Embed Size (px)
Citation preview

RESEARCH ARTICLE
Endotoxin-induced silencing of mesoderm induction and functional differentiation: role of HMGB1 in pluripotency and infection
Kavitha Sivasubramaniyan1, Rajesh Reddy Atluri1, Kanchan Sarda1, Milan Arvind1, Vishnu Balaji1 & Kaushik Dilip Deb1††Author for correspondence1Manipal University, Embryonic Stem Cell and Developmental Biology Program, Manipal Institute of Regenerative Medicine, #10 Service Road, Domlur Layout, Bangalore 560071, IndiaTel.:+1 91 80 2535 6660;Fax: +1 91 80 2535 6662;E-mail: [email protected]
A
part of
Keywords: embryoid bodies, human embryonic stem, lipopolysaccharides, mesoderm
10.2217/17460751.3.1.xxx © 2
Objectives: Mechanisms underpinning Gram-negative bacterial vaginosis-induced birth anomalies are obscure. Ethical issues limit such studies on peri-implantation-stage human embryos. Here we have used embryoid bodies (EBs) as an in vitro model to examine the effect of Gram-negative bacterial endotoxins/lipopolysaccharides (LPS) on the faithful induction of germ lineages during embryogenesis. The role of LPS-inducible cytokine and pluripotency-related DNA-binding protein HMGB1 was also studied in these EBs. Methods: EBs derived from the human embryonic stem cell (hESC) line HUES9 were exposed to 12.5 pg/ml of LPS for 48 h. The expression profile of the ectoderm, endoderm, mesoderm and trophectoderm lineage markers, such as βIII-tubulin, GATA4, BMP2, Brachury and β-hCG, were studied, by RT-PCR and immunofluorescence. Inhibition of mesoderm induction was confirmed by RT-PCR analysis for hANP, cTnT, ABCG2, GATA2, BMP4 and HAND1. Osteoblast differentiation was induced in the EBs, and confirmed by von Kosa and Alizarin red staining. A comet assay was also carried out to assess the degree of apoptosis in these EBs. Results and conclusions: We found that the LPS-treated EBs were selectively silenced for mesoderm markers and failed to differentiate into functional osteoblasts. HMGB1 expression was absent in the normal EBs and was found to be localized in the cytoplasm of the LPS-treated EBs. Overall, our data indicate that endotoxin-induced HMGB1 expression in the peri-implantation-stage embryos can bring about severe birth defects of, for example, the bone and heart. This study also indicates that HMGB1 could be involved in maintenance of pluripotency in the hESCs by impeding their differentiation.
r Pro
of
Human embryonic stem cells (hESCs) havebeen widely used to understand the molecularmechanisms underpinning human develop-ment. These pluripotent cells provide a reliablesource for studying differentiation to all thegerm layer lineages, namely ectoderm, endo-derm, mesoderm and trophectoderm lineages[1,2]. HESCs have been successfully directedtoward the formation of different tissues of var-ious lineages [3]. These cells can also be used toproduce preimplantation embryo- or blasto-cyst-like entities, known as embryoid bodies(EBs), which consist of a differentiated popula-tion of cells representing all the germ layers.These EBs, therefore, closely mimic a growingembryo, which consists of the placental precur-sors (trophectoderm) and the cells of theembryo proper (ectoderm, endoderm and mes-oderm) [4]. It is known that ectoderm forms theskin and the nervous system, the mesodermforms tissues such as the cardiomyocytes, boneand blood, and the endoderm forms , for exam-ple, the liver, lungs and intestine of thedeveloping embryo [5].
Gram-negative bacterial infections of thematernal genital tract, known as bacterial vagi-nosis, can lead to the formation of poor-qualityembryos, which fail to implant [6]. Subclinical orsilent infections of Gram-negative bacteria suchas Chlamydia trachomatis can also cause birthdefects with poorly developed tissues and organsof the fetus [7]. Ethical issues limit studies on themolecular mechanisms underlying such patho-genesis in human embryos. Endotoxin lipopoly-saccaharides (LPS) is the main antigeniccomponent of Gram-negative bacterial cell walland is regularly shed in the surrounding environ-ment. LPS is known to cause various perinatalcomplications [8]. In previous studies we haveestablished the role of various proinflamatoryand other LPS-inducible cytokines and growthfactors, such as IL-1α, IL-1β, TNF-α and CSF1during embryo implantation and in subsequentpregnancy loss [9–11]. However, the molecularevents underlying poor fetal development andbirth defects during silent infections are notknown. We hypothesize that the presence of LPSin the environment of the developing fetus may
utho
008 Future Medicine Ltd ISSN 1746-0751 Regen. Med. (2008) 3(1), xxx–xxx 1

RESEARCH ARTICLE – Sivasubramaniyan, Atluri, Sarda, Arvind, Balaji & Deb
2
selectively inhibit the induction of one or moreof the lineages during early pregnancy.
In this study we have used EBs as an in vitromodel to examine the effect of LPS on the differ-entiation and faithful induction of the germ lin-eages during peri-implantation embryonicdevelopment. The expression of LPS-inducibleand pluripotency-related gene high-mobilitygroup box 1 (HMGB1) was studied to assess itspossible involvement in the aberrant differentia-tion of the LPS-treated EBs [12,13]. HMGB1 isexplicitly expressed by the cells of the inner cellmass and is absent in the trophectoderm cells ofthe blastocyst [14]. HMGB1 is also known as aDNA-binding protein that can regulate expres-sion of genes [12]. Owing to its versatile rolesboth during development and in response toendotoxins, we hypothesized that HMGB1 maybe a key player in mediating LPS-induced devel-opmental defects.
In previous studies we have shown that LPSexposure can render the preimplantation embryoor 5-day-old blastocyst inefficient for implanta-tion [15]. We therefore used early-stage 5-day-oldEBs to closely mimic the peri-implantation stageof embryonic development (day 4–5). We foundthat LPS exposure for 48 h inhibited functionalmesoderm formation in these EBs. LPS-inducedHMGB1 expression in these EBs also indicatesits possible role in silencing mesoderm induc-tion. These findings indicate for the first timethat the presence of endotoxins in the maternalenvironment can lead to predictable mesodermtissue-specific birth defects, such as malforma-tion of bones. This study also indicates thatHMGB1 is related to pluripotency in hESCsand that its expression silences mesoderm-specific genes and differentiation.
Material & methodsCulture of hESCs & production of EBshESC line HUES-9 was obtained from HarvardUniversity (MA, USA) and was used after insti-tutional ethics committee approval. It was main-tained on mouse embryonic feeder (MEF) cells.HUES-9 was maintained in embryonic stem cellmedium (ES medium) consisting of 80%KnockOut DMEM and 20% KnockOut serumreplacement (KSR), supplemented with 2 mML-glutamine, 1% nonessential amino acid solu-tion, 0.1 mM β-mercaptoethanol, 4 ng/mlhuman recombinant basic fibroblast growth fac-tor (βFGF), and penicillin-streptomycin50 U/ml (all from Invitrogen, CA, USA). Forinduction of EB formation, the hESC were
seeded on a low-adherent 60-mm plate (BD Bio-sciences, CA, USA) containing ES media with-out FGF2. Human ESCs from three confluent35-mm dishes were collected after trypsinizationand used for inducing EBs in each l60-mm low-adherent dish. The EBs formed were screenedfor their typical round morphology under a bin-ocular microscope (Nikon). On day 2.5 theywere collected and washed three times in the cul-ture media and finally transferred to fresh media.This step was carried out to dispose of the dyingMEFs and also to select the healthy-looking EBs.
Selection of time points & dose of LPSThe induction of all the germ lineages in the EBsand effect of LPS was tested at two different timepoints (day 4.5 and 9.5) and by using differentdoses of LPS. Early EBs at day 2.5 were washedin the culture media and were divided intogroups of approximately 30 each. Each groupwas then exposed to a dose of 5, 10, 15 or 20pg/ml LPS. A control group of EBs unexposed toLPS was also taken. Following 48 h of incuba-tion the EBs were harvested on day 4.5 or 9.5.The RNA was isolated from each group andRT-PCR-based screening was carried out for theexpression of the ectoderm, endoderm, meso-derm and trophectoderm lineage markers. Allthe doses (ranging from 5 to 20 pg LPS/ml) andboth the time points studied exerted similareffects on the expression pattern of the lineagemarkers. The median-dose 12.5 pg LPS/ml andthe earlier time point of complete lineage induc-tion, for example, day 4.5 was chosen as a repre-sentative dose and time for this study. Allsubsequent studies and analyses were carried outusing this dose and time point.
Exposure of EBs to LPSEBs at day 2.5 were exposed to 12.5 pg/ml ofendotoxin/LPS (Sigma) for 48 h supplementedin culture medium. The normal and the endo-toxin-treated EBs were harvested on day 4.5.Post-exposure, the control and endotoxin-treated EBs were divided into two groups. Onegroup (n = 30) was lysed in TRIZOL for RNAisolation and the other group (n = 90) was fixedin 4% parafomaldehyde for immunofluores-cence. The expression profile of the ectoderm,endoderm, mesoderm and trophectodermlineage markers, such as βIII-tubulin, GATA4,BMP2, Brachury and β-hCG, were studied byRT-PCR and immunofluorescence. Theexpression of the LPS-inducible and pluri-potency-related DNA binding protein
Author Pro
of
Regen. Med. (2008) 3(1) future science groupfuture science group

Endotoxins induce inhibition of mesoderm induction in EBs cells, Osteoblast, HMGB1, differentiation
future science groupfuture science group
A
HMGB1 was also studied in both the controland treated EBs.
RT-PCRTotal RNA from cells was isolated using TRI-ZOL-LS reagent (Invitrogen) as per the manu-facturer’s protocol. Complementary DNA wassynthesized using the SuperScript III First-Strand Synthesis System (Invitrogen) as per themanufacturer’s instructions. PCR was carriedout using 1U Taq DNA polymerase (Sigma) andMgCl2 to a final concentration of 1.5 mM in atotal volume of 25 µl/reaction. GAPDH wasused as the housekeeping control. PCR cyclesconsisted of an initial denaturation at 95ºC for5 min followed by 35 amplification cycles ofdenaturation at 94°C for 45 s, annealing for 45 sand extension at 72°C for 45 s and final exten-sion at 72°C for 10 min. The RT-PCR primers,amplicon sizes and their annealing temperaturesare given in Table 1.
Immunofluorescence & cell countingHESCs were grown on coverslips coated withMEFs and then fixed with 4% paraformalde-hyde (Sigma) followed by permeabilization in0.2% Triton X100 (Sigma). The slides werethen incubated with primary antibodies 1:500dilution of SSEA4 (Chemicon, CA, USA),5 µg/ml Nanog (Santa Cruz Biotechnology,CA, USA), 10 µg/ml Brachury (R&D SystemsInc., MN, USA), and 1.5 µg/ml HMGB1(Sigma) overnight at 4°C. After washing threetimes with PBS, fluorescein isothiocy-anate/Texas red- labeled secondary antibodiesagainst the primary goat/rabbit/mouse wereadded as 1:500 dilutions and incubated for 2 h.DAPI (Sigma) was used for nuclear stainingand then washed with PBS. The negative con-trols were done without primary antibodies.Slides were mounted with DABCC (Sigma)and images were acquired using Nikon Eclipse90i microscope (Nikon Corporation, Japan)and Image-Pro Express software (Media Cyber-netics, Inc., MD, USA). Similarly, the LPS-treated (n = 30) and control (n = 30) EBs weredivided and checked from the immunolocaliza-tion of Brachury and HMGB1. Approximatelyten EBs were checked for each antibody. Theexpression and localization of these proteins inthe EBs were also checked on day 9.5 to seewhether their expression was delayed or alteredas a result of LPS treatment. The results werethen compared with the control cells (non-LPS-treated EBs). To count the number of cells per
EB, the number of DAPI-stained nuclei werecounted in ten each of the control and LPS-treated EBs.
Osteoblast differentiationTo assess the differentiating potential of EBstoward tissues of mesoderm origin, EBs wereproduced and exposed to LPS until day 4.5, asdescribed previously. The normal and LPS-treated EBs, 30 each, were then subjected toosteoblast differentiation from day 8 onward[16]. To stimulate differentiation into osteogeniccells, the EBs were plated on regular 35-mm tis-sue culture dishes, and ES medium containing10–8 M dexamethasone, 50 µg/ml L-ascorbicacid and 5 mM sodium-β-glycophosphate wasused. The medium was changed every 2–3 daysand the differentiation was continued up to15 days. The osteoblast differentiation wascharacterized by identifying mineralized areasusing von Kossa and Alizarin red staining [17].These were visualized and acquired using aNikon Eclipse 90i microscope.
Comet assayDetection of DNA damage in individual EBswas carried out with a slight modification of themethod described by [18]. Comet tail length wascalculated by measuring the streak of DNAcomet tail between the edge of the EBs and theend of tail using Nikon Eclipse 90i microscopeand Image-Pro Express software.
Results Effect of time points & doses of LPS
Induction of all the germ lineages were stud-ied in control or normal EBs on days 4.5 and9.5. Complete lineage induction for the ecto-derm, endoderm, mesoderm and trophectodermwas found as early as day 4.5 by positive expres-sions of the markers (βIII-tubulin, GATA4,BMP2, Brachury and β-hCG) detected byRT-PCR. The day-9.5 EBs were also positive forthe expression of these germ lineage markers. Allthe doses of LPS tested (5–20 pg/ml) inducedspecific silencing of BMP2 and Brachury, themesoderm markers, when tested in EBs at days4.5 and 9.5. Since the induction of all the line-ages could be seen on day 4.5 onward, we con-sidered this as equivalent to a developing peri-implantation blastocyst, and used it for furtherstudies. Similarly, since all the tested doses spe-cifically silenced mesoderm induction, wedecided to use the median dose 12.5 pg/ml forfurther studies.
uthor Pro
of
3www.futuremedicine.com
RESEARCH ARTICLE – Sivasubramaniyan, Atluri, Sarda, Arvind, Balaji & Deb
4
Table 1. List of gene
Gene
Oct 4
Nanog
GAPDH
HMGB1
βIIITubulin
GATA4
Brachury
BMP2
hANP
cTNT
ABCG2
GATA2
HAND1
BMP4
βhCG
Effect of LPS on the expression of pluripotency, germ lineages markers & HMGB1 in the EBsIn this study we have used 5-day-old EBs asentities equivalent to peri-implantation-stageblastocysts. The effect of endotoxins/LPS onthe development and induction of lineages inthe EBs were examined. The hESC lineHUES9 was grown and passaged every 5 days.Figure 1a shows normal phase contrast picturesof this cell line grown on supporting MEF cells.The pluripotency of these cells were checked byRT-PCR analysis for Oct4, SSEA4 and Nanog(Figure 2). The expression of HMGB1 was alsoconfirmed by RT-PCR (Figure 2). We found pos-itive mRNA expression for Oct4, Nanog,SSEA4 and HMGB1 in these normal hESCs atday 5. The localization and expression ofSSEA4, Nanog and HMGB1 was confirmed by
immunofluorescence (Figure 3a, b & c). SSEA4was found to be surface localized, whereasNanog and HMGB1 were localized in thenucleus of the normal HUES9 cells. ThemRNA expression of ectoderm, endoderm,mesoderm and trophectoderm lineage markers,such as βIII-tubulin, GATA4, BMP2, Brachuryand β-hCG, were found to be negative in theHUES9 cells, indicating their undifferentiatedstatus.
The HUES9 cells were harvested on day 5and used for induction of EBs. The control EBswere collected on day 4.5 of culture (Figure 1b).The LPS-treated EBs were also collected on day4.5 and were compared with the normal EBsfor morphological changes under the micro-scope (Figure 1c). The normal and LPS-treatedEBs did not show any visible morphologicaldifferences in terms of their shape, size or num-
s and RT-PCR primers used.
Sequence Annealing temperature (°C)
Product size (bp)
CGACCATCTGCCGCTTTGAGCCCCCTGTCCCCCATTCCTA
57 572
CCTCCTCCATGGATCTGCTTATTCACAGGTCTTCACCTGTTTGTAG
57 262
GGGCGCCTGGTCACCAGGGCTGGGGGCCATCCACAGTCTTCTG
60 531
GCAGATGACAAGCAGCCTTATTTGCTGCATCAGGCTTTCC
60 104
CTTGGGGCCCTGGGCCTCCGAGGCTTCCTGCAGTGGTACACGGGCG
60 174
TCCAAACCAGAAAACGGAAGCTGTGCCCGTAGTGAGATGA
60 187
ACCCAGTTCATAGCGGTGACATGAGGATTTGCAGGTGGAC
60 216
TGTATCGCAGGCACTCAGGTCAGAAGTCTGGTCACGGGGAAT
60 328
GAACCAGAGGGGAGAGACAGAGCCCTCAGCTTGCTTTTTAGGAG
60 383
GGCAGCGGAAGAGGATGCTGAAGAGGCACCAAGTTGGGCATGAACGA
65 151
GTTTATCCGTGGTGTGTCTGGCTGAGCTATAGAGGCCTGGG
55 652
TGACTTCTCCTGCATGCACTAGCCGGCACCTGTTGTGCAA
60 244
TGCCTCAGAAAGAGAACCAGATGGCAGGATGAACAAACAC
60 274
GTCCTGCTAGGAGGCGCGAGGTTCTCCAGATGTTCTTTCG
60 339
GCTACTGCCCCACCATGACCATGGACTCGAAGCGCACATC
55 95
Author Pro
of
Regen. Med. (2008) 3(1) future science groupfuture science group

Endotoxins induce inhibition of mesoderm induction in EBs cells, Osteoblast, HMGB1, differentiation
future science groupfuture science group
Figure 1. Phase conembryoid bodies.
Panel (A) shows morphomouse embryonic feederlipopolysaccharides-treatacquired at 10× magnific
A
A
bers. The normal and LPS-treated EBs werescreened for pluripotency and the germ lineagemarkers. Positive expression for ectoderm,endoderm, mesoderm and trophectoderm line-ages markers βIII-tubulin, GATA4, BMP2,Brachury and β-hCG were found in all the nor-mal EBs on day 4.5 (Figure 2). HMGB1 mRNAexpression was not found in the normal EBs.The LPS-treated EBs showed positive mRNAexpression for HMGB1, βIII-tubulin, GATA4and β-hCG (Figure 2). We found no mRNAexpressions for the two mesoderm markers Bra-chury and BMP2 in these treated EBs. Inhibi-tion of mesoderm induction was furtherconfirmed by the absence of mRNA expressionsfor six other mesoderm markers: BMP4,GATA2, ABCG2, cTnT, hANP and HAND1(Figure 2). The LPS-treated EBs also showed apositive or induced expression of HMGB1(Figure 2).
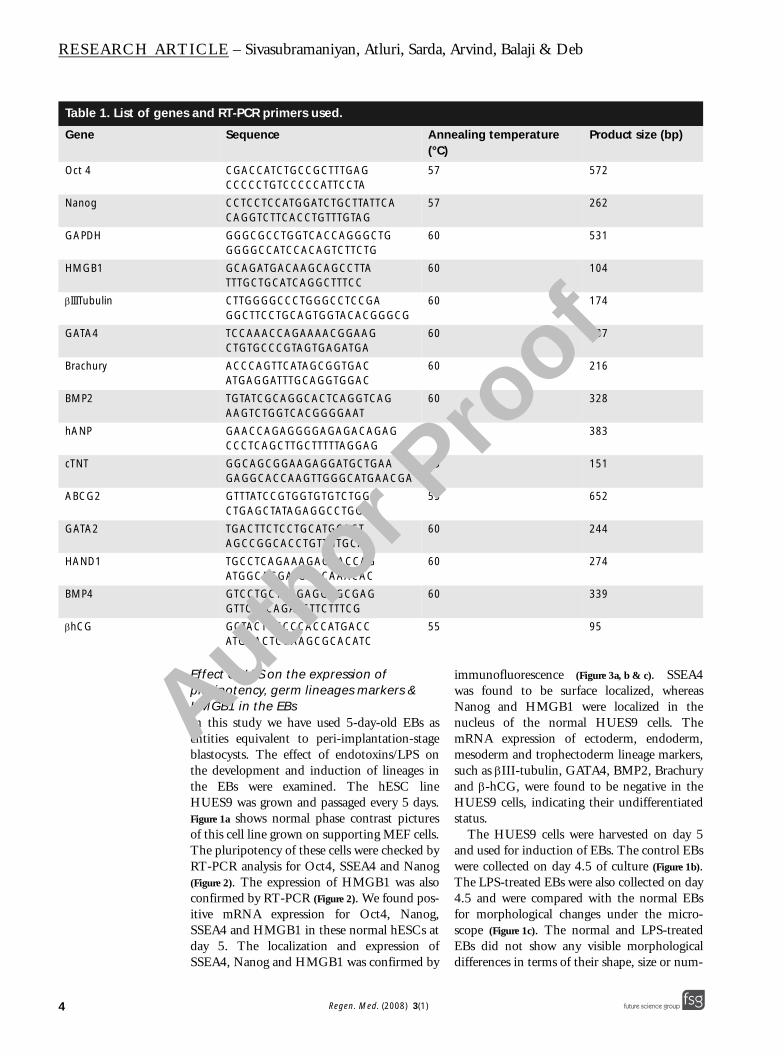
Immunofluorescence localization ofHMGB1 and the mesoderm marker Brachurywas performed to check the protein expressions.Ten EBs each were checked for Brachury andHMGB1 localization in normal and LPS-treated groups. All the normal/control EBswere (100%) positive for the expression of Bra-chury on days 4.5 and 9.5 and, similarly, 100%were negative for HMGB1 (Figure 3d & f). Wedid not see any differential expression of theseproteins across the EBs within the groups. Allthe LPS-treated EBs (100%) showed clear cyto-plasmic localization of HMGB1; however, nosignals for Brachury expression was found inthe EBs on day 4.5 (Figure 3e & g). These EBs,when further tested for Brachury expression onday 9.5, did not show a positive signal. Thisindicates that the expression of Brachury wasactually silenced or downregulated and notmerely delayed by LPS.
Effect of LPS on differentiation of EBs to osteoblastsWe found that the normal EBs could be success-fully differentiated to osteoblast cells, whichwere characterized by mineral depositions con-firmed by Alizarin Red and von Kossa staining atthe end of 15 days of differentiation. The nor-mal EBs could be successfully differentiated asevidenced by positive staining for Alizarin Redand von Kossa (Figure 3h & j). The LPS-treatedEBs (8-day old) failed to differentiate into func-tional osteoblast, as indicated by the absence ofmineral depositions with no positive signals forAlizarin Red and von Kossa (Figure 3l & k).
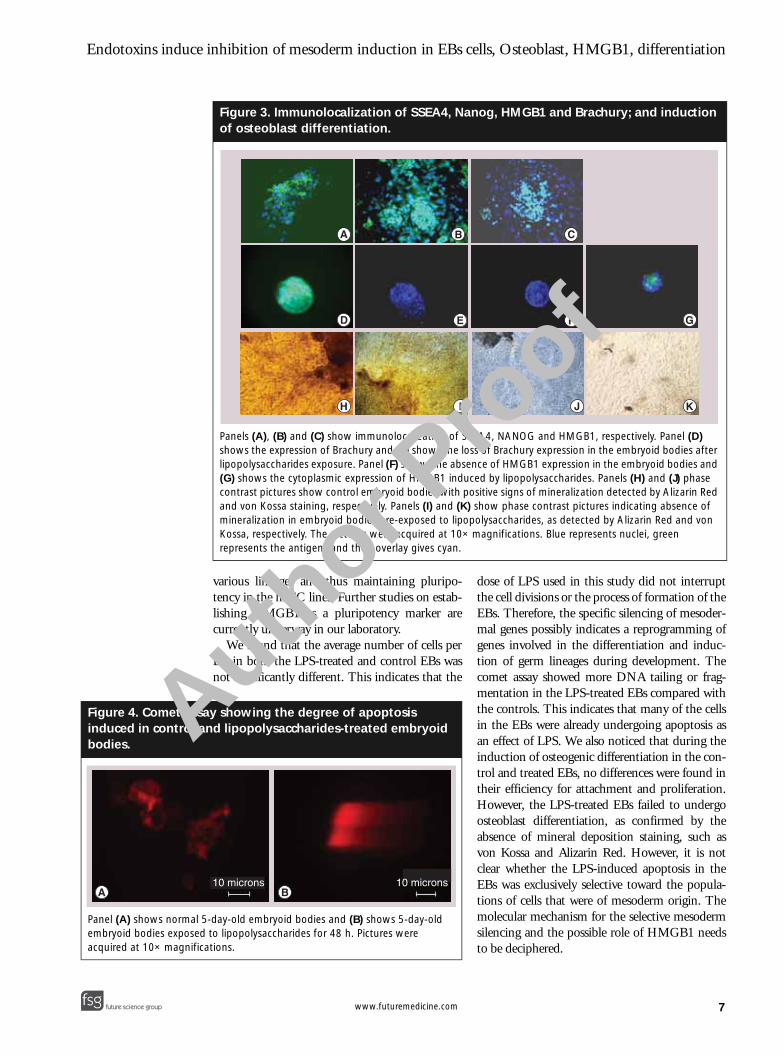
Cell numbers & DNA fragmentationFor a count of the average number of cells perEB, the DAPI-stained nuclei were counted inindividual control and LPS-treated EBs underepifluorescence. The average number of cells/EB(as mean ± standard deviation) in the controlwere 142.33 ± 48.41 cells/EB, and in the LPS-treated group were 175 ± 75.47 cells/EB. Thesevalues did not differ significantly (p = 0.57), asanalyzed by a Students t-test. However, the LPS-treated EBs showed significantly (p = 0.028)more DNA tailing or fragmentation(28.64 ± 14.36 µm average) compared with thecontrol EBs, with an average tailing of2.48 ± 1.0701 µm (Figure 4).
DiscussionGenital tract infection is a predominant cause ofbirth anomalies, both in cases of normal concep-tion or after assisted reproductive techniques [19].Several of these infections are caused by Gram-negative bacteria, such as C. trachomatis, whichare asymptomatic and cause chronic upper tractinfections [20]. In 70% of birth defect cases theunderlying causes are unknown. Here we havestudied the effect of Gram-negative bacterialvaginosis on aberrant fetal development using anembryonic stem cell model. During such injec-tions the preimplantation embryos are exposedto bacterial endotoxins/LPS in the environment[8,11]. The effect of LPS on preimplantationembryonic development and subsequent failureof implantation has been widely studied in ani-mal and rodent models [10,11]. Studies on theunderpinning mechanisms leading to develop-mental abnormalities in human embryos are notpossible owing to ethical limitations in the use ofhuman embryos.
We started with characterization of the day4.5- and 9.5-old EBs for the presence of all the
trast pictures of HUES-9 colonies and
logy of undifferentiated HUES9 colonies growing on . Panels (B) and (C) show morphologies of normal and ed day 5 embryoid bodies, respectively. Pictures were ations.
B C
uthor Pro
of
5www.futuremedicine.com

RESEARCH ARTICLE – Sivasubramaniyan, Atluri, Sarda, Arvind, Balaji & Deb
6
Figure 2. RT-PCR anand ectoderm, endotrophectoderm mar
(A) pluripotency (OCT4, endoderm (GATA4); (D) GATA2, HAND1, BMP4); cells, embryoid bodies, alineage markers were fouembryoid bodies showedlipopolysaccharides-treatmesoderm markers.
Mesoderm
Pluripotency
Endoderm
Ectoderm
Trophectoderm
germ layer lineages. The positive expression ofβIII-tubulin, GATA4, BMP2, Brachury and β-hCG indicated the presence of all the germ layerlineages (the endo-, ecto-, meso- and trophecto-derm) in these EBs on both days. This also estab-lished the fact that the day-4.5-old EBs wereequivalent to developing peri-implantation-stageembryos or blastocysts in terms of their constitu-ent cells representing all the germ lineages. Thesesimilarities between developing embryos andEBs derived from hESCs have been establishedby previous workers [4]. In the present study, LPSwas supplemented in the culture media at a con-centration of 12.5 pg/ml. This dose is more thantwice that of a report, which showed that as lowas 2–5 pg/ml LPS was sufficient to cause altera-tions in the proliferation of hematopoeticprecursor cells in culture [21].
The effect of LPS on the induction of thegerm lineages in the EBs was studied byRT-PCR and immunoflurescence analyses of thelineage-specific markers. We found a specificsilencing of all the eight mesoderm markers,namely Brachury, BMP2, hANP, cTnT,ABCG2, GATA2, BMP4 and HAND1. Wefound that the mesoderm markers were silencedin the day 4.5 and 9.5 EBs after the initial 48-hLPS exposure. This indicates that the expressionof the mesoderm markers in the EBs were reallysilenced for a long time and not merely delayedby the exposure to LPS. The failure to induceosteoblast differentiation after 15 days of osteo-induction in these EBs further strengthens thesuggestion that the effect was long term. Themesoderm lineage is known as a precursor for tis-sues such as the osteogenic cells and hematopoi-etic precursor cells [22]. Birth defects in bone andmuscles are very common and their underlyingcauses largely remain unknown. This study pro-vides for the first time an in vitro human modelfor such studies and indicates a role of LPS insuch abnormalities.
In this study we found that HMGB1 was notexpressed in the normal EBs and its expressionwas induced in the EBs treated with LPS. How-ever, it is also possible that the expression ofHMGB1 is downregulated to undetectable levelsin the normal/control EBs. This reduced sensitiv-ity could be due to the RT-PCR conditions andthe low number (n = 30) of EBs that we screenedper reaction. HMGB1 is also shown to be silencedin the trophectoderm cells of human blastocysts[14]. HMGB1 is a known LPS-inducible cytokine[12] and its cytoplasmic expression in the LPS-treated EBs indicates its possible role as a nonclas-sical proinflamatory cytokine in causing the meso-dermal defects. Anti-HMGB1 antibodies can beused to treat lethal endotoxemia and sepsis [12].Whether this intervention could be effective forprotecting the developing fetus from the adverseeffects of endotoxins is not known. At the sametime, the observed nuclear localization andexpression of HMGB1 in the pluripotent hESCsand its loss of expression in a differentiated popu-lation of cells in the EBs indicate its probableinvolvement in maintenance of stemness in thehESCs. This observation is in support of a previ-ous study, which showed that HMGB1 is specifi-cally expressed in the inner cell mass of theblastocyst [14]. Our data also indicate that nuclearor DNA binding forms of HMGB1 may beinstrumental in silencing differentiation to the
alysis for the expression of pluripotency derm, mesoderm and
kers.
NANOG and HMGB1); (B) ectoderm (βIII Tubulin); (C) mesoderm (Brachury, BMP2, ANP, cTnT, ABCG2, and (E) trophectoderm (βhCG) genes in normal HUES9 nd lipopolysaccharides-treated embryoid bodies. The nd to be absent in the HUES9 cells. The normal expression for all the lineage markers. The ed embryoid bodies showed no expression for the
GAPDH
OCT4
NANOG
HMGB1
β III tubulin
GATA4
Brachury
BMP2
ANP
cTnT
ABCG2
GATA2
HAND1
BMP4
βhCG
HUES-9
EBsLPS tr
eate
d EBs
Author Pro
of
Regen. Med. (2008) 3(1) future science groupfuture science group
Endotoxins induce inhibition of mesoderm induction in EBs cells, Osteoblast, HMGB1, differentiation
future science groupfuture science group
Figure 4. Comet assinduced in control abodies.
Panel (A) shows normal embryoid bodies exposedacquired at 10× magnific
A
A
various lineages and thus maintaining pluripo-tency in the hESC lines. Further studies on estab-lishing HMGB1 as a pluripotency marker arecurrently underway in our laboratory.We found that the average number of cells perEB in both the LPS-treated and control EBs wasnot significantly different. This indicates that the
dose of LPS used in this study did not interruptthe cell divisions or the process of formation of theEBs. Therefore, the specific silencing of mesoder-mal genes possibly indicates a reprogramming ofgenes involved in the differentiation and induc-tion of germ lineages during development. Thecomet assay showed more DNA tailing or frag-mentation in the LPS-treated EBs compared withthe controls. This indicates that many of the cellsin the EBs were already undergoing apoptosis asan effect of LPS. We also noticed that during theinduction of osteogenic differentiation in the con-trol and treated EBs, no differences were found intheir efficiency for attachment and proliferation.However, the LPS-treated EBs failed to undergoosteoblast differentiation, as confirmed by theabsence of mineral deposition staining, such asvon Kossa and Alizarin Red. However, it is notclear whether the LPS-induced apoptosis in theEBs was exclusively selective toward the popula-tions of cells that were of mesoderm origin. Themolecular mechanism for the selective mesodermsilencing and the possible role of HMGB1 needsto be deciphered.
Figure 3. Immunolocalization of SSEA4, Nanog, HMGB1 and Brachury; and induction of osteoblast differentiation.
Panels (A), (B) and (C) show immunolocalization of SSEA4, NANOG and HMGB1, respectively. Panel (D) shows the expression of Brachury and (E) shows the loss of Brachury expression in the embryoid bodies after lipopolysaccharides exposure. Panel (F) shows the absence of HMGB1 expression in the embryoid bodies and (G) shows the cytoplasmic expression of HMGB1 induced by lipopolysaccharides. Panels (H) and (J) phase contrast pictures show control embryoid bodies with positive signs of mineralization detected by Alizarin Red and von Kossa staining, respectively. Panels (I) and (K) show phase contrast pictures indicating absence of mineralization in embryoid bodies pre-exposed to lipopolysaccharides, as detected by Alizarin Red and von Kossa, respectively. The pictures were acquired at 10× magnifications. Blue represents nuclei, green represents the antigens and their overlay gives cyan.
A B C
D E F G
H I J K
ay showing the degree of apoptosis nd lipopolysaccharides-treated embryoid
5-day-old embryoid bodies and (B) shows 5-day-old to lipopolysaccharides for 48 h. Pictures were ations.
10 microns 10 micronsB
uthor Pro
of
7www.futuremedicine.com

RESEARCH ARTICLE – Sivasubramaniyan, Atluri, Sarda, Arvind, Balaji & Deb
8
Executive summary
• Asymptomatic and silof mesoderm origin.
• Lipopolysaccharides (Lvery low concentratio
• Embroid bodies (EBs) successfully employedlineages.
• HMGB1, which is explocalized and is associ
• Induction of EBs fromhave significantly dow
• LPS induces HMGB1 e
• LPS-induced HMGB1
• This study indicates thcaused by silent genit
ConclusionOur study demonstrates for the first time a corre-lation between Gram-negative bacterial LPS andbirth defects related to formation of tissues ofmesoderm origin, such as the bones, blood and/orheart muscles. We have also shown that early EBscould be effectively employed as a model systemto study fetal abnormalities caused by maternalinfections or due to new drugs. Expression andcytoplasmic localization of the DNA-bindingcytokine HMGB1 in the EBs after LPS exposureindicates its probable involvement in the forma-tion of developmentally compromised embryosduring such infections. At the same time, ourfinding strongly indicates that nuclear localizationof HMGB1 maintains pluripotency in hESCs byinhibiting the faithful induction of all the germlayer lineages.
Financial & competing interests disclosure The authors have no relevant affiliations or financialinvolvement with any organization or entity with a finan-cial interest in or financial conflict with the subject matteror materials discussed in the manuscript. This includesemployment, consultancies, honoraria, stock ownership oroptions, expert testimony, grants or patents received orpending, or royalties.
No writing assistance was utilized in the production ofthis manuscript.
Ethical conduct of research The author states that they have obtained appropriate insti-tutional review board approval or have followed the princi-ples outlined in the Declaration of Helsinki for all human oranimal experimental investigations. In addition, for investi-gations involving human subjects, informed consent has beenobtained from the participants involved.
ent Gram-negative genital tract infections can lead to birth defects, specifically of the bone or other tissues
PS), the most common antigen from Gram-negative bacterial cell wall, can cause such birth defects, even at ns.
derived from human embryonic stem cells (hESCs) closely mimic a developing embryo and can be to study developmental toxicity of LPS or any other molecule/drug on differentiation or induction of the
ressed by the inner cell mass of a human blastocyst is also expressed by the pluripotent hESCs is nuclear ated with maintenance of pluripotency.
hESCs, which consist of a mixed population of differentiated cells of all the lineages, do not express, or n-regulated expression of HMGB1, indicating that HMBG1 can be involved in maintenance of pluripotency.
xpression in the cells of the EBs, and its protein is localized in the cytoplasm of these cells.
expression is involved in silencing of mesoderm induction.
at several of the unexplained birth defects during normal pregnancy and after in vitro fertilization can be al tract infections of Gram-negative bacteria.
4. Carpenedo RL, Sargent CY, McDevitt TC: 7. Romero R, Espinoza J, Mazor: Can
uthor Pro
of
Bibliography1. Thomson JA, Itskovitz-Eldor J, Shapiro SS
et al.: Embryonic stem cell lines derived from human blastocysts. Science 282, 1145–1147 (1998).
2. Reubinoff BE, Pera MF, Fong CY et al.: Embryonic stem cell lines from human blastocysts: somatic differentiation in vitro. Nat. Biotechnol. 18, 399–404 (2000).
3. Deb KD, Devi Prakash A, Sharma V, Totey S: Embryonic stem cells: from markers to market. Rejuvenation Res. (2007) (Epub ahead of print).
Rotary suspension culture enhances the efficiency, yield and homogeneity of embryoid body differentiation. 25(9), 2224–2234 Stem Cells (2007)(In Press).
5. Dvash T, Benvenisty N: Human embryonic stem cells as a model for early human development. Best Pract. Res. Clin. Obstet. Gynaecol. 18(6), 929–940 (2004).
6. Deb K, Chatturvedi MM, Jaiswal YK: Gram-negative bacterial endotoxin induced infertility: a birds eye view. Gynecol. Obstet. Invest. 57, 224–232 (2004).
endometrial infection/inflammation explain implantation failure, spontaneous abortion, and preterm birth after in vitro fertilization? Fertil. Steril. 82(4), 799–780 (2004).
8. Deb K, Chaturvedi MM, Jaiswal YK: Comprehending the role of LPS in Gram-negative bacterial vaginosis: ogling into the causes of unfulfilled child-wish. Arch. Gynecol. Obstet. 270(3), 133–146 (2004).
9. Deb K, Chaturvedi MM, Jaiswal YK: A ‘minimum dose’ of LPS required for implantation failure: assessment of its effect on the maternal reproductive organs and IL-
A
Regen. Med. (2008) 3(1) future science groupfuture science group

Endotoxins induce inhibition of mesoderm induction in EBs cells, Osteoblast, HMGB1, differentiation
1α expression in mouse. Reproduction 128(1), 87–97 (2004).
10. Deb K, Chaturvedi MM, Jaiswal YK: The role of TNF-α in Gram-negative bacterial LPS induced implantation failure. Reprod. Med. Biol. 4, 79–88 (2005).
11. Deb K, Chaturvedi MM, Jaiswal YK: Gram-negative bacterial LPS induced poor uterine receptivity and implantation failure in mouse: alterations in IL-1β expression in the preimplantation embryo and uterine horns. Infect. Dis. Obstet. Gynecol. 13(3), 125–133 (2005).
12. Yang H, Wang H, Czura CJ, Tracy KJ: The cytokine activity of HMGB1. J. Leukoc. Biol. 78, 1–8 (2005).
13. Skottman H, Mikkola M, Lundin K et al.: Gene expression signatures of seven individual human embryonic stem cell lines. Stem Cells 23, 1343–1356 (2005).
14. Adjaye J, Huntriss J, Herwig R et al.: Primary differentiation in the human
blastocyst: comparative molecular portraits of inner cell mass and trophectoderm cells. Stem Cells 23(10), 1514–1525 (2005).
15. Jaiswal YK, Chaturvedi MM, Deb K: Effect of bacterial endotoxins on superovulated mouse embryos in vivo: is CSF-1 involved in endotoxin induced pregnancy loss? Infect. Dis. Obstet. Gynecol. 32050, 1–9 (2006).
16. Duplomb L, Dagouassat M, Jourdon P et al.: Embryonic stem cells: new tool to study osteoblast and osteoclast differentiation. Stem Cells 25(3), 544–552 (2007).
17. Karp JM, Ferraeira LS, Ali K et al.: Cultivation of human embryonic stem cells without the embryoid body step enhances ostogenesis in vitro. Stem Cells 24, 835–843 (2006).
18. Takahashi M, Saka N, Takahashi H et al.: Assessment of DNA damage in individual hamster embryos by comet assay. Mol. Reprod. Dev. 54, 1–7 (1999).
19. Edwards RG, Ludwig M: Are major defects in children conceived in vitro due to innate problems in patients or to induced genetic damage? Reprod. Biomed. 7(2), 131–138 (2003).
20. Ingalls RR, Rice AP, Qureshi N et al.: The inflammatory cytokine response to chlamydia trachomatis infection is endotoxin mediated. Infect. Immun. 3125–3130 (1995).
21. Rinehart JJ, Keville L: Effects of endotoxin on proliferation of human hematopoietic cell precursors. Cytotechnology 24(2), 153–159 (1997).
22. Hyslop LA, Armstrong L, Stojkovic M, Lako M: Human embryonic stem cells: biology and clinical implications. Expert Rev. Mol. Med. 7(19), 1–21 (2005).
Author Pro
of
9future science groupfuture science group www.futuremedicine.com





























