasculine phenotype than that of the left (Agate et al.,2003). Since both halves of the brain are exposed to thesame circulating hormones, this phenomenon indicates thatthe genetic sex of the brain cells contributes to sexual differ-entiation. Moreover, both gonadal sex and incubation tem-perature affect male-typical sexual behavior in the leopardgecko (Eublepharis macularius), a lizard with temperature-dependent sex determination (Flores et al., 1994; Rhen andCrews, 1999). This may suggest that incubation tempera-ture directly affects brain organization of the sexual behav-ior of the leopard gecko.
Various studies have been conducted to investigate theexistence of brain autonomous sexual differentiation(Dewing et al., 2003; Scholz et al., 2006). Extragonadal ste-roidogenesis, for example of neurosteroids, is thought to beone process involved in the regulation of gonad-indepen-dent brain sexual differentiation. Neurosteroids are steroidssynthesized in the nervous system either de novo or by thein situ metabolism of precursors from other tissues. Theyare able to regulate not only adult brain functions but alsothe sexual differentiation of the brain independently fromgonads (Plassart-Schiess and Baulieu, 2001; Hollowayand Clayton, 2001). The expression of P450arom in thebrain has been reported in many species, including reptilesand birds (Willingham et al., 2000; Trant et al., 2001; Perl-man and Arnold, 2003). However, few studies have focusedon early development, or on other genes, such as the genefor cholesterol side-chain cleavage enzyme (P450scc),which catalyzes the first step of steroidogenesis and pro-duces pregnenolone, a major neurosteroid (Nelson et al.,1993).
In the present study, the leopard gecko was chosen as anappropriate experimental subject in order to elucidatewhether sex steroid hormones can be produced or receivedin extragonadal tissues during development. There are anumber of advantages in using this species. As previouslymentioned, it has been suggested that incubation tempera-ture has a direct effect on the leopard gecko brain (Floreset al., 1994; Rhen and Crews, 1999). Thus, it is interestingto analyze brain development in this species. Furthermore,
past research on reptiles often relied on wild animals andthis created problems in obtaining sufficient samples fromidentical conditions. However, the leopard gecko can beeasily maintained and sampled under controlled laboratoryconditions, and is also easy to breed. In addition, it reachessexual maturation within one year, a relatively short timecompared with other experimental reptiles, turtles, andcrocodiles, which further facilitates research into the devel-opment of the reproductive systems in this species. Anotheradvantage of using the leopard gecko is the availability ofmolecular information on sex-related genes in the species,such as those for sex steroid hormone receptors (Rhenand Crews, 2001), P450arom (Endo and Park, 2005),GnRH (Ikemoto and Park, 2003; Ikemoto et al., 2004),prolactin (Kato et al., 2005).
In the present study, we identified the cDNA sequenceof the leopard gecko, including the full-length of P450scccoding region. The expression of two steroidogenicenzymes, P450scc and P450arom, and four sex steroid hor-mone receptors, the androgen receptor, estrogen receptor aand b, and the progesterone receptor, were analyzed invarious embryonic tissues.
2. Materials and methods
2.1. Animals
All animals were treated according to the guidelines of the BiologicalScience Committee at the University of Tokyo. Adult fertile male andfemale leopard geckos (15 months of age) were used to identify the partialcDNA of P450scc and examined for tissue distribution of P450scc. Theywere hatched at 29 �C and raised at the same temperature in our labora-tory. Crickets were provided as the main diet three times per week andshelter, water, and a calcium supplement were supplied ad libitum. Ani-mals were maintained on a 14:10 h light/dark cycle at 29 �C. They wereanesthetized with sodium pentobarbital (50 mg/kg) and killed by rapiddecapitation, followed by complete bleeding. The following tissues werequickly removed, frozen immediately in liquid nitrogen and stored at�70 �C until required: from the female adult leopard gecko, whole brain,pituitary gland, ovary, oviduct, liver, kidney, heart, lung, skin, and adrenalgland; and from the male adult leopard gecko, whole brain, testis, vas def-erens, and kidney.
Fertilized eggs of the leopard gecko were acquired less than 24 h afteroviposition from the colony maintained in our laboratory and were incu-bated at 26, 32, and 34 �C. In the leopard gecko colony used in this study,sex of almost all embryos incubated at 26 and 34 �C was determined tofemale and more than 80% of the embryos at 32 �C were to male asdescribed previously (Viets et al., 1993). At 26 �C, six embryos at stage32 and 33, seven embryos at stage 34 and 35, three embryos at stage 36,37, and five embryos at stage 38 and 39 were collected from days 8 to19 of incubation, from 15 to 28, from 28 to 30, and from 30 to 39, respec-tively, according to the criteria of Dufaure and Hubert (1961). At 32 �C,four embryos at stage 32 and 33, six embryos at stage 34 and 35, threeembryos at stage 36, 37, and four embryos at stage 38 and 39 were col-lected from days 5 to 9 of incubation, from 12 to 20, at 20, and from 20to 23, respectively, and at 34 �C, five embryos at stage 32 and 33, sixembryos at stage 34 and 35, four embryos at stage 36, 37, and fourembryos at stage 38 and 39 were collected from days 6 to 8 of incubation,from 7 to 15, from 15 to 16, and from 18 to 19, respectively. Various tis-sues including brain and gonad–adrenal–mesonephric complex (GAM)were obtained from these embryos. All tissues were frozen immediatelyin liquid nitrogen and stored at �70 �C until required.

72 D. Endo et al. / General and Comparative Endocrinology 155 (2008) 70–78
2.2. RNA extraction and cDNA synthesis
Total RNA was extracted using ISOGEN (Nippon Gene, Tokyo,Japan). The cDNAs used as templates for RT-PCR were synthesized fromdenatured total RNA using 100 pmol oligo(dT) primer and 100 U ofM-MLV reverse transcriptase (Promega, Madison, WI) in a 20 ll reactionvolume with incubation at 42 �C for 1.5 h. After incubation, the reactionswere diluted to 25 ng/ll and used as templates. The adrenal gland cDNA,used for rapid amplification of cDNA ends (RACE; see Section 2.3), wassynthesized from 3 lg of total RNA using a SMART RACE cDNAAmplification Kit (BD Biosciences Clontech, Palo Alto, CA) accordingto the manufacturer’s instructions.
2.3. Molecular cloning of leopard gecko P450scc cDNA by
RT-PCR and RACE
RT-PCR was carried out to obtain partial leopard gecko P450scccDNA from adrenal gland cDNA using degenerate primers, lgscc-dSE01and lgscc-dAS01, deduced from conserved regions. All the followingPCR amplifications were performed in a 20 ll reaction mixture containingeach primer at 1 lM, 0.25 u of TaKaRa Ex Taq (TaKaRa, Shiga, Japan),each dNTP, and Ex Taq buffer (TaKaRa). The reaction conditions forRT-PCR were as follows: 94 �C for 5 min, 35 cycles of 94 �C for 40 s,58 �C for 30 s, 72 �C for 1 min, and 72 �C for 7 min. The amplified prod-ucts were separated by electrophoresis in 1.5% agarose gel and visualizedusing ethidium bromide staining. DNA fragments were extracted using aQIA Quick Gel Extraction Kit (Qiagen K.K., Tokyo, Japan) and directlysequenced using a dRhodamine terminator cycle sequencing FS ReadyReaction Kit (Applied Biosystems, Tokyo) by primers lgscc-dSE01 andlgscc-dAS01. After sequencing, sense and antisense gene-specific primers,lgscc-SE01, lgscc-SE02, and lgscc-AS01 were designed based on the
Table 1Oligonucleotide primers used for degenerate PCR, RT-PCR, RACE, and sequ
Gene Name Nucleotide sequencea
lgP450scc lgscc-dSE01 5 0-ATHTACAGGGAlgscc-dAS01 5 0-GGNTCWCGRCClgscc-SE01 5 0-GTGAAGCTTGGlgscc-SE02 5 0-GGGACACGCTTAlgscc-SE03 5 0-CAGAACTGCTGClgscc-SE04 5 0-TCAAGGAGACAlgscc-SE05 5 0-GAATTTATGCCAlgscc-SE06 5 0-CATCTTTTTCTGlgscc-SE07 5 0-CCTTGGCTAACClgscc-AS01 5 0-CTCAGCAACGGGlgscc-AS02 5 0-ATTCAGGAGAGlgscc-AS03 5 0-GCATGTAGACAClgscc-AS04 5 0-GTGTAAATTGGGlgscc-AS05 5 0-CAGTTTCTCCCTlgscc-AS06 5 0-GGGAATTCCTGG
lgP450arom lgarom-SE01 5 0- CAATTTTGAGAlgarom-AS01 5 0-GGATGGGATGA
lgERa lgERa-SE01 5 0-GATTCGGAAAGlgERa-AS01 5 0-TGGCTCGGCATC
lgERb lgERb-SE01 5 0-GTGGAACACAGlgERb-AS01 5 0-GGGCTTGTGCAG
lgAR lgAR-SE01 5 0-ATGAAGCAGGGlgAR-AS01 5 0-GCAGGTTACGG
lgPR lgPR-SE01 5 0-CTGGCATGGTAClgPR-AS01 5 0-GACTACACACAA
N represents all four nucleotides.a Abbreviations for degenerate nucleotides: Y, C or T; R, G or A; K, G or
sequences. Using these primers, the inner part of the cDNA fragmentsamplified by the primers, lgscc-dSE01 and lgscc-dAS01, were sequenced.After determining the partial sequence of P450scc cDNA, sense and anti-sense gene-specific primers were designed based on the sequence (Table 1).RACE was carried out to obtain the complete sequence. 3 0- and 5 0-RACEwas performed with SE03 and the Nested Universal Primer (NUP) of thekit (see Section 2.2), and NUP and AS02, respectively. Each PCR condi-tion was as follows: 94 �C for 5 min, 35 cycles of 94 �C for 40 s, 64 �C 30 s,72 �C for 2 min, and 72 �C 7 min. The amplified products were sequencedas described above.
2.4. Comparison of the amino acid sequences of various P450scc
The CLUSTAL X program (version 1.81) was downloaded from ftp://ftp-igbmc.u-strasbg.fr/pub/ClustalX/ and used with default settings toalign the deduced amino acid sequences of P450scc of the leopard geckoand other species in relation to each other (Thompson et al., 1997). Theamino acid identity was calculated using GeneDoc software (version2.6.002) (Nicholas and Nicholas, 1997). The GenBank accession numbersof P450scc used in the comparisons and phylogenic analysis are as follows:human NM000781; mouse NM019779; American alligator DQ007995;chicken NM0010017; zebra finch AY633556, and zebra fish NM152953.
2.5. Molecular phylogenic analysis
The amino acid sequences of the entire ORFs of P450scc in the leopardgecko and several species from all vertebrate classes were aligned usingCLUSTAL X with default settings. The alignment of the amino acidsequences was used to generate the phylogenic tree, using the neighbor-joining method (Saitou and Nei, 1987). Bootstrap values were calculatedwith 1000 replications to estimate the robustness of internal branches.
encing
Usage
GAARNTNGG-30 For degenerate PCRCATRGCRTA-30 For degenerate PCRCGCCATGACCGCCTG-30 For sequencing
GGGCTCCTGCAGG-3 0 For sequencing and RT-PCRAAAAGGTGATATG-3 0 For RACE and sequencing
ATAAGGCTCCACCC-30 For sequencingTGGGTCGGGACCC-3 0 For sequencing
ATCCATATGCTCG-3 0 For sequencingATGGGATTCAAG-3 0 For sequencingACCGATTTCAGCAC-30 For sequencing and RT-PCR
GCACAAATTTGTCC-3 0 For RACE and sequencingGCTTGACAAAGTC-3 0 For sequencingCCAAATTTCTGG-3 0 For sequencing
GTAAATTGGCCCA-3 0 For sequencingAGATTTGTAGG-30 For sequencing
AAAGTGTCCCCCAC -3 0 For RT-PCRCAGTTTTTCGTTGG -3 0 For RT-PCR
ACCGCAGAGGTGG-3 0 For RT-PCRTAGCAAGGCACTG-30 For RT-PCR
TCCAATCTATCCC-3 0 For RT-PCRTCACTGCCGCTG-3 0 For RT-PCR
ATGACCCTTGGAGC-3 0 For RT-PCRAATCCTGGTAAGGC-30 For RT-PCR
TTGGGGGTCGAAAG-30 For RT-PCRAGAAGCTGCCTCTC-3 0 For RT-PCR
T; H, A, T or C; W, A or T.
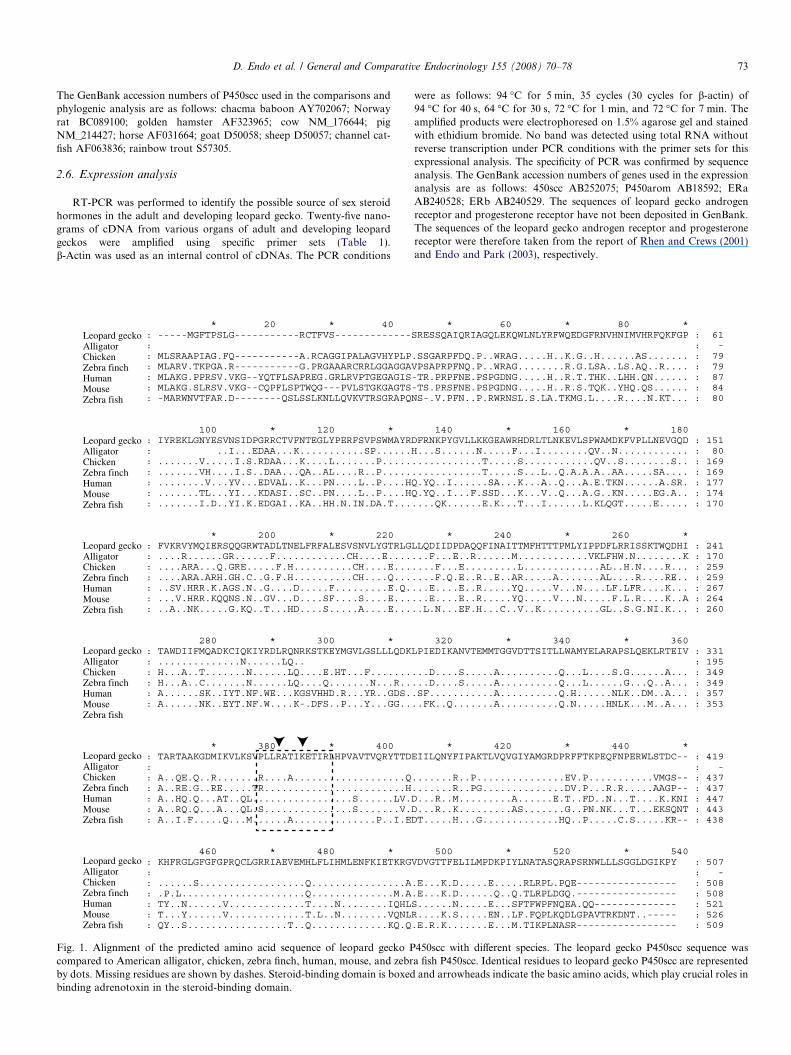
D. Endo et al. / General and Comparative Endocrinology 155 (2008) 70–78 73
The GenBank accession numbers of P450scc used in the comparisons andphylogenic analysis are as follows: chacma baboon AY702067; Norwayrat BC089100; golden hamster AF323965; cow NM_176644; pigNM_214427; horse AF031664; goat D50058; sheep D50057; channel cat-fish AF063836; rainbow trout S57305.
2.6. Expression analysis
RT-PCR was performed to identify the possible source of sex steroidhormones in the adult and developing leopard gecko. Twenty-five nano-grams of cDNA from various organs of adult and developing leopardgeckos were amplified using specific primer sets (Table 1).b-Actin was used as an internal control of cDNAs. The PCR conditions
:::::::
* 20 * 40-----MGFTPSLG-----------RCTFVS-------------
MLSRAAPIAG.FQ-----------A.RCAGGIPALAGVHYPLPMLARV.TKPGA.R-----------G.PRGAAARCRRLGGAGGAMLAKG.PPRSV.VKG--YQTFLSAPREG.GRLRVPTGEGAGISMLAKG.SLRSV.VKG--CQPFLSPTWQG---PVLSTGKGAGTS-MARWNVTFAR.D--------QSLSSLKNLLQVKVTRSGRAPQ
:::::::
100 * 120 *IYREKLGNYESVNSIDPGRRCTVFNTEGLYPERFSVPSWMAYR
..I...EDAA...K...........SP.............V.....I.S.RDAA...K....L.......P............VH....I.S..DAA...QA..AL....R..P.............V...YV...EDVAL..K...PN....L..P....H.......TL...YI...KDASI..SC..PN....L..P....H.......I.D..YI.K.EDGAI..KA..HH.N.IN.DA.T...
:::::::
* 200 * 220FVKRVYMQIERSQQGRWTADLTNELFRFALESVSNVLYGTRLG....R......GR......F............CH....E........ARA...Q.GRE.....F.H..........CH....E.......ARA.ARH.GH.C..G.F.H..........CH....Q.....SV.HRR.K.AGS.N..G....D.....F.........E.Q....V.HRR.KQQNS.N..GV...D....SF....S....E.....A..NK.....G.KQ..T...HD....S.....A....E...
::::::
280 * 300 *TAWDIIFMQADKCIQKIYRDLRQNRKSTKEYMGVLGSLLLQDK..............N......LQ..H...A..T.......N......LQ....E.HT...F.......H...A..C.......N......LQ....Q.......N...R..A......SK..IYT.NF.WE...KGSVHHD.R...YR..GDS.A......NK..EYT.NF.W....K-.DFS..P...Y...GG..
:::::::
* 380 * 400TARTAAKGDMIKVLKSVPLLRATIKETIRLHPVAVTVQRYTTD
A..QE.Q..R.......R....A...................QA..RE.G..RE.....TR........................HA..HQ.Q...AT..QL.................S......LV.A..RQ.Q...A...QL.S...............S.......V.A..I.F.....Q...M......A..............P..I.E
:::::::
460 * 480 *KHFRGLGFGFGPRQCLGRRIAEVEMHLFLIHMLENFKIETKRG
......S..................Q................A
.P.L.....................Q..............M.ATY..N......V.............T....N........IQHLT...Y......V.............T.L..N........VQNLQY..S.................T..Q.............KQ.Q
Leopard geckoAlligatorChickenZebra finchHumanMouseZebra fish
Leopard geckoAlligatorChickenZebra finchHumanMouseZebra fish
Leopard geckoAlligatorChickenZebra finchHumanMouseZebra fish
Leopard geckoAlligatorChickenZebra finchHumanMouseZebra fish
Leopard geckoAlligatorChickenZebra finchHumanMouseZebra fish
Leopard geckoAlligatorChickenZebra finchHumanMouseZebra fish
Fig. 1. Alignment of the predicted amino acid sequence of leopard gecko Pcompared to American alligator, chicken, zebra finch, human, mouse, and zebrby dots. Missing residues are shown by dashes. Steroid-binding domain is boxedbinding adrenotoxin in the steroid-binding domain.
were as follows: 94 �C for 5 min, 35 cycles (30 cycles for b-actin) of94 �C for 40 s, 64 �C for 30 s, 72 �C for 1 min, and 72 �C for 7 min. Theamplified products were electrophoresed on 1.5% agarose gel and stainedwith ethidium bromide. No band was detected using total RNA withoutreverse transcription under PCR conditions with the primer sets for thisexpressional analysis. The specificity of PCR was confirmed by sequenceanalysis. The GenBank accession numbers of genes used in the expressionanalysis are as follows: 450scc AB252075; P450arom AB18592; ERaAB240528; ERb AB240529. The sequences of leopard gecko androgenreceptor and progesterone receptor have not been deposited in GenBank.The sequences of the leopard gecko androgen receptor and progesteronereceptor were therefore taken from the report of Rhen and Crews (2001)and Endo and Park (2003), respectively.
* 60 * 80 *SRESSQAIQRIAGQLEKQWLNLYRFWQEDGFRNVHNIMVHRFQKFGP
.SSGARPFDQ.P..WRAG.....H..K.G..H......AS.......VPSAPRPFNQ.P..WRAG........R.G.LSA..LS.AQ..R....-TR.PRPFNE.PSPGDNG.....H..R.T.THK..LHH.QN......-TS.PRSFNE.PSPGDNG.....H..R.S.TQK..YHQ.QS......NS-.V.PFN..P.RWRNSL.S.LA.TKMG.L....R....N.KT...
: 61: -: 79: 79: 87: 84: 80
140 * 160 * 180DFRNKPYGVLLKKGEAWRHDRLTLNKEVLSPWAMDKFVPLLNEVGQDH...S......N.....F...I........QV..N........................T.....S............QV..S........S..............T.....S...L..Q.A.A.A..AA.....SA....Q.YQ..I......SA...K...A..Q...A.E.TKN......A.SR.Q.YQ..I...F.SSD...K...V..Q...A.G..KN.....EG.A......QK......E.K...T...I......L.KLQGT.....E.....
: 151: 80: 169: 169: 177: 174: 170
* 240 * 260 *LLQDIIDPDAQQFINAITTMFHTTTPMLYIPPDFLRRISSKTWQDHI...F...E..R......M............VKLFHW.N........K....F...E.........L.............AL..H.N....R.......F.Q.E..R..E..AR.....A.......AL....R....RE.....E....E..R.....YQ.....V...N....LF.LFR....K......E....E..R.....YQ.....V...N.....F.L.R....K..A..L.N...EF.H...C..V..K..........GL..S.G.NI.K...
: 241: 170: 259: 259: 267: 264: 260
320 * 340 * 360LPIEDIKANVTEMMTGGVDTTSITLLWAMYELARAPSLQEKLRTEIV
...D....S.....A..........Q...L....S.G......A...
...D....S.....A..........Q...L......G...Q..A...
.SF...........A..........Q.H......NLK..DM..A...
..FK..Q.......A..........Q.N.....HNLK...M..A...
: 331: 195: 349: 349: 357: 353
* 420 * 440 *EIILQNYFIPAKTLVQVGIYAMGRDPRFFTKPEQFNPERWLSTDC--
.......R..P...............EV.P...........VMGS--
.......R..PG..............DV.P...R.R.....AAGP--D...R..M.........A......E.T..FD..N...T....K.KNID...R..K.........AS.......G..PN.NK...T...EKSQNTDT.....H...G.............HQ..P.....C.S.....KR--
: 419: -: 437: 437: 447: 443: 438
500 * 520 * 540VDVGTTFELILMPDKPIYLNATASQRAPSRNWLLLSGGLDGIKPY
.E...K.D.....E.....RLRPL.PQE-----------------
.E...K.D......Q..Q.TLRPLDGQ.-----------------S......N.....E...SFTFWPFNQEA.QQ--------------R....K.S.....EN..LF.FQPLKQDLGPAVTRKDNT..-----.E.R.K.......E...M.TIKPLNASR-----------------
: 507: -: 508: 508: 521: 526: 509
450scc with different species. The leopard gecko P450scc sequence wasa fish P450scc. Identical residues to leopard gecko P450scc are representedand arrowheads indicate the basic amino acids, which play crucial roles in

74 D. Endo et al. / General and Comparative Endocrinology 155 (2008) 70–78
3. Results
3.1. Cloning of the partial sequence of leopard gecko
P450scc cDNA and comparison with other vertebrate
P450scc
We designed degenerate primers based on the sequenceof the highly conserved region. Using these primers,711 bp of the partial sequence was acquired. Based on thispartial sequence, gene-specific primers for 3 0- and 5 0-RACEwere designed. The bands acquired by RACE weresequenced directly using the primers SE03, SE04, SE05,SE06, SE07, AS02, AS03, AS04, AS05, and AS06. Thelength of the leopard gecko P450 aromatase cDNA identi-fied in this study was 1645 bp, excluding the poly(A) tail,with an open reading frame encoding a 507 amino acidprotein. As shown in Fig. 1, the deduced amino acidsequence of the leopard gecko P450scc showed moderate
whole brain (fem
ale)
whole brain (m
ale)pituitary glandovaryoviducttestisvas deferes
β-actin-
P450scc-
Fig. 3. Expression analysis of P450scc in various tissues of the adult leopardgland, ovary, oviduct, testis, vas deferens, hemipenis, liver, kidney, heart, lung,were subjected to PCR for the leopard gecko P450scc. b-Actin was also analyzefrom both males and females: testis and vas deferens were from males and ot
0.1
baboonhuman
1000
horsepig
cowsheepgoat
952
1000
1000948
560
hamster
rat
mouse 8211000
zebra fish
troutcatfish
1000
1000
Leopard gecko
zebra finch
chicken
alligator
777940
917
1000
Fig. 2. Phylogenic tree of vertebrate P450scc, based on the amino acidhomology. Numbers at the branches represent the confidence limitscomputed by the bootstrap procedure (1000 trials). For details onaccession numbers in the GenBank databases, see Section 2.
identity with that of other vertebrates: 61% with chickens,57% with zebra finch, 46% with humans, 45% with mice,and 51% with zebra fish. The predicted protein sequencecontains two basic amino acids that play crucial roles inbinding adrenotoxin in the steroid-binding domain (Wadaand Waterman, 1992). In the steroid-binding domain,almost all amino acids were conserved (10/12 with chick-ens, 12/12 with humans, 11/12 with zebra fish). Thesequence of the cDNA clone was deposited in GenBank(Accession No. AB252075).
3.2. Molecular phylogenic analysis
The phylogenic tree was calculated using an amino acidalignment of the P450scc of 17 selected species. The resultis shown in Fig. 2. The P450scc of reptiles (leopard geckoand American alligator) and birds (chicken and zebrafinch) were clustered together, and that of the leopardgecko was first to branch off from the cluster.
3.3. Expression analysis of P450scc in the adult leopard
gecko
The tissue distribution of P450scc was investigated byRT-PCR. The genomic structure of P450scc has not yetbeen characterized in the leopard gecko, but there areintrons in the homologous region of the target of the pri-mer set in the genome of humans (Morohashi et al.,1987). No band was detected using a primer set with thegenomic DNA of the leopard gecko as a template (datanot shown). Thus, the effects of genomic DNA contamina-tion of cDNA could be excluded. As shown in Fig. 3,strong signals of P450scc were detected in the pituitarygland, ovary, testis, liver, kidney, heart, and adrenal gland,and moderate signals were detected in the whole brain, ovi-duct, vas deferens hemipenis, and skin.
3.4. Expression of two steroidogenic enzymes in embryos of
the leopard gecko
Tissue distributions of P450scc and P450arom were exam-ined to determine steroid hormone-producing tissues in theleopard gecko embryos from stages 32 to 39. The tempera-
kidney (female)
kidney (male)
liver
heart
skin
adrenal glandR
T(-)
hemipenis
lung
gecko. Twenty-five nanograms of cDNA from the whole brain, pituitaryskin, adrenal gland, and total RNA without reverse transcription (RT(�))d as quality control of cDNAs. The whole brain and kidney were obtainedher tissues were removed from females.

P450scc-
P450arom-
β-actin-
P450scc-
P450arom-
β-actin-
P450scc-
P450arom-
β-actin-
P450scc-
P450arom-
β-actin-
whole
brain
GA
M
olfactory
bulb
heart
liver
tail
whole
brain
GA
M
olfactory
bulb
heartliver
tail
lung
hemipenis
whole
brain
GA
M
olfactory
bulb
heartliver
tail
lung
hemipenis
kidney
whole
brain
GA
M
olfactory
bulb
heartliver
tail
lunghem
ipeniskidney
fat body
26
P450scc P450arom β-actin
32-33
34-35
36-37
38-39
P450scc P450arom β-actin
(stage) 32-33
34-35
36-37
38-39
(stage)32 34 26 32 34 26 32 34 26 32 34 26 32 34 26 32 34
Fig. 4. Expression analysis of two steroidogenic enzymes in various tissues of leopard gecko embryos. Twenty-five nanograms of cDNA from (a) thewhole brain, olfactory bulb, GAM, heart, liver, and tail of the embryo at stage 33; (b) whole brain, olfactory bulb, GAM, heart, liver, lung, hemipenis, andtail of the embryo at stage 34; (c) whole brain, olfactory bulb, GAM, heart, liver, lung, hemipenis, kidney, tail, and skin of the embryo at stage 37; and (d)whole brain, olfactory bulb, GAM, heart, liver, lung, hemipenis, kidney, tail, body fat, and skin of the embryo at stage 39 were subjected to PCR forleopard gecko P450scc and P450arom. b-Actin was also analyzed as quality control of cDNAs. These are results from representative individuals incubatedat 32 �C. The expression analysis of P450scc, P450arom, and b-actin in the (e) GAM and (f) brain from each stage of representative embryos incubated attwo female-producing temperature (26,34 �C) and male-biased temperature (32 �C) are shown. Sample numbers for this analysis were shown in Section 2.
D. Endo et al. / General and Comparative Endocrinology 155 (2008) 70–78 75
ture-sensitive period of the leopard gecko has not been deter-mined accurately; at stage 33 the sex of the embryo is indeter-minate, but it is thought to be determined at stage 38 (Bull,1987). mRNA expression of P450scc and P450arom wasobserved in the brain and GAM prior to the temperature-sensitive period at stage 33 (Fig. 4a). Expression in the brainand GAM was also observed at stages 34, 37, and 39(Fig. 4b–d). Furthermore, mRNA expression of theseenzymes was observed at stage 34 in other tissues, such asthe lung and tail. P450arom was observed in the olfactorybulbs at all stages; however, only the brain and GAMexpressed both enzymes at all stages examined. As shownin Fig. 4e and f, the expression of both enzymes was observedin the brain and GAM of embryos incubated at 26, 32, and34 �C. Additionally, while all GAMs studied expressedP450scc, some did not express P450arom mRNA (stage 32incubated at 26 �C and stage 37 incubated at 34 �C).
3.5. Expression of four sex steroid hormone receptors in
leopard gecko embryos
Sex steroid hormone receptor mRNAs showed a widerdistribution compared with steroidogenic enzymes
(Fig. 5a–d). The expression of estrogen receptor a wasdetected in all tissues examined from stage 33, whereasthe expression of estrogen receptor b and androgen recep-tor was not observed in all tissues. The expression of foursex steroid hormone receptors was detected in the brainat all stages and all incubation temperatures (Fig. 5f).Expression was also detected in the GAM; however, pro-gesterone receptor signals were weak or undetected untilstage 35 (Fig. 5e).
4. Discussion
It has recently been suggested that a gonad-indepen-dent sex differentiation mechanism exists in the brainof reptiles and birds (Agate et al., 2003; Flores et al.,1994; Rhen and Crews, 1999). Non-gonadal steroid hor-mones, especially neurosteroids, may regulate gonad-independent sex differentiation of the brain (Plassart-Schiess and Baulieu, 2001; Holloway and Clayton,2001). To examine the possibility of the formation of anon-gonadal steroid hormone signaling system aroundthe period of gonadal sex determination, we analyzedthe expression of genes related to sex steroid hormones

AR-
ERa-
ERb-
PR-
AR-
ERa-
ERb-
PR-
AR-
ERa-
ERb-
PR-
AR-
ERa-
ERb-
PR-
whole
brain
GA
M
olfactory
bulb
heart
liver
tail
whole
brain
GA
M
olfactory
bulb
heartliver
tail
lung
hemipenis
whole
brain
GA
M
olfactory
bulb
heartliver
tail
lung
hemipenis
kidney
whole
brain
GA
M
olfactory
bulb
heartliver
tail
lunghem
ipeniskidney
fat body
32-33
34-35
36-37
38-39
(stage)
AR ERa ERb PR AR ERa ERb PR
32-33
34-35
36-37
38-39
(stage)26 32 34 26 32 34 26 32 34 26 32 34 26 32 34 26 32 34 26 32 34 26 32 34
Fig. 5. Expression analysis of four sex steroid hormone receptors in various tissues of leopard gecko embryos. Twenty-five nanograms of cDNAs from(a) the whole brain, olfactory bulb, GAM, heart, liver, and tail of the embryo at stage 33; (b) whole brain, olfactory bulb, GAM, heart, liver, lung,hemipenis, and tail of the embryo at stage 34; (c) whole brain, olfactory bulb, GAM, heart, liver, lung, hemipenis, kidney, tail, and skin of the embryo atstage 37; and (d) whole brain, olfactory bulb, GAM, heart, liver, lung, hemipenis, kidney, tail, body fat, and skin of the embryo at stage 39 were subjectedto PCR for leopard gecko estrogen receptor a (ERa) and b (ERb), androgen receptor (AR), and progesterone receptor (PR). Representative results of theexpression analysis of the four sex steroid hormone receptors in the GAM (e) and brain (f) from each stage of embryos incubated at two female-producingtemperature (26,34 �C) and male-biased temperature (32 �C) are shown. Sample numbers for this analysis were shown in Section 2. All samples in thisfigure were from same individuals with Fig. 4.
76 D. Endo et al. / General and Comparative Endocrinology 155 (2008) 70–78
in various tissues obtained from leopard gecko embryos.The leopard gecko is a good experimental animal tostudy the molecular mechanisms of reptilian brain sexdifferentiation because of its availability in the labora-tory, early attainment of sexual maturation, and the dif-ference in sexual behavior patterns between female adultsexposed to different incubation temperatures duringdevelopment (Flores et al., 1994; Rhen and Crews, 1999).
Initially, 1645 bp of the partial cDNA sequence wasidentified, including the full-length of the coding regionfor the leopard gecko P450scc. Almost all amino acids inthe steroid-binding domain of P450scc are well conservedamong vertebrates, including the leopard gecko, and twobasic residues in the domain were present in all vertebratespecies examined (Fig. 1) (Wada and Waterman, 1992).Thus, the cDNA sequence acquired in this study is thoughtto be part of the cDNA of functional leopard geckoP450scc. This is the first identification of the cDNAsequence, including the full-length of coding region forP450scc, in reptiles.
In the phylogenic tree drawn using the amino acidsequences of P450scc of species from all vertebrate phylum,
the P450scc amino acid sequences of reptiles and birds wereclustered together and those of the leopard gecko branchedoff first (Fig. 2). It is well known that crocodilians are moreclosely related to birds than to Squamates. Thus, in a com-parative study of vertebrates, Squamates should be treatedas an independent group of amniotes, similar to mammali-ans and birds, and not as a group of reptiles.
In the adult leopard gecko, mRNA expression ofP450scc was observed in various tissues, including thebrain, by RT-PCR (Fig. 3). P450scc synthesizes pregneno-lone, which is the first step of steroid hormone biosynthe-sis, and pregnenolone itself is thought to serve as aneurosteroid (Nelson et al., 1993). The expression ofP450scc in the brain suggests the capacity for steroid hor-mone synthesis in the leopard gecko, as well as in the rat,zebra finch, and other species (Le Goascogne et al., 1987;Tsutsui et al., 1999). In most species, the gonads and adre-nal gland are the main steroidogenic organs and strongexpression was reported in these organs (Miller, 1988).The expression of P450scc has been reported in other tis-sues, such as the brain, skin, lung, and epididymis (LeGoascogne et al., 1987; Slominski et al., 1996; Vilchis

D. Endo et al. / General and Comparative Endocrinology 155 (2008) 70–78 77
et al., 2002). However, there is some difference in theexpression pattern between species. For example, strongexpression was observed in the liver and kidney of the leop-ard gecko, but not in the hamster (Vilchis et al., 2002).Thus, steroidogenesis outside the gonads and adrenal glandmay reflect a diverse role in different species.
The expression of P450scc and P450arom was examinedin leopard gecko embryos by RT-PCR. It was found thatexpression of P450scc was restricted in the brain, andembryonic GAM differed from adult tissues (Figs. 3 and4). Moreover, strong expression of P450scc was observedin the liver and heart of the adult leopard gecko, but notin the embryo. Embryonic expression of P450arom wasobserved in various tissues, including the brain, GAM,olfactory bulb, lung, and tail. However, constant expres-sion of both steroidogenic enzymes at each of the stagesexamined was only found in the brain and GAM(Fig. 4a–d). Thus, it cannot be decided in which tissuesthe mRNA of these steroidogenic enzymes was expressed.Expression of both enzymes was also observed in allembryonic brain incubated at 26, 32, and 34 �C (Fig. 4f).P450scc was also detected in all GAMs studied during allstages of development, regardless of the incubation temper-ature. These results suggest that the brain and GAM arethe main areas of sex steroid biosynthesis during develop-ment of the leopard gecko. Though expression ofP450arom was observed in all brain tissue examined, itwas barely detected in some GAMs, even at the female-producing incubation temperature (26 and 34 �C) beforestage 37 (Fig. 4e). The expression of P450arom in the allbrain examined from stage 32, prior to gonadal sex deter-mination, may indicate an important role for the brain-derived steroid hormone, estrogen, in brain sexualdifferentiation.
Unlike steroidogenic enzymes, sex steroid hormonereceptors were expressed in a variety of tissues. The mRNAof estrogen receptor a was expressed in all tissues at all stagesexamined (Fig. 5). Androgen receptor, estrogen receptor b,and progesterone receptor mRNAs were expressed in vari-ous, but not all tissues. In almost all the brains and GAMsstudied, the four receptors were expressed at all stages, inde-pendent of incubation temperature. It is notable that all foursex steroid hormone receptors, as well as the two steroido-genic enzymes, P450scc and P450arom, were expressed inthe brain before the sex-determining period. This suggeststhat the steroid hormones synthesized in the brain candirectly affect its differentiation. In the leopard gecko, itwas suggested that the incubation temperature indepen-dently affects sexual behavior as well as gonadal sex (Floreset al., 1994; Rhen and Crews, 1999). The expression level ofestrogen synthetase, P450arom, is stimulated in the gonad atthe female-producing temperature and promotes female sexdifferentiation in crocodilians (Smith and Joss, 1994; Gabrielet al., 2001). As in the gonads, the expression of sex steroidhormone-related genes in the brain may also be affecteddirectly by incubation temperature and thus contribute tobrain differentiation.
In the present study, we cloned the cDNA for leop-ard gecko P450scc and analyzed the expression of twosteroidogenic enzymes, P450scc and P450arom, and foursex steroid hormone-receptors in the developing brainsof the leopard gecko prior to gonadal sex differentia-tion. Our results strongly suggest that the embryonicbrain is able to synthesize and respond to steroid hor-mones. Although past research of the molecular mecha-nisms of temperature-dependent sex determination hasfocused only on the gonad, future research should alsoinvestigate the effect of incubation temperature on thebrain sex steroid hormone signaling system to investi-gate the possibility of the brain autonomous sexdifferentiation.
Acknowledgments
We are grateful to Prof. Y. Oka, Ms. M. Kyokuwa,Dr. M. Enomoto, Mr. K. Kato, Ms. M. Utsumi, Ms. T.Kato, Ms. E. Kurakata, and Dr. T. Ikemoto, Departmentof Biological Sciences, Graduate School of Science, TheUniversity of Tokyo, for valuable discussion throughoutthis study. This work was supported by Grants-in-Aidfrom the Japan Society for the Promotion of Science.
References
Agate, R.J., Grisham, W., Wade, J., Mann, S., Wingfield, J., Schanen, C.,Palotie, A., Arnold, A.P., 2003. Neural, not gonadal, origin of brainsex differences in a gynandromorphic finch. Proc. Natl. Acad. Sci.USA 100, 4873–4878.
Akazome, Y., Abe, T., Mori, T., 2002. Differentiation of chicken gonadas an endocrine organ: expression of LH receptor, FSH receptor,cytochrome P450c17 and aromatase genes. Reproduction 123, 721–728.
Bull, J.J., 1987. Temperature-sensitive periods of sex determination in alizard: similarities with turtles and crocodilians. J. Exp. Zool. 241, 143–148.
Bull, J.J., Gutzke, W.H., Crews, D., 1988. Sex reversal by estradiol inthree reptilian orders. Gen. Comp. Endocrinol. 70, 425–428.
Chardard, D., Dournon, C., 1999. Sex reversal by aromataseinhibitor treatment in the newt Pleurodeles waltl. J. Exp. Zool.283, 43–50.
Cooke, B., Hegstrom, C.D., Villeneuve, L.S., Breedlove, S.M., 1998.Sexual differentiation of the vertebrate brain: principles and mecha-nisms. Front. Neuroendocrinol. 19, 323–362.
Couse, J.F., Hewitt, S.C., Bunch, D.O., Sar, M., Walker, V.R., Davis,B.J., Korach, K.S., 1999. Postnatal sex reversal of the ovaries inmice lacking estrogen receptors alpha and beta. Science 286, 2328–2331.
Coveney, D., Shaw, G., Renfree, M.B., 2001. Estrogen-induced gonadalsex reversal in the tammar wallaby. Biol. Reprod. 65, 613–621.
Dewing, P., Shi, T., Horvath, S., Vilain, E., 2003. Sexually dimorphic geneexpression in mouse brain precedes gonadal differentiation. Brain Res.Mol. Brain Res. 118, 82–90.
Dufaure, J.P., Hubert, J., 1961. Table de developement du lezard vivipare:Lacerta (Zootica) vivipara Jacquin. Arch. Anat. Microsc. Morphol.Exp. 50, 309–328.
Endo, D., Park, M.K., 2003. Quantification of three steroid hormonereceptors of the leopard gecko (Eublepharis macularius), a lizard withtemperature-dependent sex determination: their tissue distributionsand the effect of environmental change on their expressions. Comp.Biochem. Physiol. B. Biochem. Mol. Biol. 136, 957–966.

78 D. Endo et al. / General and Comparative Endocrinology 155 (2008) 70–78
Endo, D., Park, M.K., 2005. Molecular cloning of P450 aromatase fromthe leopard gecko and its expression in the ovary. J. Steroid Biochem.Mol. Biol. 96, 131–140.
Flores, D., Tousignant, A., Crews, D., 1994. Incubation temperatureaffects the behavior of adult leopard geckos (Eublepharis macularius).Physiol. Behav. 55, 1067–1072.
Gabriel, W.N., Blumberg, B., Sutton, S., Place, A.R., Lance, V.A., 2001.Alligator aromatase cDNA sequence and its expression in embryos atmale and female incubation temperatures. J. Exp. Zool. 290, 439–448.
Ganesh, S., Choudhary, B., Raman, R., 1999. Temporal differencebetween testis and ovary determinations with possible involvementof testosterone and aromatase in gonadal differentiation in TSDlacking lizard, Calotes versicolor. J. Exp. Zool. 283, 600–607.
Holloway, C.C., Clayton, D.F., 2001. Estrogen synthesis in the male braintriggers development of the avian song control pathway in vitro. Nat.Neurosci. 4, 170–175.
Ikemoto, T., Park, M.K., 2003. Identification and characterization of thereptilian GnRH-II gene in the leopard gecko, Eublepharis macularius,and its evolutionary considerations. Gene 316, 157–165.
Ikemoto, T., Enomoto, M., Park, M.K., 2004. Identification andcharacterization of a reptilian GnRH receptor from the leopardgecko. Mol. Cell. Endocrinol. 214, 137–147.
Kato, K., Ikemoto, T., Park, M.K., 2005. Identification of the reptilianprolactin and its receptor cDNAs in the leopard gecko, Eublepharis
macularius. Gene 346, 267–276.Kobayashi, T., Kajiura-Kobayashi, H., Nagahama, Y., 2003. Induction of
XY sex reversal by estrogen involves altered gene expression in ateleost, tilapia. Cytogenet. Genome Res. 101, 289–294.
Koopman, P., Gubbay, J., Vivian, N., Goodfellow, P., Lovell-Badge, R.,1991. Male development of chromosomally female mice transgenic forSry. Nature 351, 117–121.
Le Goascogne, C., Robel, P., Gouezou, M., Sananes, N., Baulieu, E.E.,Waterman, M., 1987. Neurosteroids: cytochrome P-450scc in rat brain.Science 23, 1212–1215.
Matsuda, M., Nagahama, Y., Shinomiya, A., Sato, T., Matsuda, C.,Kobayashi, T., Morrey, C.E., Shibata, N., Asakawa, S., Shimizu, N.,Hori, H., Hamaguchi, S., Sakaizumi, M., 2002. DMY is a Y-specificDM-domain gene required for male development in the medaka fish.Nature 417, 559–563.
McCarthy, M.M., Konkle, A.T., 2005. When is a sex difference not a sexdifference? Front. Neuroendocrinol. 26, 85–102.
Miller, W.L., 1988. Molecular biology of steroid hormone synthesis.Endocr. Rev. 9, 295–318.
Morohashi, K., Sogawa, K., Omura, T., Fujii-Kuriyama, Y., 1987. Genestructure of human cytochrome P-450(SCC), cholesterol desmolase. J.Biochem. (Tokyo) 101, 879–887.
Nelson, D.R., Kamataki, T., Waxman, D.J., Guengerich, F.P., Esta-brook, R.W., Feyereisen, R., Gonzalez, F.J., Coon, M.J., Gunsalus,I.C., Gotoh, O., et al., 1993. The P450 superfamily: update on newsequences, gene mapping, accession numbers, early trivial names ofenzymes, and nomenclature. DNA Cell Biol. 12, 1–51.
Nicholas, K.B., Nicholas Jr., H.B., 1997. Genedoc: a tool for editingand annotating multiple sequence alignments. Distributed by theauthor.
Perlman, W.R., Arnold, A.P., 2003. Expression of estrogen receptor andaromatase mRNAs in embryonic and posthatch zebra finch brain. J.Neurobiol. 55, 204–219.
Plassart-Schiess, E., Baulieu, E.E., 2001. Neurosteroids: recent findings.Brain Res. Brain Res. Rev. 37, 133–140.
Rhen, T., Crews, D., 1999. Embryonic temperature and gonadal sexorganize male-typical sexual and aggressive behavior in a lizard withtemperature-dependent sex determination. Endocrinology 140, 4501–4508.
Rhen, T., Crews, D., 2001. Distribution of androgen and estrogen receptormRNA in the brain and reproductive tissues of the leopard gecko,Eublepharis macularius. J. Comp. Neurol. 437, 385–397.
Saitou, N., Nei, M., 1987. The neighbor-joining method: a new method forreconstructing phylogenic trees. Mol. Biol. Evol. 4, 406–425.
Sarre, S.D., Georges, A., Quinn, A., 2004. The ends of a continuum:genetic and temperature-dependent sex determination in reptiles.Bioessays 26, 639–645.
Scholz, B., Kultima, K., Mattsson, A., Axelsson, J., Brunstrom, B.,Halldin, K., Stigson, M., Dencker, L., 2006. Sex-dependent geneexpression in early brain development of chicken embryos. BMCNeurosci. 7, 12.
Sinclair, A.H., Berta, P., Palmer, M.S., Hawkins, J.R., Griffiths, B.L.,Smith, M.J., Foster, J.W., Frischauf, A.M., Lovell-Badge, R., Good-fellow, P.N., 1990. A gene from the human sex-determining regionencodes a protein with homology to a conserved DNA-binding motif.Nature 346, 240–244.
Slominski, A., Ermak, G., Mihm, M., 1996. ACTH receptor, CYP11A1,CYP17 and CYP21A2 genes are expressed in skin. J. Clin. Endocrinol.Metab. 81, 2746–2749.
Smith, C.A., Joss, J.M., 1994. Steroidogenic enzyme activity ovariandifferentiation in the saltwater crocodile, Crocodylus porosus. Gen.Comp. Endocrinol. 93, 232–245.
Smith, C.A., Andrews, J.E., Sinclair, A.H., 1997. Gonadal sexdifferentiation in chicken embryos: expression of estrogen recep-tor and aromatase genes. J. Steroid Biochem. Mol. Biol. 60,295–302.
Smith, C.A., Sinclair, A.H., 2004. Sex determination: insights from thechicken. Bioessays 26, 120–132.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmouginm, F., Higgins,D.G., 1997. The CLUSTAL_X windows interface: flexible strategiesfor multiple sequence alignment aided by quality analysis tools.Nucleic Acids Res. 25, 4876–4882.
Tousignant, A., Crews, D., 1994. Effect of exogenous estradiol applied atdifferent embryonic stages on sex determination, growth, and mortalityin the leopard gecko (Eublepharis macularius). J. Exp. Zool. 268, 17–21.
Trant, J.M., Gavasso, S., Ackers, J., Chung, B.C., Place, A.R., 2001.Developmental expression of cytochrome P450 aromatase genes(CYP19a and CYP19b) in zebrafish fry (Danio rerio). J. Exp. Zool.290, 475–483.
Tsutsui, K., Ukena, K., Takase, M., Kohchi, C., Lea, R.W., 1999.Neurosteroid biosynthesis in vertebrate brains. Comp. Biochem.Physiol. C. Pharmacol. Toxicol. Endocrinol. 124, 121–129.
Viets, B.E., Tousignant, A., Ewert, M.A., Nelson, C.E., Crews, D., 1993.Temperature-dependent sex determination in the leopard gecko,Eublepharis macularius. J. Exp. Zool. 265, 679–683.
Vilchis, F., Chavez, B., Larrea, F., Timossi, C., Montiel, F., 2002. ThecDNA cloning and tissue expression of the cytochrome P450scc fromSyrian hamster (Mesocricetus auratus). Gen. Comp. Endocrinol. 126,279–286.
Wada, A., Waterman, M.R., 1992. Identification by site-directed muta-genesis of two lysine residues in cholesterol side chain cleavagecytochrome P450 that are essential for adrenodoxin binding. J. Biol.Chem. 267, 22877–22882.
Wibbels, T., Crews, D., 1995. Steroid-induced sex determination atincubation temperatures producing mixed sex ratios in a turtle withTSD. Gen. Comp. Endocrinol. 100, 53–60.
Willingham, E., Baldwin, R., Skipper, J.K., Crews, D., 2000. Aroma-tase activity during embryogenesis in the brain and adrenal-kidney-gonad of the red-eared slider turtle, a species with temperature-dependent sex determination. Gen. Comp. Endocrinol. 119, 202–207.





























