Embed Size (px)
Citation preview

R E S EA RCH AR T I C L E
Biofilm-growing intestinal anaerobic bacteria
Gianfranco Donelli1, Claudia Vuotto1, Rita Cardines2 & Paola Mastrantonio2
1Microbial Biofilm Laboratory, IRCCS Fondazione Santa Lucia, Rome, Italy; and 2Department of Infectious, Parasitic and Immune-mediated
Diseases, Istituto Superiore di Sanita, Rome, Italy
Correspondence: Gianfranco Donelli,
Microbial Biofilm Laboratory (LABIM), IRCCS
Fondazione Santa Lucia, Via Ardeatina 306,
00179 Rome, Italy. Tel.: +39 06 51501305/
+39 06 516501307; fax: +39 06 51501306;
e-mails: [email protected] or
Received 14 October 2011; revised 1 March
2012; accepted 12 March 2012.
Final version published online 23 April 2012.
DOI: 10.1111/j.1574-695X.2012.00962.x
Editor: Thomas Bjarnsholt
Keywords
biofilm; adherence; anaerobes; intestine.
Abstract
Sessile growth of anaerobic bacteria from the human intestinal tract has been
poorly investigated, so far. We recently reported data on the close association
existing between biliary stent clogging and polymicrobial biofilm development
in its lumen. By exploiting the explanted stents as a rich source of anaerobic
bacterial strains belonging to the genera Bacteroides, Clostridium, Fusobacteri-
um, Finegoldia, Prevotella, and Veillonella, the present study focused on their
ability to adhere, to grow in sessile mode and to form in vitro mono- or dual-
species biofilms. Experiments on dual-species biofilm formation were planned
on the basis of the anaerobic strains isolated from each clogged biliary stent,
by selecting those in which a couple of anaerobic strains belonging to different
species contributed to the polymicrobial biofilm development. Then, strains
were investigated by field emission scanning electron microscopy and confocal
laser scanning microscopy to reveal if they are able to grow as mono- and/or
dual-species biofilms. As far as we know, this is the first report on the ability
to adhere and form mono/dual-species biofilms exhibited by strains belonging
to the species Bacteroides oralis, Clostridium difficile, Clostridium baratii, Clos-
tridium fallax, Clostridium bifermentans, Finegoldia magna, and Fusobacterium
necrophorum.
Introduction
Anaerobes contribute to form the largest and most diver-
sified microbial community of the human body, that is,
that of the gastrointestinal tract, exceeding by 2–4 log
units the aerobic flora. In the last years, metagenomic
experiments have shown that the vast majority of intesti-
nal bacteria belong to the phyla Firmicutes and Bacteroi-
detes (Fakhry et al., 2009). As the small intestine is
concerned, bacterial concentration ranges from 105 to 109
bacteria per gram of intestinal content and anaerobes are
mainly represented by Bacteroides, Bifidobacterium, Clos-
tridium, Finegoldia (formerly Peptostreptococcus), Fusobac-
terium, Prevotella, and Veillonella species (Berg, 1996).
The predominant anaerobes are represented by Bactero-
ides species that are bile-resistant, non-spore-forming,
Gram-negative rods (Wexler, 2007). It has been reported
that Bacteroides fragilis exhibits a high tolerance to bile
salts as well as a great ability to utilize a broad spectrum
of polysaccharides and to vary surface antigens to evade
host immune responses; all these features presumably
explain its persistence in high numbers in the intestine
(Pumbwe et al., 2007). Bifidobacteria are non-spore-form-
ing, filamentous Gram-positive anaerobic rods that inha-
bit the gastrointestinal tract and the vaginal mucosa.
Recent studies have suggested that their interaction with
intestinal epithelial cells is effective on the integrity of
mucosa that is protected from inflammation presumably
by metabolites produced by these anaerobes (Amit-
Romach et al., 2010). Clostridia are spore-forming, micro-
aerophilic Gram-positive rods, and the species Clostridium
baratii, Clostridium bifermentans, and Clostridium perfrin-
gens are frequently isolated at the intestinal level while the
sporadically occurring microorganism Clostridium difficile
is known to overgrow in the intestine of patients treated
with broad-spectrum antibiotics (Gorbach, 1996). A
recent characterization of multidrug-resistant C. difficile
clinical isolates showed that antibiotic resistance provides
this pathogen with potential advantages over the co-resi-
dent gut flora (Spigaglia et al., 2011). Thus, C. difficile,
ª 2012 Federation of European Microbiological Societies FEMS Immunol Med Microbiol 65 (2012) 318–325Published by Blackwell Publishing Ltd. All rights reserved
IMM
UN
OLO
GY
& M
EDIC
AL
MIC
ROBI
OLO
GY

also for the recent emergence of new hypervirulent epi-
demic strains, is considered an increasingly alarming
nosocomial enteric pathogen (Bartlett, 2010). Finegoldia,
formerly known as Peptostreptococcus, is a genus consist-
ing of Gram-positive anaerobic cocci, occurring in short
chains, in pairs or as single cells. Species belonging to the
genus Finegoldia, slow-growing commensals in the mouth
and in the intestinal tract, can cause septicemia and
abscesses in the brain, lungs, and liver of immunosup-
pressed patients (Brook, 2008). The genus Fusobacterium
includes 13 species of Gram-negative, strictly anaerobic,
non-spore forming, and spear-shaped bacilli. The most
frequent isolates in clinical specimens are Fusobacterium
necrophorum and Fusobacterium nucleatum, both belong-
ing to the normal flora of the oral cavity, intestinal tract,
and vagina. Particularly, F. necrophorum accounts for
25% of the isolates from liver abscesses (Huggan & Mur-
doch, 2008).
Prevotella genus, constituted by nonmotile, Gram-nega-
tive anaerobic rods, includes 20 different species known
to contribute in causing periodontitis, abscesses, bactere-
mia, wound, and urogenital tract infections (Alauzet
et al., 2010). Veillonella genus consists of small, strictly
anaerobic Gram-negative cocci lacking of flagella, spores,
and capsule. In the human intestine, Veillonella spp. con-
tribute to dehydroxylation of bile acids and have been
suggested as causative agents of opportunistic infections
(Verma et al., 2010).
The ability of the above-mentioned anaerobic species
to form biofilm and/or co-aggregate has been rarely
reported in the intestinal tract if compared with the num-
ber of studies on the development of anaerobes as multi-
species biofilms in the oral cavity (Kolenbrander, 2011;
Marsh et al., 2011) and in several chronic infections
including chronic wounds, cystic fibrosis, and otitis media
(James et al., 2008; Burmølle et al., 2010; Thornton et al.,
2011).
Sandra MacFarlane group reported (Ahmed et al.,
2007) on the intestinal occurrence of Bacteroides and Bifi-
dobacteria as microcolonies identified by fluorescence in
situ hybridization and on their distribution throughout
the mucus layer observed by confocal laser scanning
microscopy (CLSM). Then, we reported results on the
development of a multi-species biofilm in the lumen of
clogged biliary stents as consequence of the ascending
colonization from duodenum of aerobic and anaerobic
bacteria (Guaglianone et al., 2008). More recently, How-
ard Ceri group proposed a model for culturing mucosal
anaerobic bacteria recovered from colonic biopsies to
develop multi-species biofilms (Sproule-Willoughby et al.,
2010). The identification by conventional and molecular
techniques of both culturable and nonculturable sessile-
growing bacterial and fungal species in the biliary sludge
has revealed the occurrence of anaerobes in the 57% of
the examined biliary stents and has confirmed the poly-
microbial nature of the biofilm developing in their lumen
(Guaglianone et al., 2010).
Using the clogged biliary stent as a model of multi-
species biofilm development and exploiting the large
number of explanted stents as a generous source of anaer-
obic strains belonging to the genera Bacteroides, Clostrid-
ium, Fusobacterium, Finegoldia, Prevotella, and Veillonella,
the present study investigated on their ability to adhere,
to grow in sessile mode, and to form in vitro mono- or
dual-species biofilms.
Materials and methods
Bacterial strains
The anaerobic strains, here identified by the codes
assigned when isolated from the explanted biliary stents,
are the following: C. baratii strain CbaBs33, C. bifermen-
tans strain CbiBs1, C. difficile strain CdiBs21, Clostridium
fallax strain CfaBs3, C. perfringens strain CpeBs31, Fine-
goldia magna strain FmBs21, F. magna strain FmBs12,
B. fragilis strain BfBs12, Bacteroides oralis strain BoBs32,
F. necrophorum strain FnBs4, Parabacteroides distasonis
strain PdBs7, Prevotella intermedia strain PiBs18, Veillo-
nella spp. strain VBs4.
Culture conditions
All anaerobic strains, maintained on Brucella agar supple-
mented with vitamin K (0.5 mg L�1), haemin (5 mg L�1),
and 5% defibrinated sheep red blood cells, were routinely
cultured in brain heart infusion broth (BHI) containing
the above-mentioned supplements. All bacterial cultures
were anaerobically grown at 37 °C in an anaerobic
chamber.
Quantitative biofilm production assay
Bacterial strains were grown anaerobically at 37 °C in pre-
reduced BHI broth for a time ranging from 24 to 72 h,
depending on the strain. Each well of a 96-well flat-
bottomed plastic tissue culture plate (three wells for each
strain) was filled with 20 lL of the broth culture (adjusted
to 0.5 McF) and 180 lL of fresh BHI supplemented with
1% glucose. As a control, a well with fresh BHI supple-
mented with 1% glucose without bacteria has been used.
The plate was covered with a lid and incubated anaerobi-
cally for 48 h at 37 °C. Then, the content of each well was
removed, and the wells were carefully washed three times
with 200 lL of PBS. The plate was dried for 1 h at
60 °C and stained for 5 min with 150 lL of 2% Hucker’s
FEMS Immunol Med Microbiol 65 (2012) 318–325 ª 2012 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Biofilm-growing intestinal anaerobic bacteria 319

crystal violet. Excess stain was rinsed off by rinsing the
plate under tap water, and the plate was dried for 10 min
at 60 °C. Each assay was performed in triplicate and
repeated three times. The dye bound to the adherent cells
was solubilized with 150 lL of 33% (v/v) glacial acetic
acid per well. The optical density (OD) of each well was
measured at 570 nm using a microplate photometer (Mul-
tiscan FC; Thermo Scientific). The cut-off OD (ODc) is
defined as three standard deviations above the mean OD
of the negative control. According to the defined ODc,
all the strains were classified on the basis of their adher-
ence ability into the following categories: nonadherent
(OD � ODc), weakly adherent (ODc < OD � 29ODc),
moderately adherent (2ODc < OD � 49ODc), and
strongly adherent (49ODc < OD) (Stepanovic et al., 2000).
To further investigate by field emission scanning elec-
tron microscopy (FESEM) and CLSM the ability of single
bacterial strains to form biofilm, each well of a 24-well
plastic tissue culture plate, with a 13-mm diameter glass
coverslip placed on the bottom, was filled with 200 lL of
a broth culture (adjusted to 0.5 McF) of each strain and
1.8 mL of prereduced BHI broth supplemented with 1%
glucose and incubated for 48 h at 37 °C.As the ability to grow in a mixed biofilm of the couples
of anaerobic strains isolated from biliary stents (Veillonel-
la spp. strain VBs4 + F. necrophorum strain FnBs4; B. fra-
gilis strain BfBs12 + F. magna strain FmBs12; C. difficile
strain CdiBs21 + F. magna strain FmBs21), the following
protocol was applied: 200 lL from each broth culture of
the two strains, at the same OD (adjusted to 0.5 McF),
were mixed in a well with 1.6 mL of prereduced BHI
broth supplemented with 1% glucose and incubated
anaerobically for 48 h at 37 °C. After incubation, the
content of each well was removed and the wells were
washed carefully three times with PBS.
Field emission scanning electron microscopy
(FESEM)
Bacterial biofilms, obtained as described earlier, were fixed
with 2.5% glutaraldehyde in 0.1 M cacodylate buffer (pH
7.4) at room temperature for 30 min, postfixed with 1%
OsO4 in 0.1 M phosphate buffer for 20 min and dehy-
drated through graded ethanol (30°, 50°, 70°, 85°, 95°,100°). After critical point drying in hexamethyldisilazane
and gold coating by sputtering, biofilm samples were
examined by a field emission scanning electron micro-
scope (Sigma-Zeiss) at an accelerating voltage of 5 kV.
Confocal laser scanning microscopy (CLSM)
Biofilms grown on coverslips were fixed with 3.7% para-
formaldehyde at room temperature for 30 min and
stained with the LIVE/DEAD® BacLightTM Bacterial Via-
bility Kit (Invitrogen, Molecular Probes®) by adding, in
each well of a 24-well plate, 3 lL of the dye mixture in
1 mL of distilled water for 15 min at room temperature
in the dark. The stain was aspirated, and the coverslips
was gently washed twice with distillate water. This kit
employs two nucleic acid stains differing in their ability to
penetrate healthy bacterial cells: green-fluorescent SYTO®
9 stain and red-fluorescent propidium iodide stain. When
used alone, SYTO® 9 stain labels both live and dead
bacteria. In contrast, propidium iodide penetrates only
bacteria with damaged membranes, reducing SYTO® 9
fluorescence when both dyes are present. Thus, live bacte-
ria with intact membranes fluoresce green, while dead
bacteria with damaged membranes fluoresce red. The exci-
tation/emission maxima for these dyes are about 480/
500 nm for the SYTO® 9 and 490/635 nm for propidium
iodide. Fluorescence from stained biofilms was viewed
using a CLSM (Nikon C1si), the mounted specimens were
observed using a 109 lens and the acquired images of the
biofilms were at a resolution of 512 9 512 pixels.
Results
Anaerobic strains were investigated for their ability to
adhere in vitro, and the relative results are shown in
Table 1 and Fig. 1. Among the Gram-negative anaerobic
strains tested for their quantitative biofilm production,
those belonging to the species B. fragilis, F. necrophorum,
P. intermedia, and Veillonella spp. were strongly adherent;
B. oralis was moderately adherent; and P. distasonis was
weakly adherent. As the Gram-positive anaerobic strains
are concerned, those belonging to the species C. baratii,
C. fallax, C. perfringens, C. bifermentans, and F. magna
were strongly adherent and only the C. difficile strain was
moderately adherent.
Particularly, the ability of different species to coexist in
a unique microbial community or to develop as a dual-
species biofilm was investigated by focusing our experi-
ments on the couples of anaerobic strains belonging to
different species occurring within the same biliary stent.
Then, the skill of these strains to grow together and/or
to co-aggregate in a mutualistic mode was explored by
FESEM (Figs 2–4).As the F. necrophorum strain FnBs4 + Veillonella spp.
strain VBs4-mixed biofilm is concerned, the scanning
electron microscopy analysis reveals (Fig. 2c) a thick bio-
film characterized, after 48 h of incubation, by an EPS
matrix denser than that observed in the respective mono-
species biofilms (Fig. 2a and b).
Scanning electron micrographs of the mono-species
biofilms of F. magna strain FmBs12 (Fig. 3a) and
B. fragilis strain BfBs12 (Fig. 3b) revealed, after 48-h
ª 2012 Federation of European Microbiological Societies FEMS Immunol Med Microbiol 65 (2012) 318–325Published by Blackwell Publishing Ltd. All rights reserved
320 G. Donelli et al.
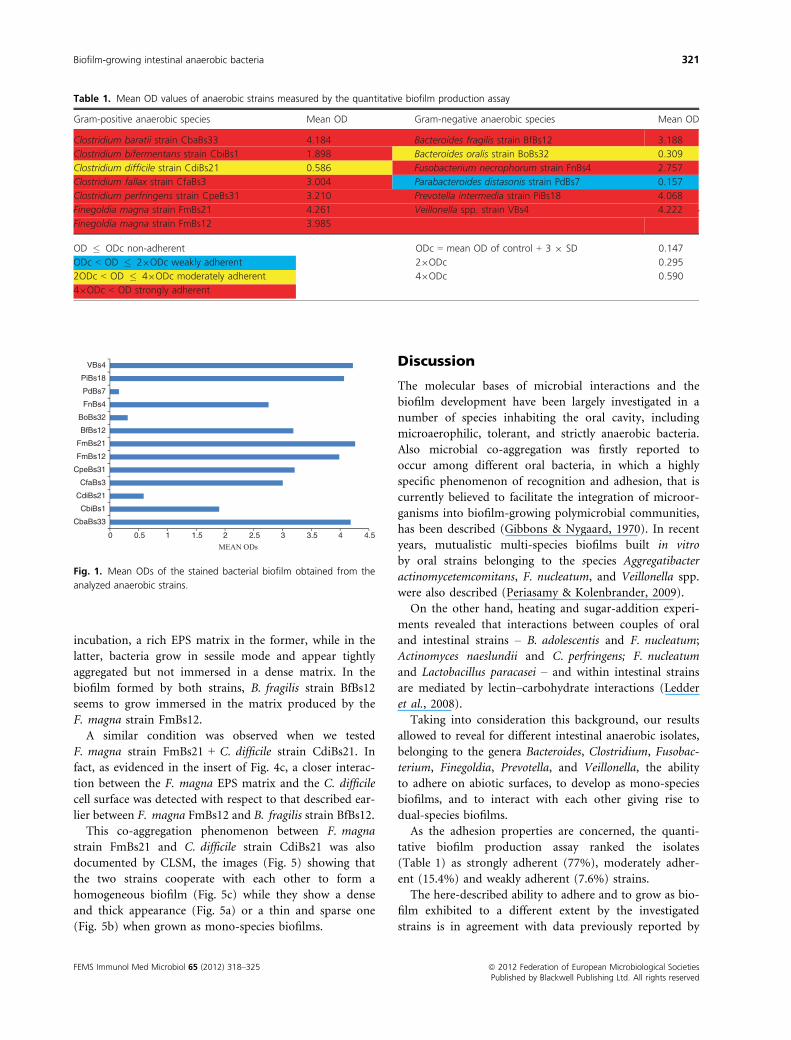
incubation, a rich EPS matrix in the former, while in the
latter, bacteria grow in sessile mode and appear tightly
aggregated but not immersed in a dense matrix. In the
biofilm formed by both strains, B. fragilis strain BfBs12
seems to grow immersed in the matrix produced by the
F. magna strain FmBs12.
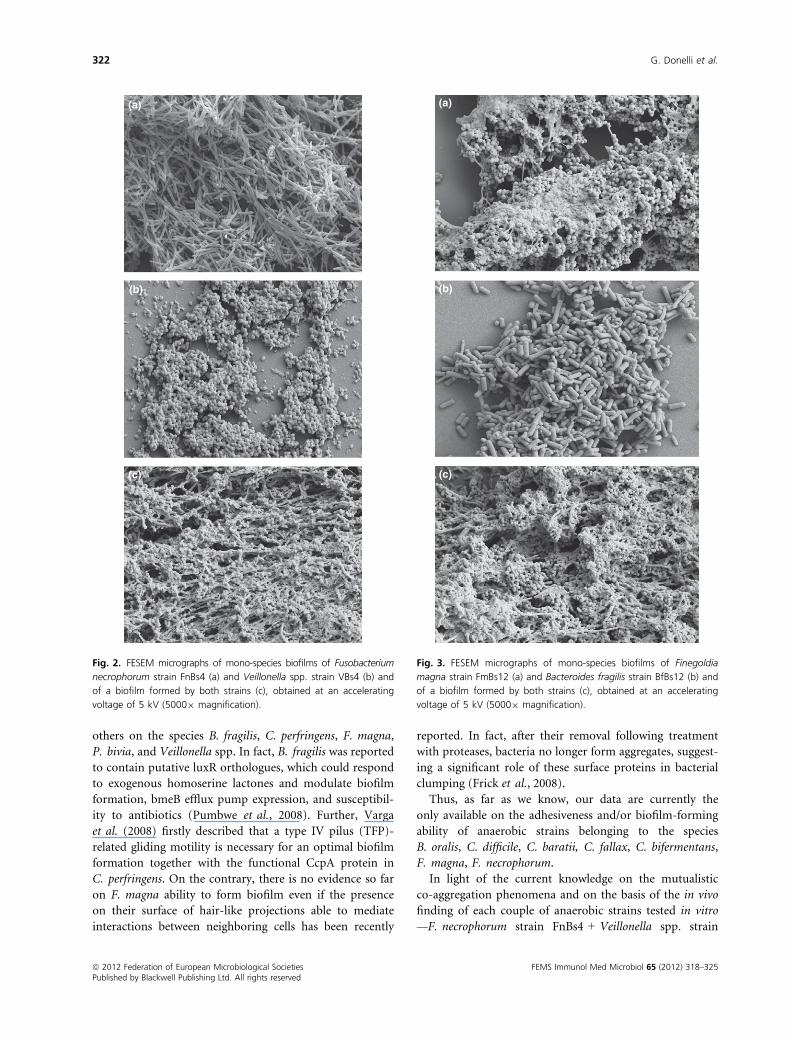
A similar condition was observed when we tested
F. magna strain FmBs21 + C. difficile strain CdiBs21. In
fact, as evidenced in the insert of Fig. 4c, a closer interac-
tion between the F. magna EPS matrix and the C. difficile
cell surface was detected with respect to that described ear-
lier between F. magna FmBs12 and B. fragilis strain BfBs12.
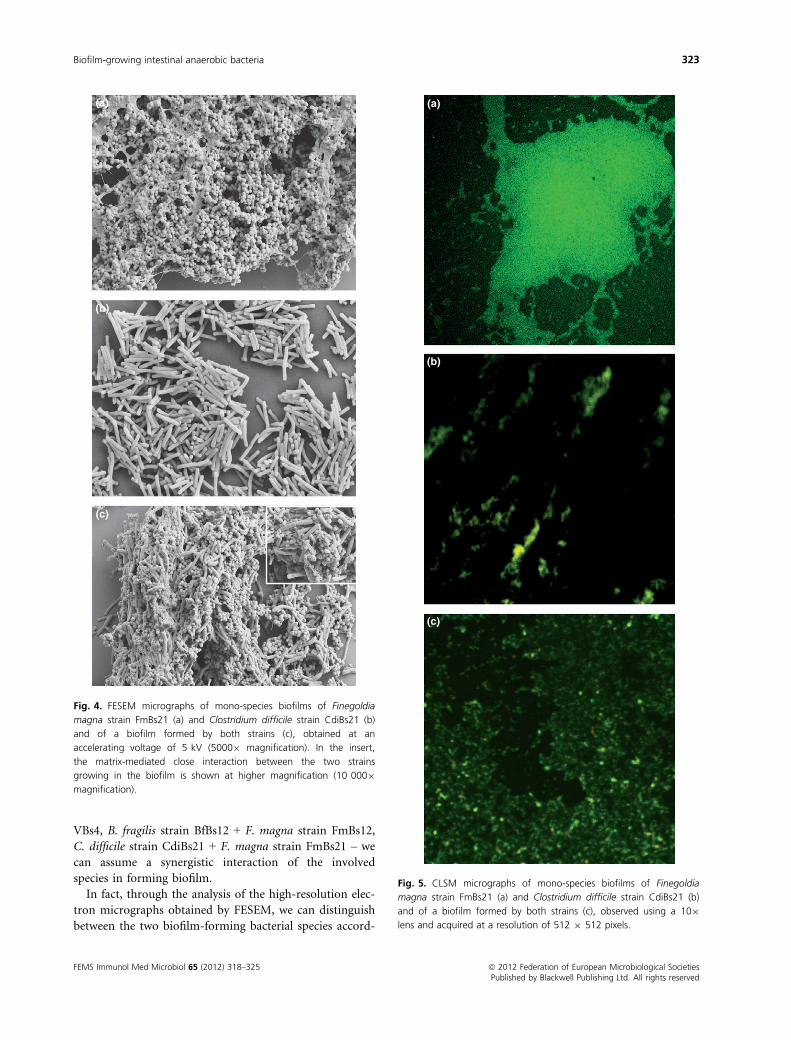
This co-aggregation phenomenon between F. magna
strain FmBs21 and C. difficile strain CdiBs21 was also
documented by CLSM, the images (Fig. 5) showing that
the two strains cooperate with each other to form a
homogeneous biofilm (Fig. 5c) while they show a dense
and thick appearance (Fig. 5a) or a thin and sparse one
(Fig. 5b) when grown as mono-species biofilms.
Discussion
The molecular bases of microbial interactions and the
biofilm development have been largely investigated in a
number of species inhabiting the oral cavity, including
microaerophilic, tolerant, and strictly anaerobic bacteria.
Also microbial co-aggregation was firstly reported to
occur among different oral bacteria, in which a highly
specific phenomenon of recognition and adhesion, that is
currently believed to facilitate the integration of microor-
ganisms into biofilm-growing polymicrobial communities,
has been described (Gibbons & Nygaard, 1970). In recent
years, mutualistic multi-species biofilms built in vitro
by oral strains belonging to the species Aggregatibacter
actinomycetemcomitans, F. nucleatum, and Veillonella spp.
were also described (Periasamy & Kolenbrander, 2009).
On the other hand, heating and sugar-addition experi-
ments revealed that interactions between couples of oral
and intestinal strains – B. adolescentis and F. nucleatum;
Actinomyces naeslundii and C. perfringens; F. nucleatum
and Lactobacillus paracasei – and within intestinal strains
are mediated by lectin–carbohydrate interactions (Ledder
et al., 2008).
Taking into consideration this background, our results
allowed to reveal for different intestinal anaerobic isolates,
belonging to the genera Bacteroides, Clostridium, Fusobac-
terium, Finegoldia, Prevotella, and Veillonella, the ability
to adhere on abiotic surfaces, to develop as mono-species
biofilms, and to interact with each other giving rise to
dual-species biofilms.
As the adhesion properties are concerned, the quanti-
tative biofilm production assay ranked the isolates
(Table 1) as strongly adherent (77%), moderately adher-
ent (15.4%) and weakly adherent (7.6%) strains.
The here-described ability to adhere and to grow as bio-
film exhibited to a different extent by the investigated
strains is in agreement with data previously reported by
MEAN ODs0 0.5 1 1.5 2 2.5 3 3.5 4 4.5
CbaBs33
CbiBs1
CdiBs21
CfaBs3
CpeBs31
FmBs12
FmBs21
BfBs12
BoBs32
FnBs4
PdBs7
PiBs18
VBs4
Fig. 1. Mean ODs of the stained bacterial biofilm obtained from the
analyzed anaerobic strains.
Table 1. Mean OD values of anaerobic strains measured by the quantitative biofilm production assay
FEMS Immunol Med Microbiol 65 (2012) 318–325 ª 2012 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Biofilm-growing intestinal anaerobic bacteria 321
others on the species B. fragilis, C. perfringens, F. magna,
P. bivia, and Veillonella spp. In fact, B. fragilis was reported
to contain putative luxR orthologues, which could respond
to exogenous homoserine lactones and modulate biofilm
formation, bmeB efflux pump expression, and susceptibil-
ity to antibiotics (Pumbwe et al., 2008). Further, Varga
et al. (2008) firstly described that a type IV pilus (TFP)-
related gliding motility is necessary for an optimal biofilm
formation together with the functional CcpA protein in
C. perfringens. On the contrary, there is no evidence so far
on F. magna ability to form biofilm even if the presence
on their surface of hair-like projections able to mediate
interactions between neighboring cells has been recently
reported. In fact, after their removal following treatment
with proteases, bacteria no longer form aggregates, suggest-
ing a significant role of these surface proteins in bacterial
clumping (Frick et al., 2008).
Thus, as far as we know, our data are currently the
only available on the adhesiveness and/or biofilm-forming
ability of anaerobic strains belonging to the species
B. oralis, C. difficile, C. baratii, C. fallax, C. bifermentans,
F. magna, F. necrophorum.
In light of the current knowledge on the mutualistic
co-aggregation phenomena and on the basis of the in vivo
finding of each couple of anaerobic strains tested in vitro
—F. necrophorum strain FnBs4 + Veillonella spp. strain
(a)
(b)
(c)
Fig. 2. FESEM micrographs of mono-species biofilms of Fusobacterium
necrophorum strain FnBs4 (a) and Veillonella spp. strain VBs4 (b) and
of a biofilm formed by both strains (c), obtained at an accelerating
voltage of 5 kV (50009 magnification).
(a)
(b)
(c)
Fig. 3. FESEM micrographs of mono-species biofilms of Finegoldia
magna strain FmBs12 (a) and Bacteroides fragilis strain BfBs12 (b) and
of a biofilm formed by both strains (c), obtained at an accelerating
voltage of 5 kV (50009 magnification).
ª 2012 Federation of European Microbiological Societies FEMS Immunol Med Microbiol 65 (2012) 318–325Published by Blackwell Publishing Ltd. All rights reserved
322 G. Donelli et al.
VBs4, B. fragilis strain BfBs12 + F. magna strain FmBs12,
C. difficile strain CdiBs21 + F. magna strain FmBs21 – we
can assume a synergistic interaction of the involved
species in forming biofilm.
In fact, through the analysis of the high-resolution elec-
tron micrographs obtained by FESEM, we can distinguish
between the two biofilm-forming bacterial species accord-
(a)
(b)
(c)
Fig. 4. FESEM micrographs of mono-species biofilms of Finegoldia
magna strain FmBs21 (a) and Clostridium difficile strain CdiBs21 (b)
and of a biofilm formed by both strains (c), obtained at an
accelerating voltage of 5 kV (50009 magnification). In the insert,
the matrix-mediated close interaction between the two strains
growing in the biofilm is shown at higher magnification (10 0009
magnification).
(a)
(b)
(c)
Fig. 5. CLSM micrographs of mono-species biofilms of Finegoldia
magna strain FmBs21 (a) and Clostridium difficile strain CdiBs21 (b)
and of a biofilm formed by both strains (c), observed using a 109
lens and acquired at a resolution of 512 9 512 pixels.
FEMS Immunol Med Microbiol 65 (2012) 318–325 ª 2012 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Biofilm-growing intestinal anaerobic bacteria 323

ing to their highly different features (rod- or spear-shaped
bacilli vs. cocci). This morphological approach allows
evaluating the more or less balanced presence of each
species in the mixed biofilm and their skill to interact
with each other.
Particularly, a mature biofilm, exhibiting a ‘common’
EPS matrix denser than that produced by Veillonella spp
strain VBs4 alone, was developed within 48 h by F. necro-
phorum strain FnBs4 + Veillonella spp. strain VBs4.
On the other hand, our data on F. necrophorum strain
FnBs4 biofilm seem to confirm that the production of an
extra-cellular polysaccharide matrix is not an intrinsic
feature of the species belonging to the genus Fusobacteri-
um, as already known for F. nucleatum, nevertheless
reported to be able to co-adhere and form biofilm (Zilm
& Rogers, 2007).
Anyway, a synergy between Fusobacterium and Veillo-
nella in developing a dual-species biofilm is presumably
present. In fact, as it is already known, Fusobacterium has
strong adhesive properties because of the presence of lec-
tins that mediate not only the adhesion to epithelia but
also the co-agglutination with other bacteria (Roberts,
2000) by playing its pivotal role of ‘bridging microorgan-
ism’ in inter-species adherence and multi-species oral
biofilm (Kaplan et al., 2009). On the other hand, Veillo-
nella is not able to catabolize sugars, so its growth
depends on acetic, propionic, butyric, and lactic acids
provided by Fusobacterium.
As the interaction between the FESEM investigated
strains of B. fragilis and F. magna is concerned, a syner-
gistic co-aggregation can be hypothesized on the basis of
the largely increased production of EPS matrix in the
mixed biofilm.
On the basis of data obtained by FESEM and CLSM
investigations, the sticky biofilm constituted by C. difficile
strain CdiBs21 + F. magna strain FmBs12 appears quite
different from the thin and sparse biofilm developed by
C. difficile alone or the dense and thick one exhibited by
F. magna. Thus, according to our data, two different
hypotheses can be taken into consideration: (1) C. difficile
strain CdiBs21 coexists in biofilm growing together with
the strong biofilm-producer F. magna strain FmBs12; or
(2) C. difficile strain CdiBs21, classified by the quantita-
tive biofilm production assay as a weakly adherent strain,
became able to grow as biofilm as consequence of a close
interaction with the F. magna, direct or mediated by
a biofilm-promoting substance released by the latter. It
should be considered that both the coexistence and the
possibly induced growth as biofilm of C. difficile could
protect the microorganism from the action of antimicro-
bial drugs thus causing the failure of the targeted anti-
biotic therapies. In fact, our findings could well explain
why C. difficile relapses occur in 15–20% of CDI patients.
As a whole, our data suggest the possibility that non- or
weak biofilm-producing bacteria could benefit from living
in biofilms developed by other strong biofilm-forming
species.
According to our knowledge, this is the first report on
adherence and/or biofilm formation displayed by strains
belonging to the anaerobic species B. oralis, C. difficile,
C. baratii, C. fallax, C. bifermentans, F. magna, F. necro-
phorum.
Our intention is to continue in studying the mecha-
nisms of mono- and mixed-biofilms formation of anaero-
bic bacteria to elucidate their scarcely investigated role in
a number of acute or chronic severe infections in
humans.
Acknowledgements
The authors are indebted to Dr Fabrizio Barbanti and
Dr Emilio Guaglianone for their continuous advice and
skilled assistance in performing experiments in anaerobic
chamber and in scanning electron microscopy investiga-
tions, respectively. The authors report no conflicts of
interest. The authors alone are responsible for the content
and writing of the paper.
References
Ahmed S, Macfarlane GT, Fite A, McBain AJ, Gilbert P &
Macfarlane S (2007) Mucosa-associated bacterial diversity in
relation to human terminal ileum and colonic biopsy
samples. Appl Environ Microbiol 73: 7435–7442.Alauzet C, Marchandin H & Lozniewski A (2010) New insights
into Prevotella diversity and medical microbiology. Future
Microbiol 5: 1695–1718.Amit-Romach E, Uni Z & Reifen R (2010) Multistep
mechanism of probiotic bacterium, the effect on innate
immune system. Mol Nutr Food Res 54: 1–8.Bartlett JG (2010) Clostridium difficile: progress and challenges.
Ann NY Acad Sci 1213: 62–69.Berg RD (1996) The indigenous gastrointestinal microflora.
Trends Microbiol 4: 430–435.Brook I (2008) Microbiology and management of abdominal
infections. Dig Dis Sci 53: 2585–2591.Burmølle M, Thomsen TR, Fazli M et al. (2010) Biofilms in
chronic infections – a matter of opportunity – monospecies
biofilms in multispecies infections. FEMS Immunol Med
Microbiol 59: 324–336.Fakhry S, Manzo N, D’Apuzzo E, Pietrini L, Sorrentini I, Ricca
E, De Felice M & Baccigalupi L (2009) Characterization of
intestinal bacteria tightly bound to the human ileal
epithelium. Res Microbiol 160: 817–823.Frick IM, Karlsson C, Morgelin M, Olin AI, Janjusevic R,
Hammarstrom C, Holst E, de Chateau M & Bjorck L (2008)
Identification of a novel protein promoting the colonization
ª 2012 Federation of European Microbiological Societies FEMS Immunol Med Microbiol 65 (2012) 318–325Published by Blackwell Publishing Ltd. All rights reserved
324 G. Donelli et al.

and survival of Finegoldia magna, a bacterial commensal
and opportunistic pathogen. Mol Microbiol 70: 695–708.Gibbons RJ & Nygaard M (1970) Interbacterial aggregation of
plaque bacteria. Arch Oral Biol 15: 1397–1400.Gorbach SL (1996) Microbiology of the Gastrointestinal Tract.
Medical Microbiology, 4th edn (Baron S, ed.), chapter 95.
University of Texas Medical Branch at Galveston, Galveston,
TX.
Guaglianone E, Cardines R, Mastrantonio P, Di Rosa R, Penni
A, Puggioni G, Basoli A, Fiocca F & Donelli G (2008) Role
of multispecies microbial biofilms in the occlusion of biliary
stents. Microb Ecol Health Dis 20: 207–209.Guaglianone E, Cardines R, Vuotto C, Di Rosa R, Babini V,
Mastrantonio P & Donelli G (2010) Microbial biofilms
associated with biliary stent clogging. FEMS Immunol Med
Microbiol 59: 410–420.Huggan PJ & Murdoch DR (2008) Fusobacterial infections:
clinical spectrum and incidence of invasive disease. J Infect
57: 283–289.James GA, Swogger E, Wolcott R, Pulcini E, Secor P, Sestrich
J, Costerton JW & Stewart PS (2008) Biofilms in chronic
wounds. Wound Repair Regen 16: 37–44.Kaplan CW, Lux R, Haake SK & Shi W (2009) The
Fusobacterium nucleatum outer membrane protein RadD is
an arginine-inhibitable adhesin required for inter-species
adherence and the structured architecture of multi-species
biofilm. Mol Microbiol 71: 35–47.Kolenbrander PE (2011) Multispecies communities:
interspecies interactions influence growth on saliva as sole
nutritional source. Int J Oral Sci 3: 49–54.Ledder RG, Timperley AS, Friswell MK, Macfarlane S & McBain
AJ (2008) Coaggregation between and among human
intestinal and oral bacteria. FEMS Microbiol Ecol 66: 630–636.Marsh PD, Moter A & Devine DA (2011) Dental plaque
biofilms: communities, conflict and control. Periodontology
2000 55: 16–35.Periasamy S & Kolenbrander PE (2009) Aggregatibacter
actinomycetemcomitans builds mutualistic biofilm
communities with Fusobacterium nucleatum and Veillonella
species in saliva. Infect Immun 7: 3542–3551.
Pumbwe L, Skilbecka CA, Nakanoa V, Avila-Camposc MJ,
Piazzad RMF & Wexlera HM (2007) Bile salts enhance
bacterial co-aggregation, bacterial-intestinal epithelial cell
adhesion, biofilm formation and antimicrobial resistance of
Bacteroides fragilis. Microb Pathog 43: 78–87.Pumbwe L, Skilbeck CA & Wexler HM (2008) Presence of
quorum-sensing systems associated with multidrug
resistance and biofilm formation in Bacteroides fragilis.
Microb Ecol 56: 412–419.Roberts GL (2000) Fusobacterial infections: an underestimated
threat. Br J Biomed Sci 57: 156–162.Spigaglia P, Barbanti F & Mastrantonio P; on behalf of the
European Study Group on Clostridium difficile (ESGCD)
(2011) Multidrug resistance in European Clostridium difficile
clinical isolates. J Antimicrob Chemother 66: 2227–2234.Sproule-Willoughby KM, Stanton MM, Rioux KP, McKay
DM, Buret AG & Ceri H (2010) In vitro anaerobic biofilms
of human colonic microbiota. J Microbiol Methods 83: 296–301.
Stepanovic S, Vukovic D, Dakic I, Savic B & Svabic-Vlahovic
M (2000) A modified microtiter-plate test for quantification
of staphylococcal biofilm formation. J Microbiol Methods
40: 175–179.Thornton RB, Rigby PJ, Wiertsema SP, Filion P, Langlands J,
Coates HL, Vijayasekaran S, Keil AD & Richmond PC
(2011) Multi-species bacterial biofilm and intracellular
infection in otitis media. BMC Pediatr 11: 94.
Varga JJ, Therit B & Melville SB (2008) Type IV pili and the
CcpA protein are needed for maximal biofilm formation
by the Gram-positive anaerobic pathogen Clostridium
perfringens. Infect Immun 76: 4944–4951.Verma R, Dhamija R, Ross SC, Batts DH & Loehrke ME
(2010) Symbiotic bacteria induced necrotizing pancreatitis
JOP. J Pancreas 11: 474–476.Wexler HM (2007) Bacteroides: the good, the bad, and the
nitty-gritty. Clin Microbiol Rev 20: 593–621.Zilm PS & Rogers AH (2007) Co-adhesion and
biofilm formation by Fusobacterium nucleatum in response
to growth pH. Anaerobe 13: 146–152.
FEMS Immunol Med Microbiol 65 (2012) 318–325 ª 2012 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Biofilm-growing intestinal anaerobic bacteria 325





























