Embed Size (px)
Citation preview

INTRODUCTION
A cascade of inductive interactions establishes the dorsoventralaxis of the vertebrate embryo (Harland and Gerhart, 1997;Moon and Kimelman, 1998; Nieto, 1999). In amphibians, theinduction of dorsal structures is initiated by a dorsovegetalsignaling center called the Nieuwkoop center (Harland andGerhart, 1997). The analogous region in teleosts correspondsto blastomeres at the dorsal margin and the dorsal yolksyncytial layer (YSL), an extraembryonic structure formedwhen marginal blastomeres fuse with the yolk at themidblastula stage (Long, 1983; Kimmel et al., 1995; Mizunoet al., 1996; Schier and Talbot, 1998).
The Nieuwkoop center is distinguished by the nuclearlocalization of maternal β-catenin protein (Schneider et al.,1996; Larabell et al., 1997), a transcriptional effector in theWnt signaling pathway that is required for the initiation ofdorsal development (Heasman et al., 1994; Behrens et al.,1996; Molenaar et al., 1996; Fagotto et al., 1997; Pelegri and
Maischen, 1998). In zebrafish, β-catenin specificallyaccumulates in nuclei of dorsal blastomeres and the dorsalYSL (Schneider et al., 1996), where it activates, directly orindirectly, the expression of several zygotic genes that mediatethe development of dorsal axial structures (Wylie et al., 1996).The homeobox gene bozozok(boz; also known as dharmaandnieuwkoid) is among the first genes activated by β-catenin(Yamanaka et al., 1998; Koos and Ho, 1998; Fekany et al.,1999). Soon after the onset of zygotic transcription, boz isexpressed in blastomeres at the dorsal margin. In the lateblastula and early gastrula, bozis expressed in the dorsal YSL.Prechordal plate, notochord, floor plate and forebrain arevariably disrupted inbozmutants, indicating that bozfunctionsin the development of dorsal mesoderm and ventral neural tube(Schier et al., 1996; Solnica-Krezel et al., 1996; Fekany et al.,1999).
In addition to the transcriptional regulator Boz, a number ofsecreted proteins play key roles in the development of dorsalmesoderm. One such protein is encoded by squint (sqt), a
2583Development 127, 2583-2592 (2000)Printed in Great Britain © The Company of Biologists Limited 2000DEV3155
In vertebrate embryos, maternal β-catenin proteinactivates the expression of zygotic genes that establish thedorsal axial structures. Among the zygotically acting geneswith key roles in the specification of dorsal axial structuresare the homeobox gene bozozok(boz) and the nodal-related(TGF-β family) gene squint (sqt). Both genes are expressedin the dorsal yolk syncytial layer, a source of dorsalmesoderm inducing signals, and mutational analysis hasindicated that boz and sqt are required for dorsalmesoderm development. Here we examine the regulatoryinteractions among boz, sqt and a second nodal-relatedgene, cyclops(cyc). Three lines of evidence indicate that bozand sqt act in parallel to specify dorsal mesoderm andanterior neuroectoderm. First, bozrequires sqt function toinduce high levels of ectopic dorsal mesoderm, consistentwith sqt acting either downstream or in parallel to boz.Second, sqt mRNA is expressed in blastula stage bozmutants, indicating that boz is not essential for activationof sqttranscription, and conversely, bozmRNA is expressed
in blastula stage sqtmutants. Third, boz;sqt double mutantshave a much more severe phenotype than bozand sqt singlemutants. Double mutants consistently lack the anteriorneural tube and axial mesoderm, and ventral fates aremarkedly expanded. Expression of chordin and noggin1 isgreatly reduced in boz;sqtmutants, indicating that the bozand sqt pathways have overlapping roles in activatingsecreted BMP antagonists. In striking contrast to boz;sqtdouble mutants, anterior neural fates are specified inboz;sqt;cyc triple mutants. This indicates that cycrepressesanterior neural development, and that boz and sqtcounteract this repressive function. Our results support amodel in which boz and sqt act in parallel to inducedorsalizing BMP-antagonists and to counteract therepressive function of cyc in neural patterning.
Key words: bozozok, squint, cyclops, Nodal, Zebrafish, Dorsalmesoderm, Neural pattern
SUMMARY
bozozok and squint act in parallel to specify dorsal mesoderm and anterior
neuroectoderm in zebrafish
Howard I. Sirotkin 1, Scott T. Dougan 1, Alexander F. Schier 2 and William S. Talbot 1,*1Department of Developmental Biology, Stanford University School of Medicine, 279 Campus Drive, Beckman Center B300,Stanford, CA 94305, USA2Developmental Genetics Program, Skirball Institute of Biomolecular Medicine, New York University School of Medicine, New York,NY 10016, USA*Author for correspondence (e-mail: [email protected])
Accepted 30 March; published on WWW 23 May 2000

2584
member of the nodal-related subclass of the TGF-βsuperfamily that is essential for early steps in dorsal mesodermdevelopment (Feldman et al., 1998). sqt is expressed in dorsalmarginal blastomeres soon after the midblastula transition, andthe expression domain then expands to include the dorsal YSL(Erter et al., 1998; Feldman et al., 1998). Analysis of doublemutants has revealed that sqt and a second zebrafish nodal-related gene, cyclops(cyc), have partially redundant functionsin mesoderm development (Feldman et al., 1998; Rebagliati etal., 1998a; Sampath et al., 1998; reviewed in Schier and Shen,2000).
Antagonists of ventralizing bone morphogenetic protein(BMP) signals comprise a second class of secreted factorsrequired for the development of dorsal structures. Members ofthe BMP subclass of the TGF-β superfamily, including Bmp2,Bmp4 and Bmp7, have potent ventralizing activity in explantsand when overexpressed in embryos (Dale et al., 1992; Jones etal., 1992; Hemmati-Brivanlou and Thomsen, 1995; Nishimatsuand Thomsen, 1998). Mutational analysis shows that thesefactors are required for the formation of ventral cell types(Kishimoto et al., 1997; Nguyen et al., 1998; Dick et al., 2000;Schmid et al., 2000). β-catenin and factors downstream of itinduce expression of proteins that oppose the action ofventralizing BMPs, including Chordin (Sasai et al., 1994;Piccolo et al., 1996; Pelegri and Maischein, 1998), Noggin(Smith and Harland 1992; Zimmerman et al., 1996) andFollistatin (Hemmati-Brivanlou et al., 1994).
The cross-regulatory interactions among the genes requiredfor development of dorsal mesoderm are not well defined. Forexample, overexpression of boz or sqt within the YSL non-autonomously induces dorsal mesoderm formation within theoverlying blastoderm (Yamanaka et al., 1998; Koos and Ho,1998; Feldman et al., 1998), and overexpression of mousenodalcan rescue dorsal mesoderm development in bozmutants(Fekany et al., 1999). Thus boz may activate sqt to inducedorsal mesoderm. Nevertheless, these results do not excludethe possibility that sqt and boz act in parallel during dorsalmesoderm induction, since it has not been determined whetherboz is necessary for sqt expression. Additional complexityarises from the observations that boz and sqt can inducechordinexpression in zebrafish embryos and Xenopusexplants,respectively (Koos and Ho, 1998; Rebagliati et al., 1998b),suggesting that β-catenin could activate chordin through apathway involving boz, sqtor both.
In order to elucidate the regulatory relationships among thegenes required for the development of dorsal structures, wehave analyzed the genetic interactions among boz, sqtand cyc.In contrast to previous models in which boz activates sqtexpression, we report that the activation of sqt transcriptiondoes not require boz function. Instead, we find that boz andsqt are activated in parallel pathways required for dorsaldevelopment. Both genes are required for proper expression ofnoggin1while either gene is sufficient for the expression ofchordin. Ventral fates are markedly expanded in boz;sqtmutants, consistent with the loss of BMP antagonists in theseembryos. Whereas boz and sqt single mutants have variableeffects on forebrain and axial mesoderm development, boz;sqtdouble mutants have a severe truncation of the anterior neuraltube and a complete loss of axial mesoderm. In strikingcontrast to boz;sqt double mutants, anterior neural fates arespecified in boz;sqt;cyc triple mutants. This indicates that cyc
represses anterior neural development, and that boz and sqtcounteract this repressive function. Our results support a modelin which bozand sqt act in parallel to induce dorsalizing BMP-antagonists and to counteract the repressive function of cyc inneural patterning.
MATERIALS AND METHODS
Mutant alleles and embryo collectionThe mutations utilized in this study, bozm168, sqtcz35and cycm294, havebeen described previously (Feldman et al., 1998; Rebagliati et al.,1998a; Sampath et al., 1998; Fekany et al., 1999). Embryos werecollected from natural matings, raised at 28°C, and staged bymorphology as described (Kimmel et al., 1995). Ages in hourspostfertilization (h) were normalized to reflect times for a givenmorphological stage as reported by Kimmel et al. (1995).
RNA microinjectionTo obtain sense boz mRNA for microinjection, the pCMV-sportdharma plasmid (Yamanaka et al., 1998) was linearized with HpaI andtranscribed with SP6 polymerase using the mMessage mMachine kit(Ambion). sqtmRNA was transcribed from the pCS2+ squint plasmid(Feldman et al., 1998) using SP6 polymerase after linearization withNotI. cyc mRNA was transcribed from the pCS2+Cyclops plasmid(Rebagliati et al., 1998a) using T7 polymerase after linearization withNotI. RNA was diluted in 5 mg/ml Phenol Red, 0.2 M KCl prior tomicroinjection. 0.5 nl of RNA was microinjected into 1-4 cellembryos that had been dechorionated by Pronase treatment or 4-8 cellembryos were injected through the chorion. At the appropriate stageembryos were fixed in 4% paraformaldehyde in phosphate-bufferedsaline (PBS) and stored in methanol at –20°C.
In situ hybridizationEmbryos collected for in situ hybridization were fixed in 4%paraformaldehyde in PBS at 4°C and manually dechorionated.Antisense RNA was synthesized and hybridizations were carried outas described (Thisse et al., 1993). Constructs used to synthesizeantisense probes for sqt, chordin, cyc, boz, gsc, axial, hgg1, otx2,pax6.1, pax2.1, emx1and krox20 have been previously described(Krauss et al., 1991a,b; Oxtoby and Jowett, 1993; Strähle et al., 1993;Mori et al., 1994; Thisse et al., 1994; Morita et al., 1995; Miller-Bertoglio et al., 1997; Schulte-Merker et al., 1997; Feldman et al.,1998; Sampath et al., 1998; Yamanaka et al., 1998). Antisensenoggin1 probe was transcribed with SP6 polymerase from SalI-linearized IMAGE zebrafish EST clone fc23h06, which correspondsto the noggin1gene (Furthauer et al., 1999).
Photography and genotypingLive embryos were manually dechorionated, anesthetized inTricaine and photographed in 2.5-3.5% methycellulose in embryomedium as described (Kimmel et al., 1995). Following in situhybridization, embryos were stored in methanol at –20°C. Prior tophotography embryos were cleared in benzyl benzoate:benzylalcohol (2:1), mounted in Canada balsam:methyl salicylate (40:1)and photographed using a Zeiss Axioplan microscope. Forgenotyping after photography, embryos were washed twice in benzylbenzoate: benzyl alcohol (2:1), 2-3 times in 100% methanol andonce in PBS with 0.1% Tween-20. Genomic DNA was extractedusing the QiaAmp Tissue Kit (Qiagen). Following extraction,genomic DNA was precipitated and resuspended in 50-80 µl H20(embryos younger than 40% epiboly were resuspended in 50µl). 5µl of the genomic DNA was used as a template in each PCR reaction.Primers and conditions for genotyping sqtcz35, cycm294 andbozm168
have been described (Feldman et al., 1998; Sampath et al., 1998;Fekany et al., 1999).
H. I. Sirotkin and others

2585boz and sqt in dorsal axial development
RESULTS
boz requires Nodal-related signals to induce dorsalmesodermPrevious studies have shown that boz and sqt are expressed inthe YSL and dorsal marginal blastomeres and that both bozand sqt can induce dorsal mesoderm in wild-type embryos(Erter et al., 1998; Feldman et al., 1998; Koos and Ho, 1998;Yamanaka et al., 1998). To begin to understand therelationships between boz, sqt and cyc, we tested whether theactivity of boz requires Nodal-related signals (Fig. 1). Weinjected synthetic boz mRNA into wild-type, cyc, sqt andsqt;cyc embryos and assayed the homeobox gene goosecoid(gsc) as a marker for dorsal mesoderm. Microinjection of 75 pg
of boz mRNA strongly induces gscin wild-type embryos (Fig.1B; Koos and Ho, 1998; Yamanaka et al., 1998) and cycmutants (data not shown). In contrast, bozcan induce gsconlyweakly in sqt mutants (Fig. 1D), and no gsc expression isdetectable in sqt;cyc mutants injected with boz mRNA (Fig.1F). While boz is unable to induce dorsal mesoderm in theabsence of Nodal signaling, it can still dorsalize sqt;cycmutants, as indicated by induction of chordin (chd) (Fig. 1H),which is expressed in axial and paraxial mesoderm and dorsalectoderm in wild-type zebrafish embryos (Miller-Bertoglio etal., 1997; Schulte-Merker et al., 1997). Because boz cannotinduce dorsal mesoderm in the absence of Nodal-relatedsignals, we conclude that sqt functions either downstream ofbozor that sqt and bozact in parallel pathways required for thedevelopment of dorsal mesoderm.
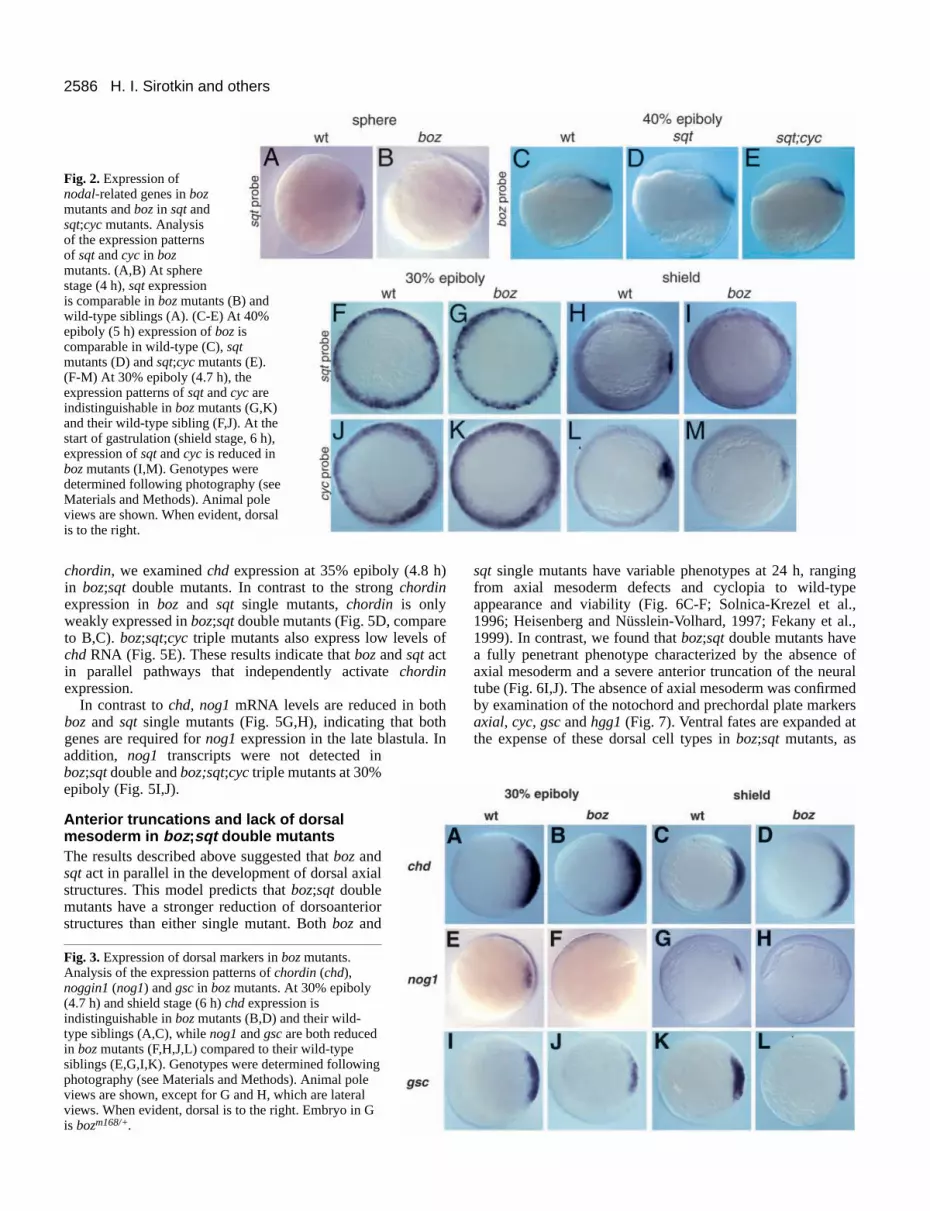
boz and sqt are not required to initiate each other’sexpressionAs one test to distinguish whether Nodal-related signals actdownstream of or parallel to boz, we examined sqt and cycexpression in boz mutants. The early phases of sqt and cycexpression are intact in bozmutants (Fig. 2B,G,K), while laterexpression is decreased (Fig. 2I,M). In the reciprocal set ofexperiments, we examined the expression of boz in sqt andsqt;cyc double mutants. At 40% epiboly (5 h) the expressionof boz is intact in both sqt and sqt;cyc double mutants (Fig.2D,E). Thus early expression of sqtand cyc is independent ofboz function, and bozexpression does not require the activityof Nodal-related signals. These results support the conclusionthat the expression of bozand sqt is initiated in parallel.
Expression of noggin1 and goosecoid , but notchordin , requires bozboz does not regulate the earliest phases of sqt or cycexpression, suggesting that it controls other genes that inducethe development of dorsal cell types. To determine if boz isrequired for the expression of chd or noggin1(nog1) (Miller-Bertoglio et al., 1997; Schulte-Merker et al., 1997; Furthaueret al., 1999), we analyzed their expression in bozmutants. At30% epiboly (4.7 h) and shield stage (6 h), expression of chdin boz mutants is comparable to wild type (Fig. 3B,D). Incontrast, at 30% epiboly, nog1mRNA is not detectable in bozmutants (Fig. 3F), and expression is reduced or absent at shieldstage (Fig. 3H). Thus nog1 expression is dependent on bozfunction, but other regulators must activate chordinexpression.As in previous reports (Solnica-Krezel et al., 1996; Fekany etal., 1999), expression of gscis reduced in bozmutants at 30%epiboly (Fig. 3J) and shield stage (Fig. 3L).
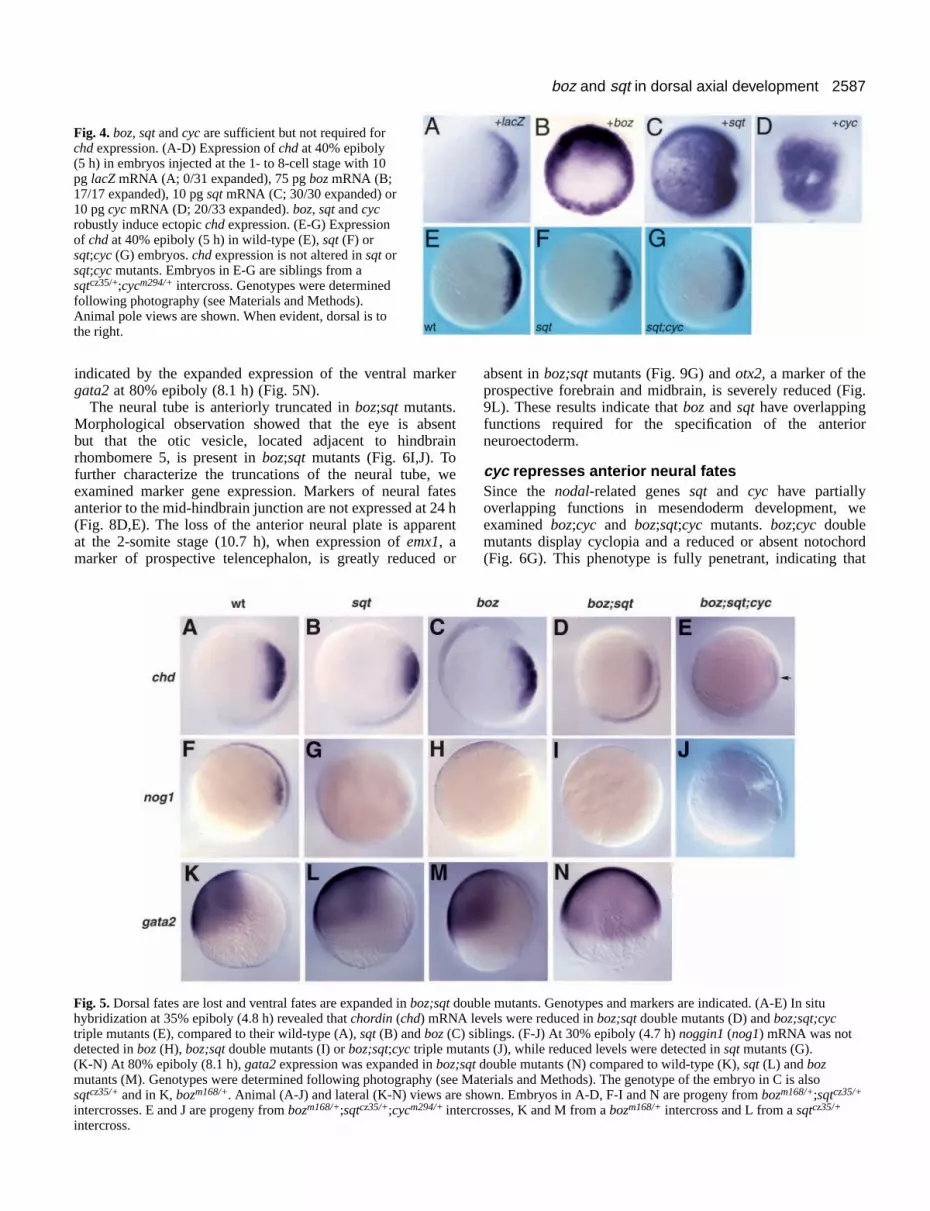
boz and Nodal-related signals act in parallel toregulate chordinWhile expression of chd does not require boz, overexpressionof boz strongly induces chd (Koos and Ho, 1998; Fig. 4B).Similarly, both sqt (Rebagliati et al., 1998b; Fig. 4C) and cyc(Fig. 4D) can inducechd, but expression ofchd is not alteredin sqt or sqt;cyc mutants at 40% epiboly (Fig. 4F,G). Thusoverexpression of boz, sqt or cyc is sufficient to induce chdexpression, but analysis of bozmutants and single and doublemutants for the nodal-related genes shows that they are notessential for chd expression in the late blastula.
To determine if bozand sqtmight act redundantly to activate
Fig. 1. bozozokrequires Nodal-related signals to induce dorsalmesoderm. (A-H) Expression of gsc(A-F) at 60% epiboly (7 h) orchd(G,H) at shield stage (6 h) in embryos injected at the 1-4 cellstage with 75 pg lacZ mRNA (A,C,E,G) or 75 pg bozmRNA(B,D,F,H). Genotypes of embryos are indicated. bozcan readilyinduce dorsal mesoderm in wild-type embryos (B), but only weaklyin sqt mutants (D; 10/11 sqtmutants had little or no ectopicexpression), and not at all in sqt;cycdouble mutants (F; 6/6 noexpression). Ectopic chd is induced in sqt;cycdouble mutants (H;Among 32 embryos from sqtcz35/+; cycm294/+ intercross, all hadexpanded chd, and 1 of 10 genotyped embryos was a sqt;cycmutant).Embryos in A-D, E,F and G,H are siblings from three sqtcz35/+;cycm294/+ intercrosses. Animal pole views are shown. When evident,dorsal is to the right. Genotypes were determined followingphotography (see Materials and Methods). Genotype of wild-typeembryo in (A) is cycm294/+and (B) is sqtcz35/+.
2586
chordin, we examined chd expression at 35% epiboly (4.8 h)in boz;sqt double mutants. In contrast to the strong chordinexpression in boz and sqt single mutants, chordin is onlyweakly expressed in boz;sqt double mutants (Fig. 5D, compareto B,C). boz;sqt;cyc triple mutants also express low levels ofchdRNA (Fig. 5E). These results indicate that bozand sqt actin parallel pathways that independently activate chordinexpression.
In contrast to chd, nog1mRNA levels are reduced in bothboz and sqt single mutants (Fig. 5G,H), indicating that bothgenes are required for nog1expression in the late blastula. Inaddition, nog1 transcripts were not detected inboz;sqtdouble and boz;sqt;cyctriple mutants at 30%epiboly (Fig. 5I,J).
Anterior truncations and lack of dorsalmesoderm in boz;sqt double mutantsThe results described above suggested that bozandsqtact in parallel in the development of dorsal axialstructures. This model predicts that boz;sqt doublemutants have a stronger reduction of dorsoanteriorstructures than either single mutant. Both boz and
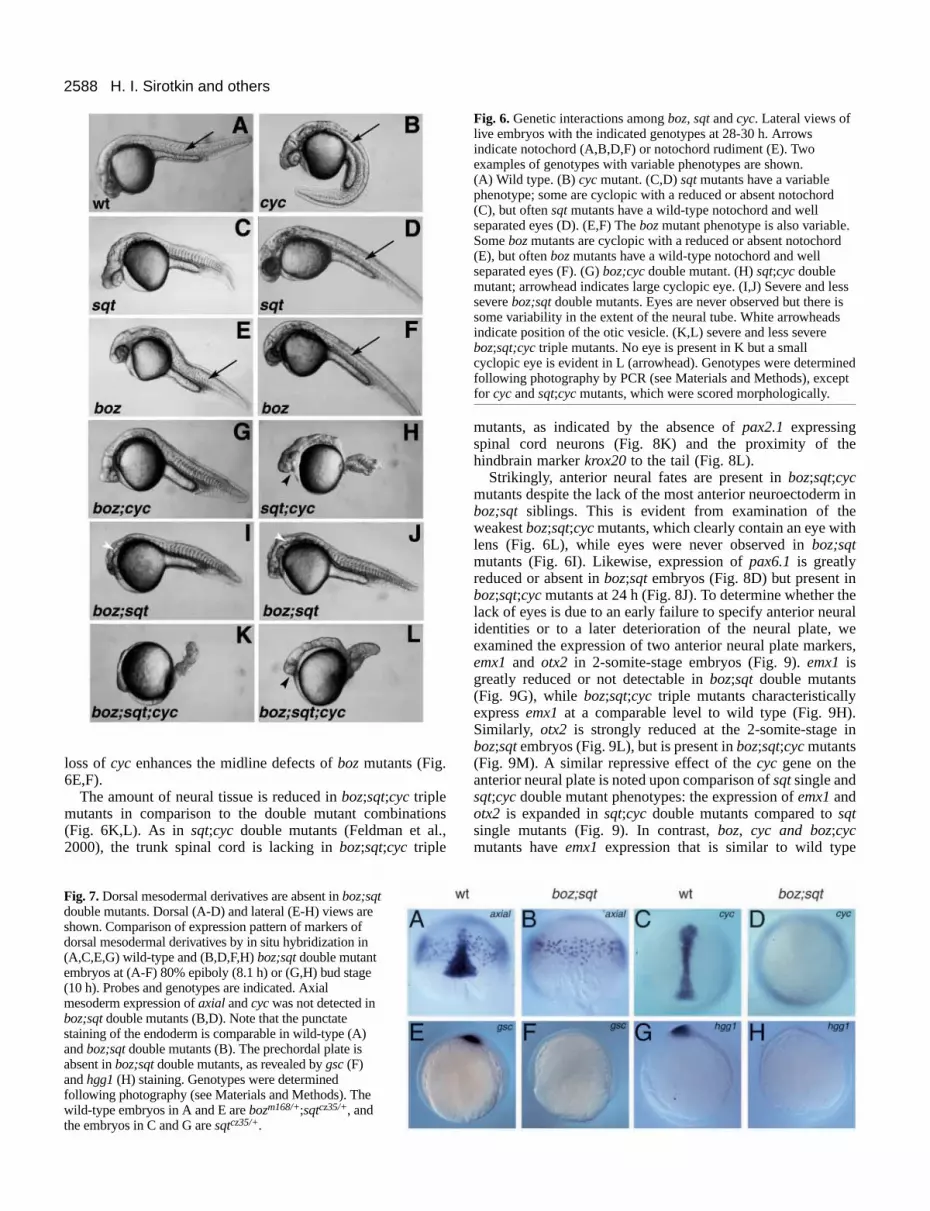
sqt single mutants have variable phenotypes at 24 h, rangingfrom axial mesoderm defects and cyclopia to wild-typeappearance and viability (Fig. 6C-F; Solnica-Krezel et al.,1996; Heisenberg and Nüsslein-Volhard, 1997; Fekany et al.,1999). In contrast, we found that boz;sqt double mutants havea fully penetrant phenotype characterized by the absence ofaxial mesoderm and a severe anterior truncation of the neuraltube (Fig. 6I,J). The absence of axial mesoderm was confirmedby examination of the notochord and prechordal plate markersaxial, cyc, gscand hgg1(Fig. 7). Ventral fates are expanded atthe expense of these dorsal cell types in boz;sqt mutants, as
H. I. Sirotkin and others
Fig. 2. Expression ofnodal-related genes in bozmutants and bozin sqtandsqt;cycmutants. Analysisof the expression patternsof sqtand cyc in bozmutants. (A,B) At spherestage (4 h), sqtexpressionis comparable in bozmutants (B) andwild-type siblings (A). (C-E) At 40%epiboly (5 h) expression of boziscomparable in wild-type (C), sqtmutants (D) and sqt;cycmutants (E).(F-M) At 30% epiboly (4.7 h), theexpression patterns of sqtand cycareindistinguishable in bozmutants (G,K)and their wild-type sibling (F,J). At thestart of gastrulation (shield stage, 6 h),expression of sqt and cyc is reduced inbozmutants (I,M). Genotypes weredetermined following photography (seeMaterials and Methods). Animal poleviews are shown. When evident, dorsalis to the right.
Fig. 3. Expression of dorsal markers in bozmutants.Analysis of the expression patterns of chordin (chd),noggin1(nog1) and gscin bozmutants. At 30% epiboly(4.7 h) and shield stage (6 h) chdexpression isindistinguishable in bozmutants (B,D) and their wild-type siblings (A,C), while nog1and gscare both reducedin bozmutants (F,H,J,L) compared to their wild-typesiblings (E,G,I,K). Genotypes were determined followingphotography (see Materials and Methods). Animal poleviews are shown, except for G and H, which are lateralviews. When evident, dorsal is to the right. Embryo in Gis bozm168/+.
2587boz and sqt in dorsal axial development
indicated by the expanded expression of the ventral markergata2at 80% epiboly (8.1 h) (Fig. 5N).
The neural tube is anteriorly truncated in boz;sqt mutants.Morphological observation showed that the eye is absentbut that the otic vesicle, located adjacent to hindbrainrhombomere 5, is present in boz;sqt mutants (Fig. 6I,J). Tofurther characterize the truncations of the neural tube, weexamined marker gene expression. Markers of neural fatesanterior to the mid-hindbrain junction are not expressed at 24 h(Fig. 8D,E). The loss of the anterior neural plate is apparentat the 2-somite stage (10.7 h), when expression of emx1, amarker of prospective telencephalon, is greatly reduced or
absent in boz;sqt mutants (Fig. 9G) and otx2, a marker of theprospective forebrain and midbrain, is severely reduced (Fig.9L). These results indicate that bozand sqt have overlappingfunctions required for the specification of the anteriorneuroectoderm.
cyc represses anterior neural fatesSince the nodal-related genes sqt and cyc have partiallyoverlapping functions in mesendoderm development, weexamined boz;cyc and boz;sqt;cyc mutants. boz;cyc doublemutants display cyclopia and a reduced or absent notochord(Fig. 6G). This phenotype is fully penetrant, indicating that
Fig. 4. boz, sqtand cycare sufficient but not required forchdexpression. (A-D) Expression of chd at 40% epiboly (5 h) in embryos injected at the 1- to 8-cell stage with 10pg lacZ mRNA (A; 0/31 expanded), 75 pg bozmRNA (B;17/17 expanded), 10 pg sqtmRNA (C; 30/30 expanded) or10 pg cycmRNA (D; 20/33 expanded). boz, sqt and cycrobustly induce ectopic chdexpression. (E-G) Expressionof chd at 40% epiboly (5 h) in wild-type (E), sqt(F) orsqt;cyc(G) embryos. chdexpression is not altered in sqtorsqt;cyc mutants. Embryos in E-G are siblings from asqtcz35/+;cycm294/+ intercross. Genotypes were determinedfollowing photography (see Materials and Methods).Animal pole views are shown. When evident, dorsal is tothe right.
Fig. 5. Dorsal fates are lost and ventral fates are expanded in boz;sqt double mutants. Genotypes and markers are indicated. (A-E) In situhybridization at 35% epiboly (4.8 h) revealed that chordin(chd) mRNA levels were reduced in boz;sqt double mutants (D) and boz;sqt;cyctriple mutants (E), compared to their wild-type (A), sqt (B) and boz(C) siblings. (F-J) At 30% epiboly (4.7 h) noggin1(nog1) mRNA was notdetected in boz (H), boz;sqt double mutants (I) or boz;sqt;cyc triple mutants (J), while reduced levels were detected in sqtmutants (G).(K-N) At 80% epiboly (8.1 h), gata2expression was expanded in boz;sqt double mutants (N) compared to wild-type (K), sqt(L) and bozmutants (M). Genotypes were determined following photography (see Materials and Methods). The genotype of the embryo in C is alsosqtcz35/+and in K, bozm168/+. Animal (A-J) and lateral (K-N) views are shown. Embryos in A-D, F-I and N are progeny from bozm168/+;sqtcz35/+
intercrosses. E and J are progeny frombozm168/+;sqtcz35/+;cycm294/+intercrosses, K and M from a bozm168/+ intercross and L from a sqtcz35/+
intercross.
2588
loss of cyc enhances the midline defects of bozmutants (Fig.6E,F).
The amount of neural tissue is reduced in boz;sqt;cyc triplemutants in comparison to the double mutant combinations(Fig. 6K,L). As in sqt;cyc double mutants (Feldman et al.,2000), the trunk spinal cord is lacking in boz;sqt;cyc triple
mutants, as indicated by the absence of pax2.1 expressingspinal cord neurons (Fig. 8K) and the proximity of thehindbrain marker krox20to the tail (Fig. 8L).
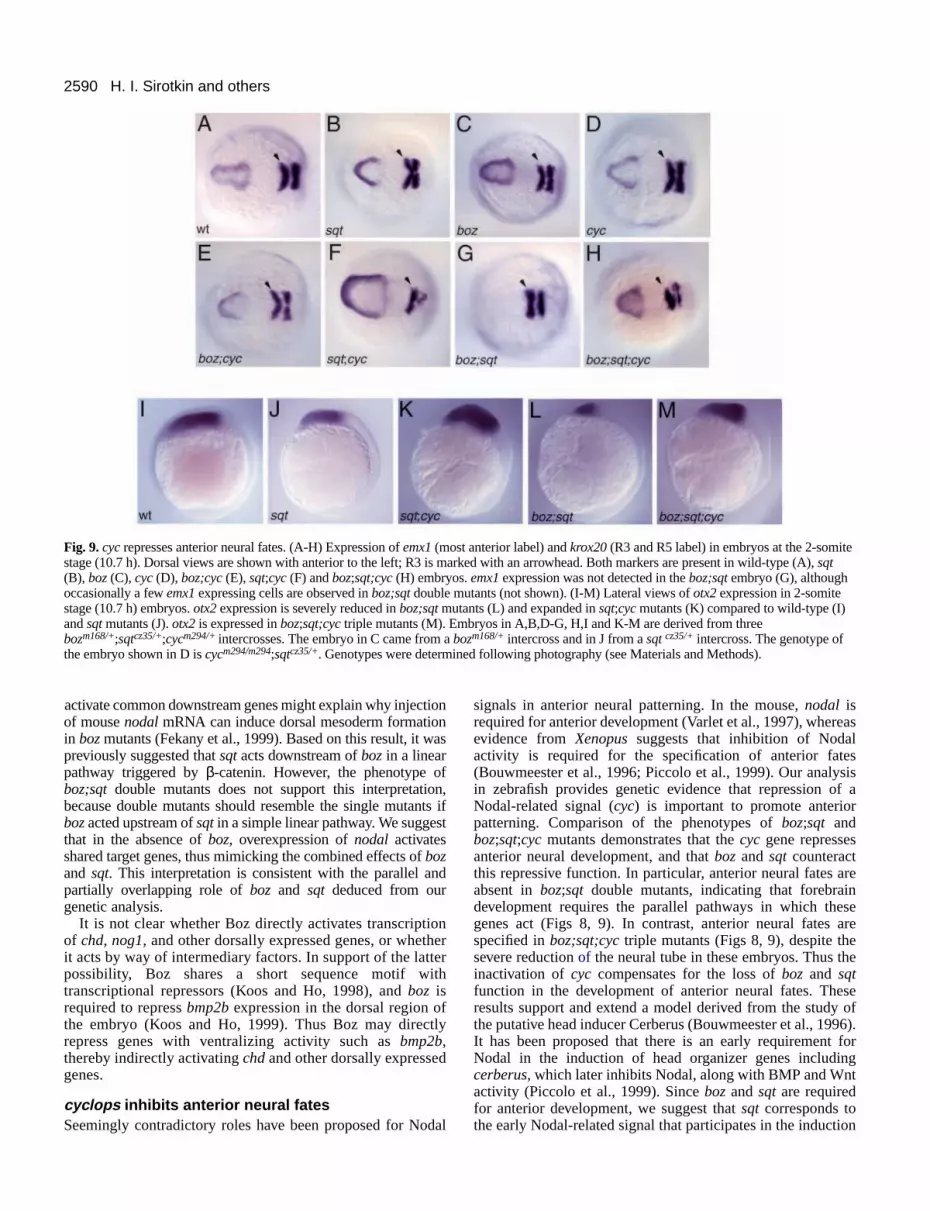
Strikingly, anterior neural fates are present in boz;sqt;cycmutants despite the lack of the most anterior neuroectoderm inboz;sqt siblings. This is evident from examination of theweakest boz;sqt;cycmutants, which clearly contain an eye withlens (Fig. 6L), while eyes were never observed in boz;sqtmutants (Fig. 6I). Likewise, expression ofpax6.1 is greatlyreduced or absent in boz;sqt embryos (Fig. 8D) but present inboz;sqt;cycmutants at 24 h (Fig. 8J). To determine whether thelack of eyes is due to an early failure to specify anterior neuralidentities or to a later deterioration of the neural plate, weexamined the expression of two anterior neural plate markers,emx1and otx2 in 2-somite-stage embryos (Fig. 9). emx1 isgreatly reduced or not detectable in boz;sqt double mutants(Fig. 9G), while boz;sqt;cyc triple mutants characteristicallyexpress emx1at a comparable level to wild type (Fig. 9H).Similarly, otx2 is strongly reduced at the 2-somite-stage inboz;sqt embryos (Fig. 9L), but is present in boz;sqt;cycmutants(Fig. 9M). A similar repressive effect of the cyc gene on theanterior neural plate is noted upon comparison of sqtsingle andsqt;cycdouble mutant phenotypes: the expression of emx1andotx2 is expanded in sqt;cyc double mutants compared to sqtsingle mutants (Fig. 9). In contrast, boz, cyc and boz;cycmutants have emx1 expression that is similar to wild type
H. I. Sirotkin and others
Fig. 6. Genetic interactions among boz, sqtand cyc. Lateral views oflive embryos with the indicated genotypes at 28-30 h. Arrowsindicate notochord (A,B,D,F) or notochord rudiment (E). Twoexamples of genotypes with variable phenotypes are shown.(A) Wild type. (B)cycmutant. (C,D)sqtmutants have a variablephenotype; some are cyclopic with a reduced or absent notochord(C), but often sqt mutants have a wild-type notochord and wellseparated eyes (D). (E,F) Thebozmutant phenotype is also variable.Some bozmutants are cyclopic with a reduced or absent notochord(E), but often bozmutants have a wild-type notochord and wellseparated eyes (F). (G)boz;cyc double mutant. (H) sqt;cycdoublemutant; arrowhead indicates large cyclopic eye. (I,J) Severe and lesssevere boz;sqt double mutants. Eyes are never observed but there issome variability in the extent of the neural tube. White arrowheadsindicate position of the otic vesicle. (K,L) severe and less severeboz;sqt;cyctriple mutants. No eye is present in K but a smallcyclopic eye is evident in L (arrowhead). Genotypes were determinedfollowing photography by PCR (see Materials and Methods), exceptfor cycand sqt;cycmutants, which were scored morphologically.
Fig. 7. Dorsal mesodermal derivatives are absent in boz;sqtdouble mutants. Dorsal (A-D) and lateral (E-H) views areshown. Comparison of expression pattern of markers ofdorsal mesodermal derivatives by in situ hybridization in(A,C,E,G) wild-type and (B,D,F,H) boz;sqt double mutantembryos at (A-F) 80% epiboly (8.1 h) or (G,H) bud stage(10 h). Probes and genotypes are indicated. Axialmesoderm expression of axial and cycwas not detected inboz;sqt double mutants (B,D). Note that the punctatestaining of the endoderm is comparable in wild-type (A)and boz;sqt double mutants (B). The prechordal plate isabsent in boz;sqt double mutants, as revealed by gsc(F)and hgg1(H) staining. Genotypes were determinedfollowing photography (see Materials and Methods). Thewild-type embryos in A and E are bozm168/+;sqtcz35/+, andthe embryos in C and G are sqtcz35/+.

2589boz and sqt in dorsal axial development
(Fig. 9). Taken together, comparison of different mutantcombinations provides genetic evidence that the cycgene actsto inhibit anterior neural development and that the anteriortruncations in boz;sqtdouble mutants can be suppressed by theelimination of cyc function.
DISCUSSION
boz and sqt act in parallel pathways required for thespecification of dorsal mesoderm and anteriorneural fatesThe functions ofbozand sqt are required for early events indorsal mesoderm development in zebrafish (Solnica-Krezel etal., 1996; Heisenberg and Nüsslein-Volhard, 1997; Feldman etal., 1998; Koos and Ho, 1998; Yamanaka et al., 1998; Fekanyet al., 1999). As the regulatory interactions between thesegenes were unclear, we performed a genetic analysis to studytheir role in dorsal development and neural patterning. Threelines of evidence indicate that bozand sqt act in parallel andhave overlapping roles in the development of dorsal mesodermand anterior neuroectoderm. First, boz requires Nodal-relatedsignals to induce dorsal mesoderm, because the ability of bozoverexpression to activate ectopic gscexpression is reduced insqt and eliminated in sqt;cyc mutants (Fig. 1). Second, sqtexpression at the late blastula stage is not reduced in bozmutants (Fig. 2), demonstrating that boz is not an essentialearly regulator of sqt transcription. Conversely, bozexpressionin the blastula is not affected in sqtor sqt;cycmutants (Fig. 2).Third, whereas boz and sqt single mutants have variable
phenotypes, and some homozygous bozand sqtsingle mutantssurvive to adulthood, boz;sqtdouble mutants have a strong andfully penetrant loss of axial mesoderm and anterior structures.These results demonstrate that expression of sqt and boz isactivated in parallel and exclude models in which sqtfunctionsdownstream of boz in a simple linear pathway.
Our analysis suggests that the boz and sqt pathwaysconverge on a common target, the secreted BMP antagonistChordin. sqt is a potent inducer of chordin expression(Rebagliati et al., 1998b; Fig. 4), but chordinmRNA levels arenot reduced in sqt or sqt;cyc mutants at blastula stages.Similarly, boz function is not required for chordin expressionin the late blastula (Fig. 3), despite the fact that boz activateschordin expression (Fig. 4; Koos and Ho, 1998). In boz;sqtdouble mutants, chordin expression is markedly reduced.These results indicate that sqtand bozhave redundant roles inmaintaining normal chordinexpression levels.
chordinis not the only dorsalizing factor that is a target of theboz and sqtpathways. noggin1expression is greatly reduced inbozand sqtsingle mutants, and in boz;sqtdouble and boz;sqt;cyctriple mutants (Fig. 5). These results indicate that both sqt andboz are required for noggin1 expression, whereas chordinexpression requires either boz or sqt. Thus it seems that thedorsalizing activities of bozand sqtare mediated, at least in part,by the activation of BMP antagonists. In addition, inhibition ofWnt signals may be an important consequence of bozand sqtaction, because expression of dkk1, which encodes a secretedWnt antagonist with potent head-inducing activity, is reduced ingastrula-stage bozand sqtmutants (Glinka et al., 1998; Niehrs,1999; Hashimoto et al., 2000). The finding that boz and sqt
Fig. 8. Analysis of neural markers in boz;sqt,sqt;cycand boz;sqt;cyc mutants. Geneexpression was examined by whole-mount insitu hybridization of (A-C) wild-type, (D-F)boz;sqt, (G-I) sqt;cyc and (J-L) boz;sqt;cycembryos at 24 h. Lateral views are shown.(A,D,G,J) Expression of pax6.1, which isabsent or severely reduced in boz;sqt mutants(D). (B,E,H,K)pax2.1marks the optic stalk,mid-hindbrain border (arrowhead) andhindbrain in wild-type (B) and sqt;cycembryos (H). The optic stalk is absent inboz;sqt double mutants (E) and the mid-hindbrain boundary appears to be at the mostanterior part of the neural tube. In boz;sqt;cycmutants (K), the optic stalk expression isabsent but the neural tube extends anterior tothe mid-hindbrain junction. (C,F,I,L)krox20marks hindbrain rhombomeres R3 and R5 inwild-type (C), boz;sqt(F), sqt;cyc (I) andboz;sqt;cyc(L) embryos.
2590
activate common downstream genes might explain why injectionof mouse nodalmRNA can induce dorsal mesoderm formationin bozmutants (Fekany et al., 1999). Based on this result, it waspreviously suggested that sqt acts downstream of bozin a linearpathway triggered by β-catenin. However, the phenotype ofboz;sqt double mutants does not support this interpretation,because double mutants should resemble the single mutants ifboz acted upstream of sqt in a simple linear pathway. We suggestthat in the absence of boz, overexpression ofnodal activatesshared target genes, thus mimicking the combined effects of bozand sqt. This interpretation is consistent with the parallel andpartially overlapping role of boz and sqt deduced from ourgenetic analysis.
It is not clear whether Boz directly activates transcriptionof chd, nog1, and other dorsally expressed genes, or whetherit acts by way of intermediary factors. In support of the latterpossibility, Boz shares a short sequence motif withtranscriptional repressors (Koos and Ho, 1998), and boz isrequired to repress bmp2bexpression in the dorsal region ofthe embryo (Koos and Ho, 1999). Thus Boz may directlyrepress genes with ventralizing activity such as bmp2b,thereby indirectly activating chdand other dorsally expressedgenes.
cyclops inhibits anterior neural fatesSeemingly contradictory roles have been proposed for Nodal
signals in anterior neural patterning. In the mouse, nodal isrequired for anterior development (Varlet et al., 1997), whereasevidence from Xenopussuggests that inhibition of Nodalactivity is required for the specification of anterior fates(Bouwmeester et al., 1996; Piccolo et al., 1999). Our analysisin zebrafish provides genetic evidence that repression of aNodal-related signal (cyc) is important to promote anteriorpatterning. Comparison of the phenotypes of boz;sqt andboz;sqt;cyc mutants demonstrates that the cyc gene repressesanterior neural development, and that boz and sqt counteractthis repressive function. In particular, anterior neural fates areabsent in boz;sqt double mutants, indicating that forebraindevelopment requires the parallel pathways in which thesegenes act (Figs 8, 9). In contrast, anterior neural fates arespecified in boz;sqt;cyctriple mutants (Figs 8, 9), despite thesevere reduction of the neural tube in these embryos. Thus theinactivation of cyc compensates for the loss of boz and sqtfunction in the development of anterior neural fates. Theseresults support and extend a model derived from the study ofthe putative head inducer Cerberus (Bouwmeester et al., 1996).It has been proposed that there is an early requirement forNodal in the induction of head organizer genes includingcerberus,which later inhibits Nodal, along with BMP and Wntactivity (Piccolo et al., 1999). Since bozand sqt are requiredfor anterior development, we suggest that sqt corresponds tothe early Nodal-related signal that participates in the induction
H. I. Sirotkin and others
Fig. 9. cyc represses anterior neural fates. (A-H) Expression of emx1(most anterior label) and krox20(R3 and R5 label) in embryos at the 2-somitestage (10.7 h). Dorsal views are shown with anterior to the left; R3 is marked with an arrowhead. Both markers are present in wild-type (A),sqt(B), boz(C), cyc(D), boz;cyc(E), sqt;cyc(F) and boz;sqt;cyc(H) embryos. emx1expression was not detected in the boz;sqt embryo (G), althoughoccasionally a few emx1expressing cells are observed in boz;sqt double mutants (not shown). (I-M) Lateral views of otx2expression in 2-somitestage (10.7 h) embryos. otx2expression is severely reduced in boz;sqt mutants (L) and expanded in sqt;cyc mutants (K) compared to wild-type (I)and sqtmutants (J). otx2is expressed in boz;sqt;cyctriple mutants (M). Embryos in A,B,D-G, H,I and K-M are derived from threebozm168/+;sqtcz35/+;cycm294/+intercrosses. The embryo in C came from a bozm168/+ intercross and in J from a sqt cz35/+ intercross. The genotype ofthe embryo shown in D is cycm294/m294;sqtcz35/+. Genotypes were determined following photography (see Materials and Methods).

2591boz and sqt in dorsal axial development
of the head organizer. In contrast, the later activity of Cycrepresses anterior development, and inhibition of this functionallows specification of anterior fates. cyc mRNA is notexpressed in boz;sqt mutants at 80% epiboly (Fig. 7D),suggesting that the cyc gene acts to repress anteriordevelopment prior to midgastrulation.
If the cycgene acted to repress the neural inducers chdandnog1, this could explain the restoration in boz;sqt;cyc triplemutants of anterior neural territories that are absent in boz;sqtdouble mutants. However, our results do not support thispossibility, because these BMP antagonists are not expressedin boz;sqt;cyc triple mutants at higher levels than in boz;sqtdouble mutants at the late blastula stage (Fig. 5). An alternativeexplanation derives from the finding that nonaxial mesodermemits diffusible signals that induce posterior neural identities(Woo and Fraser, 1997). Because axial mesoderm is absent inboz;sqt mutants (Fig. 7), and ventral fates are expanded (Fig.5), it is possible that increased activity of these posteriorizingsignals accounts for the loss of anterior neural fates in boz;sqtmutants. According to this view, anterior fates are restored inthe triple mutant by the reduction in the activity of theposteriorizing signals that results from the diminution ofmesoderm in sqt;cycand boz;sqt;cycmutants.
In summary, our results suggest that the homeobox gene bozand the nodal-related gene sqt act in a complex regulatorynetwork that controls dorsal development in zebrafish. Thesegenes act in parallel to activate the BMP antagonist Chordin,and both genes are required for noggin1expression. bozandsqt also activate pathways that block Nodal signaling, thusallowing the development of anterior neural fates.
We thank the members of the Talbot and Schier laboratories forhelpful discussions and Rory Feeney, Lauren Jow and MicheleMittman for fish care. H.I.S. is supported by postdoctoral fellowshipF32 HD08420 from the NIH. S.T.D. is supported by a fellowship fromthe ACS. This work was supported by NIH grants GM57825 (W.S.T.)and GM56211 (A.F.S.). A.F.S. is a Scholar of the McKnightEndowment Fund for Neuroscience. W.S.T. is a Pew Scholar in theBiomedical Sciences.
REFERENCES
Behrens, J., von Kries, J. P., Kuhl, M., Bruhn, L., Wedlich, D., Grosschedl,R. and Birchmeier, W. (1996). Functional interaction of beta-catenin withthe transcription factor LEF-1. Nature382, 638-642.
Bouwmeester, T., Kim, S., Sasai, Y., Lu, B. and De Robertis, E. M. (1996).Cerberus is a head-inducing secreted factor expressed in the anteriorendoderm of Spemann’s organizer. Nature382, 595-601.
Dale, L., Howes, G., Price, B. M. and Smith, J. C. (1992). Bonemorphogenetic protein 4: a ventralizing factor in early Xenopusdevelopment. Development115, 573-585.
Dick, A., Hild, M., Bauer, H., Imai, Y., Maifeld, H., Schier, A. F., Talbot,W. S., Bouwmeester, T. and Hammerschmidt, M.(2000). Essential roleof Bmp7 (snailhouse) and its prodomain in dorsoventral patterning of thezebrafish embryo. Development127, 343-354.
Erter, C. E., Solnica-Krezel, L. and Wright, C. V. (1998). Zebrafish nodal-related 2 encodes an early mesendodermal inducer signaling from theextraembryonic yolk syncytial layer. Dev. Biol.204, 361-372.
Fagotto, F., Guger, K. and Gumbiner, B. M. (1997). Induction of the primarydorsalizing center in Xenopus by the Wnt/GSK/beta-catenin signalingpathway, but not by Vg1, Activin or Noggin. Development124, 453-460.
Fekany, K., Yamanaka, Y., Leung, T., Sirotkin, H. I., Topczewski, J.,Gates, M. A., Hibi, M., Renucci, A., Stemple, D., Radbill, A., Schier, A.F., Driever, W., Hirano, T., Talbot, W. S. and Solnica-Krezel, L. (1999).The zebrafish bozozok locus encodes Dharma, a homeodomain protein
essential for induction of gastrula organizer and dorsoanterior embryonicstructures. Development126, 1427-1438.
Feldman, B., Gates, M. A., Egan, E. S., Dougan, S. T., Rennebeck, G.,Sirotkin, H. I., Schier, A. F. and Talbot, W. S. (1998). Zebrafish organizerdevelopment and germ-layer formation require nodal-related signals. Nature395, 181-185.
Feldman, B., Dougan, S. T., Schier, A. F. and Talbot, W. S. (2000). Nodal-related signals establish mesendodermal fate and trunk neural identity inzebrafish. Curr. Biol. 10, 531-534.
Furthauer, M., Thisse, B. and Thisse, C. (1999). Three different noggingenes antagonize the activity of bone morphogenetic proteins in thezebrafish embryo. Dev. Biol.214, 181-196.
Gates, M. A., Kim, L., Egan, E. S., Cardozo, T., Sirotkin, H. I., Dougan,S. T., Lashkari, D., Abagyan, R., Schier, A. F. and Talbot, W. S. (1999).A genetic linkage map for zebrafish: comparative analysis and localizationof genes and expressed sequences. Genome Res.9, 334-347.
Glinka, A., Wu, W., Delius, H., Monaghan A. P., Blumenstock, C. andNiehrs, C. (1998). Dickkopf-1 is a member of a new family of secretedproteins and functions in head induction. Nature391, 357-362.
Harland, R. and Gerhart, J. (1997). Formation and function of Spemann’sorganizer. Annu. Rev. Cell Dev. Biol.13, 611-667.
Hashimoto, H., Itoh, M., Yamanaka, Y., Yamashita, S., Shimizu, T.,Solnica-Krezel, L., Hibi, M. and Hirano, T. (2000). Zebrafish Dkk1functions in forebrain specification and axial mesendoderm formation. Dev.Biol. 217, 138-152.
Heasman, J., Crawford, A., Goldstone, K., Garner-Hamrick, P.,Gumbiner, B., McCrea, P., Kintner, C., Noro, C. Y. and Wylie, C.(1994). Overexpression of cadherins and underexpression of beta-catenininhibit dorsal mesoderm induction in early Xenopus embryos. Cell 79,791-803.
Heisenberg, C. P. and Nüsslein-Volhard, C. (1997). The function ofsilberblick in the positioning of the eye anlage in the zebrafish embryo. Dev.Biol. 184, 85-94.
Hemmati-Brivanlou, A. and Thomsen, G. H. (1995). Ventral mesodermalpatterning in Xenopus embryos: expression patterns and activities of BMP-2 and BMP-4. Dev. Genet.17, 78-89.
Hemmati-Brivanlou, A., Kelly, O. G. and Melton, D. A. (1994). Follistatin,an antagonist of activin, is expressed in the Spemann organizer and displaysdirect neuralizing activity. Cell 77, 283-285.
Jones, C. M., Lyons, K. M., Lapan, P. M., Wright, C. V. and Hogan, B. L.(1992). DVR-4 (bone morphogenetic protein-4) as a posterior-ventralizingfactor in Xenopus mesoderm induction. Development115, 639-647.
Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B. and Schilling,T. F. (1995). Stages of embryonic development of the zebrafish. Dev. Dyn.203, 253-310.
Kishimoto, Y., Lee, K. H., Zon, L., Hammerschmidt, M. and Schulte-Merker, S. (1997). The molecular nature of zebrafish swirl: BMP2 functionis essential during early dorsoventral patterning. Development124, 4457-466.
Koos, D. S. and Ho, R. K. (1998). The nieuwkoid gene characterizes andmediates a Nieuwkoop-center-like activity in the zebrafish. Curr. Biol. 8,1199-1206.
Koos, D. S. and Ho, R. K. (1999). The nieuwkoid/dharma homeobox gene isessential for bmp2b repression in the zebrafish pregastrula. Dev. Biol.215,190-207.
Krauss, S., Johansen, T., Korzh, V., Moens, U., Ericson, J. U. and Fjose,A. (1991a). Zebrafish pax[zf-a]: a paired box-containing gene expressed inthe neural tube. EMBO J. 10, 3609-3619.
Krauss, S., Johansen, T., Korzh, V. and Fjose, A. (1991b). Expression of thezebrafish paired box gene pax[zf-b] during early neurogenesis. Development113, 1193-1206.
Larabell, C. A., Torres, M., Rowning, B. A., Yost, C., Miller, J. R., Wu,M., Kimelman, D. and Moon, R. T. (1997). Establishment of thedorsoventral axis in Xenopus embryos is presaged by early asymmetries inbeta-catenin that are modulated by the Wnt signaling pathway. J. Cell Biol.136, 1123-1136.
Long, W. L. (1983). The role of the yolk syncytial layer in determination ofthe plane of bilateral symmetry in the Rainbow Trout, Salmo gairdneriRichardson. J. Exp. Zool.228, 91-97.
Miller-Bertoglio, V. E., Fisher, S., Sanchez, A., Mullins, M. C. andHalpern, M. E. (1997). Differential regulation of chordin expressiondomains in mutant zebrafish. Dev. Biol. 192, 537-550.
Mizuno, T., Yamaha, E., Wakahara, M., Kurolwa, A. and Takeda, H.(1996). Mesoderm induction in zebrafish. Nature383, 131-132.

2592
Molenaar, M., van de Wetering, M., Oosterwegel, M., Peterson-Maduro,J., Godsave, S., Korinek, V., Roose, J., Destree, O. and Clevers, H.(1996). XTcf-3 transcription factor mediates beta-catenin-induced axisformation in Xenopus embryos. Cell 86, 391-399.
Moon, R. T. and Kimelman, D. (1998). From cortical rotation to organizergene expression: toward a molecular explanation of axis specification inXenopus. BioEssays20, 536-545.
Mori, H., Miyazaki, Y., Morita, T., Nitta, H. and Mishina, M. (1994).Different spatio-temporal expressions of three otx homeoprotein transcriptsduring zebrafish embryogenesis. Brain Res. Mol. Brain Res.27, 221-231.
Morita, T., Nitta, H., Kiyama, Y., Mori, H. and Mishina, M. (1995).Differential expression of two zebrafish emx homeoprotein mRNAs in thedeveloping brain. Neurosci Lett.198, 131-134.
Nguyen, V. H., Schmid, B., Trout, J., Connors, S. A., Ekker, M. andMullins, M. C. (1998). Ventral and lateral regions of the zebrafish gastrula,including the neural crest progenitors, are established by a bmp2b/swirlpathway of genes. Dev. Biol.199, 93-110.
Niehrs, C.(1999). Head in the WNT: the molecular nature of Spemann’s headorganizer. Trends Genet. 15, 314-319.
Nieto, M. A. (1999). Reorganizing the organizer 75 years on. Cell 98, 417-425.
Nishimatsu, S. and Thomsen, G. H. (1998). Ventral mesoderm induction andpatterning by bone morphogenetic protein heterodimers in Xenopusembryos. Mech. Dev.74, 75-88.
Oxtoby, E. and Jowett, T. (1993). Cloning of the zebrafish krox-20 gene (krx-20) and its expression during hindbrain development. Nucleic Acids Res. 21,1087-1095.
Pelegri, F. and Maischein, H. M. (1998). Function of zebrafish beta-cateninand TCF-3 in dorsoventral patterning. Mech. Dev.77, 63-74.
Piccolo, S., Agius, E., Leyns, L., Bhattacharyya, S., Grunz, H.,Bouwmeester, T. and De Robertis, E. M. (1999). The head inducerCerberus is a multifunctional antagonist of Nodal, BMP and Wnt signals.Nature397, 707-710.
Piccolo, S., Sasai, Y., Lu, B. and De Robertis, E. M. (1996). Dorsoventralpatterning in Xenopus: inhibition of ventral signals by direct binding ofchordin to BMP-4. Cell 86, 589-598.
Rebagliati, M. R., Toyama, R., Haffter, P. and Dawid, I. B. (1998a). cyclopsencodes a nodal-related factor involved in midline signaling. Proc. Natl.Acad. Sci. USA95, 9932-9937.
Rebagliati, M. R., Toyama, R., Fricke, C., Haffter, P. and Dawid, I. B.(1998b). Zebrafish nodal-related genes are implicated in axial patterning andestablishing left-right asymmetry. Dev. Biol.199, 261-272.
Sampath, K., Rubinstein, A. L., Cheng, A. M., Liang, J. O., Fekany, K.,Solnica-Krezel, L., Korzh, V., Halpern, M. E. and Wright, C. V. E.(1998). Induction of the zebrafish ventral brain and floorplate requirescyclops/nodal signalling. Nature395, 185-189.
Sasai, Y., Lu, B., Steinbeisser, H., Geissert, D., Gont, L. K. and DeRobertis, E. M. (1994). Xenopus chordin: a novel dorsalizing factoractivated by organizer- specific homeobox genes. Cell 79, 779-790.
Schier, A. F., Neuhauss, S. C., Harvey, M., Malicki, J., Solnica-Krezel, L.,Stainier, D. Y., Zwartkruis, F., Abdelilah, S., Stemple, D. L., Rangini,Z., Yang, H. and Driever, W. (1996). Mutations affecting the developmentof the embryonic zebrafish brain. Development123, 165-178.
Schier, A. S. and Shen, M. M.(2000). Nodal signalling in vertebratedevelopment.Nature403, 385-389.
Schier, A. F. and Talbot, W. S. (1998). The zebrafish organizer. Curr. Opin.Genet. Dev. 8, 464-471.
Schmid, B., Furthauer, M., Connors, S. A., Trout, J., Thisse, B., Thisse, C.and Mullins, M. C. (2000). Equivalent genetic roles for bmp7/snailhouseand bmp2b/swirl in dorsoventral pattern formation. Development127, 957-967.
Schneider, S., Steinbeisser, H., Warga, R. M. and Hausen, P. (1996). Beta-catenin translocation into nuclei demarcates the dorsalizing centers in frogand fish embryos. Mech. Dev.57, 191-198.
Schulte-Merker, S., Lee, L. J., McMahon, A. P. and Hammerschmidt, M.(1997). The zebrafish organizer requires chordino. Nature387, 862-863.
Smith, W. C. and Harland, R. M. (1992). Expression cloning of noggin, anew dorsalizing factor localized to the Spemann organizer in Xenopusembryos. Cell 70, 829-840.
Solnica-Krezel, L., Stemple, D. L., Mountcastle-Shah, E., Rangini, Z.,Neuhauss, S. C., Malicki, J., Schier, A. F., Stainier, D. Y., Zwartkruis,F., Abdelilah, S. and Driever, W. (1996). Mutations affecting cell fates andcellular rearrangements during gastrulation in zebrafish. Development123,67-80.
Strähle, U., Blader, P., Henrique, D. and Ingham, P. W. (1993). Axial, azebrafish gene expressed along the developing body axis, shows alteredexpression in cyclops mutant embryos. Genes Dev.7, 1436-1446.
Thisse, C., Thisse, B., Halpern, M. E. and Postlethwait, J. H. (1994).goosecoid expression in neurectoderm and mesendoderm is disrupted inzebrafish cyclops gastrulas. Dev. Biol.164, 420-429.
Thisse, C., Thisse, B., Schilling, T. F. and Postlethwait, J. H. (1993).Structure of the zebrafish snail1 gene and its expression in wild-type,spadetail and no tail mutant embryos. Development119, 1203-1215.
Varlet, I., Collignon, J. and Robertson, E. J.(1997). nodal expression in theprimitive eododerm is required for the specification of the anterior axisduring mouse gastrulation. Development124, 1033-1044.
Wylie, C., Kofron, M., Payne, C., Anderson, R., Hosobuchi, M., Joseph,E. and Heasman, J. (1996). Maternal beta-catenin establishes a ‘dorsalsignal’ in early Xenopus embryos. Development122, 2987-2996.
Woo, K. and Fraser, S. E.(1997). Specification of the zebrafish nervoussystem by nonaxial signals. Science277, 254-257.
Yamanaka, Y., Mizuno, T., Sasai, Y., Kishi, M., Takeda, H., Kim, C. H.,Hibi, M. and Hirano, T. (1998). A novel homeobox gene, dharma, caninduce the organizer in a non-cell-autonomous manner. Genes Dev.12,2345-2353.
Zimmerman, L. B., De Jesus-Escobar, J. M. and Harland, R. M. (1996).The Spemann organizer signal noggin binds and inactivates bonemorphogenetic protein 4. Cell 86, 599-606.
H. I. Sirotkin and others





























