Embed Size (px)
Citation preview

Stimulators and Inhibitors ofHepatic Porphyrin Formation in Human Sera
AR1EENB. RiF'KmD, SHIGERU SASSA, IRWiN R. MimiZ, ROBERTWINCHESTER,LEONARDHARBER, and ATrALLAH KAPPAS
From the Rockefeller University, NewYork 10021, the Department ofObstetrics and Gynecology, Cornell University Medical College, New York 10021,and the Department of Dermatology, NewYork University Medical School,NewYork 10016
A B S T R A C T Human sera were found to contain fac-tors that stimulate and factors that inhibit porphyrinformation by cultured avian liver cells. The capacity ofsera to stimulate or inhibit porphyrin formation variedin different hormonal states and in the porphyrias. Serafrom 31 post partum women, eight of whom were notlactating, inhibited porphyrin formation to a mean level30% below the level in control cultures and also in-hibited drug and steroid stimulation of porphyrin for-mation. In contrast, mean porphyrin formation com-pared to control cultures was increased between 9 and21% by sera from 52 normal subjects, 16 women on oralcontraceptives, and 11 pregnant women. It was increased193% by sera from nine subjects with acute intermittentporphyria and 172% by sera from 13 subjects with por-phyria cutanea tarda. Heated sera or ethanol extracts ofsera from all groups of subjects further increased themean porphyrin stimulation by sera and, for the postpartum subjects, eliminated the inhibitory effect. Ethanolextracts of sera from 28 oral contraceptive-treatedwomen caused significantly greater mean stimulation ofporphyrin formation than did extracts of sera from 30normal women. While sera from 17 out of 22 porphyricsubjects contained both stimulatory and inhibitory fac-tors, 5 out of 22 had no evidence of an inhibitory com-ponent. There appeared to be heterogeneity in the oc-currence of the factors among porphyrics.
The factor(s) in sera responsible for porphyrin stimu-lation were heat-stable and insensitive to trypsin; werepresent in the supernates after ethanol precipitation ofplasma proteins; were extractable in ethyl acetate andnondialyzable; and they migrated with the albumin-containing fraction of serum during electrophoresis.
Received for publication 17 January 1973 and in revisedform 19 July 1973.
The factor (s) responsible for porphyrin inhibition wereheat labile, sensitive to trypsin, and resistant to neu-raminidase; were present in the ethanol precipitates ofsera and were nondialyzable; and they migrated withthe gamma globulin fraction of serum during electro-phoresis. Inhibition of porphyrin formation was not at-tributable to heme, free or bound as hemoglobin, hemo-pexin, or hemalbumin.
INTRODUCTIONExtracts of plasma from patients with acute intermit-tent porphyria (AIP)1 and from women ingesting oralcontraceptive steroids have been found to stimulateporphyrin formation in cultured chick embryo liver cells(1). The chemical nature of the factor, or factors, insera that increase the accumulation of porphyrins hasnot been identified. Among the many different drugsand steroids that have been shown to stimulate porphy-rin formation, both in cultured liver cells and in ani-mals and humans in vivo (2-9), steroid hormones andtheir metabolites are the only compounds occurringnaturally in man. Well-documented variations in clini-cal activity of porphyria with respect to sex and en-dogenous hormonal alterations (10), and evidence ofincreased urinary excretion of steroids with the ca-pacity to stimulate porphyrins in AIP (11, 12), makeparticularly interesting the possibility that circulatinghormonal substances are involved in the pathogenesisof porphyria.
1Abbreviations used in this paper: AIA, allylisopropyl-acetamide; AIP, acute intermittent porphyria; ALA, 8-aminolevulinic acid; ALAS, 8-aminolevulinic acid synthe-tase; DDC, 3,5-diethyl-1,4-dihydrocollidine dicarboxylate;PCT, porphyria cutanea tarda.
The Journal of Clinical Investigation Volume 53 April 1974-1167-1177 1167

The present study was designed to examine the ef-fects of sera fronm subjects in a variety of hormonialstates on liver cell porphyrin accumulation, and tocompare these to the effects of sera from normal andporphyric subjects. We found that human sera containfactors that stimulate and factors that inhibit porphyrinformation by liver cells. The amount and relative pro-portion of those factors vary in different hormonalstates anid in the porphyrias. This report describesthese findinigs and our progress in separating andcharacterizing the serum conmponents responsible forthe stiniulatory and inihibitor) plhenonlelnea.
METHODSSubjects aitd blood samples. The subjects studied in-
cluded 52 normal men and women, 28 women ingesting oralcontraceptive steroids, 11 pregnant women, 31 post partumwomen, nine patients with AIP, and 13 patients withporphyria cutanea tarda (PCT). The normal subjects werecomposed of two groups. The first group, designated "nor-mal subjects A," consisted of 33 women between the agesof 20 and 35 who attended the obstetrics and gynecologyclinic at The New York Hospital for routine examina-tions. Their sera and sera extracts were used for the datain Fig. 1. The second group, designated "normal subjectsB," consisted of 11 women and eight men, between theages of 21 and 35, who were laboratory personnel andvolunteers for this study. Their sera were used for thedata in Fig. 4. All normal subjects were healthy and deniedhaving ingested any drugs within the month preceding thestudy.
The contraceptive-treated group consisted of 28 women,ages 18-38, attending the obstetrics and gynecology clinicat The New York Hospital. Each subject was taking theusually recommended doses of Ortho-novum (norethin-drone and mestranol, Ortho Pharmaceutical Corp., Raritan,N. J.), Ovulen (ethynodiol diacetate and mestranol, G. D.Searle and Co., Chicago, Ill.), or Norlestrin (norethin-drone acetate and ethinyl estradiol, Parke, Davis and Co.,Detroit, Mich.). They had been taking the contraceptivesfrom 3 to 30 mo.
The pregnant group consisted of 11 women, ages 23-32,attending the prenatal clinic at The New York Hospital.They were all between the 30th and 38th wk of gestationand, therefore, were all in the 3rd trimester. Six wereprimigravidas and five were gravida twos.
Sera were obtained from 20 post partum women, 3-4days after normal spontaneous deliveries at The NewYork Hospital. Most of the women selected (17 subjects)were lactating, thus excluding any effect from hormonesgiven to suppress lactation. Sera from three nonlactatingwomen were also examined. Blood samples were also ob-tained at routine examinations, 6 wk post partum, from 11women, 6 of whom had breast fed their infants and fiveof whom had not. The post partum women were be-tween 14- and 25-yr old. Of the 31 subjects in this group,there were 17 primigravidas, 9 gravida twos, 2 gravidathrees, and one each gravida four, five, and seven.
9 subjects with AIP and 13 with PCT were patientsat the Rockefeller University Hospital or University Hos-pital (New York University). Diagnoses were based oncharacteristic history, clinical observations, and measure-ments of urinary porphyrins and porphyrin precursors, ac-
cording to the criteria described by Marver and Schmid(10). The AIP patients included one man and eight women,ranging in age from 22 to 56. The PCT patients includednine men and four women; they ranged in age from 30to 65. During the study, two AIP inpatients receivedmeperidine for pain. The others with AIP and those withPCT had received no drugs within at least 1 mo bef oregiving the serum samples.
Blood was drawn into tubes without anticoagulants.Clots were allowed to form for 2 h, and then the sampleswere spuni at 2,000 rpm f or 20 mim at 4°C. 2-ml aliquotsof sera were placed in -lass vials and kept at 0°C untilused.
Experiiettal miethods. Chick embryo liver cells weregrown in primary cultures according to the method ofGranick (2). In brief, livers from 14- or 15-day-old chickembryos were minced and then incubated at 37'C for 20-30min in 9 ml 1% trypsin (Gallard-Schlesinger ChemicalMfg. Corp., Carle Place, N. Y.) and 3 ml 1o pangestin(Difco Laboratories, Detroit, Mich.). Approximately 5 X105 cells were added to 19 x 65-mm glass shell vials, ineach of which a glass cover slip had been placed. Eagle'spowdered basal medium (Grand Island Biological Co.,Grand Island, N. Y.) in distilled water was supplementedper liter, as follows: 3.0 g sodium bicarbonate, 100 mlfetal bovine serum (Microbiological Associates, Inc., Beth-esda, Md.), 10 ml amino acid concentrate (MicrobiologicalAssociates, Inc.), and antibiotics (streptomycin, 100 mg,penicillin, 100,000 U, and Mycostatin [E. R. Squibb andSons, Princeton, N. J.], 25,000 U). pH was adjusted to7.4. 1 ml medium was added to each vial. The vials wereincubated at 37°C in an atmosphere of 95% air and 5%CO2 for 24 h, after which the media were replaced withfresh media, and additions to the vials (sera, extracts ofsera, modified sera, drugs) were made. The vials were in-cubated for an additional 20-24 h and then frozen.
Porphyrins were measured by the method of Granick(2), modified by Sassa. The modifications involved lyophi-lizing the cells and media, measuring total porphyrin ac-cumulation in the combined cells and media (rather thanin the cells alone), and using an R-136 photomultipliertube to enhance red-light sensitivity of the fluorimeter.Porphyrins were extracted from the lyophilized materialby adding 5 ml of a solution of 1 N perchloric acid: metha-nol (1: 1, vol/vol) to each vial in the dark, stirring ona Vortex mixer (Scientific Industries, Inc., Queens Village,N. Y.), and filtering through Whatman paper no. 2 after30 min. Porphyrins were measured in the filtrate, usinga Hitachi model MPF 2 A spectrophotofluorimeter (ThePerkin-Elmer Corp., Norwalk, Conn.) at an excitationwave length of 400 nm and an emission wave length of 654nm. In some experiments, emission spectra from 560 to 700nm were obtained to characterize the type of porphyrinformed. A coproporphyrin III reference standard was in-cluded in each assay to permit quantification of the por-phyrins. Mean recovery of added coproporphyrin III was90%O.
In control experiments, 100 ul untreated or heated serafrom normal, post partumi, and porphyric individuals, (a)yielded no measurable porphyrins when incubated withmedia in the absence of cells (b) did not produce anychanges in porphyrin formation when added to cells at theend of the incubation period, and (c) did not interferewith the measurement of porphyrins. Therefore, the serumeffects on liver cell porphyrin formation we observedcould not be attributed to porphyrins in sera, to direct
1168 Rifkind, Sassa, Merkatz, Winchester, Harber, and Kappas

effects of sera on porphyrin formation, or to interferenceby sera with the extraction or measurement of porphyrins.
Two types of controls were routinely included in eachculture experiment. The first was the inclusion of 10-15vials to which no additions were made, based on our ob-servation that 100 Al of water, 0.9% saline, 0.1 M phos-phate buffer at pH 7.4, Earle's balanced salt solution, andthe residue of 10 ml evaporated ethanol in 1 ml water,added to 1 ml medium, had no effect on porphyrin accumu-lation. The second control was the inclusion of 10-15vials containing 30 Aig allylisopropylacetamide (AIA), adrug which is a known stimulator of porphyrin formation.The vials with AIA were included in each experiment,both to ensure that the cells were capable of increasingporphyrin formation in response to a drug and to allowmeasurement of the extent of responsiveness of each par-ticular cell culture preparation. In early experiments weincluded several doses of AIA to construct AIA dose-response curves but later used multiple vials with onestandard dose of AIA (30 ,g) which we found equallyeffective for judging the responsiveness of each cell cul-ture preparation.
Each cell culture preparation consisted of 150-250 vials,and the data reported here are derived from approximately60 cell culture experiments. Each experimental group in-cluded five to ten replicate vials. The mean porphyrin ac-cumulation in each group of replicate vials was calculated,as well as the SE, SD, and total variance.
Within each cell culture experiment, the mean variationbetween duplicate vials was small (about 10%). Therewas, however, greater variation in responsiveness amongseparate cell culture experiments. For example, in 60 ex-periments, 30 ,ug AIA resulted in porphyrin stimulationranging from 5 to 33 times control values, with a meanvalue of 10 times control. To facilitate comparisons amongresponses of sera tested in different cell culture experi-ments, the data have been normalized as follows: the ratiobetween the observed response to AIA (30 ;Lg) and theaverage AIA response (10 times control) was derived; theexperimental results were then multiplied by that ratio toobtain corrected results; the control values were taken as100%; and the percent increase or decrease in porphyrinformation after the addition of serum was calculated. Forexample, if control values were 5 pmol porphyrin/vial,and AIA (30 ,g) resulted in porphyrin formation of 40,pmol/vial, the ratio, [50 (expected) - 5 (control) ]/[40(observed) - 5 (control) ] = 1.29, would be calculated. Ahypothetical serum response of 12 pmol/vial would bemultiplied by the factor, 1.29. The corrected result, 15.5,would be used to calculate the percent increase in porphy-rin formation compared to controls. When the observedporphyrin stimulation by 30 ,g AIA exceeded 10 timescontrol, the serum responses were correspondingly de-creased by the correction procedure. For inhibitory effectsof sera in cultures in which porphyrin formation was stim-ulated by AIA, the data were corrected as described above.For inhibitory effects in control cultures, no adjustmentswere made because the amount of inhibition of the al-ready low porphyrin levels in control cultures could notbe shown to be related to the induction response of theparticular culture to AIA. Statistical significance of dif-ferences from the appropriate control and statistical sig-nificance of differences in mean responses of sera fromdifferent groups of subjects were determined by Student'st test. Differences were accepted as being statistically sig-nificant if the P value was < 0.05.
The normalized data are presented for purposes of com-
paring results from several cell culture experiments. Wewish to emphasize, however, that the normalization proce-dure did not result in any qualitative or any statisticallysignificant quantitative changes in the findings, since themean results when calculated from the normalized datawere, in no instance, significantly different from the resultscalculated from the raw data. The normalization proceduredemonstrates that variations in responsiveness of cell cul-ture preparations cannot account for the findings reported.
Six lots of fetal bovine sera were used in these experi-ments. Each lot had been pretested and accepted if it hadno harmful effect on cell morphology and if it permittedat least a 10-fold stimulation of porphyrin formation by30 ,Ag AIA. There were no significant differences in therange of responses to AIA in cell cultures using the dif-ferent lots of bovine sera. Moreover, sera from eachgroup of subjects were examined with each lot of bovinesera, and the behavior, of the sera remained true to type,i.e., extracts of sera from oral contraceptive-treated andporphyric subjects increased porphyrin accumulation, whilepost partum sera inhibited porphyrin formation, regardlessof the lot of sera used. Sera from eight individuals weretested in cell culture experiments using two-four differentlots of bovine sera. The mean difference in response wasonly 15%±o0.03 (SE). Therefore, since different lots of fetalbovine sera did not introduce a major variable into thesystem, no adjustment was made for the different lots offetal bovine sera. 100 Al fetal bovine sera, an amount equiva-lent to the amount of human sera usually added to cultures,did not affect control or AIA-induced porphyrin formation.100 /4 fetal bovine sera heated to 60'C for 10 min resultedin porphyrin formation 1.21 times the level in controlcultures.
8-Aminolevulinic acid synthetase (ALAS) activity wasmeasured in the mitochondrial subfractions from liver, cellsgrown in 10-cm Petri dishes (Falcon Plastics, Div. of Bio-quest, Oxnard, Calif.) containing 20 times the amount ofmedia, cells, and sera used in the vials (13). Reactionswere carried out in duplicate in incubation volumes of 200pl and were stopped with 200 ,ul 10% trichloroacetic acid.Each reaction mixture contained one-fifth of the mito-chondrial (10,000g) pellet derived from cells scraped to-gether from two replicate Petri dishes. A zero-time con-trol was always included. DNA was measured by themethod of Kissane and Robins (114). Hemoglobin wasmeasured by difference spectroscopy, using the method ofLevere, Swerdlow, and Garavoy (15).
Ethanol extracts of sera were prepared by adding 10vol of cold absolute ethanol to 1 or 2 ml sera (1). Aftercentrifugation at 2,000 rpm for 10 min, the ethanolic super-nate was removed and the ethanol evaporated in vacuo at37° C. The dry residue was brought back to the originalserum volume with Earle's balanced salt solution. It is thisresolubilized dried ethanolic supernate that is referred toas "ethanol extract" in the text. Usually, 100 IAI sera,ethanol extracts of sera, or sera otherwise modified (seebelow) were added per culture vial.
To extract and partially purify the proteins precipitatedby ethanol, the precipitates were extracted twice with 4vol of 10%o ammonium acetate in 70%o ethanol. 2 vol of10% ammonium acetate in 100%o ethanol were added to thecombined supernates, which were allowed to remain at4°C overnight to reprecipitate the protein (16). The re-sulting fine precipitate was washed with ether, dried underair, and dissolved in a vol of Earle's balanced salt solutionequal to the volume of the original serum sample.
For experiments examining the effect of neuraminidase
Stimulators and Inhibitors of Hepatic Porphyrin Formation in Human Sera 1169
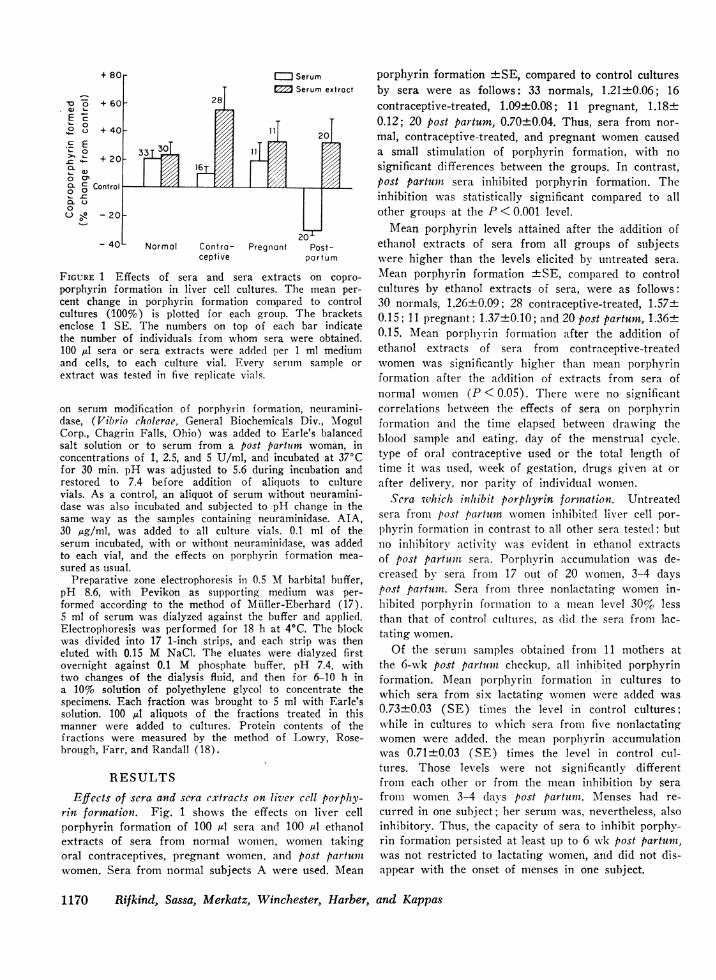
+ 80r I= SerumE7 Serum extroct
Normal Contra- Pregnont Post-ceptive portum
FIGURE 1 Effects of sera and sera extracts on copro-
porphyrin formation in liver cell cultures. The mean per-
cent change in porphyrin formation compared to controlcultures (100%v) is plotted for each group. The bracketsenclose 1 SE. The numbers on top of each bar indicatethe number of individuals from whom sera were obtained.100 ,ul sera or sera extracts were added per 1 ml mediumand cells, to each culture vial. Every sertim sample or
extract was tested in five replicate vials.
on serum modification of porphyrin formation, neuramini-dase, (Vibrio cholerae, General Biochemicals Div., MogulCorp., Chagrin Falls, Ohio) was added to Earle's balancedsalt solution or to serum from a post partutm woman, inconcentrations of 1, 2.5, and 5 U/ml, and incubated at 37'Cfor 30 min. pH was adjusted to 5.6 during incubation andrestored to 7.4 before addition of aliquots to culturevials. As a control, an aliquot of serum without neuramini-dase was also incubated and subjected to pH change in thesame way as the samples containing neuraminidase. AIA,30 ,ug/ml, was added to all culture vials. 0.1 ml of theserum incubated, with or without neuraminidase, was addedto each vial, and the effects on porphyrin formation mea-
sured as usual.Preparative zone electrophoresis in 0.5 M barbital buffer,
pH 8.6, with Pevikon as supporting medium was per-formed according to the method of Miiller-Eberhard (17).5 ml of serum was dialyzed against the buffer and appliecl.Electroplhoresis was performed for 18 h at 4°C. The blockwas divided into 17 1-inch strips, and each strip was theneluted with 0.15 M NaCI. The eluates were dialyzed firstovernight against 0.1 M phosphate buffer, pH 7.4, withtwo changes of the dialysis fluid, and then for 6-10 h ina 10% solution of polyethylene glycol to concentrate thespecimens. Each fraction was brought to 5 ml with Earle'ssolution. 100 ,ul aliquots of the fractions treated in thismanner were added to cultures. Protein contents of thefractions were measured by the method of Lowry, Rose-brough, Farr, and Randall (18).
RESULTS
Effects of sera and sera extracts on liver cell porphy-rin formation. Fig. 1 shows the effects on liver cellporphyrin formation of 100 Al sera and 100 Al ethanolextracts of sera from normal women, women takingoral contraceptives, pregnant women, and post partumwomen. Sera from normal subjects A were used. Mean
porphyrin formation +SE, compared to control culturesby sera were as follows: 33 normals, 1.21-+-0.06; 16contraceptive-treated, 1.09+0.08; 11 pregnant, 1.18±0.12; 20 post partum, 0.70±0.04. Thus, sera from nor-mal, contraceptive-treated, and pregnant women causeda small stimulation of porphyrin formation, with nosignificant differences between the groups. In contrast,post partumin sera inhibited porphyrin formation. Theinhibition was statistically significant compared to allother groups at the P < 0.001 level.
Mean porphyrin levels attained after the addition ofethanol extracts of sera from all groups of subjectswere higher than the levels elicited by untreated sera.Mean porphyrin formation 4±SE, compared to controlcultures by ethanol extracts of sera, were as follows:30 normals, 1.26+0.09; 28 contraceptive-treated, 1.57+0.15; 11 pregnant; 1.37±0.10; and 20 post partum, 1.36+0.15. Mean porphyrin formation after the addition ofethanol extracts of sera from contraceptive-treatedwomen was significantly higher than mean porphyrinformation after the addition of extracts from sera ofnormal women (P < 0.05). There were no significantcorrelations between the effects of sera on porphyrinformation and the time elapsed between drawing theblood sample and eating, day of the menstrual cycle.type of oral contraceptive used or the total length oftime it was used, week of gestation, drugs given at orafter delivery, nor parity of individual women.
Scra zcthich inihibit porphyrin formation. Untreatedsera from post partumi women inhibited liver cell por-phyrin formation in contrast to all other sera tested; butno inhibitory activity was evident in ethanol extractsof post partunt sera. Porphyrin accumulation was de-creased by sera from 17 out of 20 women, 3-4 dayspost partumi. Sera from tlhree nonlactating women in-hibited porphyrin formationi to a mean level 30%' lessthan that of control cuilttures, as did the sera from lac-tating women.
Of the serum samples obtained from 11 mothers atthe 6-wk post partumiii checkup, all inhibited porphyrinformation. Mean porphyrin formation in cultures towhich sera from six lactating women were added was0.73±0.03 (SE) times the level in control cultures;while in cultures to which sera from five nonlactatingwomen were added, the mean porphyrin accumulationwas 0.71±0.03 (SE) times the level in control cul-tures. Those levels were not significantly differentfrom each other or from the mean inhibition by serafrom women 3-4 days post partumn. AMenses had re-curred in one subject; her serum was, nevertheless, alsoinhibitory. Thus, the capacity of sera to inhibit porphy-rin formation persisted at least up to 6 wk post partum,was not restricted to lactating women, and did not dis-appear with the onset of menses in one subject.
1170 Rifkind, Sassa, Merkatz, Winchester, Harber, and Kappas
o3 + 60.Ia) L-E:
0o + 40
.' Et-0
>, + 200.. a)o 0'
. c) Control0 0
0U g~ - 20

TABLE I
Serum Inhibition of Porphyrin Formation Induced by Drugs and Steroids*
Number Coproporphyrin/ InhibitionAddition to culture vials of vials vial iSE by serum
pmol %None (control) 15 11.040.7Serum 11 5.14:0.2T -54AIA 30 ,pg 7 164.44:7.0AIA 30 pg + serum 7 16.3 412. 1 -90Etiocholanolone 5 pg 7 45.2:4:4.9Etiocholanolone 5 pug + serum 7 8.040.3$ -8211-Ketopregnanolone 5 pAg 7 21.1 4 1.611-Ketopregnanolone 5 pg + serum 7 4.8410.61 -77DDC10 ,g 7 56.44:3.6DDC10 pAg + serum 7 36.3413.1§ -36Secobarbital 20 ,g 7 43.2 42.4Secobarbital 20 ,g + serum 7 10.440.8t -76Mephenytoin 20 jug 7 124.0+431.2Mephenytoin 20 pAg + serum 7 14.3 0.2t -88
* Each culture vial contained 1 ml medium and approximately 5 X 105 liver cells. Serum wasalways added in concentrations of 100 pl/vial. All drugs resulted in significant stimulation ofporphyrin formation compared to control cultures (P < 0.001).t§ Significantly less porphyrin formation than in conmparable cultures without serumt (P < 0.001), or § (P < 0.05).
To amplify the inhibitory effect, 100 A post partumsera, together with 30 pg AIA, were added to cultures.Mean porphyrin formation in cultures to which 23 postpartum sera were added was 0.40±0.06 (SE), whilein cultures to which sera from 12 normal women wereadded, it was 0.91±0.08 (SE) of the AIA-inducedcontrols. The difference was significant at the P < 0.001level. Post partumn sera also inhibited the stimulation ofporphyrin formation by several different drugs andsteroid hormones, including secobarbital (Seconal, EliLilly and Co., Indianapolis, Ind.), mephenytoin (Mesan-toin, Sandoz Pharmaceuticals, East Hanover, N. J.),3,5-diethyl-1,4-4dihydrocollidine dicarboxylate (DDC),5P-androstan-3a-ol-17-one (etiocholanolone) and 5f-androstan-3a-ol-1 1,20-dione (11-ketopregnanolone). Theresults of a characteristic experiment are shown inTable I.
The effect of sera which inhibited porphyrin forma-tion on ALAS activity in liver cell cultures was mea-sured (Table II). Compared to control cultures, serafrom two post partum subjects, which had inhibitedporphyrin formation by 54 and 32%, depressed ALASactivity by 61 and 29%, respectively, whereas a serumsample from a normal male that had not inhibited por-phyrin formation did not inhibit ALAS. The depressionof ALAS activity was not attributable to a direct in-hibitory effect of the sera on ALAS, since the samesera added to chick embryo liver homogenates in vitrodid not alter the activity of control or drug-inducedALAS.
When 5 pg 8-aminolevulinic acid (ALA) was addedto the culture vials, a mean increase of p.orphyrins to20 times the level in control cultures resulted. Sera fromeight post partum subjects caused a mean inhibition ofconversion of 5 ug ALA to porphyrins of 34%±0.06(SE). Sera from the same eight subjects inhibited theinduction of porphyrin formation by 30 pg AIA, by55%+0.06 (SE). The decrease in AIA induction wassignificantly greater than the decrease in ALA con-version (P < 0.05).
TABLE I IEffects of Post Partum Sera on ALAS Activity in
Cultured Liver Cells
Additions to cultures* ALAS activity
pmol ALA formedl,Ag DNAper h
Experiment 1None (control) 32.7sPost partum serum A 12.7Normal male serum 30.3
Experiment 2None (control) 30.4Post partum serum B 21.8
* Serum was added to Petri dishes containing 20 ml media andapproximately 101 liver cells in a concentration of 0.1 ml/mlmedium.I Each result is the mean of duplicate determinations on livercells from two Petri dishes.
Stimulators and Inhibitors of Hepatic Porphyrin Formation in Human Sera 1171
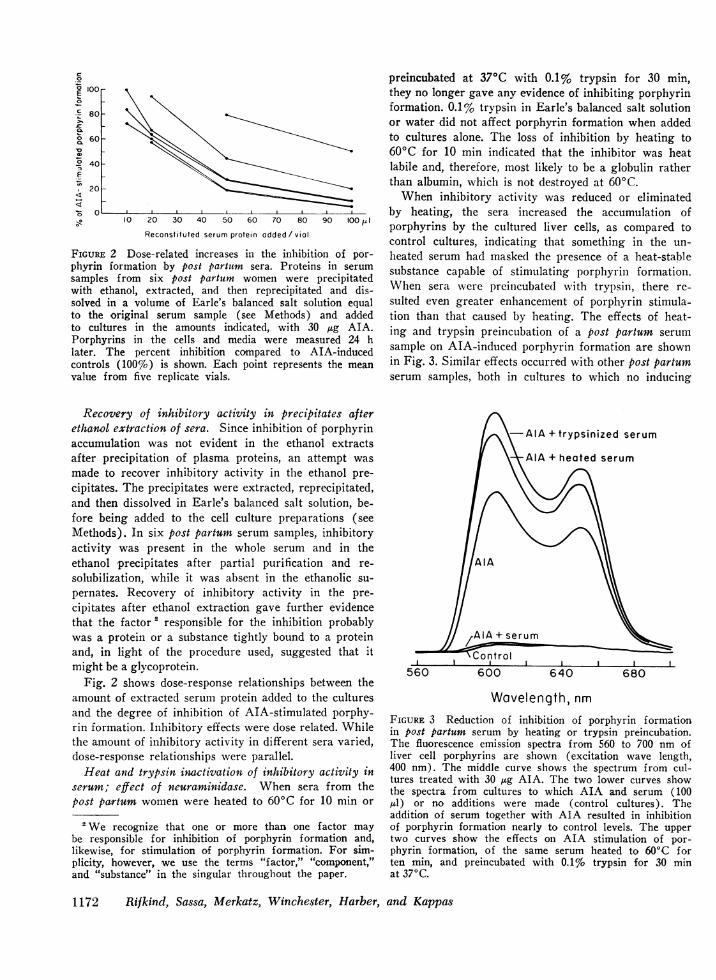
c0
0 100E
,c 80
° 60
°2 40F
2020o O
10 20 30 40 50 60 70 80 90
Reconstituted serum protein added/ vial
100 81I
FIGURE 2 Dose-related increases in the inhibition of por-phyrin formation by post partutm sera. Proteins in serumsamples from six post partum women were precipitatedwith ethanol, extracted, and then reprecipitated and dis-solved in a volume of Earle's balanced salt solution equalto the original serum sample (see Methods) and addedto cultures in the amounts indicated, with 30 ,ug AIA.Porphyrins in the cells and media were measured 24 hlater. The percent inhibition compared to AIA-inducedcontrols (100%o) is shown. Each point represents the meanvalue from five replicate vials.
Recovery of inhibitory activity in precipitates afterethanol extraction of sera. Since inhibition of porphyrinaccumulation was not evident in the ethanol extractsafter precipitation of plasma proteins, an attempt wasmade to recover inhibitory activity in the ethanol pre-cipitates. The precipitates were extracted, reprecipitated,and then dissolved in Earle's balanced salt solution, be-fore being added to the cell culture preparations (seeMethods). In six post partum serum samples, inhibitoryactivity was present in the whole serum and in theethanol precipitates after partial purification and re-solubilization, while it was absent in the ethanolic su-pernates. Recovery of inhibitory activity in the pre-cipitates after ethanol extraction gave further evidencethat the factor 2 responsible for the inhibition probablywas a protein or a substance tightly bound to a proteinand, in light of the procedure used, suggested that itmight be a glycoprotein.
Fig. 2 shows dose-response relationships between theamount of extracted serum protein added to the culturesand the degree of inhibition of AIA-stimulated porphy-rin formation. Inhibitory effects were dose related. Whilethe amount of inhibitory activity in different sera varied,dose-response relationships were parallel.
Heat and trypsin inactivation of inhibitory activity inserum; effect of neuraminidase. When sera from thepost partum women were heated to 60°C for 10 min or
2 We recognize that one or more than one factor maybe responsible for inhibition of porphyrin formation and,likewise, for stimulation of porphyrin formation. For sim-plicity, however, we use the terms "factor," "component,"and "substance" in the singular throughout the paper.
preincubated at 37°C with 0.1% trypsin for 30 min,they no longer gave any evidence of inhibiting porphyrinformation. 0.1% trypsin in Earle's balanced salt solutionor water did not affect porphyrin formation when addedto cultures alone. The loss of inhibition by heating to60°C for 10 min indicated that the inhibitor was heatlabile and, therefore, most likely to be a globulin ratherthan albumin, which is not destroyed at 600C.
When inhibitory activity was reduced or eliminatedby heating, the sera increased the accumulation ofporphyrins by the cultured liver cells, as compared tocontrol cultures, indicating that something in the un-heated serum had masked the presence of a heat-stablesubstance capable of stimulating porphyrini formation.When sera were preincubated with trypsin, there re-sulted even greater enhancement of porphyrin stimula-tion than that caused by heating. The effects of heat-ing and trypsin preincubation of a post partum serumsample on AIA-induced porphyrin formation are shownin Fig. 3. Similar effects occurred with other post partumserum samples, both in cultures to which no inducing
*AIA + trypsinized serum
*AlA + heated serum
IA + serumj _r
C~~ontrolIL- I I I.,
560 600 640 680
Wavelength, nmFIGURE 3 Reduction of inhibition of porphyrin formationin post partum serum by heating or trypsin preincubation.The fluorescence emission spectra from 560 to 700 nm ofliver cell porphyrins are shown (excitation wave length,400 nm). The middle curve shows the spectrum from cul-tures treated with 30 ,ug AIA. The two lower curves showthe spectra f rom cultures to which AIA and serum (100,ul) or no additions were made (control cultures). Theaddition of serum together with AIA resulted in inhibitionof porphyrin formation nearly to control levels. The uppertwo curves show the effects on AIA stimulation of por-phyrin formation, of the same serum heated to 60°C forten min, and preincubated with 0.1% trypsin for 30 minat 370C.
1172 Rifkind, Sassa, Merkatz, Winchester, Harber, and Kappas
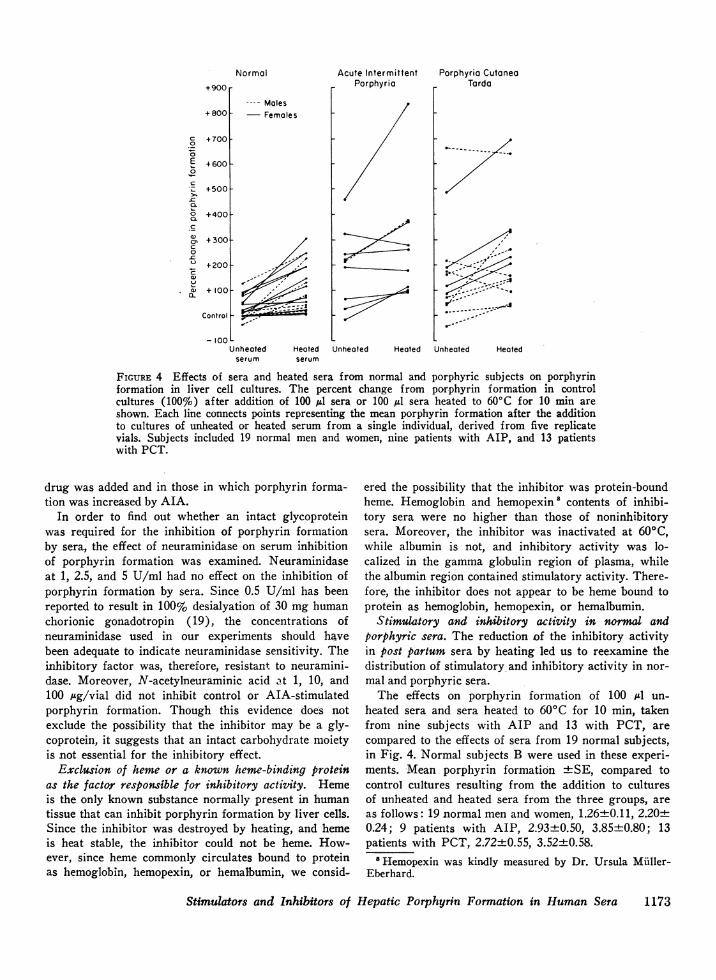
Acute IntermittentPorphyria
Unheated Heated
Porphyria CutoneaTarda
V, :-
------
Unheated Heated
FIGURE 4 Effects of sera and heated sera from normal and porphyric subjects on porphyrinformation in liver cell cultures. The percent change from porphyrin formation in controlcultures (100%) after addition of 100 Al sera or 100 IL sera heated to 600C for 10 min areshown. Each line connects points representing the mean porphyrin formation after the additionto cultures of unheated or heated serum from a single individual, derived from five replicatevials. Subjects included 19 normal men and women, nine patients with AIP, and 13 patientswith PCT.
drug was added and in those in which porphyrin forma-tion was increased by AIA.
In order to find out whether an intact glycoproteinwas required for the inhibition of porphyrin formationby sera, the effect of neuraminidase on serum inhibitionof porphyrin formation was examined. Neuraminidaseat 1, 2.5, and 5 U/ml had no effect on the inhibition ofporphyrin formation by sera. Since 0.5 U/ml has beenreported to result in 100% desialyation of 30 mg humanchorionic gonadotropin (19), the concentrations ofneuraminidase used in our experiments should havebeen adequate to indicate neuraminidase sensitivity. Theinhibitory factor was, therefore, resistant to neuramini-dase. Moreover, N-acetylneuraminic acid at 1, 10, and100 gg/vial did not inhibit control or AIA-stimulatedporphyrin formation. Though this evidence does notexclude the possibility that the inhibitor may be a gly-coprotein, it suggests that an intact carbohydrate moietyis not essential for the inhibitory effect.
Exclusion of heme or a known heme-binding proteinas the factor responsible for inhibitory activity. Hemeis the only known substance normally present in humantissue that can inhibit porphyrin formation by liver cells.Since the inhibitor was destroyed by heating, and hemeis heat stable, the inhibitor could not be heme. How-ever, since heme commonly circulates bound to proteinas hemoglobin, hemopexin, or hemalbumin, we consid-
ered the possibility that the inhibitor was protein-boundheme. Hemoglobin and hemopexin' contents of inhibi-tory sera were no higher than those of noninhibitorysera. Moreover, the inhibitor was inactivated at 60°C,while albumin is not, and inhibitory activity was lo-calized in the gamma globulin region of plasma, whilethe albumin region contained stimulatory activity. There-fore, the inhibitor does not appear to be heme bound toprotein as hemoglobin, hemopexin, or hemalbumin.
Stimulatory and inhibitory activity in normal and
porphyric sera. The reduction of the inhibitory activityin post partum sera by heating led us to reexamine thedistribution of stimulatory and inhibitory activity in nor-
mal and porphyric sera.
The effects on porphyrin formation of 100 Al un-
heated sera and sera heated to 60(C for 10 min, takenfrom nine subjects with AIP and 13 with PCT, are
compared to the effects of sera from 19 normal subjects,in Fig. 4. Normal subjects B were used in these experi-
ments. Mean porphyrin formation +SE, compared tocontrol cultures resulting from the addition to culturesof unheated and heated sera from the three groups, are
as follows: 19 normal men and women, 1.26±0.11, 2.20±0.24; 9 patients with AIP, 2.93+0.50, 3.85+0.80; 13patients with PCT, 2.72±0.55, 3.52±0.58.
'Hemopexin was kindly measured by Dr. Ursula Muller-Eberhard.
Stimulators and Inhibitors of Hepatic Porphyrin Formation in Human Sera
Normal+900 r
+800F---- Males- Females
00
zE0
0
.
c
0'
0L
+700
+ 600 F
+500f
+400[
+300[
+200
+ 1ool-
Control _
- IOOLUnheated
serumHeatedserum
1173

701r60-
50
40k-C
E0
,,-I
2
-. 3
-=56
200,4
20k
10
10 20 50 100
Serum added/ vial
FIGURE 5 Dose-related increases in stimulation of por-pllyrin formation by sera. Sera from six subjects, (1, 2, 3,PCT; 4, AIP; 5, pregnant; 6, normal female), wereheated to 60°C for 10 min and then added to culture vialsin the amounts indicated. Porphyrin accumulation in thecells and media were measured 24 h later. Each pointrepresents the mean value for five replicate vials. Meanporphyrin formation in 17 control vials (no added sera)was 6.29±0.54 (pmol coproporphyrin±2 SE).
In all 19 normal subjects, lheated sera produced higherlevels of porphyrins than unheated sera; in 14 out of 19the stimulation was statistically significant. There wasno significant difference between the effects of serafrom males and females. Thus, sera from normal indi-viduals appeared to contain a component that inhibitedand a component that stimulated porphyrin formation;heating reduced the inhibition and resulted in the dis-play of stimulatory activity.
Untreated sera from patients with AIP and PCTcaused greater mean stimulation of porphyrin formationthan did heated sera from normal subjects; and heatingof AIP and PCT sera caused a further increase in meanporphyrin accumulation. In 5 out of 9 subjects withAIP, and in 5 out of 13 subjects with PCT, greaterstimulatory activity than that found in sera of anynormal individual was present in unheated sera. Serafrom the other porphyric subjects could not be distin-guished from normal sera. Both the AIP subject andthe PCT subject whose sera had the greatest abilityto stimulate porphyrin formation were clinically symp-tomatic. However, among the remaining subjects, nocorrelation was observed between symptoms and thecapacity of serum to stimulate porphyrin accumulation.No effects could be attributed to meperidine in thissmall sample.
Heating did not increase the capacity to stimulateporphyrin accumulation in the sera from two subjectswith AIP and one with PCT. Heating sera from twoother patients with PCT resulted in significantly de-creased stimulation of porphyrin formation, suggesting
that in certain individuals with porphyria, the inhibitorycomponent may be mssing, and moreover, that in someporphyrics the factor responsible for stimulation ofporphyrin formation may also be p)artly heat labile.
To examine the dose-response relationships of thestimulatory effect, sera from six individuals (one nor-mal, one pregnant, one with AIP, and three with PCT)were heated to 60°C for 10 min to inactivate the inhibi-tory component, and aliquots from 10 to 200 A4 wereadded to culture vials. The resulting dose-response re-lationships are shown in Fig. 5. Adding increasingamounts of heated serum to ctultutres resulted in increas-ing porphyrin formation.
Sera from two subjects with AIP were separatelyextracted with three organic solvents: ethyl acetate,dichloromethane, and petroleum ether. The stimulatoryactivities in both sera were fully extractable in ethylacetate and not extractable in dichloromethane or pe-troleum ether, suggesting that polar compounds wereresponsible for the stimulation of porphyrin formationby the sera.
- Control culture(no additions) + fractions
--- Etiocholonolone 5Es9 +fractions
40
30 \i.-
cl0
ECL
20
10
A
Sonfrol _oddit1
S I^E
2500 c
r a2 a, Alb Pre olb
2 4 6 8 10 12 14 16 18
Serum froctions
FIGURE 6 SeparatioIn by electrophoresis of the serum com-ponents responsible for inhibition and stimulation of por-phyrin formation. Each point on the upper two graphs indi-cates the mean porphyrin formation in five vials to each ofwhich 100 ,ul of one of the serum fractions derived fromelectrophoresis was added. The brackets and the shadedareas define ±2 SE. The bottom graph indicates the pro-tein content of the fractions. Inhibitory activity is evidentin the gamma region and stimulatory activity in the albu-min region.
1I74 Rifkind, Sassa, Merkatz, Winchester, Harber, and Kappas
30
cr'n

Separation of the serum stimulatory and inhibitoryfactors. An attempt was made to separate the serumcomponents responsible for stimulation and inhibition ofporphyrin formation by zone electrophoresis. A serumsample from a post partum woman which caused pro-nounced inhibition of porphyrin accumulation whenuntreated and strong stimulation of porphyrin accumu-lation when heated, was used. After dialysis, the effectsof 17 serum fractions on liver cell porphyrin formationwere examined both in control cultures and in culturesin which porphyrin formation was stimulated by etio-cholanolone. Additional controls contained dialyzed bar-bital buffer. The electrophoresis procedure effectivelyseparated the stimulatory and inhibitory activities. Stim-ulatory activity was located in the albumin region andinhibitory activity in the gamma region; both zoneswere broad (Fig. 6). Moreover, since stimulatory andinhibitory activities were both evident after exhaustivedialysis, the responsible factors were not dialyzable.
The effects on porphyrin formation of dilutions ofwhole serum were compared to the effects of dilutionsof the gamma globulin fraction which contained the in-hibitory activity (Fig. 7). As the concentration of wholeserum, was diluted by adding decreasing amounts ofserum to a constant volume of medium, the inhibitoryactivity declined, and stimulatory activity became ap-parent. However, when the fractions which inhibitedporphyrin formation were progressively diluted, theinhibitory effect diminished, but no stimulation was ob-served. Moreover, while heated whole serum stimulatedthe formation of porphyrins, the heated fractions con-taining the inhibitory activity did not; the inhibitoryeffect was merely diminished. Thus, gel electrophoresishad separated the two components, and the inhibitory
+ 250 rDilution
- +2000
0oE ° 150
-0 E
c.. + 100
o + 50o
_0 -
Q 'l00o -
~a- 50-
Heating
1)00p1 50,41 IOO/LI 50l 20/LI OO/I1 tOO/LI
Whole -globulin Whole y-globulinserum froction serum fraction
FIGURE 7 Effects of diluting and heating both serum andthe inhibitory fractions derived from electrophoresis (gammaregion) on porphyrin formation. Each result represents themean effect from five replicate vials. The effects of dilutionare shown on the left, and of heating, on the right.
component gave no evidence of contamination with stim-ulatory activity.
Stability of factors. Serum inhibitory and stimula-tory activities were unaltered after three repeated cyclesof freezing and thawing, and remained stable at 0°C forat least 1 yr.
DISCUSSIONThese studies show that human sera contain separablefactors which respectively stimulate and inhibit porphyrinformation in cultured chick embryo liver cells. The fac-tor responsible for porphyrin stimulation in most serawas heat stable, insensitive to trypsin, present in super-nates after ethanol precipitation of plasma proteins,absent in the precipitates, extractable in ethyl acetate,not extractable in less polar organic solvents, not dialyz-able, and migratory with the albumin fraction of serumon electrophoresis. The factor responsible for inhibitoryactivity was heat labile, sensitive to trypsin, resistantto neuraminidase, present in the precipitates after ethanolprecipitation of serum proteins, absent in the supernates,nondialyzable, and migratory with the gamma fractionof serum on electrophoresis. The stimulatory componentdoes not appear to be a protein, unless it is a proteinthat is soluble in ethanol and insensitive to trypsin.However, since the stimulatory component was presentin the albumin fraction after electrophoresis and couldnot be dialyzed, it is probably normally bound to aserum protein. Since ethanol extracts as well as thealbumin-containing fraction of sera stimulated porphyrinformation, the factor responsible for the stimulation maybe active either in the free or protein-bound state.
Strand and Marver (20) reported that normal serumacquired the capacity to stimulate porphyrin formationafter f-glucuronidase hydrolysis, indicating that sub-stances in sera which stimulate porphyrins may be con-jugated to glucuronic acid and that the stimulatoryfactor is probably inactive in the conjugated state.Together, our data and those of Strand and Marversuggest that the stimulator may be a compound of smallmolecular weight, normally conjugated and proteinbound. The inhibitory factor, on the other hand, ap-pears to be a protein or a substance requiring tightbinding to a protein for activity.
There have been no previous reports of substances inhuman sera that inhibit liver cell porphyrin formation.Sardesai and Orten (21) found an inhibitor of por-phyrin formation by acetone powder preparations ofbeef liver in the plasma of dogs. Like the inhibitoryfactor we have observed in human sera, the canine factorwas heat labile.
Substances that inhibit erythroid heme synthesis inhuman sera have been reported. The sera of certainpatients with red cell aplasia (22) have been found to
Stimulators and Inhibitors of Hepatic Porphyrin Formation in Human Sera 1175

contain gamma globulin inhibitors of erythropoiesiswhich have recently been shown to act directly onerythroid cells (23, 24). It is possible that the inhibitorof heme synthesis in erythroid cells, and the inhibitorof porphyrin accumulation in hepatic cells (the latteralso associated with the gamma globulin fraction ofsera) may be related. Inhibitors of ervthropoietic henleformation have also been reported in sera from individut-als at high altitudes (25), newborn babies (26), anemicrats (27), liypertransfused rats (28), and in homoge-nates of rabbit liver, kidney, and spleen (29); how-ever most have not been characterized. The last factormentioned (29) was heat stable, which distinguishes itfrom the serum inhibition of porphyrin formation wehave observed.
The pronounced inhibition of porphyrin formationby post partum sera is not, in all likelihood, attributableto prolactin, since human prolactin falls to prepregnantlevels within 1 or 2 wk post partumn (30), wlhile stronginhibition of porphyrin accumulation persiste(d for atleast 6 wk post partumn, in both lactatinig anid non-lactating women. MIoreover, bovine prolactin added toliver cell cultures did not inhibit porphyrin formation.The possibility that the serum factor that inhlibits por-phyrin formation may be an antibody whose productioniis increased post partuhn must be consideredl in light ofthe association of inhibitory activity witlh the serumgammaglobulins. Elevated levels of IgM in sera of sev-eral post partum women have been reported (31). Theprocesses occurring during uterine involution in humansare not well understood, and it is also possible that theinhibitory factor may be a myoglobin derivative or an-other compound produced as a result of uterine involu-tion.
Post partum sera inhibited porphyrin formation incontrol cultures, inhibited the stimulation of porphyrinformation by AIA and other drugs, and inhibited ac-tivity of ALAS. They also caused some inhibition ofthe conversion of ALA to porphyrins, indicating thatthe inhibitory effect was not exerted solely at the levelof ALAS. The inhibitory factor could result in inhibi-tion of porphyrin formation by: (a) directly affectingboth ALAS and a subsequent step in porphyrin biosyn-thesis, (b) binding to both ALA and chemical inducingagents, or (c) affecting cellular metabolism in a waythat only indirectly results in alteration of porphyrinformation. Whatever their mode of action may be, theserum factors studied resulted in gross alterations ofporphyrin formation by hepatic cells. Since the factorshave the capacity to modify heme biosynthesis, theiroccurrence in serum would seem to have clinical im-plications.
More than 200 chemical agents are known whichstimulate hepatic porphyrin or heme formation or both(2-8, 32). Normal individuals may develop altered ca-
pacity to metabolize drugs as a consequence of ex-posure to such agents, while certain genetically sus-ceptible individuals, such as porphyrics, may developsevere clinical symptoms. The factors involved in thenormal and abnormal regulation of hepatic heme forma-tion are not yet well uinderstoo(l. The serutn componentthat inihibits porphyrin formiiation may mo(llllate the ef-fect of inducers of ALAS present in the circulation inexcess and thus re(duce increases in porphyrini formationiin response to the drugs and chemicals to which allpeople are commonly exposed.
The number of porphyric subjects studied here was toosmall to draw conclusions about the role of the factorsin the porphyrias. Although stimulatory activity ap-pears to be increased in sera from patients with bothAIP and PCT, high levels of stimulatory activity andclinical manifestations are not always related. Althougha substance with the capacity to inhibit porphyrin for-mation could be detected in the sera of most porphyricswe studied, two subjects xvith AIP and three with PCTseemed to lack the inhibitory component. It is possiblethat porphyrias are .heterogeneous with respect to oc-currence of the stimulatory and inhibitory factor and thatthe absence of the inhlibitory factor may play a patho-genetic role in certain patients with the disorders. Astudy of the effect onl porphyrin formation of serumfrom relatives of porphyria patients might show whetherthe occurrence of the factors was genetically deter-mined and might aid in the understanding of the bio-chemical phenomenology of the porphyrias. It is alsopossible that a component in serum that can inhibitporphyrin formation may eventually prove useful in thetherapeutic management of certain cases of porphyria.
ACKNOWLEDGMENTS
We thank Judith Fitzpatrick, Jan Curry, and Enid Sternfor their valuable technical assistance.
This work was supported by U. S. P. H. S. Grant HR04313, Special Fellowship FO 3 HD 40904, American CancerSociety Award BC-98, and New York City Health Re-search Council Grant U-2286.
REFERENCES
1. Kappas, A., C. S. Song, S. Sassa, R. D. Levere, andS. Granick, 1969. The occurrence of substances in hu-man plasma capable of inducing the enzyme 8-amino-levulinate synthetase in liver cells. Proc. Natl. Acad.Sci. U. S. A. 64: 557.
2. Granick, S. 1966. The induction in vitro of the syn-thesis of 8-aminolevulinic acid synthetase in chemicalporphyria. A response to certain drugs, sex hormonesand foreign chemicals. J. Biol. Chemii. 241: 1359.
3. Granick, S., and A. Kappas. 1967. Steroid inductionof porphyrin synthesis in liver cell culture. I. Struc-tural basis and possible physiological role in the con-trol of heme formation. J. Biol. Chem. 242: 4587.
4. Racz, W. J., and G. S. Marks. 1972. Drug-induced
1176 Rifkind, Sassa, Merkatz, Winchester, Harber, and Kappas

porphyrin biosynthesis. IV. Investigation of the differ-ences in response of isolated liver cells and the liverof the chick embryo to porphyria-inducing drugs. Bio-chem. Pharmacol. 21: 143.
5. Kappas, A., C. iS. Song, R. D. Levere, R. A. Sachson,and S. Granick. 1968. The induction of 5-aminolevu-linic acid synthetase in vivo in chick embryo liver bynatural steroids. Proc. Natl. Acad. Sci. U. S. A. 61:509.
6. Rif kind, A. B., P. N. Gillette, C. S. Song, and A.Kappas. 1970. Induction of hepatic 5-aminolevulinic acidsynthetase by oral contraceptive steroids. J. Clin. En-docrinol. Metab. 30: 330.
7. Wada, O., Y. Yano, G. Urata, and K. Nakao. 1968.Behavior of hepatic microsomal cytochromes aftertreatment of mice with drugs known to disturb porphy-rin metabolism in liver. Biochem. Pharmacol. 17: 595.
8. Rif kind, A. B., P. N. Gillette, C. S. Song, and A.Kappas. 1973. Drug stimulation of 8-aminolevulinicacid synthetase and cytochrome P-450 in vivo in chickembryo liver. J. Pharmacol. Exp. Ther. 185: 214.
9. Koskelo, P., A. Eisalo, and I. Toivonen. 1966. Urinaryexcretion of porphyrin precursors and coproporphyrinin healthy females on oral contraceptives. Br. Med. J.1: 652.
10. Marver, H. S., and R. Schmid. 1972. The porphyrias.In Metabolic Basis of Inherited Disease. J. B. Stan-bury, J. B. Wyngaarden, and D. S. Fredrickson, edi-tors. McGraw-Hill Book Co., New York. 3rd edition.1087.
11. Goldberg, A., M. R. Moore, A. D. Beattie, P. E. Hall,J. McCallum, and J. K. Grant. 1969. Excessive urinaryexcretion of certain porphyrinogenic steroids in humanacute intermittent porphyria, Lancet. 1: 115.
12. Kappas, A., H. L. Bradlow, P. N. Gillette, and T. F.Gallagher. 1972. Studies in porphyria. I. A defect inthe reductive transformation of natural steroid hor-mones in the hereditary liver disease, acute intermittentporphyria. J. Exp. Mled. 136: 1043.
13. Poland, A., and E. Glover. 1973. 2,3,7,8-Tetrachlorodi-benzo-p-dioxin: a potent inducer of 8-aminolevulinicacid synthetase. Science (Wash. D. C.). 179: 476.
14. Kissane, J. M., and E. Robins. 1958. The fluorimetricmeasurement of deoxyribonucleic acid in animal tissueswith special reference to the central nervous system.J. Bio. Chem. 233: 184.
15. Levere, R. D., F. Swerdlow, and M. R. Garavoy.1971. Measurement of human plasma hemoglobin bydifference spectrophotometry. J. Lab. Clin. Med. 77:168. 1
16. Albert, A., J. Kobi, J. Leiferman, and I. Derner. 1961.Purification of pituitary gonadotropin from urine ofnormal men. J. Clin. Endocrinol. Metab. 21: 1.
17. Miiller-Eberhard, H. J. 1960. A new supporting me-
dium for preparative electrophoresis. Scand. J. Clin.Lab. Invest. 12: 33,
18. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folinphenol reagent. J. Biol. Chenii. 193: 265.
19. Van Hall,- E. V., J. L. Vaitukaitis, G. T. Ross, J. W.Hickman, and G. Ashwell. 1971. Immunological andbiological activity of HCG following progressive de-sialyation. Endocrinology. 88: 456.
20. Strand, L. J., and H. Marver. 1970. Determination ofA-aminolevulinic acid synthetase (ALA-S) in cell cul-ture: naturally occurring inducers in normal humanplasma. Clin. Res. 18: 345.
21. Sardesai, V. M., and J. M. Orten. 1965. Role of aplasma factor in porphyriti biosynthesis. Proc. Soc.Exp. Biol. Med. 120: 552.
22. Krantz, S. B., and V. Kao. 1967. Studies on red cellaplasia, I. Demonstration of a plasma inhibitor toheme synthesis and an antibody to erythroblast nuclei.Proc. Natl. Acad. Sci. U. S. A. 58: 493.
23. Zaentz, S. D., and S. B. Krantz. 1972. Demonstrationof cytolytic antibody specific for erythroblasts in purered cell aplasia (PRCA). Clin. Res. 20: 504.
24. Zalusky, R., E. D. Zanjani, A. S. Gidari, and J. Ross.1973. Site of action of a serum inhibitor of erythro-poiesis. J. Lab. Clin. Med. 81: 867.
25. Reynafarje, C., J. Ramos, J. Faura, and D. Villa-viencio. 1964. Humoral control of erythropoietic ac-tivity in man during and after altitude exposure. Proc.Soc. Exp. Biol. Med. 116: 649.
26. Skjaelaaen, P., and S. Halvorsen. 1971. Inhibition oferythropoiesis by plasma from newborn infants. ActaPaediatr. Scatnd. 60: 301.
27. Molinari, P. F., F. F. Menninger, Jr., and H. Rosen-krantz. 1970. Detection of an inhibitor of hemoglobinsynthesis in rat plasma. J. Lab. Clin. Med. 76: 466.
28. Whitcomb, W. H., and M. Z. Moore. 1965. The inhibi-tory effect of plasma from hypertransfused animals onerythrocyte iron incorporation in mice. J. Lab. Clin.Med. 66: 641.
29. Erslev, A. J., and L. A. Kazal. 1968. Inactivation oferythropoietin by tissue homogenates. Proc. Soc. Exp.Biol. Med. 129: 845.
30. Hwang, P., H. Guyda, and H. Friesen. 1971. A radio-immunoassay for human prolactin. Proc. Natl. Acad.Sci. U. S. A. 68: 1902.
31. Gusdon, J. P., Jr. 1969. Fetal and maternal immuno-globulin levels during pregnancy. Am. J. Obstet. Gy-necol. 103: 895.
32. Conney, A. H. 1971. Environmental factors influencingdrug metabolism. In Fundamentals of Drug Metabolismand Drug Disposition. B. N. LaDu, H. G. Mandel, andE. L. Way, editors. The Williams & Wilkins Co.,Baltimore. 253.
Stimulators and Inhibitors of Hepatic Porphyrin Formation in Human Sera 1177


![19. FINAL Novocure- PIOM 10.4.15[1] - Food and Drug ... of active electronic devices include deep brain stimulators, spinal cord stimulators, vagus nerve stimulators, pacemakers, defibrillators](https://img.dokumen.tips/doc/110x75/5b0be9927f8b9ae61b8eae77/19-final-novocure-piom-104151-food-and-drug-of-active-electronic-devices.jpg)


























