Embed Size (px)
Citation preview

7
SADRŽAJ / CONTENTS
Uvodna predavanja / Introductory lecture
VIRUSNE INFEKCIJE PERADI – GLOBALNI PROBLEM VIRAL INFECTIONS OF POULTRY – THE GLOBALLY CHALLENGING SITUATION Vladimir Savić ................................................................................................................................................. 11
IZAZOVI I OČEKIVANJA EU I PERADARSTVA HRVATSKE U SREDNJOROČNOM RAZDOBLJU DO 2030. GODINE CHALLENGES AND EXPECTATIONS OF THE EU AND CROATIAN POULTRY IN THE MEDIUM TERM BY 2030 Branko Bobetić ............................................................................................................................................... 17
DJELATNOSTI LABORATORIJA CENTRA ZA PERADARSTVO TIJEKOM 2017. I 2018. GODINE ACTIVITIES OF THE POULTRY CENTRE LABORATORIES IN 2017 AND 2018 Tajana Amšel Zelenika, Vladimir Savić, Borka Šimpraga, Marina Tišljar, Mirta Balenović, Marijana Sokolović, Tihomir Zglavnik, Fani Krstulović, Luka Jurinović, Marija Berendika ............................................ 24 Usmena izlaganja / Lectures
CHARACTERISTICS OF AVIAN BOTULISM TYPE C OUTBREAKS IN COMMERCIAL BROILERS IN SLOVENIA ZNAČAJKE POJAVE PTIČJEG BOTULIZMA TIPA C U TOVNIH PILIĆA U SLOVENIJI Olga Zorman Rojs, Rahela Juršič Cizerl, Liljana Štalcer, Tea Poljanec, Brigita Slavec ................................ 33
ZNAČENJE BAKTERIJA RODA CHLAMYDIA SP. U JATIMA PERADI I NJIHOV ZOONOTSKI POTENCIJAL SIGNIFICANCE OF CHLAMYDIA SP. IN POULTRY FLOCKS AND ITS ZOONOTIC POTENTIAL Danijela Horvatek Tomić, Ana Marquiza Quilicot, Željko Gottstein, Maja Lukač, Estella Prukner-Radovčić . 38
POJAVNOST OŠTEĆENJA PRSNIH KOSTI KOKOŠI NESILICA U ALTERNATIVNOM UZGOJU I OBOGAĆENIM KAVEZIMA THE PREVELENCE OF KEEL BONE DAMAGE IN LAYING HENS IN ALTERNATIVE AND ENRICHED CAGES REARING Zlatko Janječić, Zlata Kralik, Dalibor Bedeković ............................................................................................. 44
FIELD EXPERIENCES CONCERNING THE LONG TERM USE OF MS-H AND MS LIVE VACCINES ON PARENT STOCK FARM IN HUNGARY TERENSKA ISKUSTVA S DUGOTRAJNOM PRIMJENOM ŽIVIH CJEPIVA MS-H I MS NA FARMI RODITELJSKOG JATA U MAĐARSKOJ László Kőrösi, János Povazsán, Attila Jacsó, Chris Morrow, Miklós Gyuranecz ........................................... 49
ALTERNATIVNO LIJEČENJE BOLESTI UZROKOVANE PROTOZOOM HISTOMONAS MELEAGRIDIS U ZAGORSKOG PURANA ALTERNATIVE TREATMENT OF PROTOZOAN DISEASE CAUSED BY HISTOMONAS MELEAGRIDIS IN ZAGORJE TURKEY Tihomir Zglavnik, Tajana Amšel Zelenika, Marina Tišljar, Borka Šimpraga, Željko Kolar, Anđelko Ferek Jambrek .......................................................................................................................................................... 52

8
INAKTIVACIJA MIKOTOKSINA U HRANI ZA ŽIVOTINJE INACTIVATION OF MYCOTOXINS IN ANIMAL FEED Marijana Sokolović .......................................................................................................................................... 58
SOJA U HRANI ZA PERAD SOYBEAN IN POULTRY FEED Marija Berendika, Marijana Sokolović, Tihomir Zglavnik, Gabrijela Krivec .................................................... 69
UČINAK DODATKA ALUVIJALNOG DEPOZITNOG HOLOCENSKOG MINERALA (ADNEM) U KRMU TOVNIH PILIĆA FARMSKOG UZGOJA NA TVORBU KOLAGENA U NJIHOVU MIŠIĆJU EFFECT OF ADDING ALLUVIAL DEPOSIT HOLOCENE MINERALS (ADNEM) TO FARM BREEDING BROILER FEED ON COLLAGEN FORMATION IN THEIR MUSCLES Hrvoje Mazija, Zlatko Janječić, Helga Medić, Dalibor Bedeković, Mirta Balenović, Marijan Andrašec, Bratko Filipič ............................................................................................................................................................... 73
INFEKCIJA KONZUMNIH NESILICA BAKTERIJOM MANNHEIMIA HAEMOLYTICA LAYING HEN INFECTION WITH MANNHEIMIA HAEMOLYTICA Borka Šimpraga, Fani Krstulović, Luka Jurinović, Marina Tišljar, Marijana Sokolovć .................................... 83
ASSESSMENT OF MOLECULAR DIVERSITY OF SOME POTENTIAL PATHOGENIC E. COLI ISOLATED BY ERIC-PCR SCREENING PROCJENA MOLEKULARNE RAZNOVRSNOSTI NEKIH POTENCIJALNO PATOGENIH IZOLATA E. COLI POMOĆU ERIC-PCR PROBIRA Camelia Tulcan, Marijana Sokolović, Sorina Popescu, Ioan Hutu, Simona Marc, Oana Maria Boldura, Calin Mircu ...................................................................................................................................................... 90
PROIZVODNI POKAZATELJI NAKON PRIMJENE AUTOGENOG CJEPIVA PROTIV PTIČJIH PATOGENIH SOJEVA E. COLI I POBOLJŠANJA PROGRAMA CIJEPLJENJA U JATIMA TEŠKE LINIJE KOKOŠI PRODUCTION PARAMETERS AFTER AUTOGENOUS APEC E. COLI VACCINE APPLICATION AND IMPROVEMENT OF VACCINATION PROGRAM IN BROILER BREEDER FLOCK Željko Gottstein, Liča Lozica, Danijela Horvatek Tomić, Maja Lukač, Estella Prukner-Radovčić, Metka Vlahek, Nada Dolenčić, Željko Glavak ........................................................................................................... 98
RAPID DIFFERENTIATION OF INFECTIOUS BRONCHITIS VIRUS STRAINS BY RESTRICTION FRAGMENT LENGTH POLYMORPHISM ANALYSIS BRZA METODA IDENTIFIKACIJE I DIFERENCIJACIJE SOJEVA VIRUSA PTIČJEG ZARAZNOG BRONHITISA METODOM PCR-RFLP Katarina Huić Babić, Leonida Kutle, Lana Ljuma Skupnjak, Anto Vrdoljak .................................................... 104
PROVJERA SIGURNOSTI I UČINKOVITOSTI CJEPIVA PROTIV ZARAZNE BOLESTI BURZE PREMA ZAHTJEVIMA EUROPSKE FARMAKOPEJE SAFETY AND EFFICACY ASSESSMENT OF THE VACCINE AGAINST INFECTIOUS BURSAL DISEASE ACCORDING TO THE EUROPEAN PHARMACOPOEIA REQUIREMENTS Anto Vrdoljak, Olga Zorman Rojs, Gert Jan Boelm, Lana Ljuma Skupnjak ................................................... 113

9
DEVELOPMENT OF LIVE ATTENUATED VACCINE AGAINST INFECTIOUS BRONCHITIS VIRUS ACCORDING TO CURRENT EUROPEAN REGULATIVE RAZVOJ ŽIVOG ATENUIRANOG CJEPIVA PROTIV PTIČJEG ZARAZNOG BRONHITISA U SKLADU S EUROPSKOM REGULATIVOM Lana Ljuma Skupnjak, Gert Jan Boelm, Anto Vrdoljak .................................................................................. 120 Poster prezentacije / Poster presentations
UTJECAJ VISINE STELJE NA JAČINU KONTAKTNOG DERMATITISA NA JASTUČIĆIMA NOGU I TARZALNIM ZGLOBOVIMA TOVNIH PILIĆA EFFECT OF LITTER DEPTH ON SEVERITY OF FOOTPAD DERMATITIS AND HOCK BURNS IN BROILERS Ivana Sabolek, Srebrenka Nejedli, Ivan Vlahek, Kristina Matković, Sven Menčik, Aneta Piplica, Željko Pavičić, Mario Ostović .................................................................................................................................... 130
UČINAK GUSTOĆE NASELJENOSTI NA KONFORMACIJU TOVNIH PILIĆA EFFECT OF STOCKING DENSITY ON BODY CONFORMATION OF BROILERS Almira Softić, Aida Kavazović, Aida Kustura, Velija Katica, Behija Dukić, Amina Hrković-Porobija, Maja Varatanović, Ajla Ališah .................................................................................................................................. 135
PRISUTNOST RODA CAMPYLOBACTER U KOŽAMA VRATOVA TOVNIH PILIĆA TIJEKOM LJETNOG RAZDOBLJA 2018. GODINE PRESENCE OF THE GENUS CAMPYLOBACTER IN BROILER NECK SKIN DURING THE SUMMER PERIOD OF 2018 Sanja Furmeg, Vesna Jaki Tkalec, Maja Kiš, Jadranka Sokolović, Darko Majnarić, Željko Cvetnić ............. 140
UTJECAJ GODIŠNJEG RAZDOBLJA NA POJAVNOST GLJIVICA U HRANI ZA TOVNE PILIĆE SEASONAL IMPACT ON THE OCCURRENCE OF FUNGI IN BROILER FEED Ivica Ravić, Mario Ostović, Matija Kovačić, Anamaria Ekert Kabalin, Kristina Matković, Ivana Sabolek, Željko Pavičić, Ivona Žura Žaja, Hrvoje Valpotić, Danijela Horvatek Tomić ................................................... 145
PREVALENCE OF GALLIBACTERIUM ANATIS AND ESCHERICHIA COLI ON BROILER BREEDER FARMS IN CROATIA PROŠIRENOST BAKTERIJA GALLIBACTERIUM ANATIS I ESCHERICHIA COLI NA FARMAMA RODITELJSKIH JATA TEŠKIH LINIJA KOKOŠI U HRVATSKOJ Liča Lozica, Danijela Horvatek Tomić, Željko Gottstein ................................................................................. 150
ISPITIVANJE PRISUTNOSTI SALMONELLA SPP. U UZORCIMA SVJEŽEG PORCIONIRANOG PILEĆEG MESA U MALOPRODAJI TESTING FOR SALMONELLA SPP. PRESENCE IN RETAIL FRESH PORTIONED CHICKEN MEAT SAMPLES Zdenko Mlinar, Ivančica Kovaček, Ana Vasiljević, Vedran Prahin ................................................................. 155
MONITORING REZIDUA VETERINARSKO-MEDICINSKIH PROIZVODA I OSTALIH TVARI U PERADI I JAJIMA U EUROPSKOJ UNIJI U RAZDOBLJU 2009. - 2016. GODINE MONITORING OF VETERINARY MEDICINAL PRODUCT RESIDUES AND OTHER SUBSTANCES IN POULTRY AND EGGS IN THE EUROPEAN UNION DURING THE 2009-2016 PERIOD Damir Pavliček, Marija Denžić Lugomer ......................................................................................................... 159

10
MIKROBIOLOŠKA KVALITETA HRANE ZA ŽIVOTINJE U REPUBLICI HRVATSKOJ MICROBIOLOGICAL QUALITY OF ANIMAL FEED IN THE REPUBLIC OF CROATIA Marijana Sokolović, Marija Berendika, Borka Šimpraga, Fani Krstulović ....................................................... 166
UTJECAJ DODATKA ETERIČNIH ULJA LIMUNA I KOMORAČA U HRANU NA TOVNA I KLAONIČKA SVOJSTVA TE SENZORSKA SVOJSTVA MESA TOVNIH PILIĆA ADDITION OF LEMON AND FENNEL ESSENTIAL OIL TO FEED INFLUENCES BROILER PERFORMANCE, CARCASS COMPOSITION AND SENSORY MEAT QUALITY Brigita Hengl, Gordana Kralik, Slobodan Lilić, Estella Prukner Radovčić, Mislav Đidara, Marcela Šperanda 172

PERADARSKI DANI 2019. 11
VIRUSNE INFEKCIJE PERADI – GLOBALNI PROBLEM
Vladimir Savić
Hrvatski veterinarski institut, Centar za peradarstvo, Zagreb, Hrvatska Sažetak Peradarska industrija se suočava s brojnim izazovima, među kojima su virusne infekcije od posebne važnosti. Virusi se mogu brzo širiti, nisu osjetljivi na antibiotike i uzrokuju enormne gospodarske gubitke u peradarskoj industriji širom svijeta. Zajednička tržišta i prisutnost međunarodnih peradarskih tvrtki omogućavaju lakše i brže širenje virusnih infekcija peradi u različite zemlje, regije pa čak i kontinente. Uz to, nedavna prelijevanja određenih virusa iz peradi u populacije ptica selica dovela su do nepredvidivih pojava bolesti u udaljenim peradarskim pogonima. U radu su razmotrene najvažnije virusne bolesti koje danas predstavljaju velik izazov za peradarsku industriju. Niti jedna od njih dosad nije stavljena pod zadovoljavajuću kontrolu na globalnoj razini, naprotiv, sve više se pojavljuju jače patogeni ili antigenski varijantni virusni sojevi. Cijepljenje je moguće za mnoge virusne infekcije peradi, ali ono ne jamči uvijek i potrebnu zaštitu. Nedvojbeno je da pravilna uporaba cjepiva za perad putem odgovarajućih programa cijepljenja ima dragocjen učinak na kontrolu i prevenciju virusnih infekcija, a time i na smanjenje gospodarskih gubitaka. Ipak, loše planirana i nepravilno provedena cijepljenja mogu potaknuti pojavu i pospješiti prijenos virulentnih sojeva virusa. Također, cijepljenje ne može biti zamjena za biosigurnost i održavanje higijene, iako postoje takve zablude. Visoka konkurentnost i zahtjevi za nižim troškovima proizvodnje peradi promijenili su sustave upravljanja, što je rezultiralo smanjenim naglaskom na biosigurnost i druge osnovne mjere sprječavanja zaraznih bolesti. Stoga se može zaključiti da su današnji sustavi upravljanja peradarskom proizvodnjom koji zanemaruju temeljna načela kontrole i iskorjenjivanja bolesti u velikoj mjeri odgovorni za pojavu i širenje virusnih infekcija. Sve ovo čini virusne infekcije peradi globalno opterećujućim problemom i zahtijeva uključivanje veterinara specijaliziranih za peradarstvo kako u razmatranje bolesti tako i u upravljačka tijela još u fazi planiranja novih peradarskih pogona. Ključne riječi: influence ptica, newcastleska bolest, zarazni bronhitis, zarazna bolest burze, gumborska bolest, cijepljenje, biosigurnost Uvod
Brojni su izazovi s kojima se suočava peradarska industrija, bilo izravno kroz učinkovitost i ekonomičnost proizvodnje mesa i jaja ili neizravno kroz tržišne probleme, sigurnost hrane i javno zdravstvo, percepciju potrošača, dobrobit životinja, povećanje zakonskih zahtjeva i tako dalje. Pojava i širenje zaraznih bolesti imaju golem utjecaj na peradarsku industriju i predstavljaju ozbiljnu prijetnju zdravlju ljudi i divljih ptica (Wang i sur., 2013.). Naročito zabrinjavaju virusne infekcije. Virusi se mogu brzo širiti, nisu osjetljivi na antibiotike i uzrokuju enormne gospodarske gubitke u peradarskoj industriji širom svijeta. Cijepljenje je moguće za mnoge virusne infekcije peradi, ali ono ne jamči uvijek i potrebnu zaštitu. U mnogim slučajevima, čak i kada je zaštićena od kliničke pojave bolesti, cijepljena i naizgled zdrava perad može, poput trojanskog konja, prenositi terenski virus potajno šireći infekciju.
Peradarska proizvodnja prednjači pred svim poljoprivrednim granama u integraciji prema većim proizvodnim jedinicama i korporativnim tvrtkama. Zajedno s osnivanjem i nasta-janjem međunarodnih tvrtki i tržišta, ovo rezultira lakšim i bržim širenjem virusnih infekcija peradi na više peradarskih farmi, regija, zemalja pa i čak kontinenata. Uz to, nedavna prelijevanja određenih virusa iz peradi u populacije ptica selica rezultirala su nepredvidivim izbijanjima infekcija u udaljenim peradarskim pogonima. U ovom radu su raz-motrene najvažnije virusne bolesti koje danas predstavljaju najveći izazov za peradarsku industriju.
Influenca ptica
Virusi influence ptica (IP) su vrlo zarazni, izrazito promje-njivi virusi koji su široko rasprostranjeni u pticama. Virusi influence koji inficiraju perad mogu se podijeliti u dvije skupine. Iznimno virulentni virusi uzrokuju visoko patogenu
doc. dr. sc. Vladimir Savić, dr.med.vet., Hrvatski veterinarski institut, Centar za peradarstvo, Laboratorij za virusologiju i serologiju, Heinzelova 55, 10000 Zagreb, Hrvatska; e-mail: [email protected]

VIRUSNE INFEKCIJE PERADI – GLOBALNI PROBLEM 12
influencu ptica (VPIP), a smrtnost zaraženih jata je i do 100%. Ti virusi su ograničeni na podtipove H5 i H7, iako svi virusi H5 i H7 ne uzrokuju VPIP. Svi ostali virusi IP uzrokuju blažu, prije svega respiratornu bolest, osim ako se simptomi pogoršaju zbog sekundarnih infekcija (Capua i Alexander, 2009.). Virusi IP mogu zaraziti sve vrste peradi i zatočenih ptica u svim dijelovima svijeta. Učestalost pojave primarne infekcije obično ovisi o stupnju kontakta s divljim pticama. Sekundarno širenje je obično povezano s ljudskim aktivnostima, bilo preseljavanjem peradi i njihovih proiz-voda ili prijenosom infektivnog fecesa sa zaraženih na osjetljiva jata, ali u sekundarnom širenju potencijalno mogu biti uključene i divlje ptice (Alexander i Capua, 2008.). Donedavno su se virusi VPIP rijetko izolirali iz divljih ptica, ali se situacija s VPIP podtipa H5 dramatično promijenila u proteklih 15-ak godina. Velika rasprostranjenost virusa VPIP podtipa H5N1 u peradi na Dalekom Istoku neminovno je rezultirala prelijevanjem ovoga virusa u populacije divljih ptica u svibnju 2005. (Alexander i Capua, 2008.) te postoje čvrsti dokazi da ga divlje ptice mogu širiti na velike udaljenosti (Normile, 2006.). Ovaj se virus proširio tijekom 2005. i 2006. godine na perad i divlje ptice u velikom broju europskih i afričkih zemalja (Salzberg i sur., 2007.; Hesterberg i sur., 2009.). Slična situacija se ponovila i 2014. godine kada se virus VPIP podtipa H5N8 širio divljim pticama selicama s Dalekog Istoka u Europu i u Sjevernu Ameriku (Lee i sur., 2015.). Dalekoistočni virus podtipa H5N8 i njegove lokalne presloženice izazvali su najgoru epizootiju IP u povijesti SAD-a, što je rezultiralo uništenjem više od 47 milijuna kokoši, purana i drugih komercijalno uzgojenih ptica s procijenjenim gospodarskim gubitkom na više od tri milijarde dolara samo u SAD-u. Nakon epizootija H5N1 (2005.-2006.) i H5N8 (2014.-2015.), drugi virus VPIP također podtipa H5N8 je 2016. godine unesen divljim ptica selicama u Europu. Do kolo-voza 2017. zabilježeno je 1.112 pojava ovoga virusa u peradi i 955 u divljih ptica u 30 europskih zemalja. Radilo se o najvećoj ikad zabilježenoj epizootiji virusa VPIP na europ-skom kontinentu s obzirom na broj izbijanja bolesti u peradi, zemljopisni opseg i broj mrtvih divljih ptica. (Brown i sur., 2017.; Napp i sur., 2018.). Virus je nastavio cirkulirati u peradi u pojedinim područjima u Europi čak i 2019. godine. U budućnosti se mogu očekivati nova unošenja virusa VPIP divljim pticama selicama iz udaljenih područja s poslije-dičnim infekcijama peradi. Iako niskopatogeni virusi IP uzrokuju blažu bolest, infekcija peradi virusom podtipa H9N2 je rezultirala velikim gubi-cima tijekom posljednjih nekoliko desetljeća. Od sredine 1990. godine ova infekcija je zabilježena u peradi u nekoliko europskih zemalja, Južnoj Africi, SAD-u, Južnoj Koreji i Kini, a infekcija je nedavno postala raširena i endemična u peradi širom Azije, Bliskog Istoka i Sjeverne Afrike (Capua i Alexander, 2009.). S obzirom na to da ne postoji obvezan nadzor peradi na viruse IP osim za podtipove H5 i H7, tre-nutno postoji ozbiljan rizik od unosa, širenja i ukorjenjivanja virusa H9N2 u peradi u zemljama koje su trenutno slobodne od ove infekcije. Cijepljenje protiv IP je moguće i danas se uglavnom cijepi protiv virusa H9N2, no cijepljenje protiv
podtipova H5 i H7 nije iznimka, osobito u borbi protiv virusa VPIP. Međutim, cijepljenje protiv IP je složeno i zahtjevno zbog velike raznolikosti podtipova influence pa čak i zbog značajnih antigenskih razlika među virusima istoga podtipa. Ovo može rezultirati slabom zaštitom čak i nakon pravilne uporabe cjepiva istoga podtipa kao što je i terenski virus. Optimalno cijepljenje protiv IP pravilno primijenjenim adekvatnim cjepivima spriječit će pojavu kliničkih znakova i uginuće, umanjit će razinu i trajanje izlučivanja terenskog virusa te povećati otpornost prema infekciji na način da će biti potrebna veća količina virusa da bi se cijepljene ptice inficirale (Capua i sur., 2004.). Međutim, moguće je da će suboptimalno cijepljenje umanjiti ili spriječiti pojavu bolesti, ali neće umanjiti širenje terenskog virusa te će doći do ukorijenjene bolesti (Webster i sur., 2006.), osobito ako se loše provode ili se ne provode ostale mjere za kontrolu IP. Zbog toga bi cijepljenje trebalo biti samo dio strategije kontrole i suzbijanja IP, koja mora uključivati poboljšanu biosigurnost i otkrivanje jata izlo-ženih terenskom virusu unutar cijepljene populacije (Capua i Marangon, 2007.). Određeni virusi IP imaju značajan zoonotski potencijal iako nijedan virus IP nije prilagođen ljudima i dosad ne postoji učinkovit prijenos s čovjeka do čovjeka. Većina osoba koje su se zarazile virusom IP bila je u dodiru sa zaraženom pe-radi, uglavnom u enzootskim područjima, no zaražavanje ljudi putem mesa peradi i jaja dosad nije zabilježeno. Ipak, percepcija potrošača često može biti suprotna. Na primjer, virus VPIP podtipa H5N1 je tijekom 2005. i 2006. godine u Hrvatskoj otkriven isključivo u divljim pticama, no ovo je imalo značajan utjecaj na lokalnu peradarsku industriju zbog smanjenja potrošnje mesa peradi za čak 20% na vrhuncu krize (Savić i sur., 2007.).
Newcastleska bolest
Newcastleska bolest (NB) je još jedna pogubna bolest peradi koja uzrokuje smrtnost do 100% u potpuno osjetljivim jati-ma kokoši. Nasuprot VPIP, gotovo da nema komercijalnog jata peradi na koje NB nema utjecaja, barem zbog mjera kontrole i zaštite protiv ove bolesti. U mnogim područjima gdje se perad komercijalno uzgaja kontrola NB se oslanja na cijepljenje, ali ova bolest ipak predstavlja velik ograničava-jući čimbenik za razvoj peradarske proizvodnje u mnogim zemljama (Alexander, 2001.). Kao i za VPIP, gospodarska važnost NB se ne odnosi samo na izravne gubitke u zara-ženim jatima te troškove cijepljenja, nego i na utjecaj koji može nastati zbog trgovinskih ograničenja i embarga na područja i zemlje u kojima su se pojavile epizootije ove bolesti (Aldous i Alexander, 2001.). Iako su takva ogra-ničenja općenito opravdana, u mnogim slučajevima nisu vođena epizootiološkim razlozima, već predstavljaju poli-tičku odluku zemlje uvoznice. Svjedoci smo slučajeva kada zemlje s ukorijenjenom NB zabranjuju uvoz peradi i proiz-voda od peradi iz zemlje s ograničenim izbijanjem NB koja je bolest odmah stavila pod kontrolu uklanjanjem zaražene peradi i potpuno odvojenim držanjem i proizvodnjom zdrave peradi.

VIRUSNE INFEKCIJE PERADI – GLOBALNI PROBLEM 13
Bolest je stalno prisutna u mnogim zemljama širom svijeta, a u nekima uzrokuje sporadične pojave ili iznenadne epizootije, dok je u drugim zemljama ukorijenjena. Za NB, uz još nekoliko drugih virusnih bolesti peradi, obvezna je prijava Svjetskoj organizaciji za zdravlje životinja (Inter-national International Epizooties - OIE). Samo dvije virusne bolesti peradi, VPIP i NB, su na bivšoj listi A koja pred-stavlja zarazne bolesti od velike važnosti u međunarodnoj trgovini životinjama i životinjskim proizvodima. Unatoč tome, čini se da se NB jako malo prijavljuje OIE-u, budući da su neke zemlje formalno slobodne od ove bolesti, dok se istodobno u renomiranim znanstvenim člancima opisuju ozbiljne i dugotrajne pojave NB u tim zemljama. Nedostatak dijagnostičkih kapaciteta i praćenja ove infekcije također može biti razlog njenom neprijavljivanju međunarodnim organizacijama. Kao što je spomenuto za IP, virulentni vi-rusi mogu se zaraziti i umnožavati u cijepljenim pticama bez izazivanja kliničke bolesti (Parede i sur., 1990, Capua i sur., 1993., Guittet i sur., 1993.), a to može dovesti do ukorijenjenosti na način da cijepljenje prikriva infekciju te ovo značajno koči pasivni nadzor koji se oslanja na kliničke simptome i ugibanje zaražene peradi. Za razliku od virusa IP, virusi NB su antigenski jedinstveni i pripadaju serotipu nazvanom ptičji avulavirus 1 (Avian Avulavirus-1 ili AAvV-1; bivši ptičji paramiksovirus 1 ili aPMV-1). Zbog toga je cijepljenje daleko jednostavnije, budući da praktički ne postoji antigenska nesukladnost između terenskih virusa i cjepnih sojeva. Živa i inaktivirana cjepiva protiv NB se naširoko koriste još od 1950-ih godina. Ako se pravilno primjene zdravim jedinkama, živa i vektorska cjepiva mogu zaštititi od kliničke bolesti i uginuća uslijed infekcije virulentnim virusima NB. Za razliku od cjepiva protiv IP, dostupnost živih cjepiva protiv NB omo-gućuje masovnu primjenu u peradarskim nastambama, što podrazumijeva manje rada i troškova. Treba napomenuti da se cjepiva protiv NB razlikuju po tropizmu i sposobnosti repliciranja u neimunim jedinkama pa i izbor cjepiva treba odgovarati lokalnim epozootiološkim okolnostima. Cjepiva i cijepljenje protiv NB su nedavno u preglednom radu de-taljno prikazali Dimitrov i sur. (2017.).
Zarazni bronhitis
Zarazni bronhitis (ZB) je akutna i vrlo zarazna virusna bolest koja pogađa kokoši svih dobi. Sveprisutan je u većini svijeta gdje se perad uzgaja i smatra se najkužnijom bolesti peradi. Pobol obično doseže 100%, ali je smrtnost niska i doseže do 25% pa čak i više u pilića mlađih od šest tjedana, no obično je beznačajna u starijih jedinki (Cavanagh i Gelb, 2008.). Unatoč niskoj smrtnosti, gospodarska važnost ZB najvjerojatnije nadilazi gospodarsku važnost NB. Prema istraživanju objavljenom u Godišnjaku o zdravlju životinja OIE-a od 2006. do 2009. godine provedenom u 176 zemalja, ZB je među svim bolestima peradi na drugom mjestu po uzrokovanim gubicima, odmah iza VPIP, a slijede ga niskopatogena IP, NB i zarazna bolest burze (ZBB) (Anonymous, 2011.).
Unatoč sugestivnom nazivu, ZB uz bolest dišnoga sustava uzrokuje različite kliničke oblike. Infekcija jajovoda u vrlo mladoj dobi može rezultirati njegovim trajnim oštećenjem, dok infekcija odraslih kokoši obično uzrokuje pad ili pre-stanak nesivosti, kao i proizvodnju jaja s tankom i defor-miranom ljuskom, obično s gubitkom pigmentacije. Odre-đeni sojevi virusa ZB mogu značajno oštetiti bubrege i izazvati akutni nefritis, urolitijazu i uginuće, osobito u pilića (Jackwood i de Wit, 2013.). Druga važna činjenica je da virusi ZB postoje u mnogim različitim antigenskim ili genskim tipovima, tj. serotipovima ili genotipovima, koje obično nazivamo varijantama. Virusi ZB imaju golemu sposobnost promjene kako kroz spontane mutacije tako i genskim preslagivanjem (Cavanagh i Gelb, 2008.). Takve promjene mogu rezultirati pojavom novih varijanti od kojih će većina na kraju iščeznuti, neke će postati enzootične ili će imati ograničeno značenje, dok će samo manji broj varijanti postati dominantan, tj. biti raširen u većim regijama, čak i kontinentima. Ove nove varijante obično su proizvoljno nazivane korištenjem punog ili skraćenog naziva geografske lokacije ili godine izolacije, ponekad u kombinaciji s rednim brojevima, i tako dalje. Kao rezultat toga, postoji nekoliko serotipova/genotipova koje su znanstvenici različito nazvali, što je dovelo do velike konfuzije. Na primjer, oznake LX4, D388, A2, korejski-II (K-II), japanski-III (JP-III) i QX-like se odnose na istu varijantu ZB poznatiju kao QX. Drugi dobar primjer je gdje se za istu varijantu koriste oznake 4/91, 793B ili CR88. Nedavni pokušaj usklađivanja klasi-fikacije razvrstava viruse ZB u šest genotipova (ili G-a) označenih rimskim brojevima (GI do GVI), koji ukupno uključuju 32 različite linije označene arapskim brojevima (Valastro i sur., 2016.). Prema ovoj klasifikaciji, varijanta QX treba biti nazivana GI-19, dok se oznaka GI-13 odnosi na varijantu 4/91. Unatoč sustavnoj klasifikaciji i nume-ričkim oznakama još uvijek se preferiraju stariji proizvoljni nazivi za varijante virusa ZB (de Witt i sur., 2018.). Stroga biosigurnost i sustav sa samo jednom dobi peradi na farmi predstavljaju osnovne mjere kontrole, ali cijepljenje je obično neizbježno kako bi se postigla otpornost pilića prema virusu ZB. Dostupna su živa atenuirana i inaktivirana cje-piva, ali budući da postoje brojne antigene varijante virusa ZB, cijepljenje nije ni približno jednostavno kao što je u slučaju NB. Cjepiva određenog serotipa ili genotipa virusa ZB mogu zaštititi pravilno cijepljenu perad od homolognog tipa ZB. Međutim, često postoji samo djelomična zaštita od sojeva drugih serotipova ili genotipova pa zaštita može varirati od visoke do niske (de Wit et al., 2011.). Danas su na raspolaganju cjepiva različitih serotipova/ge-notipova, ali ne i svih trenutno cirkulirajućih serotipo-va/genotipova. To zapravo i nije potrebno, jer određeni cjepni sojevi pružaju visoku križnu zaštitu od određenih heterolognih serotipova ili genotipova (Bijlenga i sur., 2004.). Stoga uz klasifikaciju u serotipove i genotipove, sojeve virusa ZB koji pružaju visoku križnu zaštitu jedni prema drugima svrstavamo u isti protektotip (de Wit i sur., 2011.). U nekoliko je studija pokazano da cijepljenje dvama antigenski različitim živim atenuiranim cjepivima može rezultirati širokom križnom zaštitom protiv mnogih različitih

VIRUSNE INFEKCIJE PERADI – GLOBALNI PROBLEM 14
varijanti virusa ZB. To znači da su alati za uspješno cijep-ljenje protiv većine varijanti virusa ZB dostupni, kao što je planiranje cijepljenja uporabom protektotipova i kombina-cija antigenski različitih cjepiva. Ipak, ključan je način na koji se koriste ti alati, tj. kako osmisliti učinkovit program cijepljenja protiv ZB za određene epizootiološke prilike, u suprotnom i obilni programi cijepljenja mogu zakazati. Poznavanje točnih epizootioloških prilika zahtijeva sustavni i trajni nadzor jata i točnu dijagnozu kako bi se razlikovalo cjepne od cirkulirajućih terenskih sojeva virusa ZB (Savić, 2017.).
Zarazna bolest burze (ZBB, gumborska bolest)
Infekcija virusom ZBB uzrokuje kliničku bolest samo u pilića mlađih od 10 tjedana, dok starije jedinke obično ne pokazuju kliničke znakove. Mogu se zaraziti purani, patke, biserke, fazani i nojevi, ali klinička bolest nije prisutna u ovih vrsta (Eterradossi i Saif, 2013.). Iako su problemi uzrokovani ZBB ograničeni na jednu vrstu peradi i njihovu kratku dob prijemčivosti za virus ZBB, ova bolest je i dalje glavni uzrok gospodarskih gubitaka u peradarskoj industriji. Povijesno gledano, virus ZBB je uzrokovao uglavnom sub-kliničku bolest u tovnih pilića s neznatnim uginućem, ali s imunosupresijom koja je rezultirala uglavnom neizravnim gubicima zbog sekundarnih infekcija, zaostajanja u rastu i povećanog klaoničkog otpada. Situacija se drastično promijenila sredinom 1980-ih kada su u Zapadnoj Europi otkriveni vrlo virulentni sojevi koji su uzrokovali smrtnost od 5% do 15% s povremenim ekstremima od 25% u tovnih pilića i do 60% smrtnosti u pilenki (van den Berg i sur., 1991.). Ovi vrlo virulentni sojevi (vvZBB) su se brzo proširili diljem svijeta uzrokujući dramatične gubitke (van den Berg, 2000.). Infekcija virusima vvZBB mogla se lako prepoznati na terenu zbog akutnog oblika bolesti, tipičnih promjena i osobito povećane smrtnosti, ostavljajući dojam da su virusi vvZBB u potpunosti zamijenili klasične sojeve, čime je klasična ZBB postala zanemarenom. Ipak, nedavne studije ukazuju na prisutnost blažih terenskih sojeva virusa ZBB u Europi, koji se lako mogu previdjeti zbog fokusiranja na klinički bolesne piliće (de Wit i sur., 2018.). Stoga je, uz strogu biosigurnost i provođenje higijenskih mjera, cijep-ljenje ključna metoda ne samo za sprječavanje kliničke bolesti i uginuća, nego i za umanjivanje gospodarskih šteta koje mogu nastati zbog neopažene subkliničke infekcije virusom ZBB (McIlroy i sur., 1992.). Slično kao i kod NB, virusi ZBB pripadaju jednom serotipu, ali cijepljenje protiv ZBB nije tako jednostavno. Glavni razlog je vrlo jaka inter-ferencija majčinskih protutijela kako s cjepnim tako i s terenskim sojevima, što znači da cijepljenje živim cjepivima u prisutnosti majčinskih protutijela može biti dijelom ili potpuno neučinkovito. Pilići pasivno stječu protutijela od svojih roditelja te ona uskoro nakon izlijeganja počinju propadati i traju svega nekoliko tjedana, ovisno ponajprije o njihovoj početnoj razini i tipu pilića. Budući da su pilići najpodložniji virusu ZBB u prvim tjednima života, aktivni imunitet treba biti induciran što prije nakon izlijeganja (Müller i sur., 2012.). Živa cjepiva protiv ZBB pokazuju
različit stupanj atenuacije. Općenito, više atenuirana cjepiva mogu biti neutralizirana nižim razinama protutijela, dok se više virulentni sojevi mogu probiti kroz više razine protu-tijela. Stoga su pilići ranije podložni infekciji terenskim vi-rusom nego cijepljenju atenuiranim cjepivima. Cilj cijep-ljenja živim cjepivima protiv ZBB treba biti njihova što ranija primjena, ali dovoljno kasno da se izbjegne neutra-lizacija majčinskim protutijelima. Stoga, u jatima sa zna-čajno neujednačenim razinama majčinskih protutijela mogu biti potrebna dva cijepljenja, prvo za piliće s početno nižim razinama i drugo za piliće s početno višim razinama maj-činskih protutijela. Unatoč svemu, postoji vremenski okvir kada pilići postaju podložni terenskoj infekciji, osobito virusima vvZBB, ali još nisu podložni cijepljenju živim cjepivima. Ovo osjetljivo razdoblje je čak i dulje, jer je potrebno nekoliko dana za razvoj aktivnog imuniteta na cijepljenje. Moguće je donekle skratiti ovo razdoblje primjenom manje atenuiranih cjepiva, poput „intermedijarnih“ i „interme-dijarnih plus (vrućih)“ cjepiva koja se mogu probiti kroz više razine majčinskih protutijela te su prikladna za pri-mjenu u ranijoj dobi nego „blaga“ cjepiva. "Intermedijarna" i "intermedijarna plus" cjepiva potiču i jači imunosni odziv nakon cijepljenja, ali povećana učinkovitost živih cjepiva protiv ZBB je prvenstveno povezana s umanjenom ne-škodljivošću. To znači da poglavito "intermedijerna plus" cjepiva mogu izazvati imunosupresiju usporedivu s onom koju izazivaju klasični terenski sojevi virusa ZBB. U novije vrijeme su na raspolaganju vektorska cjepiva protiv ZBB s odličnom kombinacijom učinkovitosti i neškodljivosti. Po-godna su za cijepljenje in ovo 18. dana inkubacije ili jednodnevnih pilića u valionici bez obzira na prisutnost majčinskih protutijela. Imunokompleksna cjepiva se mogu primijeniti na isti način, iako su prilično različita od vek-torskih cjepiva u svojim temeljnim biološkim svojstvima. Takva napredna vektorska i imunokompleksna cjepiva su općenito skuplja i moraju se primijeniti injekcijom pojedi-načno svakom piletu za razliku od konvencionalnih živih cjepiva protiv ZBB koja su prikladna za masovno cijepljenje na farmi.
Razmatranje i zaključci
Uz navedene virusne infekcije peradi, nekoliko drugih kon-stantno predstavlja problem za peradarsku industriju, ovisno o vrsti i tipu peradi, dobi i donekle geografskom području. To su, na primjer, zarazni laringotraheitis, puranski rinotra-heitis, virusni hepatitis pataka, boginje peradi, Marekova bolest i druge. Iako je ovdje razmatrano samo nekoliko najvažnijih virusnih infekcija peradi, sve one dijele neke zajedničke karakteristike. Nijedna od ovih bolesti dosad nije bila zadovoljavajuće kontrolirana na globalnoj razini, na-protiv, sve se više pojavljuju jače patogeni ili antigenski varijantni virusni sojevi. Nova cjepiva djelotvorna protiv novih sojeva se kontinuirano razvijaju, dok se upotreba dostupnih cjepiva primjenjuje u industriji peradi već deset-ljećima. Nedvojbeno je da pravilna uporaba cjepiva za perad kroz odgovarajuće programe cijepljenja ima značajan utjecaj

VIRUSNE INFEKCIJE PERADI – GLOBALNI PROBLEM 15
na kontrolu i prevenciju virusnih infekcija, a time i na sma-njenje gospodarskih gubitaka. Međutim, loše i nepravilno cijepljenje može pogodovati širenju virulentnih uzročnika, dok cjepiva koja ne sprječavaju prijenos terenskih sojeva mogu stvoriti uvjete koji potiču nastanak jače patogenih sojeva koji uzrokuju težu bolest u necijepljenih domaćina (Read i sur., 2015.). Čak i neka živa atenuirana cjepiva mo-gu povratiti virulenciju nakon prijenosa s jedinke na jedinku i zatim mogu uzrokovati tešku bolest u necijepljenih jedinki (Coppo i sur., 2013.). Dakle, loše planirana i nepravilno provedena cijepljenja predstavljaju dvosjekli mač s neže-ljenim posljedicama koje se neće nužno pojaviti u kratkom roku, ali dugoročno gledano definitivno hoće. Drugi problem je činjenica da cijepljenje ne može biti zamjena za biosigurnost i održavanje higijene, iako postoje takve zablu-de. Velika konkurentnost i zahtjevi za nižim troškovima proizvodnje peradi promijenili su sustave upravljanja, što je rezultiralo smanjenim naglaskom na biosigurnost i druge osnovne prakse kontrole bolesti. Mnoge nove i moderne farme izgrađene su kao kompleksi višestrukih jata bez mogućnosti jednokratnog useljenja i pražnjenja i držanja samo jedne dobne skupine peradi na farmi. Umjesto toga, kontinuirana proizvodnja u takvim farmama s više dobnih skupina omogućuje kontinuiranu cirkulaciju virusa sa sta-rijih na mlađa jata. Obilna cirkulacija terenskog virusa s jedinke na jedinku u takvim kompleksima pogoduje na-stanku novih antigenskih varijanti ili jače patogenih sojeva. Također je vrlo teško, ako ne i nemoguće, osmisliti učinkovit i siguran program cijepljenja na takvim farmama, osobito ako je infekcija već prisutna. Razrjeđivanje jata tovnih pilića, tj. odvoz na klanje dio po dio jata u cilju povećanja profita druga je praksa koja uvelike doprinosi širenju virusnih infekcija među jatima i farmama. Može se zaključiti da su sustavi upravljanja peradarskom proizvod-njom koji zanemaruju temeljna načela kontrole i iskorje-njivanja bolesti u velikoj mjeri odgovorni za pojavu i širenje virusnih infekcija u današnje vrijeme. Čak i u slučajevima masovnih epizootija VPIP u peradi u SAD-u i Europi koje su se dogodile nakon unosa virusa divljim pticama selicama treba imati na umu da je taj virus nastao u peradi, iako se izvorno pojavio na drugoj strani svijeta. Također, prisutnost međunarodnih tvrtki i zajedničkih tržišta omogućava lakše i brže širenje virusnih infekcija između udaljenih lokacija. Sve to čini virusne infekcije peradi globalnim problemom koji ugrožava perad na lokalnoj razini i zahtijeva uključi-vanje veterinara specijaliziranih za peradarstvo kako u raz-matranje bolesti tako i u upravljačka tijela još u fazi plani-ranja novih peradarskih farmi ili rekonstrukcije postojećih peradarskih pogona.
Napomena
Ovaj rad je izvorno napisan i prezentiran kao plenarno predavanje na 15. Europskom peradarskom kongresu (The XVth European Poultry Conference) održanom od 17. do 21. rujna 2018. godine u Dubrovniku u organizaciji Hrvatskoga ogranka Svjetske udruge peradara u znanosti i struci (World Poultry Science Association – Croatian Branch). Rad je u
izvornom obliku na engleskom jeziku dostupan u Zborniku Kongresa na mrežnoj stranici: https://pure.au.dk/portal/files/140225746/sbornik_docladov_epc_2018.pdf
Literatura
ALDOUS, E. W., D. J. ALEXANDER (2001): Detection and dif-ferentiation of Newcastle diseasevirus (avian paramyxovirus type 1). Avian Pathology, 30: 117-128.
ALEXANDER, D. J. (2001): Newcastle disease - The Gordon Memorial Lecture. British Poultry Science, 42: 5-22.
ALEXANDER, D. J., I. CAPUA (2008): Avian influenza in poultry. World's Poultry Science Journal, 64: 513-532.
ANONYMOUS (2011): World Livestock Disease Atlas: A Quanti-tative Analysis of Global Animal Health DataA (2006–2009). World Bank, Washington, DC and TAFS Forum, Bern.
BIJLENGA, G., J. K. COOK, J. GELB, J. J. DE WIT (2004): Development and use of the H strain of avian infectious bronchitis virus from the Netherlands as a vaccine: a review. Avian Pathology, 33: 550-557.
BROWN, I., P. MULATTI, K. SMIETANKA, C. STAUBACH, P. WILLEBERG, C. ADLHOCH, D. CANDIANI, C. FABRIS, C. ZANCANARO, J. MORGADO, F. VERDONCK (2017): Avian influenza overview October 2016-August 2017. EFSA Journal, 15: 5018.
CAPUA, I., D. J. ALEXANDER (2009): Avian influenza infection in birds: A challenge and opportunity for the poultry vete-rinarian. Poultry Science, 88: 842-846.
CAPUA, I., S. MARANGON (2007): The challenge of controlling notifiable avian influenza by means of vaccination. Avian Diseases, 51: s1 317-322.
CAPUA, I., M. SCACCHIA, T. TOSCANI, V. CAPORALE (1993): Unexpected isolation of virulent Newcastle disease virus from commercial embryonated fowls’ eggs. Journal of Veterinary Medicine B, 40: 609-612.
CAPUA, I., C. TERREGINO, G. CATTOLI, A. TOFFAN (2004): Increased resistance of vaccinated turkeys to experimental infection with an H7N3 low-pathogenicity avian influenza virus. Avian Pathology 33: 158-163.
CAVANAGH, D., J. GELB (2008): Infectious bronchitis. In: SAIF, Y. M., FADLY, A. M., GLISSON, J. R., MCDOUGALD, L. R., NOLAN, L. K. and SWAYNE, D. E. (Eds) Diseases of poultry, 12th ed. pp. 117-135 (Blackwell Publishing, Ames, Iowa).
COPPO, M. J., A. H. NOORMOHAMMADI, G. F. BROWNING, J. M. DEVLIN (2013): Challenges and recent advancements in infectious laryngotracheitis virus vaccines. Avian Patho-logy, 42:195-205.
DE WIT, J. J., C. CAZABAN, R. DIJKMAN, G. RAMON, Y. GARDI (2018): Detection of different genotypes of infectious bronchitis virus and of infectious bursal disease virus in European broilers during an epidemiological study in 2013 and the consequences for the diagnostic approach. Avian Pathology, 47: 140-151.
DE WIT, J. J., J. K. COOK, H. M. VAN DER HEIJDEN (2011): Infectious bronchitis virus variants: a review of the history, current situation and control measures. Avian Pathology, 40: 223-235.
DIMITROV, K. M., C. L. AFONSO, Q. YU, P. J. MILLER (2017): Newcastle disease vaccines-A solved problem

VIRUSNE INFEKCIJE PERADI – GLOBALNI PROBLEM 16
or a continuous challenge? Veterinary Microbiology, 206: 126-136.
ETERRADOSSI, N., Y. M. SAIF (2013): Infectious bursal disease. In: SWAYNE, D. E., GLISSON, J. R., MCDOUGALD, L. R., NOLAN, L. K., SUAREZ, D. L. and NAIR, V. (Eds) Diseases of poultry, 13th ed. pp. 209-246 (Blackwell Publishing, Ames, Iowa).
GUITTET, M., H. LE COQ, M. MORIN, V. JESTIN, G. BEN-NEJEAN (1993): Distribution of Newcastle disease virus after challenge in tissues of vaccinated broilers. Proceedings of the 10th World Veterinary Poultry Association Congress, Sydney, 179.
HESTERBERG, U., K. HARRIS, D. STROUD, V. GUBERTI, L. BUSANI, M. PITTMAN, V. PIAZZA, A. COOK, I. BROWN (2009): Avian influenza surveillance in wild birds in the European Union in 2006. Influenza and Other Respiratory Viruses, 3: 1-14.
JACKWOOD, M. W., J. J. DE WIT (2013): Infectious Bronchitis. In: SWAYNE, D. E., GLISSON, J. R., MCDOUGALD, L. R., NOLAN, L. K., SUAREZ, D. L., NAIR, V. (Eds) Diseases of poultry, 13th ed. pp. 139-159 (Blackwell Publishing, Ames, Iowa).
LEE, D. H., M. K. TORCHETTI, K. WINKER, H. S. IP, C. S. SONG, D. E. SWAYNE (2015): Intercontinental spread of Asian-origin H5N8 to North America through Beringia by migratory birds. Journal of Virology, 89: 6521-6524.
MCILROY, S. G., E. A. GOODALL, D. W. BRUCE, R. M. MCCRACKEN, M. S. MCNULTY (1992): The cost benefit of vaccinating broiler flocks against subclinical infectious bursal disease. Avian Pathology, 21: 65-76.
MÜLLER, H., E. MUNDT, N. ETERRADOSSI, M. R. ISLAM (2012): Current status of vaccines against infectious bursal disease. Avian Pathology, 41: 133-139.
NAPP, S., N. MAJÓ, R. SÁNCHEZ-GÓNZALEZ, J. VERGARA-ALERT (2018): Emergence and spread of highly pathogenic avian influenza A(H5N8) in Europe in 2016-2017. Transboundary and Emerging Diseases, 1-10.
NORMILE, D. (2006): Evidence points to migratory birds in H5N1 spread. Science, 311: 1225.
PAREDE, L., P. L. YOUNG (1990): The pathogenesis of velogenic Newcastle disease virus infection of chickens of
different ages and different levels of immunity. Avian Diseases, 34: 803-808
READ, A. F., S. J. BAIGENT, C. POWERS, L. B. KGOSANA, L. BLACKWELL, L. P. SMITH, D. A. KENNEDY, S. W. WALKDEN-BROWN, V. NAIR, C. FRASER (2015): Imperfect vaccination can enhance the transmission of highly virulent pathogens. PLoS Biology, 13: e1002198.
SALZBERG, S. L., C. KINGSFORD, G. CATTOLI, D. J. SPIRO, D. A. JANIES, M. M. ALY, I. H. BROWN, E. COUACY-HYMANN, G. M. DE MIA, D. H. DUNG, A. GUERCIO, T. JOANNIS, A. S. M. ALI, A. OSMANI, I. PADALINO, M. D. SAAD, V. SAVIĆ, N. A. SENGAMALAY, S. YINGST, J ZABORSKY, O. ZORMAN-ROJS, E. GHEDIN, I. CAPUA (2007): Genome Analysis Linking Recent European and African Influenza (H5N1) Viruses. Emerging Infectious Diseases, 13: 713-718.
SAVIĆ, V. (2017) Monitoring of IBV circulation and prevalence. Zootechnica International, 10: 48-51.
SAVIĆ, V., R. RAGUŽĆ-ĐURIĆ, G. KRIVEC, B. ŠIMPRAGA, M. TIŠLJAR, M. MIKEC, M. SOKOLOVIĆ, M. BALE-NOVIĆ, T. AMŠEL ZELENIKA (2006): Peradarstvo u Hrvatskoj i svijetu u 2005. i 2006. Stočarstvo, 61: 213-229.
VALASTRO, V., E. C. HOLMES, P. BRITTON, A. FUSARO, M. W. JACKWOOD, G. CATTOLI, I. MONNE (2016): S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infection, Genetics and Evolution, 39: 349-364.
VAN DEN BERG, T. P. (2000): Acute infectious bursal disease in poultry: a review. Avian Pathology, 29: 175-194.
VAN DEN BERG, T. P., M. GONZE, G. MEULEMANS (1991): Acute infectious bursal disease in poultry: isolation and characterisation of a highly virulent strain. Avian Pathology, 20: 133-143.
WANG, Y., Z. JIANG, Z. JIN, H. TAN, B. XU (2013): Risk factors for infectious diseases in backyard poultry farms in the Poyang Lake area, China. PLoS One, 8: e67366.
WEBSTER, R. G., M. PEIRIS, H. CHEN, Y. GUAN (2006): H5N1 outbreaks and enzootic influenza. Emerging Infectious Diseases, 12: 3-8.
VIRAL INFECTIONS OF POULTRY – THE GLOBALLY CHALLENGING SITUATION
Instead of summary
This work was originally written and presented as a plenary lecture at the XVth European Poultry Conference held in Dubrovnik from 17 to 21 September 2018, organized by the World Poultry Science Association – Croatian Branch. The Conference Proceedings book is available online, for example at: https://pure.au.dk/portal/files/140225746/ /sbornik_docladov_epc_2018.pdf.

PERADARSKI DANI 2019. 17
IZAZOVI I OČEKIVANJA EU I PERADARSTVA HRVATSKE U SREDNJEROČNOM RAZDOBLJU DO 2030. GODINE
Branko Bobetić
Gospodarsko interesno udruženje „Croatiastočar“, Zagreb, Hrvatska Proizvodno tržišni i potrošni trendovi u sektorima mesa peradi i konzumnih jaja
Meso peradi
U razdoblju od 2012. do 2018. godine ukupna europska proizvodnja svih vrsta mesa peradi dosegnula je količinu od 14,889 milijuna tona u odnosu na 2012. godinu od 12,717 milijuna tona, što je povećanje za 17%. Proizvodnja je povećana kod starih članica za 9% na 10,800 milijuna tona u odnosu na 9.855 tona u 2012. godini, dok su novopridošle članice EU 13 u prošloj godini ostvarile proizvodnju od 4,089 milijuna tona u odnosu na 2,862 tone, što je povećanje proizvodnje za 43%. Hrvatska proizvodnja mesa peradi u prošloj godini, prema bilancama jednodnevne peradi iz valionica, dosegnula je količinu od cca 85.000 tona, što je u odnosu na proizvodnju od 79.000 tona u 2012. godini povećanje za 9% (tablice 1.-3.). Europska komisija predviđa povećanje proizvodnje mesa peradi kod EU članica do 2030. godine za 4% na ukupnu količinu od cca 15,475 milijuna tona. Predviđeno povećanje u srednjoročnom razdoblju kod 15 zemalja EU je za 1%, dok je projekcija povećanja mesa peradi kod 13 zemalja EU za 10% na količinu od 5,509 milijuna tona. U istom razdoblju Europska komisija predviđa pad proizvodnje svinjskog mesa za 2%, goveđeg mesa za 6% te povećanje proizvodnje ovčjeg mesa za 7%. Trendovi proizvodnje mesa peradi u Hrvatskoj trebali bi biti definirani strategijom hrvatske poljoprivrede do 2030. godine, izrada koje je u tijeku pri Ministarstvu poljoprivrede i Svjetske banke. U prošloj godini od ukupne EU proizvodnje mesa peradi udio pilećeg mesa iznosio je 81%, purećeg mesa 14% te ostalih vrsta mesa peradi 5%. U Hrvatskoj je udio pilećeg mesa bio cca 78%, udio purećeg mesa 15%, dok je 7% bio udio kokošjeg mesa, mesa pataka i gusaka. U odnosu na EU proizvodnju mesa peradi udio uvoza EU članica bio je 5%,
dok je udio izvoza bio 11%. EU uvoz mesa peradi iz trećih zemalja najvećim dijelom se odnosi na meso pilećih prsiju te panirane proizvode od prsiju peradi, dok EU izvoz sadrži najvećim dijelom kategorije mesa peradi niže cjenovne vrijednosti. Udio uvoza mesa peradi u Hrvatskoj u prošloj godini u odnosu na domaću proizvodnju iznosio je 26%, dok je udio izvoza iznosio cca 10%. Samodostatnost u proizvodnji i potrošnji mesa peradi EU članica u prošloj godini iznosila je 105%, a u Hrvatskoj 89%. Maloprodajna potrošnja mesa peradi EU članica iznosila je cca 24 kg p/c, a u Hrvatskoj je cca 21 kg p/c u prošloj godini.
Konzumna jaja
Ukupna EU proizvodnja konzumnih jaja u prošloj godini ostvarena je u količini od 6,932 milijuna tona, što je u odnosu na proizvodnju 2012. godine u količini od 6,240 tona povećanje proizvodnje za cca 11%. U Hrvatskoj je procijenjena proizvodnja konzumnih jaja u prošloj godini bila cca 40.000 tona, što je povećanje za oko 14% u odnosu na 2012. godinu kada je proizvodnja iznosila cca 35.000 tona (tablice 4.-5.). Udio EU uvoza konzumnih jaja u odnosu na proizvodnju iznosio je 0,3%, dok je udio EU izvoza iznosio 3%. U Hrvatskoj je udio uvoza konzumnih jaja u odnosu na ostvarenu proizvodnju u prošloj godini iznosio 12%, dok je udio izvoza bio 4%. Prosječna potrošnja konzumnih jaja EU članica bila je cca 13,1 kg p/c, dok je u Hrvatskoj iznosila cca 11 kg p/c. Europska komisija predviđa povećanje proizvodnje konzumnih jaja do 2030. godine za cca 11% na ukupnu količinu od 7,700 milijuna tona. Članice EU, prema zadnjim pokazateljima, imaju instaliranih kapaciteta za cca 397 milijuna konzumnih nesilica od čega na kavezni sustav otpada cca 53%, podni sustav držanja nesilica 26%, sustav free range 15% i sustav organske proizvodnje 5%. Hrvatska ima instaliranih kapaciteta za cca 1,858 milijuna nesilica od čega 73% otpada na kavezni uzgoj, 25% na podni uzgoj te svega 2% na free range i organski uzgoj.
Branko Bobetić, dr. med. vet., Gospodarsko interesno udruženje "Croatiastočar", Ivana Šibla 9, 10000 Zagreb, Hrvatska; tel: +385(0)1 6604990; faks: +385(0)1 6604998; E-mail: [email protected]
IZAZOVI I OČEKIVANJA EU I PERADARSTVA HRVATSKE U SREDNJEROČNOM RAZDOBLJU DO 2030. GODINE 18
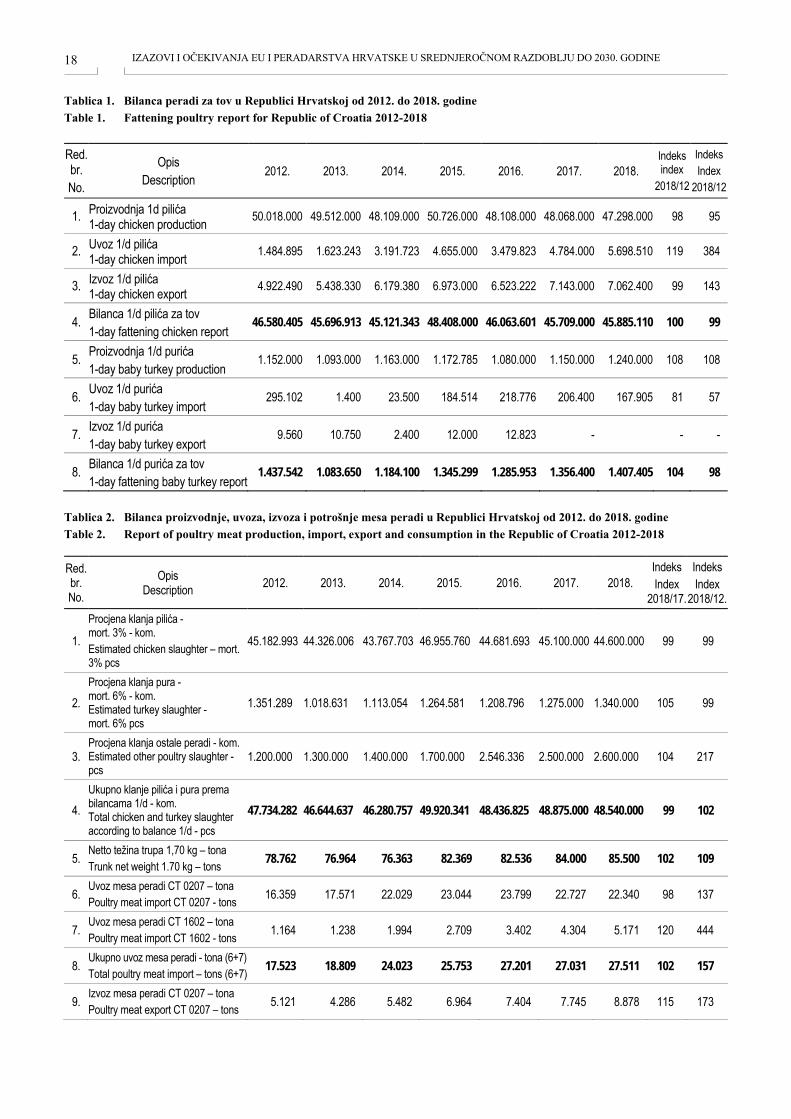
Tablica 1. Bilanca peradi za tov u Republici Hrvatskoj od 2012. do 2018. godine Table 1. Fattening poultry report for Republic of Croatia 2012-2018
Red. br. No.
Opis Description 2012. 2013. 2014. 2015. 2016. 2017. 2018.
Indeks index
2018/12
Indeks Index
2018/12
1. Proizvodnja 1d pilića 1-day chicken production 50.018.000 49.512.000 48.109.000 50.726.000 48.108.000 48.068.000 47.298.000 98 95
2. Uvoz 1/d pilića 1-day chicken import 1.484.895 1.623.243 3.191.723 4.655.000 3.479.823 4.784.000 5.698.510 119 384
3. Izvoz 1/d pilića 1-day chicken export 4.922.490 5.438.330 6.179.380 6.973.000 6.523.222 7.143.000 7.062.400 99 143
4. Bilanca 1/d pilića za tov 1-day fattening chicken report
46.580.405 45.696.913 45.121.343 48.408.000 46.063.601 45.709.000 45.885.110 100 99
5. Proizvodnja 1/d purića 1-day baby turkey production
1.152.000 1.093.000 1.163.000 1.172.785 1.080.000 1.150.000 1.240.000 108 108
6. Uvoz 1/d purića 1-day baby turkey import
295.102 1.400 23.500 184.514 218.776 206.400 167.905 81 57
7. Izvoz 1/d purića 1-day baby turkey export
9.560 10.750 2.400 12.000 12.823 -
- -
8. Bilanca 1/d purića za tov 1-day fattening baby turkey report
1.437.542 1.083.650 1.184.100 1.345.299 1.285.953 1.356.400 1.407.405 104 98
Tablica 2. Bilanca proizvodnje, uvoza, izvoza i potrošnje mesa peradi u Republici Hrvatskoj od 2012. do 2018. godine Table 2. Report of poultry meat production, import, export and consumption in the Republic of Croatia 2012-2018
Red. br. No.
Opis Description 2012. 2013. 2014. 2015. 2016. 2017. 2018.
Indeks Index
2018/17.
Indeks Index
2018/12.
1.
Procjena klanja pilića - mort. 3% - kom. Estimated chicken slaughter – mort. 3% pcs
45.182.993 44.326.006 43.767.703 46.955.760 44.681.693 45.100.000 44.600.000 99 99
2. Procjena klanja pura - mort. 6% - kom. Estimated turkey slaughter - mort. 6% pcs
1.351.289 1.018.631 1.113.054 1.264.581 1.208.796 1.275.000 1.340.000 105 99
3. Procjena klanja ostale peradi - kom. Estimated other poultry slaughter - pcs
1.200.000 1.300.000 1.400.000 1.700.000 2.546.336 2.500.000 2.600.000 104 217
4. Ukupno klanje pilića i pura prema bilancama 1/d - kom. Total chicken and turkey slaughter according to balance 1/d - pcs
47.734.282 46.644.637 46.280.757 49.920.341 48.436.825 48.875.000 48.540.000 99 102
5. Netto težina trupa 1,70 kg – tona Trunk net weight 1.70 kg – tons 78.762 76.964 76.363 82.369 82.536 84.000 85.500 102 109
6. Uvoz mesa peradi CT 0207 – tona Poultry meat import CT 0207 - tons 16.359 17.571 22.029 23.044 23.799 22.727 22.340 98 137
7. Uvoz mesa peradi CT 1602 – tona Poultry meat import CT 1602 - tons 1.164 1.238 1.994 2.709 3.402 4.304 5.171 120 444
8. Ukupno uvoz mesa peradi - tona (6+7) Total poultry meat import – tons (6+7) 17.523 18.809 24.023 25.753 27.201 27.031 27.511 102 157
9. Izvoz mesa peradi CT 0207 – tona Poultry meat export CT 0207 – tons 5.121 4.286 5.482 6.964 7.404 7.745 8.878 115 173

IZAZOVI I OČEKIVANJA EU I PERADARSTVA HRVATSKE U SREDNJEROČNOM RAZDOBLJU DO 2030. GODINE 19
Red. br. No.
Opis Description 2012. 2013. 2014. 2015. 2016. 2017. 2018.
Indeks Index
2018/17.
Indeks Index
2018/12.
10. Izvoz mesa peradi CT 1602 – tona Poultry meat export CT 1602 – tons 5.268 4.900 5.772 5.827 6.467 6.863 6.662 97 126
11. Ukupno izvoz mesa peradi – tona (9+10) Total poultry meat export – tons (9+10)
10.389 9.186 11.254 12.791 13.871 14.608 15.540 106 150
12. Ukupna potrošnja mesa peradi (5+8-11) – tona Total poultry meat consumption (5+8-11) – tons
85.896 86.587 89.132 95.331 99.828 96.423 97.471 101 113
13. Populacija stanovništva (x000) Country population (x000) 4.287 4.272 4.256 4.240 4.210 4.189 4.150 99 97
14. Potrošnja - kg p/c CWE Consumption – kg p/c CWE 20,04 20,27 20,94 22,48 23,71 23,00 23,50 102 117
15. Uvoz žive peradi tona CWE - R=71%/ Live poultry import tons CWE – R=71%
234 275 873 1.119 1.922 2.189 2.821 129 1.206
16. Izvoz žive peradi tona CWE – R=71% Live poultry export tons CWE – R=71%
1.222 913 2.723 2.276 3.067 4.078 5.304 130 434
17. Bruto domaća proizvodnja - tona 5+16-15/12/ Gross domestic production - tons 5+16-15/12
79.750 77.602 78.213 83.196 83.682 85.889 87.983 102 110
18. Samodostatnost 17/12 Self-sufficiency 17/12 92% 89% 86% 86% 84% 89% 89% 100 97
19. Ukupno klanje peradi u klaonicama DZS Total poultry slaughter in DZS slaughterhouses
35.608.514 34.866.946 35.732.332 37.969.971 38.206.661 38.500.000 38.800.000 101 109
20. Razlika (19-4) - Difference (19-4) -12.125.768 -11.777.691 -10.548.425 -11.950.370 -10.230.164 -10.375.000 -9.710.000 94 80
Izvor podataka: DZS Source of data: Bureau of Statistics Tablica 3. Inkubacija rasplodnih jaja i procjena proizvodnje jednodnevnih pilića u Republici Hrvatskoj od 2012. do 2018.
godine Table 3. Incubation of breeder eggs and estimate of 1-day chicken production in the Republic of Croatia 2012-2018
Red. br. No.
Opis Description 2012. 2013. 2014. 2015. 2016. 2017. 2018.
Indeks Index
2018/12.
Indeks Index
2018/17.
1. Obuhvat broja valionica Number of hatcheries 28 29 24 23 25 24 23 82 96
2. Ukupno uloženih rasplodnih jaja LL + mješovito Total breeder eggs input LL + mixed
3.322.000 2.872.000 2.129.000 2.907.000 2.615.000 3.491.000 2.985.000 90 86
3.
Procjena proizvodnje 1/d nesilica Estimated 1/d laying hen production
1.329.000 1.148.000 851.000 1.163.000 1.046.000 1.396.000 1.194.000 90 86

IZAZOVI I OČEKIVANJA EU I PERADARSTVA HRVATSKE U SREDNJEROČNOM RAZDOBLJU DO 2030. GODINE 20
Tablica 4. Procjena uzgoja pilenki lakih linija i ukupnog broja kokošiju u Republici Hrvatskoj od 2012. do 2018. godine Table 4. Estimated light-line hen breeding and total number of hens in the Republic of Croatia 2012-2018
Red. br. No.
Opis/Description 2012. 2013. 2014. 2015. 2016. 2017. 2018. Indeks/ Index
2018/12.
Indeks/ Index
2018/17.
1.
Procjena proizvodnje 1/d nesilica u RH/ Estimated 1/d laying hen production in RH
1.329.000 1.148.000 851.000 1.163.000 1.046.000 1.396.000 1.194.000 113 86
2.
Uvoz jednodnevnih nesilica CT 010511/91 Import of 1-day laying hens CT 010511/91
608.304 417.314 381.679 679.479 617.211 1.366.000 1.050.000 173 77
3.
Ukupna bilanca jednodnevnih nesilica (1+2) Total balance of 1-day laying hens (1+2)
1.937.304 1.565.314 1.232.679 1.842.479 1.663.211 2.651.000 2.244.000 135 85
4.
Procjena uzgoja pilenki - mortalitet 3%/ Estimated hen breeding - mortality 3%
1.879.185 1.518.355 1.195.699 1.787.205 1.379.000 2.570.000 2.177.000 135 85
5. Uvoz pilenki CT 01059400 Import of hens CT 01059400
116.605 173.223 226.304 244.823 406.722 164.175 198.000 170 121
6. Procjena ukupnih pilenki LL (4+5) Total hen estimate LL (4+5)
1.995.790 1.691.578 1.422.003 2.032.028 1.785.722 2.952.000 2.375.000 137 80
7. Ukupan broj kokošiju – DZS Total number of hens – DZS
3.700.000 3.800.000 3.722.000 3.017.000 3.858.000 4.100.000 3.900.000 137 95
8. Razlika 7-6 - Difference 7-6 1.704.210 2.108.422 2.299.997 984.972 2.072.278 1.148.000 1.525.000 137 133
9.
Kapaciteti nesilica kod registr. proizvođača Laying hen capacity at registered producers
2.000.000 2.052.849 1.698.911 1.786.661 1.850.000 1.800.000 1.860.000 137 103
Tablica 5. Procjena proizvodnje konzumnih jaja te bilanca uvoza, izvoza i potrošnje u Republici Hrvatskoj od 2012. do 2018.
godine Table 5. Estimated egg production and import, export and consumption balance for Republic of Croatia 2012-2018
Red. br. No.
Opis/Description 2012. 2013. 2014. 2015. 2016. 2017. 2018. Indeks/ Index
2018/12.
Indeks/ Index
2018/17.
1. Proizvodnja konzumnih jaja LL - robni proizvođači (3-2) Production of eggs LL - producers (3-2)
419.957.000 450.553.000 437.003.000 444.305.000 544.000.000 575.000.000 570.000.000 136 99
2. Samoopskrbna proizvodnja Self-supply production
165.000.000 155.000.000 135.000.000 120.000.000 110.000.000 100.000.000 80.000.000 48 80
3. Procjena nacionalne proizvodnja konz. jaja Estimate of national egg production
584.957.000 605.553.000 572.003.000 564.305.000 654.000.000 675.000.000 650.000.000 111 96

IZAZOVI I OČEKIVANJA EU I PERADARSTVA HRVATSKE U SREDNJEROČNOM RAZDOBLJU DO 2030. GODINE 21
Red. br. No.
Opis/Description 2012. 2013. 2014. 2015. 2016. 2017. 2018. Indeks/ Index
2018/12.
Indeks/ Index
2018/17.
4. Uvoz konzumnih jaja (kom) CT 04072100 Egg import (pcs) CT 04072100
16.672.000 11.840.000 49.296.000 99.136.000 115.000.000 90.000.000 88.000.000 528 98
5. Izvoz konzumnih jaja (kom) CT 04072100 Egg export (pcs) CT 04072100
9.056.000 8.368.000 8.656.000 10.544.000 23.000.000 15.000.000 25.000.000 276 167
6. Potrošnja konzumnih jaja Egg consumption 592.573.000 609.025.000 612.643.000 652.897.000 746.000.000 750.000.000 713.000.000 120 95
7. Populacija broja stanovnika u 000 Population in 000
4.287 4.272 4.256 4.240 4.210 4.189 4.150 97 99
8. Potrošnja kom. jaja - p/c Egg consumption, pcs - p/c 138 143 144 154 177 179 172 124 96
9. Razlika 6-3 - Difference 6-3 7.616.000 3.472.000 40.640.000 88.592.000 92.000.000 75.000.000 63.000.000 827 84
10. Samodostatnost proizvodnja/potrošnja Production/consumption self-sufficiency
99% 99% 93% 86% 88% 89% 91% 92 102
Izvor podataka: DZS Napomena: Za procjenu proizvodnje jednodnevnih nesilica primijenjena je valivost od 80%, a kod uzgoja pilenki mortalitet od 3%. Procjena proizvodnje izračunata je na bazi bilanci pilenki uz proizvodnju od oko 275 jaja godišnje po ulaznoj nesilici lake linije. Source of data: Bureau of Statistics Note: Assessment of 1-day laying hen production is based on 80% hatchery rate, and hen production on 3% mortality rate. Assessment of production is based on hen balance with production of about 275 eggs per year per light-line laying hen.
Tržišna konkurentnost u sektorima mesa i konzumnih jaja
Svake druge godine EU krovna asocijacija AVEC naručuje studiju konkurentnosti najvećih EU proizvođača mesa peradi na globalnom tržištu. Prema prošlogodišnjoj studiji nizozemskog poljoprivrednog instituta iz Wageningena prosječna EU cijena koštanja kg žive vage najvećih EU
proizvođača pilića u 2017. godini (slika 1.) iznosila je cca 0,82 eura/kg, što je za 29% veća cijena u odnosu na prosječnu cijenu koštanja najvećih svjetskih izvoznika na globalnom tržištu (USA, Tajland, Brazil, Argentina i Ukrajina) koja je iznosila 0,69 eura/kg. Prosječna EU cijena pilećih trupova grill obrade sa svim troškovima klanja iznosila je 1,46 eura, što je za 24% veća cijena u odnosu na globalne svjetske izvoznike koja je iznosila 1,18 eura.

IZAZOVI I OČEKIVANJA EU I PERADARSTVA HRVATSKE U SREDNJEROČNOM RAZDOBLJU DO 2030. GODINE 22
Godina/Year 2012. 2013. 2014. 2015. 2016. 2017. 2018. Indeks/Index 109 101 97 98 101 101 101
Opis - Description EU RH Indeks RH/EU - RH/EU index Prosječna cijena 2012.-2018. - Average price 2012-2018 187,31 189,81 101
Slika 1. Tržišne cijene pilića grill obrade u RH i EU od 2012. do 2018. godine (€/100 kg) Figure 1. Grill chicken meat market price in RH and EU, 2012-2018 (€/100 kg) Slična je situacija u trošku proizvodnje konzumnih jaja (slika 2.) gdje je EU cijena iznosila cca 0,89 eura/kg, što je za 23% veća cijena koštanja u odnosu na prosječnu cijenu najvećih izvoznika proizvoda od jaja na globalnom tržištu (USA, Ukrajina, Argentina, Indija), a iznosila je 0,73 eura/kg. Na globalnom tržištu uglavnom se trguje pro-izvodima od jaja pa je prema zadnjoj studiji AVEC-a EU trošak proizvodnje jaja u prahu iznosio 5,41 eura/kg, što je za 21% veći trošak u odnosu na 4,46 eura/kg koliki je trošak proizvodnje najvećih izvoznika proizvoda od jaja na globalom tržištu. Na EU troškove proizvodnje u odnosu na globalne izvoznike najvećim dijelom utječe trošak EU legislative u proizvodnji mesa peradi s udjelom od 6% u ukupnom trošku s naslova zdravlja i sigurnosti hrane, zaštite okoliša i dobrobiti peradi, a dijelom i veći EU trošak rada. Udio EU legislative u ukupnom trošku proizvodnje kon-zumnih jaja iznosi visokih 16%. U proteklih sedam godina prosječna veleprodajna cijena pilića grill obrade EU članica iznosila je 1,87 eura/kg, dok je u Hrvatskoj bila veća za cca 1% ili 1,90 eura. Nakon 2012. godine EU tržne cijene grill pilića padale su do 2015. godine za 7%, dok je pad tržnih cijena u Hrvatskoj bio prisutan uzastopno četiri godine do 2016. Godine, što je rezultiralo padom tržnih cijena u odnosu na 2012. za 16%. Prosječna veleprodajna EU cijena konzumnih jaja bila je cca 1,36 eura/kg, dok je u Hrvatskoj bila cca 1,64 eura/kg.
Nadolazeći izazovi peradarske industrije
U studenom 2018. godine u američkom gradu Atlanti prvi puta je održan skup znanstvenika, istraživača, inovatora i predstavnika najvećih proizvođača mesa peradi iz 20 ze-malja na temu očekivanih izazova na globalnom tržištu mesom peradi te primjeni inovativne tehnologije. Na skupu su analitičari respektabilne nizozemske RABO banke iznijeli opću procjenu trendova rasta potrošnja animalnih bjelan-čevina u idućih 20 godina za više od 25%. Najveći prosječni godišnji rast proizvodnje očekuje se u peradarskoj industriji i to kod mesa peradi od 2%, jaja za 1,6%, goveđeg mesa za 1,1% i svinjskog mesa za 1%. Analitičari također predviđaju pojačanu volatilnost globalnog tržišta koja će biti uzrokova-na brzorastućom potrošnjom zbog rasta populacije stanov-ništva i povećanja kupovne moći kod sadašnjih siromašnih regija u svijetu. Na volatilnost tržišta i dalje će utjecati poremećaji uzrokovani bolestima peradi kao što je to bio slučaj nakon izbijanja influence peradi. Pretpostavlja se također da će na volatilnost djelomice utjecati i sami po-trošači, naročito potrošači nove generacije koji u sve većoj mjeri preferiraju sigurnost i zdravlje hrane s obzirom na sve veće značenje sigurnosti i zdravlja potrošača, dobrobit u uzgoju, zaštitu okoliša, transparentnost u cjelokupnom lancu od proizvodnje do potrošnje te sve veći udio inovativnih proizvoda ne samo kroz asortiman nego i proizvoda bez prisustva antibiotika, non GMO, bez alergena te s manje soli i E brojeva.

IZAZOVI I OČEKIVANJA EU I PERADARSTVA HRVATSKE U SREDNJEROČNOM RAZDOBLJU DO 2030. GODINE 23
Godina/Year 2012. 2013. 2014. 2015. 2016. 2017. 2018. Indeks/Index 108 134 126 128 143 114 115
Opis - Description EU RH Indeks RH/EU - RH/EU index Prosječna cijena 2012.-2018. - Average price 2012-2018 133,59 164,02 123
Slika 2. Pregled neto prodajnih cijena konzumnih jaja RH i EU od 2012. do 2018. godine (€/100 kg) Figure 2. Net shelf price for eggs in RH and EU 2012-2018 (€/100 kg) Za europsko peradarstvo svakako je izazov vezan za BREXIT s obzirom na to da je Ujedinjeno Kraljevstvo iza Njemačke drugi najveći kupac mesa peradi europskih proizvođača u godišnjoj količini većoj od 500.000 tona s intra tržišta Na hrvatsku industriju peradskog mesa i konzumnih jaja značajno će utjecati i nova politika CAP-a do 2027. godine s obzirom na to da su posjednici peradi u Hrvatskoj vrlo malo koristili predpristupne fondove i proračunska sredstva sa-dašnjeg programskog razdoblja za ulaganje u modernizaciju kako kapaciteta primarne proizvodnje u peradarstvu tako i u prerađivačke kapacitete. Peradarsku industriju na globalnoj razini u idućim godinama u segmentu sigurnosti hrane očekuje niz već i sada prisutnih izazova. U procesnoj industriji očekuje se aplikacija nove generacije bio-informatičkih alata za otkrivanje i inden-tifikaciju serotipova Salmonella spp. kroz kontinuirano uzimanje briseva na procesnoj opremi i trupovima. Razvija se nova generacija prototipova antimikrobnih probiotika radi eradikacije uzročnika salmoneloza, kampilobakterioza i klostridioza. Isto tako u prerađivačkim pogonima predviđa
se zamjena klasičnih materijala, poglavito lijevanih kućišta, novim kompozitnim materijalima koji su otporni na habanja i omogućuju efikasnije održavanje i sanitaciju. U primarnoj proizvodnji očekuje se sve veća primjena robota i auto-matizacije u objektima koji bi slikovnim i ostalim vrstama senzora uočili sva odstupanja, što bi rezultiralo pravo-dobnom intervencijom radi otklanjanja uzroka proizvodnih odstupanja. Osobit izazov za cjelokupni lanac od uzgoja do tržišta je primjena digitalnih metoda koje bi trebale rezultirati optimalnim proizvodno tržišnim bilancama trupova peradi. U posljednjih nekoliko godina najaktualnija tehnologija u svijetu je primjena tzv. umjetne inteligencije. U peradarstvu je to izazov integriranja brojnih i složenih podataka u svim proizvodnim fazama, što bi trebalo re-zultirati najvišim proizvodno sigurnosnim standardima, proizvodima namirnica visoke nutritivne vrijednosti, do-stupnosti u najširem smislu uz najviši stupanj dobrobiti peradi i zaštite okoliša. Izvor podataka: FAOStat, Eurostat, DZS i Europska komisija, AVEC

PERADARSKI DANI 2019. 24
DJELATNOSTI LABORATORIJA CENTRA ZA PERADARSTVO TIJEKOM 2017. I 2018. GODINE
Tajana Amšel Zelenika, Vladimir Savić, Borka Šimpraga, Marina Tišljar, Mirta Balenović, Marijana Sokolović, Tihomir Zglavnik, Fani Krstulović, Luka Jurinović, Marija Berendika
Hrvatski veterinarski institut, Centar za peradarstvo, Zagreb, Hrvatska
Sažetak
Osnovna zadaća Centra za peradarstvo je laboratorijska i terenska dijagnostika zaraznih, nametničkih i drugih bolesti peradi, praćenje njihove pojave, širenja i suzbijanja, ispitivanje zdravstvene ispravnosti hrane za životinje te sirovina i aditiva koje služe za njezinu proizvodnju. U radu je prikazana djelatnost laboratorija Centra za peradarstvo te učestalost pojave pojedinih virusnih, bakterijskih i parazitskih bolesti peradi te bolesti koje su nastale posljedično pogreškama u tehnologiji i hranidbi tijekom 2017. i 2018. godine.
Ključne riječi: dijagnostika, epizootiologija, peradarstvo, Hrvatska Uvod Od prvih zapisa o razvoju peradarstva u hrvatskoj literaturi iz 15. stoljeća do danas pristup proizvodnji se mijenjao od ekstenzivno držane peradi preko golemih ulaganja u tehno-logiju proizvodnje i selekciju pasmina peradi, postizanja intenzivne proizvodnje koja u vrlo kratkom vremenu omo-gućava kvalitetan proizvod – meso i jaja pa do današnjih alternativnih načina uzgoja i sve češćeg i cijenjenijeg povratka ka slobodnom ekološkom uzgoju peradi. Centar za peradarstvo osnovan je 1961. godine u okviru Veterinarskog fakulteta kao specijalizirana institucija koja će zajedničkim koordiniranim radom osiguravati i razvijati peradarsku proizvodnju u svim njenim tehnološkim segmen-tima, a 1993. godine postaje sastavni dio Hrvatskoga veteri-narskog instituta, kao znanstveno-dijagnostička institucija koja u svakom trenutku može odgovoriti potrebama pera-darske proizvodnje. Laboratoriji Centra za peradarstvo su akreditirani prema normi HRN EN ISO/IEC 17025. Za razliku od ostalih di-jagnostičkih i analitičkih laboratorija, Centar za peradarstvo ima i savjetodavnu ulogu u hrvatskom peradarstvu te na taj način održava bliske kontakte s peradarskim proizvođačima. Osnovna i najznačajnija zadaća Centra za peradarstvo je laboratorijska i terenska dijagnostika zaraznih, nametničkih i drugih bolesti peradi, praćenje njihove pojave, širenja i suzbijanja, ispitivanje zdravstvene ispravnosti hrane za
životinje te sirovina i aditiva koje služe za njezinu proizvodnju. Danas Centar za peradarstvo broji 20 djelatnika od kojih je 7 doktora znanosti, 1 magistar znanosti i 1 magistar specijalist te dva djelatnika visoke stručne spreme. Velika pozornost polaže se na znanstvena, razvojna i primijenjena istraživanja u području veterinarstva i pera-darstva te se stečena znanja i dostignuća prenose u praksu. Upravo zbog toga neprestano se radi na uvođenju novih laboratorijskih metoda za dijagnostiku, analitiku, preventivu i terapiju peradi. Iznimno dobri rezultati međulaboratorijskih ispitivanja u kojima sudjeluju laboratoriji Centra za peradarstvo iz godine u godinu ukazuju na visoku kvalitetu, razvijenost i oprem-ljenost laboratorija. Cilj ovoga rada je, kroz prikaz djelatnosti laboratorija Centra za peradarstvo, ukazati na učestalost pojave pojedinih virusnih, bakterijskih i parazitskih bolesti peradi te bolesti nastalih zbog pogrešaka u tehnologiji i hranidbi tijekom 2017. i 2018. godine.
Laboratorij za patologiju
Osnovna djelatnost Laboratorija za patologiju je pato-loškomorfološka dijagnostika bolesti domaće peradi i divljih ptica. Prepoznavanje i praćenje pojavnosti i rasprostranje-nosti bolesti temelji se na razudbenom nalazu, pato-loškohistološkoj i citološkoj pretrazi, a prema potrebi
dr. sc. Tajana Amšel Zelenika, dr. med. vet., Hrvatski veterinarski institut, Centar za peradarstvo, Heinzelova 55, 10000 Zagreb, Hrvatska; e-mail: [email protected]

DJELATNOSTI LABORATORIJA CENTRA ZA PERADARSTVO TIJEKOM 2017. I 2018. GODINE 25
nadopunjuje se pretragom elektronskim mikroskopom, pre-traživanjem molekularnim metodama i imunohistokemij-skom pretragom. Od 2018. godine Laboratorij za patologiju uključen je u Nacionalnu grupu za sprječavanje i prevenciju trovanja divljih životinja. Time je Centar za peradarstvo aktivno pristupio pokretanju Nacionalnog akcijskog plana ujedno sudjelujući i doprinoseći ostvarivanju pristupa
„jednog zdravlja“. Tijekom 2017. i 2018. godine Laboratorij za patologiju zaprimio je 711 lešina domaće peradi (tovnih pilića, konzumnih i rasplodnih nesilica), tovnih purana (intenzivna proizvodnja i obiteljska gospodarstva) i zagor-skih purana te ukrasnih i autohtonih pasmina peradi i ptica. U tablicama 1.-4. prikazane su bolesti pojedinih vrsta i kategorija peradi tijekom 2017. i 2018. godine.
Tablica 1. Bolesti tovnih pilića, rasplodnih i konzumnih nesilica uzrokovane pretežito mikroorganizmima u 2017. i 2018. godini Table 1. Diseases in broiler chickens, breeding and laying hens caused mostly by microorganisms in 2017 and 2018
Bolest / Disease Tovni pilići
Broiler Rasplodne nesilice
Breeding hens Konzumne nesilice
Laying hens 2017. 2018. 2017. 2018. 2017. 2018.
Omfalitis, infekcija žumanjčane vrećice; upala skočnih zglobova Omphalitis, yolk sac infection; hock joint inflammation 16 110
Hepatitis s uklopinama (tipa adenovirusnih uklopina) Hepatitis with inclusions (adenovirus inclusions) 4
Gnojnofibrinozna upala gornjih dišnih prohoda; poliserozitis; sepsa Purulent fibrinous inflammation of upper airways; polyserositis; sepsis 9 16
Bolesti organa za reprodukciju / Reproductive organ diseases 19 61 6 Kronična dišna bolest / Chronic respiratory disease 3 15 Sindrom zarazne kržljavosti /Infectious stunting syndrome 4 3 13 Sindrom zarazne kržljavosti, nekroza glave bedrene kosti; erozija kutikule mišićnog želuca; kokcidioza; sepsa / Infectious stunting syndrome, femur head necrosis; gizzard erosion; coccidiosis; sepsis
54 62
Sindrom otečene glave / Swollen head syndrome 8 Marekova bolest / Marek's disease 2 1 23 19 Boginje / Fowl pox 1 1 Sepsa / Sepsis 4 1 Nekrotični enteritis / Necrotic enteritis 5 Aspergiloza / Aspergillosis 1 1 Ukupno / Total 83 65 33 119 101 61 Tablica 2. Bolesti tovnih pilića i konzumnih nesilica uzrokovane pretežito pogreškama u tehnologiji hranidbe i uzgoja u 2017. i
2018. godini Table 2. Diseases in broiler chickens and laying hens caused mostly by errors in nutrition and breeding technology in 2017 and
2018
Bolesti / Disease Tovni pilići / Broiler Konzumne nesilice / Laying hens
2017. 2018. 2017. 2018. Kanibalizam; kljucanje / Cannibalism; Pecking 5 2 Periferna neuropatija / Peripheral neuropathy 12 Ascites – sindrom plućne hipertenzije Ascites – pulmonary hypertension syndrome (PHS) 7 2 1
Neuropatija / Neuropathy 8 Pretilost / Overweight 2 Suspektno trovanje / Suspected poisoning 1 Ukupno / Total 7 2 29 2

DJELATNOSTI LABORATORIJA CENTRA ZA PERADARSTVO TIJEKOM 2017. I 2018. GODINE 26
Tablica 3. Bolesti tovnih pilića, rasplodnih i konzumnih nesilica, uzrokovane pretežno parazitskom invazijom u 2017. i 2018. godini
Table 3. Diseases in broiler chickens, breeding and laying hens caused mostly by parasitic invasion in 2017 and 2018
Bolest / Disease Tovni pilići / Broiler Rasplodne nesilice
Breeding hens Konzumne nesilice
Laying hens 2017. 2018. 2017. 2018. 2017. 2018.
Glistavost / Ascaridiasis 1 4 Tekutijaza / Red mite infestation 2 4 1 Knemidokoptoza / Knemidocoptosis 1 1 Ukupno / Total 2 1 9 2 Tablica 4. Bolesti purana (intenzivna proizvodnja; obiteljska gospodarstva i zagorski puran) uzrokovane pretežito mikro-
organizmima (bakterije, gljivice, virusi i paraziti), pogreškama u tehnologiji uzgoja i hranidbi u 2017. i 2018. godini Table 4. Diseases in turkeys (intensive production; family farms and Zagorje turkey) caused mostly by microorganisms
(bacteria, fungi, viruses and parasites) in 2017 and 2018
Bolest / Disease Tovni puran
Broiler turkey Zagorski puran Zagorje turkey
2017. 2018. 2017. 2018. Limfomatozna hiperplazija u bubrezima / Lymphomatous hyperplasia of the kidneys 1 Nekroza glave bedrene kosti; sindrom malapsorpcije; nekrotični miokarditis; sepsa Femur head necrosis; malabsorption syndrome; necrotic myocarditis; sepsis 27
Nekrotični enteritis / Necrotic enteritis 12 Sepsa / Sepsis 2 Bordeteloza purana / Turkey bordetellosis 5 Marekova bolest / Marek's disease 2 2 Histomonijaza / Histomoniasis 4 Singamoza / Syngamosis 1 Leukom rožnice; kaheksija / Corneal leukoma; Cachexia 1 Ukupno / Total 29 7 1 20 U patologiji tovnih pilića protekle dvije godine prevladavao je sindrom zarazne kržljavosti, često praćen nekrozom glave bedrene kosti, erozijama mišićnog želuca uz sepsu. U rasplodnih nesilica najčešće je dijagnosticirana bolest organa za reprodukciju te omfalitis i infekcija žumanjčane vreće. Nešto rjeđe utvrđen je sindrom otečene glave te pojedinačni slučajevi hepatitisa s uklopinama, Marekove bolesti i aspergiloze. Tijekom dvogodišnjeg razdoblja u konzumnih nesilica najčešće su dijagnosticirane bolesti organa za repro-dukciju, bolesti dišnih prohoda (kronična respiratorna bolest i gnojnofibrinozna upala gornjih dišnih prohoda). I dalje su prisutne bolesti uzrokovane pogreškama u tehnologiji hranidbe i uzgoja uz povremenu infestaciju parazitima. Laboratorijska dijagnostika bolesti u purana ukazuje na veći broj jedinki s nalazom nekroze glave bedrene kosti u kombinaciji sa sindromom malapsorpcije, miokarditisom i sepsom. U uzgojima pataka i gusaka dijagnosticirane su bakterijske infekcije, no broj pretraženih uzoraka bio je izrazito mali pa
je teško zaključiti koje bolesti prevladavaju u ovim uz-gojima.
Laboratorij za virusologiju i serologiju
Dijagnostika svih značajnih virusnih bolesti peradi izravno temeljem dokaza uzročnika ili neizravno dokazom specifičnih protutijela u necijepljenih ptica kao i procjena postvakcinalne imunosti u farmske i slobodno držane peradi osnova su rada Laboratorija za virusologiju i serologiju. Uz virusne bolesti laboratorij radi i serološku i molekularnu dijagnostiku nekih bakterijskih bolesti peradi. Kao nacionalni referentni laboratorij, uz dijagnostiku virusnih i bakterijskih bolesti peradi na zahtjev kupca, laboratorij obavlja dijagnostiku influence ptica i newcast-leske bolesti. Prema „Programu pretraživanja virusa influ-ence ptica u divljih ptica“ i „Programu pretraživanja virusa influence u peradi“ laboratorij organizira i provodi pri-kupljanje i testiranje uzoraka divljih ptica za potrebe nacionalnog monitoringa zbog utvrđivanja rizika od uno-

DJELATNOSTI LABORATORIJA CENTRA ZA PERADARSTVO TIJEKOM 2017. I 2018. GODINE 27
šenja virusa influence ptica u uzgoje domaće peradi, kao i serološki nadzor nisko patogenih podtipova virusa influence ptica H5 i H7 u različitih vrsta peradi i visokopatogenog podtipa H5N8 u pataka i gusaka. Laboratorij provodi i "Program nadziranja groznice za-padnog Nila na području Republike Hrvatske" serološkim pretraživanjem ekstenzivno držanih kokoši i dokazom uzročnika molekularnim pretraživanjem uzoraka tkiva uginulih ili odstrijeljenih ptica. U okviru "Plana za odobravanje objekata za uzgoj i valjenje peradi" Ministarstva poljoprivrede, Centar provodi i se-rološka testiranja na serovarove Salmonella gallinarum, Salmonella pullorum, Mycoplasma gallisepticum i Myco-plasma meleagridis. Temeljem TCP/RER/3004 programa Svjetske organizacije za hranu i poljoprivredu (FAO) određen je kao referentni laboratorij za influencu ptica za zemlje Istočne Europe i Kavkaza te u okviru ovoga programa proizvodi i referentne reagense za influencu ptica i provodi edukaciju za labo-ratorijsku dijagnostiku ove bolesti. Razvoj novih metoda dijagnostike u Laboratoriju za virusologiju i serologiju tijekom 2017. i 2018. godine Zbog iznimno učestalih i brzih mutacija koje se događaju u genomu virusa influence ptica uspješno su učinjene brojne modifikacije za molekularne metode u cilju detaljne identifikacije visoko patogenih virusa podtipova H5N5 i H5N8 koji su cirkulirali tijekom epizootije 2017. godine u Hrvatskoj.
Tijekom 2017. i 2018. godine uvedena je automatizirana ekstrakcija RNA i DNA, RT-PCR za detekciju virusa avijarnog encefalomijelitisa i klasični PCR za genotip VII virusa newcastleske bolesti. Laboratorij za virusologiju i serologiju Centra za pera-darstvo zaprimio je tijekom 2017. godine 1.307, a tijekom 2018. godine 1.217 uzoraka na dokaz odnosno kvanti-fikaciju virusa i mikoplazmi u peradi i divljih ptica. Najveći broj uzoraka odnosi se na program monitoringa influence A u peradi i divljih ptica. Tablica 5. prikazuje broj pretražene domaće peradi na prisutnost virusa influence A. Pretrage za dokaz virusa i mikoplazmi u domaće peradi prikazane su u tablici 6. Tablica 5. Broj pretražene domaće peradi na prisutnost
virusa influence A u 2017. i 2018. godini Table 5. Number of domestic poultry searched for
influenza A virus in 2017 and 2018
Influenca A Influenza A
Broj pretražene peradi Number of poultry searched 2017. 2018.
Kokoši / Chickens 208 575
Patke / Ducks 54 32
Pure / Turkeys 6 3
Guske / Geese 10 29
Tablica 6. Pretrage za dokaz/kvantifikaciju virusa i mikoplazmi domaće peradi u 2017. i 2018. godini (pozitivno/pretraženo) Table 6. Tests performed for virus and mycoplasma verification/quantification in domestic poultry in 2017 and 2018
(positive/examined)
Uzročnik / Cause Konzumne nesilice
Laying hens Rasplodne nesilice
Breeding hens Tovni pilići Broilers Purani Turkeys
Patke Ducks
Guske Geese
2017. 2018. 2017. 2018. 2017. 2018. 2017. 2018. 2017. 2018. 2017. 2018. Newcastleska bolest Newcastle disease 0/13 0/1 0/2 0/3
Zarazni bronhitis Infectious bronchitis 2(1*)/7 0/3 1/1 13(11*)/2
3 6(2*)/15
Zarazna bolest burze Infectious bursal disease 0/2 0/1 1*/5
Ptičji reovirusi Avian reoviruses 3/5 0/2 0/7
Leukoza peradi Avian leukosis 0/1
Ptičji adenovirusi Avian adenoviruses 14/23 9/34 2/2 2/8
Marekova bolest Marek's disease 0/1
Boginje peradi / Fowl pox 1/1 2/3 Zarazni laringotraheitis Infectious laryngotracheitis 2*/3

DJELATNOSTI LABORATORIJA CENTRA ZA PERADARSTVO TIJEKOM 2017. I 2018. GODINE 28
Uzročnik / Cause Konzumne nesilice
Laying hens Rasplodne nesilice
Breeding hens Tovni pilići Broilers Purani Turkeys
Patke Ducks
Guske Geese
2017. 2018. 2017. 2018. 2017. 2018. 2017. 2018. 2017. 2018. 2017. 2018. Mycoplasma gallisepticum 5/7 4/11 0/1 0/4 0/2 0/4 Mycoplasma synoviae 6/13 0/11 0/1 0/3 Mycoplasma meleagridis 0/2 0/3
Parvovirus 0/1 Ptičji astrovirusi / Avian astroviruses 0/2
Ptičji pneumovirus Avian pneumovirus 0/1 2/2 (tip B) 0/1
Dersijeva bolest Derzsy's disease 0/1
Zarazna anemija kokoši Chicken infectious anemia 0/5 0/14
*Broj uzoraka u kojem je dokazan cijepni soj virusa / Number of samples with vaccine virus strain verified Tablica 7. Broj pretraženih uzoraka seruma na prisutnost protutijela na određene virusne bolesti, mikoplazme i salmonelu
skupine D u 2017. i 2018. godini Table 7. Number of samples examined for the presence of antibodies to particular viral diseases, mycoplasma and
Protutijela (Metoda) / Antibodies (Method) 2017. 2018. Influenca ptica, podtip H5 (IHA) / Influenza, subtype H5 (IH test) 3.748 4.434 Influenca ptica, podtip H7 (IHA) / Influenza, subtype H7 (IH test) 3.748 4.434 Influenca ptica, H5N8 (IHA) / Influenza, H5N8 (IH test) 391 592 Newcastleska bolest (IHA) / Newcastle disease (IH test) 3.461 1.084 Encefalitis zapadnog Nila (ELISA) / West Nile encephalitis (ELISA) 1.732 478 Hemoragični enteritis purana (IDG) 1.296 1.132 Zarazna bolest burze (ELISA) / Infectious bursal disease (ELISA) 556 408 Sindrom pada nesivosti (IHA) / Egg drop syndrome (IHA)
1.363 810
Zarazni bronhitis (ELISA) / Infectious bronchitis (ELISA) 606 390 Ptičji pneumovirus (ELISA) / Avian pneumovirus (ELISA) 540 260 Leukoza (ELISA) / Leukosis (ELISA) 315 255 Salmonella D skupine (BSA) / Group D Salmonella (BSA) 4.205 4.373 Mycoplasma gallisepticum (BSA) 6.531 6.512 Mycoplasma synoviae (BSA) 1.589 1.197 Mycoplasma meleagridis (BSA) 1.884 1.584 Ukupno / Total 31.965 27.943 Serološkim metodama pretraženo je u 2017. godini 31.965 seruma, a u 2018. godini 27.943 seruma na prisutnost protu-tijela na određene virusne bolesti, mikoplazme i salmonelu skupine D. Najveći broj pretraženih seruma napravljen je na osnovi programa koji propisuje Ministarstvo poljoprivrede i odnosi se na detekciju protutijela za influencu A, newcast-
lesku bolest, encefalitisa Zapadnog Nila, salmonelu skupine D i mikoplazme (tablica 7.). Dokaz visokopatogene influence ptica uzrokovane podtipo-vima H5N5 i H5N8 u divljih ptica i domaće peradi zasi-gurno je obilježilo 2017. godinu. Dokazana je i prisutnost ptičjeg adenovirusa u domaće peradi, reovirusa, pneumo-

DJELATNOSTI LABORATORIJA CENTRA ZA PERADARSTVO TIJEKOM 2017. I 2018. GODINE 29
virusa, boginja peradi, infekcija peradi virusom Zapadnog Nila te infekcija bakterijom Mycoplasma gallisepticum. U 2018. godini dokazana je infekcija ptičjim adenovirusima, virusom Zapadnog Nila i bakterijama Mycoplasma gallisep-ticum i Mycoplasma synoviae u farmski držane peradi te infekcija virusom zaraznog bronhitisa u seoske peradi. Tijekom 2018. godine serološkim pretraživanjem uzoraka na prisutnost protutijela na influencu ptica u seoske peradi utvrđena je prisutnost protutijela na H7 podtip influence, što je rezultiralo uzimanjem većeg broja brisova na sumnjivom području (područje Veterinarske stanice Našice), no virus nije detektiran. Laboratorij za bakteriologiju U Laboratoriju za bakteriologiju provodi se dijagnostika značajnih bolesti peradi bakterijske i gljivične etiologije, identifikacija i biokemijska karakterizacija izdvojenih bakte-rija i gljivica pomoću identifikacijskog sustava ATB, mole-kularne metode za dokazivanje bakterija roda Campylo-bacter, bakterije roda Salmonella (posebno serovarova Gallinarum/Pullorum) te za vrste Enterococcus faecalis i Enterococcus faecium. Laboratorij za bakteriologiju imeno-van je nacionalnim referentnim laboratorijem za salmonele i kampilobaktere u peradi od strane Ministarstva poljopri-vrede Republike Hrvatske.
Razvoj novih metoda dijagnostike u Laboratoriju za bakteriologiju tijekom 2017. i 2018. godine U Laboratoriju za bakteriologiju u potpunosti je uređen prostor za molekularnu dijagnostiku gdje je uz dokazivanje ranije navedenih bakterija uvedena i genotipizacija izolata kampilobaktera metodom MLST (Multi Locus Sequence Typing). Ovu metodu moguće je primijeniti za mnoge bakterijske vrste, a kao izrazito reproducibilna metoda predstavlja odličan alat u epidemiologiji. Tablice 8. i 9. prikazuju ukupan broj pretraženih uzoraka u Laboratoriju za bakteriologiju tijekom 2017. i 2018. godine. Od ukupno 15.506 pretraženih uzoraka izdvojen je 181 izolat bakterija roda Salmonella (iz uzoraka tovnih pilića 122, konzumnih nesilica 41, rasplodnih nesilica 5, rasplod-nih i tovnih purana po jedan te iz uzoraka fazana 6 izolata). Najviše izdvojenih salmonela pripadalo je serovaru Infantis i to iz uzoraka prikupljenih od tovnih pilića. Na drugom mjestu bila je S. enteritidis koja je najzastupljenija u uzorcima podrijetlom od konzumnih nesilica, zatim slijede serovarovi Mbandaka, Typhimurium, Coeln te manje za-stupljeni Corvalis, Senftenberg, Teddingtone, Derby, Bovis-morbificans, Livingstone, Singapore i Szentes. Serovarovi Blockley i Typhimurium dokazani su u jatu rasplodnih purana dok su u fazana dokazani Blockley, Infantis i Typhimurium.
Tablica 8. Ukupan broj pretraženih uzoraka u laboratoriju za bakteriologiju u 2017. godini Table 8. Total number of samples tested at Laboratory of Bacteriology in 2017
Vrsta uzorka / Sample type Broj
uzoraka Number of samples
Pretrage / Tests Bakteriološke / Bacteriologic Mikološke
Mycologic SAL CL PAS STA EC PSA ENT CAMP
Izmet / navlake ze obuću / Faeces/overboots 14.025 14.025 Perad / uzrok uginuća / Poultry/cause of death 168 168 84 25 168 160 168 48 28 Perad/pretraga organa na salmonele Poultry/organ testing for salmonella 250 250
R. jaja/ljuske jaja / B. eggs/egg shell 305 305 1 1 1 1 1 K. jaja / C. eggs 38 38 Brisovi/nastambe za perad, valionice Swabs from farming facilities 196 24 172 172 172 172 172
Brisovi nečisnica / Cloacal swabs 35 35 Prašina iz nastambi za perad Dust from farming facilities 15 15
Slijepa crijeva / Appendices 367 367 Bakterijske kulture / Bacterial cultures 102 102 Pretraga na rezidue antimikrobnih lijekova Testing for antimicrobial residues 5
Ukupno / Total 15.506 14.967 84 25 341 333 341 221 367 201 R. jaja – rasplodna jaja / breeding eggs, K. jaja – konzumna jaja / eggs for consumption, SAL – Salmonella, CL – Clostridium, PAS – Pasteurella, STA – Stahylococcus, EC – Escherichia coli, PSA – Pseudomonas aeruginosa, ENT – Enterococcus, CAMP – Campylobacter, Brisovi – brisovi uzorkovani sa površina i opreme u nastambama i valionicama za perad / Swabs – swabs sampled from surfaces and equipment in poultry houses and hatcheries

DJELATNOSTI LABORATORIJA CENTRA ZA PERADARSTVO TIJEKOM 2017. I 2018. GODINE 30
Tablica 9. Ukupan broj pretraženih uzoraka u Laboratoriju za bakteriologiju u 2018. godini Table 9. Total number of samples tested at Laboratory of Bacteriology in 2018
Vrsta uzorka / Sample type Broj
uzoraka Number of samples
Pretrage / Tests Bakteriološke / Bacteriologic Mikološke
Mycologic SAL CL PAS STA EC PSA ENT CAMP Izmet / navlake ze obuću / Faeces/overboots 13.160 13.160 Perad / uzrok uginuća / Poultry/cause of death 129 129 106 28 129 129 129 77 6 Perad/pretraga organa na salmonele Poultry/organ testing for salmonella 263 263
R. jaja/ljuske jaja / B. eggs/egg shell 641 641 K. jaja / C. eggs 26 26 Brisovi/nastambe za perad, valionice Swabs from farming facilities 219 40 179 179 179 179 179
Brisovi nečisnica / Cloacal swabs 32 32 Prašina iz nastambi za perad Dust from farming facilities 9 9
Slijepa crijeva / Appendices 333 333 Bakterijske kulture / Bacterial cultures 96 96 Pretraga na rezidue antimikrobnih lijekova Testing for antimicrobial residues 3
Ukupno / Total 14.911 14.396 106 28 308 308 308 256 333 185 R. jaja – rasplodna jaja / breeding eggs, K. jaja – konzumna jaja / eggs for consumption, SAL – Salmonella, CL – Clostridium, PAS – Pasteurella, STA – Stahylococcus, EC – Escherichia coli, PSA – Pseudomonas aeruginosa, ENT – Enterococcus., CAMP – Campylobacter Brisovi – brisovi uzorkovani sa površina i opreme u nastambama i valionicama za perad Swabs – swabs sampled from surfaces and equipment in poultry houses and hatcheries Iz organa pretražene peradi zbog utvrđivanja uzroka uginuća izdvojeno je 33 izolata bakterija roda Clostridium, 115 izo-lata bakterije E. coli, 33 izolata bakterija roda Enterococcus, 11 izolata bakterije Erysipelothrix rhusiopathiae te 10 izolata bakterije Mannheimia haemolytica. Od ukupno pretraženih 367 uzoraka slijepih crijeva tovnih pilića izdvojena su 243 izolata bakterija roda Campylo-bacter. Od toga je 138 izolata pripadalo vrsti C. jejuni, a 105 vrsti C. coli. Od ukupno 14.911 uzoraka pretraženih u 2018. godini iz-dvojeno je 205 bakterijskih sojeva roda Salmonella. Izdvo-jeno je i serološki tipizirano 47 salmonela iz konzumnih nesilica, četiri iz rasplodnih nesilica, 141 iz tovnih pilića te pet salmonela iz tovnih purana, dvije salmonele u rasplodnih purana te po tri soja u pataka i fazana. Najviše izdvojenih salmonela pripadalo je serovaru Infantis (95) i to iz uzoraka prikupljenih od tovnih pilića te Mban-daka u 40 jata. Slijedi S. enteritidis koja je najzastupljenija u uzorcima podrijetlom od konzumnih nesilica. Utvrđena je manja prisutnost serovarova Typhimurium, Mbandaka, Bovismorbificans, Corvalis, Coeln, Senftenberg, Blockley. Serovarovi Schwarzengrund, Bovismorbificans i Kottbus izdvojeni su iz uzoraka podrijetlom od purana. Iz uzoraka podrijetlom od fazana izoliran je po jedan serovar
Typhimurium, Blockley i Agona. U uzorcima iz jata pataka dokazan je serovar Infantis. Iz organa pretražene peradi zbog utvrđivanja uzroka uginuća izdvojeno je 59 izolata bakterija roda Clostridium, 69 izolata bakterije E. coli, 45 izolata bakterija roda Enterococcus, pet izolata bakterije Hafnia alvei te tri izolata bakterije Mannheimia haemolytica. Od ukupno pretražena 333 uzorka slijepih crijeva izdvojeno je 199 sojeva bakterija roda Campylobacter. Od toga je 117 sojeva pripadalo vrsti C. jejuni, a 82 vrsti C. coli. Iz uzoraka slijepih crijeva zbog praćenja proširenosti indi-katorskih bakterija E. coli i Enterococcus faecalis i faecium izdvojeno je 277 izolata bakterije E. coli i 212 sojeva bakterija E. faecalis (54) i faecium (158).
Laboratorij za analitiku stočne hrane Kvaliteta i sigurnost hrane i hrane za životinje i dalje je značajan čimbenik u osiguranju zdravlja ljudi i životinja. Uz neophodne nutritivne karakteristike važne za proizvodnost, zdravlje, ponašanje i dobrobit životinja, sigurnost hrane važan je čimbenik u sprječavanju ulaska nepoželjnih pato-gena i tvari u hranidbeni lanac. Proizvođači hrane i hrane za životinje kao „subjekti u poslovanju s hranom“ imaju stoga važnu ulogu u uzgoju životinja i proizvodnji proizvoda
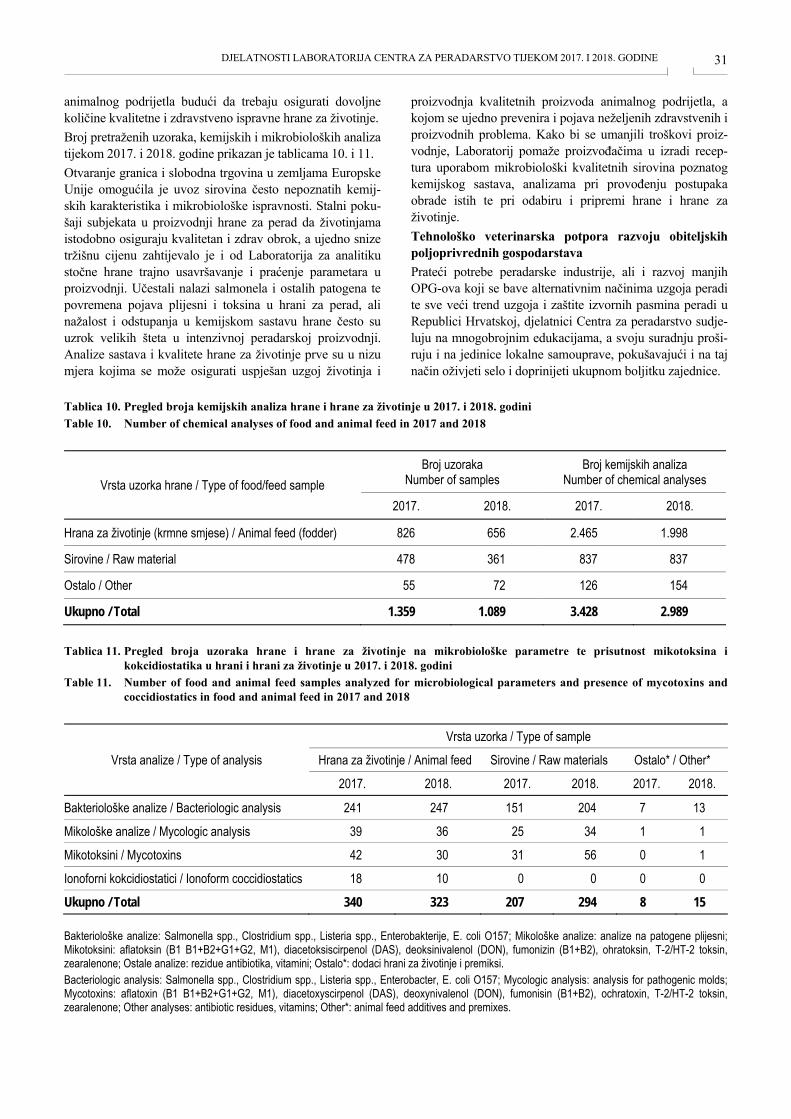
DJELATNOSTI LABORATORIJA CENTRA ZA PERADARSTVO TIJEKOM 2017. I 2018. GODINE 31
animalnog podrijetla budući da trebaju osigurati dovoljne količine kvalitetne i zdravstveno ispravne hrane za životinje. Broj pretraženih uzoraka, kemijskih i mikrobioloških analiza tijekom 2017. i 2018. godine prikazan je tablicama 10. i 11. Otvaranje granica i slobodna trgovina u zemljama Europske Unije omogućila je uvoz sirovina često nepoznatih kemij-skih karakteristika i mikrobiološke ispravnosti. Stalni poku-šaji subjekata u proizvodnji hrane za perad da životinjama istodobno osiguraju kvalitetan i zdrav obrok, a ujedno snize tržišnu cijenu zahtijevalo je i od Laboratorija za analitiku stočne hrane trajno usavršavanje i praćenje parametara u proizvodnji. Učestali nalazi salmonela i ostalih patogena te povremena pojava plijesni i toksina u hrani za perad, ali nažalost i odstupanja u kemijskom sastavu hrane često su uzrok velikih šteta u intenzivnoj peradarskoj proizvodnji. Analize sastava i kvalitete hrane za životinje prve su u nizu mjera kojima se može osigurati uspješan uzgoj životinja i
proizvodnja kvalitetnih proizvoda animalnog podrijetla, a kojom se ujedno prevenira i pojava neželjenih zdravstvenih i proizvodnih problema. Kako bi se umanjili troškovi proiz-vodnje, Laboratorij pomaže proizvođačima u izradi recep-tura uporabom mikrobiološki kvalitetnih sirovina poznatog kemijskog sastava, analizama pri provođenju postupaka obrade istih te pri odabiru i pripremi hrane i hrane za životinje. Tehnološko veterinarska potpora razvoju obiteljskih poljoprivrednih gospodarstava Prateći potrebe peradarske industrije, ali i razvoj manjih OPG-ova koji se bave alternativnim načinima uzgoja peradi te sve veći trend uzgoja i zaštite izvornih pasmina peradi u Republici Hrvatskoj, djelatnici Centra za peradarstvo sudje-luju na mnogobrojnim edukacijama, a svoju suradnju proši-ruju i na jedinice lokalne samouprave, pokušavajući i na taj način oživjeti selo i doprinijeti ukupnom boljitku zajednice.
Tablica 10. Pregled broja kemijskih analiza hrane i hrane za životinje u 2017. i 2018. godini Table 10. Number of chemical analyses of food and animal feed in 2017 and 2018
Vrsta uzorka hrane / Type of food/feed sample Broj uzoraka
Number of samples Broj kemijskih analiza
Number of chemical analyses
2017. 2018. 2017. 2018.
Hrana za životinje (krmne smjese) / Animal feed (fodder) 826 656 2.465 1.998
Sirovine / Raw material 478 361 837 837
Ostalo / Other 55 72 126 154
Ukupno / Total 1.359 1.089 3.428 2.989 Tablica 11. Pregled broja uzoraka hrane i hrane za životinje na mikrobiološke parametre te prisutnost mikotoksina i
kokcidiostatika u hrani i hrani za životinje u 2017. i 2018. godini Table 11. Number of food and animal feed samples analyzed for microbiological parameters and presence of mycotoxins and
coccidiostatics in food and animal feed in 2017 and 2018
Vrsta analize / Type of analysis
Vrsta uzorka / Type of sample
Hrana za životinje / Animal feed Sirovine / Raw materials Ostalo* / Other*
2017. 2018. 2017. 2018. 2017. 2018.
Bakteriološke analize / Bacteriologic analysis 241 247 151 204 7 13
Mikološke analize / Mycologic analysis 39 36 25 34 1 1 Mikotoksini / Mycotoxins 42 30 31 56 0 1 Ionoforni kokcidiostatici / Ionoform coccidiostatics 18 10 0 0 0 0
Ukupno / Total 340 323 207 294 8 15
Bakteriološke analize: Salmonella spp., Clostridium spp., Listeria spp., Enterobakterije, E. coli O157; Mikološke analize: analize na patogene plijesni; Mikotoksini: aflatoksin (B1 B1+B2+G1+G2, M1), diacetoksiscirpenol (DAS), deoksinivalenol (DON), fumonizin (B1+B2), ohratoksin, T-2/HT-2 toksin, zearalenone; Ostale analize: rezidue antibiotika, vitamini; Ostalo*: dodaci hrani za životinje i premiksi. Bacteriologic analysis: Salmonella spp., Clostridium spp., Listeria spp., Enterobacter, E. coli O157; Mycologic analysis: analysis for pathogenic molds; Mycotoxins: aflatoxin (B1 B1+B2+G1+G2, M1), diacetoxyscirpenol (DAS), deoxynivalenol (DON), fumonisin (B1+B2), ochratoxin, T-2/HT-2 toksin, zearalenone; Other analyses: antibiotic residues, vitamins; Other*: animal feed additives and premixes.

DJELATNOSTI LABORATORIJA CENTRA ZA PERADARSTVO TIJEKOM 2017. I 2018. GODINE 32
Mnoge jedinice lokalne samouprave aplicirajući putem EU fondova, no i iz vlastitih sredstava potiču razvoj OPG-ova koji se bave proizvodnjom peradi. Međutim, zaštitom izvor-nih pasmina peradi (Zagorski puran, Kokoš Hrvatica itd.) počinje ozbiljnije uključivanje većeg broja peradara u uzgoj te se time povećava i potreba za zdravstvenim unapređe-njem, terenskom edukacijom i savjetovanjem o podizanju konkurentnosti OPG-ova koji se bave uzgojem izvornih pasmina peradi. Prepoznavši rad Centra za peradarstvo, došlo je do aktivne suradnje te je na području zasad jedne županije uspostav-ljena suradnja u procesu razvoja uzgoja Zagorskog purana. Uvidom u tehnologiju uzgoja i držanja Zagorskog purana u velikom broju OPG-ova zatečeni su značajni tehnološki nedostatci (neprimjereni objekti, slabo čišćenje i raskužba, nesnalaženje u procesu sakupljanja i skladištenja jaja, ne-primjereni ispusti, hranidba itd.), uginuća purića tijekom tzv. „tople faze“, često zbog nekvalitetne i neprimjerene prehrane u prvim danima života. Ukupno je nadzirano 19 obiteljskih gospodarstava koja se bave uzgojem Zagorskog purana. Tablica 12. prikazuje broj bolesnih jata prema vrsti uzročnika. Tablica 12. Broj pozitivnih jata zagorskog purana na prisut-
nost mikroorganizama i parazita Table 12. Number of Zagorje turkey flocks positive for
microorganisms and parasites
Uzročnik / Causative agent Broj pozitivnih jata Number of positive flocks
Mycoplasma gallisepticum 10 Escherichia coli 11 Clostridium perfingens 6 Histomonas meleagridis 4 Corona virus 1 Enterococcus 1 Glistavost / Worm infestation 1 Velika greška koja se javlja u gotovo svih uzgajivača Zagor-skog purana je učestala i nasumična upotreba antimikrobnih lijekova koje imaju na OPG-u ili njima raspolažu područne Veterinarske stanice (ambulante). Nekontroliranom, često
nepotrebnom i krivom upotrebom antimikrobnih lijekova (bez laboratorijske dijagnostike, a vrlo često i bez kliničkog pregleda purana) stvara se rezistencija bakterija koja u ko-načnici može uzrokovati velika uginuća i nesagledive štete u gospodarstvima. Na temelju provedenih testova antimikrob-ne osjetljivosti tijekom protekle godine u Centru za pera-darstvo utvrđena je vrlo različita osjetljivost izdvojenih sojeva bakterija pa se nasumičnim davanjem antimikrobnih lijekova nanosi šteta kako na porast rezistencije mikroorga-nizama u životinja tako i potencijalna opasnost rezistencije na antimikrobne lijekove u suzbijanju obolijevanja ljudi. Također, međusobna nekontrolirana prodaja rasplodnih jaja, miješanje jaja u „divljim“ valionicama, razmjena jednodnev-nih purića i zamjena purana između uzgajivača doprinijela je širenju mikoplazmoze u uzgojima, što rezultira trenutnim, ali i mogućim izrazito velikim problemima u budućnosti. Mikoplazmoza kao teško izlječiva bolest koja je zahvatila velik broj jata slobodno držanih purana predstavlja potenci-jalnu opasnost u intenzivnoj peradarskoj proizvodnji. Tako-đer, pojava i prisutnost histomonijaze, za koju nažalost na području Europske Unije nema dozvoljene učinkovite tera-pije, mogla bi stvarati veće probleme s obzirom na dugotraj-nost infestacije pašnjaka. Stoga je važna kontrola i stalna prisutnost veterinarskog nadzora te pravodobno prepozna-vanje i liječenje kako bi se spriječile štete većih razmjera.
Imunoprofilaksa
Širok spektar djelatnosti navedenih u ovom radu zasigurno zaokružuje i izrada programa imunoprofilakse. Naime, uz razvoj i velika ulaganja u tehnologiju proizvodnje i selekciju pasmina peradi razvijala se i proizvodnja i primjena cjepiva od osnovnih zakonom propisanih cjepiva do današnjih pristupa imunoprofilaksi čitavog niza bolesti. Sve je to zahtijevalo trajnu edukaciju i praćenje svjetskih dostignuća u imunologiji kako bi se peradarima dao pravovaljani odgovor na pitanje prema kojem programu cijepiti i hoće li program biti učinkovit te perad zaštićena od bolesti. Za uspješnu peradarsku proizvodnju uz opremu, hranidbu i znanje djelatnika na farmi od iznimne je važnosti veteri-narski nadzor, pravodobna dijagnostika i sagledavanje cjelokupne proizvodnje. Svakodnevno ulaganje u znanje i osposobljenost djelatnika u svim procesima proizvodnje učinila je Centar za peradarstvo već više od pola stoljeća nezaobilaznom karikom u peradarstvu Hrvatske.
ACTIVITIES OF THE POULTRY CENTRE LABORATORIES IN 2017 AND 2018
Summary
The main task of the Poultry Centre is laboratory and field diagnosis of infectious, parasitic and other poultry diseases, monitoring of their occurrence, spread and control, testing of safety of animal feed, raw materials and feed additives used in production. The paper presents activities of the Poultry Centre laboratories, as well as the prevalence of particular viral, bacterial and parasitic poultry diseases, and diseases due to technological and feeding errors in 2017 and 2018. Key words: diagnosis, epizootiology, poultry farming, Croatia

PERADARSKI DANI 2019. 33
CHARACTERISTICS OF AVIAN BOTULISM TYPE C OUTBREAKS IN COMMERCIAL BROILERS IN SLOVENIA
Olga Zorman Rojs1, Rahela Juršič Cizerl2, Liljana Štalcer2, Tea Poljanec2, Brigita Slavec1
1 University of Ljubljana, Veterinary faculty, Institute for Poultry, Birds, Small Mammals and Reptiles, Ljubljana, Slovenia 2 Veterinarska ambulanta Perutnina Ptuj, Veterinarstvo d.o.o., Ptuj, Slovenia
Summary
Avian botulism is a flaccid paralytic disease caused by ingestion of the botulinum neurotoxin produced by Clostridium botulinum. Although intoxications could be relatively often found in waterfowl, outbreaks are sporadic in commercial poultry. In the last two years, several outbreaks of botulism occurred in broiler flocks aged from two to six weeks. In all flocks, higher sudden mortality, often seen only in one part of the barn, was reported by poultry keepers. Most prominent clinical signs included leg weakness and flaccid paralysis of the neck, drooping wings and tail; in some cases also laboured breathing was observed. In all outbreaks, type C botulinum toxin was identified in serum samples taken from diseased birds. Higher mortality lasted from two days to more than one week depending on the rapidity of interventions taken. Although several control measurements were applied to prevent recurrent outbreaks, on some farms intoxications were confirmed in two to four following fattening periods. The most important step to minimise the severity of intoxication within the affected flock was rapid collection and disposal of dead and diseased birds. Key words: avian botulism, poultry, broilers
Introduction Botulism is an intoxication that affects animals and humans. Up to now, eight different serotypes of C. botulinum neurotoxins (BoNT) are described: A, B, C1, C2, D, E, F and G. Intoxications are a significant cause of morbidity and mortality in free-living waterfowl, game birds and poultry. Outbreaks in poultry were reported in broilers, layers, turkeys, pheasants and ostriches. The majority of botulism in poultry was caused by C (C1/C2) toxin from type C strains of C. botulinum, although C/D mosaic type has also been reported in some Asian and European countries (Anniballi et al., 2013; Jang et al., 2014; Popp et al., 2012; Skarin et al., 2010; Skarin et al., 2013). Outbreaks may be induced after the ingestion of toxin by cannibalization of carcasses; maggots are a particularly rich source of toxin. Another pathway is toxicoinfection associated with cecal colonisation with C. botulinum, production of BoNT and ingestion of BoNT present in droppings. Neurotoxin inhibits acetylcholine release in cholinergic nerve endings and intoxication is clinically
characterized by ascending paralysis that begins with legs and progressively moves anteriorly to involve wings, neck and eyelids. The severity of clinical signs is proportional to the intoxication dose with mortality ranges from 1% to more than 40% (Skarin et al., 2013). The purpose of this study was to analyse 13 cases of avian botulism that occurred in 2017 and 2018 in Slovenia. We report the clinical presentation in broiler flocks aged from two to six weeks, along with diagnostic analyses.
Material and Methods
Affected broiler flocks From 2017 to 2018, 13 poultry flocks associated with botulism outbreaks were investigated. Epidemiological data for each flock/farm are shown in Table 1. Veterinarians reported onset of high mortality with clinical signs of depression and leg weakness. From affected flocks, various samples were collected and sent for further laboratory investigations (Table 1).
Prof. dr. Olga Zorman Rojs, tel.:+386 1 4779 242, [email protected]

CHARACTERISTICS OF AVIAN BOTULISM TYPE C OUTBREAKS IN COMMERCIAL BROILERS IN SLOVENIA 34
Table 1. Data on affected flocks, samples received and diagnostic tests performed
Case number
Farm ID1
Date of receipt
Age at onset (days) Population Samples received Samples tested by
MLA2 Additional diagnostic
investigations 630/17 1 27.3.2017
24 27.800 Carcasses, blood samples, cloacal
and tracheal swabs Serum, liver Necropsy, histology, AI,
adeno
841/17 2 4.5.2017 31 13.400 Carcasses, blood samples Serum Necropsy, bacteriology 1107/17 3 28.6.2017
23 29.900 Blood samples (healthy and moribund
birds), tracheal and cloacal swabs, feed, organs
Serum, feed Histology, AI, TRTV, MG, MS, adeno
1232/17 1 26.7.2017
28 27.800 Blood samples, gizzard content, feed Serum, gizzard content, feed
/
1253/17
4 1.8.2017 32 18.960 Carcasses, blood samples feed, gizzard content
Serum, feed, gizzard content
Necropsy, bacteriology, AI, ILT, adeno, IBV
1283/17
5 11.8.2017 22 33.500 Carcasses, blood samples, feed
Serum Necropsy, bacteriology, AI, ILT, TRT, IBV
129/18 6 19.2.2018 23 14.000 Blood samples Serum / 519/18 3 1.5.2018 12 30.000 Carcasses, blood samples Serum, liver Necropsy, histology,
bacteriology, AI, 850/18 1 10.7.2018 18 27.200 Carcasses,
blood samples Serum, liver
Necropsy, AI
886/18 7 18.7.2018 28 31.800 Carcasses, blood samples
Serum Necropsy, AI, bacteriology
1079/18 1 30.8.2018 13 27.200 Carcasses, blood samples
Serum Necropsy
1345/18 8 26.9.2018 18 11.300 Blood samples Serum / 1489/18 9 23.10.2018 34 14.400 Blood samples Serum /
1identification 2mouse lethality bioassay Detection of C. botulinum toxins and determination of toxin type For each flock, 5 to 10 serum samples were pooled to a single sample and sent to the Institute for Microbiology and Parasitology of Veterinary Faculty to analyse it by standard mouse lethality bioassay test. The toxin type was determined by neutralization of the toxin with specific antitoxins. Samples of feed, content of gizzard and organs were also examined by the same methods. In addition, pooled serum sample obtained from healthy chickens from one of the affected flock was tested. Other investigations performed Pathological examinations were performed on dead birds from 8 affected flocks. Various organs (liver, spleen, heart, kidney, bursa of Fabricius, thymus, small intestine) were collected during necropsy and sent for further histopathological and bacteriological examinations. Cloacal, oropharyngeal swabs and organs were analysed by previously published molecular methods for the detection of pathogens: avian influenza virus (Spackman et al., 2002),
herpesvirus (Van Devanter et al., 1996), coronavirus (Muradrasoli et al., 2009), adenovirus (Wellehan et al., 2004) and mycoplasmas (Raviv and Kleven, 2009). In two outbreaks, feed samples were analysed for the level of the coccidiostats narasin and nicarbasin. Examinations were performed at the Institute of Food Safety, Feed and Environment of Veterinary Faculty.
Results and Discussion
Botulism in poultry occurs sporadically. Outbreaks were reported in broiler flocks, meat type turkeys, and recently also in adult hens (Dohms et al., 1982; Popp et al., 2012; Souillard el al., 2014; Sato el al., 2016; Trampel et al., 2005). In the past, botulism outbreaks in Slovenia were rare, but the situation has been changed in the last two years. This study describes 13 recent botulism outbreaks in commercial broilers which occurred in 2017 and 2018. In all outbreaks, C. botulinum type C toxin was identified by standard mouse bioassay using pooled serum samples taken from moribund birds. Serum samples collected from 10

CHARACTERISTICS OF AVIAN BOTULISM TYPE C OUTBREAKS IN COMMERCIAL BROILERS IN SLOVENIA 35
healthy birds from one of the affected flocks were negative. In five suspected cases, other types of samples were also examined. C. botulinum type C toxin was confirmed only in one liver sample (out of four examined), while feed (3 samples) and gizzard content (2 samples) were negative. Although the mouse bioassay is considered as the golden standard for botulinum confirmation, it is time consuming and requires the use of animals. Real-time PCR has recently been developed for the detection C. botulinum producing type C, D C/D and D/C toxins (Lindberg et al., 2010; Lindström and Korkeala, 2006; Skarin et al., 2010); unfortunately, these diagnostic techniques were not used in our study. Results showed serum to be the most suitable type of samples if no other methods are available. The negative result in the pooled serum sample from healthy birds indicated the importance of appropriate sampling
procedure. It is essential to take serum samples from clinically affected or moribund birds. Most outbreaks (11/13) occurred during late spring and autumn, and only in two flocks botulism occurred in winter months (February, March), which is in agreement with other reports of botulism in poultry (Dohms et al., 1982; Popp et al., 2012; Souillard et al., 2014). Clinical signs at the flock level were always related to a sudden onset of high mortality. In the beginning of the illness, dead or dying birds were not seen throughout the buildings, but were mostly localized in one area. Affected chickens were depressed and pale, they could not walk, and later on drooping head, wings and tail were displayed. Individual birds also showed laboured breathing before death. Dead birds were found laid on the chest with head crawled in litter. In half of the affected flocks, diarrhoea was also noticed.
Table 2. Duration of illness, mortality due to botulism outbreak, additional infections confirmed, total mortality and feed
conversion in affected flocks
Case number Farm ID Duration of higher mortality (days of age)
Mortality due to botulism (%)
Additional infections confirmed Feed conversion
Total mortality (%)
630/17 1 14 days (24-37) 11.36 Coccidiosis,
adenovirus 1.65 18.96
841/17 2 4 days (31-34) 2.82 Neg. 1.83 6.66
1107/17 3 9 days (23-32) 12.38 Neg. 1.86 17.13
1232/17 1 13 days (28-41) 12.53 / 1.87 14.48
1253/17 4 10 days (32-42) 7.70 Staphylococcus sp. 1.73 10.69
1283/17 5 10 days (22-32) 6.12 Neg. 1.70 8.09
129/18 6 8 days (23-31) 2.1 / 1.80 4.55
519/18 3 15 days (12-27) 7.04 E. coli 1.74 14.94
850/18 1 2 days (28-29) 1.7 Neg. 1.71 8.44
886/18 7 15 days (28-43) 9.33 Neg. 14.58
1079/18 1 3+6 days (13-15 and 32-37) 4.41 / 1.77 6.45
1345/18 8 5 days (18-22) 6.57 / 1.67 18.15
1489/18 9 6 days (34-39) 5.13 / 1.73 8.05

CHARACTERISTICS OF AVIAN BOTULISM TYPE C OUTBREAKS IN COMMERCIAL BROILERS IN SLOVENIA 36
Routine diagnostic gross examination was performed on 56 carcases from 8 affected flocks. All necropsied chickens were in good condition. The majority had pale combs and conjunctivae, some were cyanotic. Urates and faecal material adhered to cloaca, beak, nears and eyelids. Blood was not clotted. The crop was empty in all examined birds, although feed particles mixed with litter and feathers were occasionally found in gizzard. Liver was enlarged and hyperaemic, while spleen was of normal size with subs-capular petechial haemorrhages noted. Kidneys were slightly enlarged and pale. Hyperaemia of the proximal part of trachea was seen, and in individual birds lungs were also hyperaemic. In the small intestine, thickening of the wall was noticed and petechial haemorrhages were present in the majority of necropsied birds. In caeca and colon, no specific gross lesions were seen. In one outbreak (case number 630/17), gross lesions specific for coccidiosis (E. acervulina and E. tenella) were observed. On microscopic analyses, no specific lesions were found. Mild degeneration of tubulo-cytes and myocytes was noticed. Liver was congested and large haemorrhages could be present. In the intestine, desquamation and karyorrhexis of epithelial cells were regularly found. No microscopic lesions were noticed in other organs (bursa of Fabricius, thymus, brain, pancreas, oesophagus, proventriculus). Although clinical symptoms and absence of specific post mortem changes could be indicative of botulism, additional investigations were performed to exclude other infections. All samples were negative for AIV, herpes and corona virus. Only in one flock (case number 630/17), adenovirus was detected. Bacteriology was performed on liver and heart samples collected from dead birds of seven affected flocks. Staphylococcus spp. and E. coli (some colonies) were isolated from organs (liver, heart) taken from dead birds from two outbreaks, while all other samples were negative. In 2017, Maxiban was used in broiler feed. It is known that nicarbazin medicated broilers may show reduced heat tolerance if exposed to high temperature and humidity. In two flocks, high mortality occurred after new feed had been delivered and feed samples were analysed for the level of narasin and nicarbasin. The results did not indicate that higher mortality could be related to this particular coccidiostat. Higher mortality lasted from two days to more than two weeks. In one flock, higher mortality occurred twice in the same fattening period. The mortality due to botulism varied from 1.70% to 12.53%. The highest and lowest mortality due to botulism was recorded in two flocks of four-week-old broilers. At the end of the production period, total losses varied from 6.45% to almost 19%. Economic losses were high, although feed conversion was not affected (Table 2). On all farms, reinforced biosecurity measures were implemented. Farmers were told to rapidly remove carcasses, which was not easy to do due to the high number of dead birds and ongoing mortality. It seems that this was the most important step to minimise the severity of intoxication within the affected flock. Detailed plan of
cleaning and disinfection of houses and surrounding area was applied, preceded by rodent control. Flocks were treated with amoxicillin, but it looks that the treatment had no effect on mortality or duration of the illness. The epidemiology of botulism in poultry is still poorly understood. C. botulinum is an anaerobic organism found in the soil and spores can actively survive decades in the environment. Uptake of the toxin, in particular through maggots on carcasses, which become available during periods of high mortality, is understood as an important factor in wild birds. An important role of carcasses has been suspected also in poultry botulism (Dohms et al., 1982), although recent studies indicated that healthy poultry do not normally harbour toxigenic C. botulinum (Hardy and Kaldhusdal, 2013). During the two-year period, altogether nine production farms were affected. At one farm (Farm ID 1), it was not the first botulism outbreak. This particular farm had an outbreak in August 2016. The farm is located near the forest. Beside commercial broilers, pigs, rabbits and cats were also seen walking freely along the premises on the same location, but none of the animals died. After the outbreak in 2016, several interventions were taken. The house and its surrounding was cleaned and disinfected, and rodent control was progressed. The house was empty for 6 months and the farmer was strongly requested to restrict movements of personnel and domestic animals. The next botulism outbreak on the same farm (Farm 1) was confirmed after one production cycle without any health problems. In the observed two-year period, three additional outbreaks were recorded on the same farm, although not in subsequent production cycles. It seems that C. botulinum spores had remained on the farm in spite of all interventions taken. A very similar situation was also seen on another farm (Farm ID 3), where the first outbreak occurred in June 2017 and the second one a year later, after six production cycles with no clinical signs of botulism. Recent epidemiological studies show that darkling beetles, drinking water, dust and litter are the main contaminated areas. In addition, C. botulinum was detected after cleaning and disinfection of poultry houses; most frequently, samples taken from ventilation systems were positive (Souillard et al., 2017). A mouse bioassay, which was used for botulism confirmation, is not appropriate for running a large number of samples. Due to this limitation, no further epidemio-logical studies were performed and as in most of the outbreaks described, the original source of C. botulinum intoxication remained unknown.
Conclusions
Outbreaks of botulism in poultry are known to be associated with high mortality and to cause significant economic losses. In outbreaks with low mortality or where the illness is not recognised, botulism may never be suspected. As such, it needs to be included in the differential diagnosis in all cases of the sudden onset mortality in commercial poultry flocks.

CHARACTERISTICS OF AVIAN BOTULISM TYPE C OUTBREAKS IN COMMERCIAL BROILERS IN SLOVENIA 37
References
ANNIBALLI, F., A. FIORE, C. LOFSTROM, H. SKARIN, B. AURICCHIO, C. WOUDSTRA, L. BANO, B. SEGER-MAN, M. KOENE, V. BAVERUD, T. HANSEN, P. FACH, A. TEVELL, M. HEDELAND, E. OLSSON, D. DE MEDICI (2013): Management of animal botulism outbreaks: from clinical suspicion to practical countermeasures to prevent or minimize outbreaks. Biosecurity and Bio-terrorism, 11, 191–199.
DOHMS, J. E., P. H. ALLEN, J. K. ROSENBERGER (1982): Cases of type C botulism in broiler chickens. Avian Dis., 26, 204–210.
HARDY, S. P., M. KALDHUSDAL (2013): Type C and C/D toxigenic Clostridium botulinum is not normally present in the intestine of healthy broilers. Vet. Microbiol., 165 (3–4), 466-468.
JANG, I., M. S. KANG, H. R. KIM, J. Y. OH, J. I. LEE, H. S. LEE, Y. K. KWON (2014): Occurrence of avian botulism in Korea during the period from June to September 2012. Avian Dis., 58, 666-669.
LINDBERG, A., H. SKARIN, R. KNUTSSON, G. BLOMQVIST, V. BÅVERUD (2010): Real-time PCR for Clostridium botulinum type C neurotoxin (BoNTC) gene, also covering a chimeric C/D sequence - Application on out-breaks of botulism in poultry. Vet. Microbiol., 146, 118-123.
LINDSTRÖM, M., H. KORKEALA (2006): Laboratory Diag-nostics of Botulism. Clinical Microbiol. Rev., 19, 298-314.
MURADRASOLI, S., N. MOHAMED, A. HORNYÁK, J. FOHLMAN, B. OLSEN, S. BELÁK, J. BLOMBERG (2009): Broadly targeted multiprobe QPCR for detection of coronaviruses: Coronavirus is common among mallard ducks (Anas platyrhynchos). J. Virol. Methods, 159, 277-287.
POPP, C., R. HAUCK, W. GAD, H. M. HAFEZ (2012): Type C botulism in a commercial Turkey farm: a case report. Avian Dis., 56, 760–763.
RAVIV, Z., S. H. KLEVEN (2009): The development of diagnostic real-time TaqMan PCRs for the four pathogenic avian mycoplasmas. Avian Dis., 53, 103-107.
SATO, Y., W. L. WIGLE, S. GALLAGHER, A. L. JOHNSON, R. W. SWEENEY, P. S. WAKENELL (2016): Outbreak of
Type C Botulism in Commercial Layer Chickens. Avian Dis., 60, 90-94.
SKARIN H., A. LINDBERG, G. BLOMQVIST, A. ASPÁN, V. BÅVERUD (2010): Molecular characterization and comparison of Clostridium botulinum type C avian strains. Avian Pathol., 39, 511-518.
SKARIN, H., G. BLOMQVIST, V. BÅVERUD, V. (2013): Botulism. In Diseases of Poultry 13th Ed. (D.E. Swayne, J. R. Glisson, L. R. McDougald, L. K. Nolan, D. L. Suarez & V. Nair, Eds.), Ames, IA: Blackwell Publishing, 953–957.
SOUILLARD, R., C. WOUDSTRA, C. LE MARÉCHAL, M. DIA, M. H. BAYON-AUBOYER, M. CHEMALY, P. FACH, S. LE BOUQUIN (2014): Investigation of Clostridium botulinum in commercial poultry farms in France between 2011 and 2013. Avian Pathol., 43, 458-464.
SOUILLARD,R., C. LE MARÉCHAL, V. BALLAN, S. ROUXEL, D. LÉON, L. BALAINE, T. POËZEVARA, E. HOUARD, B. ROBINEAU, C. ROBINAULT, M. CHEMALY, S. LE BOUQUIN (2017): Investigation of a type C/D botulism outbreak in free-range laying hens in France. Avian Pathol., 46, 195-201.
SPACKMAN, E., D. A. SENNE, T. J. MYERS, L. L. BULAGA, L. P. GARBER, M. L. PERDUE, K. LOHMAN, L. T. DAUM, D. L. SUAREZ (2002): Development of a real-time reverse transcriptase PCR assay for type A influenza virus and the avian H5 and H7 hemagglutinin subtypes. J. Clin. Microbiol. 40, 3256-3260.
TRAMPEL, D. W., S. R. SMITH, T. E. ROCKE (2005): Toxicoinfectious botulism in commercial caponized chickens. Avian Dis., 49, 301-303.
VAN DEVANTER, D. R., P. WARRENER, L. BENNETT, E. R. SCHULTZ, S. COULTER, R. L. GARBER, T. M. ROSE (1996): Detection and analysis of diverse herpesviral species by consensus primer PCR. J. Clin. Microbiol., 34, 1666-1671.
WELLEHAN, J. F., A. J. JOHNSON, B. HARRACH, M. BEN-KÖ, A. P. PESSIER, C. M. JOHNSON, M. M. GARNER, A. CHILDRESS, E. R. JACOBSON (2004): Detection and analysis of six lizard adenoviruses by consensus primer PCR provides further evidence of a reptilian origin for the atadenoviruses. J Virol., 78, 13366-1369.
ZNAČAJKE POJAVE PTIČJEG BOTULIZMA TIPA C U TOVNIH PILIĆA U SLOVENIJI
Sažetak
Ptičji botulizam je mlohava paraliza uzrokovana unošenjem botulinum neurotoksina koji proizvodi bakterija Clostridium botulinum. Iako se intoksikacija relativno često nalazi u divljih ptica, sporadične epidemije bilježe se kod komercijalne peradi. U posljednje dvije godine zabilježeno je nekoliko epidemija botulizma u jatima brojlera u dobi od dva do šest tjedana. U svim jatima peradari su izvijestili o povišenoj stopi iznenadne smrtnosti, često u samo jednom dijelu nastambe. Najizraženiji klinički znaci bili su slabost nogu i mlohava paraliza vrata, obješena krila i rep; u nekim slučajevima zapaženo je i otežano disanje. U svim tim epidemijama utvrđen je botulinum toksin tipa C u uzorcima seruma uzetih od oboljelih ptica. Povišena smrtnost potrajala je od dva dana do više od jednog tjedna, ovisno o brzini provedbe intervencija. Iako je provedeno više mjera kako bi se spriječile opetovane epidemije, na nekim gospodarstvima je intoksikacija potvrđena u dva do četiri sljedeća tovna razdoblja. Najvažniji korak za smanjenje težine intoksikacije u zahvaćenom jatu bilo je brzo prikupljanje i uklanjanje uginulih i oboljelih ptica. Ključne riječi: ptičji botulizam, perad, brojleri

PERADARSKI DANI 2019. 38
ZNAČENJE BAKTERIJA RODA CHLAMYDIA SP. U JATIMA PERADI I NJIHOV ZOONOTSKI POTENCIJAL
Danijela Horvatek Tomić1, Ana Marquiza Quilicot2, Željko Gottstein1, Maja Lukač1, Estella Prukner-Radovčić1
1 Sveučilište u Zagrebu, Veterinarski fakultet, Zagreb, Hrvatska 2 College of Veterinary Medicine, Visayas State University, ViSCA, Baybay City, Leyte, Filipini
Sažetak
Klamidioza peradi, često smatrana emergentnom bolesti, najčešće je povezana s bakterijom Chlamydia psittaci (C. psittaci), ali u novije vrijeme i s novo opisanom vrstom C. gallinacea. Obje spomenute bakterije imaju ekonomsko značenje u uzgojima peradi, dok je C. psittaci poznata i kao uzročnik s iznimnim zoonotskim potencijalom. Ovaj rad prikazuje pojavnost klamidija u peradi diljem svijeta, ali i u Hrvatskoj, gdje je u zadnjem istraživanju dokazana prisutnost C. gallinacea u čak 46,3% pretraženih uzoraka. Ključne riječi: Chlamydiaceae, C. psittaci, C. gallinacea, perad, zoonoza Uvod Klamidiozu peradi uzrokuju bakterije roda Chlamydia, najčešće C. psittaci. Prema taksonomiji iz 2015. godine, ovaj rod sastoji se od 11 vrsta, od kojih su u ptica, uz spomenute, opisani i nalazi C. abortus, C. pecorum, C. trachomatis, C. suis i C. muridarum te dvije nove vrste, C. avium i C. gallinacea (Pantchev i sur., 2009.; Sachse i sur., 2014.; Guo i sur., 2016.; Szymańska-Czerwińska i sur., 2017.). Dok je većina vrsta uglavnom vrsno specifična, C. psittaci može uzrokovati bolesti u različitih vrsta životinja (osobito značajni pobačaji u kobila; Polkinghorne i Greub, 2017.; Jenkins i sur., 2018.; Jelocnik i sur., 2018.), ali je opisan i nalaz ove bakterije u klinički zdravih goveda, ovaca i svinja (Kauffold i sur., 2006.; Kemmerling i sur., 2009.; Lenzko i sur., 2011.). Bolest koju ova bakterija uzrokuje u ljudi najčešće se naziva psitakoza ili ornitoza, a kao najčešći izvor bolesti za ljude spominju se papige ili perad (purani i patke) (Petrovay i Balla, 2008.; Beeckman i Vanrompay, 2009.). Donedavno se smatralo da je C. psittaci jedina koja uzrokuje bolest u peradi, ali i u ptica općenito. Novija istraživanja ukazuju na to da se C. gallinacea može smatrati predominantom vrstom u peradi, a dosad je opisana u domaće peradi, purana i pataka (Guo i sur., 2016.; Quilicot i Prukner-Radovčić., 2018.). S druge strane, C. avium
pretežno je nađena u golubova i papiga (Sachse i sur., 2014.). Patogenost dviju novih vrsta klamidija za ptice još nije do kraja istražena, no sa sigurnošću se može tvrditi da je C. avium uzrokovala dišne simptome u papiga i golubova (Sachse i sur., 2014.). Ipak, dosad nije opisana pojava drugih kliničkih znakova vezanih za prisutnost C. gallinacea u peradi, osim gubitka težine (Guo i sur., 2016.; Li i sur., 2017.). Stoga se ova bakterija može smatrati komenzalom u prirodnom domaćinu, peradi (Sachse i Laroucau, 2015.). Zoonotski potencijal klamidija dobro je poznat, no onaj C. gallinacea i C. avium još uvijek nije u potpunosti potvrđen. Opisani su slučajevi obolijevanja ljudi koji su bili u kontaktu s pozitivnom peradi u klaonici (Laroucau i sur., 2009a.), no nakon toga nije bilo novih dokaza o obolijevanju ljudi zbog C. gallinacea.
Etiologija i patogeneza
Bakterije roda Chlamydia imaju karakterističan bifazični razvojni ciklus koji uključuje razvoj elementarnih i reti-kularnih te intermedijarnih tjelešaca (Matsumoto i Manier, 1970.). Elementarna tjelešca su male okrugle forme koje predstavljaju infektivni oblik te se prihvaćaju za različite stanice domaćina, od epitelnih do makrofaga (Knittler i Sachse, 2015.). Unutar stanice stvaraju metabolički aktivna
Izv. prof. dr. sc. Danijela Horvatek Tomić; Zavod za bolesti peradi s klinikom, Veterinarski fakultet, Sveučilište u Zagrebu, Heinzelova 55, 10000 Zagreb, Hrvatska; Tel: 012390281, fax: 012390410; Email: [email protected]

ZNAČENJE BAKTERIJA RODA CHLAMYDIA SP. U JATIMA PERADI I NJIHOV ZOONOTSKI POTENCIJAL 39
retikularna tjelešca koja nakon binarne diobe opet prelaze u elementarna tjelešca i zaražavaju susjedne stanice. Osim specifičnog razvojnog ciklusa, klamidije su značajne i zbog svojih razlika u virulentnosti. Bakterija C. psittaci jedna je od genetski najraznovrsnijih klamidija te je tako opisano 15 različitih genotipova, temeljeno na ompA genu (Sachse i sur., 2008.). Potrebno je i napomenuti da su ranijih godina spominjani serovarovi zapravo ekvivalenti genotipovima klamidija (Vanrompay i sur., 1997a.; Sachse i sur., 2008.). Te genotipove možemo povezivati i s određenim vrstama ptica, no razlikuju se i po svojoj patogenosti za ljude. Poznato je da se genotip A prvenstveno javlja u psitacina, genotip B u golubova, C u pataka i gusaka, D u purana, E i E/B u golubova odnosno pataka, F u psitacina i purana, WC u goveda te M56 u glodavaca (Harkinezhad i sur., 2009.; Vanrompay, 2013.). Zoonotski potencijal uzrokovanja bolesti u ljudi naročito pokazuju genotipovi A, B i E/B (Heddema i sur., 2006.; Vanrompay i sur., 2007.). Primje-nom molekularnih metoda dokazano je da se i C. gallinacea, temeljeno na ompA genu, genetski dosta razlikuje. Tako su Guo i sur. (2016.) opisali najmanje 13 genetskih varijanti (najvjerojatnije 13 različitih serovarova) ove bakterije, a Szymańska-Czerwińska i sur. (2017.) devet genetskih vari-janti. U Sjedinjenim Američkim Državama (državi Alaba-ma), Li i sur. (2017.) opisali su 14 biotipova C. gallinacea u 16 sekvenciranih uzoraka. Uz pojedinačni nalaz čest je opis i miješanih infekcija klamidijama. Opisani su nalazi C. psittaci uz C. avium i C. gallinacea, ne samo u istom jatu, već i u pojedinim jedinkama istodobno (Krautwald-Jung-hanns i sur., 2013.; Laroucau i sur., 2014.). U peradi, naročito kod purana, opisane su i miješane infekcije drugim bakterijama, npr. Ornithobacterium rhinotracheale (van Loock i sur., 2005.) ili virusima (van Lock i sur., 2006). Na-čin prijenosa klamidioze je najčešće direktnim kontaktom, osobito kontaminiranim izmetom, iscjetkom iz nosa ili konjunktiva, a klamidije ulaze u domaćina dišnim putem (Harkinezhad i sur., 2009.). S druge strane, dokazan je i vertikalni prijenos klamidija u jatima purana i pataka (Lublin i sur., 1996.). Značajnu ulogu u prijenosu uzročnika, ali i u težini kliničkih znakova mogu imati i ektoparaziti kao što su tekuti (Circella i sur., 2011.) te divlje ptice (Sachse i Laroucau, 2015.).
Klinička slika i patoanatomski nalaz
Pojavnost klamidioze u peradi uzrokuje značajne ekonom-ske gubitke, a sama bolest može biti fatalna i za ptice i za ljude. Klinički znakovi u peradi su većinom nespecifični i uključuju anoreksiju, dišne poremetnje, zeleni proljev, ko-njunktivitis, smanjenu proizvodnju jaja i pad tjelesne težine, a težina kliničke slike ovisi o vrsti klamidije i patogenosti genotipa, dobi i vrsti peradi (Andersen i Vanrompay, 2000.; Vanrompay, 2013.). U pataka i gusaka moguć je nalaz kliničkih znakova živčanog sustava, a u purana unilateralno ili bilateralno otečenje iznad očiju (Shivaprasad, 2015.). Velik problem predstavljaju inaparentne infekcije koje se naročito javljaju u peradi, a takve životinje dulje vrijeme mogu biti kliconoše (Guo i sur., 2016.). Patoanatomski,
nalazi se hepato- i splenomegalija, uz konjunktivitis, upalu zračnih vrećica, perikarditis, perihepatitis i peritonitis, no moguće su i teže promjene ako se uz klamidije pojave i drugi mikroorganizmi (Shivaprasad, 2015.). Patohistološke promjene nisu patognomonične, no tipičan je nalaz elemen-tarnih ili retikularnih tjelešaca u citoplazmi makrofaga ili epitelnih stanica obojenih posebnim bojenjima (Shivaprasad, 2015.). Kada je u pitanju C. gallinacea, dosad je opisan jedino gubitak težine pa se zapravo o patogenosti ove vrste još uvijek nedovoljno zna (Guo i sur., 2016.).
Dijagnostika
Radi postavljanja etiološke dijagnoze najčešće se zaživotno uzimaju skupni uzorci izmeta odnosno obrisci konjunktiva, ždrijela i kloake (tzv. trostruki obrisci) (Quilicot i sur., 2017.a). U današnje vrijeme dijagnostika se najčešće temelji na dokazu DNK molekularnim metodama, konvencional-nom te lančanom reakcijom polimerazom u stvarnom vre-menu (PCR), čime se omogućava brzi, osjetljivi i specifični dokaz klamidija (Anon., 2018.). Kako C. psittaci više nije jedina vrsta koja može uzrokovati bolest u peradi, neophod-no je ne samo dokazati prisutnost bakterija roda Chlamydia, već i odrediti vrstu specifičnim početnicama (Ehricht i sur., 2006; Zocevic i sur., 2012.). U dijagnostici moguće je kori-stiti i sekvencijsku DNA-čip tehnologiju (DNA microarray) koja je iznimno prikladna metoda za dokaz pojedine vrste klamidija (Sachse i sur., 2005.). Iako je ranije zlatni standard dijagnostike predstavljalo umnažanje klamidija u kokošjim zametcima ili kulturi stanica, danas se ove metode puno rjeđe koriste, osobito zbog opasnosti za zdravlje djelatnika u laboratorijima. Stoga je pridržavanje biosigurnosnih mjera neophodno u laboratorijima u kojima se radi s potencijalnim zoonotskim mikroorganizmima. Ako molekularne metode nisu dostupne, može se primijeniti citološka pretraga uz spe-cifična bojenja otisnih preparata organa ili eksudata, imuno-histokemijske metode i tzv. enzyme-linked immunosorbent assay (ELISA) testove (Anon., 2018.). Dokaz specifičnih protutijela nije naročito značajna metoda u dijagnostici klamidioze zbog dugotrajne prisutnosti protutijela i visokog postotka pojavnosti ovih bakterija u jatima peradi. Stoga je neophodno pretražiti parne serume, no dokaz specifičnih protutijela ne mora upućivati na akutnu infekciju, a mogući su i lažno negativni rezultati (Anon., 2018.). Ako se ipak dokazuju protutijela, najčešće se primjenjuje reakcija veza-nja komplementa te ELISA testovi, no u novije vrijeme opisana je i uporaba ekspresijske DNA-čip tehnologije (Sachse i sur., 2018.).
Pojavnost
Iako je klinički manifestna klamidioza naročito zabilježena u jatima purana i pataka (Newman i sur., 1992.; Vanrompay i sur., 1997b.; Dickx i Vanrompay, 2001.; van Loock i sur., 2005.), sve je češći nalaz C. psittaci i u kokoši (Zhang i sur., 2008.; Zhou i sur., 2010.; Yin i sur., 2013.). Nažalost, u velikoj većini C. psittaci zarazila je i ljude, naročito one koji su bili u kontaktu sa zaraženom peradi na farmama ili u

ZNAČENJE BAKTERIJA RODA CHLAMYDIA SP. U JATIMA PERADI I NJIHOV ZOONOTSKI POTENCIJAL 40
klaonicama (Pelle-Duporte i Gendre, 2001.; Tiong i sur., 2007.; Gaede i sur., 2008.; Petrovay i Balla, 2008.; Dickx i Vanrompay, 2011.). Klamidioza pataka osobito je česta u zemljama s intenzivnom proizvodnjom, što je najčešće u Aziji i Europi, naročito u Francuskoj te pokazuje izniman zoonotski potencijal (Haas i sur., 2007.; Laroucau i sur., 2009.a,b; Yin i sur., 2015.). Novo otkrivene vrste, C. avium i C. gallinacea, dosad su opisane u golubova i papiga, odnosno peradi. Iako su takva istraživanja još uvijek dosta rijetka, moguće je uvidjeti da je C. gallinacea prisutna u jatima peradi diljem svijeta. Čak i ranije kad su se ove bakterije nazivale „atipičnim klamidijama“ dokazano je da su prisutne u jatima peradi u Europi i Aziji (Zocevic i sur., 2012.), dok novija istraživanja ovu bakteriju nazivaju endemskom u peradi. Tako je C. gallinacea dokazana u 80% pozitivnih uzoraka koji su potjecali od 129 jata peradi u Francuskoj (Hulin i sur., 2015.), a dokazana je i kao pre-dominantna vrsta klamidija u ptica u Kini (63,8% svih pozi-tivnih uzoraka i čak 81,2% pozitivnih uzoraka podrijetlom od peradi) (Guo i sur., 2016.), Poljskoj (C. gallinacea nađena u 65,5% pozitivnih jata i u 73,0% pozitivne peradi (Szymańska-Czerwińska i sur., 2017.) i Francuskoj (16 od 33 jata peradi bila su pozitivna na C. gallinacea (Hulin i sur., 2015.). U Nizozemskoj u istraživanju koje su proveli Heijne i sur. (2018.) u skupnim uzorcima izmeta prikuplje-nim sa 151 farme kokoši nesilica ustanovljen je 71 pozitivan uzorak na prisutnost C. gallinacea, dok ni u jednom nije dokazana C. psittaci. Isto tako, pretražena perad nije po-kazivala znakove bolesti. I jata slobodno držane peradi često su pozitivna na klamidije. Tako je u Italiji opisana prisutnost C. gallinacea u 15% uzoraka uzetih od 160 ptica (Donati i sur., 2018.) te u 12,4% obrisaka ždrijela peradi u SAD, država Alabama (Li i sur., 2017.). Dosadašnja istraživanja u Hrvatskoj provedena su na jatima purana, kokoši nesilica i slobodno držanoj peradi. Ranijih godina u dijagnostici rabili su se uglavnom ELISA testovi te su tako Prukner-Radovčić i sur. (2006a.) dokazali specifični antigen i protutijela protiv C. psittaci u 22,2% odnosno 60% uzoraka podrijetlom od purana. Te iste godine opisan je i slučaj pojave dišnih simptoma uz pad nesivosti u jatu kokoši nesilica, a metodom ELISA dokazan je specifičan antigen klamidija u 78,5% tzv. trojnih obrisaka (Prukner-Radovčić i sur., 2006b.). Sekvenciranjem ompA i 16S gena potvrđen je nalaz atipičnih klamidija u jatu tovnih purana s dišnim simptomima (Prukner-Radovčić i sur., 2012.). U jatima slobodno-živuće peradi opisan je nalaz C. psittaci metodom PCR u stvarnom vremenu te su 4 jata bila pozitivna, uz dokaz genotipa B (Quilicot i sur., 2018.). U istraživanju kojem je cilj bio procjena učestalosti Chlamydia sp. u uzgojima slobodno držane peradi u Hrvatskoj i na Filipinima opisana je prisutnost C. gallinacea u 46,30% pretraženih uzoraka metodom PCR u stvarnom vremenu (Quilicot i Prukner-Radovčić, 2018.).
Liječenje
Liječenje klamidioze peradi uglavnom se temelji na primjeni različitih skupina antibiotika. Klamidije su osjetljive na
tetracikline i fluorokinolone pa se stoga klortetraciklin, doksiciklin i enrofloksacin najčešće primjenjuju u liječenju peradi (Vanrompay, 2013.; Shivaprasad, 2015.). Nažalost, liječenje je dugotrajno, a ptice često ostaju kliconoše. S dru-ge strane, pokusi provedeni na peradi pokazuju mogućnost razvoja cjepiva protiv C. psittaci. Kako se radi o DNA cjepivima, istraživanja su još uvijek u tijeku te cjepivo zasad nije komercijalno dostupno, no svakako bi bilo od velike koristi u suzbijanju ove opasne zoonoze (Quilicot i sur., 2017b.). Najbolja preventiva ogleda se u strogom pridrža-vanju biosigurnosnih mjera, smanjenom kontaktu s divljim pticama i drugim životinjama (Shivaprasad, 2015.).
Zoonotski potencijal
Psitakoza/ornitoza u ljudi posljedica je udisanja kontamini-ranog aerosola. Iako se najčešće kao izvor bolesti za ljude spominju papige, perad također predstavlja izvor ovoga zoonotskog mikroorganizma (Pelle-Duporte i Gendre, 2001.; Laroucau i sur., 2009.; Tiong i sur., 2007.). Najčešće obolijevaju veterinari te radnici na farmama, u valionicama ili klaonicama (Petrovay i Balla, 2008.; Laroucau i sur., 2009a,b.; Dickx i Vanrompay, 2011.). Isto tako, bitno je poštivanje mjera biosigurnosti u laboratorijima u kojima se provodi dijagnostika klamidioze. Inkubacija u ljudi obično traje 5-14 dana, a bolest se može manifestirati kao inapa-rentna ili opasna sistemska infekcija s pojavom pneumonije i u najgorem slučaju smrti (Petrovay i Balla, 2008.; Anon., 2018). Dijagnostika može biti otežana zbog nespecifičnih simptoma te seronegativnosti koja se može pojaviti u nekih bolesnika. Lijekovi izbora najčešće su tetraciklin, doksi-ciklin i azitromicin (Anon., 2018.). U dosad spomenutim slučajevima bolest u ljudi uzrokovala je C. psittaci, no daljnja istraživanja su neophodna kako bi se dokazala stvarna patogenost novih vrsta, C. gallinacea i C. avium.
Zaključak
Opisom dviju novih vrsta klamidija u peradi i ptica općenito nameće se potreba za detaljnijom dijagnostikom ove zoonoze, poglavito s aspekta postavljanja točne etiološke dijagnoze i detaljnije diferencijalne dijagnostike. Isto tako, neophodna su i daljnja istraživanja patogenosti i virulencije C. gallinacea, s posebnim osvrtom na epidemiološka istra-živanja i dokaz zoonotskog potencijala. Istraživanja pojav-nosti C. gallinacea sve su učestalija, no razlikuju se po postotku prevalencije, od 12% do čak 80%. Vrlo vjerojatno te razlike ovise o vrsti pretraženih uzoraka, načinu držanja peradi i primijenjenim biosigurnosnim mjerama na farma-ma. Na kraju, potrebno je naglasiti da bez obzira na vrstu klamidije, u slučaju nalaza unutarstaničnih patogena u pe-radi ili ptica općenito svakako se treba pridržavati strogih mjera biosigurnosti zbog njihovog mogućeg zoonotskog potencijala.
Literatura
ANDERSEN, A. A., D. VANROMPAY (2000): Avian chlamy-diosis. Rev. Sci. Tech., 19, 396-404.

ZNAČENJE BAKTERIJA RODA CHLAMYDIA SP. U JATIMA PERADI I NJIHOV ZOONOTSKI POTENCIJAL 41
ANNONYMOUS (2018): Avian Chlamydiosis. Section 3.3. Chapter 3.3.1. OIE Terrestrial Manual, 2018.
BEECKMAN, D. S. A., D. C. G. VANROMPAY (2009): Zoonotic Chlamydophila psittaci infections from a clinical perspective. Clin. Microbiol. Inf., 15, 1, 11-17.
CIRCELLA, E. N. PUGLIESE, G. TODISCO, M. A. CAFIERO, O. A. E. SPARAGANO, A. CAMARDA (2011): Chlamydia psittaci infection in canaries heavily infested by Dermanyssus gallinae. Exp Appl Acarol. 55, 329–338. DOI 10.1007/s10493-011-9478-9
DICKX, V., D. VANROMPAY (2011): Zoonotic transmission of Chlamydia psittaci in a chicken and turkey hatchery. J. Med. Microbiol., 60, 775–779.
DONATI, M., K. LAROUCAU, A. GUERRINI, A. BALBONI, D. SALVATORE, E. CATELLI, C. LUPINI, A. LEVI, A. DI FRANCESCO (2018): Chlamydiosis in Backyard Chickens (Gallus gallus) in Italy. Vector Borne Zoonotic Dis., 18 (4), 222-225.
EHRICHT, R., P. SLICKERS, S. GOELLNER, H. HOTZEL, K. SACHSE (2006): Optimized DNA microarray assay allows detection and genotypingof single PCR-amplifiable target copies. Mol. Cell. Probes, 20, 60–63.
GAEDE, W., K. F. RECKLING, B. DRESENKAMP, S. KENKLIES, E. SCHUBERT, U. NOACK, H. M. IRMSCHER, C. LUDWIG, H. HOTZEL, K. SACHSE (2008): Chlamydophila psittaci infections in humans during an outbreak of psittacosis from poultry in Germany. Zoon. Pub. Health, 55, 184–188.
GUO, W., J. LI, B. KALTENBOECK, J. GONG, W. FAN, C. WANG (2016): Chlamydia gallinacea, not C. psittaci, is the endemic chlamydial species in chicken (Gallus gallus). Sci. Rep. 18, 19638. doi: 10.1038/srep19638.
HAAS, W. H., C. M. SWAAN, A. MEIJER, G. NEVE, U. BUCHHOLZ, M. BEER, J. E. VAN STEENBERGEN, G. KRAUSE (2007): A Dutch case of atypical pneumonia after culling of H5N1 positive ducks in Bavaria was found infected with Chlamydophila psittaci. Euro Surveill. 12 (11), E071129071123.17.
HARKINEZHAD, T., T. GEENS, D. VANROMPAY (2009): <Chlamydophila psittaci infections in birds: a review with emphasis on zoonotic consequences. Vet. Microbiol., 135, 68–77.
HEDDEMA, E. R., E. J. VAN HANNEN, B. DUIM, C. M. J. E. VANDENBROUCKE-GRAULS, Y. PANNEKOEK (2006): Genotyping of Chlamydophila psittaci in Human Samples. Emerg. Infect. Dis., 12,1989–1990.
HEIJNE, M., J. A. VAN DER GOOT, H. FIJTEN, J. W. VAN DER GIESSEN, E. KUIJT, C. B. M. MAASSEN, A. VAN ROON, B. WIT, A. P. KOETS, H. I. J. ROEST (2018): A cross sectional study on Dutch layer farms to investigate the prevalence and potential risk factors for different Chlamydia species. PLoS One. 13 (1), e0190774. doi: 10.1371/jour-nal.pone.0190774.
HULIN, V., S. OGER, F. VORIMORE, R. AAZIZ, B. DE BARBEYRAC, J. BERRUCHON, K. SACHSE, K. LAROUCAU (2015): Host preference and zoonotic potential of Chlamydia psittaci and Chlamydia gallinacea in poultry. Pathog. Dis., 73 (1), 1-11. doi: 10.1093/femspd/ftv005.
JELOCNIK, M., C. JENKINS, B. O'ROURKE, J. BARNWELL, A. POLKINGHORNE (2018): Molecular evidence to suggest pigeon-type Chlamydia psittaci in association with an equine
foal loss. Transbound. Emerg. Dis. 65, 911-915. doi: 10.1111/tbed.12817.
JENKINS, C., M. JELOCNIK, M.L. MICALLEF, F. GALEA, A. TAYLOR-BROWN, D. R. BOGEMA, M. LIU, B. O'ROURKE, C. CHICKEN, J. CARRICK, A. POLKING-HORNE (2018): An epizootic of Chlamydia psittaci equine reproductive loss associated with suspected spillover from native Australian parrots. Emerg. Microbes Infect., 16, 88. doi: 10.1038/s41426-018-0089-y.
KAUFFOLD, J., F. MELZER, A. BERNDT, G. HOFFMANN, H. HOTZEL, K. SACHSE (2006): Chlamydiae in oviducts and uteri of repeat breeder pigs. Theriogen., 66, 1816-1823.
KEMMERLING, K., U. MULLER, M. MIELENZ, H. SAUER-WEIN (2009): Chlamydophila species in dairy farms: polymerase chain reaction prevalence, disease association, and risk factors identified in a cross-sectional study in western Germany. J. Dairy Sci., 92, 4347-4354.
KNITTLER, M. R., K. SACHSE (2015): Chlamydia psittaci: update on an underestimated zoonotic agent, Pathog Dis., 73, 1–15.
KRAUTWALD-JUNGHANNS, M. E., J. STOLZE, V. SCHMIDT, J. BÖHME, K. SACHSE, K. CRAMER (2013): Efficacy of doxycycline for treatment of chlamydiosis in flocks of racing and fancy pigeons. Tierarztliche Praxis. Ausgabe K, Kleintiere/Heimtiere 41, 392-398.
LAROUCAU, K., B. DE BARBEYRAC, F. VORIMORE, M. CLERC, C. BERTIN, T. HARKINEZHAD, K. VERMINNEN, F. OBENICHE, I. CAPEK, C. BÉBÉAR, B. DURAND, G. ZANELLA, D. VANROMPAY, B. GARIN-BASTUJI, K. SACHSE (2009a): Chlamydial infections in duck farms associated with human cases of psittacosis in France. Vet. Microbiol. 135, (1–2), 82–89. doi:10.1016/j.vetmic.2008.09.048.
LAROUCAU, K., F. VORIMORE, R. AAZIZ, A. BERNDT, E. SCHUBERT, K. SACHSE (2009b): Isolation of a new chlamydial agent from infected domestic poultry coincided with cases of atypical pneumonia among slaughterhouse workers in France. Infect. Genet. Evol. 9, 1240-1247. doi: 10.1016/j.meegid.2009.08.005.
LAROUCAU, K., R. AAZIZ, L. MEURICE, V. SERVAS, I. CHOSSAT, H. ROYER, B. DE BARBEYRAC, V. VAILLANT, J. L. MOYEN, F. MEZIANI, K. SACHSE, P. ROLLAND (2014): Outbreak of psittacosis in a group of women exposed to Chlamydia psittaci-infected chickens. Euro. Surveill., 20, 21155.
LENZKO, H., U. H. MOOG, K. HENNING, R. LEDERBACH, R. DILLER, C. MENGE, K. SACHSE, L. D. SPRAGUE (2011): High frequency of chlamydial co-infections in clinically healthy sheep flocks. BMC Vet. Res., 7, 29.
LI, L., M. LUTHER, K. MACKLIN, D. PUGH, J. LI, J. ZHANG, J. ROBERTS, B. KALTENBOECK, C. WANG (2017): Chlamydia gallinacea: a widespread emerging Chlamydia agent with zoonotic potential in backyard poultry. Epidemiol. Infect. 145, 2701-2703. doi: 10.1017/S0950268817001650.
LUBLIN, A., G. SHUDARI, S. MECHANI, Y. WEISMAN (1996): Egg transmission of Chlamydia psittaci in turkeys. Vet. Rec., 139, 300.
MATSUMOTO, A., G. P. MANIRE (1970): Electron microscopic observations on the effects of penicillin on the morphology of Chlamydia psittaci. J. Bacteriol., 101, 278–285.

ZNAČENJE BAKTERIJA RODA CHLAMYDIA SP. U JATIMA PERADI I NJIHOV ZOONOTSKI POTENCIJAL 42
NEWMAN, C. P., S. R. PALMER, F. D. KIRBY, E. O. CAUL (1992): A prolonged outbreak of ornithosis in duck processors. Epidemiol. Infect., 108, 203–210.
PANTCHEV, A., R. STING, R. BAUERFEIND, J. TYCZKA, K. SACHSE (2009): New real-time PCR tests for species-specific detection of Chlamydophila psittaci and Chlamydophila abortus from tissue samples.Vet. J., 181, 145-150. doi: 10.1016/j.tvjl.2008.02.025.
PELLE-DUPORTE, D., I. GENDRE (2001): Ornithosis epidemics in apoultry slaughterhouse. INRS, Documents for the occupational physician, 85, 49–57.
PETROVAY, F., E. BALLA (2008): Two fatal cases of psittacosis caused by Chlamydophilla psittaci. J. Med. Microbiol., 57, 1296-1298.
POLKINGHORNE, A., G. GREUB (2017): A new equine and zoonotic threat emerges from an old avian pathogen, Chlamydia psittaci. Clin. Microbiol. Infect. 23, 693-694. doi: 10.1016/j.cmi.2017.05.025.
PRUKNER-RADOVČIĆ, E., Z. BIĐIN, D. HORVATEK (2006a): Prevalence of C. psittaci in turkey parent flocks in Croatia: Detection of antigen and antibodies. 6th International Symposium on Turkey Diseases. 11-13.05.2006., Berlin, Njemačka, 77.
PRUKNER-RADOVČIĆ, E., D. HORVATEK, Ž. MIŠKOVIĆ, H. MAZIJA (2006b): Chlamydophilosis in laying hens: a case report. Annual Workshop of COST Action 855. 03.-05.09.2006., Edinburgh, Velika Britanija, 82-83.
PRUKNER-RADOVČIĆ, E., D. HORVATEK TOMIĆ, K. LAROUCAU (2012): Detection of a new Chlamydiaceae strain from turkeys in Croatia. 9th International Symposium on Turkey Diseases. 21-23.06.2012., Berlin, Njemačka, 84-87.
QUILICOT, A. M. M., D. HORVATEK TOMIĆ, Ž. GOTTSTEIN, M. LUKAČ, E. PRUKNER-RADOVČIĆ (2017a): Com-parison of sampling procedures for the detection of Chlamydiaceae in free-range chicken flocks. Veterinary Science and Profession. 05.-07.10.2017. Zagreb, Hrvatska, 58.
QUILICOT, A. M. M., Ž. GOTTSTEIN, D. HORVATEK TOMIĆ, E. PRUKNER-RADOVČIĆ (2017b): Progress in Chlamydia psittaci vaccine development in poultry. WPS Journal, 73, 3, 505-514.
QUILICOT, A. M., E. PRUKNER-RADOVČIĆ (2018): Chla-mydia gallinacea in chickens. Book of Abstracts VI Mediter-ranean Poultry Summit., 18.-20. 06. 2018., Torino, Italija, 96.
QUILICOT, A. M., D. HORVATEK TOMIĆ, Ž. GOTTSTEIN, E. PRUKNER-RADOVČIĆ (2018): Chlamydia psittaci in free-range chicken flocks in Croatia. 16. Deutscher Chlamydien Workshop. 21-23.03.2018., Dusseldorf, Njemačka.
SACHSE, K., H. HOTZEL, P. SLICKERS, T. ELLINGER, R. EHRICHT (2005): DNA microarray-based detection and identification of Chlamydia and Chlamydophila spp. Mol. Cell Probes., 19, 41-50.
SACHSE, K., K. LAROUCAU, H. HOTZEL, E. SCHUBERT, R. EHRICHT, P. SLICKERS (2008): Genotyping of Chlamy-dophila psittaci using a new DNA microarray assay based on sequence analysis of ompA genes. BMC Microbiol., 8, 63.
SACHSE, K., K. LAROUCAU, K. RIEGE, S. WEHNER, M. DILCHER, H. H. CREASY, M. WEIDMANN, G. MYERS, F. VORIMORE, N. VICARI, S. MAGNINO, E. LIEBLER-TENORIO, A. RUETTGER, P. M. BAVOIL, F. T. HUFERT, R. ROSSELLÓ-MÓRA, M. MARZ (2014): Evidence for the existence of two new members of the family Chlamydiaceae and proposal of Chlamydia avium sp. nov. and Chlamydia
gallinacea sp. nov. Syst. Appl. Microbiol. 37, 79-88. doi: 10.1016/j.syapm.2013.12.004.
SACHSE, K., K. LAROUCAU (2015): Two more species of Chlamydia - does it make a difference? Pathog Dis., 73, 1–3.
SACHSE, K., K. S. RAHMAN, C. SCHNEE, E. MÜLLER, M. PEISKER, T. SCHUMACHER, E. SCHUBERT, A. RUETTGER, B. KALTENBOECK, R. EHRICHT (2018): A novel synthetic peptide microarray assay detects Chlamydia species-specific antibodies in animal and human sera. Sci., Rep. 16, 4701. doi: 10.1038/s41598-018-23118-7.
SHIVAPRASAD, H. L. (2015): Avian chlamydiosis. In: Manual of Poultry Diseases (J. Brugere-Picoux, J.-P. Vaillancourt, Eds.), Toppan Printing Leefung, 273-278.
SZYMAŃSKA-CZERWIŃSKA, M., A. MITURA, K. ZARĘBA, C. SCHNEE, A. KONCICKI, K. NIEMCZUK (2017): Poultry in Poland as Chlamydiaceae Carrier. J. Vet. Res. 61, 411-419. doi: 10.1515/jvetres-2017-0072.
VAN LOOCK, M., T. GEENS, L. DE SMIT, H. NAUWYNCK, P. VAN EMPEL, C. NAYLOR C, H. M. HAFEZ, B. M. GODDEERIS, D. VANROMPAY (2005): Key role of Chlamydophila psittaci on Belgian turkey farms in association with other respiratory pathogens. Vet. Microbiol., 107, 91–101. doi: 10.1016/j.vetmic.2005.01.009.
VAN LOOCK, M. V., K. LOOTS, S. V. ZANDE, M. V. HEERDEN, H. NAUWYNCK, B. M. GODDEERIS, D. VANROMPAY (2006): Pathogenic interactions between Chlamydophila psittaci and avian pneumovirus infections in turkeys. Vet. Microbiol., 112, 53–63.
VANROMPAY, D., P. BUTAYE, C. SAYADA, R. DUCA-TELLE, F. HAESEBROUCK (1997a): Characterization of avian Chlamydia psittaci strains using omp1 restriction mapping and serovar-specific monoclonal antibodies. Res. Microbiol., 148, 327-333.
VANROMPAY, D., P. BUTAYE, A. VAN NEROM, R. DUCATELLE, F. HAESEBROUCK (1997b): The prevalence of Chlamydia psittaci infections in Belgian commercial turkey poults. Vet. Microbiol., 54, 85–93.
VANROMPAY, D., T. HARKINEZHAD, M. VAN DE WALLE,
D. BEECKMAN, C. VAN DROOGENBROECK, K. VERMINNEN, R. LETEN, A. MARTEL, K. CAUWERTS (2007): Chlamydophila psittaci Transmission from Pet Birds to Humans. Emerg. Infect Dis., 13, 1108–1110.
VANROMPAY, D. (2013): Avian Chlamydiosis. In: Diseases of Poultry 13th edition. (D. Swayne, Ed.). Ames: Wiley- Blackwell Publishing, 1055-1073.
TIONG, A., T. VU, M. COUNAHAN, J. LEYDON, G.TALLIS, S. LAMBERT (2007): Multiple sites of exposure in an outbreak of ornithosis in workers at a poultry abattoir and farm. Epidemiol. Infect., 135, 1184–1191.
YIN, L., I. D. KALMAR, S. LAGAE, S. VANDENDRIESSCHE, W. VANDERHAEGHEN, P. BUTAYE, E. COX, D. VANROMPAY (2013): Emerging Chlamydia psittaci in-fections in the chicken industry and pathology of Chlamydia psittaci genotype B and D strains in specific pathogen free chickens. Vet. Microbiol., 162, 740–749.
YIN, L., I. KALMAR, J. BODEN, D. VANROMPAY (2015): Chlamydia psittaci infections in Chinese poultry: A literature review. WPS Journal 71, 473-482. doi:10.1017/ /S0043933915002226
ZHANG, F., S. LI, J. YANG, W. PANG, L. YANG, C. HE (2008): Isolation and characterization of Chlamydophila psittaci

ZNAČENJE BAKTERIJA RODA CHLAMYDIA SP. U JATIMA PERADI I NJIHOV ZOONOTSKI POTENCIJAL 43
isolated from laying hens with cystic oviducts. Avian Dis., 52, 74–78.
ZHOU, J., C. QIU, G. LIN, X. CAO, F. ZHENG (2010): Isolation of Chlamydophila psittaci from laying hens in China.Vet. Res., 3, 43–45.
ZOCEVIC, A., F. VORIMORE, C. MARHOLD, D. HORVATEK, D. WANG, B. SLAVEC, Z. PRENTZA, G. STAVIANIS, E.
PRUKNER-RADOVČIĆ, A. DOVČ, V. I. SIARKOU, K. LAROUCAU (2012): Molecular characterization of atypical Chlamydia and evidence of their dissemination in different European and Asian chicken flocks by specific real-time PCR. Environ. Microbiol., 14, 2212–2222. doi:10.1111/j.1462-2920.2012.02800.x
SIGNIFICANCE OF CHLAMYDIA SP. IN POULTRY FLOCKS AND ITS ZOONOTIC POTENTIAL
Summary
Poultry chlamydiosis, an emerging zoonotic disease, mostly is caused by C. psittaci, but recently also with a newly described species, C. gallinacea. Both bacteria cause economic losses to poultry production, while C. psittaci also has a significant zoonotic potential. This paper presents the presence and prevalence of C. gallinacea all over the world and in Croatia, where it was detected in 46.3% of examined samples. Key words: Chlamydiaceae, C. psittaci, C. gallinacea, poultry, zoonosis

PERADARSKI DANI 2019. 44
POJAVNOST OŠTEĆENJA PRSNIH KOSTI KOKOŠI NESILICA U ALTERNATIVNOM UZGOJU I OBOGAĆENIM KAVEZIMA
Zlatko Janječić1, Zlata Kralik2, Dalibor Bedeković1
1 Sveučilište u Zagrebu, Agronomski fakultet, Zagreb, Hrvatska 2 Sveučilište Josipa Jurja Strossmayera u Osijeku, Fakultet agrobiotehničkih znanosti, Osijek, Hrvatska
Sažetak
Napuštanjem držanja kokoši nesilica u klasičnim kavezima te prijelazom na nove alternativne sustave držanja i obogaćene kaveze očekivalo se da će doći do znatnog poboljšanja dobrobiti nesilica. Omogućen im je veći prostor za boravak i kretanje unutar kaveza i samog objekta te im je u slobodnom sustavu osiguran izlaz u ispuste. No, ipak pojavilo se i nekoliko problema za koje još nema rješenja i tek se na njima započelo raditi. Jedan od tih problema je i oštećenje prsne kosti koje se javlja kod kokoši nesilica, a može biti iskazano devijacijom prsne kosti što je blaži oblik i lomovima prsne kosti koje je teži oblik i prouzrokuje životinji veliku bol i patnju. Cilj ovoga rada je prikazati dosadašnje spoznaje vezane uz pojavnost ovih poteškoća u držanju kokoši nesilica u alternativnim uzgojima i obogaćenim kavezima. Iz dosadašnjih je spoznaja vidljivo da su oštećenja prsnih kosti najviše izražena kod nesilica koje su držane u volijerama i podnom uzgoju, dok je znatno manje oštećenja zabilježeno u obogaćenim kavezima. Neka od mogućih rješenja ovoga problema usmjerena su prema promjenama u opremi za držanje nesilica, genetici odnosno stvaranju novih robusnijih hibrida te hranidbi s ciljem ojačavanja koštanog sustava. Ključne riječi: kokoši nesilice, oštećenja prsne kosti, alternativni uzgoj, obogaćeni kavezi Uvod Začetak moderne proizvodnje konzumnih jaja datira još u davnim dvadesetim i tridesetim godinama dvadesetog stoljeća kada su u SAD-u razvijeni prvi žičani baterijski kavezi za smještaj kokoši nesilica kako bi se kokoši zaštitile od grabežljivaca, bolesti i ozljeda. Takvi su kavezi pojedno-stavili uzgoj povećavajući učinkovitost kokoši nesilica i smanjujući potrebu za prostorom u kojem su kokoši držane (Leaders, 2012.). Sustavi za smještaj kokošima moraju osi-gurati prostor, hranu, vodu, svjetlo, kvalitetu zraka i sani-tarne uvjete koji promiču dobro zdravlje i dobrobit kokoši te sigurnost hrane. Krajem osamdesetih i devedesetih godina prošloga stoljeća, kada su međunarodni propisi počeli ogra-ničavati uporabu konvencionalnih kaveza, započela su razmišljanja o prijelazu na nove sustave smještaja kokoši nesilica koji su za cilj imali uravnotežiti potrebe životinja za povećanjem njihove dobrobiti te i dalje osigurati profitabil-nost proizvodnje konzumnih kokošjih jaja. Alternativni sustavi proizvodnje su slobodno držanje, polu-intenzivno držanje, držanje na dubokoj prostirci i etažno držanje u staji.
Oni su sa sustavom držanja kokoši nesilica u obogaćenim kavezima obvezni načini držanja kokoši nesilica u zemljama EU od 1. siječnja 2012. godine kada je stupila na snagu Direktiva 1999/74 / EC (Čižmak i Bišćan, 2017.). Te pro-mjene su se možda čak i najviše odrazile na proizvodnju konzumnih jaja u RH u odnosu na ostale proizvodnje jaja u EU. Ulaskom RH u članstvo EU 1. srpnja 2013. godine hrvatski proizvođači konzumnih jaja su nakon turbulentnih 1990.- ih gotovo preko noći stavljeni pred zid te su svoju dosadašnju proizvodnju u klasičnim baterijskim kavezima morali zamijeniti jednim od sustava dopuštenih gore nave-denom Direktivom. Neke proizvodnje su se tada naprasno ugasile, a ostali su krenuli s opremanjem i adaptacijom po-stojećih objekata, a drugi od temelja s novim objektima i opremom najčešće kreditnim zaduživanjem i jednim dijelom uz potporu EU sredstava iz različitih tada dostupnih fondova. Između ostaloga, to je rezultiralo i time da smo od nekadašnje samodostatne proizvodnje još i danas uvoznici gotovo 20% konzumnih kokošjih jaja (Croatiastočar, 2018.). Napuštanjem držanja kokoši nesilica u klasičnim kavezima te prijelazom na alternativne sustava držanja i obogaćene
Prof. dr. sc. Zlatko Janječić, Sveučilište u Zagrebu, Agronomski fakultet, Zavod za hranidbu životinja, Svetošimunska 25, 10 000 Zagreb, Republika Hrvatska; tel.: +385 (0)1 239 3951; fax: +385 (0)1 239 3932; e-mail: [email protected]

POJAVNOST OŠTEĆENJA PRSNIH KOSTI KOKOŠI NESILICA U ALTERNATIVNOM UZGOJU I OBOGAĆENIM KAVEZIMA 45
kaveze očekivalo se je da će doći do znatnog poboljšanja dobrobiti nesilica. Omogućen im je veći prostor za boravak i kretanje unutar kaveza i samog objekta te im je u slobodnom sustavu osiguran izlaz u ispuste. Međutim, sve se više ukazuje na probleme koji se u tim sustavima proizvodnje pojavljuju. Jedan od tih problema su i oštećenja prsnih kosti kod kokoši nesilica u vidu devijacija i lomova koja mogu biti bolna za kokoši te time ugrožavaju dobrobit i smanjuju produktivnost (Harlander i sur., 2015.). Stoga je cilj ovoga rada ukazati na pojavnost oštećenja prsnih kosti kokoši nesilica u alternativnom uzgoju i obo-gaćenim kavezima te prikazati neka od rješenja da tih pojava bude što manje.
Sustav držanja kokoši u obogaćenim kavezima
Obogaćeni kavezi moraju osigurati najmanje 750 cm2 povr-šine poda za svaku nesilicu, od toga 6002 cm iskoristivo. Visina kaveza na mjestima osim iskoristivog prostora mora biti najmanje 20 cm u svakoj točki. Minimalna površina kaveza mora iznositi 2.000 cm2. U kavezu kokošima treba osigurati gnijezdo za nesenja jaja odijeljeno visećim traka-ma, prostirku za kljucanje i «kupanje», prečku za sjedenje dužine najmanje 15 cm po kokoši. Kavezi moraju biti opremljeni priborom za skraćivanje/brušenje noktiju. Kon-strukcija baterija se izvodi kao samostojeća. Vrata kaveza su takvog oblika i veličine da se iz kaveza može izvaditi ili u njega staviti odrasla kokoš, a da joj se tim postupkom ne prouzroče patnje ili ozljede. Svi metalni dijelovi najčešće su galvanizirani. Stranice i podovi kaveza su od žičane mreže, a omogućavaju dobar pregled životinja i nesmetano provjetra-vanje. Kokoši se hrane iz žljebastih hranilica koje su smještene s vanjske strane svakog reda kaveza. Raspodjela hrane u hranidbene žljebove provodi se lancem. Za napa-janje kokoši koristi se automatski nipl sustav. Izmet iz svake etaže kaveza pada kroz žičani pod na horizontalnu poli-propilensku traku ispod svakog reda kaveza. Traka klizi na specijalnim nosačima i bočno je zaštićena pregradama koje onemogućavaju ispadanje gnoja u hranidbene žljebove. Na kraju svake etaže nalaze se noževi od inoksa za čišćenje polipropilenskih traka. Prolaz između dva reda kaveza mora biti razmaknut najmanje 90 cm, a najmanja visina od poda do prvog reda kaveza mora biti 35 cm.
Alternativni sustavi držanja kokoši nesilica
Kod slobodnog sistema držanja kokoši nesilice moraju imati neprekidan pristup otvorenom prostoru, zemljište na koje životinje imaju pristup mora biti uglavnom pokriveno vege-tacijom, najveća naseljenost ne smije biti veća od 1000 kokoši po hektaru zemljišta dostupnog kokošima odnosno po jednoj kokoši mora biti osigurano 10 m2, a unutrašnjost zgrade mora biti uređena tako da zadovolji uvjete speci-ficirane za držanje na dubokoj stelji ili etažnog sustava držanju u štali. Polu-intenzivni sistem držanja kokoši nesilica razlikuje se od slobodnog samo u broju nesilica po m2 zatravnjenog ispusta, odnosno najveća naseljenost ne smije biti veća od
4000 kokoši po hektaru zemljišta dostupnog kokošima, od-nosno po jednoj kokoši mora biti osigurano 2,5 m2. Vanjski dio ispusta koji služi za napasivanje kokošima u slobodnom i polu-intenzivnom uzgoju treba biti ograđen žičanom mrežom visine najmanje 1,5 m. Ograda služi za kontrolu proizvodnje te za sprječavanje neželjenog ulaza različitih predatora. Najveća naseljenost kokoši nesilica kod podnog sistema držanja na dubokoj prostirci ne smije biti veća od 7 kokoši po m2 iskoristivog podnog prostora, najmanje 1/3 podnog prostora mora biti pokrivena prostirkom (slama, drvena strugotina) i mora biti osiguran dovoljno velik prostor poda za sakupljanje izmeta. Ovisno o broju kokoši nesilica objekti za proizvodnju jaja s etažnim sistemom držanja u staji imaju određen broj volijera s gnijezdima koja mogu biti opremljena takozvanim istjeri-vačem peradi, određen broj linija hranjenja i pojenja te linija prečki za sjedenje. Kako bi se životinje podijelile u manje skupine čitav se objekt može podijeliti u više odjeljaka žičanim pregradama. Pregrade su visine zida i svaki hodnik između volijera ima vrata. Ovakvo formiranje manjih sku-pina zahtijeva i EU Direktiva 98/58/EC o zaštiti životinja na farmama kojom se dozvoljava najviše naseljeno 6000 nesi-lica po odjeljku. Podovi etaža se izrađuju od žičane rešetke. Ispod podnih rešetki, kao i ispod gnijezda nalaze se trake za izgnojavanje. Projektirane volijere trebaju imati najmanje 10 cm hranidbenog prostora po nesilici, 15 cm prečki za sjedenje po nesilici i ne više od 25 nesilica po m2. Prostora za čeprkanje u sistemu volijera kokoši imaju napretek, bu-dući da je cijela podna površina prekrivena prostirkom. Grupna gnijezda u proizvodnim objektima se smještaju na prvoj i drugoj razini i to naizmjenično, tako da se ispred ulaza u gnijezdo nalazi veći prostor s hranom i vodom na kojem se zadržava više životinja te im se skraćuje put do gnijezda kad osjete potrebu za nesenjem. Na svaki m2 gnijezda dolazi 120 nesilica.
Dosadašnje spoznaje o pojavnosti oštećenja prsnih kosti
Jedan od najvećih izazova s kojim se suočavaju proizvođači konzumnih kokošjih jaja je pojavnost oštećenja prsnih kosti. (FAWC, 2010.; 2013.). U posljednjem desetljeću brojne studije koje su uglavnom provođene u zemljama EU bilježe devijacije i lomove prsnih kosti koje se kreću od 5% pa do čak 97%, ovisno o sustavu držanja i dobi nesilica. Stvarni raspon broja zahvaćenih kokoši jako varira i vjerojatno ovisi o nizu čimbenika, uključujući između ostalog dob, okoliš uzgoja, hranidbu, genetsku liniju i sistem smještaja. Iako ptice s prijelomom ili bez njega mogu djelovati slično (osim ako koristite posebne metode promatranja), prijelomi se obično mogu lako identificirati palpacijom. Vjeruje se da prijelomi, pogotovo kada su svježi i u stanju u kojem kosti nisu imobilizirane već su u zasebnim komadima, kod kokoši uzrokuju bolove i smanjuju živahnost i proizvodnost. Čini se da su učinci na proizvodnost naročito veliki u kokoši starijih od 50 tjedana.

POJAVNOST OŠTEĆENJA PRSNIH KOSTI KOKOŠI NESILICA U ALTERNATIVNOM UZGOJU I OBOGAĆENIM KAVEZIMA 46
Tako su Rodenburg i sur. (2008.) proveli istraživanje koje je za cilj imalo usporediti dobrobit kokoši nesilica u oboga-ćenim kavezima i držanju na dubokoj prostirci. U dobi kokoši od 60 tjedana kokoši držane u obogaćenim kavezima imale su uz smanjenu smrtnost i znatno nižu učestalost prijeloma prsnih kosti. Cilj istraživanja koji su proveli Wilkins i sur. (2011.) bio je između ostalog procijeniti oštećenost prsnih kosti kod kokoši nesilica držanih u obogaćenim kavezima i volijerama. Na kraju proizvodnog razdoblja procijenjeno je da je u obogaćenim kavezima 36% kokoši nesilica imalo oštećenja prsnih kosti, dok je kod volijera taj broj iznosio više od 80%. Riber i Hinrichsen (2016.) su proveli istraživanja u Danskoj s ciljem procjene oštećenja prsne kosti u komercijalnim jatima kokoši nesilica držanih u različitim sustavima i pri različitoj dobi. Zabilježili su veća oštećenja prsnih kosti kod nesilica starih 62 tjedna u odnosu na one stare 32 tjedna. Kokoši u volijerama imale su veća oštećenja prsnih kosti u odnosu na one držane podnim načinom držanja; 11,6%:4,9%. Heerkens i sur. (2016.) su kod 47 jata kokoši nesilica starih 60 tjedana, a držanih u volijerama u Belgiji utvrdili 82,5% lomova i 58,9% devijacija prsnih kosti. Nasr i sur. (2013.) zaključuju da kokoši nesilice bez oštećenja prema onima s lomovima prsnih kosti nesu veći broj jaja (91,7% prema 84,9%), jaja su teža (61,9 g prema 60,2 g), jedu manje hrane (139 g prema 151 g) i piju manje vode (212 mL prema 237 mL). Kappeli i sur. (2011.) iznose da je pojavnost oštećenja prsnih kosti na švicarskim farmama iz-nosila i do 83% u višeetažnim sustavima uzgoja, volijerama. Struelens i Tuyttens (2009.) ukazuju na prečke za sjedenje kokoši koje propisuje Direktiva EU 1999/74, dizajn i izvedba kojih mogu utjecati na veću pojavnost oštećenja prsnih kosti. U istraživanju koje su proveli Đukić-Stojčić i sur. (2017.) je zaključeno da je pojavnost oštećenja prsne kosti kod obogaćenih kaveza bez prečki 4,3%, dok je kod obogaćenih kaveza s prečkama ona iznosila 39%. Slično su zaključili i Rodenburg i sur. (2008.), te Wilkins i sur. (2011.). Hester i sur. (2013.) su izvijestili da je većina pro-blema sa skeletnim sustavom povezana s osteoporozom koja je osobito naglašena kod držanja kokoši nesilica u obo-gaćenim kavezima, a pojačan udio oštećenja je zabilježen kod nesilica koje su u kavezima imale metalne prečke. Stratmann i sur. (2015.) zaključuju da mekane poliuretanske prečke značajno smanjuju pojavnost oštećenja prsnih kosti kod višeetažnih sustava držanja kokoši nesilica u odnosu na metalne prečke. Heerkens i sur. (2016.) su nakon prove-denog istraživanja zaključili da ugrađivanje rampi između prečaka značajno smanjuje oštećenja prsnih kosti te da treba razmišljati o modifikaciji dosadašnjih sustava višeetažnog držanja kokoši nesilica. Cilj istraživanja koje su proveli Rufener i sur. (2018.). bio je istražiti utjecaj lomova prsne kosti na produktivnost kokoši nesilica. Zaključili su da su kokoši s prijelomima u 61. tjednu nesivosti imale za 16,2% nižu nesivost nego kokoši bez prijeloma. Riber i sur. (2018.) u svom revijalnom radu iznose trenutne spoznaje o utjecaju oštećenja prsnih kosti na dobrobit kokoši nesilica. Zaključuju da su oštećenja prsnih kosti zabilježena u svim tipovima komercijalne proizvodnje konzumnih
kokošjih jaja, s različitim postotkom pojavnosti ovisno o sustavu, dobi i hibridu kokoši nesilica. Kokoši nesilice sa slomljenim prsnim kostima pokazuje razlike u ponašanju koje je vidljivo u korištenju gnijezda, prpošenju i smanjenoj mobilnosti. Strogo ukazuju na činjenicu da je lom prsne kosti izvor boli i tijekom više tjedana nakon što je do njega došlo. Negativan utjecaj lomova prsne kosti vidljiv je u smanjenoj proizvodnji jaja. Zbog manjeg broja provedenih istraživanja utjecaj devijacija prsne kosti na kondiciju kokoši nesilica još nije dovoljno jasan. No, nađeno je da devijacija prsne kosti svakako ima negativan utjecaj na dobrobit kokoši nesilica. Preporučuju da je potrebno provesti daljnja istraživanja koja će se usredotočiti na utjecaj genetskih linija i genetske selekcije, držanja i hranidbe na razvoj, pojavnost oštećenja prsnih kosti kod kokoši nesilica.
Intervencije za smanjenje pojavnosti oštećenja prsnih kosti
Iz dosadašnjih spoznaja izrađen je letak za držaoce kokoši nesilica kojim se želi ukazati na mogućnosti smanjenja po-javnosti oštećenja prsnih kosti (CA 15224, 2018.). Upute su fokusirane na aktivnosti tijekom uzgoja pilenki, razdoblje nesenja jaja, a najveći je naglasak dan hranidbi. Poznato je da pilenke kontinuirano razvijaju svoje koštane i kognitivne mehanizme potrebne za svoj daljnji život, tako da je raz-doblje uzgoja vrlo značajno za osiguranje jakih kosti kod kokoši nesilica kako bi se mogle samostalno kretati kroz sisteme proizvodnje bez ozljeda. Kada je moguće, okoliš uzgoja i nesenja jaja treba biti što sličniji, osobito kod kaveznog i ne kaveznog sistema proizvodnje jaja. Tijekom uzgoja dodatnu opremu treba uvoditi postupno tako da se kokoši mogu naučiti kako ih koristiti na odgovarajući način, što dovodi do preciznijih pokreta. Generalno je poznato da povećana aktivnost vodi ka boljem zdravlju kosti. Proiz-vođači bi također trebali razmotriti specifične intervencije uključujući raniji pristup prečkama i/ili rampama. Poznato je da kokoši nisu dobri letači te da su bolje u hodanju i izvođenju kratkih skokova kako bi se okomito kretale. Iz tog razloga kokošima treba osigurati alternativne načine za oko-mito kretanje pomoću rampi ili prečki postavljenih blizu jedne drugima, no opet da se ograniči nakupljanje izmeta ili spavanje (na rampama). Tijekom nesenja kokoši se sudaraju s opremom u njihovom okruženju, što dovodi do prijeloma. Sudaranja se događaju u kaveznim i ne kaveznim sustavima gdje uzroci mogu biti panika ili promet visoke gustoće, kao što je u sumrak dok se ptice kreću prema gornjoj etaži. Također se može sumnjati da su uzroci u ubrzanom mahanju krilima tijekom panike ili dok se ptice pokušavaju vratiti u svoj prvobitni položaj nakon pada. Stoga bi proizvođači trebali pomoći pri tim pokretima i spriječiti paniku. Neke specifične preporuke za smanjenje panike uključuju postup-no kretanje uzgajivača kroz svoje nastambe u odjeći različite obojanosti, iz različitih smjerova i u različito doba dana. U početku bi se proizvođači trebali polako kretati, a nakon što se kokoši priviknu započeti s bržim i redovitijim kretanjem. Idealno, to bi trebalo započeti što je ranije moguće, uključujući i razdoblje tijekom uzgoja i/ili unutar prvog

POJAVNOST OŠTEĆENJA PRSNIH KOSTI KOKOŠI NESILICA U ALTERNATIVNOM UZGOJU I OBOGAĆENIM KAVEZIMA 47
tjedna nakon useljenja. Radio aparati ili neki drugi izvori buke također mogu biti od pomoći, poglavito ako se koriste od početka još u uzgojnim objektima. Za pomoć pri kretanju (više se odnosi na ne kavezni sustav) od početka uzgoja potrebno je da prečke budu postavljene tako da pomažu kretanje u okomitom smjeru. Generalno, prečke trebaju biti postavljene jedna od druge pod kutom manjim od 450 i s razmacima manjim od 50 cm okomito i 75 cm vodoravno. Pokazalo se da rampe povećavaju broj kontroliranog kre-tanja kokoši, što dovodi do smanjenja sudaranja tijekom ključnog razdoblja, sumraka. Pojavnost prijeloma prsnih ko-sti bila je manja za 9% u komercijalnim volijerama s ram-pama (64%) u odnosu na volijere bez rampi (73%), a tako-đer je utvrđeno je da su veće koristi od rampi u smanjenju prijeloma ako ih se uključi i u uzgoju i u razdoblju nesenja. Hranidba je vrlo zahtjevna glede davanja preporuka s obzi-rom na to da su se potrebe hibridnih nesilica tijekom vreme-na značajno promijenile, kao što su i proizvodnja jaja i duljina ciklusa nesenja dramatično porasle. Uz to, različiti sustavi držanja poput slobodnog držanja imaju veće zahtjeve za hranjivim tvarima, dok će varijacije u izloženosti sunče-voj svjetlosti, sadržaju kalcija u vodi i drugi okolišni čimbenici imati utjecaj na specifične hranidbene zahtjeve. S obzirom na mnogobrojne čimbenike konačni obrok treba biti sastavljen u suradnji s nutricionističkim stručnjakom. Opće preporuke temeljene na provedenim istraživanjima ukazuju na to da tijekom uzgoja pilenke trebaju dobivati minimalno 9 g/kg između 0. i 14. tjedna starosti i tada slijedi povećanje do 35 g/kg kalcija u obroku, a dodatak vitamina K (12 mg/kg) dovodi do povećanja volumena kosti, što koristi boljem skladištenju kalcija. Ističe se da tijekom nesenja krmna smjesa s 0,11% do 0,26% nefitinskog fosfora dopu-njena fitazom može poslužiti boljem zadržavanju kalcija. Pokazalo se da primjena linolenske kiseline (npr. iz lanenog sjemena) od 23. do 30. tjedna starosti poboljšava čvrstoću kosti i rezultira manjom stopom prijeloma, iako proizvođači trebaju biti oprezni zbog učinaka na kvalitetu jaja. Premda je malo vjerojatno da će dnevno povećanje kalcija u obroku rezultirati time da kokoši apsorbiraju više kalcija, dokazano je da bi davanje većih čestica kalcija u razdoblju od 2 do 3 sata neposredno prije sutona dovelo do toga da kokoš apsorbira više kalcija.
Zaključak
Lomovi i devijacije prsne kosti smanjuju dobrobit kokoši nesilica, jer kod njih izazivaju bol, a primjetni su i slabiji proizvodni pokazatelji. Iz dosadašnjih je spoznaja vidljivo da su oštećenja prsnih kosti zabilježena u svim tipovima ko-mercijalne proizvodnje konzumnih kokošjih jaja, s različitim postotkom pojavnosti ovisno o sustavu, dobi i hibridu kokoši nesilica. Isto tako se može zaključiti da su lomovi i devijacije prsnih kosti izraženije kod ne kaveznih sustava držanja (volijere i podni uzgoj), dok je znatno manje oštećenja zabilježeno u obogaćenim kavezima. Neka od mo-gućih rješenja ovoga problema usmjerena su prema promje-nama u opremi za držanje nesilica, genetici odnosno stva-ranju novih robusnijih hibrida te hranidbi s ciljem ojačava-nja koštanog sustava.
Zahvala
Ovaj je rad izrađen sudjelovanjem u COST akciji Identifyng Causes and Solutions of Keel Bone Damage in Laying Hens, CA15224.
Literatura
BOBETIĆ, B. (2018): Financijski rezultati poslovanja u 2017. godini te proizvodni pokazatelji sektora proizvodnje mesa peradi i proizvoda od mesa peradi te konzumnih jaja u periodu 2012. do 2017. godine. GIU Croatiastočar, Zagreb.
CA 15224 (2018): Interventions to reduce keel bone fractures. Leaflet, http://www.keelbonedamage.eu
ČIŽMAK, VALERIJA, TIHANA BIŠĆAN (2017): Držanje kokoši nesilica prema standardima EU. https://www.savjetodavna.hr/ /adminmax/publikacije/nesiliceEU_762017.pdf
ĐUKIĆ STOJČIĆ, MIRJANA, LIDIJA PERIĆ, RENATA RELIĆ, IVANA BOŽIČKOVIĆ, VESNA RODIĆ, VIDA REZAR (2017): Oštećenje grudne kosti kod kokoši nosilja gajenih u različitim sistemima držanja u Srbiji. Biotechnology in Animal Husbandry, 33 (4): 487-492.
FAWC (2010): Opinion on Osteoporosis and Bone Fractures in Laying Hens. Farm Animal Welfare Council. London, UK.
FAWC (2013): Keel bone fractures in laying hens. Farm Animal Welfare Council London, UK.
HARLANDER-MATAUSCHEK, A., T. B. RODENBURG, V. SANDILANDS, B. W. TOBALSKE, M. J. TOSCANO (2015): Causes of keel bone damage and their solutions in laying hens. World's Poultry Science Journal, 71: 461–472.
HEERKENS, J. L., E. DELEZIE, T. B. RODENBURG, I. KEMPEN, J. ZOONS, B. AMPE, F. A. TUYTTENS (2016): Risk factors associated with keel bone and foot pad disorders in laying hens housed in aviary systems. Poult Sci., 95 (3): 482-488.
HESTER, P. Y., S. A. ENNEKING, B. K. HALEY, H. W. CHENG, M. E. EINSTEIN, D. A. RUBIN (2013): The effect of perch availability during pullet rearing and egg laying on musculoskeletal health of caged White Leghorn hens. Poultry Science, 92: 1972-1980.
KÄPPELI, S., S. G. GEBHARDT-HENRICH, E. FRÖHLICH, A. PFULG, M. H. STOFFEL (2011): Prevalence of keel bone deformities in Swiss laying hens. Br Pou. Sci., 52 (5):531-536.
LEADERS, M. A. (2012): Welfare Implications of Laying Hen Housing. https://www.avma.org/KB/Resources/LiteratureReviews/Documents/laying_hen_housing_bgnd.pdf
NASR, M. A. F., J. MURRELL, L. J. WILKINS, C. J. NICOL (2012): The effect of keel fractures on egg-production parameters, mobility and behaviour in individual laying hens. Animal Welfare, 21 (1): 127-135.
STRUELENS, E., F. A. M. TUYTTENS (2009): Effects of perch design on behaviour and health of laying hens. Animal Welfare, 18 (4): 533-538.
RIBER, A. B., L. K. HINRICHSEN (2016): Keel-bone damage and foot injuries in commercial laying hens in Denmark. Animal Welfare, 25: 179-184.
RIBER, A. B., T. M. CASEY-TROTT, M. S. HERSKIN (2018): The Influence of Keel Bone Damage on Welfare of Laying Hens. Front. Vet. Sci., 28; 5:6.

POJAVNOST OŠTEĆENJA PRSNIH KOSTI KOKOŠI NESILICA U ALTERNATIVNOM UZGOJU I OBOGAĆENIM KAVEZIMA 48
RUFENER, C., S. BAUR, A. STRATMANN, M. J. TOSCANO (2018): Keel bone fractures affect egg laying performance but not egg quality in laying hens housed in a commercial aviary system. Poultry Science; https://doi.org/10.3382/ps/pey544
RODENBURG, T. B., F. A. M. TUYTTENS, K. DE REU, L. HERMAN, J. ZOONS, B. SONCK (2008): Welfare asses-sment of laying hens in furnished cages and non-cage systems: An on-farm comparison. Animal Welfare, 17: 363–373.
STRATMANN, A., E. K. FRÖHLICH, A. HARLANDER-MATAUSCHEK, L. SCHRADER, M. J. TOSCANO, H. WÜRBEL, S. G. GEBHARDT-HENRICH (2015): Soft
perches in an aviary system reduce incidence of keel bone damage in laying hens. PLoS One., 10 (3): e0122568.
STRUELENS, E., F. A. M. TUYTTENS (2009): Effects of perch design on behaviour and health of laying hens. Animal Welfare, 18: 533-538.
WILKINS, L. J., J. L. MCKINSTRY, N. AVERY, T. G. KNOWLES, S. N. BROWN, J. TARLTON, C. NICOL (2011): Influence of housing system and design on bone strength and keel bone fractures in laying hens. Veterinary Record,169: 414.
THE PREVELENCE OF KEEL BONE DAMAGE IN LAYING HENS IN ALTERNATIVE AND ENRICHED CAGES REARING
Summary
By leaving the rearing of laying hens in classic cages and moving to new alternative systems and enriched cage rearing, it was expected that there would be a significant improvement in the welfare of laying hens. They have a larger space for staying and moving inside the cage and in the free range system they have exit to open area. However, there have been several problems that still have no solution and are yet to be managed. One of these problems is damage to the keel bones that occurs in laying hens and can manifest by a mild form of bone deviation or fractures of the keel bone as a severe form, causing great pain and suffering in laying hens. The aim of this paper is to present the knowledge about the prevalence of these difficulties in keeping laying hens in alternative rearing and enriched cages. From the results so far, it is seen that damage to the bones is most pronounced in laying hens held in barn and floor rearing, whereas a significantly less damage is recorded in enriched cages. Some of the possible solutions to this problem are geared towards changing equipment for keeping laying hens, genetics, or creating new, more robust hybrids, along with nutrition aimed at strengthening their bone system. Key words: laying hens, keel bone damage, alternative rearing, enriched cages

PERADARSKI DANI 2019. 49
FIELD EXPERIENCES CONCERNING THE LONG TERM USE OF MS-H AND MS LIVE VACCINES ON PARENT STOCK FARM IN HUNGARY
László Kőrösi1, János Povazsán1, Attila Jacsó1, Chris Morrow2, Miklós Gyuranecz3
1 Rhone Vet Kft., Budapest, Hungary 2 Bioproperties Pty Ltd., Australia
3 Institute for Veterinary Medical Research, Centre for Agricultural Research, Hungarian Academy of Sciences, Budapest, Hungary
Summary
Mycoplasma synoviae (MS) infection in parent stock (PS) flocks significantly impairs economic parameters, can reduce egg production, predisposes to Escherichia coli peritonitis, and causes egg apical abnormality decreasing the hatchability. Vertically infected offspring can have reduced performance. Two consecutive broiler PS flocks of a multi-age farm of the company were vaccinated with live MS vaccines. One flock was immunized with MS-H vaccine by eye drop and the other flock was vaccinated with MS Live by spray. According to the study results, the multi-age farm of the company was infected with wild MS. Both used live MS vaccines able to protect the offspring. Detectability of the MS-H strain was longer, stronger in the flock vaccinated by MS-H, and even superseded the MS1 strain of the MS Live vaccinated flock in the older age of the flock.
Key words: Mycoplasma synoviae (MS), live mycoplasma vaccines, parent stock, broiler
Introduction
Mycoplasma synoviae (MS) eradication is not easy, particularly in the multi-age parent stock (PS) farms, but flocks vaccinated with MS live vaccine could provide higher profitability of the farms (Kőrösi et al., 2015). The company has a history of MS infection confirmed by institute examinations and some of their flocks had high mortality due to peritonitis (Raviv et al., 2007). The multi-age production farm is very close to two broiler houses (about 200 m), so they are using live MS vaccines. In Hungary, two live MS vaccines are authorized, one thermosensitive (MS-H) and one attenuated, freeze dried (MS Live; MS1). Materials and methods Studied farms and flocks A company located in north-east Hungary that produces and sells day old (DO) broilers was studied. The company has one single-age rearing farm and one multi-age production farm as well. The company hatchery is very close to the production farm. The rearing farm has 2 houses which are located around 5 km from the production farm. The production farm has 3 units: unit 1 has one house with four pens, unit 2 has two houses, and unit 3 has two houses.
Due to the limited rearing capacity, they are moving the reared pullets to the production farm at about 14 weeks of age.
Vaccination, treatment and sampling Two consecutive broiler PS flocks were vaccinated and compared. One flock (Flock A) was immunized with MS-H vaccine by eye drop at 5 weeks (DO placement on 09.02.2016, vaccination on 16.03.2016). At 14 weeks of age (18-19.05.2016) the flock was moved into the production farm. The other flock (Flock B) was vaccinated with MS Live by spray at 7 weeks of age (DO placement on 07.06.2016, vaccination on 26.07.2016) and moved into the production farm at 14 weeks of age (14-15.09.2016). Flock A houses are about 50 m far from the Flock B house. The company uses preventive treatment in each PS flock at 15 weeks (after moving) and at 21 weeks. The treatment at 15 weeks includes amoxiclav for 4 days and at 21 weeks enrofloxacin for 4 days. Their broiler farmers treat DO chickens with enrofloxacin for 3 days and at around 2-3 weeks of age with amoxicillin for 4-5 days. We tested the PS flocks in their laying cycle at 34-40-45-55-60 weeks of age, and then we tested their progeny from different hatches at the hatch and at the killing age. The sampling protocols are summarized in Table 1.
László Kőrösi +36309820054, [email protected]

FIELD EXPERIENCES CONCERNING THE LONG TERM USE OF MS-H AND MS LIVE VACCINES ON PARENT STOCK FARM IN HUNGARY
50
Table 1. Sampling protocol of vaccinated flocks
Sampling (date), age (wks)
Flock A
Sampling Sampling 1 Sampling 2 Sampling 3 Sampling 4 Sampling 5
Age, date 34 weeks 03.10.2016
40 weeks 14.11.2016
45 weeks 19.12.2016
55 weeks 25.02.2017
60 weeks 31.03.2017
Hatch (DO) 24.10.2016 16.12.2016 26.01.2017 21.03.2017 19.04.2017
Killing 11.29.2016 26.01.2017 25.02.2017 03.05.2017 22.05.2017
Flock B
Age, date 34 weeks 26.01.2017
40 weeks 11.03.2017
45 weeks 19.04.2017
55 weeks 24.06.2017
60 weeks 01.08.2017
Hatch (DO) 24.02.2017 31.03.2017 15.05.2017 17.07.2017 22.08.2017
Killing 31.03.2017 03.05.2017 24.06.2017 22.08.2017 26.09.2017 Laboratory testing The aim of the examination of Flock A was to test tracheal swabs for the presence of MS-H vaccine strain and wild-type MS strains with molecular biology methods. The aim of the examination of Flock B was to test tracheal swabs for the presence of MS1 vaccine strain, MS-H vaccine strain and wild-type MS strains with molecular biology methods. Twenty-five samples were taken at each sampling time and 5 tracheal swabs were pooled. DNA extractions from each pool were performed with the Qiagen DNA Mini kit (Qiagen Inc., Valencia, CA, USA) in accordance with the manufacturer's instructions. A PCR including melt-MAMA assay targeting the point mutation of the obg gene of MS was used to detect and differentiate the MS-H vaccine strain, MS Live vaccine strain and wild-type MS (Kreizinger et al., 2015; Kreizinger et al., 2017). All testings throughout the
trial were performed in the Institute for Veterinary Medical Research, Centre for Agricultural Research, Hungarian Academy of Sciences, Hungary. Results Flock A The MS-H strain persisted in the flock throughout the study period, however, wild strain appeared at older age. MS1 strain was not detectable in the flock (Table 2). Flock B The MS1 strain was not detectable at 34 weeks of age, but was demonstrated at 40 and 45 weeks of age. Later, the MS-H strain superseded MS1 strain. Wild strain also appeared also at older age (Table 3).
Table 2. PCR test results in Flock A (MS-H vaccinated flock)
PS age 34 wks 40 wks 45 wks 55 wks 60 wks
Pools* 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5
PS Date 03.10.2016 14.11.2016 19.12.2016 24.02.2017 31.03.2017
Result V V V V V V V V V V V V V V V V V V V W
V V V V V V W
Hatch Date 24.10.2016 16.122016 26.01.2017 21.03.2017 19.04.2017
Result N N N N N N N N N N N N N N N N N N N N N N N N N
Brojler Date 29.11.2016 26.01.2017 25.02.2017 03.05.2017 22.05.2017
Result N N N N N N N N N N N N N N N N N N N N N N N N N *pool = 5 trachea/choana swabs V = MS-H vaccine W = wild type MS N = negative

FIELD EXPERIENCES CONCERNING THE LONG TERM USE OF MS-H AND MS LIVE VACCINES ON PARENT STOCK FARM IN HUNGARY
51
Table 3. PCR test results in Flock B (vaccinated MS Live vaccine)
PS age 34 wks 40 wks 45 wks 55 wks 60 wks Pools* 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 PS Date 26.01.2017 11.03.2017 19.04.2017 24.06.2017 01.08.2017
Result N N N N N V2
V2
N V2
V2
V2
N V2
V2
V2
V
V2 V
V2 V
V
V V W
V V V V W
Hatch Date 24.02.2017 31.03.2017 15.05.2017 17.07.2017 22.08.2017 Result N N N N N N N N N N N N N N N N N N N N N N N N N
Broiler Date 31.03.2017 03.05.2017 24.06.2017 22.08.2017 26.09.2017 Result N N N N N N N N N N N N N N N N N N N N N N N N N
*pool = 5 trachea/choana swabs V2 = MS Live vaccine W = wild type MS N = negative V = MS-H vaccine Discussion According to the results, the wild MS was still playing a role in the production farm, but both vaccines were able to protect the offspring. Faberwe et al. (2017) showed the rate of circulation to be decreased in MS-H vaccinated birds compared to unvaccinated birds. The rearing farm was always negative before vaccinations. All the 25 pools were positive for the vaccine strain in the PS flock vaccinated with MS-H (at 55 and 60 weeks there was 1-1 wild MS pool) and all the 25-25 pools of the offspring were negative at DO and at killing age. Ten pools were positive and 7 pools were negative for the vaccine strain in the PS flock vaccinated with MS Live. Eight pools were positive for MS-H vaccine strain from 55 weeks of age (at 60 weeks there were 2 wild MS pools) and all the 25-25 pools were negative at DO and at killing age. References FEBERWEE, A., J. J. DE WIT, W. J. LANDMAN (2009): Induction of eggshell apex abnormalities by Mycoplasma synoviae: field and experimental studies. Avian Pathol., 38 (1): 77-85
KŐRÖSI, L., J. POVAZSÁN, C. NEUBAUER, M. HESS (2015): Field experiences concerning the long term use of the live, apathogenic, thermosensitive MS vaccine on commercial layer farm in Hungary, WVPA Congress presentation, Cape Town KREIZINGER, Z., K. M. SULYOK, A. PÁSZTOR, K. ERDÉLYI, O. FELDE, J. POVAZSÁN, L. KŐRÖSI, M. GYURANECZ (2015): Rapid, simple and cost-effective molecular method to differentiate the temperature sensitive (ts+) MS-H vaccine strain and wild-type Mycoplasma synoviae isolates, Plos One, 2015 Jul 24;10 (7) KREIZINGER, Z., K. M. SULYOK, D. GRÓZNER, K. BEKŐ, Á. DÁM, Z. SZABÓ, M. GYURANECZ (2017): Development of mismatch amplification mutation assays for the differentiation of MS1 vaccine strain from wild-type Mycoplasma synoviae and MS-H vaccine strains, Plos One, 2017 Apr 18;12 (4) RAVIV, Z., N. FERGUSSON-NOEL, V. LAIDINIS, R. WOOTEN, S. H. KLEVEN (2007): Role of Mycoplasma synoviae in commercial layer Escherichia coli peritonitis syndrome. Avian Diseases, 51:685-690
TERENSKA ISKUSTVA S DUGOTRAJNOM PRIMJENOM ŽIVIH CJEPIVA MS-H I MS NA FARMI RODITELJSKOG JATA U MAĐARSKOJ Sažetak Infekcija matičnih jata (MJ) bakterijom Mycoplasma synoviae (MS) značajno pogoršava ekonomske parametre, smanjuje proizvodnju jaja, predisponira razvoj peritonitisa uzrokovanog bakterijom Escherichia coli te uzrokuje defekte jaja, što umanjuje nesivost. Vertikalno zaraženi potomci mogu slabije napredovati. Dva uzastopna MJ brojlera na farmi s više dobnih skupina cijepljena su živim MS cjepivima. Jedno jato imunizirano je cjepivom MS-H kapima za oči, a drugo jato cjepivom MS Live u spreju. Rezultati su pokazali da je farma s više dobnih skupina zaražena divljim tipom MS. U oba jata su živa MS cjepiva zaštitila potomke. Detektabilnost soja MS-H trajala je duže i bila je snažnija kod jata cijepljenog cjepivom MS-H i čak je nadmašila soj MS1 u jatu cijepljenom cjepivom MS Live u starijoj skupini tog jata. Ključne riječi: Mycoplasma synoviae (MS), živa cjepiva MS, roditeljsko jato, brojler

PERADARSKI DANI 2019. 52
ALTERNATIVNO LIJEČENJE BOLESTI UZROKOVANE PROTOZOOM HISTOMONAS MELEAGRIDIS U ZAGORSKOG PURANA
Tihomir Zglavnik1, Tajana Amšel Zelenika1, Marina Tišljar1, Borka Šimpraga1, Željko Kolar2, Anđelko Ferek Jambrek2
1Hrvatski veterinarski institut, Centar za peradarstvo, Zagreb, Hrvatska 2Krapinsko-zagorska županija, Krapina, Hrvatska
Sažetak
U radu su prikazana četiri slučaja izbijanja simptoma histomonijaze u obiteljskim poljoprivrednim gospodarstvima (OPG), uzgajivačima zagorskog purana. U svim OPG-ima primijenjeno je uobičajeno liječenje antimikrobnim lijekovima i dehelmintizacija uz pojačanje mjera biosigurnosti. Na jednom OPG-u uz navedeno liječenje primijenjeni su i alternativni oblici terapije jabučnim octom, probiotikom i dodatkom do 10% ljekovitog autohtonog bilja u komercijalne smjese. Postotak uginuća na kraju uzgoja od 1% ukazuje na dobar učinak alternativnog oblika liječenja, za razliku od 20% odnosno 30% u slučajevima tretiranim isključivo primjenom antimikrobnih lijekova i dehelmintizacijom uz pojačanje mjera biosigurnosti. Ključne riječi: Crnoglavost, Histomonas meleagridis, zagorski puran, probiotik, jabučni ocat, liječenje antimikrobnim lijekovima, biosigurnosne mjere, Stolisnik (Achillea millefolium),Velika kopriva (Urtica dionica), Maslačak (Taraxacum officinale), Pelin (Artemisia absinthum), Neven (Calendula officinalis), Gospina trava (Hypericum perforatum), Matičnjak (Melisa officinalis), Kadulja (Salvia officinalis) Uvod Zagorski puran je najpoznatiji autohtoni proizvod koji 2016. godine dobiva oznaku zemljopisnog podrijetla. Poseb-nosti zagorskog purana proizlaze iz načina držanja, odnosno ekstenzivnog uzgoja ispašom na otvorenom. Također, za razliku od intenzivno uzgojenih purana koji brzo rastu i uzgajaju se u više ciklusa godišnje, zagorski puran uzgaja se u jednom cjelogodišnjem, prirodnom ciklusu s minimalno 6 mjeseci uzgoja do spremnosti za klaoničku obradu. Zagorski puran na tržištu uvijek je bio prepoznat kao specijalitet, meso posebne kvalitete. Krapinsko-zagorska županija već nekoliko godina dodjeljuje potporu za povećanje uzgoja zagorskog purana na svom po-dručju. Prema izvješću stručnih služba Krapinsko-zagorske županije šteta prouzročena raznim bolestima u uzgojima zagorskog purana iznosila je tijekom 2017. godine oko 700.000 HRK. Histomonijazu je prvi opisao Cushman 1893. godine kao bolest koja je u Americi nanosila iznimno veliku štetu te prepolovila proizvodnju s 6,5 na 3,6 milijuna purana. Ova
bolest i danas stvara velike ekonomske gubitke na tom području. Liebhart i sur. (2016.) u svojem istraživanju tijekom 2014. i 2015. godine bilježe pojavu histomonijaze u Austriji u deset jata komercijalnih farmi purana s velikom smrtnošću, naro-čito u muških purana te zagađivanje velikih pašnih površina protozoom Histomonas meleagridis. Histomonas meleagridis pripada rodu Histomonas, obitelji Dientamoebidae, redu Tritrichomonadida. Protozooa H. meleagridis je veličine od 3 do 21 µm, a nalazimo ga u dva oblika: bičastom s 1-2 biča u lumenu slijepih crijeva i tkiv-nom obliku bez bičeva u stijenci slijepih crijeva i jetri (Hess i McDougald, 2013.). Bolest je poznata u purana, kokoši, čukare, ogrličaste snježnice, dok u blažem obliku obolije-vaju paun, biserka, virđinijska prepelica i fazan. Smrtnost u purana može iznositi do 100% u lošim uvjetima držanja, smještaja i u pokusnim uvjetima (Hauck i Hafez, 2013.), dok u pilića može nanijeti 5%-20% gubitaka. Često se pojavljuje u subkliničkoj formi u peradi te predstavlja značajan izvor zaraze za purane u mješovitim uzgojima (McDouglad, 2005.).
Tihomir Zglavnik, dr. med. vet., Hrvatski veterinarski institut, Centar za peradarstvo, Zagreb, Hrvatska; e-mail: [email protected]

ALTERNATIVNO LIJEČENJE BOLESTI UZROKOVANE PROTOZOOM HISTOMONAS MELEAGRIDIS U ZAGORSKOG PURANA 53
Prenosi se izravno s purana na purana oralnim i kloakalnim putem. Uzročnik u okolišu relativno brzo propada. Lofti i sur. (2012.) utvrdili su da u vodi i fecesu najdulje opstaje do 9 sati. Ako je prisutan Heterakis gallinarum (crv koji para-zitira u cekumu kokoši, purana, lještarke, biserke, jarebice, fazana i prepelice) histomonas biva incistiran u jaja hete-rakisa i zaštićen jajnom ovojnicom te takav može opstati mjesecima i godinama, čemu dodatno pogoduje ako zara-žena jaja proguta gujavica ili puž. Stoga je u prevenciji ove bolesti vrlo važan naglasak na provedbi mjera biosigurnosti i dehelmintizacije. Van der Gulden i Van Erp (1972.) i Capo (2018.) preporučuju tekuću i završnu raskužbu perklorocte-nom kiselinom, karbarilom, tetraklorvinosom, natrijevim hipokloritom i natrijevim dikloroizocijanuratom. Uzročnika mogu širiti insekti poput muha i skakavaca koji prelaskom preko fekalija brzo kontaminiraju okoliš ili sami bivaju pojedeni s uzročnikom. Primarno histomonas se umnaža u crijevima, krvlju dolazi do jetre, ali su zabilježeni i izolati iz bubrega, Fabricijeve burze, voljke, želuca, gušte-rače i pluća (Huck i sur., 2017.). U crijevima izaziva upalne promjene, koje su naročito izražene u slijepim crijevima gdje se očituju čirevima s mogućom perforacijom, posljedič-nom upalom potrbušnice i drugih organa. U jetri se nađu opsežna patognomonična nekrotična žarišta veličine do 4 cm okruglog oblika. U početnoj fazi žarišta su žutomrke do zelene, a u starijih promjena narančasto-žute boje, od kojih po organizaciji procesa stvaraju žućkasto-sivo-bijele ožilj-kaste promjene. Crv Heterakis gallinarum svojom prisut-nošću u sluznici i submukozi izaziva upalu i odebljane sti-jenke slijepih crijeva pojačavajući patogeni učinak parazita Histomonas meleagridis. Opisane promjene se kompliciraju intenzivnim porastom sekundarnih bakterija poput Esche-richia coli, Bacillus subtilis, Clostridum perfingens i dr. Posljedično dolazi do enzimno-funkcionalnog oštećenja crijeva i jetre s proljevom žuto-pjenušave boje izrazito ne-ugodna mirisa. Klinički se bolest očituje 7-12 dana nakon infestacije. Obo-ljeli purani su potišteni, opuštenih krila, zatvorenih očiju, ukočenog hoda, spuštene ili glave smještene ispod krila. Purani zaostaju u rastu, pokazuju loše proizvodne rezultate uz kaheksiju i nisku konverziju hrane. Diferencijalno dijagnostički lezije na jetri treba razlikovati od tuberkuloze, leukoze, trihomonijaze i mikotičnih pro-mjena, a zbog sličnosti promjena u crijevima treba isključiti kokcidiozu. Broj prijavljenih slučajeva histomonijaze diljem svijeta raste nakon 2013. godine (CAC, 2013.) kada je prema Codex alimentarius Organizacije za poljoprivredu i hranu ujedinje-nih naroda i Svjetske zdravstvene organizacije preporučeno nekorištenje nitromidazola. Tijekom 1995. godine Europska unija (EU) također zabranjuje korištenje nitromidazola (Uredbom komisije 1798/95 (CEC, 1995.)). U 2002. godini stavljen je izvan uporabe i nifursol, jedino preostalo sredstvo u borbi protiv histomonijaze (Uredba 1756/2002 (CEC; 2002.)). Odlukom Europske agencije za lijekove od 2009. godine sustavno se prati prisutnost rezidua antihistomo-nijaka na području EU, ali i u zemljama izvoznicama na tržište EU (EMA, 2009.).
S obzirom na nedostatak učinkovitog lijeka potrebno je pristupiti alternativnim mogućnostima liječenja. Tako po-znati francuski travar Maurice Messegue (1988.) navodi da za svaku bolest u okolišu postoji barem jedna ljekovita biljka. Hafez i Hauck (2006.) u svojem su pokusu koristili biljke i postigli dobre preventivne i terapijske rezultate u slučajevima histomonijaze njihovim dodavanjem u hranu za purane. Dokazali su 30% manju smrtnost u pokusno infe-stiranih purana kojima je u hranu dodan cimet, češnjak, limun i ružmarin, za razliku od uobičajeno hranjene pokusno infestirane skupine. Također Willford (1974.) i Treben (2004.) preporučuju dostupne autohtone ljekovite biljke poput stolisnika, velike koprive, maslačka, pelina, nevena, Gospine trave, matičnjaka i kadulje. Hrvatsko zagorje obi-luje brojnom i raznolikom florom od 1346 vrsta krito-sjemenjača na području Parka prirode Medvednica i njenim rubnim dijelovima (Židrovec i sur., 2006.), što omogućava njihovu primjenu kod alternativnih oblika liječenja histo-monijaze. Sekundarnu bakterijsku infekciju potrebno je liječiti anti-mikrobnim lijekovima. Međutim, zbog velike raznolikosti sojeva i rastuće rezistencije bakterija vrlo je važno napraviti test osjetljivosti, što su opisali Šimpraga i sur. (2009.). Istra-žili su antimikrobnu osjetljivost sojeva E. coli izoliranih u industrijski uzgojenih purana na 32 antibiotika. Zaključili su da postoji znatno odstupanje u otpornosti pojedinih sojeva E. coli. Kempf i sur. (2013.) navode da je liječenje paromomicin sulfatom moguće, jer lijek ima dobar profilaktički učinak, međutim, u terapijskom smislu dolazi do razvoja rezistent-nih sojeva crijevnih bakterija pa naglašavaju važnost pra-vodobne dijagnoze. Pristup u liječenju primjenom paromo-micin sulfatom navode kao mogućnost kada nije dostupan nijedan učinkovit lijek uz naglašen metafilaktički (terapija zdravih i oboljelih) učinak u sprječavanju histomonijaze.
Prikaz slučaja
Zbog većih zdravstvenih problema i gubitaka u uzgojima zagorskog purana u 2017. godini Krapinsko-zagorska župa-nija tijekom 2018. godine ugovara suradnju sa Centrom za peradarstvo, koja je za cilj imala unaprjeđenje uzgoja i umanjivanje broja oboljelih i uginulih purana. Sklopljeni ugovor s Krapinsko-zagorskom županijom obuhvatio je te-renski obilazak uzgajivača tijekom mjeseca svibnja i lipnja (1. obilazak), srpnja (2. obilazak) i rujna (3. obilazak). Uz uvid u veličinu proizvodnje, tehnologiju uzgoja i hranidbu utvrđivani su zdravstveni problemi te se pristupilo njihovom rješavanju. U uzgojima zagorskog purana na području Krapinsko-zagorske županije dominirale su dvije bolesti: zarazni sinusitis i histomonijaza. U ovom radu opisan je slučaj izbijanja histomonijaze te mo-gućnost njenog liječenja. Od ukupno trideset i tri uzgajivača u četiri uzgoja klinički je postavljena sumnja na histo-monijazu. Patoanatomski i patohistološki histomonijaza je potvrđena u dva uzgoja, dok su u jednom klinički sum-njivom uzgoju promjene na jetri utvrđene prilikom klanja. U

ALTERNATIVNO LIJEČENJE BOLESTI UZROKOVANE PROTOZOOM HISTOMONAS MELEAGRIDIS U ZAGORSKOG PURANA 54
četvrtom OPG-u klinički sumnjiv puran prilikom prve naznake bolesti izoliran je u karantenu i neškodljivo uklonjen te je tako spriječeno širenje bolesti .
Slučaj 1. OPG (br. 1) je u trenutku obilaska imao sto purića u dobi od deset tjedna. Purići su bili smješteni u čvrsti dio objekta podijeljenog na dva dijela s natkrivenim ispustom te ispustom na travnate površine ograđene pletenom žicom. Dio čvrstog objekta bio je prilagođen uzgoju purića u toploj fazi, dok su u drugom dijelu bile smještene kokoši s ma-tičnim jatom purana (1 mužjak i 4 ženke). U trenutku prvog obilaska tijekom lipnja primijećeno je da kokoši zbog nedo-statno postavljene ograde borave na hranilicama zajedno s puranima. Prema anamnestičkim podatcima pojedini purići bili su nujni, nezainteresirani za hranu i vodu te su se skupljali na hrpe. Pregledom natkrivenog dijela ispusta uočen je pjenu-šavo vodenasti proljev s primjesama krvi. Predložene su dodatne laboratorijske pretrage koje vlasnici nisu prihvatili. Organoleptičkim pregledom smjese u hranilicama uočeno je da smjesa po svojim fizikalno-kemijskim svojstvima ne odudara od poželjnih karakteristika, a pregledom deklaracija utvrđeno je da sastav odgovara uzrasnoj kategoriji purića. S obzirom na nedostatak dodatnih laboratorijskih pretraga, a prema simptomima i nalazu krvi u izmetu postavljena je sumnja na veći broj mogućih bolesti (kokcidioza, koliba-ciloza, histomonijaza). Na osnovi anamneze i kliničkih simptoma preporučena je promjena ugrađenog kokcidiosta-tika u smjesi, dodatak vitamina i probiotika kako bi se uravnotežila crijevna mikroflora uz provedbu učestalije te-kuće dezinfekcije prostora u kojem obitavaju purani. Vla-sniku je savjetovano da u slučaju širenja simptoma unutar jata što prije kontaktira djelatnike Centra za peradarstvo. Na poziv vlasnika već sedam dana kasnije utvrđeno je da oko 20% jata pokazuje znakove nezainteresiranosti za hranu i vodu, nevoljkost, potištenost, skupljanje u hrpe. Pojedini purani imali su glavu smještenu ispod krila, a u dva purana bila je izražena cijanoza glave i vrata. Pregledom natkri-venog dijela betonskog ispusta utvrđena je prisutnost vo-denastog pjenušavog izmeta žute boje neugodna mirisa s tragovima krvi i oljuštenog crijevnog epitela te jedan izmet čvršće konzistencije sa slabo probavljenim sadržajem crijeva i oljuštenim crijevnim epitelom na površini. Jedan od dva purana s cijanozom glave i vrata odavao je izrazito loše zdravstveno stanje. U dogovoru s vlasnikom učinjeno je žrtvovanje i razudba. Rezultat patoanatomske pretrage bio je sljedeći: kataralni enteritis, zadebljanje stijenke slijepih crijeva s ljuštenjem sluznice i kraterastim udubljenjima u kojima nalazimo ma-nju količinu gnojnofibrinskog sadržaja. Na vrhu jednoga jetrenog režnja nalazi se nekrotično žarište veličine do 1 cm okruglog oblika žutomrke do zelene boje. U plućima su uočene blage upalne promjene. S obzirom na nalaz postavljena je osnovana sumnja na histomonijazu te su uzorkovani pluća, voljka, slijepa crijeva i promijenjeni dio režnja jetre za bakteriološku analizu. Jetra i crijeva uzeta su i za patohistološku i parazitološku analizu.
Patohistološki nalaz ukazao je na degenerativne i upalne promjene hepatocita, pretežito limfoidnu hiperplaziju i diseminiranu žarišnu nekrozu. U nekrotičnom tkivu utvrđeni su oblici koji su odgovarali razvojnim oblicima protozoe Histomonas meleagridis. Utvrđene upalne promjene odgo-varale su dvanaestom danu infekcije, što odgovara i kliničkoj slici. Parazitološkom pretragom u crijevima je utvrđena Eimeria spp. Bakteriološkom pretragom iz jetre, crijeva i voljke izdvojena je bakterija E. coli, a anaerobnom pretragom crijeva i voljke izdvojene su bakterije roda Clostridium. Mikološkom pretragom pluća izdvojene su kvasnice. Vlasniku su predložene mjere podizanja biosigurnosti, tera-pija kokoši i purana Dehelmanom 10%® (Krka d.d., levamizol), zakiseljavanje vode domaćim jabučnim octom vrijednosti do pH 6. Kako se vlasnik bavi prikupljanjem lijekovitog bilja, preporučen je obrok koji uz komercijalnu smjesu za puriće sadrži 10%-tni udio autohtonih ljekovitih biljaka (stolisnik, velika kopriva, maslačak i dr.). Pobol u jatu iznosio je 20%, a uginuće 1%. Slučaj 2. Tijekom rujna vlasnik OPG-a (br. 2) telefonskim putem zatražio je savjet zbog povećanih uginuća purana. OPG posjeduje jato od 140 purana dobi 5-6 mjeseci u mješovitom uzgoju s kokošima na zajedničkom ispustu. Vlasnik navodi da je provodio liječenje sulfonamidima i tilozin tartaratom (kupljenim u obližnjoj veterinarskoj ambulanti) i to počet-kom i krajem kolovoza u trajanju od 5 odnosno 3 dana. U tom razdoblju vlasnik bilježi 18%-tno (25/140) uginuće. Kako prema anamnestičkim podatcima nije bilo moguće po-staviti dijagnozu vlasniku je preporučeno donošenje svježe uginulih ili izrazito bolesnih purana na patoanatomsku pretragu i potrebne dopunske laboratorijske analize. Patoanatomskom analizom žrtvovanog purana utvrđena je izrazita kaheksija, povećanje oba jeterena režnja s rasijanim, pretežito okruglo žućkasto-crvenim nekrotičnim žarištima u kojima se sporadično može vidjeti uvučenost središnjeg i ispupčenost perifernog dijela žarišta približna promjera do 1 cm i gnojnofibrinska upala slijepih crijeva. Takav nalaz upućivao je na histomonijazu. Vlasnik nije prihvatio dopun-ske laboratorijske pretrage. Na osnovi patološko anatomskog nalaza vlasniku je prepo-ručena provedba mjera prevencije i biosigurnosti. Kako bi umanjili bol i patnju purana uzrokovanu sekundarnom bak-terijskom infekcijom preporučeno je liječenje enrofloksa-cinom u trajanju od 10 dana te trokratna dehelmintizacija Dehelmanom 10%® (Krka d.d., levamizol) kokoši i purana. Prilikom obilaska polovinom mjeseca rujna utvrđeno je da je vlasnik proveo terapiju i uklonio kokoši. Vlasniku je pre-poručeno korištenje probiotika i prebiotika kako bi se uravnotežila crijevna mikroflora te vapnjenjenje ispusta i preraspodjelu purana na nove pašne površine. Do kraja proizvodnje vlasnik bilježi još 19 uginulih purana. Ukupno uginuće tijekom proizvodnje doseglo je 31% (44/140) purana.

ALTERNATIVNO LIJEČENJE BOLESTI UZROKOVANE PROTOZOOM HISTOMONAS MELEAGRIDIS U ZAGORSKOG PURANA 55
Slučaj 3. Tijekom redovitog obilaska OPG-a (br. 3) koncem srpnja u uzgoju je bilo oko 150 purana dobi oko 5 mjeseci. Vlasnik je kroz posljednja dva tjedna pustio purane na ispašu na kojoj borave i kokoši. Kliničkim pregledom utvrđeno je oko 10% purana sa znakovima nezainteresiranosti za hranu i vodu, nevoljkost, potištenost, a na travnatom ispustu primijećen je žut pjenušav proljev neugodna mirisa. Postavljena je sumnja na prisutnost histomonijaze i sekundarnih bakterijskih infek-cija u jatu. Vlasnik nije odobrio razudbu i dodatne labo-ratorijske pretrage. Na osnovi kliničkih simptoma preporučena je višekratna dehelmintizacija Dehelmanom 10%® (Krka d.d., levamizol) i suzbijanje sekundarnih bakterija oksitetraciklinom u traja-nju od 10 dana. OPG je zabilježio 20%-tno uginuće (30/150) purana do kraja proizvodnje. Prilikom klanja u studenom i prosincu vlasnik zabrinut za zdravstvenu kakvoću mesa purana telefonskim putem opisuje za histomonijazu tipične promjene u jetri koje je utvrdio u oko 10% jata (12/120).
Slučaj 4. Tijekom obilaska OPG-a (br. 4) koji je u uzgoju imao 100 purana u dobi od 5 mjeseci vlasnik napominje velike pro-bleme tijekom 2017. godine kada je uočio vodenast izmet sumporastog mirisa, a kod uginulih purana primijetio je promjene na jetri. U trenutku obilaska na OPG-u je u odvojenoj nastambi bilo smješteno 10-ak kokoši. Vlasnik je u uzgoju primijetio purana sa simptomima bolesti sličnima onim u 2017. godini te ga je izolirao u zaseban objekt. Kako je puran već dosegnuo tjelesnu masu potrebnu za klanje preporučeno je ekonomsko iskorištavanje. Pre-ostali purani i kokoši liječeni su Dehelmanom 10%® (Krka d.d., levamizol) uz vapnjenjenje površina i pojačane mjere biosigurnosti. Prilikom klanja vlasnik je fotografirao jetru te fotografiju proslijedio djelatniku Centra za peradarstvo. Promjene na jetri izgledom su odgovarale mogućoj prisut-nosti histomonijaze.
Rasprava
Histomonijaza kao tvrdokorna bolest teško je iskorjenjiva zbog zabrane uporabe učinkovitih lijekova poput nitromi-dazola, nitrofurana, dimetrinizadola, ipronidazola, ronida-zola, furazolidona, metronidazola, karnidazola i dr. Oteža-vajuća okolnost je i nedostatak materijalnih sredstava u malim OPG-ovima za provođenje laboratorijske dijagno-stike bolesti te kupnju lijekova. Sve je to utjecalo na najbolji mogući pristup borbi protiv ove bolesti i smanjenju velikih gospodarskih gubitaka u uzgojima zagorskog purana. Prije svega bilo je potrebno podići biosigurnosne mjere na najvišu moguću razinu u datim uvjetima. Prvenstveno se to odnosilo na sprječavanje suživota kokoši i purana kao i prijenos uzročnika onečišćenom obućom, opremom i sl. te na izolaciju oboljelih purana. Vrlo je važno bilo provesti čišćenje objekata i ispusta, pre-lijevanje vrućom vodom i posipanje hidratiziranim vapnom. Ispusti moraju biti osunčani i izloženi vjetru kako bi se
potakla prirodna sterilizacija. Preporučeno je smanjivanje gustoće naseljenosti purana po m2 i rotacija pašnih površina, čime bi se umanjila opterećenost histomonasom. Od velikog značenja je provesti terapiju antiparazitikom kako bi se suzbio prijenosnik Heterakis gallinarum i time skratila mogućnost preživljavanja za Histomonas melegaridis u njegovim jajima kroz više godina. Razmatranjem slučaja broj 1. možemo primijetiti da je parazitološkom pretragom dobiven negativan rezultat na prisutnost histomonasa. Međutim, negativan nalaz nije isključio postojanje bolesti s obzirom na to da je zbog dugog razdoblja od uzorkovanja do početka parazitološke pretrage nastupilo propadanje Histomonas meleagridis. Stoga je i dio promijenjene jetre proslijeđen na patohistološku pretragu. U crijevima je parazitološki utvrđeno pet Eimeria spp. što je nedovoljno za razvoj bolesti, međutim, zbog prisutnosti oljuštenog crijevnog epitela i čireva postojali su značajni pogodovni čimbenici za razvoj kokcidioze. Patohistološkom pretragom (slučaj 1.) utvrđene su promjene koje odgovaraju dvanaestom danu infekcije protozoom Histomonas meleagridis. Uz to, bakteriološkom pretragom izdvojene su bakterija E. coli, bakterije roda Clostridium za koje je napravljen test osjetljivosti. Izdvojene bakterije po-kazale su vrlo različitu osjetljivost na antimikrobne lijekove. Najučinkovitiji se pokazao enrofloksacin. Preporučena je terapija u trajanju od 10 dana u vodi za piće. Odmah nakon očitovanja kliničkih simptoma, prije prove-dene patohistološke dijagnostike vlasniku je preporučena primjena ljekovitog bilja s dodatkom od 10% na ukupnu količinu komercijalne smjese. Najveći udio činili su stolisnik, velika kopriva i maslačak, dok je udio ostalih biljaka poput pelina, nevena, Gospine trave, matičnjaka i kadulje bio svega 1%. Kao što je već napomenuto, Willford (1974.) i Treben (2004.) ukazali su na vrlo ljekovit učinak ovih biljaka. Stolisnik (Achillea millefolium) djeluje kod bolesti želuca i crijeva umanjujući grčeve i nadutost crijeva, poboljšava apetit i aktivnost crijevnih žlijezda, otklanja smetnje u radu jetre, umanjuje žučne grčeve, pospješuje rad gušterače. Velika kopriva (Urtica dionica) ublažava crijevne grčeve, zaustavlja krvarenja, uravnotežuje rad gušterače, pozitivno djeluje na jačanje opće otpornosti. Bogata je i brojnim nutrijentima poput željeza, magnezija, natrija, kalija, kalcija, fosfora i obiluje vitaminom A. Maslačak (Taraxacum officinale) poboljšava izlučivanje želučanih so-kova, potiče rad jetre i izlučivanje žuči, ublažava kronične upalne promjene na jetri. Pelin (Artemisia absinthum) pospješuje probavu i apetit, umanjuje grčeve te katar želuca i crijeva, umanjuje otečenje jetre, djeluje antiparazitno na crijevne crve. Neven (Calendula officinalis) ima učinak kod bolesti crijeva i želuca, crijevnih grčeva, proljeva, crijevne upale (kolitis), u liječenju bolesti jetre, djeluje antiparazitno na crijevne crve te jača opću otpornost. Ulje Gospine trave (Hypericum perforatum) djeluju umirujuće i antiparazitno na crijevne crve. Matičnjak (Melisa officinalis) djeluje umiru-juće i umanjuje crijevne grčeve. Kadulja (Salvia officinalis) djeluje protiv upala crijeva, želuca, jetre, ima izvrsno djelo-vanje na oštećenu jetru, otklanja crijevne smetnje i proljeve, olakšava izlučivanje žuči i poboljšava apetit.

ALTERNATIVNO LIJEČENJE BOLESTI UZROKOVANE PROTOZOOM HISTOMONAS MELEAGRIDIS U ZAGORSKOG PURANA 56
Također vlasniku je preporučeno zakiseljavanje vode do-maćim jabučnim octom vrijednosti do pH 6. Učinak ja-bučnog octa očituje se blagim antiseptičnim djelovanjem u pojilicama, a istodobno ima prebiotsko djelovanje sniža-vanjem ukupnog pH u probavnom traktu. Međutim, pritom je važno paziti na održavanje acidobazne ravnoteže u purića. Petog dana nakon provedene antimikrobne terapije s obzirom na narušen sastav crijevne mikroflore purićima je uskraćena voda noću, a u odstajalu vodu dodana je ampula od 6 mL suspenzije Prolife® (Zeta Farmaceutici) koja sadrži žive bakterijske kulture Lactobacillus acidophilus, Bacillus coagulans, Lactobacillus sporogenes, Bifidobacterium lactis, Bifidobacterium lactis, Lactobacillus plantarum sa žlicom saharoze na 5 L vode kroz tri dana ujutro. Usporedi li se slučaj 1. gdje je uz uobičajenu antimikrobnu terapiju i dehelmintizaciju uz pojačane mjere biosigurnosti primijenjen i alternativni oblik liječenja, vidljivo je da je po-stotak uginuća na kraju uzgoja od 1% ukazao na uspješan učinak alternativnog oblika terapije, za razliku od 20% od-nosno 30% u slučajevima tretiranim isključivo antimikrob-nom terapijom i dehelmintizacijom uz pojačanje mjera biosigurnosti.
Zaključak
S obzirom na izrazito dobre rezultate u slučaju 1. u kojem je pobol purana u trenutku dijagnoze iznosio 20% možemo zaključiti da je pravodobna dijagnoza, terapija kokoši i pu-rana Dehelmanom® (Krka d. d., levamizol) i enrofloksa-cinom, zakiseljavanje vode domaćim jabučnim octom do pH 6 te obrok koji je uz komercijalnu smjesu za puriće sa-državao i 10%-tni udio autohtonih ljekovitih biljaka uz po-većanje biosigurnosnih mjera i primjenu probiotika rezulti-rao uginućem od svega 1%. Važnu ulogu odigrala je primjena antimikrobnih lijekova u suzbijanju sekundarne bakterijske infekcije, čime je uma-njena smrtnost u sva tri slučaja bolesti. Zakiseljavanje jabučnim octom i upotreba probiotika imala je dobar učinak u stabilizaciji crijevne mikroflore. Zaključno se može reći da su potrebna daljnja istraživanja kako bi se ispravno valorizrala terapijska doza odnosno učinak svake od navedenih biljaka kao i njihovih sastojaka.
Literatura
CAC. (2013): Joint FAO/WHO food standard program codex alimentarius commission, residues of veterinary drugs in Food. (Codex Classification of Foods and Animal Feeds).
CEC. (1995). Commission Regulation (EC) No 1798/95 of July 25, 1995 amending Annex IV to Council Regulation (EEC) No 2377/90 layingd own a Community procedure for the establishment of maximum residue limits of veterinary medicinal products in foodstuffs of animal origin. Off. J., L 174, 20–21.
CEC. (2002). Council Regulation (EC) No 1756/2002 of 23 September 2002 amending Directive 70/524/EEC concer-ning additives in feeding stuffs as regards with draw al of the authorisation of an additive and amending Commission Regulation (EC) No 2430/1999. Off. J., L 181, 1–2
CUPO, K. L. (2018.): Methods for Identification and Eradication of Heterakis gallinarum Vectors on Poultry Facilities to Prevent Blackhead Disease Outbreaks. Master of Science in
Poultry Science, Raleigh, North Carolina CUSHMAN, S. (1893). The production of turkeys. Rhode Island
Agricultural Experiment Station Bulletin, 25: 89–123. EMA. (2009): European Medicines Agency. Status of MRL
procedures MRL assessmentsin the context of Council Regulations (EEC) No 2377/90. EMEA/CVMP/765/99-Rev. 23, 1–19.
HAFEZ, H. M., R. HAUCK (2006). Efficacy of a herbal product against Histomonas
Meleagridis after experimental infection of turkey poults. Arch. Anim. Nutrit. 60, 436-442..
HAUCK, R.,H. M. HAFEZ, (2013): Experimental infections with the protozoan parasite Histomonas meleagridis: a review. Paras. Res., 112: 19–34.
HAUCK, R., S. STOUTE, P. R. CHAIN, C. G. SENTIES-CUE, H. L. SHIVAPRASED, (2017.): Retrospective study of Histomonas (Balachead) in California turkey flocks 2000 – 2014. Av. Dis., 62 (1): 94-100..
HESS, M., L. R. MC DOUGALD (2013). Histomoniasis (black-head) and other protozoan diseases of the intestinal tract. In D. E. Swayne, J. R. Glisson, L. R. McDougald, L. K. Nolan, D. L. Suarez, V. Nair (Eds.), Dis. Poult. 13th edn (pp. 1172–1182). Ames, IA: Wiley-Blackwell
KEMPF, I., R. A. LE, A PERRIN-GUYOMARD, G. MOURAND, D. L. LE, S BOUGEARD, P. RICHEZ, P. G. LE, N. ETERRADOSSI (2013). Effect of in-feed paromo-mycin supplementation on antimicrobial resistance of enteric bacteria in turkeys. Vet. J., 198: 398–403
LIEBHART, D., P. GANAS, T. SULEJMANOVIĆ, M. HESS (2016): Histomonosis in
poultry: previous and current strategies for prevention and therapy, Av. Pathol., 46 (1), 1-18.
LOTFI, A. R., E. M., ABDELWHAB, H. M HAFEZ, (2012). Persistence of Histomonas meleagridis in or on materials used in poultry houses. Av. Dis., 56: 224-226.
MESSEGUE M.(1988.): O ljudima i travama. Nakladnik: Otokar Keršovani, Opatija
MC DOUGALD L. R. (2005.): Blackhead disease (Histomo-niasis) inpoultry: A critical review. Av. Dis., 49(4): 462-476.
ŠIMPRAGA, B., M. SOKOLOVIĆ, F. KRSTULOVIĆ (2009.): Osjetljivost sojeva bakterije E. coli izdvojenih iz tovnih purana na antibiotike. Simpozij „Peradarski dani 2009.“ s međunarodnim sudjelovanjem Poreč, Hrvatska, 25-28. 03. 2009.. Zbornik radova.
TREBEN, M. (2004.): Zdravlje iz Božje ljekarne: savjeti i isku-stva u liječenju ljekovitim biljem, prijevod Gesundheitaus Apotheke Gottes. Mozaik knjiga.
VAN DER GULDEN, W. J., A. J. VAN ERP (1972). The effect of paraceticacid as a disinfectant on worm eggs. Laboratory Animal Science, 22: 225–226.
WILIFORT, R.(1974.) Ljekovito bilje i njegova upotreba. Izda-vačko knjižarsko poduzeće Mladost.
ŽIDOVEC, V., I. VRŠEK, B. ANČIĆ, S. GRZUNOV (2006.): Tradicijski seoski vrtovi sjeverozapadne Hrvatske. Sjeme-narstvo, 23: 273-283.

ALTERNATIVNO LIJEČENJE BOLESTI UZROKOVANE PROTOZOOM HISTOMONAS MELEAGRIDIS U ZAGORSKOG PURANA 57
ALTERNATIVE TREATMENT OF PROTOZOAN DISEASE CAUSED BY HISTOMONAS MELEAGRIDIS IN ZAGORJE TURKEY
Summary
Four cases of histomoniasis symptoms on family farms of Zagorje turkey breeders are presented. Standard therapy with antimicrobial agents, dehelminthisation and reinforced biosecurity measures were applied on all these farms. Alternative modes of therapy with apple vinegar, probiotics and addition of up to 10% of autochthonous medicinal herbs to commercial feed were used on one of the farms. The 1% rate of deaths recorded on this farm at the end of the breeding process indicated the favourable effect of alternative treatment, as compared with 20% and 30% rate in the animals treated exclusively with antimicrobials, dehelminthisation and increased biosecurity measures. Key words: Blackhead disease, Histomonas meleagridis, Zagorje turkey, probiotic, apple vinegar, therapy with anti-microbials, biosecurity measures, Achillea millefolium, Urtica dionica, Taraxacum officinale, Artemisia absinthum, Calendula officinalis, Hypericum perforatum, Melisa officinalis, Salvia officinalis

PERADARSKI DANI 2019. 58
INAKTIVACIJA MIKOTOKSINA U HRANI ZA ŽIVOTINJE
Marijana Sokolović
Hrvatski veterinarski institut, Centar za peradarstvo, Zagreb, Hrvatska
SAŽETAK
Patogene plijesni i njihovi sekundarni metaboliti, mikotoksini, često su prisutni u sirovinama, hrani i hrani za životinje. Pojedini mikotoksini toksični su za ljude i životinje već u vrlo niskim koncentracijama te stoga njihova pojavnost u hrani može nepovoljno utjecati na zdravlje i proizvodnost životinja te zdravlje ljudi. Zbog velike učestalosti mikotoksina te pojave trovanja i obolijevanja u ljudi i životinja koje se mogu povezati s plijesnima i mikotoksinima provedena su brojna istraživanja kojima je cilj smanjenje njihovog potencijalnog toksičnog učinka. Novija istraživanja usmjerena su ne samo na sprječavanje rasta i širenja patogenih plijesni, već i na razvoj postupaka i metoda inaktivacije mikotoksina u manje toksične metabolite. Iako su mikotoksini relativno stabilni te se ne uništavaju prilikom obrade i prerade sirovina i hrane, mogu se ipak inaktivirati nekim postupcima pri čemu nastaju metaboliti slabije toksičnosti. Najviše su ispitivani različiti fizikalni, kemijski i biološki postupci detoksikacije učinkovitost kojih ovisi o vrsti toksina, njihovoj rasprostranjenosti i razini kontaminacije. Uspješnost inaktivacije umanjuje istodobna prisutnost različitih mikotoksina te neke toksikološke interakcije. Kako bi se nepoželjni učinak mikotoksina u životinja i ljudi sveo na najmanju moguću mjeru, sprječavanje njihova nastanka i dalje se smatra najboljom preventivnom mjerom. Ipak, zbog nedostupnosti odgovarajućih sirovina, ekonomskih čimbenika i zahtjeva tržišta u praksi se često, s više ili manje uspjeha, primjenjuju različiti postupci inaktivacije mikotoksina. U radu se daje pregled postupaka inaktivacije za najznačajnije mikotoksine s obzirom na njihovu učestalost i potencijalne štetne učinke u životinja.
Ključne riječi: Mikotoksini, plijesni, hrana za životinje, inaktivacija
Uvod Infekcija žitarica toksikogenim vrstama plijesni (prije ili poslije žetve, tijekom transporta i skladištenja) dovodi do kontaminacije sirovina i hrane mikotoksinima. Mikotoksini su sekundarni metaboliti filamentoznih plijesni koji mogu uzrokovati toksikoze u ljudi i životinja. Danas je poznato više od 400 strukturno različitih mikotoksina od kojih su kao kontaminanti najčešće detektirani aflatoksini, trikoteceni, zearalenon, fumonizini i drugi. Proizvode ih plijesni rodova Aspergillus, Penicillium, Fusarium te rjeđe plijesni drugih rodova (Sokolović i sur., 2008., Bryła i sur., 2016.). Brojna istraživanja ukazala su na povezanost prisutnosti miko-toksina u hrani i hrani za životinje s pojavom akutnih ili kroničnih simptoma, uključujući nastanak karcinogenih, neurotoksičnih i imunomodulacijskih štetnih učinaka. Hra-nidba životinja kontaminiranom hranom može uzrokovati loše proizvodne rezultate kao što su smanjeni unos hrane, slabija konverziju i pad tjelesne mase te povećanu učestalost bolesti zbog imunomodulacijskih učinaka toksina i drugo. Uz navedeno, hranjenje kontaminiranom hranom može dovesti do pojave rezidua mikotoksina u jajima, mlijeku i
mesu predstavljajući i opasnost za zdravlje ljudi (Pestka, 2007., Streit i sur., 2012., Marin i sur., 2013., Omotayo i sur., 2019.). Prisutnost plijesni i mikotoksina u sirovinama i gotovoj hrani ozbiljan je problem u proizvodnji hrane. Zbog klimatskih promjena, globalne trgovine i zahtjeva tržišta kao i dostupnih preciznih metoda detekcije mikotoksina u da-našnje vrijeme gotovo je nemoguće nabaviti sirovinu koja ne sadrži barem jedan mikotoksin (Klich, 2007., Miraglia i sur., 2009.). Budući da hrana za životinje sadrži velik udio sirovina, važno je znati jesu li iste potencijalni izvor miko-toksina. Pritom je količina mikotoksina u sirovinama u pravilu veća u odnosu na gotov proizvod budući da se isti u formulaciji gotovih proizvoda ipak koriste u manjoj mjeri (kombiniranje sirovina s ostalim sastojcima gotove hrane za životinje), ali i stoga što pojedini postupci obrade hrane mogu umanjiti broj plijesni i mikotoksina. Zbog neizbježne kontaminacije, a u cilju smanjenja po-tencijalnog štetnog učinka plijesni i mikotoksina, mnoge zemlje svijeta uvele su stroge zakonske propise za kontrolu mikotoksina u hrani i hrani za ljude. U zemljama Europske Unije prisuntost mikotoksina pri proizvodnji i uporabi hrane
Dr. sc. Marijana Sokolović, dr. med. vet, Hrvatski veterinarski insitut, Centar za peradarstvo, Laboratorij za analitiku stočne hrane, Heinzelova 55., 10000 Zagreb, Hrvatska; Tel.: +385 (0)1 2440-214; Fax: +385 (0)1 2441-396; E-mail: [email protected]

INAKTIVACIJA MIKOTOKSINA U HRANI ZA ŽIVOTINJE 59
regulirana je Uredbom komisije (EZ) br. 1881/2006. i nekoliko drugih propisa (2002/32/EZ, 2004/852/EC, 2005/949/EU, 2006/401/EC, 2006/576/EZ, 2006/583/EC, 2013/165/EU, 2014/519/EU). Najveće dopuštene količine (NDK) mikotoksina u hrani procijenjene su na temelju rezultata istraživanja toksičnosti i izloženosti pojedinim toksinima, a izražene su u dnevnom obroku hrane za živo-tinje s udjelom vlage od 12% (2002/32/EK). Od miko-toksina u hrani za životinje NDK postoji samo za aflatoksin. Za druge mikotoksine na razini Europske Unije propisane su Smjernce o prisuthosti deoksinivalenola, zearalenona, ohra-toksina A, T-2 i HT-2 toksina te fumonizina u hrani za ži-
votinje (2006/576/EC). Pri hranidbi životinja žitaricama i proizvodima od žitarica koje sadrže mikotoksine u „doz-voljenim koncentracijama“ te su u skladu sa zakonskim propisima i smjernicama posebnu pozornost treba usmjeriti na udio takvih žitarica u dnevnom obroku životinje kako ne bi došlo do izlaganja životinja visokim koncentracijama mikotoksina. U slučaju da neka hrana za životinje sadrži količine nepoželjnih tvari koje prekoračuju najvišu do-puštenu količinu definiranu u direktivi (tablica 1.) ili smjernicama, ona se ne smije miješati s istim ili drugim proizvodima namijenjenim za hranu za životinje kako bi se „razrijedila“ količina mikotoksina u konačnom proizvodu.
Tablica 1. Najveće dopuštene količine (NDK) mikotoksina u hrani za životinje (2002/32/EZ, Prilog 1.)
Mikotoksin Proizvodi namijenjeni za hranu za životinje NDK (mg/kg)* Aflatoksin B1 Krmiva s iznimkom:
0,05 kikirikija, kopre, ploda kokosove palme, palminih koštica, sjemenki pamuka, oraha
palme babassu, kukuruza i proizvoda dobivenih prera�om istih
0,02
Potpuna krmna smjesa za goveda, ovce i koze s iznimkom: 0,05 mliječnih goveda
0,005 teladi i janjadi
0,01 Potpuna krmna smjesa za svinje i perad
(osim mladih životinja) 0,02
Ostala potpuna krmna smjesa 0,01 Dopunska krmna smjesa za goveda, ovce i koze
(osim dopunska krmna smjesa za životinje za proizvodnju mlijeka, telad i janjad) 0,05
Dopunska krmna smjesa za svinje i perad (osim mladih životinja) 0,03
Ostala dopunska krmna smjesa 0,005
*Najveća dopuštena količina u mg/kg (ppm) u odnosu na hranu za životinje s udjelom vlage od 12% Table 1. Maximum content of mycotoxins in feedingstuff (2002/32/EC, Annex 1)
Mycotoxin Products intended for animal feed MC (mg/kg)* Aflatoxin B1 Feed materials with the exception�of:
0.05 groundnut, copra, palm-kernel, cotton seed, babassu, maize and products derived from the
processing thereof
0.02
Complete feedingstuffs for cattle, sheep and goats with the exception of: 0.05 dairy cattle
0.005 calves and lambs
0.01 Complete feedingstuffs for pigs and poultry
(except young animals) 0.02
Other complete feedingstuffs 0.01 Complementary feedingstuffs for cattle, sheep and goats (except complementary feedingstuffs
for dairy animals, calves and lambs) 0.05
Complementary feedingstuffs for pigs and poultry (except young animals) 0.03
Other complementary feedingstuffs 0.005
*Maximum content in mg/kg (ppm) relative to a feedingstuff with a moisture content of 12%

INAKTIVACIJA MIKOTOKSINA U HRANI ZA ŽIVOTINJE 60
Međutim, zbog globalnih klimatskih promjena i njihovog utjecaja na poljoprivredu i uzgoj životinja te spomenutih zahtjeva tržišta, kontaminirana sirovina (u količinama iznad NDK vrijednosti propisanih zakonom) često je jedini izbor. Proizvođači hrane moraju stoga prvenstveno otkriti izvor kontaminacije te prema potrebi i mogućnostima primijeniti odgovarajuće postupke dekontaminacije. Cilj dobre pro-izvođačke prakse je proizvodnja zdrave, nepatvorene hrane koja ne predstavlja opasnost za zdravlje ljudi, životinja i/ili okoliša. Postupci detoksifikacije/inaktivacije mikotoksina Prisutnost plijesni i mikotoksina u hrani može se izbjeći na nekoliko načina: prevencija kontaminacije različitim agro-tehnološkim mjerama, uklanjanje kontaminirane hrane, smanjenje količine mikotoksina u hrani kemijskim, fizi-kalnim i biološkim metodama te tretiranjem oboljelih ljudi i životinja. Kako je vrlo teško umanjiti štetne učinke mi-kotoksina u praksi, najveća pozornost svakako se posvećuje sprječavanju nastanka mikotoksina te se primjenjuju brojne preventivne mjere još pri uzgoju žitarica (rotacija, raspored sadnje žitarica, uzgoj rezistentnih i/ili rano zrelih hibrida, biološka i kemijska kontrola bolesti bilja i pojave insekata i
glodavaca, sprječavanje rasta korova, dostupnost vode, primjena fertilizatora, žetva prije pune starosti, sušenje i drugo) (slika 1.). Cilj takvih postupaka je uzgoj što kvali-tetnijih žitarica (s obzirom na hranjivi sastav) uz istodobno sprječavanje stvaranja uvjeta za razvoj plijesni i/ili oštećenja zrna. Kontrola pojavnosti plijesni i mikotoksina te procjena razine kontaminacije mikotoksinima neophodne su mjere sigurnosti u cijelom proizvodnom procesu (uzgoj sirovina, prerada, trasnport i skladištenje). Navedene mjere dio su analize opasnosti i određivanja kritičnih kontrolnih točaka (HACCP, engl. Hazard Analysis Critical Control Points) (FAO, 2001., Aldered i i Magan, 2004., Edwards, 2004., Scudamore, 2005., Alberts i sur., 2006., Chulze, 2010., Jacobsen, 2014., Mesterhazy, 2014., Bryden, 2017.). Uspješnost fizikalnih, kemijskih i bioloških metoda te učinkovitost dodataka u hrani u smanjenju nepoželjnih štetnih učinaka mikotoksina ovisi o brojnim čimbenicima, ali tek poznavanje mehanizma detoksifikacije omogućuje odabir pravilne metode (tablica 2.). U praksi, najčešći je odabir kombinacija fizikalnih, kemijskih i bioloških metoda koja s većim ili manjim uspjehom i uz prihvatljiv ekonomski trošak može umanjiti nepoželjne posljedice kontaminacije mikotoksinima (Basappa i Shanta., 1996., Karlovsky i sur., 2016.).
Slika 1. Prikaz dostupnih metoda prevencije i dekontaminacije sirovina, hrane i hrane za životinje prije i poslije infekcije plijesnima i kontaminacije mikotoksinima (Hojnik i sur., 2017)
Figure 1. Overview of available prevention and decontamination methods taken before and after fungal and mycotoxin contamination of raw materials, food and feed (Hojnik i sur., 2017)

INAKTIVACIJA MIKOTOKSINA U HRANI ZA ŽIVOTINJE 61
Tablica 2. Prikaz fizikalnih i kemijskih metoda za smanjenje koncentracije pojedinih mikotoksina u žitaricama (Prilagođeno od Karlovsky, 2016)
Table 2. Overview of physical and chemical processes for minimising of mycotoxin content in cereals (Addapted from Karlovsky, 2016)
Postupak/ treatement
Vrsta hrane/ Feed Type
AFLA OA ZON FUM TCT
Fizikalni postupci/Physical treatement Sortiranje/ Sorting Kukuruz/Corn X D D X X
Žitarice/Cereals NA D D NA X Prosijavanje/ Siewing
Kukuruz/Corn D D D D D Žitarice/Cereals NA D D NA X
Flotacija/ Flotation
Kukuruz/Corn X NA NA X X Žitarice/Cereals NA NA NA NA X
Pranje/ Washing
Kukuruz/Corn D NA D D D Žitarice/Cereals NA NA D NA D
Ljuštenje/ Dehulling
Kukuruz/Corn X NA NA NA X Žitarice/Cereals NA NA X NA X
Namakanje/ Steeping
Kukuruz/Corn X X X X X Žitarice/Cereals NA D D NA D
Mljevenje/ Milling
Kukuruz/Corn X NA X X X Žitarice/Cereals NA X X NA X
Zagrijavanje/ Heating
Kukuruz/Corn X X X X X Žitarice/Cereals NA X X NA X
UV zračenje/ UV radiation
Kukuruz/Corn X D NP NP NP Žitarice/Cereals NA D NP NA NP
Y zračenje/ Y radiation
Kukuruz/Corn X D D X D Žitarice/Cereals NA D D NA D
Hladna plazma/ Cold Plasma
Kukuruz/Corn D D D D D Žitarice/Cereals NA D D NA D
Kemijski postupci/ Chemical treatment Kiseline/ Acid
Kukuruz/Corn X D D D NA Žitarice/Cereals NA D NA NA D
Lužine/ Alkaline
Kukuruz/Corn X D X D X Žitarice/Cereals NA D X NA X
Oksidacija/ Oxidation
Kukuruz/Corn X D X D D Žitarice/Cereals NA D D NA D
Redukcija/ Reduction
Kukuruz/Corn X NP NP NP NA Žitarice/Cereals NA NP NP NA D
Amonizacija/ Ammoniation
Kukuruz/Corn X X X NP X Žitarice/Cereals NA X X NA D
NP Nije primjenjivo/Not applicable AFLA – Aflatokisni/Aflatoxins; OA- Ohratoksin A/Ochratoxin A; ZON – Zearalenon/Zearalenone; FUM – Fumonizini/Fumonisins; TCT – Trihoteceni/Trichothecenes
D Djelomično uspješno/Partially applicable X Eksperimentalno dokazano/Experimental
NA Nema dovoljno informacija/Not enough information

INAKTIVACIJA MIKOTOKSINA U HRANI ZA ŽIVOTINJE 62
Fizikalne metode
Fizikalne metode dekontaminacije uključuju različite po-stupke kao što su čišćenje, mljevenje, sortiranje, kuhanje, kuhanje u alkalnim uvjetima, pečenje, prženje, fermentaciju, ekstrudiranje, zračenje, primjenu hladne plazme i drugo. Čišćenje, sortiranje. Nakon žetve, osnovna mjera preven-tive rasta toksikogenih plijesni i proizvodnje mikotoksina je kontrola kvalitete sirovina. Pritom se nastoje izbjeći ošte-ćenja zrna, gubitak kvalitete te spriječiti onečišćenja mikro-organizmima i prisutnost insekata i glodavaca. Strogi pro-grami higijene uvelike će doprinijeti smanjenju potencijalne kontaminacije plijesnima i mikotoksinima. Čišćenje podra-zumijeva održavanje higijene skladišta i okoliša, prijevoznih sredstava i sve opreme kako bi se u slučaju da i postoji one-čišćenje spriječilo njegovo širenje (FAO, 2002., Eeckhout i sur., 2013.). Pojedini postupci mogu utjecati na smanjenje količine mikotoksina u hrani, ali učinkovitost nije jednako uspješna za sve mikotoksine. Primjerice, ispiranje vodom ili natrijevim karbonatom pokazalo se uspješnim u smanjenju koncentracija zearalenona i fumonizina B1 u žitaricama. Isto je primjenjivo za mikotoksine koji su topljivi u vodi ili u alkalnim otopinama. Smanjenje ohratoksina postignuto je primjenom postupaka uklanjanja kontaminiranih zrna (Ka-bak i sur., 2006.). Općenito se smatra da je uklanjanje ošte-ćenih i nezrelih zrna jedna od najučinkovitijih metoda smanjenja kontaminacije mikotoksina. Razlog navedenom je to što se mikotoksini najčešće zadržavaju upravo u ošteće-nim i slomljenim zrnima. Sortiranje može biti ručno, zrakom i gravitacijom. Primjerice, uklanjanje zrna kontaminiranih fumonizinima postupkom sortiranja prosijavanjem (sita veli-čine 3 mm) prije daljnje obrade smanjilo je ukupnu razinu fumonizina u rasponu od 26,2% do 69,4% (Sydenham i sur., 1994., Bullerman i Bianchini, 2007.). Sortiranje oštećenih zrna kukuruza radi uklanjanja deoksinivalenola i zearale-nona smanjilo je kontaminaciju toksina za 73% i 79%. Ipak, metoda se ne smatra uspješnom, jer je istodobno bilo odbačeno i 69% ukupne količine kukuruza (Trenholm i sur., 1991.). Razdvajanje zrna provodi se ne samo uklanjanjem oštećenih zrna i zrna vidljivo zahvaćenih plijesnima, već i na temelju oblika, boje, veličine, gustoće i drugog. U pravilu, način sortiranja ovisi o vrsti žitarice/hrane. U praksi se je često primjenjivalo sortiranje zrna kukuruza pregledom pod UV svjetlom (365 nm ili više). Pritom nalaz fluorescencije smatrao se dokazom prisutnosti aflatoksina. Navedena fluo-rescencija nastaje zbog reakcije enzima biljne peroksidaze koji reagira s kojičnom kiselinom koju proizvodi plijesan Aspergillu flavus. Problem kod ove metode je to što ne proizvodi svaka vrsta plijesni navedenu kiselinu te time neće ni doći do pojave fluorescencije, a zbog visokih temperatura dolazi i do kemijskih promjena kojične kiseline te također neće nastati fluorescencija. Metoda UV sortiranja isto tako nije jednako uspješna za sve žitarice i zrna, a česta je i pojava lažno pozitivnih i lažno negativnih rezultata (Bothast i Hesseltine, 1975.). Općenito, sortiranje kao metoda ukla-njanja potencijalne kontaminacije mikotoksinima može re-zultirati značajnim smanjenjem kontaminacije, ali se ista zbog dugotrajnog i napornog postupka u praksi rjeđe pro-
vodi (Castells i sur., 2007., van der Westhuizen i sur., 2011., Khatibi i sur., 2014., Matumba i sur., 2015.). Novija metoda sortiranja je metoda NIRS (engl. near infrared spectro-scopy). Princip metode je mjerenje valne duljine i intenziteta apsorpcije infracrvenog zračenja u ispitivanom uzorku uz apsorpciju elektromagnetskog zračenja na valnim duljinama u rasponu od 500 do 2500 nm kako bi se odredile funk-cionalne skupine molekula. Metoda je neinvazinva i korisna za kontrolu kvalitete hrane tijekom i pri završetku proiz-vodnje, a pokazala je i dobre rezultate za otkrivanje mikotoksina u orašastim plodovima i žitaricama. Usprkos tome što je metoda relativno jednostavna i financijski isplativa, nije se pokazala dovoljno preciznom s obzirom na definirane zakonske propise (Alander i sur., 2013., McMullin i sur., 2015., Levasseur-Garcia, 2018.). Sušenje i prerada. Usprkos brojnim čimbenicima koji utje-ču na rast plijesni i proizvodnju mikotoksina, temperatura, udio vlage u hrani i aktivnost vode (aw) smatraju se najzna-čajnijim čimbenicima. Zbog navedenog, sušenje žitarica po-trebno je provesti u što kraćem roku nakon žetve, a skladištenje mora biti u uvjetima kontrolirane vlage i temperature (FAO, 2002.). Aktivnost vode (aw) označava "slobodnu vodu" koja je dostupna za rast plijesni. U prirodi postoji velik broj plijesni koje trebaju drugačije uvijete za rast (kserofilne, higrofilne i mezofilne plijesni). Hrana koja sadrži veće razine aw u pravilu pogoduje rastu toksikogenih plijesni. Smatra se da higrofilne plijesni koje najčešće rastu u žitaricama prije žetve (npr. Alternaria spp., Fusarium spp.) zahtijevaju 1,00 aw, mezofilne plijesni (npr. Clado-sporium spp, Verticillium spp., Geotrichum spp.) koje najčešće rastu tijekom žetve 0,95-1,00 aw, a kserofilne plijesni (npr. Aspergillus spp., Penicillium spp.) koje rastu nakon žetve <0,95 aw. Kako na rast mikotoksikogenih plijesni uz vodu i temperaturu utječe više čimbenika, može se smatrati da će i bez osiguranja optimalnih uvjeta plijesni rasti u hrani s udjelom vlage od 13% do 22% i u širokom rasponu temperatura. Smanjeni rast plijesni bit će kada udio vlage padne ispod 8% (Mannaa i Kim, 2017.). Mikotoksini su u pravilu vrlo stabilne tvari i ne uništavaju ih standadni postupci obrade i prerade hrane. Ipak, neki postupci i primjena temperatura mogu utjecati na količinu toksina u hrani. Primjerice, namakanje zrna kukuruza kroz 36-50 sati pri temperaturi od 50 ºC u vodi koja sadrži 0.1% - 0.2% SO2 postupak je kojim se potiče stvaranje mliječne kiseline te dolazi do pucanja proteinskog matriksa. Ovaj postupak je prvi korak pri vlažnom mljevenju kukuruza, a pokazao se uspješnim u uklanjanju aflatoksina, fumonizina, zearalenona i deoksinivalenola (Aly, 2002., Lefyedi i Taylor, 2006.). Nadalje, meljevenjem se također može sma-njiti kontaminacija mikotoksinima, jer se uklanjaju dijelovi koji sadrže više mikotoksina (aflatoksini i plijesni se naj-češće zadržavaju na površini zrna, dok se mikotoksini poput deoksinivalenola i zearalenona češće nalaze u dijelovima žitarica koje sadrže vlakna). Primjerice, smanjenje količine mikotoksina uočeno je pri proizvodnji brašna koje je bilo značajno manje kontaminirano aflatoksinima u odnosu na mekinje (Cheli i sur., 2013., Kilonzo i sur., 2014., Schwake-

INAKTIVACIJA MIKOTOKSINA U HRANI ZA ŽIVOTINJE 63
Anduschus i sur., 2015., Tibola i sur., 2015.). Primjena visokih temperatura (iznad 100 ºC) u postupcima tostiranja, prženja, pečenja i ekstrudiranja također se pokazala uspješ-nom u uklanjanju mikotoksina kao što su aflatoksini i fu-monizini (Hahn i sur., 2015.). Oliveira i suradnici (2013.) su pri ekstrudiranju uočili smanjenje količine aflatoksina za 50%-80%. Smanjenje od 40%-80% aflatoksina u uzorcima kukuruza postignuto je i pečenjem te pri temperaturama iznad 160 °C (Conway i sur., 1978., Raters i Matissek, 2008.). Učinkovitost ovih postupaka ovisila je o vrsti tok-sina, temperaturi obrade i udjelu vlage u uzorcima hrane. Uz aflatoksine, primjena temperature je uspješna i za smanjenje količine fumonizina, zearalenona i citrinina, dok je slabije učinkovita za inaktivaciju ohratoksina, deoksinivalenola i nivalenola (Trivede i sur., 1992., Scudamore i sur., 2004., Bullerman i Bianchini, 2007., Scudamore i sur., 2008., EFSA, 2014., Karlovsky i sur., 2016.). Zračenje. Neionizirajuće zračenje (sunčevo svjetlo, UV i mikrovalovi) i ionizirajuće zračenje (gama zračenje) mogu također djelomice ukloniti patogene mikroorganizme iz hra-ne, a tako i mikotoksine. Razgradnja aflatoksina sunčevim svjetlom smanjila je razinu toksina za 40% nakon tri sata te čak 75% nakon 30 sati, a zearalenona čak i do 90%. Sunčevo svjetlo pokazalo se je uspješnijim i od mikrovalova i gama zračenja. Tretiranje pšenice kontaminiratne aflatok-sinima UV zrakama (254 nm i 362 nm) u trajanju od 30 minuta u potpunosti je inaktiviralo aflatoksin (Atalla i sur., 2004.). Mikrovalno zračenje i konvekcijsko grijanje mogu smanjiti količinu mikotoksina za 32% nakon 30 minuta djelovanja, odnosno 40% ako su postignute više temperature (150-175 ºC). Za gama zračenje provedena su različita ispitivanja te je zabilježeno 43%-tno smanjenje količine aflatoksina u dozi od 25 kGy, ali svega 11%-21% u dozi od 15 kGy. Za fumonizine doza od 15 kGy rezulitrala je smanjenjem količine toksina od 20%, a smanjivanje količine postignuto je i za deoksinivalenol (Refai i sur., 1996., Visconti i sur., 1996., Bretz i sur., 2006., Herzallah i sur., 2008., Köppen i sur., 2012.). UV zračenje se također po-kazalo uspješnim u uklanjanju patogenih mikroorganizama i mikotoksina. Za razliku od drugih metoda, ono ne koristi temperaturu (te tako ne dolazi do kemijskih promjena), a ekonomski je isplativije. Hladna plazma. Obrada hrane primjenom hladne plazme (CAP, engl. atmospheric pressure plasma) postupak je de-kontaminacije mikotoksina bez primjene topline. Plazma kao četvrto stanje tvari nastaje povisivanjem energetske razine tvari od čvrstog stanja preko tekućeg i plinovitog do ioniziranog stanja plina specifičnih fizikalnih i kemijskih karakteristika s rezultirajućim oksidirajućim učinkom. Ne-davna istraživanja ukazala su na brojne primjene ove tehnologije u polju biologije, medicine i u industriji hrane. Uspješno je primijenjenna za dekontaminaciju bakterijskih patogena u hrani (Salmonella typhimurium, Staphylococcus aureus, Escherichia coli, Listeria monocytogenes i druge) te štetnih tvari poput fenola, pesticida i slično (Doubla i sur., 2007., Heo i sur., 2014., Misra, 2015., Min i sur., 2016.). Tehnologija CAP uspješno je primijenjena za inaktivaciju
plijesni rodova Aspergillus, Cladosporium, Penicillium, Fusarium i Alternaria na žitaricama, orašastim plovima i drugoj hrani te je uočeno i značajno (više od 50%) sma-njenje količine aflatoksina, fumonizina, deoksinivalenola, zearalenona i ohratoksina (Basaran i sur., 2008., Selcuk i sur., 2008., Ito i sur., 2012., Pignata i sur., 2014., Ouf i sur., 2015., Park i Ha., 2015., Dasan i sur., 2016, ten Bosch i sur., 2017.). Kemijske metode dekontaminacije podrazumijevaju pri-mjenu različitih kemijskih tvari u cilju inaktivacije miko-toksina. Dekontaminacija je s više ili manje uspjeha po-stignuta s kiselinama (fosforna, limunska, octena, tartarna i mliječna kiselina) i bazama (amonijak, kalcijev i natrijev hidroksid), oksidirajućim tvarima (vodikov peroksid, ozon), reducirajućim tvarima (bisulfiti, šećeri), klorirajućim tva-rima (klor), solima (natrijev hipoklorit, natrijev bikarbonat), formaldehidom i drugim. Za inaktivaciju aflatoksina uspješ-nim su se pokazali amonijak i natrijev bisulfit te limunska, octena i mliječna kiselina u postupcima koji simuliraju ku-hanje. Oksidirajući agensi poput ozona uspješno su razorili strukturu aflatoksina stvarajući spojeve slabije toksičnosti. Na ohratoksin su djelovali amonijak, natrijev hipoklorit, mravlja, propionska i sorbinska kiselina. Fumonizini su re-ducirani primjenom natrij hidrogen karobonata, kalijeva hidroksida i vodikovog peroksida, dok su trikotecene inaktivirali kalcij-hidroksid monometilamin i natrij bisulfit (Chełkowski i sur., 1981., Trenholm i sur., 1992., McKenzie i sur., 1997., Palencia i sur., 2003., Inan i sur., 2007., Lee i sur., 2015., Aiko i sur., 2016., Verheecke i sur., 2016.). Uz navedene tvari u praksi su se koristile i karboksilne kiseline kao konzervansi u hrani, jer uspješno inhibiraju rast plijesni. Budući da kemijske tvari mogu uzrokovati promjenu struk-ture mikotoksina, razina toksičnosti treba se provjeriti nakon tretiranja. U praksi je dozvoljena uporaba samo onih ke-mikalija koje su dozvoljene zakonskim propisima (Codex Alimentarius General Standards for Food Additives i ostali zakoni o hrani). Zakonski propisi dozvoljavaju tretiranje hrane za životinje kiselinama, lužinama, reducirajućim i oksidirajućim agensima. Oni se najčešće koriste za smanje-nje količine mikotoksina postupcima imerzije i fumigacije. Uz navedeno, ne smije se zaboraviti kako tretiranje kemi-kalijama može promijeniti i hranidbeni sastav te je stoga isti potrebno provjeriti nakon tretiranja.
Biološke metode. Biološka detoksifikacija mikotoksina po-kazala se uspješnom u uvjetima umjerene klime okoliša. Detoksifikacija u ovom slučaju podrazumijeva djelovanje mikroorganizama na molekule mikotoksina i proizvodnju metabolita slabije toksičnosti. Pojedine bakterije, kvasci i plijesni proizvode enzime koji imaju sposobnost inaktivacije mikotoksina. Primjerice, plijesni roda Aspergillus, Eurotium rubrum, Phoma sp., Penicillium raistrickii, Dactylium den-droiedes, Mucor spp., Absidia ripens, Trichoderma viridae, Rhizopus spp. i druge mogu inaktivirati aflatoksine. Isto tako, aflatoksine mogu inaktivirati neke vrste bakterija kao što su Nocardia corynebacterioides, Nocardia asteroides,

INAKTIVACIJA MIKOTOKSINA U HRANI ZA ŽIVOTINJE 64
Corynebacterium rubrum, Rhodococcus arythropolis, Myco-bacterium spp., Pseudomonas aeruginosa, Bacillus spp., s učinkovitošću od 28,7% do gotovo 99% (Ciegler i sur., 1966., Vekiru i sur., 2010., Popiel i sur., 2014., Ji i sur., 2016.). Fumonizine mogu inaktivirati bakterije roda Sphingomonas, a zearalenon plijesan Clonostachys rosea. Slični rezultati zabilježeni su i pri fermentaciji, jer pritom pojedine bakterije i plijesni mijenjaju strukturu hrane te ujedno inaktiviraju pojedine mikotoksine. Primjerice, bakte-rije rodova Lactobacillus, Propionibacterium, Lactococcus i Bifidobacterium (bakterije mliječno-kiselinog vrenja) mogu inaktivirati aflatoksine u fermentiranim proizvodima i kruhu (Moss i Long, 2002., Heinl i sur., 2010., Popiel i sur., 2014., Ahlberg i sur., 2015., Saladino i sur., 2016.). Ispitivanja učinkovitosti kvasca Saccharomyces cervisiae u inaktivaciji aflatoksina i ohratoksina rezultirala su proizvodnjom i komercijalnih adsorbensa mikotoksina koji s više ili manje uspjeha mogu djelovati na većinu mikotoksina u hrani za životinje (Moss i Long, 2002., Alberts i sur., 2006., Topcu i sur., 2010., Petruzzi i sur., 2014., He i sur., 2016.).
Dodaci hrani
Uporaba organskih i anorganskih tvari koje vežu miko-toksine može se opisati kao fizikalno-kemijska metoda smanjenja koncentracije mikotoksina. Početna istraživanja pokazala su se jako uspješnima, ali zbog istodobnog vezivanja vitamina i minerala, kao i umanjenog djelovanja nekih antimikrobnih tvari rezultati nisu zadovoljavajući (Huwig i sur., 2001., Kolosova i Stroka, 2011., De Mil i sur., 2015.). Filosilikatne gline su u istraživanju Phillipsa i sur. (1988.) uspješno vezale aflatoksine. U tu svrhu ispitivani su i aluminosilikati, bentoniti i slično. Uspješnost apsorpcije drugih mikotoksina znatno je manja. Dodavanje aditiva biljnog podrijetla (začini, biljke) koristi se tijekom obrade hrane. Primjerice, izotiocijanati su uspješno inhibirali rast plijesni roda Aspergillus te smanjili količinu aflatoksina u brašnu (>80%) (Aiko i Mehta, 2015., Hontanaya i sur., 2015.). Na smanjenje količine aflatoksina također je djelovala i primjena ekstrakta biljaka Azadirachta indica („neem“), Ocimum tenuiflorum i Adhatoda, pričem je uspješnost aktivacije rasla s porastom temperature (Panda i Mehta., 2013., Vijayanandraj i sur., 2014., Aiko i Mehta., 2015., Sultana i sur., 2015.). Primjena reducirajućih šećera te nekih enzima (amilaze, proteaze, ksilanaze, peptidaze, laktaze) pokazala se uspješnom u smanjenju količine fumonizina, deoksinivalenola i ohratoksina A (Fernandez-Surumay i sur., 2005., Abrunhosa i sur., 2010., Scarpari i sur., 2014., Vanhoutte i sur., 2016.). Iako primjena enzima djeluje obećavajuće, zbog proteinske naravi postoji opasnost alergijeske reakcije te je stoga prije njihove primjene neophodno provjeriti mogu li izazvati neželjeni učinak kao alergeni (EFSA, 2009.). Posebnu skupinu dodataka hrani čine brojni komercijalni proizvodi za vezivanje mikotoksina koji se vrlo često i koriste u praksi. Isti su razvijeni na temelju poznavanja toksikogenosti plijesni, metabolizma mikotoksina, olakšavajući uzgajivačima životinja osiguranje odgovarajućih proizvodnih rezultata i zdravlje životinja.
Zaključci
U hrani za životinje najčešće su prisutne plijesni rodova Aspergillus, Penicillium, Fusarium, dok su druge plijesni rjeđe zastupljene. Od mikotoksina, najznačajniji su afla-toksini, ohratoksin A, fumonizin, zearalenon i trikoteceni. U slučaju da neka hrana za životinje sadrži količine nepoželjnih tvari koje prekoračuju najvišu dopuštenu količinu definiranu zakonskim propisima, ona se ne smije miješati s istim ili drugim proizvodima namijenjenim za hranu za životinje kako bi se „razrijedila“ količina miko-toksina u konačnom proizvodu. Međutim, zbog globalnih klimatskih promjena i njihovog utjecaja na poljoprivredu i uzgoj životinja te spomenutih zahtjeva tržišta kontaminirana sirovina (u količinama iznad dozvoljenih vrijednosti pro-pisanih zakonom) često je jedini izbor. Proizvođači hrane moraju stoga prvenstveno otkriti izvor kontaminacije te prema potrebi i mogućnostima primijeniti odgovarajuće postupke dekontaminacije. Mikotoksini su u pravilu vrlo stabilne tvari i ne uništavaju ih standadni postupci obrade i prerade hrane. Ipak, neki po-stupci i primjena temperatura mogu utjecati na količinu toksina u hrani. Za dekontaminaciju krmiva koriste se fizikalne, kemijske i mikrobiološke metode, dok se štetni učinak u organizmu nastoji prevenirati dodacima hrani. U praksi, najčešći je odabir primjena svih dostupnih preven-tivnih mjera te kombinacija fizikalnih, kemijskih i bioloških metoda koja s većim ili manjim uspjehom uz prihvatljiv ekonomski trošak može umanjiti nepoželjne posljedice kon-taminacije mikotoksinima. Cilj dobre proizvođačke prakse je proizvodnja zdrave, nepatvorene hrane koja ne predstavlja opasnost za zdravlje ljudi, životinja i/ili okoliša.
Literatura
ABRUNHOSA, L., R. R. M. PATERSON, A. VENÂNCIO (2010): Biodegradation of ochratoxin A for food and feed decontamination. Toxins., 2 (5), 1078–1099.
AHLBERG, S. H., V. JOUTSJOKI, J. J. KORHONEN (2015): Potential of lactic acid bacteria in aflatoxin risk mitigation. Int. J. Food. Microbiol. 207, 87-102.
AIKO, V. A. MEHTA (2015): Occurence and detoxification of mycotoxins. J. Biosci., 40 (5), 943-954.
AIKO, V. P. EDAMANA, A. MEHTA (2016): Decomposition and detoxification of aflatoxin B1 by lactic acid. J. Sci. Food Agric., 96 (6), 1959-1966.
ALANDER, J. T., V. BOCHKO, B. MARTINKAUPPI, S. SARANWONG, T. MANTERE (2013): A review of optical nondestructive visual and near-infrared methods for food quality and safety. Int. J. Spectr. https://www.hindawi.com/journals/ijs/2013/341402/abs/ [30.03.2019.]
ALBERTS, J. F., Y. ENGELBRECHT, P. S. STEYN, W. HOLZAPFEL, W. H. VAN ZYL (2006): Biological degradation of aflatoxin B1 by Rhodococcus erythropolis cultures. Int, J, Food Microbiol., 109 (1-2), 121-126.
ALDRED, D., N. MAGAN (2004): Prevention strategies for trichothecenes. Toxicol. Lett., 153 (1), 165-171.

INAKTIVACIJA MIKOTOKSINA U HRANI ZA ŽIVOTINJE 65
ALY, S. E. (2002): Distribution of aflatoxins in product and by-products during glucose production from contaminated corn. Nahrung., 46 (5), 341-344.
ATALLA, M. M., N. M. HASSANEIN, A. A. EL-BEIH, Y. A. YOUSSEF (2004): Effect of fluorescent and UV light on mycotoxin production under different relative humidities in wheat grains. Int. J. Agric. Biol., 6, 1006-1012.
BASAPPA, S. C., T. SHANTHA (1996): Methods for detoxi-fication of aflatoxins in foods and feeds - a critical appraisal. J. Food Sci. Technol., 33 (2), 95-107.
BASARAN,P. N. BASARAN-AKGUL, L. OKSUZ (2008): Eli-mination of Aspergillus parasiticus from nut surface with low pressure cold plasma (LPCP) treatment. Food Microbiol., 25 (4), 626-632.
BOTHAST, R. J., C. W. HESSELTINE (1975): Bright greenish-yellow fluorescence and aflatoxin in agricultural commodities. Appl. Microbiol., 30 (2), 337-338.
BRETZ, M., M. BEYER, B. CRAMER, A. KNECHT, H. U. HUMPF (2006): Thermal degradation of the Fusarium mycotoxin deoxynivalenol. J. Agric. Food Chem., 54 (17), 6445-6451.
BRYDEN, W. L. (2017). Mycotoxins sin the food chain: human health implications. Asia. Pac. J. Clin. Nutr., 16 (1), 95-101.
BRYŁA, M., A. WASKIEWICZ, G. PODOLSKA, K, SZYMCZYK, R. JEDRZEJCZAK, K. DAMAZIAK, A. SUŁEK (2016): A. Occurrence of 26 Mycotoxins in the grain of cereals cultivated in Poland. Toxins., 8 (6), E160.
BULLERMAN, L. B., A. BIANCHINI (2007): Stability of mycotoxins during food processing. Int. J. Food Microbiol., 119 (1-2), 140-146.
CASTELLS, M., A. J. RAMOS, V. SANCHIS, S. MARÍN (2007): Distribution of total aflatoxins in milled fractions of hulled rice. J .Agric. Food Chem., 55 (7), 2760-2764.
CHELI, F., L. PINOTTI, L. ROSSI, V. DELL’ORTO (2013): Effect of milling procedures onmycotoxin distribution in wheat fractions: a review. LWT – Food Sci. Technol., 54 (2), 307-314.
CHEŁKOWSKI J, GOLIŃSKI P, GODLEWSKA B, RADO-MYSKA W, SZEBIOTKO K, WIEWIÓROWSKA M (1981) Mycotoxins in cereal grain. Part IV. Inactivation of ochratoxin A and other mycotoxins during ammoniation. Nahrung., 25 (7) M, 631-637.
CHULZE, S. N. (2010). Strategies to reduce mycotoxin levels in maize during storage: a review. Food Addit. Contam. Part A. Chem. Anal. Control Expo. Risk Assess., 27 (5), 651-657.
CIEGLER, A., E. B. LILLEHOJ, R. E. PETERSON, H. H. HALL (1966): Microbial detoxification of aflatoxin. Appl. Micro-biol., 14 (6), 934-939.
CODEX ALIMENTARIUS (2015): General Standard for Food Additives (GSFA) www.fao.org/gsfaonline/docs/ /CXS_192e.pdf [30.03.2019.]
Commission Recommendation of 17 August 2006 on the pre-vention and reduction of Fusarium toxins in cereals and cereal products (2006/583/EC).
CONWAY, H. F., R. A. ANDERSON, E. B. BAGLEY (1978): Detoxification of aflatoxin-contaminated corn by roasting. Cereal Chem., 55, 115-117.
DASAN, B. G., I. H. BOYACI, M. MUTLU (2016): Inactivation of aflatoxigenic fungi (Aspergillus spp.) on granular food model, maize, in an atmospheric pressure fluidized bed plasma system. Food Control., 70, 1-8.
DE MIL, T. M. DEVREESE, N. BROEKAERT, S. FRAEYMAN, P. DE BACKER, S. CROUBELS (2015): In Vitro Adsorption and in Vivo Pharmacokinetic Interaction between Doxycycline and Frequently used Mycotoxin Binders in Broiler Chickens. J. Agric. Food Chem., 63 (17), 4370-4375.
Directive 2002/32/EC of 7 May 2002 on undesirable substances in animal feed. Off. J. Eur. Communities., 140, 10-21.
DOUBLA, A., S. LAMINSI, S. NZALI, E. NJOYIM, J. KAMSU-KOM, J. L. BRISSET (2007): Organic pollutants abatement and biodecontamination of brewery effluents by a non-thermal quenched plasma at atmospheric pressure. Chemosphere., 69 (2), 332-337.
EDWARDS, S. G. (2004) Influence of agricultural practices on Fusarium infection of cereals and subsequent contamination of grain by trichothecene mycotoxins. Toxicol. Lett., 153 (1), 29-36.
EECKHOUT, M., G. HAESAERT, N. LANDSCHOOT, S. DESCHUYFFELEER, S. DE LAETHAUWER (2013): Guidelines for prevention and control of mould growth and mycotoxin production in cereals. Annex I to D8.1. Report Guidelines on Prevention Measures. Mycohunt project. www.mycohunt.eu [30.03.2019.]
EFSA, European Food Safety Authority (2009): Guidance on the submission of a Dossier on food enzymes for safety evaluation by the scientific panel of food contact material, enzymes, flavourings and processing aids. EFSA J., 1305:1-26.
EFSA, European Food Safety Authority. Panel on Contaminants in the Food Chain (CONTAM) (2014): EFSA scientific opinion on the risks for human and animal health related to the presence of modified forms of certain mycotoxins in food and feed. EFSA J 12:3916.
European Commission. Recommendation 2006/576/EC of 17 august 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products inten-ded for animal feeding. Off. J. Eur. Union., 229, 7-9.
European Commission. Recommendation 2013/165/EU of 27 March 2013 on the presence of T-2 and HT-2 toxin in cereals and cereal products. Off. J. Eur. Union., 91, 12-15.
European Commission. Regulation (EC) no 1881/2006 of 19 de-cember 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union., 364, 5-24.
FAO, Food and Agriculture Organization of the United Nations (2001): Manual on the Application of the HACCP System in Mycotoxin Prevention and Control. Rome: Food and Agri-culture Organization of the United Nations (FAO Food and Nutrition Paper No. 73). http://www.fao.org/docrep/005/ /y1390e/y1390e00.htm. [30.03.2019.]
FAO, Food and Agriculture Organization of the United Nations (2002). Good Agricultural Practices, second version. Rome: Food and Agriculture Organization of the United Nations. http://www.fao.org/ag/magazine/GAP-V2-June02.pdf. [30.03.2019.]
FERNANDEZ-SURUMAY, G., G. D. OSWEILER, M. J. YAEGER, G. E. ROTTINGHAUS, S. HENDRICH, L. K. BUCKLEY, P. A. MURPHY (2005): Fumonisin B glucose reaction products are less toxic when fed to swine. J. Agric. Food Chem., 53 (10), 4264-4271.
HAHN, I., V. NAGL, H. E. SCHWARTZ-ZIMMERMANN, E. VARGA, C. SCHWARZ, V, SLAVIK, N. REISINGER, A. MALACHOVA, M. SIRLINI, S. GENEROTTI, C. DALL'ASTA, R. KRSKA, W. D. MOLL, F. BERTHILLER (2015): Effects of orally administered fumonisin B1 (FB1),

INAKTIVACIJA MIKOTOKSINA U HRANI ZA ŽIVOTINJE 66
partially hydrolysed FB1, hydrolysed FB1 and N-(1-deoxy-D-fructos-1-yl) FB1 on the sphingolipid metabolism in rats. Food Chem. Toxicol., 76, 11-18.
HE, J. W., Y. I. HASSAN, N. PERILLA X. Z. LI, G. J. BOLAND, T. ZHOU (2016): Bacterial epimerization as a route for deoxynivalenol detoxification: the influence of growth and environmental conditions. Front. Microbiol., 7, 572.
HEINL, S., D. HARTINGER, M. THAMHESL, E. VEKIRU, R. KRSKA, G. SCHATZMAYR, W. D. MOLL, R. GRABHERR (2010): Degradation of fumonisin B1 by the consecutive action of two bacterial enzymes. J. Biotechnol., 145 (2), 120-129.
HEO, N.S., M. K. LEE, G. W. KIM, S. J. LEE, J. Y. PARK, T. J. PARK (2014): Microbial inactivation and pesticide removal by remote exposure of atmospheric air plasma in confined environments. J. Biosci. Bioeng., 117 (1), 81-85.
HERZALLAH, S., K. ALSHAWABKEH, A. AL FATAFTAH (2008): Aflatoxin decontamination of artificially contaminated feeds by sunlight, γ-radiation, and microwave heating. J. Appl. Poult. Res., 17 (4), 515-520.
HOJNIK, N., U. CVELBAR, G. TAVČAR-KALCHER, J. L. WALSH, I. KRIŽAJ (2017): Mycotoxin Decontamination of Food: Cold Atmospheric Pressure Plasma versus „Classic“ Decontamination. Toxins (Basel)., 9 (5), 151, 1-19
HONTANAYA, C., G. MECA, F. B. LUCIANO, J. MANES, G. FONT (2015): Inhibition of aflatoxin B1, B2, G1, and G2 production by Aspergillus parasiticus in nuts using yellow and oriental mustard flours. Food Control., 47, 154-160.
HUWIG A., S. FREIMUND, O. KAPPELI, H. DUTLER (2001): Mycotoxin detoxication of animal feed by different adsor-bents. Toxicol. Lett., 122 (2), 179-188.
INAN, F., M. PALA, I. DOYMAZ (2007); Use of ozone in detoxification of aflatoxin B1 in red pepper. J. Stored Prod. Res., 43 (4), 425-429.
ITO, M., I. SATO, M. ISHIZAKA, S. YOSHIDA, M. KOITABASHI, S. YOSHIDA, S. TSUSHIMA (2013):, Bac-terial cytochrome P450 system catabolizing the Fusarium toxin deoxynivalenol. Appl. Environ. Microbiol., 79 (5), 1619-1628.
JACOBSEN, B. J. (2014): Good agricultural and harvest practices to reduce mycotoxin contamination in wheat in temperate countries. In: Leslie JF, Logrieco AF (Ees) Mycotoxin reduction in grain chains. Wiley Blackwell, New Delhi, pp. 209-219
JI, C., Y. FAN, L. ZHAO (2016): Review on biological degradation of mycotoxins. Anim. Nutr., 2 (3), 127-133.
KABAK, B., A. D. DOBSON, I. VAR (2006): Strategies to prevent mycotoxin contamination of food and animal feed. Crit. Rev. Food Sci. Nutr., 46 (8), 593-619.
KARLOVSKY, P., M. SUMAN, F. BERTHILLER, J. DE MEESTER, G. EISENBRAND, i. PERRIN, I. P. OSWALD, G. SPEIJERS, A. CHIODINI, T. RECKER, P DUSSORT (2016): Impact of food processing and detoxification treatments on mycotoxin contamination. Mycotoxin Res., 32 (4), 179-205.
KHATIBI, P. A., G. BERGER, J. WILSON, W. S. BROOKS, N. MCMASTER, C. A. GRIFFEY, K. B. HICKS, N. P. NGHIEM, D. G. SCHMALE 3RD (2014): A comparison of two milling strategies to reduce the mycotoxin deoxynivalenol in barley. J Agric. Food Chem., 62 (18), 4204-13.
KILONZO, R. M., J. K. IMUNGI, W. M. MUIRU, P. O. LAMUKA, P. M. NJAGE (2014): Household dietary exposure
to aflatoxins from maize and maize products in Kenya. Food Addit. Contam., 31 (12), 2055-2062.
KLICH, M. A. (2007): Environmental and developmental factors influencing aflatoxin production by Aspergillus flavus and Aspergillus parasiticus. Mycoscience, 48 (2),71-80.
KOLOSOVA, A., J. STROKA (2011).:Substances for reduction of the contamination of feed by mycotoxins: a review. World Mycotoxin J., 4 (3), 225-256.
KÖPPEN, R., J. RIEDEL, M. PROSKE, S. DRZYMALA, T. RASENKO, V. DURMAZ, M. WEBER, M. KOCH (2012): Photochemical trans-/cis-isomerization and quantitation of zearalenone in edible oils. J Agric. Food Chem., 60 (47)M, 11733-11740.
LEE, J., J. Y. HER, K. G. LEE (2015): Reduction of aflatoxins (B1, B2, G1, and G2) in soybean-based model systems. Food Chem., 189, 45-51.
LEFYEDI, M. L., J. R. N. TAYLOR (2006): Effect of dilute alkaline steeping on the microbial contamination, toxicity and diastatic power of sorghum malt. J Inst Brew., 112, 108-116.
LEVASSEUR-GARCIA, C. (2018): Updated Overview of Infrared Spectroscopy Methods for Detecting Mycotoxins on Cereals (Corn, Wheat, and Barley. Toxins (Basel), 10 (1), 38.
MANNAA, M. , K. D. KIM (2017): Influence of Temperature and Water Activity on Deleterious Fungi and Mycotoxin Pro-duction during Grain Storage. Mycobiology, 45 (4), 240-254.
MARIN, S., A. J. RAMOS, G. CANO-SANCHO, V. SANCHIS (2013): Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol., 60, 218-237.
MATUMBA, L., C. VAN POUCKE, E. NJUMBE EDIAGE, B. JACOBS, S. DE SAEGER (2015): Effectiveness of hand sorting, flotation/washing, dehulling and combinations thereof on the decontamination of mycotoxin-contaminated white maize. Food Addit. Contam. Part A. Chem. Anal. Control Expo. Risk. Assess., 32 (6), 960-969.
McKENZIE, K. S., A. B. SARR, K. MAYURA, R. H. BAILEY, D. R. MILLER, T. D. ROGERS, W. P. NORRED, K. A. VOSS, R. D. PLATTNER, L. F. KUBENA, T. D. PHILLIPS (1997): Oxidative degradation and detoxification of myco-toxins using a novel source of ozone. Food Chem. Toxicol., 35 (8), 807-820.
McMULLIN, D., B. MIZAIKOFF, R. KRSKA (2015): Advance-ments in IR spectroscopic approaches for the determination of fungal derived contaminations in food crops. Anal. Bioanal. Chem., 407 (3), 653-660.
MESTERHAZY, A. (2014): Breeding for resistance to Fusarium head blight in wheat. In: J. F: LESLIE, A. F. LOGRIECO (Eds) Mycotoxin reduction in grain chains. Wiley Blackwell, New Delhi, pp. 189-208.
MIN, S.C., S. H. ROH, B. A. NIEMIRA, J. E. SITES, G. BOYD, A. LACOMBE (2016): Dielectric barrier discharge atmos-pheric cold plasma inhibits Escherichia coli o157:H7, Salmo-nella, Listeria monocytogenes, and tulane virus in romaine lettuce. Int. J. Food Microbiol., 237, 114-120.
MIRAGLIA, M., H. J. P. MARVIN, G. A. KLETER, P. BATTILANI, C. BRERA, E. CONI, F. CUBADDA, L. CROCI, B. DE SANTIS, S. DEKKERS, L. FILIPPI, R. W. HUTIES, M. Y. NOORDAM, M. PISANTE, G. PIVA, A. PRANDINI, L. TOTI, G. J. VAN DEN BORN, A. VESPERMANN (2009): Climate change and food safety: An emerging issue with special focus on europe. Food Chem. Toxicol., 47 (5), 1009-1021.

INAKTIVACIJA MIKOTOKSINA U HRANI ZA ŽIVOTINJE 67
MISRA, N. N. (2015): The contribution of non-thermal and advanced oxidation technologies towards dissipation of pesticide residues. Trends Food Sci. Technol., 45 (2), 229-244.
MOSS, M. O., M. T. LONG (2002): Fate of patulin in the presence of the yeast Saccharomyces cerevisiae. Food Add. Contam., 19 (4), 387-399.
OLIVEIRA, G., D. M. DA SILVA, R. G. F. A. PEREIRA, L. C. PAIVA, G. PRADO, L. R. BATISTA (2013). Effect of different roasting levels and particle sizes on ochratoxin a concentration in coffee beans. Food Control., 34 (2), 651-656.
OMOTAYO, O. P., A. O. OMOTAYE, M. MWANZA, O. O. BABALOLA (2019): Prevalence of Mycotoxins and Their Consequences on Human Health. Toxicol. Res., 35 (1), 1-7.
OUF, S. A., A. H. BASHER, A. A. MOHAMED (2015): Inhibitory effect of double atmospheric pressure argon cold plasma on spores andmycotoxin production of Aspergillus niger contaminating date palm fruits. J. Sci. Food Agric., 95 (15), 3204-3210.
PALENCIA, E., O. TORRES, W. HAGLER, F. I. MEREDITH, L. D. WILLIAMS, R. T. RILEY (2003): Total fumonisins are reduced in tortillas using the traditional nixtamalization method of mayan communities. J. Nutr., 133 (10), 3200-3203.
PANDA, P., A. MEHTA (2013): Aflatoxin detoxification potential of Ocimum tenuiflorum. J, Food Saf, 33 (3), 265-272.
PARK, S. Y., S. D. HA (2015): Application of cold oxygen plasma for the reduction of Cladosporium cladosporioides and Peni-cillium citrinum on the surface of dried filefish (Stephanolepis cirrhifer) fillets. Int. J. Food Sci. Technol., 50 (4), 966-973.
PESTKA, J. J. (2007): Deoxynivalenol: toxicity, mechanisms and animal health risks. Anim. Feed Sci. Technol., 137 (3-4), 283-298.
PETRUZZI, L., A. BEVILACQUA, A. BAIANO, L. BENEDUCE, M. R. CORBO, M. SINIGAGLIA (2014): In vitro removal of ochratoxin a by two strains of Saccharomyces cerevisiae and their performances under fermentative and stressing conditions. J. Appl. Microbiol., 116 (1), 60-70.
PHILLIPS, T. D., L. F. KUBENA, R. B. HARVEY, D. R. TAYLOR, N. D. HEIDELBAUGH (1988): Hydrated sodium calcium aluminosilicate: High affinity sorbent for aflatoxin. Poult. Sci., 67 (2), 243-247.
PIGNATA, C., D. D’ANGELO, D. BASSO, M. C. CAVALLERO, S. BENEVENTI, D. TARTARO, V. MEINERI, G. GILLI (2014): Low-temperature, low-pressure gas plasma application on Aspergillus brasiliensis, Escherichia coli and pistachios. J. Appl. Microbiol., 116 (5), 1137-1148.
POPIEL, D., G. KOCZYK, A. DAWIDZIUK, K. GROMADZKA, L. BLASZCZYK, J. CHELKOWSKI (2014): Zearalenone lactone hydrolase activity in Hypocreales and its evolutionary relationships within the epoxide hydrolase subset of a/b-hydrolases. BMC Microbiol., 14:82.
Provedbena uredba komisije (EU) 2015/949 оd 19. lipnja 2015. o odobrenju pregleda određene hrane koje određene treće zemlje provode prije izvoza u pogledu prisutnosti određenih miko-toksina.
RATERS, M., R. MATISSEK (2008): Thermal stability of afla-toxin B1 and ochratoxin A. Mycotoxin Res., 24 (3), 130-134.
REFAI, M. K., N. H. AZIZ, F. EL-FAR, A.A. HASSAN (1996): Detection of ochratoxin produced by Aspergillus ochraceus in feedstuffs and its control by -radiation. Appl. Radiat. Isot., 47 (7), 617-621.
SALADINO, F., C. LUZ, L. MANYES, M. FERNANDEZ-FRANZ, G. MECA (2016): In vitro antifungal activity of lactic acid bacteria against mycotoxigenic fungi and their
application in loaf bread shelf life improvement. Food Control, 67, 273-277.
SCARPARI, M., C. BELLO, C. PIETRICOLA, M. ZACCARIA, L. BERTOCCHI, A. ANGELUCCI, M. R. RICCIARDI, V. SCALA, A. PARRONI, A. A. FABBRI, M. REVERBERI, S. ZIALIC, C. FANELLI (2014): Aflatoxin control in maize by Trametes versicolor. Toxins, 6 (12), 3426-3437.
SCHWAKE-ANDUSCHUS, C., M. PROSKE, E. SCIURBA, K. MUENZING, M. KOCH, r. MAUL (2015): Distribution of deoxynivalenol, zearalenone, and their respective modified analogues in milling fractions of naturally contaminated wheat grains. World Mycotoxin J., 8 (4), 433-443.
SCUDAMORE, K. A., R. C. E. GUY, B. KELLEHER, S. J. MACDONALD (2008): Fate of the fusarium mycotoxins, deoxynivalenol, nivalenol and zearalenone, during extrusion of wholemeal wheat grain. Food Addit. Contam. Part A., 25 (11), 331-337.
SCUDAMORE, K. AA, J. N. BANKS, R. C. E. GUY (2004): Fate of ochratoxin a in the processing of whole wheat grain during extrusion. Food Addit Contam., 21 (5), 488-497.
SCUDAMORE, K. A. (2005):. Principles and Applications of Mycotoxin Analysis. The Mycotoxin Blue Book, Edited by Duerte Diaz, 157-185, Nottingham University Press.
SELCUK, M., L. OKSUZ, P. BASARAN (2008): Deconta-mination of grains and legumes infected with Aspergillus spp. and Penicillum spp. by cold plasma treatment. Bioresour. Technol., 99 (11), 5104-5109.
SOKOLOVIĆ, M., V. GARAJ-VRHOVAC, B. ŠIMPRAGA (2008): T-2 toxin: incidence and toxicity in poultry. Arh. Hig. Rada. Tokiskol., 59 (1), 43-52.
STREIT, E., G. SCHATZMAYR, P. TASSIS, E. TZIKA, D. MARIN, I. TARANU, C. TABUC, A. NICOLAU, I. APRODU, O. PUEL, I. P. OSWALD (2012): Current Situation of Mycotoxin Contamination and Co-occurence in Animal Feed - Focus on Europe. Toxins (Basel)., 4 (10), 788-809.
SULTANA, B., R. NASEER, P. NIGAM (2015): Utilization of agro-wastes to inhibit aflatoxins synthesis by Aspergillus parasiticus: a biotreatment of three cereals for safe long-term storage. Bioresour Technol .,197, 443-450.
SYDENHAM, E. W., L. VAN DER WESTHUIZEN, S. STOCKENSTROM, G. S. SHEPHARD, P. G. THIEL (1994): Fumonisin-contaminated maize: physical treatment for the partial decontamination of bulk shipments. Food Addit. Contam., 11 (1), 25-32.
TEN BOSCH, L., K. PFOHL, G. AVRAMIDIS, S. WIENEKE, W. VIÖL, P. KARLOVSKY (2017): Plasma-based degradation of mycotoxins produced by Fusarium, Aspergillus and Alternaria species. Toxins, 9 (3), 97.
TIBOLA, C. S., J. M. CUNHA FERNANDES, E. M. GUARIENTI, M. NICOLAU (2015): Distribution of Fusarium mycotoxins in wheat milling process. Food Control, 53, 91-95.
TOPCU, A., T. BULAT, R. WISHAH, I. H. BOYACI (2010): Detoxification of aflatoxin B1 and patulin by Enterococcus faecium strains. Int. J. Food Microbiol., 139 (3), 202-205.
TRENHOLM, H. L., L. L. CHARMLEY, D. B. PRELUSKY, R. M. WARNER (1992): Washing procedures using water or sodium carbonate solutions for the decontamination of three cereals contaminated with deoxynivalenol and zearalenone. J. Agric. Food Chem., 40 (11), 2147-2151.
TRENHOLM, H. L., L. L. CHARMLEY, D. B. PRELUSKY, R. M. WARNER (1991): Two physical methods for the deconta-

INAKTIVACIJA MIKOTOKSINA U HRANI ZA ŽIVOTINJE 68
mination of four cereals contaminated with deoxynivalenol and zearalenone. J. Agric. Food Chem., 39 (2), 356-360.
TRIVEDE, A. B., E. DOI, N. KITABATAKE (1992): Cytotoxicity of citrinin heated at temperatures above 100 °C. Biosci. Biotech. Biochim., 56 (3), 423-426.
Uredba (EZ) br. 852/2004 Europskog Parlamenta i Vijeća od 29. travnja 2004. o higijeni hrane.
Uredba komisije (EU) br. 519/2014 оd 16. svibnja 2014. o izmjeni Uredbe (EZ) br. 401/2006 u pogledu metoda uzorkovanja veli-kih serija, začina i dodataka prehrani, kriterija učinkovitosti za toksine T-2, HT-2 i citrinin te orijentacijskih metoda analize.
Uredba komisije (EZ) br. 401/2006 od 23. veljače 2006. o utvrđivanju metoda uzorkovanja i analize za službenu kontrolu razina mikotoksina u hrani.
VAN DER WESTHUIZEN L., G. S. SHEPHARD, J. P. RHEEDER, H. M. BURGER, W. C. A. GELDERBLOM, C. P. WILD, Y. Y. GONG (2011). Optimising sorting and washing of home-grown maize to reduce fumonisin contami-nation under laboratory-controlled conditions. Food Control, 22, 396-400.
VANHOUTTE, I., K. AUDENAERT, L. DE GELDER (2016): Biodegradation of mycotoxins: tales from known and unexplored worlds. Front. Microbiol., 7, 561. PMC4843849.
VEKIRU, E., C. HAMETNER, R. MITTERBAUER, J. RECHT-HALER, G. ADAM, G. SCHATZMAYR, R. KRSKA, R. SCHUHMACHER (2010): Cleavage of zearalenone by Trichosporon mycotoxinivorans to a novel nonestrogenic metabolite. Appl . Environ. Microbiol., 76 (7), 2353-2359.
VERHEECKE, C. T. LIBOZ, F. MATHIEU (2016): Microbial degradation of aflatoxin B1: Current status and future advances. Int. J. Food. Microbiol., 21, 237, 1-9.
VIJAYANANDRAJ, S., R. BRINDA, K. KANNAN, R. ADHITHYA, S. VINOTHINI, K. SENTHIL, R. R. CHINTA, V. PARANIDHARAN, R. VELAZHAHAN (2014): Detoxi-fication of aflatoxin B1 by an aqueous extract from leaves of Adhatoda vasica Nees. Microbiol. Res., 169 (4), 294-300.
VISCONTI, A., M. SOLFRIZZO, M. B. DOKO, A. BOENKE, M. PASCALE (1996): Stability of fumonisins at different storage periods and temperatures in gamma-irradiated maize. Food Addit. Contam., 13 (8), 929-938.
INACTIVATION OF MYCOTOXINS IN FEED
Summary
Pathogenic moulds and their secondary metabolites, mycotoxins, are common contaminants of raw materials, feed and food. Certain mycotoxins are toxic for humans and animals at a relatively low concentration. Therefore, their incidence in food can have detrimental effects on productivity of animals and health of animals and humans. High occurrence of mycotoxins and episodes of intoxication in humans and animals related to mycotoxins resulted in numerous researches on the development of methods for minimizing their potential toxic effects. New studies are focused not only on the prevention of mould growth but also on different mycotoxin inactivation methods that will produce less toxic metabolites. Mycotoxins are relatively stable substances that are not destroyed during most food processing procedures. However, certain methods can transform mycotoxins into less toxic substances. Detoxification of mycotoxin by different physical, chemical and biological methods has been used most often. Their efficiency depends on the type, incidence and level of toxin contamination, as well as on co-occurrence and certain toxic interactions. In order to avoid or minimize harmful effects of mycotoxins in animals and humans, prevention of their production is still the best preventive measure. In reality, demand for high quality raw materials, economic factors and unavailability of mycotoxin-free materials results in the application of different inactivation strategies with more or less success. This paper summarizes different inactivation procedures for the most often mycotoxins that can have potential toxic effects in animals. Key words: mycotoxins, moulds, feed, inactivation

PERADARSKI DANI 2019. 69
SOJA U HRANI ZA PERAD
Marija Berendika1, Marijana Sokolović1, Tihomir Zglavnik1, Gabrijela Krivec2
1Hrvatski veterinarski institut, Centar za peradarstvo, Zagreb, Hrvatska 2Vanjski suradnik, Prilaz Gjure Deželića 16, Zagreb, Hrvatska
Sažetak
Iako je kukuruz najrasprostranjenije krmivo u hranidbi svih vrsta i dobi peradi, soja je jedna od najznačajnijih. Ovisno o kategoriji i dobi životinja ona čini 18%-30% smjese za perad. Glavna značajka joj je visok udio bjelančevina, ima dobar omjer aminokiselina najsličnijih životinjskima i bogata je mastima. Da bi hranidba domaćih životinja sojom dala zadovoljavajuće rezultate sirova soja mora se termički obraditi (tostiranjem, ekstrudiranjem ili kuhanjem). Tim fizikalnim postupkom se može smanjiti većina antinutritivnih tvari do količine koja nije štetna za hranidbu životinja i poboljšati fizikalno-kemijske karakteristike i nutritivnu vrijednost soje. Ključne riječi: soja, sirove bjelančevine, hranidba peradi, perad, hrana za životinje Uvod Većina sirovina koje se koriste u proizvodnji hrane za živo-tinje su biljnog podrijetla te kao takve predstavljaju najveći udio u hrani kao i najveći trošak same proizvodnje. Krmiva (biljnog ili životinjskog podrijetla) prvenstveno ima-ju ulogu zadovoljiti hranidbene potrebe životinja odnosno potrebe životinja za energijom, hranjivim tvarima, minera-lima, prehrambenim vlaknima i vitaminima. Mogu biti u izvornom obliku, svježa ili konzervirana ili kao proizvodi dobiveni njihovom industrijskom preradom. Krmne smjese sastoje se od najmanje dvije vrste krmiva s dodacima hrani ili bez njih. Dijele se na potpune krmne smjese koje svojim sastavom u potpunosti udovoljavaju dnevnom obroku ili dopunske krmne smjese koje imaju visok udio određenih tvari, ali su po svom sastavu dostatne za dnevni obrok jedino u kombinaciji s drugom hranom za životinje. Da bi se isplanirala dnevna potreba životinje za hranjivim sastojcima, uz poznavanje koliko ih određena životinja tre-ba, potrebno je znati i hranjivu vrijednost pojedinog krmiva odnosno ukupan sadržaj iskoristivih hranjivih tvari koje određena životinja može iskoristiti u jedinici mase krmiva. Ovisno o vremenskim uvjetima, plodnosti tla, gnojidbi, klimi, bolestima, načinu prerade i uvjetima čuvanja krmiva, količine hranjivih tvari u njima mogu odstupati s obzirom na vrijednosti u tablicama kemijskog sastava i hranjive vrijed-
nosti krmiva. Zbog navedenog, određivanjem osnovne ke-mijske analize hrane za životinje analizom kemijskog sa-stava krmiva kao što su vlaga ili voda, sirovi pepeo, sirove bjelančevine, sirova vlakna, sirove masti i nedušične ekstraktivne tvari, kao i analizom sadržaja ostalih hranjivih tvari (vitamini, minerali, aminokiseline, masne kiseline i sl.) možemo dobiti potpunu sliku stvarne hranjivosti pojedinog krmiva koje koristimo za hranidbu životinja (Grbeša, 2004.). U hranidbi peradi najčešće se koriste kukuruz, pšenica, pšenične posije, pšenično stočno brašno, ječam, raž, zob, tritikale, soja i sojina sačma, suncokretova sačma, bučina pogača, stočni grašak itd. Hrana za životinje ima ulogu podmirivanja potreba organizma za svim hranjivim tvarima koje su potrebne za određenu vrstu i dob životinje. Kvali-tetnim sirovinama utječe se na proizvodnost peradi, kakvoću konačnih proizvoda te na zdravstveno stanje same životinje.
Proizvodnja i potrošnja soje
Meso i jaja peradi su lako dostupni, jeftini i nutritivno zado-voljavajući izvori animalnih proteina, minerala i vitamina u ljudskoj prehrani. Uzgoj peradi, za razliku od uzgoja drugih vrsta životinja, traje kraće i brže se može doći do mesa zadovoljavajuće nutritivne vrijednosti. U proizvodnji peradi najveći dio otpada na hranu koja čini 60%-70% same proizvodnje (Dumanovski i sur., 2004.). U novije vrijeme sve je češća uporaba soje u proizvodnji hrane za perad. Iako je kukuruz najrasprostranjenije krmivo u hranidbi svih vrsta
Marija Berendika, dipl.ing.preh.teh., Hrvatski veterinarski institut, Centar za peradarstvo, Laboratorij za analitiku stočne hrane, Henizelova 55, 10000 Zagreb, Hrvatska; Tel.: +385 (0)1 2440-214; Fax: +385 (0)1 2441-396; E-mail: [email protected]

SOJA U HRANI ZA PERAD 70
i dobi peradi, soja je jedna od najznačajnijih. Ovisno o kategoriji i dobi životinja ona čini 18%-30% smjese za perad. Glavna značajka joj je visok udio bjelančevina, ima dobar omjer aminokiselina najsličnijih životinjskim te je bogata mastima (Kiš, 2004.). Soja (latinski: Glicine max) je biljka iz porodice mahunarki podrijetlom iz Azije i jedna je od najznačajnijih uljarica koja se koristi kako u prehrani ljudi tako i za hranidbu životinja, a u Hrvatskoj se počela uzgajati krajem 20. stoljeća (Vratarić i Sudarić, 2008.). Proizvodnja soje u Hrvatskoj u razdoblju od 2012. do 2016. godine povećavala se kao što je prikazano u tablici 1. (Državni zavod za statistiku). Prema FAO (Food and Agriculture Organization of the United Nations) u 2017. godini u svijetu je posijano 123 551 146 hektara soje, a svjetska proizvodnja je bila 352 643 548 tona. Najveći proizvođač soje je Amerika (Sjedinjene Američke Države, Brazil, Argentina) s 88,5%, zatim slijedi Azija sa 7,5%, Europa s 3% i Afrika s 0,9% svjetske proizvodnje. Kao što je prikazano u slici 1., u razdoblju od 2013. do 2017. godine sijanje i proizvodnja soje u svijetu kontinuirano raste.
Soja kao krmivo
Ovisno o uvjetima uzgoja i sortnim osobinama, sojino zrno može sadržavati 35%-50% bjelančevina, 18%-24% ulja,
30% i više ugljikohidrata, oko 5% pepela minerala (kalij, fosfor, sumpor, kalcij, željezo, magnezij, natrij), kao i vita-min A, vitamine B-kompleksa, D, E i K (Vratarić i Suda-rić, 2008.). Preradom sojinog zrna može se dobiti sojino ulje, sojina sačma, sojina pogača, sojino brašno, tekstu-rirani bjelančevinasti koncentrati soje te izolati koji se koriste u ljudskoj prehrani te u hranidbi životinja, kao i za sirovine u prehrambenoj industriji. Toplinski obrađena soja je soja iz koje nije uklonjeno ulje. Ispravnom termičkom obradom inaktivirane su antinutri-tivne tvari, a u isto vrijeme dobiven je proizvod bogat uljem, bjelančevinama i esencijalnim masnim kiselinama i ener-gijom. Zbog visokog udjela masti takav proizvod je sklon kvarenju. Sojina sačma dobiva se zagrijavanjem ostataka oljuštenog ili djelomice oljuštenog zrna soje, nakon prešanja i ekstrakcije ulja. Ove dvije vrste sojine sačme razlikuju se u nutritivnom sastavu, ali sadrže visok udio bjelančevina i imaju izvrstan aminokiselinski profil, visoku ješnost i inaktivirane antinutritivne tvari. Bjelančevinasti koncentrat soje dobiva se izdvajanjem topljivih ugljikohidrata iz so-jinog brašna. Takav koncentrat sadrži oko 65% proteina (Riaz, 2011.). Sojino ulje se prvenstveno proizvodi za ljudsku prehranu, ali se upotrebljava i u hrani za životinje, posebno za perad, a kako ima dobru probavljivost, dobar je izvor energije (Banaszkiewicz, 2018.).
Tablica 1. Ukupna proizvodnja soje u Hrvatskoj (Državni zavod za statistiku) Table 1. Total soybean production in Croatia (Croatian Bureau of Statistics)
Godina / Year 2012. 2013. 2014. 2015. 2016. Proizvodnja (t) / Produ ction (tons) 93 720 111 320 131 420 196 430 244 080
Tablica 2. Udio svjetske proizvodnje soje po kontinentima (FAOSTAT, 2019)
Table 2. Soybean production share according to continents (FAOSTAT, 2019)
Godina / Year 2013. 2014. 2015. 2016. 2017.
Amerika (%) / America (%) 87,5 88,0 89,1 87,6 88,5 Azija (%) / Asia (%) 9,5 8,3 7,2 8,5 7,5 Europa (%) / Europe (%) 2,2 2,9 2,9 3,1 3,0 Afrika (%) / Africa (%) 0,7 0,8 0,8 0,7 0,9
Tablica 3. Ukupna zasijana površina i proizvodnja soje u svijetu (FAOSTAT, 2019)
Table 3. Total soybean area/production in the world (FAOSTAT, 2019)
Godina / Year
2013. 2014. 2015. 2016. 2017. Posijano (hektar) Soybean area (ha) 111 018 797 117 636 333 120 791 447 121 848 237 123 551 146
Proizvedeno (tona) / Soybean production (tons) 277 533 908 306 207 046 323 195 697 335 508 753 352 643 548

SOJA U HRANI ZA PERAD 71
Slika 1. Proizvodnja (urod) soje u svijetu od 2013. do 2017. godine. Figure 1. World soybean production (yield) 2013-2017. Najzastupljenije aminokiseline u sojinom zrnu su arginin (12%-13%), lizin (6%-7%), treonin (4%-5%), serin (5%-6%), fenilalanin (5%), izoleucin (5%), valin (4%-5%), histi-din (3%), triptofan (2%), metionin (1%), cistin (1%) itd., a od masnih kiselina linolna (53%), oleinska (23%), palmi-tinska (11%), linolenska (7%-8%), stearinska (4%), arahi-dinska (0,3%), zatim palmitooleinska, miristinska i behi-ninska po 0,1% (Vratarić i Sudarić, 2000.). U sojinom zrnu prisutne su antinutritivne tvari (termo-labilne i termostabilne) koje nisu probavljive i/ili negativno utječu na probavu i uzrokuju sporiji rast životinja: inhibitori proteaza (vežu i inaktiviraju probavne enzime gastrointesti-nalnog trakta životinje) tripsin inhibitor i kimotripsin inhi-bitor, lektini (hemoglutinini), fitoestrogeni, stahioze i rafi-noze, filati, alergeni, pektini, mikotoksini, oligosaharidi (Zhang i Parsons, 1993.). Inhibitori proteaza koji inaktivi-raju enzime gušterače tripsin i kimotripsin utječu na slabiju iskoristivost ostalih enzima koji imaju ulogu u pucanju veze manjih peptida. Time se smanjuje probavljivost proteina, a gušterača se dodatno opterećuje pokušavajući nadoknaditi enzime (mogućnost pojave hipertrofije i lezija gušterače), što posljedično dovodi do gubitka aminokiselina sa sumpo-rom (metionin i cistin) (Flavin, 1982.). Soja ima dva tripsin inhibitora, Kunitzov inhibitor i Bowman-Birkov inhibitor, naziv su dobili prema znanstve-nicima koji su ih prvi izolirali i karakterizirali. Budući da je tripsin inhibitor termolabilan, može se inaktivirati toplin-skom obradom (ne u potpunosti, ali smatra se da preostala aktivnost nema nepovoljan učinak u hranidbi). Primjerice, postupkom ekstrudiranja soje smanjuje se udio tripsin inhibitora za 97%-98% (Van der Poel, 1997.). Sirova soja sadrži lektine (hemaglutinini), proteine koji se vežu za ugljikohidrate te hranidba takvom sojom može uzrokovati sporiji rast i smrtnost životinja (Kakade i sur.,1972.), ali termičkom obradom soje djelovanje lektina također se može inaktivirati. Najčešće se za određivanje količine antinutritivnih tvari u soji koriste analize na prisutnost tripsina i ureaza test. Urea
se pomoću ureaze razgrađuje na amonijak, što može biti toksično za preživače ako se hrane sirovom sojom, ali se toplinskom obradom aktivnost ureaze smanjuje približno kao i tripsin inhibitor (Detlef, 1994.). Metoda određivanja ureaze temelji se na promjeni vrijednosti pH prilikom otpuš-tanja amonijaka. Visoke vrijednosti ureaze ukazuju na to da tripsin inhibitori nisu deaktivirani. Aktivnost ureaze u sirovoj soji je oko 2,0, a smatra se da je sojino zrno pravilno obrađeno ako je aktivnost ureaze 0,05-0,2. Ako su vrijed-nosti ureaze blizu ništice to nam ukazuje da su tripsin inhibitori deaktivirani, međutim, to može značiti i da je obrada soje bila neadekvatna te su se pritom uništile i brojne aminokiseline. Drugi način utvrđivanja dobre obrade sojinog zrna i stupnja razaranja proteina je određivanjem topljivosti proteina. Toplinskom obradom soje topljivost proteina u vodi kalijevim hidroksidom (KOH) i drugim otapalima se smanjuje. Soja s visokom topljivošću u KOH i niskom aktiv-nosti ureaze zadovoljava zahtjeve za aminokiselinama za uzgoj brojlera (Willis, 2004.). Araba i Dale (1990 a,b) utvrdili su da je prirast pilića čija je hrana sadržavala punomasnu soju kao glavni izvor esenci-jalnih aminokiselina bio slabiji ako je topljivost proteina u KOH bila niža od 70% i viša od 85%. Novijim istraži-vanjima Shin (2002) preporučuje da topljivost proteina u KOH za brojlere i nesilice bude 75%-90%, a aktivnost ureaze 0,05-0,3. Dozier i Hess (2011.) su utvrdili da su vrijednosti topljivosti bjelančevina prihvatljive ako su od 78% do 84% i da ozna-čavaju optimalan postupak obrade toplinom. Za kokoši nesilice i starije brojlere topljivost bjelančevina u KOH prihvatljiva je od 84% do 89% budući da su manje osjet-ljiviji na antinutritivne tvari, a niža od 74% očituje se u padu iskoristivosti lizina za sve životinje. Anderson-Haferman i sur. (1992.) prikazali su kako se probavljivost lizina, metionina, cistina i treonina povećava kao rezultat toplinske obrade sojinog zrna (tablica 4.).
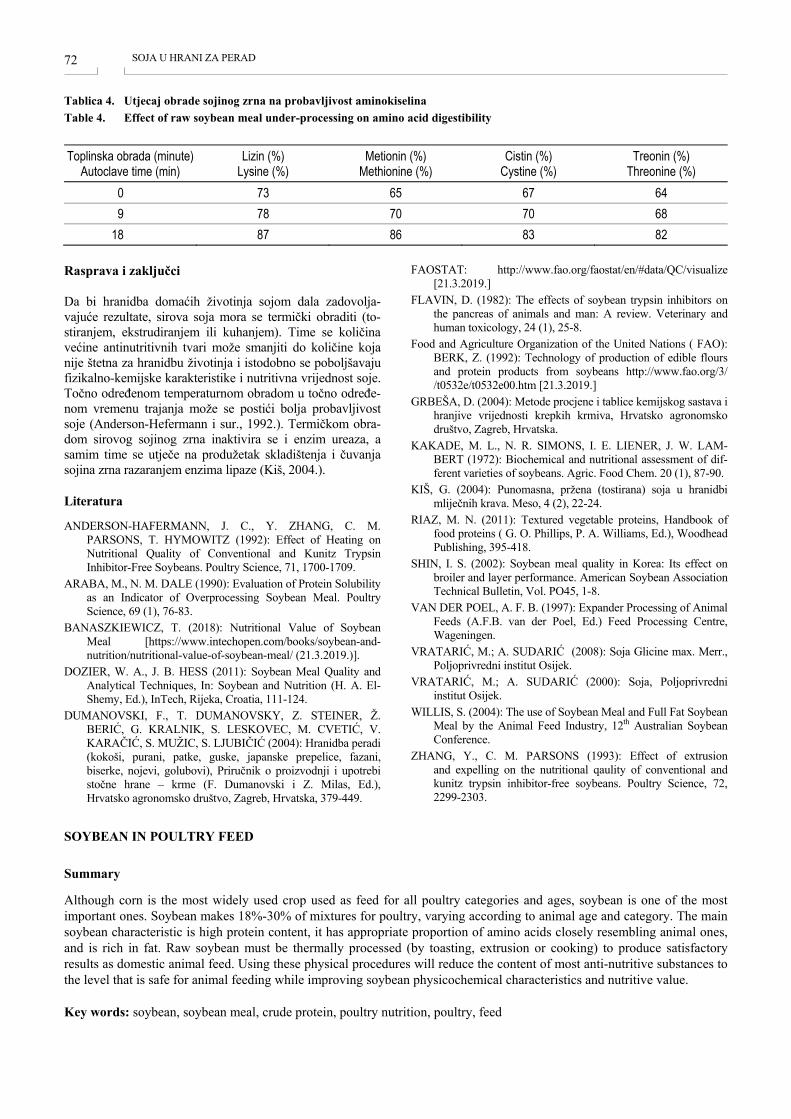
SOJA U HRANI ZA PERAD 72
Tablica 4. Utjecaj obrade sojinog zrna na probavljivost aminokiselina Table 4. Effect of raw soybean meal under-processing on amino acid digestibility
Toplinska obrada (minute) Autoclave time (min)
Lizin (%) Lysine (%)
Metionin (%) Methionine (%)
Cistin (%) Cystine (%)
Treonin (%) Threonine (%)
0 73 65 67 64 9 78 70 70 68
18 87 86 83 82 Rasprava i zaključci
Da bi hranidba domaćih životinja sojom dala zadovolja-vajuće rezultate, sirova soja mora se termički obraditi (to-stiranjem, ekstrudiranjem ili kuhanjem). Time se količina većine antinutritivnih tvari može smanjiti do količine koja nije štetna za hranidbu životinja i istodobno se poboljšavaju fizikalno-kemijske karakteristike i nutritivna vrijednost soje. Točno određenom temperaturnom obradom u točno određe-nom vremenu trajanja može se postići bolja probavljivost soje (Anderson-Hefermann i sur., 1992.). Termičkom obra-dom sirovog sojinog zrna inaktivira se i enzim ureaza, a samim time se utječe na produžetak skladištenja i čuvanja sojina zrna razaranjem enzima lipaze (Kiš, 2004.).
Literatura
ANDERSON-HAFERMANN, J. C., Y. ZHANG, C. M. PARSONS, T. HYMOWITZ (1992): Effect of Heating on Nutritional Quality of Conventional and Kunitz Trypsin Inhibitor-Free Soybeans. Poultry Science, 71, 1700-1709.
ARABA, M., N. M. DALE (1990): Evaluation of Protein Solubility as an Indicator of Overprocessing Soybean Meal. Poultry Science, 69 (1), 76-83.
BANASZKIEWICZ, T. (2018): Nutritional Value of Soybean Meal [https://www.intechopen.com/books/soybean-and-nutrition/nutritional-value-of-soybean-meal/ (21.3.2019.)].
DOZIER, W. A., J. B. HESS (2011): Soybean Meal Quality and Analytical Techniques, In: Soybean and Nutrition (H. A. El-Shemy, Ed.), InTech, Rijeka, Croatia, 111-124.
DUMANOVSKI, F., T. DUMANOVSKY, Z. STEINER, Ž. BERIĆ, G. KRALNIK, S. LESKOVEC, M. CVETIĆ, V. KARAČIĆ, S. MUŽIC, S. LJUBIČIĆ (2004): Hranidba peradi (kokoši, purani, patke, guske, japanske prepelice, fazani, biserke, nojevi, golubovi), Priručnik o proizvodnji i upotrebi stočne hrane – krme (F. Dumanovski i Z. Milas, Ed.), Hrvatsko agronomsko društvo, Zagreb, Hrvatska, 379-449.
FAOSTAT: http://www.fao.org/faostat/en/#data/QC/visualize [21.3.2019.]
FLAVIN, D. (1982): The effects of soybean trypsin inhibitors on the pancreas of animals and man: A review. Veterinary and human toxicology, 24 (1), 25-8.
Food and Agriculture Organization of the United Nations ( FAO): BERK, Z. (1992): Technology of production of edible flours and protein products from soybeans http://www.fao.org/3/ /t0532e/t0532e00.htm [21.3.2019.]
GRBEŠA, D. (2004): Metode procjene i tablice kemijskog sastava i hranjive vrijednosti krepkih krmiva, Hrvatsko agronomsko društvo, Zagreb, Hrvatska.
KAKADE, M. L., N. R. SIMONS, I. E. LIENER, J. W. LAM-BERT (1972): Biochemical and nutritional assessment of dif-ferent varieties of soybeans. Agric. Food Chem. 20 (1), 87-90.
KIŠ, G. (2004): Punomasna, pržena (tostirana) soja u hranidbi mliječnih krava. Meso, 4 (2), 22-24.
RIAZ, M. N. (2011): Textured vegetable proteins, Handbook of food proteins ( G. O. Phillips, P. A. Williams, Ed.), Woodhead Publishing, 395-418.
SHIN, I. S. (2002): Soybean meal quality in Korea: Its effect on broiler and layer performance. American Soybean Association Technical Bulletin, Vol. PO45, 1-8.
VAN DER POEL, A. F. B. (1997): Expander Processing of Animal Feeds (A.F.B. van der Poel, Ed.) Feed Processing Centre, Wageningen.
VRATARIĆ, M.; A. SUDARIĆ (2008): Soja Glicine max. Merr., Poljoprivredni institut Osijek.
VRATARIĆ, M.; A. SUDARIĆ (2000): Soja, Poljoprivredni institut Osijek.
WILLIS, S. (2004): The use of Soybean Meal and Full Fat Soybean Meal by the Animal Feed Industry, 12th Australian Soybean Conference.
ZHANG, Y., C. M. PARSONS (1993): Effect of extrusion and expelling on the nutritional qaulity of conventional and kunitz trypsin inhibitor-free soybeans. Poultry Science, 72, 2299-2303.
SOYBEAN IN POULTRY FEED
Summary
Although corn is the most widely used crop used as feed for all poultry categories and ages, soybean is one of the most important ones. Soybean makes 18%-30% of mixtures for poultry, varying according to animal age and category. The main soybean characteristic is high protein content, it has appropriate proportion of amino acids closely resembling animal ones, and is rich in fat. Raw soybean must be thermally processed (by toasting, extrusion or cooking) to produce satisfactory results as domestic animal feed. Using these physical procedures will reduce the content of most anti-nutritive substances to the level that is safe for animal feeding while improving soybean physicochemical characteristics and nutritive value. Key words: soybean, soybean meal, crude protein, poultry nutrition, poultry, feed

PERADARSKI DANI 2019. 73
UČINAK DODATKA ALUVIJALNOG DEPOZITNOG HOLOCENSKOG MINERALA (ADNEM) U KRMU TOVNIH PILIĆA FARMSKOG UZGOJA NA TVORBU KOLAGENA U NJIHOVU MIŠIĆJU
Hrvoje Mazija3, Zlatko Janječić1 Helga Medić4, Dalibor Bedeković1, Mirta Balenović2, Marijan Andrašec3, Bratko Filipič3
1 Sveučilište u Zagrebu, Agronomski fakultet, Zavod za hranidbu životinja, Zagreb, Hrvatska 2 Hrvatski veterinarski institut, Centar za peradarstvo, Zagreb, Hrvatska 3 Institut za eksperimentalnu i translacijsku onkologiju, Zagreb, Hrvatska 4 Sveučilište u Zagrebu, Prehrambeno biotehnološki fakultet, Zavod za prehrambeno-tehnološko inženjerstvo, Zagreb, Hrvatska
Sažetak
Nastavljeno je istraživanje povoljnog učinka dodatka 0,5% aluvijalnog depozitnog nanosa holocenskog minerala (ADNEM) u krmu tovnih pilića, ali na način kojim bi se spoznalo optimalno razdoblje tova za njegovu primjenu s obzirom na proizvodni uspjeh i tvorbu kolagena u prsnom mišićju. Preliminarnim ograničenim istraživanjem postignut je bolji učinak dodatka 0,5% negoli 1,0% ADNEM-a. Zato je u ovom mikropokusu ADNEM dodavan skupinama pilića bilo u početnu, završnu I. odnosno završnu II. smjesu u količini od 0,5% te su bilježeni pokazatelji uspješnosti tova. Dvije usporedne skupine pilića jele su ADNEM (0,5%) tijekom cijelog razdoblja tova odnosno uopće ga nisu jele. Pokazalo se da je ADNEM dodan u prvih 11 dana tova jednako učinkovit kao i onaj koji se davao tijekom svih 35 dana tova. Tvorba kolagena u utovljenih pilića dviju kontrolnih skupina bila je veća kod skupine koja je neprestano jela hranu s dodanih 0,5% ADNEM-a. S obzirom na relativno malen broj pretraženih uzoraka mišićja ovaj se rezultat ne može nužno pripisati povećanoj tvorbi kolagena, već i drugim čimbenicima, ponajprije mineralnom sastavu ADNEM-a, ali i dobi pilića. Uspjeh makropokusa u farmskom tovu pilića potvrdio je zapažanja u oba preliminarna pokusa. Pilići koji su jeli dodani ADNEM (34.900 pilića) postigli su europski indeks učinkovitosti (EPEF) 342, a oni koji ga nisu jeli 327, što je za 15 indeksnih jedinica manje. Također u dobi četiri tjedna tjelesna masa im je bila 1555 g prema 1438 g, što ukazuje na mogućnost skraćenja tova ako se u krmnu smjesu primiješa 0,5% ADNEM-a. Najznačajnije je zapažanje uspješnosti mehaničkog odvajanja prsnog mišićja, koja je u pilića s dodanim ADNEM-om bila potpuna, dok je usporedno u pilića bez dodanog ADNEM-a u krmnu smjesu približno 2% mišića bilo gnjecavo i neiskoristivo za stavljanje na tržište kao file. Potreban je nastavak istraživanja kako bi se razjasnili čimbenici koji dovode do poboljšanja proizvodne uspješnosti tova pilića uz primjenu ADNEM-a.
Ključne riječi: tovni pilići, aluvijalni depozitni nanos erodiranih minerala, krmna smjesa, kolagen Uvod
Glavnina holocenskih aluvijalnih mineralnih pijesaka (HAMP) zbog svojeg neutralnog djelovanja korištena je uglavnom u usporednim istraživanjima učinka mineralnih dodataka krmnim smjesama. Kao negativne kontrole uspo-ređeni su s nekim zeolitima te nisu imali učinka na zdravlje ili proizvodna svojstva kokoši nesilica (Olver, 1997.). Tek su u posljednjih nekoliko godina privukli značajniju po-zornost istraživača, ali i praktičara s obzirom na očitovane učinke što su vezani uz različitost njihova kemijskog sastava
odnosno geografskog područja iz kojeg su potekli. Pri-mijenjeni kao dodatak stočnoj hrani, napose peradi, tovnih pilića i kokoši nesilica očitovali su različite povoljne učinke. Sažeto, ti se povoljni učinci odnose općenito na zdravlje i proizvodna svojstva tovnih pilića (Trkulja, 2012., Janječić i sur., 2017.), bolju aktivnost imunosnog sustava očitovanu poboljšanjem uspjeha cijepljenja protiv newcastleske bolesti (Vidović, 2011.) odnosno Marekove bolesti (Kokić, 2012.). Također, ublažili su i tzv. sindrom crnih kostiju (Lyon i sur., 1976.). Spektar različitosti učinaka mogao se je pripisati raz-likama u kemijskom sastavu tih pijesaka, što je neposredno
prof. emeritus Hrvoje Mazija, dr. med. vet., Institut za eksperimentalnu i translacijsku onkologiju, 10000 Zagreb, Hrvatska, e-mail: [email protected]

UČINAK DODATKA ALUVIJALNOG DEPOZITNOG HOLOCENSKOG MINERALA (ADNEM) U KRMU TOVNIH PILIĆA FARMSKOG UZGOJA NA TVORBU KOLAGENA U NJIHOVU MIŠIĆJU
74
povezano s njihovim već spomenutim određenim geograf-skim podrijetlom. Porječje rijeke Drave, s obzirom na geološke različitosti ko-jim se njen sliv prostire moglo je tako sadržavati holocenske aluvijalne minerale za koje se zna da imaju neki biološki učinak. Potvrdila je to analiza pijeska s lokacije Okrugljača u blizini grada Virovitice, koja je obavljena u Hrvatskom geološkom institutu, Zavodu za mineralne sirovine, Zagreb, Sachsova 2. i u cijelosti opisana u prethodnim istraživanjima (Janječić i sur., 2017.). S obzirom na to da se radi o alu-vijalnom depozitnom nanosu erodiranih minerala, ovaj je pijesak nazvan akronimom ADNEM i kao takav registriran pa mu je izdana uporabna dozvola od strane relevantnih institucija. Nađeno je ukupno 10 mineralnih oksida odnosno 52 ele-menta. Najviše je SiO2 (88,71%), a zatim AlO3 (5,37%). Od esencijalnih elemenata za perad tu se nalaze kalcij, natrij, fosfor i klor, a od mikroelemenata mangan, željezo, bakar, cink, jod, molibden, krom i selen (Klassing i Barnes, 1988.). Moguća esencijalna uloga drugih mikroelemenata nije još dovoljno istražena, što se odnosi na bor, arsen, fluor, nikal, rubidij, vanadij i još neke rijetke minerale. Nađene količine moguće toksičnih mikroelemenata manje su od dopuštenih (NRC, 2005.). Thirulogachandar i sur. (2014) te Suganya i sur. (2016.) opisuju toksičnost nekih minerala za perad, naročito metala od kojih su neki izrazito toksični i za ljude. Zato treba voditi računa o njihovim količinama u hrani kako ne bi štetno djelovali na perad, ali i na konzumenta pro-izvoda od peradi (meso, jaja, prerađevine). U tom smislu najznačajniji su olovo, arsen, kadmij, vanadij, živa, cink i bakar. Za ADNEM je dokazano povoljno djelovanje na zdravlje i proizvodna svojstva tovnih pilića (Janječić i sur., 2017.). Primiješan krmnim smjesama za tov pilića (početna, završna I. i završna II.) u količinama 0,5% odnosno 1,0% očitovao je povoljne učinke i to napose dodanih 0,5%. Proizvodni uspjeh izražen europskim faktorom učinkovitosti proizvod-nje (EPEF, European Production Eficacy Factor) (Marcu i sur., 2013.) u kontrolnih pilića koji nisu jeli dodani ADNEM iznosio je 398,49 (35. dana tova) odnosno 362,12 (42. dana tova), a u pokusnih 35. dana 395,27 te 449,89 (dodanih 0,5% ADNEM), a 42. dana 380,05 i 427,37 (dodanih 1,0% ADNEM). Iako su obje dodane količine ADNEM-a po-voljno djelovale, dodanih 0,5% pokazalo se osobito zna-čajnim (razlika od kontrole 42. dana tova iznosi 87,77 indeksnih jedinica). Trbušne masti u obje pokusne skupine bilo je manje u odnosu na kontrolnu kojoj hranom nije primiješan ADNEM. Treba naglasiti da masa netom izleženih pilića nije u kore-laciji s masom utovljenih pilića, nego s njihovom masom u dobi sedam dana (Decuypere, 1979; Tona i sur., 2003, Tona i sur., 2005., Michalczuk i sur., 2011.). Zbog toga je treba uzeti u obzir u predviđanju konačne mase pilića i kao dokaz kvalitete jednodnevnih pilića (Deeming, 1995; Tona i sur., 2003.). Zbog povoljnih rezultata primjene ADNEM-a u pokusnim uvjetima (Janječić i sur., 2017.) u mikropokusu istražen je
mogući učinak dodatka ADNEM-a u krmnoj smjesi na količinu aminokiseline hidroksiprolin u prsnom mišićnom tkivu tovnih pilića. Ova je aminokiselina sadržana u kola-genu koji je pak jedan od najznačajnijih pokazatelja njež-nosti mesa. Kao najzastupljenija bjelančevina u organizmu kolagen čini glavninu vezivnog tkiva (Hultin, 1985., Bechtel, 1986.). Jedinstvene je strukture prilagođene prven-stveno otporu istezanja. Kolagensku nit čine tri polipeptidna lanca (tropokolagen). Njihova stabilnost određena je ion-skim i hidrofobnim međudjelovanjem (Pearson i Young, 1989.). Vezivno tkivo mišićja, međutim, ne igra glavnu ulogu u mekoći mišićja mlađe peradi (pilića dobi 6-8 tje-dana), jer križne veze kolagenih vlakana još nisu oblikovane pa zato kolagen hidrolizira kuhanjem (meso postaje mekše). Porastom dobi životinje povećava se čvrstoća mesa koja se termičkom obradom u odrasle više ne smanjuje. Različitost uspjeha primjene ADNEM-a s obzirom na dob tovnih pilića potakla nas je na nastavak istraživanja kojima se nastojalo utvrditi u kojem razdoblju tova pilića primi-ješani ADNEM djeluje najpovoljnije ne samo na proizvodna svojstva, nego posebice na kakvoću mesa odnosno njegovu nježnost. Zato se je dio istraživanja odnosio i na primjenu ADNEM-a na način da je dodavan u različitoj fazi tova u krmnu smjesu (početnu, završnu I. odnosno završnu II.). Tako se nastojalo utvrditi kada je optimalno dodavati ga s obzirom na očekivani učinak, tj. na optimalnu količinu kola-gena (hidroksiprolina) u mišićju te njihovu nježnost. Njego-va količina mogla bi biti i pokazatelj očekivane tehnološke uspješnosti otkoštavanja mišićja (napose osobito vrijednog prsnog, mm. pectorales) pri mehaničkoj klaoničkoj obradi trupova pilića. Treba naglasiti da na uspjeh mehaničke obra-de trupova značajno utječe i razvoj mrtvačke ukočenosti mesa (rigor mortis) pa ono postaje čvršće i podložnije toj obradi. U tom smislu i veličina pilića (trajanje i način tova) imat će značajnu ulogu (Cavitt i sur., 2001., Dawson i sur., 1987., Dodge i sur., 1959.). Zasebno, ovaj je pripravak korišten i u makropokusu, tj. farmskom tovu pilića (farma tovnih pilića u Istri). Uz ocjenu uspješnosti tova, u ovih je pilića ocijenjeno i svojstvo stroj-nog odvajanja prsnog mišićja. Učinjeno je to posredno, sub-jektivnom procjenom postotka odbačaja prsnog mišićja nakon mehaničke obrade trupova, zapravo usporedbom skupina pilića koje nisu odnosno jesu dobivale ADNEM primiješan (0,5%) krmnoj smjesi.
Materijal i metode
Pilići u mikropokusu Istraživanje je započeto s 200 netom izleženih pilića oba spola (tovni hibrid Ross) podrijetlom iz valionice „Valipile“, Sesvetski Kraljevec. Razvrstani su slučajnim izborom u pet pokusnih skupina te smješteni u 10 kaveza (po 20 pilića u svaki) nastambe za pokuse Zavoda za hranidbu životinja Agronomskog fakulteta Sveučilišta u Zagrebu. Prostor je pripremljen za prijam pilića prema standardima za njihov podni uzgoj, a očekivana im je tjelesna masa bila 33 kg/m2 na kraju pokusa, koji je tako određen na 35 dana. Prostor je

UČINAK DODATKA ALUVIJALNOG DEPOZITNOG HOLOCENSKOG MINERALA (ADNEM) U KRMU TOVNIH PILIĆA FARMSKOG UZGOJA NA TVORBU KOLAGENA U NJIHOVU MIŠIĆJU
75
grijan radijatorima i pomoću infracrvenih žarulja kako bi se stalno održavala optimalna temperatura prostora tijekom tra-janja pokusa. Svi su pilići nakon vaganja smješteni na pro-stirku od drvnih strugotina debljine približno 10 cm, koja je prvih sedam dana prekrivena debljim papirom kako bi se spriječio rasap krme i jedenje stelje (opasnost od opsti-pacije). Krmne smjese i hranidba pilića U ovom pokusu je istraživan utjecaj dodatka 0,5% ADNEM-a u krmne smjese u različitom razdoblju tova pilića, što je temeljeno na rezultata preliminarnih istraži-vanja u kojima se pokazala učinkovitijom negoli dodanih 1,0% (Janječić i sur., 2017.). Pet pokusnih skupina pilića u dva ponavljanja (ukupno 10 kaveza) je tijekom istraživanja jelo krmne smjese kako je prikazano u tablici 1.
Sve korištene krmne smjese izrađene su u TSH „Valipile“, Sesvetski Kraljevec, a iz već dostavljenih količina uzeti su uzorci za pretragu. Kemijski sastav (određen na Zavodu za hranidbu životinja Agronomskog fakulteta Sveučilišta u Zagrebu) prikazan je u tablici 2. Pilići su prvih sedam dana jeli krmnu smjesu ad libitum iz okruglih plastičnih podnih hranilica, a zatim do kraja istraživanja iz pocinčanih okruglih visećih hranilica kapa-citeta 10 kg. ADNEM je ručno primiješan u gotove krmne smjese u pokusnom prostoru. Pilići su prvih sedam dana pili iz okruglih pojilica kapaciteta 3 L, a zatim do kraja istraži-vanja iz visećih automatskih okruglih pojilica koje su podi-zane sukladno s porastom pilića. Na taj se način poticalo jačanje mišićja nogu, sprječavalo onečišćenje vode fecesom i pojava prsnih žuljeva (bursitis praesternalis), što umanjuje klaoničku vrijednost.
Tablica 1. Primijenjene krmne smjese tijekom tova pilića Table 1. Broiler feed mixtures applied during fattening period
Pokusna skupina / Study group A B C D K***
Krmna smjesa / Feed mixture Primijenjen ADNEM (dana) / ADNEM admixed (days)
Početna** / Initial** (0-11) S* S S (0-11) Završna I** / Final** S (12-28) S S (12-28) Završna II / Final ** S S (29-35) S (29-35) *S – standardna krmna smjesa / standard feed mixture **u smjesu primiješan kokcidiostatik / coccidiostat admixed to feeding mixture ***u dobi od 2. do 6. dana u pitku vodu dodan antibiotik enrofloksacin (Enroxil® 10%) / antibiotic enrofloxacin (Enroxil® 10%) added to drinking water at the age of 2 to 6 days Pilići A su dobivali ADNEM u prvih 11 dana / Group A received ADNEM in the first 11 days Pilići B su dobivali ADNEM od 12. do 28. dana / Group B received ADNEM from day 12 to day 28 Pilići C su dobivali ADNEM od 29. do 35. dana / Group C received ADNEM from day 29 to day 35 Pilići D ne dobivaju ADNEM tijekom trajanja tova / Group D did not receive ADNEM during fattening period Pilići K dobivaju ADNEM cijelo vrijeme tova, od 1. do 35. dana / Group K received ADNEM throughout fattening period, from day 1 to day 35 Tablica 2. Kemijski sastav krmnih smjesa korištenih u pokusu Table 2. Chemical composition of feeding mixtures used in the experiment
Pokazatelj / Indicator Jedinica / Unit Početna / Initial Završna I. / Final I Završna II. / Final II
Vlaga / Moisture g/kg 113 108 106 Pepeo / Ashes g/kg 56 56 52 Sirovi protein / Raw protein g/kg 224,8 202,4 181,1 Mast / Fat g/kg 60 53 68 Sirova vlakna / Raw fiber g/kg 17 15 19 Kalcij / Calcium g/kg 10,3 10,0 9,7 Fosfor / Phosphorus g/kg 6,5 6,4 5,8 Natrij / Sodium g/kg 1,6 1,6 1,6 Šećer / Sugar g/kg 390,6 395,8 411,2 Škrob / Starch g/kg 50,0 53,9 54,2

UČINAK DODATKA ALUVIJALNOG DEPOZITNOG HOLOCENSKOG MINERALA (ADNEM) U KRMU TOVNIH PILIĆA FARMSKOG UZGOJA NA TVORBU KOLAGENA U NJIHOVU MIŠIĆJU
76
Određivanje kolagena (hidroksiprolina) u prsnom mišićju Uzorci prsnog mišićja (mm. pectorales superficialis) uzeti su na kraju pokusa u dobi pilića 35 dana i to jedino od muških jedinki. Do pretrage čuvani su u zamrzivaču pri -18 oC. Količina kolagena određena je posredno, određiva-njem hidroksiprolina, prema normi HRN ISO 3496:1994 - Meso i mesni proizvodi (Anon.,1994.). U metodu je uključena hidroliza testnog dijela uzorka sa 7 N sumporastom kiselinom pri 105 °C, nakon čega se uzorak filtrira, hidrolizat otapa, a hidroksiprolin oksidira uz klora-min-T i 4-dimetilbenzaldehid. Postupak je opisao Kolar (1990.). Pri toj reakciji nastaje crveno obojeni p-dimetil-amino benzaldehid, a intenzitet boje (količina hidroksi-prolina) odredi se spektrofotometrijski pri valnoj dužini 558 nm. Količina ukupnog kolagena dobiva se množenjem po-stignutog nalaza faktorom 8. Makropokus – farmski tov pilića Pokusna skupina ADNEM (0,5%) primiješan je u sve tri smjese (početna, završna I. i završna II.) za tov pilića smještenih u dvije nastambe moderne farme tovnih pilića u Istri (hale 4 i 5). U svaku nastambu useljeno je po 17.200 pilića izleženih u valionici Valipile d.o.o., Sesvetski Kraljevec). Pilići u hali 4 bili su hibridi Cobb 500, podrijetlom od matičnog jata u dobi 34 tjedna. U halu 5 useljeni su pilići hibrida Ross 308 ma-tičnog jata u dobi 27 tjedana. Tako je ADNEM primijenjen u krmnim smjesama kod ukupno 34.900 pilića. Usporedna skupina Usporednu skupinu činili su pilići kojima u krmnu smjesu nije primiješan ADNEM, smješteni na istoj farmi u hale 1, 2 i 3. Ukupno u sve tri nastambe smješteno je 50.300 jedno-dnevnih pilića, svi podrijetla matičnih jata hibrida Cobb 500 u dobi 29 tjedana,17.000 pilića u halu 1 te također 17.000 pilića roditeljskog jata u dobi 28 tjedana. U halu 3 useljeno je 16.300 pilića matičnog jata u dobi 60 tjedana.
U obje skupine pilića korištena je istovjetna krmna smjesa proizvedena u TSH „Valipile“, Sesvetski Kraljevec. Njen sastav opisan je u mikropokusu s razlikom što je za farmski uzgoj pilića ADNEM (0,5%) primiješan još u TSH „Valipile“, Sesvetski Kraljevec. Tov pilića u svih pet hala trajao je 35 dana te započeo u razdoblju od 20.2.-2.3. 2017. godine, što je značilo i približno istovjetne makroklimatske pa zato i istovjetne mikroklimatske uvjete u svih pet nastambi pilića. Istraživanje je provedeno u skladu s Pravilnikom o zaštiti životinja koje se koriste u pokusima ili u druge znanstvene svrhe (NN 55/2013) koji je izdalo Ministarstvo poljopri-vrede Republike Hrvatske, a koji je u skladu s legislativom Europske Unije. Rezultati Rezultati mikropokusa Tjelesne mase pilića (g), konverziju krmne smjese (kg/kg) te uginuće (apsolutni broj) tijekom 35 dana pokusa i prema pokusnim skupinama prikazujemo u tablici 3. Najveći prirast do 27. dana života postigli su pilići skupine D (1762,56 g) kojima ADNEM uopće nije dodan u krmnu smjesu. Slično, u dobi 35 dana također najteži su pilići skupine D. Pilići koji su stalno jeli u krmnoj smjesi pri-miješani ADNEM (skupina K) lakši su od pilića skupine K za 1,84 grama. Značajan je, međutim, i prirast što su ga pilići pojedinih skupina ostvarili u razdoblju od 27. do 35. dana tova. Važno je to zbog uobičajene tjelesne mase pilića što se stavljaju na tržište. Oni lakši, „grill“, tove se do postizanja približno 1,5-1,8 kg žive vage i gospodarski su najisplativiji. Teži pilići tove se do 35 ili više dana, također ovisno o zahtjevu tržišta. Zbog navedenog zasebnom tablicom (tablica 3a.) prikazujemo ostvarene tjelesne mase pilića u razdoblju tova 27-35 dana. One upućuju na brzinu prirasta u tom razdoblju tova i odrednica su kojom bi se trebali rukovoditi proizvođači tovnih pilića.
Tablica 3. Kinetika prirasta (g), konverzija hrane (kg/kg) te uginuće* pilića tijekom mikropokusa Table 3. Weight gain kinetics (g), feed conversion (kg/kg) and broiler deaths* during micro-experiment
Dob pilića / Broiler age A B C D K
1. dan / Day 1 48,70 48,48 47,00 49,28 48,33
11. dan / Day 11 366,10 367,63 367,67 375,30 359,28
27. dan / Day 27 1757,63 1725,26 1749,36 1762,56 1709,50
35. dan / Day 35 2602,63 2614,47 2564,47 2630,79 2628,95
Konverzija krmne smjese / Feed conversion 1,45 1,45 1,46 1,47 1,48
Uginuće1.-35. dana / Deaths day 1-35 2 2 2 2 2 *od 1.do 27. dana po jedno pile uginulo u skupinama A, C i D, dva u skupini B / from day 1 to day 27, one death per group recorded in groups A, C and D, and two deaths in group B *od 28. do 35. dana po jedno pile uginulo u skupinama A, C i D te dva u skupini K / from day 28 to day 35, one death per group recorded in groups A, C and D, and two deaths in group K

UČINAK DODATKA ALUVIJALNOG DEPOZITNOG HOLOCENSKOG MINERALA (ADNEM) U KRMU TOVNIH PILIĆA FARMSKOG UZGOJA NA TVORBU KOLAGENA U NJIHOVU MIŠIĆJU
77
Tablica 3a. Postignuta tjelesna masa pilića 27. odnosno 35. dana tova i razlika ostvarenog prirasta (g) Table 3a. Broiler body mass achieved on days 27 and 35 of fattening period and weight gain difference (g)
Pokusna skupina pilića / Study group A B C D K Tjelesna masa 27. dana / Body mass on day 27 1757,63 1725,26 1749,36 1762,56 1709,50 Tjelesna masa 35. dana / Body mass on day 35 2602,63 2614,47 2564,47 2630,79 2628,95 Prirast od 27. do 35. dana (g) Weight gain from day 27 to day 35 (g)
845,00 889,21 815,11 898,23 919,45
Prirast od 27. do 35 dana (%) Weight gain from day 27 to day 35 (%)
21,99 23,35 20,90 23,63 24,17
Najveći prirast tijekom završne faze tova postigli su pilići skupine K koji dobivaju ADNEM tijekom čitavog trajanja pokusa, od 1. do 35. dana, a razlika u postignutoj tjelesnoj masi u odnosu na piliće koji nisu dobivali ADNEM iznosi 21,22 grama. Između pilića koji ne jedu (D) i jedu (K) ADNEM najznačajnija je razlika i u relativnom prirastu te iznosi 23,63% prema 24,17%. Relativno najmanji prirast u toj fazi tova (20,90%) ostvarili su pilići koji su jeli dodani ADNEM jedino u posljednjoj fazi tova (završna II. smjesa)
Iz postignutih rezultata tova pilića pojedinih skupina izračunat je i europski indeks učinkovitosti (EPEF) prema niže navedenoj jednadžbi (Marcu i sur., 2013.), i to za razdoblja tova od 1. do 27. odnosno 35 dana tova. Učinjeno je to zbog tova pilića često kraćeg od 35 dana, što je uvjetovano genetskim svojstvima uzgajanog hibrida pilića, njihovom hranidbom te zahtjevima tržišta. Rezultate prikazujemo u tablici 5.
Tablica 4. Konverzija krmne smjese pilića pokusnih skupina (kg/kg) Table 4. Feed conversion according to broiler groups (kg/kg)
Pokusna skupina / Study group A B C D K 11. dan / Day 11 1,14 1,12 1,14 1,13 1,13 27. dan / Day 27 1,45 1,49 1,43 1,45 1,46 35. dan / Day 35 1,77 1,73 1,82 1,84 1,86 1.- 35. dan / Day 1-35 1,45 1,45 1,46 1,47 1,48
Pilići A su dobivali ADNEM u prvih 11 dana / Group A received ADNEM in the first 11 days Pilići B su dobivali ADNEM od 12. do 28. dana / Group B received ADNEM from day 12 to day 28 Pilići C su dobivali ADNEM od 29. do 35. dana / Group C received ADNEM from day 29 to day 35 Pilići D ne dobivaju ADNEM tijekom trajanja tova / Group D did not receive ADNEM during fattening period Pilići K dobivaju ADNEM tijekom trajanja, od 1. do 35. dana / Group K received ADNEM throughout fattening period, from day 1 to day 35 Tablica 5. Europski indeks učinkovitosti (EPEF) u tovnih pilića kojima je u hranu primiješan 0,5% ADNEM u različitoj fazi
tova Table 5. European Production Efficiency Factor (EPEF) in broilers with ADNEM 0.5% admixed to feed
Pokusna skupina / Study group A B C D K EPEF od 1. do 27. dana tova / EPEF, fattening day 1-day 27 427,1 387,0 430,6 427,1 433,8 EPEF od 1. do 35. dana tova / EPEF, fattening day 1-day 35 461,1 454,0 450,9 460,1 457,0
Pilići A su dobivali ADNEM u prvih 11 dana / Group A received ADNEM in the first 11 days Pilići B su dobivali ADNEM od 12. do 28. dana / Group B received ADNEM from day 12 to day 28 Pilići C su dobivali ADNEM od 29. do 35. dana / Group C received ADNEM from day 29 to day 35 Pilići D ne dobivaju ADNEM tijekom trajanja tova / Group D did not receive ADNEM during fattening period Pilići K dobivaju ADNEM tijekom trajanja, od 1. do 35. dana / Group K received ADNEM throughout fattening period, from day 1 to day 35

UČINAK DODATKA ALUVIJALNOG DEPOZITNOG HOLOCENSKOG MINERALA (ADNEM) U KRMU TOVNIH PILIĆA FARMSKOG UZGOJA NA TVORBU KOLAGENA U NJIHOVU MIŠIĆJU
78
Očevidno, do dobi 27 dana najviši EPEF (433,8) postigli su pilići skupine K koji su stalno jeli hranu s dodanih 0,5% ADNEM-a. Međutim, pilići koji su ga jeli samo u prvih 11 dana života (skupina A) postigli su općenito najviši EPEF (461,1). Nalaz hidroksiprolina* (kolagena) u prsnom mišićju pilića pokusnih skupina (35. dan tova) prikazujemo u tablici 6. U odnosu na sve pokusne skupine, najviše je kolagena na-đeno u prsnom mišićju pilića kojima ADNEM nije pri-miješan u hranu tijekom trajanja tova (1-35 dana). Manje ga je u pilića skupine K kojima je ADNEM primiješan u hranu tijekom 35 dana, gdje mu količina varira od najmanje 0,3416 do 0,6112. Ovo je razumljivo s obzirom na mali broj pretraženih uzoraka (svega pet).
Rezultat tova pilića na farmi Kod tova pilića na farmi u cijelosti je poštivan temeljni princip farmskog uzgoja peradi „sve unutra – sve van“, uz istovjetnu osnovnu hranidbu i mikroklimatske uvjete pa se razlike u postignutim rezultatima mogu pripisati isključivo primjeni 0,5% ADNEM-a. Doduše, valja pripomenuti da su pilići podrijetla različitih tovnih hibrida kokoši koji su se razlikovali u dobi. Broj i tjelesna masa (g) te uginuće (%) pilića tovljenih s dodanih 0,5 ADNEM-a u krmnu smjesu ili bez njega (farmski uzgoj) prikazujemo u tablici 7. Tablica 8. prikazuje skupne rezultate (tjelesnu masu) pilića u peradnjacima gdje su s krmom dobivali ADNEM i uspo-redno s pilićima koji ga nisu jeli.
Tablica 6. Nalaz hidroksiprolina* (kolagena) u prsnom mišićju pilića pokusnih skupina (35. dan tova) Table 6. Hydroxyproline* (collagen) finding in broiler breast muscles (fattening day 35)
Pokusna skupina pililća / Study group K** K K K K D***
Nalaz kolagena (%) / Collagen finding (%) 0,6112 0,3416 0,4264 0,4552 0,4328 0,684
*aminokiselina zastupljena s 12,5% u kolagenu / *12.5% of amino acid present in collagen **K – Pilićima u hrani primiješan ADNEM tijekom trajanja tova / **K – ADNEM admixed to broiler feed during fattening period ***D – Pilićima u hranu nije primiješan ADNEM tijekom trajanja tova / ***D – ADNEM admixed to broiler feed throughout fattening period Tablica 7. Broj i tjelesna masa (g) te uginuće (%) pilića tovljenih sa ili bez dodanih 0,5% ADNEM-au krmnu smjesu (farmski
uzgoj) Table 7. Number, body mass (g) and deaths (%) of broilers fattened with or without 0.5% ADNEM added to feeding mixture
(farm breeding)
Hibrid i dob roditelja pilića (tj.)
Hybrid and parental age (wks)
Hala i broj useljenih pilića
Barn and number of
broilers
1. tjedan Week 1
2. tjedan Week 2
3. tjedan Week 3
4. tjedan Week 4
5. tjedan Week 5
Ukupno uginuće (%) Total deaths
(%)
Očekivana tjelesna masa (g) Expected body
mass (g)
Ostvarena tjelesna masa (g)
Achieved body mass (g)
Cobb 500 (29) I. 17.000 146 432 871 1457 2020 0,32 2000 2020
Cobb 500 (28) II. 17.000 152 427 847 1368 1940 0,29 2000 1940
Cobb 500 (60) III. 16.300 178 432 887 1490 2020 0,51 2000 2020
Ukupno ili Ø (hale I., II. i III.) bez ADNEM-a / Total or Ø (barns I, II and III without ADNEM)
50.300 158,7 430,3 868,3 1438 1993 0,37 2000 1993
Cobb 500 (34) IV. 17.200 192 470 934 1520 2070 0.22 2000 2070
Ross 308 (27) V. 17.700 173 462 977 1590 2040 0,58 2000 2040
Ukupno ili Ø (hale IV i V) dodan ADNEM Total or Ø (barns IV and V) - ADNEM added
34.900 182,5 466 955,5 1555 2055 0,40 2000 2055,0

UČINAK DODATKA ALUVIJALNOG DEPOZITNOG HOLOCENSKOG MINERALA (ADNEM) U KRMU TOVNIH PILIĆA FARMSKOG UZGOJA NA TVORBU KOLAGENA U NJIHOVU MIŠIĆJU
79
Tablica 8. Usporedba dinamike rasta tovnih pilića farmskog uzgoja kojima je u hranu dodan ADNEM i kontrola koje ga nisu jele (g)
Table 8. Comparison of farm broiler growth dynamics with and without ADNEM added (g)
1. tjedan / Week 1 2. tjedan / Week 2 3. tjedan / Week 3 4. tjedan / Week 4 5. tjedan / Week 5
(ADNEM+) (ADNEM+) (ADNEM+) (ADNEM+) (ADNEM+)
173+192 = Ø 182,5
462+470 = Ø 466,0
977+934 = Ø 955,5
1590+1520 = Ø 1555,5
2040+2070 = Ø 2055,0
(ADNEM-) (ADNEM-) (ADNEM-) (ADNEM-) (ADNEM-)
178+152+146 = Ø 158,7
432+427+432 = Ø 430,3
887+847+871 = Ø 868,3
1490+1368+1457= Ø 1438,0
2020+1940+2020= Ø 1993,0
Slika 1. Prosječna dinamika rasta pilića koji su jeli ADNEM dodan u krmnoj smjesi u odnosu na piliće koji ga nisu jeli Figure 1. Average growth dynamics in broilers receiving ADNEM added to feed versus those that did not receive ADNEM Počevši od prvog tjedna tova pa sve do njegova kraja u dobi 35 dana pilići koji su jeli ADNEM bili su teži. No ta razlika nije se linearno povećavala, već je relativno najmanja. Treba naglasiti da je masa pilića nakon sedam dana tova u skupini koja je jela ADNEM bila za 23,8 grama veća (182,5 g prema 158,7 g), što je bila naznaka predviđanja veće konačne mase utovljenih pilića. Iz postignutih rezultata tova pilića obiju skupina izračunati su i EPEF te oni u dobi 35 dana za skupinu koja je jela dodanih 0,5% ADNEM-a u krmi iznosi 342, a onih uspo-rednih 327, što čini značajnu razliku od 15 indeksnih jedinica. Slika 1. prikazuje prosječnu dinamiku rasta pilića koji su jeli ADNEM dodan u krmnoj smjesi u odnosu na piliće koji ga nisu jeli.
Rasprava i zaključci Uspješnost industrijskog tova pilića određena je mnoštvom čimbenika, prvenstveno genetskim svojstvima hibrida, pri-mijenjenom tehnologijom uzgoja, ipak u najznačajnijoj mje-ri hranidbom te konačno i određenom svrhom tova koja mo-že biti naznačena različitim ciljem. Uz navedeno, na uspjeh tova pilića značajno utječe zdravlje roditeljskog jata, masa rasplodnih jaja, masa netom izleženih pilića te naročito ona postignuta u sedmom danu tova (Decuypere, 1979; Tona i sur., 2003., Tona i sur., 2005.; Michalczuk i sur., 2011.). Michalczuk i sur. (2011.) istražili su učinak tjelesne mase netom izleženih pilića (Ross 308) na njihovo svojstvo tovnosti te utvrdili da je ono u negativnoj korelaciji s brzinom tova (rasta) pilića, osobito pri kraćem tovu, ali se u

UČINAK DODATKA ALUVIJALNOG DEPOZITNOG HOLOCENSKOG MINERALA (ADNEM) U KRMU TOVNIH PILIĆA FARMSKOG UZGOJA NA TVORBU KOLAGENA U NJIHOVU MIŠIĆJU
80
produženom ta razlika gubi. Slično, Iqbal i sur. (2017.) za-ključili su da masa jaja pozitivno utječe na kakvoću pilića, njihovu masu i duljinu trupa, ali ne i na konačnu tjelesnu masu, konverziju hrane i uginuće. Porastom dobi pilića ove se razlike smanjuju (Meijerhof, 2006.). Kombinacijom kakvoće hranidbe i napose trajanjem tova te primijenjenim načinom mogu se postići željeni ciljevi koje u konačnici određuju konzumenti odnosno tržište. Stremljenja općenito su postizanje što je moguće veće tjelesne mase utovljenih pilića u što kraćem vremenu, no to istodobno isključuje mogućnost postizanja najkvalitetnijeg mesa s obzirom na teksturu i brojna organoleptička svojstva („zrelost“, mekoća, okus, boja). Zrelost se postiže dugo-trajnijim tovom, također boja i okus, a mekoća skraćivanjem tova. Naime, kratkim tovom ne postiže se dovoljna tvorba kolagena pa je meso često premekano, a zato i nepodobno za mehaničku obradu (strojno odvajanje najvrjednijeg prsnog mišićja). Opisane nedostatke kraćeg tova nastoji se ublažiti ili u cije-losti ukloniti odgovarajućom hranidbom koja bi trebala u vrlo kratkom vremenu osigurati dovoljnu brzinu rasta pilića, a istodobno omogućiti razvoj mišićja. Zato se u krmne smje-se dodaju različiti pripravci, uglavnom mineralne mješavine, prirodnih izvora ili unaprijed određenog i priređenog sastava. U preliminarnim istraživanjima (Janječić i sur., 2017.) upravo se je prirodnom mješavinom različitih minerala i mikroelemenata aluvijalnog podrijetla (ADNEM) dodanoj krmnoj smjesi tovnih pilića u količini 0,5% postigao vrlo povoljan uspjeh njihova tova. Rezultat opisanog makropo-kusa u cijelosti je potvrdio opravdanost primjene ADNEM-a na farmi tovnih pilića (tablica 8.). Postignuti rezultat tova značajno premašuje skupina pilića koja je jela ADNEM dodan u krmnu smjesu. Uz navedeno, na uspjeh tova pilića značajno utječe zdravlje roditeljskog jata, masa rasplodnih jaja te masa netom izleženih pilića (Meijerhof, 2006., Mendes i sur., 2007., Wolanski i sur., 2007.). Dob roditeljskog jata također utječe na masu rasplodnih jaja pa tako posredno i na masu izle-ženih pilića, koja se općenito najčešće koristi kao pokazatelj kvalitete netom izleženih pilića (Decuypere i sur., 2002.). Rezultati mikropokusa poslužili su za ocjenu opravdanosti primjene ADNEM-a u različitim fazama tova (početna, završna I. te II. krmna smjesa). U tih je pilića određena i količina hidroksiprolina (kolagena) kojom bi se mogli protumačiti rezultati makropokusa primjene ADNEM-a u farmskom uzgoju pilića, odnosno klaonički uspjeh pri stroj-nom odvajanju prsnog mišićja. Svi pilići korišteni u pokusu bili su približno istovjetnih početnih tjelesnih masa. Ipak valja naglasiti da su pilići skupine D koji nisu jeli dodani ADNEM u krmnoj smjesi bili za 2,28 grama teži od pilića skupine C koji su jeli dodani ADNEM u dobi 20-35 dana. U tih je pilića ostvaren i najbrži prirast tijekom tova (tablica 3.). Spomenuta početna veća tjelesna masa sigurno je povoljno utjecala na brzinu rasta, kako to naglašavaju Michalczuk i sur. (2011.). Time se doduše mogu tumačiti postignuti prirasti do 27. dana tova (1762,56 g), ali ne i nakon toga, kako to ovi autori navode, jer se učinak tjelesne
mase pri leženju pilića gubi nakon četiri tjedna tova. Razliku u postignutom prirastu tjelesne mase pilića dobi od 27 do 35 dana zato treba pripisati dodanom ADNEM-u, a ta je najveća u pilića skupine K koja je jela ADNEM pridodan krmi u završnoj II. smjesi (tablica 3a.). U odnosu na skupinu D (898,23 g, bez ADNEM-a) ostvareni prirast pilića skupine K je 919,45 grama. Konverzija krmne smjese gotovo je istovjetna u pilića svih skupina i statistički se ne razlikuje. Tek je za 10 dag veća u skupini K (1,48 kg) u odnosu na skupinu D (1,47 kg). Ova razlika, međutim, nije imala učinka na konačni uspjeh tova pilića skupina D odnosno K u dobi do 27 odnosno 35 dana. Izračunati EPEF (Marcu i sur., 2013.) prikazan u tablici 5. ukazuje na različitost uspješnosti tova pilića u po-kusu s obzirom na razdoblje tova u kojem je krmi primiješan ADNEM. Do dobi 27 dana najuspješniji je tov pilića kojima je ADNEM dodavan tijekom cijelog trajanja tova (u sve tri krmne smjese) te iznosi 433,8 indeksnih jedinica. Međutim, pilići kojima je ADNEM davan u početnoj smjesi (prvih 11 dana života) postižu u dobi 35 dana najpovoljniji EPEF, što iznosi 461,1 indeksnih jedinica te je za 4,1 jedinicu veći od pilića koji stalno jedu primiješani ADNEM. Ovaj se rezultat može pripisati učinku ADNEM-a za koji je (doduše in vitro) dokazano da potiče tvorbu interferona (Šooš i sur., 2015., Filipič i sur., 2016.). Tim se učinkom može tumačiti opće-nito dobro zdravlje pilića u pokusu, koje je već opisano u preliminarnim istraživanjima (Janječić i sur., 2017.). Treba naglasiti da se pokusni uvjeti uvijek razlikuju od onih farm-skih te su i postignute vrijednosti EPEF više. Uspješnost tova pilića farmskog uzgoja dviju skupina od kojih je jedna jela u krmnu smjesu primiješan ADNEM (34.900 pilića smještenih u tri nastambe), a druga usporedna (50.300 pilića) bez dodanog ADNEM-a prikazana je tablicom 5. Prosječno najveću tjelesnu masu jednako u dobi četiri i pet tjedana postigli su pilići kojima je u hranu primiješan ADNEM (1555 g i 2055 g). Postignuto uginuće u obje skupine pilića bilo je značajno manje od 1,00% (0,40%), što je zanemarivo i upućuje na općenito visok standard uzgoja pilića. Skupni rezultat za ove dvije skupine pilića prikazan tablicom 6. jasno ukazuje na uspješnost primjene ADNEM-a. Primiješanim ADNEM-om već u četvrtom tjednu tova pilići postižu prosječno 1555,5 g tjelesne mase u usporedbi sa skupinom bez dodanog ADNEM-a koja je za 175 g lakša (1438,0 g). Ostvareni rezultat na kraju tova u dobi 35 dana još je značajniji kad pilići koji su jeli ADNEM postižu 2055,0 g, a oni koji ga nisu jeli 1993,0 grama, što je za 620 grama manje. Upravo na ovom mjestu treba spomenuti rezultat određi-vanja količine kolagena (hidroksiprolina) u prsnom mišićju muških pilića u mikropokusu (Bonifer i Froning, 1996.), zakonski propisanim postupkom (Anon., 1994.). Tablicom 4. prikazani su rezultati analize prsnog mišićja (mm. pectorales), kojim bi se trebala opravdati svrha istraživanja. Naime, u pilića koji su jeli dodani ADNEM (0,5%) strojno odvajanje prsnog mišićja bilo je u cijelosti uspješno, dok je u skupini koja ga nije jela približno 2% ostalo oštećeno

UČINAK DODATKA ALUVIJALNOG DEPOZITNOG HOLOCENSKOG MINERALA (ADNEM) U KRMU TOVNIH PILIĆA FARMSKOG UZGOJA NA TVORBU KOLAGENA U NJIHOVU MIŠIĆJU
81
(„filetirkom“) i tako izgubilo tržišnu vrijednost koju postiže prsni file. Na temelju dobivenih rezultata određivanja udjela kolagena ne može se tvrditi da je dodatak ADNEM-a imao utjecaj na udjel kolagena, već bi se to trebalo utvrditi sljedećim istraživanjem. U odnosu na sve pokusne skupine, najviše je kolagena nađeno u prsnom mišićju pilića kojima ADNEM nije pri-miješan u hranu. Manje ga je u pilića skupine K kojima je ADNEM primiješan u hranu tijekom cijelog trajanja tova (1-35 dana), gdje mu količina varira od najmanje 0,3416 do 0,6112. Ovo je razumljivo s obzirom na mali broj pretra-ženih uzoraka (svega pet). Medić i sur. (2010.) istražili su moguće djelovanje holocenskih minerala (MHM) na kakvoću mesa tovnih pilića dajući ih u hrani u količini od 0,5%, 1,0%, 1,5% i 2,0%. Dokazali su da primijenjene količine ne djeluju štetno, ali nemaju utjecaja na kakvoću mesa. U kontekstu naših istraživanja valja naglasiti da je ADNEM u odnosu na MHM što su ga spomenuti istraživači koristili (Anon., 2008.) različitog i kompleksnijeg kemijskog sastava. Najveći prirast tijekom završnog dijela tova od 27. do 35. dana ostvarili su pilići koji su jeli ADNEM stalno primiješan u krmnu smjesu, što je razvidno iz mikropokusa. U farm-skom pak uzgoju, pilići koji su jeli ADNEM ostvarili su već nakon sedam dana tova značajno veću tjelesnu masu, za 23,8 grama veća (182,5 g prema 158,7 g), što je bio po-kazatelj postizanja veće tjelesne mase na kraju tova (Michalczuk i sur., 2011.). Pri kraju tova, već u dobi 28 dana, pilići koji su jeli ADNEM postigli su tjelesnu masu (1555 g) koja se može neposredno iskoristiti za proizvodnju „grill“ pilića, dok se to ne može reći za piliće koji ga nisu jeli (1438 g). Zaključno možemo spomenuti da je ADNEM u cijelosti opravdao primjenu u tovu pilića, a vrijeme primjene u krmnoj smjesi mora biti određeno s obzirom na namjeru tova, bilo kratkotrajni ili produženi. Konačno treba ustvrditi da ADNEM dodan u količini 0,5% u krmnu smjesu tovnih pilića ne dovodi do porasta količine kolagena u prsnom mišićju pri duljem tovu (35 dana), ali rezultat mikropukusa ne može biti zaključan s obzirom na relativno mali broj pretraženih uzoraka i pokusnih skupina. Uz uspješniji tov pilića ADNEM ima i važnu gospodarsku vrijednost, jer smanjuje gubitke (približno 2%) koji nastaju pri standardnom mehaničkom odvajanju prsnog mišićja. Zahvala Istraživanja je dijelom omogućilo Valipile d.d., Ive Politea 62, 10361 Sesvetski Kraljevec. Također je potporu istraži-vanju dala šljunčara Atika d.o.o., Fra F. Pintarića 4, 33000 Virovitica. Literatura ANON. (1994): Norma HRN ISO 3496;1994 standard. Meso i
mesni proizvodi. Određivanje količine hidroksiprolina. ANON. (2008): Declaration, concerning common approaches to
water management, flood protection, hydropower utilization and nature and biodiversity conservation in the Drava River
basin. Symposium “Drava River Vision“, 23 to 25 September 2008., Maribor, Slovenia.
BECHTEL, P. J. (1986): Muscle development and contractile proteins. In: Bechtel PJ, ed., Muscle as Food. New York: Academic Press, pp. 1-35.
BONIFER, L. J., G. W. FRONING (1996): Chicken skin com-position as affected by aqueous washings. J. Food Sci., 61, 895-898.
CAVIT, L. C., C. M. OWENS, J. F. MEULLENET, R. K. GANDHAPUNENI, G. W. YOUM (2001): Rigor deve-lopment and meat quality of large and small broilers and the use of Allo-Kramere shear, needle puncture, and razor blade shear to measure texture. Poult. Sci., 80, 138 (Suppl. 1).
DAWSON, P. L., D. M., JANKY, M. G. DUKES, L. D. THOMPSON, S. A. WOODWARD (1987): Effect of post mortem boning time during simulated commercial processing on the tenderness of broiler breast meat. Poult. Sci., 66, 1331-1333.
Dodge, J. W., J. Stadelman (1959): Post-mortem agin gof poultry meat and its effect on the tenderness of rthe breast muscles. Food Technol., 13, 81-84.
DECUYPERE, E. (1979): The effect of incubation temperature patterns on morphological,
physiological and reproduction criteria in Rhode Island Red birds. Agricultura, 27, 216-230.
DECUYPERE, E., K. TONA, F. BAMELIS, C. CAREGHI, B. KEMPS, B. DE KETELAERE, J. DE BAERDEMAKER J., V. BRUGGEMAN (2002): Broiler breeders and egg factors interacting with incubation conditions for optimal hatchability and chick quality. Arch. Geflügelk Special Issue, 66, 56-57.
DEEMING, D. C. (1995): What is chicken quality? World. Poult., 11 (2), 20-23.
FILIPIČ, B., I. GRADIŠNIK, M. PETROVEC, A. PEREYRA, H. MAZIJA (2016): Could enhanced induction of IFN-γ like mo-lecules from chicken macrophages using Holocene minerals stimulate the immune response against Dermanyssus galinae infestation? Altex Procc. Linz 2016 – Eusaat 201,. pp. 34.
HULTIN, H. O. (1985): Characteristics of muscle tissue. In: Fennema OR, ed., Food Chemistry, 2nd ed. New York: Marcel Dekker, pp. 725-790.
IQBAL, J., N. MUKHTAR, Z. UR REHMAN, S. H. KHAN, T. AHMAD, M. S. ANJUM, R. H. PASHA, S. UMAR (2017): Effects of egg weight on the egg quality, chick quality, and broiler performance at the later stages of production (week 60) in broiler breeders. J. Appl. Poult. Res., 26:183-191.
JANJEČIĆ, Z., D. BEDEKOVIĆ, M. BALENOVIĆ, M. ANDRAŠEC, B. FILIPIČ, M. KREŠIĆ, H. MAZIJA (2017): Učinci aluvijalnog depozitnog nanosa erodiranih minerala primijenjenog u tovu pilića. Zbornik radova Peradrarski dani 2017., Šibenik, Hrvatka. 95-106.
KLASING, K. C., D. M. BARNES (1988): Decreased Amino Acid Requirements of Growing Chicks Due to Immunologic Stress. Journal of Nutrition, 118 (9), 1158-1164.
KOKIĆ, A. (2012): Utjecaj multikomponentnih holocenskih minerala (MHM) na tijek Marekove bolesti. Doktorski rad. Veterinarski fakultet Sveučilišta u Zagrebu. Zagreb, 2012.
KOLAR, K. (1990): Colorimetric determination of hydroxyproline as measure of collagen content in meat and meat products: NMKL collaborative study. J. Assoc. Off. Anal. Chem., 73, 54-57.

UČINAK DODATKA ALUVIJALNOG DEPOZITNOG HOLOCENSKOG MINERALA (ADNEM) U KRMU TOVNIH PILIĆA FARMSKOG UZGOJA NA TVORBU KOLAGENA U NJIHOVU MIŠIĆJU
82
LYON, C. E., W. E. TOWNSEND, R. L. WILSON JR. (1976): Objective color values of non-frozen and frozen broiler breasts and thighs. Poult. Sci., 55, 1307-1312.
MARCU, A., I. VACARU-OPRIŞ, G. DUMITRESCU, L. P. CIOCHINA, A. MARCU, M. NICULA, N. MARIOARA, P. IOAN, D. DOREL, K. BARTOLOMEU, M. COSMIN (2013): The influence of genetics on economic efficiency of broiler chickens growth. Animal Science and Biotechno-logies., 46 (2), 339-346.
MEDIĆ, H., S. VIDAČEK, N. MARUŠIĆ, A. JURIĆ, T. PETRAK, H. MAZIJA (2010): Multicomponent Holocene Mineral Compound Given in Feed does not Influence Quality of Chicken Meat. XIIth European Poultry Conference Tours, Francuska, 23-27.08.2010.
MENDES, M., D. ECMEL, E. ARSLAN (2007): Profile analysis and growth curve for body mass index of broiler chickens reared under different feed restrictions in early age. Arch. Tierz., 50, 403-411.
MICHALCZUK, M., M. STĘPIŃSKA, M. ŁUKASIEWICZ (2011): Effect of the initial body weight of Ross 308 chicken broilers on the rate of growth. Ann. Warsaw Univ. of Life Sc. SGGW, Anim. Sci., 49, 121-125.
MEIJERHOF, R. (2006): Chick size matters. World Poult., 22, 30-31.
N.R.C. (2005): Mineral tolerance of animals. 2nd ed. The National Academy, Washington, DC, 1-510.
OLVER, M. D. (1997):Effect of feeding clinoptilolite (zeolite) on the performance of three strains of laying hens. Br. Poult. Sci., 38 (2), 220-222.
PEARSON, A. M., R. B. YOUNG (1989): Muscle and Meat Biochemistry. San Diego, CA, AcademicPress.
SUGANYA, T., S. SENTHILKUMAR, K. DEEPA, J. MURALIDHARAN, P. SASIKUMAR, N. MURTHUSAMY (2016): Metal toxicosis in poultry – a review. Int. Sci. Environ. Technol., 5, 515-524.
ŠOOŠ, E., B. FILIPIČ, S. MIKO, H. MAZIJA (2015): Farmaceutski pripravak za pojačanje antiproliferativne i proapoptotske aktivnosti humanih interferona holocenskim mineralima. Hrvatski glasnik intelektualnog vlasništva 5, Konsenzualni patent PK20130650.
THIRULOGACHANDAR, M. E., M. RAJESWARI, S. RAMYA (2014): Assessment of heavy metal sin Gallus and their impacts on human. Int. J. Sci. Res. Pub., 4 (6), 1-8.
TONA, K., F. BAMELIS, B. De KETELAERE, V. BRUGGEMAN, V. M. B. MORAES, J. BUYSE, O. ONAGBEASAN, E. DECUYPERE (2003): Effects of egg stirage time on spread of hetch, chick quality, and chick juvenile growrth. Poult. Sci., 82, 736-741.
TONA, K.,O. ONAGBEASAN,B. De KETELAERE,V. BRUGGEMAN,E. DECUYPERE (2005): Interrelationship between chick quality parameters and the effect of individual parameter on broiler relative growth to 7 days of age. Arch. Geflügelk., 69 (2), S. 67-72.
TRKULJA, H. (2012): Utjecaj aluvijalnog mineralnog dodatka u hrani pilića na boju kostiju. Diplomski rad. Veterinarski fakultet Sveučilišta u Zagrebu. Zagreb, 2012.
VIDOVIĆ, M. (2011): Utjecaj aluvijalnog mineralnog dodatka u hrani na imuni odziv pilića cijepljenih protiv newcastleske bolesti. Diplomski rad. Veterinarski fakultet Sveučilišta u Zagrebu, 2011.
WOLANSKI, N. J., R. A. RENEMA, F. E. ROBINSON, V. L. CARENEY, B. I. FANCHER (2007): Relationship among egg charaxcteristics, chick measurements, and early growrth traits in ten broiler breeder strains. Poult. Sci., 86, 1784-1792.
EFFECT OF ADDING ALLUVIAL DEPOSIT HOLOCENE MINERALS (ADNEM) TO FARM BREEDING BROILER FEED ON COLLAGEN FORMATION IN THEIR MUSCLES
Summary
The present study continued investigating the favorable effects of admixing 0.5% alluvial deposit Holocene minerals (ADNEM) to fattening broiler feed, with the aim to identify optimal fattening period for its application considering productivity and collagen formation in breast muscles. The preliminary limited study had revealed better effect of adding 0.5% versus 1.0% ADNEM. Therefore, in this micro-experiment, 0.5% ADNEM was added to the initial, final I or final II feed mixture, with recording fattening performance indicators. Two comparable groups of broilers received ADNEM (0.5%) throughout the fattening period or did not receive it all. Study results showed ADNEM added to broiler feed during the first 11 days of fattening to be as effective as when added throughout the 35 days of fattening. Collagen formation in the two control groups of broilers was greater in the group that received ADNEM 0.5% throughout the fattening period. Considering the relatively small number of muscle samples analyzed, this result could not necessarily be attributed to the increased collagen formation but also to other factors, primarily ADNEM mineral contents, as well as to broiler age. The success of macro-experiment in farm broilers confirmed the observations from both preliminary experiments. The broilers that received ADNEM (34,900 broilers) achieved the European Production Efficiency Factor (EPEF) 342 versus 327 in those not receiving ADNEM, i.e. by 15 index units lower. In addition, at the age of 4 weeks, the respective body mass was 1555 g and 1438 g, suggesting the possible reduction in the length of fattening period with 0.5% ADNEM admixed to broiler feed. Most important was efficient mechanical separation of breast muscles, which could be completely performed in broilers with ADNEM added to feed, as compared with those without the addition of ADNEM where approximately 2% of muscles were clammy and unusable for marketing as fillets. The research should be continued to elucidate the factors that contribute to improved productivity of broiler fattening with the use of ADNEM. Key words: fattening broilers, tovni pilići, alluvial deposit Holocene minerals (ADNEM), feeding mixture, collagen

PERADARSKI DANI 2019. 83
INFEKCIJA KONZUMNIH NESILICA BAKTERIJOM MANNHEIMIA HAEMOLYTICA
Borka Šimpraga, Fani Krstulović, Luka Jurinović, Marina Tišljar, Marijana Sokolovć
Hrvatski veterinarski institut, Centar za peradarstvo, Zagreb, Hrvatska
Sažetak
U radu je opisana infekcija konzumnih nesilica bakterijom Mannheimia (M.) haemolytica u četiri jata konzumnih nesilica u intenzivnom i ekstenzivnom uzgoju. Temeljem kliničkih znakova bolesti (podatci dobiveni od vlasnika jata i farmi) te makroskopske patološkoanatomske pretrage iz organa nesilica uzorkovanih tijekom razudbe napravljena je bakteriološka pretraga. Vrsta M. haemolytica dokazana je biokemijskom karakterizacijom pomoću API nizova, testovima na katalazu, oksidazu i pokretljivost, mikroskopskom pretragom i rastom na MacConkey agaru. Prema rezultatima testa osjetljivosti vrsta M. haemolytica izdvojena iz organa nesilica iz sva četiri jata pokazala je osjetljivost prema norfloksacinu, enrofloksacinu, ciprofloksacinu i linkospektinu. Neosjetljivost prema ampicilinu i amoksicilinu pokazali su sojevi izdvojeni iz organa nesilica iz jata 1 i 2, dok je soj iz jata 4 pokazao neosjetljivost prema amoksicilinu s klavulanskom kiselinom, ampicilinu, amoksicilinu, oksitetraciklinu, tetraciklinu i cefalotinu. Ključne riječi: konzumne nesilice, infekcija, Mannheimia haemolytica, antimikrobna osjetljivost Uvod
Bolesti dišnog sustava čest su i značajan uzrok velikih eko-nomskih šteta u intenzivnoj peradarskoj proizvodnji izazva-nih povećanom smrtnošću i troškovima liječenja oboljele peradi (Ahmed i sur., 2016.; Jammel i sur., 2016.). Poslije-dica su infekcije bakterijskim i virusnim dišnim patogenima koji mogu biti i istodobno prisutni i utjecati jedni na druge (Marien i sur., 2007.; Siddique i sur., 2016., Zaheer i sur., 2017.). Bakterijske vrste kao što su Pasteurella multocida, Pasteurella gallinarum, Riemerella anatipestifer, Bordetella avium, Mannheimia haemolytica, Ornithobacter rhinotra-cheale, Klebsiella pneumoniae, Avibacterium paragallina-rum, Gallibacterium anatis, bakterija Escherichia coli i mnoge druge uzrokuju pojavu bolesti dišnog sustava u peradi (Hafez, 2002.; Saif i sur., 2003.; Pérez i sur., 2014.; Jones i sur.; 2013.). Vrsta Mannheimia haemolytica pripada rodu Mannheimia, jednom od 15 rodova obitelji Pasteurellaceae (Highlander, 2001.). Reklasifikacijom je nekadašnja Pasteurella haemo-lytica koja je sadržavala dva biovara temeljem razgradnje arabinoze – biovar A i trehaloze – biovar T preimenovana prema novom nazivlju, a u skladu s rezultatima DNA hibridizacije i sekvenciranja 16S RNA gena u vrstu M. haemolytica biovar A, dok su sojevi P. haemolytica biovar T
preimenovani u vrstu P. trehalosi koja je kasnije nazvana Bibersteinia trehalosi (De Ley i sur., 1990.; Dewhirst i sur., 1992..; Angen i sur., 1999.; Blackall i sur., 2007.). Serovarovi 1, 2, 5, 6, 7, 8, 9, 12, 13, 14 i 16 bivše vrste P. haemolytica pripali su vrsti M. haemolytica, serovarovi 3, 4, 10 i 15 vrsti B. trehalosi, a serovar 11 novoj vrsti M. glucosida (Mohamed i Abdelsalem, 2008.). Mannheimia haemolytica ime je dobila po njemačkom znanstveniku Walteru Mannheimu zaslužnom za doprinos novom nazivlju i reklasifikaciji porodice Pasteurellaceae (Mohamed i Abdelsalam, 2008.). To je gram-negativna, fakultativno anaerobna, nepokretna bakterija kokobacilarnog oblika, posjeduje enzime katalazu i citokrom-oksidazu i tvori kapsulu. Male sjajne kolonije izrasle na krvnom agaru nemaju mirisa i tvore usku zonu ß-hemolize. Raste na hranjivim podlogama obogaćenim krvlju ili serumom i na MacConkey agaru u aerobnim uvjetima kroz 24 sata pri 37 °C. Razgrađuje laktozu, saharozu, maltozu i d-ksilozu, a ne razgrađuje d-trehalozu i t-arabinozu (Oppermann i sur., 2017.). M. haemolytica posjeduje čitav niz čimbenika virulencije, a virulencija djelomice ovisi o imunom statusu domaćina (Highlander, 2001.). Dvanaest serovarova M. haemolytica međusobno se razlikuju u strukturi genoma, površinskim antigenima i drugim osobinama pa su Davies i sur. (1997.) na temelju istraživanja zaključili da su izolati M.
dr. sc. Borka Šimpraga, dr. med. vet., Hrvatski veterinarski institut, Centar za peradarstvo, Laboratorij za bakteriologiju, Heinzelova 55, 10000 Zagreb, Tel.: +385 (0)12440 212, email: [email protected]

INFEKCIJA KONZUMNIH NESILICA BAKTERIJOM MANNHEIMIA HAEMOLYTICA 84
haemolytica klonovi koji predstavljaju različite filogenetske linije. Imunološkom i biokemijskom karakterizacijom gen-skih proizvoda M. haemolytica identificirano je mnoštvo proteina i polisaharida koji su vrlo značajni za virulenciju. Najvažniji čimbenici virulencije su kapsula (Lo i sur., 2001.), različiti adhezini odnosno proteini vanjske membra-ne (Jaramillo i sur., 2000.), fimbrije i enzimi sialoglikopro-teaze i neuraminidaze koje potiču kolonizaciju pluća (Highlander, 2001.), endotoksin (Welch i sur., 1995.) i leukotoksin (Burrows i sur., 1993.). Kapsula omogućava otpornost na fagocitozu i komplement u serumu (Highlan-der, 2001.). Proteini vanjske membane i lipoproteini (fimbrijski adhezijski antigeni) potiču kolonizaciju pluća, a imaju ulogu i u otpornosti na komplement u serumu (Highlander, 2001.). Izvanstanični enzimi sialoglikoproteaze i neuraminidaze indirektno utječu na imuni odgovor domaćina i potiču kolonizaciju stanica pluća (Highlander, 2001.). Endotoksini oštećuju leukocite i imaju toksični učinak na endotelne stanice, a leukotoksini toksično djeluju na leukocite i stvarajući pore na staničnoj stijenci uzrokuju lizu leukocita. Enzimi iz liziranih leukocita i upalni citokini djeluju na slabljenje zaštitnih imunih mehanizama pluća te uzrokuju stvaranje upale i oštećenje plućnog tkiva. Mannheimia haemolytica uobičajena je mikroflora dišnog sustava peradi, no u uvjetima stresa izazvanog prenapuče-nošću, povećanom količinom amonijaka, povećanom ko-ličinom prašine onečišćene bakterijama, glađu, žeđi, premje-štanjem životinja, visokim ili niskim temperaturama te in-fekcijama virusa ili mikoplazmi može izazvati infekciju dišnog sustava i septikemiju ((Highlander, 2001.;.Malik i sur., 2005.; Mohamed i Abdel-Salam, 2008.; Nimmermark i sur., 2009.; Ali i sur., 2015.; Umar i sur., 2017.; Setta i sur., 2017.). Brisovi dušnika, sinusa i plućnog tkiva uginulih pi-lića bakteriološki su pretraženi na dokaz prisutnosti bakterija E. coli, Bordetella avium, Salmonella spp. i M. Haemolytica. Tijekom petogodišnjeg istraživanja od ukupno 600 uginulih pilića iz brisova njih 92 izdvojena je M. Haemolytica, što je ujedno bila i najčešće izdvojena vrsta (Malik i sur., 2005.). Pojavu bolesti s kliničkim simptomima sličnim pasterelozi u jatu rasplodnih nesilica opisali su Ali i sur. (2015.) koji su prvi dokazali spomenutu bakteriju kao primarni patogen. U jatu se pojavilo značajno smanjenje nesivosti i smrtnost od 4%. Makroskopskom patološkoanatomskom pretragom utvrđene su cijanoza krijeste i privjeska, upala zračnih vre-ćica, punokrvnost pluća, petehijska krvarenja na srcu, sple-nomegalija, nekrotična žarišta na jetri te egg-peritonitis. Bakteriološkom pretragom iz uzoraka organa izdvojena je M. haemolytica koja je dokazana biokemijskom karakteri-zacijom pomoću API nizova te mikroskopskom pretragom. Setta i sur. (2017.) opisali su pojavu bolesti u dva jata konzumnih nesilica linije WHITE SHAVER i Hi-Sex Brown dobi 195 dana u kojima je došlo do značajnog pada nesivosti, smanjenja potrošnje hrane od 40% te smrtnosti od 6%. Patološkoanatomskom pretragom utvrđeni su upala zračnih vrećica, perikarditis, perihepatitis, hepatomegalija i splenomegalija, punokrvnost pluća i crijeva i egg peritonitis. Tijekom razudbe uzorkovani su organi za bakteriološku pretragu kojom je na temelju hemolitičkih kolonija izraslih
na krvnom agaru i kolonija poraslih na MacConkey agaru, mikroskopske pretrage i molekularne identifikacije (PCR) dokazana vrsta M. haemolytica. Umar i sur. (2017.) opisali su pojavu dišnih simptoma u podno držanih konzumnih nesilica dobi 52 tjedna, koji su se očitovali otečenjem krijesta i sinusa, mukoznim iscjetkom iz kljuna, hropcima uz zelenkasti proljev. Jato nesilica držano je blizu farme mliječnih krava i bikova. Tijekom patološko-anatomske pretrage iz razuđenih nesilica uzorkovani su brisevi jetre, slezene, cekalnih tonzila, jajovoda, dušnika, pluća i srca te mukozni sadržaj iz kljuna i sinusa za bak-teriološku pretragu. Na temelju biokemijske karakterizacije hemolitičkih kolonija izraslih na krvnom agaru, razmazaka učinjenih iz bakterijske kulture i obojanih prema Gramu te molekularme identifikacije (PCR) dokazana je M. haemo-lytica. Test patogenosti učinjen je na embrioniranim ko-košjim jajima i na zdravim konzumnim nesilicama dobi 8 tjedana sojem M. haemolytica izdvojenim iz organa uginulih nesilica dobi 52 tjedna. U embrioniranim kokošjim jajima već 48 sati nakon inokulacije uočena je 100%-tna smrtnost, dok su pokusno zaražene nesilice pokazivale blage simptome u obliku anoreksije, otežanog disanja, mukoznog iscjetka iz kljuna i zelenkasti proljev. Simptomi su nestali nakon jednog dana i nije bilo uginuća. Iz dušnika, pluća, srca, jetre i cekalnih tonzila izdvojena je M. haemolytica. Analiza sekvence pokazala je 99%-tnu sličnost s rDNA iz goveđeg soja M. haemolytica serovar A2. U jatu od 170 mosušnih i 110 pekinških pataka koje su po-kazivale dišne simptome i proljev tijekom makroskopske patološkoanatomske pretrage uzorkovani su jetra, srce, sle-zena, pluća, mozak i crijeva za bakteriološku pretragu. Vrsta M. haemolytica izdvojena je iz 9 pataka, dok je P. multocida dokazana u organima 114 pataka (Afifi i El-Nesr, 2013.). Infekcije vrstom M. haemolytica (bivša Pasteurella haemo-lytica) rijetke su u ljudi. Opisan je slučaj komplikacije uzrokovane infekcijom premosnice na aorti vrstom P. haemolytica (sadašnja M. haemolytica) i ß-hemolitičkim streptokom serološke skupine C u 50-godišnjeg bolesnika (Rivera i sur., 1994.). Izvor infekcije nije utvrđen, no nakon uklanjanja premosnice i šestotjedne intravenske primjene ampicilina bolesnik se potpuno oporavio. Prvi slučaj bakterijskog krupa (bakterijski traheitis) opisan je u devetomjesečne djevojčice sa simptomima promuklog kašlja, otežanog disanja, visoke temperature i nesvjestice (Watanabe i sur.,1998.). Iako je najčešći uzočnik bakte-rijskog krupa bakterija Staphylococcus aureus, u opisanom slučaju iz sadržaja dušnika izdvojena je P. haemolytica. Izvor infekcije nije utvrđen. U radu je opisana infekcija konzumnih nesilica bakterijom M. haemolytica u četiri jata konzumnih nesilica u eksten-zivnom i intenzivnom uzgoju.
Materijali i metode
Tijekom zimskih mjeseci krajem 2017. i početkom 2018. godine u dva ekstenzivno držana uzgoja konzumnih nesilica pojavila su se sporadična uginuća nesilica praćena kratkotrajnim padom nesivosti.

INFEKCIJA KONZUMNIH NESILICA BAKTERIJOM MANNHEIMIA HAEMOLYTICA 85
U jatu 1 bilo je oko 50 ekstenzivno držanih konzumnih nesilica dobi od 7,5 mjeseci do 3 godine. Veći broj nesilica iz jata imao je izrazita otečenja u području glave i izražene neurološke simptome (tresanje glavom). Nisu poznati podatci o liniji nesilica, vakcinaciji i hranidbi jata. U jatu 2 ekstenzivno držane nesilice (nisu poznat broj, linija, podatci o vakcinaciji i hranidbi) u dobi od 24 tjedna do 1,5 godine pokazivale su nevoljkost, ubrzano disanje i nakostriješeno perje. Na farmi intenzivno držanih konzumnih nesilica linije Novogen Brown u jednoj nastambi (jato 3) od oko 70.000 ptica došlo je do pada nesivosti i povećane smrtnosti. Spomenuti simptomi u istoj nastambi uočeni su dvokratno i to u nesilica dobi 51 tjedan i u dobi 65 tjedana. Nesilice su držane u obogaćenim kavezima, hranjene komercijalnom izbalansiranom hranom i napajane vodom iz gradskog vodovoda. Cijepljene su protiv Marekove bolesti, zaraznog bronhitisa, newcastleske bolesti, zarazne bolesti burze, sindroma pada nesivosti i protiv bakterija Salmonella ser. Enteritidis i ser. Typhimurium. Zdravstveno stanje u jatu se nakon prvog i nakon drugog izbijanja simptoma nakon desetak dana normaliziralo. U jatu 4 intenzivno držanih 1.800 konzumnih nesilica linije Lohman Brown, dobi 22 tjedna uočeni su izraženi znaci potištenosti, kunjanja, pojave nosnog iscjetka i otečenosti privjeska. Nesilice su držane u kavezima, hranjene izbalansiranom komercijalnom hranom i napajane vodom iz gradskog vodovoda. Makroskopska patološkoanatomska pretraga Lešine konzumnih nesilica iz četiri jata razuđivane su u Laboratoriju za patologiju Centra za peradarstvo Hrvatskog veterinarskog instituta u Zagrebu u sklopu rutinske makro-skopske patološkoanatomske pretrage i na zahtjev vlasnika. Uzorci organa uzeti su za bakteriološku pretragu. Bakteriološka pretraga Uzorci organa sterilnom mikrobiološkom ušicom nacjepljeni su na čvrste hranjive podloge i to na Columbia agar (bio Merieux, Francuska), Columbia agar s dodatkom 5% defibrinirane ovčje krvi i na MacConkey agar (bio Merieux, Francuska). Agari su inkubirani u aerobnim uvjetima pri 37 °C tijekom 24 sata. Nakon inkubacije od sumnjivih kolonija poraslih na krvnom agaru napravljeni su razmasci i obojani prema Gramu. Rast kolonija uočen je i na pločama Mac Conkey agara. Napravljeno je ispitivanje na oksidazu, katalazu i pokretljivost te biokemijska karakterizacija pomoću API nizova ID 32E. Ispitivanje na oksidazu napravljeno je tako da su odabrane pojedinačne kolonije prebačene na filtar papir na koji je prethodno stavljena kap oksidaza reagensa (Oxidase reagent, bio Merieux, Francuska). Kao pozitivna kontrola korišten je referentni soj Pseudomonas aeruginosa ATCC 27853, a kao negativna kontrola referentni soj Escherichia coli ATCC 25922. Za ispitivanje na katalazu svaka odabrana kolonija uzeta je pomoću mikrobiološke ušice i stavljena u kapljicu
vodikovog peroksida na čisto predmetno stakalce. Kao pozitivna kontrola korišten je referentni soj Staphylococcus aureus ATCC 25923, a kao negativna kontrola referentni soj Enterococcus faecalis ATCC 23186. Za test pokretljivosti nekoliko kolonija čiste bakterijske kulture suspendirano je u 1 mL F.O. na predmetnom stakalcu i pregledano pod mikroskopom. Izdvojenoj bakteriji određena je osjetljivost prema antimikrobnim lijekovima disk-difuzijskom metodom na čvrstoj podlozi Mueller-Hinton agaru (Mueller-Hinton agar 2, bio Merieux, Francuska) s dodatkom 5% do 10% % ovčje krvi prema preporukama CLSI (2012.) prema kojima su očitavani rezultati. Kao kontrola korišteni su referentni sojevi spomenuti kod testova katalaze i oksidaze. Korišteni su sljedeći diskovi: ampicin (AMP) 10 µg, amoksicilin (AMX) 25 µg, amoksicilin s klavulanskom kiselinom (AMC) 20 µg + 10 µg, tetraciklin (TE) 30 µg, oksitetraciklin (OT), 30 µg, enrofloksacin (ENO) 5µg, ciprofloksacin (CIP) 5 µg, norfloksacin (NOR) 10 µg, cefalotin (CF) 30 µg i linkospektin (LS) 9 µg+100 µg.
Rezultati
U radu su prikazani rezultati bakteriološke i/ili makro-skopske patomorfološke pretrage konzumnih nesilica u kojih je izdvojena vrsta M. haemolytica. Iako se u pojedinim slu-čajevima na temelju makroskopske patološkoanatomskog nalaza ukazala potreba za dopunskim pretragama, učinjene su pretrage prema pisanom zahtjevu kupca (vlasnika). Razudbeni nalaz Razudbenim nalazom u nesilica iz jata 1 utvrđena je ka-heksija, izrazito otečenje u području oba oka i infraorbitalnih sinusa, konjunktivitis, zamućenje rožnice, sinusitis te juve-nilni jajnik. Razudbenim nalazom u konzumne nesilice iz jata 2 utvrđeni su konjunktivitis, rinitis, gnojnofibrinozna upala sluznice šupljine kljuna i jezika, gnojna upala sinusa, kataralna di-jelom gnojnofibrinska upala sluznice jednjaka, kataralna upala dušnika, gnojnofibrinski perikarditis, gnojnofibrinska upala zračnih vrećica, atrofija slezene i jajnika i kataralna upala crijeva. Razudbenim nalazom u svih 5 nesilica dobi 51 tjedan iz jata 3 utvrđeni su sepsa, perikarditis, miokarditis, kataralno-hemoragični enteritis te ooforitis. U četiri nesilice utvrđen je splenitis, u tri splenomegalija, gnojnofibrinozna pneumonija, kataralno-fibrinozni traheitis te nekrotični hepatitis i ruptura jetre. Razudbenim nalazom 5 nesilica dobi 65 tjedana iz jata 3 utvrđeni su hepatomegalija, degeneracija srčanog mišića i nekrotični miokarditis, hemoragična upala dušnika, fibroza zračnih vrećica, tumor slezene, upala jajnika i jajovoda te fibroza gušterače s diseminiranim submilijarnim nekrotič-nim žarištima. Razudbenim nalazom u nesilica iz jata 4 utvđeni su kaheksija, edem privjeska, gnojni sinusitis, sto-matitis, glositis, kataralni dijelom mukopurulentni laringo-traheitis, punokrvnost i edem pluća, gnojnofibrinski poli-serozitis, hiperplazija i degeneracija jetre, hiperplazija bubrega, egg-peritonitis i upala jajovoda.

INFEKCIJA KONZUMNIH NESILICA BAKTERIJOM MANNHEIMIA HAEMOLYTICA 86
Tablica 1. Patološkoanatomska dijagnoza (jato 1-jato 4) Table 1. Pathoanatomical diagnosis (flock 1-flock 4)
Jato Flock Dišni sustav
Respiratory system Probavni sustav
Gastrointestinal system
Srčanožilni susta Cardiovascular
system
Limfoidni sustav
Lymphoid system
Sustav organa za
razmnožavanje Reproductive
system
Mokraćni sustav Urinary system
1 Periorbitalni edem, konjunktivitis, zamućenje rožnice, kaheksija / Periorbital edema, conjunctivitis, cloudy cornea, cachexia
Gnojno-fibrinozni sinusitis / Purulent-fibrinous sinusitis
Juvenilni jajnik Juvenile ovary
2 Konjunktivits / Conjunctivitis
Rinitis, gnojni sinusitis, kataralni traheitis, gnojno-fibrinozni celulitis / Rhinitis, purulent sinusitis, catarrhal tracheitis, purulent-fibrinous cellulitis
Gnojno-fibrinozni stomatitis i glositis, kataralni enteritis / Purulent-fibrinous stomatitis and glossitis, catarrhal enteritis
Gnojno-fibrinozni perikarditis / Purulent-fibrinous pericarditis
Atrofija slezene / Spleen atrophy
Atrofija jajnika / Ovarian atrophy
3* __ Gnojno-fibrinozna upala pluća, gnojno-fibrinozni/hemoragični traheitis, fibroza zračnih vrećica / Purulent-fibrinous pneumonia, purulent-fibrinous/hemorrhagic tracheitis, air sac fibrosis
Kataralno-hemora-gični enteritis, nekrotični hepatitis/hepato-megalija, fibroza/nekroza gušterače / Catarrhal-hemorrhagic enteritis, necrotic hepatitis/hepatomegaly, pancreatic fibrosis/necrosis
Gnojno- fibrinozni perikarditis, degeneracija miokarda, nekrotični miokarditis / Purulent-fibrinous pericarditis, myocardial degeneration, necrotic myocarditis
Hiperplazija slezene, nekrotični splenitis / Spleen hyperplasia, necrotic splenitis
Ooforitis i oviduktitis / Oophoritis and oviductitis
4 Edem privjesaka, kaheksija Edema of appendages, cachexia
Gnojni sinusitis, kataralni mukopurulentni laringotraheitis, punokrvnost i edem pluća, gnojno-fibrinozni celulitis / Purulent sinusitis, catarrhal mucopurulent laryngotracheitis, pulmonary hemorrhage and edema, purulent-fibrinous cellulitis
Gnojni stomatitis i glositis, hiperplazija jetre / Purulent stomatitis and glossitis, liver hyperplasia
Egg peritonitis, ooforitis / Egg peritonitis, oophoritis
Hiperplazija bubrega / Kidney hyperplasia
*Jato 3 – patološkoanatomska dijagnoza ukazuje na sepsu (v. Razudbeni nalaz) / *Flock 3 – pathoanatomical diagnosis indicating sepsis (see necropsy finding)
Bakteriološkom pretragom je iz oba oka, mozga i zračnih vrećica nesilica iz jata 1 izdvojena bakterija M. haemolytica. Pretraga za dokaz bakterija Avibacterium paragallinarum i Ornithobacter rhinotracheale završila je negativnim re-zultatom.
Pretragom sadržaja gnoja iz sinusa, jetre, ovarija i jajovoda nesilice iz jata 2 izdvojene su bakterije Mannheimia haemolytica i E. coli. Bakteriološkom pretragom organa konzumnih nesilica iz jata 3 tijekom prvog izbijanja bolesti od 4 jetre, 3 slezene, 4

INFEKCIJA KONZUMNIH NESILICA BAKTERIJOM MANNHEIMIA HAEMOLYTICA 87
pluća, 5 srca, 1 bubrega, 2 dušnika, 5 ovarija i 5 crijeva M. haemolytica izdvojena je iz 3 pluća, 5 srca i 2 dušnika. Pretragom organa (3 jetre, 3 slezene, 3 srca, 3 ovarija i 3 cri-jeva) nesilica tijekom drugog izbijanja bolesti M. haemo-lytica izdvojena je iz 2 jetre, 2 slezene, 2 srca, 3 ovarija i 3 crijeva. Uz M. haemolytica izdvojena je bakterija Hafnia alvei. Pretragom organa (4 glave, 4 jetre, 4 ovarija, 4 srca, 4 slezene i 1 slijepog crijeva) nesilica iz jata 4 M. haemolytica uz E. coli izdvojena je iz 3 uzorka gnojnog sadržaja iz sinusa, 2 jetre, 2 ovarija i 2 srca. Čista bakterijska kultura M. haemolytica porasla na pločama krvog agara organima nesilica iz sva 4 jata biokemijski je pomoću API nizova ID 32E identificirana kao M. haemo-lytica. Porast kolonija uočen je i na pločama MacConkey agara. U razmazima učinjenim iz bakterijske kulture nađeni su gram-negativni kokobacili. Test na tvorbu katalaze i oksidaze bio je pozitivan, a na pokretljivost negativan. Prema rezultatima testa osjetljivosti sojevi vrste M. haemo-lytica izdvojeni iz organa nesilica iz sva četiri jata pokazali su osjetljivost prema norfloksacinu, enrofloksacinu, cipro-floksacinu i linkospektinu. Neosjetljivost prema ampicilinu i amoksicilinu pokazali su sojevi izdvojeni iz organa nesilica iz jata 1 i 2, dok je soj iz jata 4 pokazao neosjetljivost prema amoksicilinu s klavulanskom kiselinom, ampicilinu, amoksi-cilinu, oksitetraciklinu, tetraciklinu i cefalotinu. Rasprava i zaključci
Infekcija bakterijom M. haemolytica zabilježena je i opisana u pilića te u konzumnih i rasplodnih nesilica (Malik i sur., 2005.; Ali i sur., 2015.; Umar i sur., 2017.; Setta i sur., 2017.). U radu je opisana infekcija bakterijom M. haemo-lytica u četiri jata konzumnih nesilica u intenzivnom i
ekstenzivnom uzgoju. Temeljem kliničkih znakova bolesti (podatci dobiveni od vlasnika jata i farmi) te makroskopske patološkoanatomske pretrage iz organa uginulih nesilica uzorkovanih tijekom razudbe napravljena je bakteriološka pretraga. Vrsta M. haemolytica izdvojena je iz oba oka, mozga i zračnih vrećica u nesilica iz jata 1. U jatu 2 do-kazana je uz E. coli u svim pretraženim organima (jetra, ovarij, jajovod, sadržaj gnoja iz sinusa). U jatu 3 tijekom prvog izbijanja bolesti M. haemolytica izdvojena je iz svih pet pretraženih srca, tri od četiri pluća i oba dušnika. Tijekom drugog izbijanja bolesti M. haemolytica (uz bakteriju H. alvei) izdvojena je iz dvije od tri jetre, dvije od tri slezene, dva od tri srca te iz sva tri ovarija i tri crijeva. U nesilica iz jata 4 M. haemolytica izdvojena je iz tri od četiri uzorka gnoja iz sinusa, iz dvije od četiri jetre, dva od četiri ovarija i dva od četiri srca. Spomenuta vrsta dokazana je biokemijskom karakterizacijom pomoću API nizova, mikro-skopskom pretragom, testovima na katalazu i oksidazu, pokretljivosti te rastom na MacConkey agaru. Takva potvrda vrste M. haemolytica u skladu je s bakteriološkom pre-tragom i potvrdnim testovima opisanim u člancima drugih autora (Malik i sur., 2005.; Ali i sur., 2015.; Umar i sur., 2017.; Seta i sur., 2017.). Pojava infekcije bakterijom koja je uobičajeno prisutna u gornjim dišnim putovima u konzumnih nesilica u ekstenzivnom i intenzivnom načinu držanja i koja najčešće igra ulogu sekundarnog patogena može se objasniti utjecajem stresa, visoke koncentracije amonijaka, visokom ili niskom temperaturom, premješta-njem ili pak jakom infestacijom parazita (Theurer i sur., 2013.). Svi nabrojeni čimbenici imaju ulogu u patogenezi vrste M. haemolytica. U dijagnostici su učinjene bakterio-loška i mikroskopska pretraga, biokemijska karakterizacija, potvrdni testovi za katalazu i oksidazu te test pokretljivosti. Od deset antimikrobnih lijekova na koje je testirana osjet-
Tablica 2. Rezultati bakteriološke pretrage (jato 1-jato 4) Table 2. Bacteriological testing results (flock 1-flock 4)
Jato / Flock
Broj pretraženih životinja
Number of animals examined
Broj pretraženih organa Number of organs examined
Bakteriološka identifikacija Bacteriological identification
1 1 2 oka, mozak, zračne vrećice / 2 eyes, brain, air sacs M. haemolytica
2 1 Uzorak gnojnog sadržaja iz sinusa, jetra, ovarij, jajovod Sample of purulent sinus content, liver, ovary, oviduct
M. haemolytica i/and E. coli
3 (prvo izbijanje) (first outburst) 5
5 srca, 5 ovarija, 4 jetre, 4 pluća, 3 slezene, 1 bubreg, 2 dušnika, 5 crijeva / 5 hearts, 5 ovaries, 4 livers, 4 lungs, 3 spleens, 1 kidney, 2 tracheae, 5 intestines
M. haemolytica (5 srca, 3 pluća, 2 dušnika) (5 hearts, 3 lungs, 2 tracheae
3 (drugo izbijanje) (second outburst) 3
3 srca, 3 ovarija, 3 jetre, 3 slezene, 3 crijeva 3 hearts, 3 ovaries, 3 livers, 3 spleens, 3 intestines
M. haemolytica i/and H. alvei (2 srca, 3 ovarija, 2 jetre, 2 slezene, 3 crijeva) 2 hearts, 3 ovaries, 2 livers, 2 spleens, 3 intestines
4 4
4 uzorka gnojnog sadržaja iz sinusa, 4 srca, 4 jetre, 4 ovarija, 4 slezene, 1 slijepo crijevo 4 samples of purulent sinus content, 4 hearts, 4 livers, 4 ovaries, 4 spleens, 1 appendix
M. haemolytica i/and E. coli (3 uzorka gnojnog sadržaja iz sinusa, 2 srca, 2 jetre, 2 ovarija) 3 samples of purulent sinus, 2 hearts, 2 livers, 2 ovaries

INFEKCIJA KONZUMNIH NESILICA BAKTERIJOM MANNHEIMIA HAEMOLYTICA 88
ljivost vrste M. haemolytica sojevi izdvojeni iz organa nesilica iz sva četiri jata pokazali su osjetljivost prema norfloksacinu, enrofloksacinu, ciprofloksacinu i linkospek-tinu. Ovakvi rezultati djelomice su u suglasju s podatcima iz literature (Malik i sur., 2005.; Ali i sur ., 2015.). U istra-živanju Malika i sur. (2005.) uočena je vrlo visoka neosjetljivost sojeva M. haemolytica prema tetraciklinu, dok je u opisanom istraživanju prema tetraciklinu neosjetljivost pokazao samo soj M. haemolytica iz jata 4. Ujedno je soj iz jata 4 pokazao višestruku neosjetljivost i to prema pet antimikrobnih lijekova, što je djelomice u suglasju s istra-živanjem Setta i sur. (2017.) koji su utvrdili neosjetljivost izolata M. haemolytica prema svim testiranim antimi-krobnim lijekovima. Soj izdvojen iz organa nesilica iz jata 3 pokazao je osjetljivost prema svim testiranim antimikrobnim lijekovima. Kako je rast vrste M. haemolytica često inhibiran rastom drugih bakterija poželjno je zbog diferencijalne dijagnoze i znatno većeg postotka pozitivnih uzoraka u laboratoriju provoditi molekularne dijagnostičke metode (Kugadas i sur., 2014.).
Literatura
AFIFI, I. SAMIA, K. A. EL-NESR (2013): Bacteriological, pathological and immunohistochemical Studies on Pasteurella spp. isolated from ducks in Beni Suef Governorate. Egypt. J. Comp. Path&Clinic Path., 26 (1), 28-48.
AHMED, S., M. M. LAPIA, L. FENG, C. ZHEFENG, F. WEIMIN (2016): Novel Advancements in Diagnosis of Periprosthetic Joint Infection. Sci. Lett., 4, 159-164
ALI, A., N. SIDDIQUE, M. A ABBAS, A. GHAFAR, S. RAFIQUE, R. ALI, AUR MEMON, K. NAEEM (2015): Role of Mannheimia (Pasteurella) haemolytica in severe respiratory tract Infection in commercial Poultry in Pakistan. Pak. Vet. J., 35 (3), 279-282.
ANGEN, Ø., R. MUTTERS, CAUGANT A. DOMINIQUE, J. E. OLSEN, M. BISGAARD (1999): Taxonomic relationships of the [Pasteurella] haemolytica complex as evaluated by DNA-DNA hybridizations and 16S rRNA sequencing with proposal of Mannheimia haemolytica gen.nov. ,comb, nov., Man-nheimia granulomatis comb.nov., Mannheimia glucosida sp. nov., Mannheimia ruminalis sp. nov. and Mannheimia varigena sp. nov. International Journal of Systematic and Evolutionary Microbiology, 49, 67-86,
BLACKALL, P. J., A. M. BOJESEN, H. CHRISTENSEN, M. BISGAARD (2007): Reclassification of [Pasteurella] trehalosi as Bibersteinia trehalosi gen. nov., comb. nov. International Journal of Systematic and Evolutionary Microbiology, 57 (4), 666-674.
BURROWS, L. L., E. OLAH-WINFIELD, R. Y. LO (1993): Molecular analysis of the leukotoxin determinants form Pasteurella haemolitica serotypes 1 to 16. Infect. Immun., 61, 5001-5007.
Clinical and Laboratory Standards Institute (CLSI) 2012: Per-formance standards for antimicrobial susceptibility testing; Twenty-Second Informational Supplement. M100-S22, Vol.32 No.3; Replce M100-S21, Vol.31 No.1.
DAVIES, R. L., S. ARKINSAW, R. K. SELANDER (1997): Evo-lutionary genetics of Pasteurella haemolitica isolates re-covered from cattle and sheep. Infect. Immun., 65, 3585-3593.
DEWHIRST, F. E., B. J. PASTER, I. OLSEN, G. J. FRASER (1992): Phylogeny of 54 representative strains of species in the family Pasteurellaceae as determined by comparison of 16S rRNA sequences. J. Bacteriol., 174, 2002 -2013.
HAFEZ, M. H. (2002): Diagnosis of Ornithobacterium rhino-tracheale. International J of Poultry Sci., 1, 114-118.
HIGHLANDER, S. K. (2001): Molecular genetic analysis of virulence in Mannheimia (Pasteurella) haemolytica. Front. Biosci., 6, 1128-1150.
JAMEEL, A. A. B, B. R. ZHOU, D. LUO (2016): Drug therapy for Hidradenitis supprativa:an update. Sci. Lett., 4, 108-117.
JARAMILLO, L., F. DIAZ, P. HERNANDEZ, H. DEBRAY, F. TRIGO, G. MENDOZA, E. ZENTENO (2000): Purification and characterization of an adhesin from Pasteurella haemo-litica. Glycobiology, 10, 31-37.
JONES, K. H., J. K. THORNTON, Y. ZHANG, M. J. MAUEL (2013): A 5-year retrospective report of Gallibacterium anatis and Pasteurella multocida isolates from chickens in Missis-sippi. Poultry Sci., 92, 3166–3171
KUGADAS, A., J. POINDEXTER, M. L. LEE, J. BAVANANTHASIVA, D. R. CALL, K. A. BRAYTON, S. SRIKUMARAN (2014): Growth of Mannheimia haemolytica: Inhibitory agents and putative mechanism of inhibition. Vet. Microbiol., 174, 155-162.
DE LEY, J., W. MANNHEIM, R. MUTTERS, K. PIECHULLA, R. TYTGAT, P. SEGERS, l M. BISGAARD, W. FREDERIKSENP, K. H. HINZ, M. VANHOUCKE (1990): Inter- and Intrafamilial Similarities of rRNA Cistrons of the Pasteurellaceae. Int. J. Syst. Bacteriol., 40, 126-137.
LO, R. Y., L. J. MCKERRAL, T. L. HILLS, M. KOSTRZYNSKA (2001): Analysis of the capsule biosynthetic locus of Mannheimia (Pasteurella) haemolytica, A1 and proposal of a nomenclature system. Infect. Immun., 69, 4458-4464.
MALIK, Y. S., Y. CHANDER, S. C. GUPTA, S. M. GOYAL (2005): A Retrospective Study on Antimicrobial Resistance in Mannheimia (Pasteurella) haemolytica, Escherichia coli, Salmonella Species, and Bordetella avium from Chickens in Minnesota. The Journal of Applied Poultry Research, 14, 506–511, https://doi.org/10.1093/japr/14.3.506
MARIEN, M., A. DECOSTERE, L. DUCHATEAU, K. CHIERS, R. FROYMAN, H. NAUWYNCK (2007): Efficacy of enro-floxacin, florfenicol and amoxicillin against Ornithobacterium rhinotracheale and Escherichia coli O2:K1 dual infection in turkeys following APV priming. Vet. microbiol., 121, 94-104.
MOHAMED, R. A., E. B. ABDELSALAM (2008): A review on pneumonic pasteurellosis (respiratory mannheimiosis) with emphasis on pathogenesis, virulence mechanisms and predisposing factors. Bulgarian Journ. of Vet. Med., 11,139−160.
NIMMERMARK, S., L. VONNE, G. GÖRAN, E. WIJNAND (2009): Ammonia, dust and bacteria in welfare-oriented systems for laying hens. Ann. Agric. and Environ. Med., 16, 103-113.
OPPERMANN, T., N. BUSSE, P. CZEZMAK (2017): Man-nheimia haemolytica growth and leukotoxin production for vaccine manufacturing - A bioprocess review. Electornic Journal of Biotehnology, 28, 95-100.
PÉREZ, M. V. M., J. L. OCHOA, C. V. CRUZ, P. S. ALONSO, G. OLMEDO-ALVAREZ, S. VACA, E. N. ABASCAL (2014) Isolation of Actinobacillus pleuropneumoniae from Layer

INFEKCIJA KONZUMNIH NESILICA BAKTERIJOM MANNHEIMIA HAEMOLYTICA 89
Hens Showing Clinical Signs of Infectious Coryza. Avian Dis., 58, 638-641.
RIVERA, M, G. C. HUNTER, J. BROOKER, C. W. O'BERG, S. H. SMYTHE, V. M. BERNHARD (1994): Aortic graft infection due to Pasteurella haemolytica and group C beta-hemolitic streptococcus. Clin. Infect. Dis., 19, 941-943.
SAIF, Y. M., H. J. BARNES, J. R. GLISSON, A. M. FADLY, L. R. MCDOUGALD, D. E. SWAYNE (2003): Diseases of poultry 11th end. Iowa State University Press. Blackwell Publishing Company, London, United Kingdom, 849.
SETTA, A., E. RAFAELI, M. SALEM HEBA (2017): Man-nheimia (Pasteurella) Haemolytica infection in commercial layers; A case repor. J. Egypt Vet. Med. Assac., 77, 241-246.
SIDDIQUE, N., N. KHALID, A. ZAHEER, A. M. ATHAR, A. AKBAR, R. SABA, R. FAROOQ, I. BEGUM, F. RASHID (2016): Isolation and Sequence Analysis of Reassortant Low Pathogenic Avian Influenza Virus H4N6 from Duck and Chicken in Live Bird Markets from Pakistan. Pak. Vet. J., 36, 258-263.
THEURER, M. E., D. E. ANDRESON, B. J. WHITE, M. D. MIESNER, D. A. MOISER, J. F. COETZEE, J. LAKRITZ, D.
E. AMRINE (2013): Effect of Mannheimia haemolitica pneumoniae on behavior and physiologic responsen of calves during high ambient environmental temperatures. J. Anim. Sci., 91, 3917-3929.
UMAR, S., S. ASIF, A. RIAZ, M. A. ABDULLAH SHAH, M. L. SOHAIL (2017): Isolation of Mannheimia haemolitica from layer hens showing respiratory clinical signs. Pak. Vet. J. ISSN:0253-8318 (PRINT), 2074-7764 (ONLINE):Accessible at:www.pvj.com.pk.
WATANABE, T., M. SATO, T. ABE, Y. ODA (1998): Bacteria croup caused by Pasteurella haemolytica. Acta Paediatrica Japonica, 40, 360-361.
WELCH, R. A., M. E. BAUER, A. D. KENT, J. A. LEEDS, M. MOAYERI, L. B. REGASA, D. L. SWENSON (1995): Battling against host phagocytes:the wherefore of the RTX family of toxins? Infect. Agents Dis., 4, 254-272.
ZAHEER, Z., I. HUSSAIN, S. U. RAHMAN, T. YOUNAS, I. ZAHEER, G. ABBAS, M. NASIR (2017): Occurrence and antibiotic susceptibility of methicillin – resistant. Staphylo-coccus aureus recovered from oropharynx of live cockerels. Pak. Vet. J., 37, 108-110.
LAYING HEN INFECTION WITH MANNHEIMIA HAEMOLYTICA
Summary
Mannheimia (M.) haemolytica infection in four flocks of laying hens on intensive and extensive rearing is described. Bacteriological examination was based on the clinical signs of disease (data reported by the owners of the flocks and farms) and pathoanatomical examination of laying hen organs sampled during necropsy. M. haemolytica was identified by biochemical characterization using API strips, catalase, oxidase and motility tests, microscopy and growth on MacConkey agar. Results of the antimicrobial sensitivity testing revealed the M. haemolytica strain isolated for laying hen organs from all four flocks to be sensitive to norfloxacin, enrofloxacin, ciprofloxacin and lincospectin. The bacterial strains isolated from laying hen organs from flocks 1 and 2 exhibited resistance to ampicillin and amoxicillin, whereas the strain isolated from flock 4 showed resistance to amoxicillin with clavulanic acid, ampicillin, amoxicillin, oxytetracycline, tetracycline and cephalothin. Key words: laying hens, infection, Mannheimia haemolytica, antimikrobial sensitivity

PERADARSKI DANI 2019. 90
ASSESSMENT OF MOLECULAR DIVERSITY OF SOME POTENTIAL PATHOGENIC E. COLI ISOLATED BY ERIC-PCR SCREENING
Camelia Tulcan1, Marijana Sokolović2, Sorina Popescu1, Ioan Hutu1, Simona Marc1, Oana Maria Boldura1*, Calin Mircu1
1Faculty of Veterinary Medicine, Timisoara, Banat's University of Agricultural Science and Veterinary Medicine "King Michael I of Romania", Timisoara, Romania 2Croatian Veterinary Institute, Poultry Centre, Zagreb, Croatia
Summary
The majority of Escherichia (E.) coli strains are considered as harmless commensals in both human and animals. However, certain isolates can be a source of resistant genes and/or can cause infections. The emergence of multidrug resistance in E. coli has become a worldwide problem, resulting in numerous researches on antimicrobial resistance, virulence of bacterial poultry pathogens and various epidemiological studies. Use of molecular methods in these studies offer the promise of a more complete determination of the potential risk of poultry originated E. coli isolates to poultry and human health. Data on genetic diversity of bacterial strains have thus emerged as an important tool for revealing different patterns of antibiotic resistance. The aim of this study was to use molecular approach in defining the zoonotic potential of chicken-originated E. coli isolates. Total genomic DNA was purified from liquid cultures of E. coli strains isolated from chicken from different farms. Molecular diversity studies of E. coli isolates were conducted using Enterobacterial Repetitive Intergenic Consensus (ERIC) – ERIC 1 and ERIC 2 molecular markers and polymerase chain reaction (PCR) fingerprinting method. Based on molecular data collected from the fingerprinting experiment, a phylogeny Unweighted Pair Group Method with Arithmetic mean (UPGMA) dendrogram showing genetic relationship among the isolates was developed. This paper presents an initial step towards development of a genetic diversity screening method for detection of potential pathogenic E. coli strains, based on specific molecular markers. The data obtained in this study could be used in grouping multiple bacterial strains according to the development of antimicrobial resistance mechanisms. Furthermore, this approach will be used in the assessment of the degree of the aforementioned risk to public health, as well as for developing strategies that will prevent spreading of E. coli infections. Key words: Escherichia coli, PCR, ERIC sequences, pathogenic strains, variability Introduction
Escherichia (E.) coli is a gram-negative facultative, anaerobic, rod-shaped bacterium commonly found in the digestive system of warm-blooded organisms. The E. coli species comprises ubiquitous non-pathogenic commensal strains and pathogenic strains characterized by the presence of specific colonization factors, virulence factors and pathogenicity associated genes. E. coli strains are classified into more than 380 serotypes according to their heat-stable somatic (O) and flagellar (H) antigens (Karmali et al., 2010). The pathogenic nature of specific strains is better evident in classification based on the type of virulence factor present in the E. coli genome and the ability to cause disease
in the host organism. Among ten pathotypes (belonging to two main groups, diarrheagenic and extraintestinal E. coli), certain strains express their pathogenicity by the production of toxins (Cowden et al., 2001; Chang et al., 2004; Karmali et al., 2010; Croxen and Finlay, 2010; 2013). Although more than 100 serotypes highly pathogenic to humans have been identified (Tozzi et al., 2003; Beutin et al., 2004; Lynn et al., 2005), certain serotypes such as E. coli O157:H7 are more often linked with foodborne outbreaks (Nataro and Kaper, 1998; Müller et al., 2001; Karmali et al., 2010; Croxen et al., 2013). Generally, pathogenic potential of E. coli is correlated with the presence of different virulence factors resulting in various enteric and extraintestinal infections (Olsen et al., 2002). E. coli infections usually
*corresponding author e-mail: [email protected]

ASSESSMENT OF MOLECULAR DIVERSITY OF SOME POTENTIAL PATHOGENIC E. COLI ISOLATED BY ERIC-PCR SCREENING
91
result from the consumption of contaminated food/feed and/or water, or through contaminated environment (Igarashi et al., 1999). Intestinal tracts of animals (mainly ruminants) are considered to be the natural reservoir of pathogenic E. coli. Nevertheless, virulent E. coli strains have been isolated from a wide diversity of animals including cattle, poultry, pigs, sheep and goats (Lynn et al., 2005; Ateba et al., 2008; Clements et al., 2012). Avian pathogenic E. coli (APEC) has been identified as the main cause of colibacillosis (a syndrome characterized by multiple organ lesions) in poultry farms (Know et al., 2008; Solà-Ginés et al., 2015). It is a common disease in poultry flocks around the world, especially if raised in intensive farming systems (Chansiripornchai, 2009). Rahman et al. (2004) found avian colibacillosis to be widely prevalent in all age groups of chickens (9.52%-36.73%), with a parti-cularly high prevalence rate in adult layer birds (36.73%). Data from other authors show greater susceptibility in younger animals. It is often reported as a cause of mortality on chicken farms worldwide representing an economically important problem of poultry production (Johnston, 2007; Know et al., 2008; Schouler, 2012; Nolan, 2013; Geetha and Palanivel, 2018). Unlike enteric pathotypes, APEC spread to different internal organs and can cause air sacculitis, pericarditis, peritonitis, salpingitis, synovitis, osteomyelitis and/or yolk sac infection (Barnes and Gross, 1997; La Ragione and Woodward, 2002; Ewers et al., 2004; De Carli et al., 2015). It has been documented that APEC isolates share virulence genes and clonal backgrounds, while certain studies suggest that there is a genome similarity to human enterotoxigenic E. coli (Dziva et al., 2013). Considering the fact that there is a potential for high proportion of resistant bacteria in the poultry fecal microbiota (Chan et al., 2012) and that during slaughter meat can be easily contaminated with E. coli of poultry origin (Lutful Kabir, 2010), it is reasonable to question whether such strains can be a source of infection for final consumer (Khoo et al., 2010; Mitchell et al., 2015). Furthermore, even if most E. coli serotypes are not considered pathogens, they may become opportunistic pathogens causing infections in immunocompromised hosts. Modern principles and practices in intensive farming sys-tems often include usage of different additives in feedstuffs for the food-producing animals. Despite the dramatic change in food producing animal and poultry industries, previous extensive use of certain antibiotics have contributed to creation of a selection pressure and development of multidrug resistance among E. coli strains. Therefore, we are facing now increased E. coli virulence and, consequently, a high rate of treatment failures and large economic losses (Olarinmoye et al., 2013). In the last decades, several methods for tracking bacterial sources have been developed for deciphering the relationships between E. coli strains from different sources by comparing DNA fingerprinting data (Dundas et al., 2001; Brocchi et al., 2006). Among advanced molecular genetics technologies for de-tection and identification of different E. coli strains, the Enterobacterial Repetitive Intergenic Consensus Polymerase Chain Reaction (ERIC-PCR) has been proved to be of great
value because it is a fast, accurate and efficient DNA finger-print identification method due to the proven discriminatory power. ERIC sequences are reduced (127 bp) non-coding regions found in Enterobacteriaceae (Sharples and Lloyd, 1990; Hulton et al., 1991; Wilson and Sharp, 2006). ERIC-PCR uses any primer combination designed for the ERIC conserved region to generate an electrophoretic molecular fingerprint model, depending on the frequency and orien-tation of the ERIC sequences in a bacterial genome. Mole-cular genotyping by ERIC-PCR is faster and more cost-effective than pulse field gel electrophoresis or multilocus sequence typing for generating information on genetic re-latedness of bacterial strains. For many organisms, it has a higher discriminatory power than other rapid fingerprinting techniques, resulting in increased frequency of use in routine epidemiological studies (Johnson and O'Bryan, 2000; Dundas et al., 2001; Saxena et al., 2002). In this study, the ERIC-PCR method was used to amplify random DNA regions that are flanked by conserved sequen-ces to generate genetic fingerprints specific for E. coli pathogenic isolates. This paper presents an initial study with the goal of developing a method for screening genetic diversity of potentially pathogenic E. coli strains. It is based on detection of specific molecular markers, using the genetic diversity of bacterial strains.
Materials and methods
Biological material For this initial study, ten isolates of E. coli strains collected from ten different poultry farms were used as biological ma-terial. The strains were collected from Ross broiler hybrids before slaughter without clinical signs of any disease. The strains were analyzed in previous studies and some charac-terization in respect to virulence profile was available (Table 1). For the purpose of this investigation, the E. coli isolates allocated with codes ranging from Ec1 to Ec10 were analyzed. Table 1. Virulence profile of selected genes for E. coli
strains analyzed in this study
Sample Serotype (identified gene) Ec1 eae (intimin producing gene) Ec2 eae (intimin producing gene) Ec3 eae (intimin producing gene) Ec4 no available information Ec5 no available information Ec6 no available information Ec7 no available information Ec8 eae, vtx2 (intimin and verocytotoxin producing gene) Ec9 eae, vtx2 (intimin and verocytotoxin producing gene) Ec10 aatA, aggR (enterotoxins producing gene)

ASSESSMENT OF MOLECULAR DIVERSITY OF SOME POTENTIAL PATHOGENIC E. COLI ISOLATED BY ERIC-PCR SCREENING
92
DNA isolation and purification For liquid cultures, the LB Broth (Miller) media (Neogen Food Safety, USA) were used. DNA was extracted from overnight cultures of selected E. coli bacterial strains using the commercial QuickExtract™ Bacterial DNA Extraction Kit (Epicentre, USA), according to the manufacturer's instructions. The suspension obtained was used as a matrix for the following enzymatic reactions. The quality and quantity of extracted DNA was assessed by spectrophoto-metry method on a NanoDrop 8000 Spectrophotometer (ThermoScientific, Germany). ERIC-PCR amplifications Escherichia coli isolates were analyzed using the ERIC-PCR method. The primers used for the ERIC-PCR method were ERIC lR, 5'ATGTAAGCTCCTGGGGATTCAC3’ and ERIC2 5'AAGTAAGTGACTGGGGTGAGCG3 (Versalovic et al., 1991). PCR was carried out in final reaction volumes of 25 µL containing 100 ng of DNA template. The amplification mixture was prepared according to the manufacturer's instructions for GoTaq Green PCR Master Mix (2X) commercial kit (Promega, USA). The ERIC-PCR reaction was performed on a DNA Engine Peltier Thermal Cycler (MJ Research, Canada) with the following program: first denaturing step for five minutes at 94 °C, followed by 45 cycles of denaturation at 94 °C for 45 seconds, annealing at 50 °C for 45 seconds, and extension at 72 °C for two minutes, while the final step of extension was at 72 °C for five minutes. The resulting PCR products were resolved on 1.8% agarose gels in TAE buffer at 100 V for 90 minutes. The PCR products were visualized and photographed under UV light. The data obtained were analyzed with VisionWorksLC software (UVP, England). Fingerprinting data interpretation The Unweighted Pair-Group Method (UPGMA) was chosen for the interpretation of data from the fingerprint experiment. The presence of alleles was recorded in a binary model, “1” for present and ”0” if absent. The Dice coefficient was used for comparison of the sets of the variables obtained. This binary matrix was used to develop the genetic similarity matrix and genetic distance matrix, which were used in the construction of a dendrogram using the DendroUPGMA software.
Results and discussion
The phylogenetic relationship was evaluated based on the DNA fingerprint similarities of E. coli strains isolated from different chicken farms. The data can be used to assess the level of the risk posed to public health and to develop strategies to prevent the spread of E. coli infections. The images of ERIC-PCR fingerprints of analyzed E. coli strains obtained by agarose gel electrophoresis were recorded. The positions of the bands on each lane and each gel were
normalized using the 1 kb DNA ladder as an external reference standard (Figs.1 and 2). The genetic diversity data obtained from this ERIC-PCR experiment are summarized in Table 2. The method that was applied in this experiment proved to be very informative; the markers harvested a total of 84 alleles, 82 of which were polymorphic. According to the results obtained, the accuracy of the method was
confirmed. This is in accordance with the results of other authors that described ERIC-PCR as a good technique for bacterial genotyping with higher discriminatory power than other techniques such as 16S rRNA gene analysis (Carvalho et al., 2001; Soltani et al., 2012; A’Yun et al., 2015). It has been shown that tested E. coli isolates are very divergent from the genetic point of view. This finding can be
Figure 1. DNA fingerprint obtained using ERIC 1R primer:
E1-E10 Escherichia coli strains, M - molecular weight marker - 100 bp DNA Ladder RTU, GeneDirex
Figure 2. DNA fingerprint obtained using ERIC 2 primer: E1-E10 Escherichia coli strains, M - molecular weight marker - 100 bp DNA Ladder RTU, GeneDirex

ASSESSMENT OF MOLECULAR DIVERSITY OF SOME POTENTIAL PATHOGENIC E. COLI ISOLATED BY ERIC-PCR SCREENING
93
explained by the fact that bacterial hereditary material is altered in the process where microorganisms are gaining new, often more pathogenic features (Ewers et al., 2003; Mellata et al., 2003). ERIC-PCR genotyping has also been shown beneficial for intra-serotypic differentiation of E. coli isolated from different sources (Prabhu et al., 2010; Soma Sekhar et al., 2017). Since we did not have information on the serotypes of selected E. coli strains, future work will also compare the virulence factors, serotypes and fingerprinting profile. Similarity and genetic distance matrices were developed for each individual. The similarity between the strains was determined on the basis of the Dice coefficient. Based on those matrices, a dendrogram showing phylogeny rela-tionships was constructed. The dendrogram was construc-ted on the basis of the averaged similarity of the matrix with the use of the algorithm of the UPGMA method. The nearest neighbor-joining clustering method has been used to show relations between similar groups. For the present experiment, ten individuals and 84 variables for each were analyzed. The genetic similarity values are used to reveal the genetic resemblance among analyzed individuals. Thus, a value of “1” means a very high similarity, this being a theoretical value, and it is obtained only in the case when comparing an individual to itself. The medium value is around 0.5. The genetic similarity was found to be
reduced, the highest value of 0.655 was recorded between the Ec8 and Ec9 strains, while the lowest similarity of 0.241 was recorded between the Ec3 and Ec5 strains. As seen in Table 3, the values for similarity hardly exceeded the medium threshold. The results indicate that for the scored alleles, the genetic similarity has a very low value. The values of genetic distances are used to reveal genetic divergence between analyzed strains. Thus, a value of “0” means no genetic divergence, and it is obtained only in the case when comparing an individual to itself, the value “1” being a theoretical maximum threshold that is impossible to be recorded in practice. The medium value is around 0.5. Analyzing the obtained matrix, it can be observed that the individuals are, on average, very distinct, as expected for the scored alleles. Most of the obtained values were over the medium value. The minimum value of 0.345, under the medium threshold, was obtained in this case between the Ec8 and Ec9 isolates. The highest value of 0.759 was recorded between individuals Ec3 and Ec5 (Table 4). The obtained matrices were used for the construction of an UPGMA dendrogram in the form of a cladogram (Fig. 3). The grouping clusters were developed using the numerical values of similarity coefficients, whereas the length of clusters was constructed on the basis of genetic distance coefficients.
Table 2. Data obtained in the fingerprinting experiment
Primer Fragment size range (bp) Scored alleles/polymophic alleles
ERIC 1 2655-238 40/40
ERIC 2 4370-208 44/42
Total 84/82 (97%) Table 3. Similarity matrix computed with Dice coefficient
Ec1 Ec2 Ec3 Ec4 Ec5 Ec6 Ec7 Ec8 Ec9 Ec10
Ec1 1 0.5 0.37 0.456 0.419 0.328 0.281 0.281 0.339 0.328
Ec2 1 0.39 0.508 0.312 0.508 0.305 0.339 0.393 0.413
Ec3 1 0.453 0.241 0.491 0.491 0.528 0.4 0.316
Ec4 1 0.492 0.467 0.5 0.429 0.414 0.267
Ec5 1 0.369 0.459 0.361 0.476 0.308
Ec6 1 0.4 0.5 0.484 0.281
Ec7 1 0.607 0.517 0.433
Ec8 1 0.655 0.3
Ec9 1 0.419
Ec10 1

ASSESSMENT OF MOLECULAR DIVERSITY OF SOME POTENTIAL PATHOGENIC E. COLI ISOLATED BY ERIC-PCR SCREENING
94
Table 4. Distance matrix based on Dice coefficient
Ec1 Ec2 Ec3 Ec4 Ec5 Ec6 Ec7 Ec8 Ec9 Ec10 Ec1 0 0.5 0.63 0.544 0.581 0.672 0.719 0.719 0.661 0.672 Ec2 0 0.61 0.492 0.688 0.492 0.695 0.661 0.607 0.587 Ec3 0 0.547 0.759 0.509 0.509 0.472 0.6 0.684 Ec4 0 0.508 0.533 0.5 0.571 0.586 0.733 Ec5 0 0.631 0.541 0.639 0.524 0.692 Ec6 0 0.6 0.5 0.516 0.719 Ec7 0 0.393 0.483 0.567 Ec8 0 0.345 0.7 Ec9 0 0.581 Ec10 0
Figure 3. UPGMA Dendrogram of E. coli isolates, created
by DendroUPGMA program using data from the two ERIC markers systems
Dendrogram analyses revealed the tested ten E. coli isolates to be grouped into two different major clusters. The first cluster contains only Ec10 isolate, whereas the other nine isolates are forming the second major cluster. The isolate Ec10 was previously found to be an aatA and aggR genotype, the only one of this kind that was selected for this study. In this selection, we considered that this genotype could be used as a control for validation of the study. The obtained results clearly confirm that this strain is different from the other strains analyzed in this study. The isolates from Ec1 to Ec9 are also grouped into two different sub-clusters. However, it is obvious that those nine isolates, even if they are caring the same genes, are divergent. They are probably the carriers of some different type of genes. The first sub-cluster containing the isolates Ec1, Ec2 and Ec4
could be considered as a cluster of eae gene carrying serotypes. Finally, it was concluded that the isolate Ec4 is most probably an eae genotype. The second sub-cluster is formed of five isolates, namely Ec3, Ec6, Ec7 and Ec8 and Ec9. However, the organization of the sub-sub-cluster leaves place for multiple suppositions. The isolate Ec3 carrying the eae gene is very closely related to individual Ec6, but they were grouped in the sub-cluster of eae and vtx2 serotypes. The possible explanation is gaining of other unspecific genes such as antibiotic resistance genes. These genes are the most plausible candidates to reveal genetic diversity among bacterial isolates that are carrying pathogenic genes, when analyzed with the ERIC molecular markers. However, it is clear that the isolates with eae and vtx2 genotype are very closely related and are grouped individually in the constructed dendrogram. Results of this research are in accordance with earlier studies that proved the highly significant discriminatory power in characterizing the genetic diversity of E. coli strains (de Moura et al., 2001; Mohapatra et al., 2007; Soma Sekhar et al., 2017). The diversity of E. coli strains clearly indicate that patho-genic E. coli continues to evolve. The ease of transmis-sibility of pathogenic E. coli urges use of rapid and accurate technologies for monitoring these pathogens. Nowadays, new technologies such as high-throughput sequencing and whole-genome sequencing have shown their use in typing and during outbreak investigation (Eppinger et al., 2011; Chan et al., 2012; Loman et al., 2013). The results of our research confirm successful use of a combination of viru-lence profiling with PCR-based genotyping tools in the investigation of virulent E. coli strains. Further research is necessary to evaluate additional threats of pathogens and to apply the method in different scenarios that include isolates from different origins.
Conclusions
The data obtained in this study show that ERIC-PCR is a very rapid and cost-effective technique for describing the diversity of E. coli isolates. It could be easily used for

ASSESSMENT OF MOLECULAR DIVERSITY OF SOME POTENTIAL PATHOGENIC E. COLI ISOLATED BY ERIC-PCR SCREENING
95
narrowing a large amount of collected isolates when searching for a particular genotype. It can also be used as an initial screening method for selected virulence or antibiotic resistance genes. The E. coli isolates screened in the present study were chosen in a way that the differences among them could serve as a control of the method accuracy. The major implication of this research is that the ERIC PCR analysis could serve as an effective tool in the routine surveillance of E. coli strains and that their evo-lution is particularly related to the development of new traits such as gaining new virulence and/or antibiotic resistance genes.
Acknowledgments
This work was conducted in the Antioxidant Systems Laboratory and Molecular Genetic Laboratory parts of the Horia Cernescu Research Unit of Banat's University of Agricultural Sciences and Veterinary Medicine ,,King Michael I of Romania”, Timisoara, Romania. Financed by UEFISCDI in the frame of the project entitled Bioeco-nomic Approach to Antimicrobial Agents – Use and Resis-tance PN-III-P1-1.2-PCCDI-2017-0361.
References
A’YUN, Q. U., A. SUWANTO, T. BARUS (2015): Genetic profiles of Escherichia coli isolated from Indonesian tempeh based on enterobacterial repetitive intergenic consensus-polymerase chain reaction (ERIC-PCR). Microbiol. Indones., 9 (2), 2.
ATEBA, C. N., M. MBEWE, C. C. BEZUIDENHOUT (2008): The prevalence of Escherichia coli O157 strains in cattle, pigs and humans in the North-West Province, South Africa. SAJS, 104, 7-8.
BARNES, H. J., W. B. GROSS (1997): Collibacillosis. In: CALNEK, B. W., H. J. BARNES, C. W. BEARD, L. R. MCDOUGALD, Y. M. SAIF, Editors. Diseases of Poultry. 10th ed. Iowa State University Press, Ames, IA, USA. Pp. 131-141.
BEUTIN, L., G. KRAUSE, S. ZIMMERMANN, S. KAULFUSS, K. GLEIER (2004): Characterisation of Shiga toxin-producing E. coli strains isolated from human patients in Germany over a 3-year period. Clin. Microbiol., 42, 1099-1180.
BROCCHI, M., A. FERREIRA, M. LANCELLOTTI, E. G. STEHLING, T. A. CAMPOS, G. NAKAZATO, A. F. CASTRO, W. D. SILVEIRA (2006): Typing of avian patho-genic Escherichia coli strains by REP-PCR. Pesq. Vet. Bras., 26 (2), 299-311.
CARVALHO, F. A., N. BARNICH, A. SIVIGNON, C. DARCHA, C. H. F. CHAN, C. P. STANNERS, A. DARFEUILLE-MICHAUD (2009): Crohn’s disease adherent-invasive Esche-richia coli colonize and induce strong gut inflammation in transgenic mice expressing human CEACAM. J. Exp. Med., 206, 2179-2189.
CHAN, J. Z-M., M. J. PALLEN, B. OPPENHEIM, C. CONSTANTINIDOU (2012): Genome sequencing in clinical microbiology. Nat. Biotechnol., 30, 1068-1071.
CHANG, H. G., B. TSERENPUNTSAG, M. KACICA, P. F. SMITH, D. L. MORSE (2004): Haemolytic uraemic syndrome incidence in New York. Emerg. Infect. Dis., 10, 928-931.
CHANSIRIPORNCHAI, N. (2009): Comparative efficacy of enrofloxacin and oxytetracycline by different administration methods in broilers after experimental infection with avian pathogenic Escherichia coli. Thai. J. Vet. Med., 39, 231-236.
CLEMENTS, A., J. C. YOUNG, N. CONSTANTINOU, G. FRANKEL (2012): Infection strategies of enteric pathogenic Escherichia coli. Gut Microbes., 3, 71-87.
COWDEN, J. M., S. AHMED, M. DONAGHY, A. RILEY (2001): Epidemiological investigation of the central Scotland outbreak of Escherichia coli O157 infection, November to December 1996. Epidemiol. Infect., 126 (3), 335-341.
CROXEN, M. A., B. B. FINLAY (2010): Molecular mechanisms of Escherichia coli pathogenicity. Nat. Rev. Microbiol., 8, 26-38.
CROXEN, M. A., R. J. LAW, R. SCHOLZ, K. M. KEENEY, M. WLODARSKA, B. B. FINLAY (2013): Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev., 26, 822-880.
DE CARLI, S., N. IKUTA, F. K. LEHMANN, V. P. DA SILVEIRA, G. DE MELO PREDEBON, A. S. FONSECA, V. R. LUNGE (2015): Virulence gene content in Escherichia coli isolates from poultry flocks with clinical signs of colibacillosis in Brazil. Poult. Sci., 94, 2635-2640.
DE MOURA A. C., K. IRINO, M. C. VIDOTTO (2001): Genetic variability of avian Escherichia coli strains evaluated by enterobacterial repetitive intergenic consensus and repetitive extragenic palindromic polymerase chain reaction. Avian Dis., 1, 173-181.
DUNDAS, S., W. T. TODD, A. I. STEWART, P. S. MURDOCH, A. K. CHAUDHURI, S. J. HUTCHINSON (2001): The central Scotland Escherichia coli O157: H7 outbreak: Risk factors for the haemolytic uremic syndrome and death among hospitalized patients. Clin. Infect. Dis., 33, 923-931.
DZIVA, F., H. HAUSER, T. R. CONNOR, P. M. VAN DIEMEN, G. PRESCOTT, G. C. LANGRIDGE (2013): Sequencing and functional annotation of avian pathogenic Escherichia coli serogroup O78 strains reveal the evolution of E. coli lineages pathogenic for poultry via distinct mechanisms. Infect. Immun., 81, 838-849.
EPPINGER, M., M. K. MAMMEL, J. E. LECLERC, J. RAVEL, T. A. CEBULA (2011): Genomic anatomy of Escherichia coli O157:H7 outbreaks. Proc. Natl. Acad. Sci. U. S. A. 108, 20142-20147.
EWERS, C., T. JANSEN, L. H. WIELER (2003): Avian pathogenic Escherichia coli (APEC). Berl. Munch. Tierarztl. Wochenschr., 116 (9-10), 381-395.
EWERS, C., T. JANΒEN, S. KIEΒLING, H. C. PHILIPP, H. LOTHAR, H. WIELER (2004): Molecular epidemiology of avian pathogenic Escherichia coli (APEC) isolated from colisepticemia in poultry. Vet. Microbiol., 104, 91-101.
GEETHA, M., K. M. PALANIVEL (2018): Avian Colibacillosis – A Mini Review. Int. J. Pure. App. Biosci., 6 (1), 376-380.
HULTON, C. S. J., C. F. HIGGINS, P. M. SHARP (1991): ERIC sequences: a novel family of repetitive elements in the genomes of Escherichia coli, Salmonella typhimurium and other enterobacteria. Mol. Microbiol., 5, 825-834.
IGARASHI, T., J. INATOMI, A. WAKE, M. TAKAMIZAWA, H. KATAYAMA, T. IWATA (1999): Failure of pre-diarrheal antibiotics to prevent haemolytic uraemic syndrome in serologically proven Escherichia coli O157:H7 gastrointestinal infection. J. Pediat., 135, 768-769.

ASSESSMENT OF MOLECULAR DIVERSITY OF SOME POTENTIAL PATHOGENIC E. COLI ISOLATED BY ERIC-PCR SCREENING
96
JOHNSON, J. R., T. T. O’BRYAN (2000): Improved repetitive-element PCR fingerprinting for resolving pathogenic and nonpathogenic phylogenetic groups within Escherichia coli. Clin. Diagn. Lab. Immunol., 7, 265-273.
JOHNSTON, A. (2007): Current disease of ducks and their control. Poult. Int. 46, 24-29.
KARMALI, M. A., V. GANNON, J. M. SARGEANT (2010): Verocytotoxin-producing Escherichia coli (VTEC). Vet. Microbiol., 140, 360-370.
KHOO, L. L., Y. HASNAH, Y. ROSNAHY, N. SAIFUR, M. A. MASWATI, M. RAMLAN (2010): The prevalence of avian pathogenic Escherichia coli in peninsular Malaysia. Malays. J. Vet. Res., 1, 27-31.
KNOW, S. G., S. Y. CHA, E. J. CHOI, B. KIM, H. J. SONG, H. K. JANG (2008): Epidemiological prevalence of avian patho-genic Escherichia coli differentiated by multiplex PCR from commercial chickens and hatchery in Korea. J. Bacteriol. Virol., 38 (4), 179-188.
LA RAGIONE, R. M., M. J. WOODWARD (2002): Virulence factors of Escherichia coli serotypes associated with avian colisepticemia. Res. Vet. Sci., 73, 27-35.
LOMAN, N. J., C. CONSTANTINIDOU, M. CHRISTNER, H. ROHDE, J. Z.-M. CHAN, J. QUICK, J. C. WEIR, C. QUINCE, G. P. SMITH, J. R. BETLEY, M. AEPFEL-BACHER, M. J. PALLEN (2013): A culture-independent sequence-based metagenomics approach to the investigation of an outbreak of Shiga-toxigenic Escherichia coli O104:H4. JAMA, 309, 1502-1510.
LUTFUL KABIR, S. M. (2010): Avian colibacillosis and salmonellosis: a closer look at epidemiology, pathogenesis, diagnosis, control and public health concerns. Int. J. Environ. Res. Public Health, 7, 89-114.
LYNN, R.M., S. J. O’BRIEN, C. M. TAYLOR, G. K. ADAK, H. CHART, T. CHEASTY, J. E. COIA, I. A. GILLESPIE, M. E. LOCKING, W. J. REILLY (2005): Childhood haemolytic uraemic syndrome, United Kingdom and Ireland. Emerg. Infect. Dis., 11, 590-596.
MELLATA, M., M. DHO-MOULIN, C. M. DOZOIS, R. CUR-TISS, P. K. BROWN, P. ARNE, A. BREE, C. DESAUTELS, J. M. FAIRBROTHER (2003): Role of virulence factors in resistance of avian pathogenic Escherichia coli to serum and in pathogenicity. Infect. Immun., 71 (1), 536-540.
MITCHELL, N. M., J. R. JOHNSON, B. JOHNSTON, R. CURTISS III, M. MELLATA (2015): Zoonotic potential of Escherichia coli isolates from retail chicken meat products and eggs. Appl. Environ.Microbiol., 81, 1177-1187.
MOHAPATRA B. R, K. BROERSMA, A. MAZUMDER (2007): Comparison of five rep-PCR genomic fingerprinting methods for differentiation of fecal Escherichia coli from humans, poultry and wild birds. FEMS Microbiol. Lett., 277, 98-106.
MÜLLER, E.E., M. M. EHLERS, W. O. K. GRABOW (2001): The occurrence of E. coli O157:H7 in Southern African water sources intended for direct and indirect human consumption. Water Res., 35, 3085-3088.
NATARO, J.P., J. B. KAPER (1998): Diarrhoeagenic Escherichia coli. Clin. Microbiol. Rev., 11, 142-201.
NOLAN, L.K. (2013): Colibacillosis. Diseases of Poultry. 13th ed. Ch. 18. Ames: Wiley-Blackwell, Print.
OLARINMOYE, O., O. O. OLADELE, A. A. ADEDIJI, U. G. NTIWUNKA, G. O. TAYO (2013): Antibiograms of avian
pathogenic Escherichia coli isolates from commercial layers with colibacillosis in Southwest Nigeria. Malays. J. Micro-biol., 9 (4), 317-325.
OLSEN, S. J., G. MILLER, T. BREUER, M. KENNEDY, C. HIGGINS, J. WALFORD, G. MCKEE, K. FOX, W. BIBB, P. A. MEAD (2002): Waterborne outbreak of Escherichia coli O157 infections and haemolytic uraemic syndrome: Implica-tions for rural water systems, Emerg. Infect. Dis., 8, 370-375.
PRABHU V., S. ISLOOR, M. BALU, V. V. SURYA-NARAYANA, D. RATHNAMMA (2010): Genotyping by ERIC-PCR of Escherichia coli isolated from bovine mastitis cases. Indian J. Biotechnol., 9, 298-301.
RAHMAN, M. A., M. A. SAMAD, M. B. RAHMAN, S. M. L. KABIR (2004): Bacteriopathological studies on salmonellosis, colibacillosis and pasteurellosis in natural and experimental infections in chickens. Bangladesh J. Vet. Med., 2, 1-8.
SAXENA, M. K., V. P. SINGH, B. D. LAKHCHARUA, G. TAJ, B. SHARMA. (2002): Strain differentiation of Indian isolates of Salmonella by ERIC-PCR. Res. Vet. Sci., 73, 313-314.
SCHOULER, C., B. SCHAEFFER, A. BREE, A. MORA, G. DAHBI, F. BIET, E. OSWALD, J. MAINIL, J. BLANCO, M. MOULIN- SCHOULEUR, M. (2012): Diagnostic strategy for identifying avian pathogenic Escherichia coli based on four patterns of virulence genes. J. Clin. Microbiol., 50, 1673-1678.
SHARPLES, G. J., R. G. LLOYD. (1990): A novel repeated DNA sequence located in the intergenic regions of bacterial chromosomes. Nucleic Acids Res., 18, 6503-6508.
SOLÀ-GINÉS, M., K. CAMERON-VEAS, I. BADIOLA, R. DOLZ, N. MAJÓ, G. DAHBI, S. VISO, A. MORA, J. BLANCO, N. PIEDRA- CARRASCO, J. J. GONZÁLEZ-LÓPEZ, L. MIGURA-GARCIA (2015): Diversity of multi-drug resistant avian pathogenic Escherichia coli (APEC) causing outbreaks of colibacillosis in broilers during 2012 in Spain. PLoS One, 10(11): e0143191. DOI: 10.1371/ /journal.pone.0143191.
SOLTANI M, S. M. PEIGHAMBARI, M. ASKARI BADOUEI, A. SADRZADEH (2012): Molecular typing of avian Escherichia coli isolates by enterobacterial repetitive intergenic consensus sequences-polymerase chain reaction (ERIC-PCR) Iran. J. Vet. Med., 6 (3):143-148.
SOMA SEKHAR, M., N. MOHAMMAD SHARIF, T. SRINVASA RAO, M. METTA (2017): Genotyping of viru-lent Escherichia coli obtained from poultry and poultry farm workers using enterobacterial repetitive intergenic consensus-polymerase chain reaction. Vet. World., 10 (11), 1292-1296.
TOZZI, A. E., A. CAPRIOLI, F. MINELLI, A. GIANVITI, L. DE PETRIS, A. EDEFONTI, G. MONTINI, A. FERRETTI, T. DE PALO, M. GAIDO (2003): Shiga toxin-producing Escherichia coli infections associated with haemolytic uremic syndrome, Italy, 1988–2000. Emerg. Infect. Dis., 9, 106-108.
VERSALOVIC, J., T. KOEUTH, J. R. LUPSKI (1991): Distri-bution of repetitive DNA sequences in eubacteria and application to fingerprinting of bacterial genomes, Nucleic Acids Res., 19 (24), 6823-6831.
WILSON, L. A., P. M. SHARP (2006): Enterobacterial repetitive intergenic consensus (ERIC) sequences in Escherichia coli: Evolution and implications for ERIC-PCR, Molecular biology and evolution, 23 (6), 1156-1168.

ASSESSMENT OF MOLECULAR DIVERSITY OF SOME POTENTIAL PATHOGENIC E. COLI ISOLATED BY ERIC-PCR SCREENING
97
PROCJENA MOLEKULARNE RAZNOLIKOSTI NEKIH POTENCIJALNO PATOGENIH IZOLATA E. COLI POMOĆU ERIC-PCR PROBIRA
Sažetak
Većinu sojeva bakterije Escherichia (E.) coli smatra se bezopasnim nametnikom u ljudi i životinja. Međutim, određeni izolati mogu biti izvor otpornih gena i/ili mogu uzrokovati infekcije. Pojavljivanje otpornosti E. coli na više lijekova postaje problem širom svijeta, što rezultira brojnim istraživanjima antimikrobne rezistencije, virulencije bakterijskih patogena peradi te raznim epidemiološkim studijama. Primjena molekularnih metoda u ovim istraživanjima obećava sveobuhvatnije utvrđivanje mogućeg rizika od izolata E. coli podrijetlom iz peradi za zdravlje peradi i ljudi. Stoga se podaci o genetskoj raznovrsnosti bakterijskih sojeva pokazuju važnim alatom za otkrivanje različitih obrazaca rezistencije na antibiotike. Cilj ovoga istraživanja bio je primijeniti molekularni pristup u definiranju zoonotskog potencijala izolata E. coli podrijetlom iz pilića. Kompletna genomska DNA pročišćena je iz tekućih kultura sojeva E. coli izoliranih iz pilića s različitih gospodarstava. Ispitivanje molekularne raznolikosti izolata E. coli provedeno je pomoću molekularnih biljega Enterobacterial Repetitive Intergenic Consensus (ERIC) – ERIC 1 i ERIC 2 i metodom otisaka lančanom reakcijom polimeraze. Temeljem molekularnih podataka dobivenih u pokusu s otiscima izrađen je filogenetski dendrogram Unweighted Pair Group Method with Arithmetic mean (UPGMA) koji pokazuje genetski odnos među izolatima. U radu se opisuje prvi korak u razvoju metode probira na genetsku raznolikost radi otkrivanja potencijalno patogenih sojeva E. coli na osnovi specifičnih molekularnih biljega. Podaci prikupljeni u ovom istraživanju mogu poslužiti u grupiranju više bakterijskih sojeva prema mehanizmima razvoja antimikrobne rezistencije. Nadalje, ovaj će se pristup rabiti u procjeni razine gore spomenutog rizika za javno zdravlje, kao i za razvoj strategija za sprječavanje širenja infekcija bakterijom E. coli. Ključne riječi: Escherichia coli, PCR, ERIC sekvence, patogeni sojevi, raznolikost

PERADARSKI DANI 2019. 98
PROIZVODNI POKAZATELJI NAKON PRIMJENE AUTOGENOG CJEPIVA PROTIV PTIČJIH PATOGENIH SOJEVA E. COLI I POBOLJŠANJA PROGRAMA CIJEPLJENJA U JATIMA TEŠKE LINIJE KOKOŠI
Željko Gottstein1, Liča Lozica1, Danijela Horvatek Tomić1, Maja Lukač1, Estella Prukner-Radovčić1, Metka Vlahek2, Nada Dolenčić2, Željko Glavak2
1Sveučilište u Zagrebu, Veterinarski fakultet, Zagreb, Hrvatska 2Galivet d.o.o., Prelog, Hrvatska
Sažetak
Imunoprofilaksa peradi predstavlja jednu od temeljnih mjera u zaštiti zdravlja peradi, ali i čovjeka kao krajnjeg konzumenta. Ova je mjera naročito naglašena u rasplodnih jata koja dio imunosti moraju prenijeti i na potomstvo. Protiv virusnih bolesti je primjena cjepiva najčešća, no i bakterijske se bolesti može uspješno kontrolirati tim postupkom. Od bakterija, ptičji patogeni sojevi E. coli (APEC) najčešći su uzrok problema na proizvodnim farmama rasplodnih i konzumnih jaja, često u kombinaciji s Gallibacterium anatis i Mycoplasma synoviae te zaraznim bronhitisom. U istraživanju provedenom na farmi teške linije kokoši utvrđeno je postojanje značajnih gubitaka djelovanjem upravo sojeva APEC koji su uz visoku patogenost bili i rezistentni na većinu antimikrobnih pripravaka. U svrhu kontrole kolibaciloze primijenjeno je autogeno cjepivo u 5 jata. Uz navedeno cjepni program je poboljšan u imunoprofilaksi zaraznog bronhitisa, zarazne anemije pilića, reoviroze, ptičjeg rinotraheitisa i mikoplazmoze. Rezultati kod navedenih jata pokazuju kontinuirano i statistički značajno smanjenje broja uginuća u razdoblju proizvodnje s 20.74% na 9.78%, smanjen je broj antimikrobnih terapija, dok je proizvodnja prosječno povećana za 7,5 jaja po useljenoj nesilici. Navedeni rezultati pokazuju potrebu za kontinuiranim praćenjem zdravlja na farmama, poboljšanjem cjepnog programa i načina primjene cjepiva, uz kontinuirane mjere s ciljem poboljšanja biosigurnosti na farmama. Ključne riječi: roditeljsko jato teške pasmine kokoši, APEC, E. coli, autogeno cjepivo, program cijepljenja Uvod
Osnova intenzivne peradarske proizvodnje temeljena je na visokoj razini biosigurnosnih mjera te kontinuiranoj kontroli svih razina proizvodnje kako bi se na vrijeme prepoznao i uklonio potencijalni problem. Ovo je naročito naglašeno u generacijama rasplodnih jata kokoši gdje propusti mogu iz-ravno kompromitirati vlastitu proizvodnju, a poglavito onu sljedeće generacije s izravnim utjecajem na pojavu problema već u razdoblju inkubacije jaja, a potom i proizvodnje. Ovaj slijed vertikalnog prijenosa je naročito naglašen u slučaju niza zaraznih bolesti koje mogu značajno kompromitirati ka-snije proizvodne rezultate pilića. Naizgled zadovoljavajući ili pak vrhunski proizvodni rezultati roditeljskih jata mogu u pilića rezultirati značajnim gubicima. Takav očit primjer je zanemarivanje imunoprofilakse osobito virusnih bolesti, npr. zarazne anemije pilića, što može dovesti do devastirajućih
posljedica u sljedećoj generaciji (Adair, 2000.; Hagood i sur., 2000.). Bakterije se pak vrlo često izdvajaju kao uzroč-nici u tijeku proizvodnje te neizbježno sudjeluju u vertikal-nom prijenosu pri čemu je njihova etiologija i prepoznavanje često u kompleksu s drugim uzročnicima i najčešće sekun-darno uzrokuju značajne štete nastavljajući se na neki pri-marni inzult, zarazni ili tehnološki (Pires-dos-Santos i sur., 2013.). Najčešće podlogu za nastavak problema u vidu klasične bakterijske zaraze stvaraju zarazni bronhitis i mikoplazme. Takav primjer je Escherichia (E.) coli koja kao komenzal doprinosi zdravlju i ravnoteži, naročito mikro-bioma u crijevima. No, istodobno izvancrijevni sojevi ove vrste (ExPEC) kao izrazito patogeni za perad (APEC) mogu uzrokovati značajne probleme te posljedično i gubitke u svim segmentima proizvodnog ciklusa, od roditelja do kraja tova pilića. Istodobno, kao mikroorganizam s izrazitim zoonotskim potencijalom, APEC može biti prijetnja i čovje-
Izv. prof. Željko Gottstein, Sveučilište u Zagrebu, Veterinarski fakultet, Zavod za bolesti peradi s klinikom, Heinzelova 55, 10000 Zagreb, Hrvatska; Tel: +385 (0)1 2390 280, fax: +385 (0)1 2390 410; e-mail: [email protected], [email protected]

PROIZVODNI POKAZATELJI NAKON PRIMJENE AUTOGENOG CJEPIVA PROTIV PTIČJIH PATOGENIH SOJEVA E. COLI I POBOLJŠANJA PROGRAMA CIJEPLJENJA U JATIMA TEŠKE LINIJE KOKOŠI
99
ku u lancu „od polja do stola“ (Jakobsen i sur., 2012.). Bakterijske, ali i druge uzročnike moguće je držati pod kon-trolom dobrom pripremom objekata, biosigurnosnim mje-ama i kasnije racionalnom primjenom antibiotika. Nepridr-žavanjem navedenih mjera osigurava se kontinuirano zadr-žavanje mikroorganizama u objektima uz kontinuirano povećanje koncentracije, postupnu evoluciju patogenih so-jeva i njihovu daljnju „prilagodbu“ na domaćina te postupan razvoj rezistencije, što onemogućava bilo kakav anti-mikrobni tretman (Heidemann Olsen i sur., 2016.). U takvim uvjetima nemoguće se boriti i održati proizvodnju u jatima peradi u standardima genetike, a naročito ostvariti dodanu vrijednost. Osiguranje minimalnih tehnoloških uvjeta i pri-
državanje spomenutih temeljnih načela proizvodnje stvara osnovu u kojoj je moguće imunoprofilaksom ostvariti do-datnu zaštitu rasplodnih jata. To istodobno osigurava i zaštitu sljedeće generacije od vertikalnog prijenosa, kao i pasivno od horizontalne zaraze, do razvoja aktivne obrane u pilića. Upravo ovi postulati su temelj na kojima mora poči-vati proizvodnja da bi bila uspješna. Cilj ovoga istraživanja je longitudinalno praćenje proizvod-nih i zdravstvenih pokazatelja kroz 7 roditeljskih jata teške pasmine kokoši Ross 308 nakon primjene autogenog cjepiva protiv E. coli te drugih cjepiva u programu cijepljenja sa svrhom poboljšanja zdravlja i proizvodnosti na samoj farmi, kao i konačnog proizvoda, tovnog pileta.
Tablica 1. Program cijepljenja po jatima prije i nakon uvođenja promjena (+ - provedeno; x - isključeno) Table 1. Vaccination program according to flocks before and after modifications (+ - present; x - excluded)
Dan / Day (D) Tjedan / Week (T/W)
Cjepivo / Vaccine R-26/R-27 R-28 R-29 R-30, R-31, R-32
D 1 Marek's E. coli live ZB/IB
+ + +
+ x
ZB/IB Ma5
+ x
ZB/IB Ma5 D 2 S. enteritidis live + + +
D 7 Kokcidioza/Coccidiosis
+ + PRT/ART live
+ PRT/ART live
D 14 NDV
+ +
+ ZB/IB 4/91
D 18 ZBB/IBD + + + D 26 ZBB/IBD + + + D 30 ZB/IB + + x
T/W 5 E. coli live
+
x
x ZB/IB Ma5
T/W 6 S. enteritidis live + + + T/W 8 NDV + + + T/W 9 ZB/IB + x x
T/W 9/10
E. coli inact. commerc. POX+AE
E. coli auto. +
E. coli auto. +
MS-H ZAP/CAV Reo live
E. coli auto. +
MS-H ZAP/CAV Reo live
T/W 11/12 ZB/IB 4/91
T/W 18
NDV+ZBB/IBD +ZB/IB+EDS inact. Salmonella inact. E. coli inact. commerc.
+
+ E. coli auto.
+
PRT/ART inact. Reo inact.
+ E. coli auto.
+
PRT/ART inact. Reo inact.
+ E. coli auto.
T/W 30 ZB/IB Ma5 / 4/91
6 T/W interval

PROIZVODNI POKAZATELJI NAKON PRIMJENE AUTOGENOG CJEPIVA PROTIV PTIČJIH PATOGENIH SOJEVA E. COLI I POBOLJŠANJA PROGRAMA CIJEPLJENJA U JATIMA TEŠKE LINIJE KOKOŠI
100
Materijali i metode U istraživanju su praćena roditeljska jata teške pasmine kokoši (Ross 308) tijekom 4 godine, pri čemu je kroz navedeno razdoblje proizvodni ciklus završilo 7 jata. S farme su redovito dostavljane lešine kokoši i pijetlova na razudbu kako bi se utvrdio uzrok uginuća, s naglaskom na praćenju kolibaciloze. Iz lešina su izdvojeni sojevi E. coli bakteriološkom pretragom koji su, nakon potvrde metodom Maldi Tof i molekularne tipizacije (Clermont i sur., 2000.; Horvatek i sur., 2017.) korišteni za izradu autogenog cjepiva (Melamed i sur., 1991.). Cjepivo je izrađeno u dozi od 5x108 bakterija u 0,3 mL u kombinaciji s uljnim adjuvansom Montanide ISA 71 R VG (SEPPIC, Francuska) te primi-jenjeno dvokratno u dobi 9 i 19 tjedana. Također su do-stavljani i uzorci seruma za dokaz specifičnih protutijela za reovirus (REO), zaraznu anemiju pilića (ZAP), virus ptičjeg rinotraheitisa (PRT) (Gottstein i sur., 2017.b), Mycoplasma synoviae (MS) i virus zaraznog bronhitisa (ZB) primjenom komercijalnih ELISA kitova (Biochek, Nizozemska, x-OvO Limited, UK) prema uputi proizvođača. Navedenim po-
stupkom proveden je monitoring postojanja divljih sojeva u jatu za uzročnike za koje nisu korištena cjepiva te odgovor na cjepiva korištena prema programu cijepljenja (tablica 1.). Proveden je i dokaz te molekularna tipizacija virusa ZB u jatima (Nedeljković i sur., 2018.). Po jatima su prikupljani podaci o tjednim uginućima i provedenim antimikrobnim terapijama od 24. do 58. tjedna te proizvodnja jaja na kraju proizvodnog ciklusa, a rezultati su statistički obrađeni primjenom testova Kruskal-Wallis i Anova LSD koristeći računalni program Statistica 13 (Tibco, SAD). Rezultati Patomorfološkom pretragom lešina u jatima R-26 i R-27 kao polazišnim jatima utvrđene su opsežne promjene u vidu sirastih naslaga u torakoabdominalnoj šupljini, fibrinozni poliserozitis, perihepatitis i perikarditis. Istodobno je mikro-biološkom pretragom utvrđena opsežna zaraza bakterijom E. coli koja je izdvojena u čistoj kulturi iz svih organa, a navedeni izolati su potom korišteni za pripremu autogenog cjepiva. Praćenjem ostalih jata, od R-28 do R-32, opsežne siraste naslage se pri patomorfološkim pretragama gube, uz
Slika 1. Ukupno i prosječno tjedno uginuće (%) tijekom proizvodnje po jatima od R-26 do R-32 (vrijednosti prosječnog
tjednog uginuća označene različitim malim slovima abecede statistički se značajno razlikuju na razini p<0,05) Figure 1. Total and average weekly mortality (%) during production in R-26 to R-32 flocks (average weekly mortality marked
with different letters are significantly different at the level p<0.05)

PROIZVODNI POKAZATELJI NAKON PRIMJENE AUTOGENOG CJEPIVA PROTIV PTIČJIH PATOGENIH SOJEVA E. COLI I POBOLJŠANJA PROGRAMA CIJEPLJENJA U JATIMA TEŠKE LINIJE KOKOŠI
101
poneki fibrinozni poliserozitis kod kojeg je obično izdvojena E. coli i iz jetre, koštane srži i peritoneuma, a novo-izdvojeni sojevi se koriste za imunizaciju sljedećih jata. U jatima R-28 i R-29 koja su praćena od početka proizvodnog ciklusa uočen je povećan broj lažnih nesilica među dostavljenim lešinama, za razliku od R-26 i R-27 koje su bile pri samom kraju ili drugoj polovici proizvodnog ciklusa. Molekularnom dijagnostikom i tipizacijom sojeva izdvojenih u jatima utvrđena je prisutnost varijantnog soja 4/91. Temeljem navedenog uvedeno je cijepljenje u valionici sojem ZB Ma5 te potom naizmjence varijantnim sojem 4/91 tijekom uzgoja, ali i u proizvodnji (tablica 1.). Cjepiva su primijenjena raspršivanjem, što je kod kasnijih jata značajno smanjilo broj lažnih nesilica. Serološkim monitoringom uočeni su visoki titrovi za viruse REO, PRT i ZAP, kao i za MS te su također uvršteni u program cijepljenja. Rezultati praćenja postotka tjednog uginuća (slika 1.) pokazuju kontinuirani pad koji se prosječno kretao od 0,59% u R-26 pa do 0,28% u R-32. Ovaj pad je statistički značajan u jata od R-29 do R-32 u odnosu na prethodna jata. Kao rezultat navedenog sma-njenja prosječnog tjednog uginuća, ukupno uginuće rasplodnih kokoši tijekom proizvodnje od 24. do 58. tjedna smanjeno je za gotovo 11%, s 20,74% kod R-26 na 9,78% kod R-32. Istodobno, analizom broja snešenih rasplodnih jaja utvrđeno je povećanje prosječnog broja jaja po jatu za preko 230 tisuća, što na prosječni broj rasplodnih jedinki po jatu od 31.385 čini povećanje od 7,5 jaja po kokoši (tablica 2.). Broj provedenih terapija i ukupan broj kokoši pod terapijom se od R-26 do R-31 postupno smanjuje, pri čemu kod R-31 potpuno izostaje potreba za terapijom (tablica 3.).
Rasprava i zaključci
Neupitno je da je u intenzivnom peradarstvu imunoprofi-laksa osnova proizvodnje. Zbog postojanja različitih mikro-organizama koji mogu gotovo onemogućiti proizvodnju, te gubitke cijepljenje značajno smanjuje ili svodi na prihvat-ljivu razinu. Pritom cjepivo ima i važnu ulogu u sprječa-vanju cirkulacije divljih sojeva te ih postupno istiskuje s
farme, kao u slučaju MS (Kleven, 2008.). Kod nekih uzroč-nika cjepni i divlji soj postoje kontinuirano u organizmu pričem cjepivo, iako štiti, postupno doprinosi razvoju viru-lentnijih sojeva, čemu je virus Marekove bolesti klasičan primjer (Nair, 2018.). Primjena autogenih cjepiva posljed-njih nekoliko godina u peradarskoj proizvodnji je u porastu. Razlog tomu je nemogućnost razvoja univerzalnog komer-cijalnog cjepiva koje bi pružilo kvalitetnu zaštitu od lokalnih sojeva, što je naročito vidljivo kod bakterijskih cjepiva kod kojih je unakrižna zaštita najčešće slaba. Istodobno se pokazalo da autogena cjepiva od lokalnih patogenih sojeva APEC mogu osigurati dodatnu zaštitu (El Jakee i sur., 2016.), no neka istraživanja upravo na teškoj liniji umanjuju njihov učinak u zaštiti od heterolognih sojeva (Li i sur., 2017.), što se zapravo može očekivati. Kako je kolibaciloza najčešće multikauzalne naravi, potrebno je kontinuiranim monitoringom pratiti svako jato te na vrijeme prilagoditi program cijepljenja. Upravo polivalentna autogena cjepiva od bakterijskih vrsta dominantno izdvojenih iz organa ko-koši pri razudbi mogu osigurati zaštitu, dok ju komercijalna živa i inaktivirana cjepiva ne osiguravaju u dovoljnoj razini. To je naročito važno upravo kod kolibaciloze kojoj podlogu najčešće čine ZB, MS ili Gallibacterium anatis. Pokazalo se da polivalentna cjepiva od sojeva APEC E. coli i Galli-bacterium anatis imaju značajnu ulogu u zaštiti visoko-produktivnih jata kokoši nesilica konzumnih jaja (Gottstein i sur., 2018.). Ponekad je program cijepljenja nemoguće prilagoditi ako je jato, npr. pri kraju uzgoja, kad se jedno-stavno više ne može cjepivom osigurati zaštitu, jer je već zaraženo, kao u slučaju MS (Kleven, 2008.). Kontinuirani
monitoring se i na ovoj farmi pokazao kao važan alat u kontroli zdravlja jata, u odabiru odgovarajuće antimikrobne terapije te prilagodbi programa i postupka primjene cjepiva. U jata R-28 i 29 bio je uočen značajan udio lažnih nesilica među uginućima, što kod R-26 i 27 nije bilo uočeno u tolikom broju budući da je monitoring započeo već pri kraju ili u drugoj polovici njihovog proizvodnog ciklusa. Uočeno je da su lažne nesilice hijerarhijski vjerojatno na najnižoj
Tablica 2. Rezultati analize proizvodnje rasplodnih jaja po jatima prije i nakon promjena programa cijepljenja Table 2. Results of hatching eggs production according to flocks before and after modification of vaccination program
Prosječan broj jaja po jatu R-26 i R-27
Average egg number in R-26 and R-27 flocks
Prosječan broj jaja po jatu R-28 do R31
Average egg number in R-28 to R-31 flocks
Razlika Difference
Prosječan broj nesilica u jatu
Average number of hens per flock
Povećanje broja jaja po nesilici
Increase in egg production per hen
4 042 290 4 278 107 235 817 31 385 7,5
Tablica 3. Broj terapija i ukupan broj kokoši pod terapijom Table 3. Number of therapies and total number of hens on therapy
Jato/Flock R-26 R-27 R-28 R-29 R-30 R-31 R-32 Broj terapija / Number of therapies 2 2 1 1 1 0 2 Ukupan broj kokoši pod terapijom Total number of hens on therapy 35 000 60 000 5000 30 000 5000 0 20 000

PROIZVODNI POKAZATELJI NAKON PRIMJENE AUTOGENOG CJEPIVA PROTIV PTIČJIH PATOGENIH SOJEVA E. COLI I POBOLJŠANJA PROGRAMA CIJEPLJENJA U JATIMA TEŠKE LINIJE KOKOŠI
102
razini i često budu žrtve kanibalizma pa je njihova učestalost na razudbi velika, temeljem čega je i uočen problem. Prove-dena molekularna tipizacija sojeva ukazuje na postojanje varijantnog soja 4/91 koji se od cjepnog razlikuje u svega nekoliko baza (Nedeljković i sur., 2018.). Rezultati različitih istraživanja pokazuju da se divlji sojevi 4/91 često ne razli-kuju ili se vrlo malo razlikuju od cjepnih. Temeljem toga uvedeno je cijepljenje već u valionici sojem Ma5 te potom naizmjence cijepljenje sojem 4/91 ZB tijekom uzgoja, a potom i tijekom proizvodnje, uz primjenu cjepiva raspr-šivanjem. Navedena kombinacija sojeva ZB osigurava širu zaštitu koja uključuje i QX i njemu slične sojeve, a primjena cjepiva raspršivanjem pruža bolju lokalnu zaštitu na sluz-nicama dišnog trakta, što je i cilj imunoprofilakse pri-mjenom živog cjepiva (De Wit i sur., 2010.). Zbog po-stojanja visokih titrova za REO i PRT uvedeno je cijepljenje i za te bolesti, kao i za ZAP za koju su titrovi u pilenki bili niski, a počeli bi rasti tek ulaskom u proizvodnju, što bi bila osnova za vertikalni prijenos (Gottstein i sur., 2017.b; Otaki i sur., 1992.). Kako je učestalost zaraze MS u jatima peradi u Europi i u Hrvatskoj izrazito visoka (Horvatek i sur., 2018.) monitoring ove bolesti je izrazito važan zbog česte prisut-nosti u podlozi različitih bolesti, naročito kolibaciloze. Mo-nitoringom su utvrđeni visoki titrovi za MS te je uvedeno cijepljenje od jata R-30 primjenom soja MS-H. Cijepljenje je rezultiralo održavanjem titra na nižim razinama, što se ovim termoosjetljivim sojem i osigurava, štiteći lokalno na sluznici gornjih dišnih prohoda. Analiza pada uginuća koji je statistički značajan te porast proizvodnje koji je na razini značajnosti od 0,089 ukazuje na to da je poboljšanje pro-grama cijepljenja izravno doprinijelo poboljšanju zdravlja i proizvodnosti longitudinalno kroz jata. To se ogleda i u smanjenju potreba za terapijom od jata R-28 do R-32. Iznimka je jato R-29 kod kojeg je terapija provedena preventivno kako bi se smanjio vertikalni prijenos MS, dok je kod R-32 terapija provedena uslijed izrazitog nekrotičnog enteritisa, vjerojatno kao posljedice hranidbenog disbalansa. Navedenim mjerama u imunoprofilaksi smanjen je pritisak kroz različite zarazne bolesti na jedinke, uz istodobno po-boljšanje uvjeta za očitovanje genetski definirane proizvod-nosti u istim ili sličnim tehničko/hranidbenim uvjetima. Zaključno, rezultati ukazuju na to da kontinuirani moni-toring i izravne profilaktičke mjere koje iz njega proizlaze imaju izravan utjecaj na proizvodne pokazatelje, smanjenje troškova liječenja i bolje zdravstveno stanje u jatima. No, ne manje važno, navedeni rezultati, kao i oni do kojih tek treba doći iz sačuvanih npr. sojeva čine osnovu za daljnja istra-živanja u svrhu karakterizacije sojeva i njihove interakcije u organizmu kokoši, a s ciljem optimizacije imunoprofilakse i biosigurnosti na farmama.
Literatura
ADAIR, B. M. (2000): Immunopathogenesis of chicken anemia virus infection. Dev.Comp. Immunol., 24, 247-255.
CLERMONT, O., S. BONACORSI, E. BINGEN, E. (2000): Rapid and simple determination of the Escherichia coli phylogenetic group. Applied and Environ. Microbiol., 66, 4555-4558.
DE WIT, J. J., W. A. J. M. SWART, T. H. F. FABRI (2010): Efficacy of infectious bronchitis virus vaccinations in the field: association between the α-IBV IgM response, protection and vaccine application parameters. Avian Pathology, 39, 123-131.
EL JAKEE, J. K., G. M. EL AMRY, A. M. HESSAIN, H. A. HEMEG, S. M. SHAFEI, I. M. MOUSSA (2016): Production and evaluation of autogenous vaccine against avian colibacil-losis. The Journal of Animal and Plant Sciences, 26, 79-87
GOTTSTEIN, Ž., D. HORVATEK TOMIĆ, M. LUKAČ, G. NEDELJKOVIĆ, E. PRUKNER-RADOVČIĆ, L. LOZICA, I. ČELEBIĆ, M. VLAHEK, N. DOLENČIĆ, Ž. GLAVAK, Z. BANOVEC, LJ. VINDIŠ (2017a): Cijepljenje protiv APEC sojeva E. coli primjenom autogenog cjepiva na roditeljskim farmama teške linije kokoši u Hrvatskoj. Zbornik Peradarski dani 2017, Šibenik, Hrvatska, 61-66.
GOTTSTEIN, Ž., D. HORVATEK TOMIĆ, M. LUKAČ, G. NEDELJKOVIĆ, E.Z. GALINDO MEDINA, E. PRUKNER-RADOVČIĆ, M. VLAHEK, Z. BANOVEC, LJ. VINDIŠ (2017b): Serološki monitoring virusnih bolesti u roditeljskih jata teških linija kokoši u Hrvatskoj. Zbornik Peradarski dani 2017, Šibenik, Hrvatska, 112-115.
GOTTSTEIN, Ž., L. LOZICA, D. HORVATEK TOMIĆ, G. NEDELJKOVIĆ, M. LUKAČ, E. PRUKNER-RADOVČIĆ (2018): Autogenous vaccine against Escherichia coli and Gallibacterium anatis reduces losses and improves production on layer farms. Proceedings of the XVth European Poultry Conference Dubrovnik, Croatia, 149.
HAGOOD, L.T., T.F. KELLY, J. C. WRIGHT, F. J. HOERR (2000): Evaluation of chicken infectious anemia virus and associated risk factors with disease and production losses in broilers, Avian Dis., 44, 803-808.
HEIDEMANN OLSEN, R., M. BISGAARD, J. P. CHRISTENSEN, S. KABELL, H. CHRISTENSEN (2016): Pathology and Molecular Characterization of Escherichia coli Associated With the Avian Salpingitis-Peritonitis Disease Syndrome. Avian Diseases, 60, 1-7.
HORVATEK TOMIĆ, D., M. LUKAČ, G. NEDELJKOVIĆ, E. PRUKNER-RADOVČIĆ, I. TOTH, L. LOZICA, M. VLA-HEK, Z. BANOVEC, Ž. GOTTSTEIN (2017): Filogenetska raznolikost APEC sojeva izdvojenih na farmama roditeljskih jata teških linija u Hrvatskoj. Zbornik Peradarski dani 2017, Šibenik, Hrvatska, 57-60.
DU mikoplazme HORVATEK TOMIĆ, D., L. LOZICA, G. NEDELJKOVIĆ, M.
LUKAČ, E. PRUKNER- RADOVČIĆ, Ž. GOTTSTEIN (2018): Prevalence of Mycoplasma synoviae in layer poultry flocks in Croatia. Proceedings of the XVth European Poultry Conference Dubrovnik, Croatia, 530.
JAKOBSEN, L., P. GARNEAU, G. BRUANT, J. HAREL, S. S. OLSEN, L.J. PORSBO (2012): Is Escherichia coli urinary tract infection a zoonosis? Proof of direct link with production animals and meat. Eur.J.Clin.Microbiol.Infect.Dis., 31, 1121-1129.
KLEVEN, S. H. (2008): Control of Avian Mycoplasma Infections in Commercial Poultry Avian Diseases, 52, 367-374.
LI, L., I. THØFNER, J.P. CHRISTENSEN, T. RONCO, K. PEDERSEN, R. H. OLSEN (2017): Evaluation of the efficacy of an autogenous Escherichia coli vaccine in broiler breeders. Avian Pathology, 46, 300-308.

PROIZVODNI POKAZATELJI NAKON PRIMJENE AUTOGENOG CJEPIVA PROTIV PTIČJIH PATOGENIH SOJEVA E. COLI I POBOLJŠANJA PROGRAMA CIJEPLJENJA U JATIMA TEŠKE LINIJE KOKOŠI
103
MELAMED D., G. LEITNER, E. DAN HELLER (1991): A Vaccine against Avian Colibacillosis Based on Ultrasonic Inactivation of Escherichia coli. Avian Diseases, 35, 17-22.
NAIR, V. (2018): Spotlight on avian pathology: Marek’s disease. Avian Pathology, 47, 440-442.
NEDELJKOVIĆ, G., L. LOZICA, A. GAVRILOVIĆ, Ž. GOTTSTEIN (2018): Infectious bronchitis virus- neglected (persistent) troublemaker in poultry. Proceedings of 11th International Congress for Veterinary Virology- ESVV 2018., Vienna, Austria.
OTAKI, KAZUKO SAITO, M. TAJIMA, Y. NOMURA (1992): Persistence of maternal antibody to chicken anaemia agent and its effect on the susceptibility of young chickens. Avian Pathology, 21, 147-151.
PIRES-DOS-SANTOS, T., M. BISGAARD, H. CHRISTENSEN (2013): Genetic diversity and virulence profiles of Escherichia coli causing salpingitis and peritonitis in broiler breeders. Veterinary Microbiology, 162, 873-880.
PRODUCTION PARAMETERS AFTER AUTOGENOUS APEC E. COLI VACCINE APPLICATION AND IMPROVEMENT OF VACCINATION PROGRAM IN BROILER BREEDER FLOCK
Abstract
Vaccination of poultry flocks is one of the basic measures in poultry health protection, but humans as final consumer as well. This measure is highlighted in breeder flocks, which have to protect and transfer part of it to the offspring. Vaccination is most frequent against viral diseases, but bacterial diseases could be successfully controlled using vaccines as well. Of bacterial pathogens, APEC E. coli strains are the most common cause of problems on breeder and layer farms, usually in combination with infectious bronchitis, Gallibacterium anatis and Mycoplasma synoviae. In this research performed on a broiler breeder farm, a high prevalence of APEC strains was detected, together with high pathogenicity and high antimicrobial resistance. For control of colibacillosis, autogenous vaccine was applied in 5 consecutive flocks. Together with that, vaccination program was improved regarding IB, CAV, REO, ART and MS vaccination. Results showed continuous and statistically significant reduction of mortality during the production period, from 20.74% to 9.78%, reduced number of therapies, while the number of eggs per housed hen increased by 7.5 eggs on average. The results obtained show that continuous monitoring of health status on farms is needed, together with improvement of vaccination program and vaccination methods, and with continuous improvement of biosecurity measures on farms. Key words: broiler breeder flock, APEC, autogenous vaccine, vaccination program

PERADARSKI DANI 2019. 104
RAPID DIFFERENTIATION OF INFECTIOUS BRONCHITIS VIRUS STRAINS BY RESTRICTION FRAGMENT LENGTH POLYMORPHISM ANALYSIS
Katarina Huić Babić, Leonida Kutle, Lana Ljuma Skupnjak, Anto Vrdoljak
Genera d.d. (Genera d.d. part of Dechra Pharmaceuticals PLC Group), Rakov Potok, Croatia
This work was supported by the European Regional Development Fund, Cohesion Fund and The Government of the Republic of Croatia.
Summary
Avian infectious bronchitis (IB) is an acute, highly contagious, viral disease that causes significant losses to the poultry industry worldwide. Since clinical signs (respiratory distress, nephritis, reduced egg production, etc.) are not highly specific, fast and reliable diagnostic methods are required to identify infectious bronchitis virus (IBV) infections when clinical symptoms are observed in the field. Although serology and virus isolation are still considered as the gold standard in the diagnosis, they show numerous drawbacks in comparison to molecular based methods. Therefore, the objective of the present study was to develop and establish a fast and reliable PCR method for simultaneous detection and differentiation of several most important IBV genotypes in a single reaction - H120 (GI-1 lineage), 793B (GI-13 lineage), QX-like (GI-19 lineage) and Variant-2 like (GI-23 lineage). A total of three field isolates and four reference vaccinal strains were used in this study. The partial S1 glycoprotein gene of these viruses was analyzed by reverse-transcriptase-polymerase chain reaction and restriction fragment length polymorphism (RT-PCR RFLP), using the restriction enzyme HinfI. The RFLP patterns led to the classification of these isolates and vaccine strains into four distinct genotypes. In order to better characterize investigated IBV genotypes, their phenotypic characterization was performed including pathogenicity in chicken embryos, ability to grow in different cell cultures, as well as replication ability in embryos and allantoic fluid. The obtained results indicate that there are some differences in phenotypic characteristics among IBV genotypes, but this approach is not sensitive enough for an unambiguous identification. In contrast, the developed two-step PCR RFLP assay enabled a rapid and reliable tool for identifying and differentiating the most prevalent IBV genotypes in a single test run. Key words: infectious bronchitis, RFLP, poultry, virus identification, vaccine Introduction Avian infectious bronchitis (IB) is an acute, highly contagious disease of the respiratory, reproductive and urogenital tracts of chickens that causes significant losses to the poultry industry worldwide (Huang et al., 2014; Jackwood and de Wit, 2013; Kwon et al., 1993; Majdani et al., 2010). The causative agent, infectious bronchitis virus (IBV), is a single-stranded positive-sense RNA virus that belongs to the Coronaviridae family (Majdani et al., 2010; Xu et al., 2016). The genome of this enveloped virus consists of approximately 27.6 kb, and encodes major structural proteins: the glycosylated spike (S), the envelope (E), the membrane (M), and the phosphorylated nucleo-
capsid (N) protein (Huang et al., 2014). Four non-structural proteins (3a, 3b, 5a, and 5b) are also encoded (Cavanagh, 2007; Gelaye et al., 2013). IBV S glycoprotein that consists of two subunits (S1 and S2) is responsible for attachment of virus to host cells and activation of the virion membrane fusion with host cell membrane (Casais et al., 2003). Variations in S1 protein have been mostly targeted for genotypic characterization and for distinguishing among different IBV serotypes (Abdel-Moneim et al., 2006; Bande et al., 2016). In 2016, Valastro et al. suggested a new and coherent classification scheme based on the complete S1 sequence phylogenetic analysis, dividing IBV strains into 6 genotypes comprising 32 distinct lineages (Valastro et al., 2016).
dr. sc. Katarina Huić Babić, Genera Inc., Svetonedeljska cesta 2, Kalinovica, 10436 Rakov Potok, Croatia; tel. +385-1-3388646; e-mail: [email protected]

RAPID DIFFERENTIATION OF INFECTIOUS BRONCHITIS VIRUS STRAINS BY RESTRICTION FRAGMENT LENGTH POLYMORPHISM ANALYSIS
105
Since the clinical signs (respiratory disease, nephritis, poor egg production, etc.) are not highly specific to IBV, diagnostic tools are necessary to identify infections by IBV when clinical symptoms are observed in the field. These diagnostic tools may include typing of the isolated IBV in order to select the optimum vaccination program with the best chance of achieving sufficient protection of the flocks (Villarreal, 2010). Conventional and more sophisticated methods that have been used for IBV diagnosis may include virus isolation in embryonated eggs, cultivation in tracheal rings, PCR based assays, as well as indirect methods, e.g., immuno-fluorescence assay, hemagglutination inhibition test, virus neutralization test, etc. (Abro, 2012; Bande et al., 2016; De Wit, 2000). Although diagnostic methods such as serology or virus isolation are still considered as the gold standard, they are time consuming, laborious and less sensitive than molecular PCR based methods and genome sequencing (Acevedo et al., 2013; Adzhar et al., 1997; Chen et al., 2010). As distinguishing mutations are not concentrated in one genome region, not all types of conventional PCR/qRT-PCR assays are able to discriminate among different viral strains based on genomic sequence variability (Kiss et al., 2015). Currently, S1 gene sequencing is considered to be the most effective method for IBV detection and differentiation among strains, wherein 95% or more sequence similarity generally indicates that strains are related (Acevedo et al., 2013; Hewson et al., 2009; Lee et al., 2003). Although identification by sequencing is nowadays a routine analysis in many laboratories, simpler identification methods re-quiring fewer resources may still be preferred by most, especially when rapid diagnosis is crucial during an outbreak. Therefore, the aim of this work was to establish and evaluate rapid and reliable PCR-RFLP method for simultaneous detection and differentiation of several major genotypes of IBV in a single reaction; H120 (GI-1 lineage), 793B (GI-13 lineage), QX-like (GI-19 lineage) and Variant 2-like (GI-23 lineage). Moreover, phenotypic characteristics of vaccine and wild-type strains were determined to provide better IBV characterization.
Materials and methods
Viral strains and RNA extraction The IBV strains used in this study were field isolates Is1/11 isolated from a farm located in Turkey (2015), Is2/08 from broilers with respiratory disease (Germany, 2008) and Is3/15 from a breeder farm located in the north-east Croatia (2011). All field isolates were propagated in 9-10 day old specific pathogen free (SPF) embryonated chicken eggs and iden-tified by sequencing. According to the results obtained by sequencing, samples were labeled as shown in Table 1. Four different reference vaccinal IBV strains Avishield IBV H120 (Genera, Croatia), Nobilis IB 4/91 (Intervet, Holland), TAbic IB VAR206 (Phibro Vaccines, Israel) and Poulvac IBQX (Zoetis, USA) were also analyzed.
RT-PCR RNA was extracted from either allantoic fluid harvested from inoculated SPF chicken eggs or directly from freeze dried vaccine using High Pure Viral RNA Extraction Kit (Roche). RT-PCR reaction was performed using Affinity Script One-Step RT-PCR Kit (Agilent Technologies, USA). Oligonucleotide primers used to amplify an approximately 1000-bp region of the S gene were as follows: SX1+, 5’-CACCTAGAGGTTTGYTWGCAT-3’ (Fellahi et al., 2017) and S-R, 5’-TACAAAACCTGCCATAACTAACAT-3’ (Wang et al., 2015). Reverse transcription and PCR ampli-fication were carried out in 25 µL of a mix consisting of 12.5 μL Herculase II RT-PCR 2× Master Mix, 1 μL of each primer (10 pmol), 0.5 μL of AffinityScript RT/RNase Block, 3 μL of RNA template (30 ng/uL) and 7 μL of RNAse free water. RT-PCR reactions were performed in a thermocycler (Primus 25 advanced PCR, PeqLab, Germany) with the following cycles: reverse transcription at 45 °C for 30 min, initial denaturation at 92 °C for 1 min, followed by 40 cycles of denaturation, annealing and extension at 92 °C for 20 s, 52 °C for 20 s and 72 °C for 70 s, respectively, and final extension was carried out at 72 °C for 7 min. RT-PCR products were visualized by electrophoresis on 1.5% agarose gel stained with GelRed Nucleic Acid Gel Stain (10000x in H2O, Biotium) and exposure on a UV transilluminator at a wavelength of 260 nm.
Table 1. IBV strains used in the experiment Tablica 1. IBV sojevi korišteni u istraživanju
Sample ID/Oznaka uzorka Origin/Podrijetlo Classification according/ Klasifikacija prema Valastro et al. 2016
Is1/11_V2 Field isolate/Terenski izolat GI - 23
Is2/08_QX Field isolate/Terenski izolat GI - 19
Is3/15_793B Field isolate/Terenski izolat GI - 13
TAbic IB VAR206 Vaccine strain/Cjepni soj GI - 23
Poulvac IBQX Vaccine strain/Cjepni soj GI - 19
Nobilis IB 4/91 Vaccine strain/Cjepni soj GI – 13
Avishield IBV H120 Vaccine strain/Cjepni soj GI - 1

RAPID DIFFERENTIATION OF INFECTIOUS BRONCHITIS VIRUS STRAINS BY RESTRICTION FRAGMENT LENGTH POLYMORPHISM ANALYSIS
106
RFLP analysis Prior to restriction analysis, PCR amplicons were purified with EZNA Cycle Pure Kit (Omega Bio-Tek, USA) and sequenced. The sequencing was performed by Eurofins MWG Operon sequencing service. The resulting PCR product sequences were analyzed for restriction enzyme recognition sequences using the online tool NEBcutter V2.0 (http://tools.neb.com/NEBcutter2/index.php). The restriction enzyme HinfI was chosen from the restriction map to give bands with sufficiently distinguishable size difference. Restriction fragment length polymorphism (RFLP) analysis was done by digesting 5 μL of each PCR product with restriction endonuclease HinfI (New England BioLabs) at 37 °C for 1 h, followed by heat inactivation at 80 °C for 20 min. After digestion, the DNA fragments were separated in a 3% high resolution agarose gel and visualized by staining with GelRed. The gel was observed under UV light and differences in the pattern were classified.
Quantification of IBV in allantoic fluid and embryos by real-time RT-PCR Infectious bronchitis virus isolates and reference samples were inoculated into the allantoic cavities of 10-day-old SPF embryonated eggs and incubated for 30 h at 37.5 °C. Eggs were cooled at 4 °C overnight before allantoic fluid and embryos were harvested. Inoculum was 103.6 EID50 per egg. Whole embryos were harvested, diluted 1:1 (w/v) with sterile cold water for injection (WFI) and homogenized using IKA Turax high-shear homogenizer. Homogenate was centrifuged at 1,000 xg for 10 min at 4°C and the supernatant was used for RNA extraction. Harvested allantoic fluid was diluted 1:1 (v/v) with WFI prior to viral RNA isolation. RNA was isolated as described previously. Quantitative real-time PCR was performed on a LightCycler 480 II (Roche) using qScript XLT 1-step RT-qPCR ToughMix (Quantabio) according to the manufacturer's instructions. A downstream primer AIBV-fr (5’-ATGCTCAACCTTGTCCCTAGCA-3’), an upstream primer AIBV-as (5’-TCAAACTGCGGATCATCACGT-3’) and a TaqMan® probe AIBV-TM (FAM-TTGGAAGTAGAGTGACGCCCAAACTTCA-BHQ1) (Meir et al., 2010) were used to amplify a 130-bp fragment of IBV N gene. All samples were tested in duplicate to ensure reproducibility of the results. Thermal profile included the following steps: 50 °C for 25 min for reverse transcription, a pre-denaturation at 95 °C for 1 min, followed by 40 cycles of denaturation at 95 °C for 10 s and annealing/extension at 60 °C for 40s. Pathogenicity in chicken embryos In order to analyze the pathogenicity of IBV isolates and attenuated vaccine strains in chicken embryos, all samples were inoculated via the allantoic cavity (103.6 EID50 per egg) into ten 10-day-old embryonated SPF eggs per sample. Eggs were candled daily to record embryo mortality. After 7 days, the remaining eggs were chilled at 4 °C and observed for typical embryo lesions caused by IBV such as dwarfing, stunting, and curling.
Infectious bronchitis virus growth in cell culture Primary chicken embryo fibroblast (CEF) culture was prepared from 10-day-old SPF embryonated hens’ eggs. Embryo’s head, limbs and viscera were removed, the remaining tissue was squeezed through a syringe into 0.25% trypsin-EDTA solution and vortexed at 37 °C for 15 minutes. The cell suspension was filtered through a mesh and centrifuged at 670 g for 5 minutes. The pellet was suspended in the test medium, the cells were counted and seeded in 96-well plates in a concentration of 2.7×105 cells/cm2. Primary chicken embryo kidney (CEK) cell culture was prepared from 17-day-old SPF embryonated hens’ eggs. Embryo’s kidneys were removed, washed 3 times in PBS and then vortexed at 37 °C for 10 minutes in 0.25% trypsin-EDTA solution. The cell suspension was filtered through mesh and centrifuged at 670 g for 5 minutes. The pellet was suspended in the test medium, the cells were counted and seeded in 96-well plates in a concentration of 3×105 cells/cm2. Vero cells (ECACC, Cat. No. 84113001) were prepared by trypsinization of cell culture. The cells were counted and seeded in 96-well plates in a concentration of 6.3×104 cells/cm2. The three concentrations (10, 50 and 100 TCID 50 in 100 µL) of each IBV strain were prepared and cultured cells were infected with the virus and then observed daily for cytopathic effects (CPE). All cells were incubated in a humidified incubator at 37 °C and 5% CO2.
Results and discussion
As S1 gene is shown to be one of the most variable regions of IBV genome, it is often selected as the target for PCR RFLP analysis (Lee et al., 2003; Montassier et al., 2008; Wang and Huang, 2000). In the present study, RT-PCR RFLP approach was used to genotype three different field isolates and to compare them against different vaccine strains of IBV. In order to differentiate IB viral strains, oligonucleotide primers complementary to approximately 1000 bp segment were evaluated. One-step reverse transcription PCR yielded the desired PCR amplicons in all tested samples. RT-PCR products were further submitted to RFLP analysis. For this purpose, the RT-PCR products were digested with restriction enzyme HinfI. The restriction fragments were analyzed by agarose gel electrophoresis, and the obtained sizes were consistent with those theoretically predicted by the online tool NEBcutter V2.0 (Table 2). The difference in RFLP patterns allowed complete differenti-ation between four different IBV genotypes: Variant 2 (GI-23 lineage), QX-like (GI-19 lineage), 793B (GI-13 lineage) and H120 (GI-1 lineage). The presence of a single RFLP band, obtained after digestion of DNA segment from field isolate Is1/11_V2, as well as reference vaccine strain TAbic IB VAR206 indicated absence of the restriction site (Fig. 1).

RAPID DIFFERENTIATION OF INFECTIOUS BRONCHITIS VIRUS STRAINS BY RESTRICTION FRAGMENT LENGTH POLYMORPHISM ANALYSIS
107
Table 2. Fragment sizes in base pairs (bp) after HinfI digest Tablica 2. Veličine fragmenata (bp) nakon digestije s HinfI
GI -23 lineage/linija
Variant 2
GI -19 lineage/linija QX-like
GI -13 lineage/linija 793B group
GI -1 lineage/linija H120
No restriction site/ Nema mjesta cijepanja 418 824 399
317 120 279 260 52 215 103
Figure 1. Restriction fragment profiles after HinfI digest of representative IBV strains. Lane 1: 50 bp marker (Thermo
Scientific); Lane 2: TAbic IB VAR206; Lane 3: Poulvac IBQX; Lane 4: Nobilis IB 4/91; Lane 5: Avishield IB H120; Lane 6: Is1/11_V2; Lane 7: Is2/08_QX; Lane 8: Is3/15_793B
Slika 1. Restrikcijski profil nakon digestije s HinfI. Stupac 1: 50 bp marker (Thermo Scientific); Stupac 2: TAbic IB VAR206; Stupac 3: Poulvac IBQX; Stupac 4: Nobilis IB 4/91; Stupac 5: Avishield IB H120; Stupac 6: Is1/11_V2; Stupac 7: Is2/08_QX; Stupac 8: Is3/15_793B
In contrast, three restriction bands of 260, 317 and 418 bp, obtained after HinfI digestion of field strain Is2/08_QX and vaccinal strain Poulvac IBQX revealed the presence of two restriction sites. Furthermore, IBV samples Is3/15_793B and
Nobilis IB 4/91 vaccine strain generated three DNA fragments of 52, 120 and 824 bp, while reference vaccine strain Avishield IB H120 produced four DNA fragments of 103, 215, 279 and 399 bp. Based on the said differences in

RAPID DIFFERENTIATION OF INFECTIOUS BRONCHITIS VIRUS STRAINS BY RESTRICTION FRAGMENT LENGTH POLYMORPHISM ANALYSIS
108
Hinfl digestion patterns, it was possible to clearly distinguish different IB genotypes included in the study. This is in good agreement with previous findings that RFLP patterns of S1 gene may distinguish among different IBV serotypes (Callison et al., 2001; Jackwood et al., 2005; Jackwood and Kwon, 2001). Moreover, the obtained RFLP genotyping results are in complete agreement with the patterns predicted from the amino acid sequences. However, no difference in RFLP patterns was observed between field isolates and vaccine strains of the same serotype, indicating that HinfI targeted sites in the analyzed region remained unchanged. Induction of cytopathic effect in CEK, CEF and Vero cells In comparison with other coronaviruses, IBV is not easily adaptable to cell culture (Chhabra et al., 2016). As reported previously, distinct cytopathic effects and apoptosis may only be detected with few strains in certain mammalian and primary cells (Han et al., 2017; Otsuki et al., 1979), while the majority of isolates require several passages of adaptation. To determine whether there is a difference in virus tropism among investigated IBV strains, Vero cell line and two primary cell lines from SPF embryos (CEK and CEF) were used. Cultured cells were infected with viruses in three different concentrations and then observed daily for cytopathic effects. As expected, chicken embryo kidney cells were most susceptible to infection with IB virus. In this cell type, CPE was observed clearly at 48 h p.i. and was fully developed by 72 h p.i. Cytopathic effect was characterized by marked granulation of cell cytoplasm and by cell rounding. CPE was detected in all attenuated vaccine strains, except for Poulvac IB QX, whereas in H120 strain it was most pronounced. Among field isolates, only the QX-like strain produced a distinct CPE on CEK, while the other two isolates showed no CPE on any of the tested cells (Table 3). Taking into account nephropathogenicity of QX-like field strains, this was not unexpected (Terregino et al., 2008; Benyeda et al., 2009). On Vero cells, only reference strain H120 induced moderate CPE. Our results confirmed previous findings that CEK cells show the highest sensitivity for IBV strains (De Wit, 2000), but no universally susceptible cell line is available.
Quantification of viruses in allantoic fluid and embryos It is known that most IBV strains grow well in developing chicken embryos and that maximum titer in allantoic fluid is achieved 24 to 48 hours post inoculation (Bande et al., 2016; De Wit, 2000). In this study, we compared the propagation potential of tested IBV strains by measuring the virus genome copy number in allantoic fluid and embryos by quantitative RT-PCR (qRT-PCR), which has become increasingly popular for detecting and quantifying viral load in different samples because it is simple, fast and less laborious compared to other techniques (Moody et al., 2000). Following inoculation of 103.6 EID50 per egg, RNA was extracted from allantoic fluid and from homogenized embryos after 30-h incubation period. In all tested samples, the IBV copy number in allantoic fluid was higher compared to embryonic tissue (Table 4). The resulting Ct values after calculation to virus titer showed the difference in concentration between allantoic fluid and embryos ranging from 101.08 EID50 (Is1/11_V2) to 102.47 EID50 (TAbic IB VAR206) per mL. The results obtained showed that egg-attenuated vaccine strains replicated faster than non-egg-adapted field isolates, which is in good agreement with Tsai et al., 2016. Viral pathogenesis in chicken embryos The pathogenicity of three isolates and four vaccine strains was studied using the standard embryo lethality test. Ten eggs per sample were inoculated via the allantoic cavity with titer 103.6 EID50 per egg and eggs were candled daily to monitor embryo vitality. Embryo deaths within the first 24 hours after virus inoculation were regarded as nonspecific. Only two vaccinal strains (Avishield IB H120 and Poulvac IBQX) caused embryo death during seven days of incubation. In the early stage (2-4 days p.i.), mortality was seen only in Avishield IBH120 group where 3/10 embryos were found dead (Table 5). Another three Avishield IB H120 and two Poulvac IBQX vaccinated embryos died in the later stage after inoculation. After 7 days, the remaining eggs were chilled at 4 °C and observed for typical embryo lesions of IBV such as dwarfing, stunting and curling. Differences were observed between wild strains and attenuated viruses of the same genotype, but also within the same groups.
Table 3. Presence of cytopathic effect (CPE) on tested cell cultures after infection with IBV strains Tablica 3. Prisutnost citopatskog efekta (CPE) na istraživanim staničnim kulturama nakon infekcije različitim sojevima IBV
Is1/11_V2 Is2/08_QX Is3/15_793B TAbic IB VAR206 Poulvac IBQX Nobilis IB
4/91 Avishield IB
H120
CEK - ++ - ++ - + +++
CEF - - - - - - -
Vero - - - - - - ++
Marks/Oznake: – no/nema CPE; + weak/slab CPE; ++ moderate/umjeren CPE, +++ strong/jak CPE

RAPID DIFFERENTIATION OF INFECTIOUS BRONCHITIS VIRUS STRAINS BY RESTRICTION FRAGMENT LENGTH POLYMORPHISM ANALYSIS
109
Table 4. Real time PCR IBV quantification in allantoic fluid (AF) and embryos Tablica 4. Kvantifikacija IBV u alantoinskoj tekućini (AF) i embrijima pomoću kvantitativnog PCR-a u stvarnom vremenu
Sample / Uzorak Average / Prosječni Ct AF
Average/ Prosječni Ct embryos/embriji
Δ Ct (embryos-AF) (embriji – AF)
Δ titer/titar (10^ EID50) /mL
Is1/11_V2 18.80 20.06 1.26 1.08
Is2/08_QX 16.83 19.83 3.00 1.60
Is3/15_793B 18.89 22.47 3.58 1.78
TAbic IB VAR206 16.91 22.78 5.87 2.47
Poulvac IBQX 16.65 18.35 1.70 1.21
Nobilis IB 4/91 17.03 18.68 1.65 1.20
Avishield IB H120 15.67 19.03 3.37 1.71 Table 5. Death and IB specific lesions from field isolates and vaccine strains on chicken embryos Tablica 5. Smrt i promjene embrija uzrokovane infekcijom različitim terenskim izolatima i cjepnim sojevima IBV
Sample Uzorak
Infeced embryos Inficirani embriji
Nonspecific death/ Nespecifična smrt
Death number/Broj uginulih 2-7 DPI*
(virus specific/ virusno specifično)
Embyos with IB typical lesions/
Embriji s IB specifičnim promjenama
IB positive embryos IB pozitivni embriji**
(%)
Is1/11_V2 10 0 0 5 5/10 (50)
Is2/08_QX 10 1 0 7 7/9 (78)
Is3/15_793B 10 0 0 5 5/10 (50)
TAbic IB VAR206 10 0 0 5 5/10 (50)
Poulvac IBQX 10 0 2 8 10/10 (100)
Nobilis IB 4/91 10 0 0 3 3/10 (30)
Avishield IB H120 10 0 6 4 10/10 (100)
* days post infection/dana poslije infekcije; **by visual inspection/vizualni pregled Neither the wild (Is1/11_V2) nor the vaccine Variant 2 strains were lethal for embryos, but one half (5/10) of the embryos showed lesions such as dwarfing, stunting and slightly curling. However, vaccinal strain induced a higher level of specific IBV lesions than the wild strain (Fig. 2C, 3D). The remaining five embryos did not show any changes in comparison to uninoculated control. Results obtained after infection with 793B-like wild strain showed stunting and dwarfing in 4/10 and curling in 1/10 inoculated chicken embryos (Fig. 2E). Surprisingly, characteristic lesions in embryos were least visible when inoculated with vaccine strain belonging to 793B-genotype (Fig. 2F), which is more common for non-egg-adapted wild strains (De Wit, 2000; Tsai et al., 2016). Vaccinal strain H120 caused embryo death before day 7 p.i. in 60% of the inoculated embryos and the remaining four embryos were completely curled into a spherical form, which is a typical lesion caused by IB
infection (Fig. 2B) (Cavanagh and Naqi, 2003; Geerligs et al., 2011). Similar lesions were observed in eight surviving embryos infected with IB QX vaccinal strain (Fig. 2H). Conversely, the wild QX strain mostly caused less pronounced lesions such as dwarfism (Fig. 2G). These findings are in alignment with previous findings (Bijlenga et al., 2004; Tsai et al., 2016) that vaccine strains may cause embryo death and specific lesions such as dwarfing, stunting and curling to a higher extent than wild strains of the same genotype. The only exception was strain IB 4/91 (793B group), which caused visible lesions in only 30% of inoculated embryos, despite the fact that the virus propagated to high titers, as evidenced by PCR. Overall, the results revealed that differentiation of genotypes could not be based solely on the presence and nature of lesions, but it may be a useful indicator for further characterization.

RAPID DIFFERENTIATION OF INFECTIOUS BRONCHITIS VIRUS STRAINS BY RESTRICTION FRAGMENT LENGTH POLYMORPHISM ANALYSIS
110
Figure 2. Gross changes in chicken-embryos caused by IBV at 17 days of age: A - negative control; B - Avishield IB H120; C -
Is1/11_V2; D - TAbic IB VAR206; E - Is3/15_793B; F - Nobilis IB 4/91; G - Is2/08_QX; H - Poulvac IBQX Slika 2. Promjene na embrijima starim 17 dana uzrokovane IBV infekcijom: A – negativna kontrola; B - Avishield IB H120;
C - Is1/11_V2; D - TAbic IB VAR206; E - Is3/15_793B; F - Nobilis IB 4/91; G - Is2/08_QX; H - Poulvac IBQX In conclusion, our results suggest that RT-PCR RFLP provides a time-saving, sensitive and reliable method for identification and differentiation among several commonly found IBV genotypes. A significant advantage of the presented method is that it is able to detect and simultaneously genotype IBV strains without the need of propagation of the virus in cell cultures or embryonated chicken eggs. Subsequent confirmation by sequencing of the PCR products may be performed, if necessary, but the described method may suffice for most routine analyses. Further work will be focused on increasing the sensitivity of the assay in order to differentiate wild-type and vaccine IBV strains simultaneously in a single reaction. This work was supported by the European Regional Development Fund, Cohesion Fund and The Government of the Republic of Croatia.
References
ABDEL-MONEIM, A. S., F. E. MAGDY, B. S. LADMAN, J. GELB (2006): S1 gene sequence analysis of a nephropathogenic strain of avian infectious bronchitis virus in Egypt. Virol. J., 3, 78-87.
ABRO, S. H. (2012): Molecular characterization and detection of infectious bronchitis virus, Dissertation.
ACEVEDO, A. M., C. L. PERERA, A. VEGA, L. RÍOS, L. CORONADO, D. RELOVA, M. T. FRÍAS, L. GANGES, J. I. NÚÑEZ, L. J. PÉREZ (2013): A duplex SYBR Green I-based real-time RT-PCR assay for the simultaneous detection and differentiation of Massachusetts and non-Massachusetts serotypes of infectious bronchitis virus. Mol. Cel. Probe, 27, 184-192.
ADZHAR, A., R. GOUGH, D. HAYDON, K. SHAW, P. BRITTON, D. CAVANAGH (1997): Molecular analysis of the 793/B serotype of infectious bronchitis virus in Great Britain. Avian Pathol., 26, 625-640.

RAPID DIFFERENTIATION OF INFECTIOUS BRONCHITIS VIRUS STRAINS BY RESTRICTION FRAGMENT LENGTH POLYMORPHISM ANALYSIS
111
BANDE, F., S. S. ARSHAD, A. R. OMAR, M. H. BEJO, M. S. ABUBAKAR, Y. ABBA (2016): Pathogenesis and diagnostic approaches of avian infectious bronchitis. Advan. Vir., 1-11.
BENYEDA, Z., T. MATÓ, T. SÜVEGES, É. SZABÓ, V. KARDI, Z. ABONYI-TÓTH, M. RUSVAI, V. PALYA (2009): Comparison of the pathogenicity of QX-like, M41 and 793/B infectious bronchitis strains from different pathological conditions. Avian Pathol., 38, 449-456.
BIJLENGA, G., J. K. COOK, J. J. GELB, J.J. DE WIT (2004): Development and use of the H strain of avian infectious bronchitis virus from the Netherlands as a vaccine: a review. Avian Pathol., 33, 550-557.
CALLISON, S., M.W. JACKWOOD, D. HILT (2001): Molecular characterization of infectious bronchitis virus isolates foreign to the United States and comparison with United States isolates. Avian Dis., 492-499.
CASAIS, R., B. DOVE, D. CAVANAGH, P. BRITTON (2003): Recombinant avian infectious bronchitis virus expressing a heterologous spike gene demonstrates that the spike protein is a determinant of cell tropism. J. Virol., 77, 9084-9089.
CAVANAGH, D. (2007): Coronavirus avian infectious bronchitis virus. Vet. Res., 38. 281-297.
CAVANAGH, D., S. NAQI (2003): Infectious bronchitis. Dis. Poult., 11, 101-119.
CHEN, H., J. ZHANG, Y. P. MA, L. N. MA, Y. Z. DING, X. T. LIU, X. P. CAI, L. Q. MA, Y. G. ZHANG, Y. S. LIU (2010): Reverse transcription loop-mediated isothermal amplification for the rapid detection of infectious bronchitis virus in infected chicken tissues. Mol. Cell. Probe, 24, 104-106.
CHHABRA, R., S. V. KUCHIPUDI, J. CHANTREY, K. GANAPATHY (2016): Pathogenicity and tissue tropism of infectious bronchitis virus is associated with elevated apop-tosis and innate immune responses. Virology, 488, 232-241.
DE WIT, JJ. (2000): Detection of infectious bronchitis virus. Avian pathol., 29, 71-93.
FELLAHI, S., M. EL HARRAK, S. KHAYI, J. L. GUERIN, J. KUHN, M. EL HOUADFI, M. M. ENNAJI, M. DUCATEZ (2017): Phylogenetic analysis of avian infectious bronchitis virus isolates from Morocco: a retrospective study (1983 to 2014). Virol. Sin., 2, 155-158.
GEERLIGS, HJ., G. J. BOELM, C. MEINDERS, B. STUURMAN, J. SYMONS, J. TARRES-CALL, T. BRU, R. VILA, M. MOMBARG, K. KARACA (2011): Efficacy and safety of an attenuated live QX-like infectious bronchitis virus strain as a vaccine for chickens. Avian pathol., 40: 93-102.
GELAYE, E., C. E. LAMIEN, R. SILBER, E. TUPPURAINEN, R. GRABHERR, A. DIALLO (2013): Development of a cost-effective method for capripoxvirus genotyping using snapback primer and dsDNA intercalating dye. PloS one., 8, e75971.
HAN, X., Y. TIAN, R. GUAN, W. GAO, X. YANG, L. ZHOU, H. WANG (2017): Infectious bronchitis virus infection induces apoptosis during replication in chicken macrophage HD11 cells. Viruses, 9, 198-212.
HEWSON, K., A. H. NOORMOHAMMADI, J. M. DEVLIN, K. MARDANI, J. IGNJATOVIC (2009): Rapid detection and non-subjective characterisation of infectious bronchitis virus isolates using high-resolution melt curve analysis and a mathematical model. Arch. Virol., 154, 649-660.
HUANG, S.W., C. F. HO, K. W. CHAN, M. C. CHENG, J. H. SHIEN, H. J. LIU, C.Y. WANG (2014): The genotyping of infectious bronchitis virus in Taiwan by a multiplex
amplification refractory mutation system reverse transcription polymerase chain reaction. J. Vet. Diagn. Invest., 26, 721-733.
JACKWOOD, M. W., S. DE WIT (2013): Infectious bronchitis. Dis. Poult., 139-159.
JACKWOOD, M. W., D. A. HILT, C. W. LEE, H. M. KWON, S. A. CALLISON, K. M. MOORE, H. MOSCOSO, H. SELLERS, S. THAYER (2005): Data from 11 years of molecular typing infectious bronchitis virus field isolates. Avian Dis., 49, 614-618.
JACKWOOD, M. W., H. M. KWON (2001): Differentiation of avian infectious bronchitis virus serotypes. In.: Google Patents.
KISS, I., T. MATÓ, Z. G. HOMONNAY, J. KOJER, A. FARSANG, Á. BÁLINT, V. PALYA (2015): Survey indicates circulation of 4/91 and QX-type infectious bronchitis viruses in Hungary in 2014', Acta Vet. Hung., 63, 382-388.
KWON, H. M., M. W. JACKWOOD, J. GELB JR (1993): Differentiation of infectious bronchitis virus serotypes using polymerase chain reaction and restriction fragment length polymorphism analysis. Avian Dis., 194-202.
LEE, C.W., D. A. HILT, M. W. JACKWOOD (2003): Typing of field isolates of infectious bronchitis virus based on the sequence of the hypervariable region in the S1 gene. J. Vet. Diagn. Invest., 15, 344-348.
MAJDANI, R., K. MARDANI, A. MORSHEDI, M. V. MARANDI, A. TALEBI (2010): Molecular Comparison of Three Different Regions of the Genome of Infectious Bronchitis Virus Field Isolates and Vaccine Strains. In Veterinary Research Forum, 73-81. Faculty of Veterinary Medicine, Urmia University.
MEIR, R., O. MAHARAT, Y. FARNUSHI, L. SIMANOV (2010): Development of a real-time TaqMan® RT-PCR assay for the detection of infectious bronchitis virus in chickens, and comparison of RT-PCR and virus isolation. J.Virol. Meth., 163, 190-194.
MONTASSIER, M. F., L. BRENTANO, H. MONTASSIER, L. J. RICHTZENHAIN (2008): Genetic grouping of avian infectious bronchitis virus isolated in Brazil based on RT-PCR/RFLP analysis of the S1 gene. Pesqui. Vet. Brasil, 28, 190-194.
OTSUKI, K., K. NORO, H. YAMAMOTO, M. TSUBOKURA (1979): Studies on avian infectious bronchitis virus (IBV). II. Propagation of IBV in several cultured cells. Arch. Virol., 60, 115-122.
TERREGINO, C., A. TOFFAN, M. S. BEATO, R. DE NARDI, M. VASCELLARI, A. MEINI, G. ORTALI, M. MANCIN, I. CAPUA (2008): Pathogenicity of a QX strain of infectious bronchitis virus in specific pathogen free and commercial broiler chickens, and evaluation of protection induced by a vaccination programme based on the Ma5 and 4/91 serotypes. Avian Pathol., 37, 487-493.
TSAI, C. T, H. Y. CHIOU, C. H. WANG (2016): The infectious bronchitis vaccine strain virus is more pathogenic in chicken embryos than the wild virus strain 2575/98. Vet. arhiv, 86, 699-709.
VILLARREAL, L. Y. B. (2010): Diagnosis of infectious bronchitis: an overview of concepts and tools. Rev.Brasil. Ciên. Avíc., 12, 111-14.
WANG, C. H., Y. C. HUANG. (2000): Relationship between serotypes and genotypes based on the hypervariable region of the S1 gene of infectious bronchitis virus. Arch. Virol., 145, 291-300.

RAPID DIFFERENTIATION OF INFECTIOUS BRONCHITIS VIRUS STRAINS BY RESTRICTION FRAGMENT LENGTH POLYMORPHISM ANALYSIS
112
WANG, Z. X., S. M. GAO, Q.W. ZHENG, M. H. SUN, W. X. JIA, R. TAO (2015): Epidemiology and Evolutionary Charac-teristics of Avian Infectious Bronchitis Virus in China. Kafkas Universitesi Veteriner Fakultesi Dergisi, 21.
XU, G., X. Y. LIU, Y. ZHAO, Y. CHEN, J. ZHAO, G. Z. ZHANG (2016): Characterization and analysis of an infectious bronchitis virus strain isolated from southern China in 2013. Virol. J., 13, 40-48.
BRZA METODA IDENTIFIKACIJE I DIFERENCIJACIJE SOJEVA VIRUSA PTIČJEG ZARAZNOG BRONHITISA METODOM PCR-RFLP
Sažetak
Ptičji zarazni bronhitis je akutna, vrlo zarazna bolest koja uzrokuje značajne ekonomske gubitke u peradarskoj industriji širom svijeta. S obzirom na to da klinički znakovi (respiratorne smetnje, nefritis, smanjena nesivost i sl.) nisu visoko specifični za virus ptičjeg zaraznog bronhitisa (IBV), potrebne su brze i pouzdane dijagnostičke metode identifikacije virusa pri pojavi prvih simptoma na terenu. Iako se metode temeljene na serologiji odnosno izolaciji virusa još uvijek smatraju „zlatnim standardom“ u dijagnostici, one pokazuju brojne nedostatke u usporedbi s molekularnim metodama. Stoga je cilj našeg istraživanja bio razvoj i uspostava brze i pouzdane PCR metode za istodobnu detekciju i diferencijaciju četiri najznačajnija genotipa IBV u jednoj reakcijskoj smjesi – H120 (GI-1 linija), 793B (GI – 13 linija), QX (GI – 19 linija) i Variant 2 (GI – 23 linija). U tu svrhu korišteni su referentni cjepni sojevi IBV-a i terenski izolati. Fragment S gena je analiziran metodom reverzne transkripcije-lančane reakcije polimerazom i digestijom pomoću restrikcijskog enzima HinfI, što je rezultiralo uspješnom diferencijacijom istraživanih genotipova. S ciljem bolje karakterizacije cjepnih sojeva i terenskih izolata provedena je i njihova fenotipska karakterizacija koja je uključivala patogenost sojeva za embrije, sposobnost njihova rasta na različitim kulturama stanica te brzinu umnožavanja u embrioniranim SPF jajima. Dobiveni rezultati ukazuju na to da postoje značajne razlike u fenotipskim karakteristikama između nekih sojeva, no ovaj se pristup ne može samostalno koristiti u njihovoj identifikaciji. Za razliku od toga, razvijena metoda RT-PCR RFLP predstavlja brz, osjetljiv i pouzdan dijagnostički alat za identifikaciju i diferencijaciju danas najznačajnijih genotipova ptičjeg zaraznog bronhitisa. Ključne riječi: ptičji zarazni bronhitis, RFLP, perad, identifikacija virusa, cjepiva Ovaj rad je sufinanciran od strane Europskog fonda za regionalni razvoj (EFRR), Kohezijskog fonda te Državnog proračuna RH.

PERADARSKI DANI 2019. 113
PROVJERA SIGURNOSTI I UČINKOVITOSTI CJEPIVA PROTIV ZARAZNE BOLESTI BURZE PREMA ZAHTJEVIMA EUROPSKE FARMAKOPEJE
Anto Vrdoljak1, Olga Zorman Rojs2, Gert Jan Boelm3, Lana Ljuma Skupnjak1
1 Genera d.d., dio Dechra Pharmaceuticals PLC grupe, Rakov Potok, Hrvatska 2 Univerza v Ljubljani, Veterinarska fakulteta, Ljubljana, Slovenija 3 GD Animal Health, Deventer, Nizozemska
Sažetak
Cijepljenje pilića živim atenuiranim cjepivima jedna je od najučinkovitijih metoda kontrole i prevencije zarazne bolesti burze. Nova cjepiva namijenjena primjeni u zemljama Europske Unije moraju zadovoljiti zahtjeve Europske farmakopeje (Ph. Eur.) koji propisuju detaljnu in vitro i in vivo provjeru sigurnosti i učinkovitosti cjepiva, kako matičnih sojeva, tako i svake serije proizvoda. U ovom radu prikazani su rezultati in vitro karakterizacije matičnog soja te rezultati in vivo provjera sigurnosti i učinkovitosti cjepiva protiv zarazne bolesti burze temeljenog na intermedijernom soju VMG91. Opća sigurnost cjepnog soja provjerena je na SPF nesilicama prema Ph. Eur. protokolu; stostruka minimalna doza cjepiva (106.0 TCID50) u SPF pilićima nije izazvala kliničke znakove zarazne bolesti burze tijekom 21 dana nakon cijepljenja. Testom učinkovitosti cjepiva protiv newcastelske bolesti u pilićima prethodno cijepljenima VMG91 sojem pokazano je da cjepivo nema značajan imunosupresivni učinak prema kriterijima Ph. Eur., s obzirom na to da cijepljenje pomoću VMG91 nije rezultiralo statistički značajno manjom zaštitom protiv izazivačke infekcije virulentnim virusom newcastelske bolesti. Testom genetske stabilnosti cjepnog soja primijećena je jedna točkasta mutacija koja uzrokuje promjenu H253Q na VP2 genu nakon in vivo pasažiranja, pojava već ranije opisana kod sličnih intermedijarnih sojeva. Laboratorijske in vivo izazivačke infekcije cijepljenih pilića pokazale su 100% učinkovitost cjepiva i u SPF i u komercijalnim pilićima cijepljenima minimalnom preporučenom dozom. Terenskim studijama na nesilicama i brojlerima potvrđena je zadovoljavajuća sigurnost i učinkovitost cjepiva u proizvodnim uvjetima, uz očuvanje proizvodnih parametara. Rezultati ispitivanja pokazuju prikladnost cjepnog soja za korištenje u zaštiti pilića protiv virulentnih sojeva zarazne bolesti burze, prema kriterijima važećim u Europskoj Uniji. Ključne riječi: zarazna bolest burze; živa atenuirana cjepiva; Europska farmakopeja Uvod
Virus zarazne bolesti burze (engl. Infectious Bursal Disease Virus, IBDV) uzrokuje imunosupresivnu bolest mladih pilića, a primarno napada limfoidno tkivo Fabricijeve burze. Virulentni sojevi zarazne bolesti burze povećavaju smrtnost i djeluju imunosupresivno te na taj način uzrokuju značajnu ekonomsku štetu peradarskoj industriji diljem svijeta. Pilići koji obole u ranoj dobi naročito su podložni imunosupre-sivnim tegobama te lošije odgovaraju na cjepiva i podložni su infekcijama virusima i bakterijama koji su inače ne-patogeni u imunokompetentnim pticama (Berg, 2000.). Mladi komercijalni pilići obično su tijekom prvih dana ži-vota dobro zaštićeni od infekcije IBDV-om naslijeđenim
materinskim protutijelima. Kako se koncentracija mate-rinskih protutijela smanjuje, pasivna imunost više nije do-voljna za zaštitu od virulentnih cirkulirajućih sojeva, nego je potrebno cijepljenjem stvoriti aktivnu imunost (Berg, 2000.). Danas su komercijalno dostupna živa atenuirana, inak-tivirana, imuno-kompleksna te vektorska cjepiva protiv zarazne bolesti burze. Zbog lakoće primjene i relativno niže cijene, živa atenuirana cjepiva su najraširenija, a mogu se prema virulentnosti cjepnog soja razvrstati u blaga, blaga intermedijarna, intemedijarna i vruća cjepiva (Eterradossi i Saif, 2008.). Glavni problem pri upotrebi živih cjepiva je kako izbjeći interferenciju s materinskim protutijelima, tj. kako pravilno odrediti optimalnu dob cijepljenja (Jackwood, 2017.).
Dr. sc. Anto Vrdoljak; Genera d.d., dio Dechra Pharmaceuticals PLC Grupe; Svetonedeljska cesta 2, Kalinovica, 10436 Rakov Potok, Hrvatska; Tel. +385-1-3388635; Email: [email protected]

PROVJERA SIGURNOSTI I UČINKOVITOSTI CJEPIVA PROTIV ZARAZNE BOLESTI BURZE PREMA ZAHTJEVIMA EUROPSKE FARMAKOPEJE
114
Zakonski okvir za registraciju veterinarskih lijekova i cje-piva značajno se pooštrio tijekom zadnjih desetljeća, što je rezultiralo značajno poboljšanom kvalitetom i sigurnošću lijekova. Kao članica EU, Republika Hrvatska je obvezna implementirati i provoditi regulatorni okvir za proizvodnju, kontrolu i distribuciju lijekova definiran i dokumentima Europske agencije za lijekove (engl. European Medicines Agency, EMA), Europske farmakopeje (engl. European Pharmacopoeia, Ph. Eur.) i Europske komisije (engl. European Commision, EC). Iako strogi regulatorni zahtjevi povećavaju kvalitetu lijeka, oni također značajno produljuju i poskupljuju razvoj i registraciju lijeka, što se konačno odražava i na dostupnost lijeka za krajnjeg korisnika. U ovom radu prikazan je sažetak razvoja, molekularne ka-rakterizacije te laboratorijskih i terenskih ispitivanja cjepiva Avishield® IBD INT, živog atenuiranog cjepiva protiv zarazne bolesti burze temeljenog na intermedijarnom soju VMG91. Cjepivo je razvijeno i karakterizirano prema tre-nutno važećoj EU regulativi, a prikazani sažetak primjerom ilustrira kompleksnost i zahtjevnost suvremenog razvoja lijeka.
Materijali i metode
Cjepiva i virusi
U radu je korišteno intermedijarno cjepivo Avishield® IBD INT proizvođača Genera d.d. Cjepivo sadrži intermedijarni soj VMG91 i registrirano je za upotrebu u većini zemalja Europske Unije. Peta pasaža cjepiva (VMG91p) dobivena je in vivo pasažiranjem matičnog soja cjepiva u SPF pilićima (Valo BioMedia, Njemačka). Komercijalno dostupno cje-pivo koje također sadrži usporediv intermedijarni cjepni soj IBDV korišteno je kao komparator u terenskim studijama. Vrlo virulentni terenski izolat DV86 korišten je u studijama za izazivačke infekcije. Prema uputama Ph. Eur., za studije imunosupresije korišteno je komercijalno dostupno cjepivo protiv newcastelske bolesti (ND) bazirano na soju Hitchner B1, a za izazivačke infekcije vrlo virulentni izolat Herts 33/56 (Ph. Eur., 2013.).
In vitro karakterizacija cjepnog soja
Sekvenciranje dijela VP2 gena provedeno je uobičajenim tehnikama korištenjem početnica F-IBDV-576s (5’-GCCAACATCAACGACAAAATTGGG-3’) i IBDV-R (5’-ATCCTGTTGCCACTCTTTCGTAGG-3’) (Zorman Rojs i sur., 2008.; Cao i sur., 1998.). Dobivene sekvence uspoređi-vane su s javno dostupnim sekvencama cjepnih i virulentnih sojeva IBDV-a D78 (EU162087.1 i EU162090.1), Cu-1 (D00867.1), DV86 (AJ878899.1) i UK661 (X92760.1). Korišten je alat NCBI BLAST (Basic Local Alignment Search Tool).
Stabilitet cjepiva u otopini
Titar virusa u vodenoj suspenziji cjepiva praćen je pri tem-peraturi 20-25 °C tijekom 3 sata nakon rekonstitucije cjepiva
u vodi slobodnoj od klora. Uzorci su titrirani 0, 1, 2 i 3 sata nakon rekonstitucije cjepiva. Titracije su rađene korištenjem primarne kulture pilećih fibroblasta.
Studije na životinjama
Studije sigurnosti i učinkovitosti na ciljnim životinjama pro-vedene su prema smjernicama danima u odgovarajućim do-kumentima Ph. Eur. (Ph.Eur., 2013.), (Ph.Eur., 2013.), (Ph.Eur., 2017.) i EMA (EMA, 1999.), (EMA, 2001.), (EMA, 2005.), (EMA, 2000.). Sigurnost cjepiva potvrđena je inokulacijom deseterostruke maksimalne doze (106.0 TCID50) u SPF piliće oralnim i okulonazalnim putem te promatranjem pilića tijekom 21 dana od inokulacije cjepiva. Test diseminacije soja VMG91 po organima proveden je inokulacijom 105.0 TCID50 jedinica virusa SPF pilićima na 8. dan starosti. Diseminacija je praćena analizom uzoraka orga-na PCR-om (Zorman Rojs i sur., 2008.) petog dana od inokulacije. Horizontalna transmisija virusa sa zaražene na naivnu je-dinku praćena je miješanjem naivnih SPF pilića s inficira-nim jedinkama istoga jata te analizom prisustva IBD virusa PCR-om nakon 5 dana u brisevima kloake i orofarinksa. Genetska stabilnost cjepnog soja provjeravana je pasažira-njem virusa kroz skupine 8-dnevnih SPF pilića. Prema Ph. Eur. protokolu (Ph.Eur., 2013.), prva skupina inokulirana je sojem VMG91, dok je svaka sljedeća skupina primila homo-genizat burzi izuzetih od pilića prethodne skupine. Postupak je ponovljen 5 puta, a u svakom koraku prisutnost virusa potvrđivana je PCR-om. U drugom dijelu testa uspoređeni su klinički znakovi i ošte-ćenja burzi kod SPF pilića inokuliranih bilo matičnim sojem VMG91 bilo pet puta pasažiranim VMG91 sojem. Oštećenja burzi praćena su farmakopejskom metodom 7., 14., 21. i 28. Dana od inokulacije. Učinkovitost cjepiva s VMG91 sojem u SPF pilićima testi-rana je u nekoliko pokusa. Testiranjem je najprije doka-zivano vrijeme početka aktivne imunosti (engl. Onset of Immunity, OOI) te potom trajanje imunosti nakon cijepljenja (engl. Duration of Immunity, DOI). Cijepljenje je provedeno apliciranjem minimalne preporučene doze cjepiva (104.0 TCID50), a učinkovitost je potvrđivana izazivačkom infekci-jom virulentnim sojem DV86 te praćenjem pobola/smrtnosti i analizom oštećenja burze uzrokovanog izazivačkim viru-som. Početak imunosti testiran je 14. dana, a trajanje imu-nosti 28. dana nakon cijepljenja. Učinkovitost na komercijalnim životinjama pozitivnim na antitijela protiv IBD virusa potvrđena je inokulacijom minimalne preporučene doze cjepiva (104.0 TCID50) i to na optimalni dan cijepljenja kako je predviđeno deventerskom formulom (De Wit, 2001.). Ciljani titar proboja bio je 125 ELISA jedinica (prema standardnom komercijalnom testu Idexx), a pilići su cijepljeni 38. dana (test početka imunosti) odnosno 13. dana starosti (test trajanja imunosti). Terenskim studijama na brojlerima odnosno nesilicama provjeravana je sigurnost i učinkovitost soja VMG91 u

PROVJERA SIGURNOSTI I UČINKOVITOSTI CJEPIVA PROTIV ZARAZNE BOLESTI BURZE PREMA ZAHTJEVIMA EUROPSKE FARMAKOPEJE
115
proizvodnim uvjetima. U studiji na brojlerima (linija Hybrid Ross 308) jedno jato primilo je cjepivo s VMG91 sojem, a drugo jato komparatorsko cjepivo bazirano na sličnom inter-medijarnom soju. Dan cijepljenja određen je primjenom deventerske formule, a pilići su cjepivo primili na 18. dan starosti. Uz uobičajene proizvodne parametre (težina, faktor konverzije, uginuća, klinički znakovi) praćena je i sero-konverzija, tj. titar prema IBDV-u. U studiji na komercijalnim nesilicama (linija Lohmann brown) pokusno jato cijepljeno je sojem VMG91 dvaput, 24. i 56. dana. Optimalni dan prvog cijepljenja procijenjen je deventerskom formulom. Kontrolno jato primilo je kompa-ratorsko cjepivo u sličnim uvjetima. Praćeni su smrtnost, tje-lesna težina i proizvodni parametri. Planiranje i provođenje in vivo pokusa bilo je u skladu s re-gulativom Republike Hrvatske i EU o postupanju s ekspe-rimentalnim životinjama.
Statistička analiza
Razlike u težini pilića i ELISA titru protutijela analizirane se testom ANOVA. Razlike u pobolu i smrtnosti analizirane su testom proporcija. Razlike u oštećenjima burzi analizirane su Kruskal-Wallisovim testom.
Rezultati i rasprava
Molekularna karakterizacija soja IBDV VMG91
Sekvence hipervarijabilnih regija VP2 gena cjepnog soja VMG91 i uzorka in vivo pasažiranog cjepnog soja VMG91p u području 599-1189 nukleotida VP2 gena uspoređene su sa sekvencama sličnih cjepnih sojeva te relevantnih vrlo viru-lentnih sojeva. Pokazana je vrlo visoka podudarnost s drugim intermedijarnim cjepnim sojevima (100% prema soju D78, 98% prema Cu-1 soju), dok je sličnost s virulent-nim terenskim izolatima oko 94% (sojevi DV86 i UK661). U uzorku in vivo pasažiranog materijala VMG91p primi-jećena je jedna supstitucija na poziciji H253Q. Ova točkasta mutacija već je ranije opisana kod intermedijarnih cjepnih sojeva pri terenskoj primjeni (Tamás, 2009.; Qi i sur., 2009.). Smatra se da ova promjena na poziciji aa253 sama nije dovoljna da značajnije promijeni stupanj virulentnosti cjepnog IBDV-a, već je potrebna još najmanje jedna pro-mjena i to na poziciji aa284 (Qi i sur., 2009.; Olesen i sur., 2018.).
Stabilitet cjepiva u otopini
Kako bi se osiguralo da jedinke prime dozu cjepiva koja je unutar granica predviđenih specifikacijom, potrebno je provjeriti stabilitet cjepiva u suspenziji nakon rekonstitucije. Pokazano je da je titar cjepiva u otopini tijekom pro-matranog vremena od 3 sata izrazito stabilan. Tipična serija proizvoda nakon otapanja cjepiva imala je titar od 104,3TCID50/dozi. Nakon 2 sata u otopini pri sobnoj temperaturi titar pada na 104,2TCID50/dozi te ostaje stabilan do najmanje 3 sata od rekonstitucije cjepiva. Dakle, pro-sječan pad titra iznosi svega 0,033 log10 TCID50/dozi/h.
Većina virusa su relativno nestabilni u okolišu, što je i razlog zašto se živa cjepiva čuvaju u liofiliziranom obliku ili smrznuta. Nakon što otapanjem ili odmrzavanjem cjepivo ponovno dođe u tekući oblik, virus može biti izložen vanjskim negativnim čimbenicima kao što su temperatura i kvaliteta vode, osobito prisutnost klora i dezinficijensa. S obzirom na to da je za učinkovitost živih cjepiva od presudne važnosti količina živih virusnih čestica koju prima jedinka koja se cijepi, potrebno je pokazati stabilnost virusa u stvarnim uvjetima primjene, i to prema Ph. Eur. Propi-sanom protokolu (Ph.Eur., 2013.). Uobičajena je preporuka proizvođača da se cjepivo treba primijeniti u roku od dva sata od rekonstitucije. Kako bi se skratilo vrijeme aplikacije, ali i smanjili troškovi cijepljenja, masovna aplikacija cjepiva putem vode za piće ili raspršivanjem uvijek je bolji izbor nego individualna primjena. S druge strane, masovna apli-kacija može rezultirati nejednolikom primjenom, jer ptice neizbježno primaju nejednake doze cjepiva.
In vivo studije sigurnosti
U studiji sigurnosti primjene cjepiva na SPF pilićima nije bilo uginuća niti kliničkih znakova zarazne bolesti burze tijekom razdoblja od 21 dana od inokulacije, čime se potvr-đuje sigurnost primjene prema kriterijima Ph. Eur. Testom diseminacije soja VMG91 po organima SPF pilića inokuliranih na 8. dan starosti pokazana je prisutnost virusa u više tkiva. Virus je potvrđen u timusu, slezeni, bubregu, burzi, cekalnim tonzilama i želucu. Prisutnost virusa u razli-čitim organima potvrđuje poznatu činjenicu da je nisko patogene viruse IBDV-a moguće kultivirati ne samo u limfoidnom tkivu, nego i in vitro na različitim supstratima, čak i na staničnim kulturama koje nisu ptičjeg podrijetla (Tamás, 2009.). Patološkim pregledom nisu uočene nikakve promjene u morfologiji zahvaćenih organa, čime je poka-zana niska virulentnost soja. Testom horizontalne transmisije virusa pokazano je da se virus VMG91 može prenositi najmanje 3 puta s inficiranih na naivne jedinke. Ovo je očekivan rezultat za IBD virus, jer je poznato da je izlučivanje kroz kloaku najvažniji i učinkovit put širenja zaraze među jedinkama jata (Eter-radossi i Saif, 2008.). Virus izlučen u fecesu naivna jedinka može unijeti oralno ili udisanjem, što objašnjava prisutnost virusa u brisevima orofarinksa. Sposobnost da se širi hori-zontalno povećava učinkovitost cjepiva, jer omogućava da i jedinke koje nisu primile virus tijekom cijepljenja naknadno budu procijepljene izlučenim virusom. Međutim, dobra praksa nalaže da se osigura čim je više moguća procijep-ljenost jata primarnim cijepljenjem kako bi se čim prije stvorila imunost stada te smanjio na minimum broj in vivo pasaža virusa, koji može voditi ka neželjenom povratu virulencije cjepnog virusa (Olesen i sur. 2018). U testu genetske stabilnosti cjepnog soja virus je uspješno pasažiran 5 puta kroz SPF piliće. Nisu primijećeni klinički znakovi koji bi ukazivali na povrat virulencije cjepnog soja. U drugom dijelu testa, gdje su uspoređivani klinički znakovi i oštećenja burzi kod SPF pilića inokuliranih bilo matičnim

PROVJERA SIGURNOSTI I UČINKOVITOSTI CJEPIVA PROTIV ZARAZNE BOLESTI BURZE PREMA ZAHTJEVIMA EUROPSKE FARMAKOPEJE
116
sojem VMG91 bilo pet puta pasažiranim VMG91 sojem, niti kod jedne skupine nisu uočeni klinički znakovi zarazne bolesti burze. Oštećenja burzi bila su veća kod skupine koja je primila pasažirani materijal, ali je uočeno da tijekom 28 dana od inokulacije virusa dolazi do potpune repopulacije limfocita i strukture burze. Relativno veća oštećenja burze uzrokovana pasažiranim virusom možda se mogu djelomice objasniti točkastom mutacijom H253Q, promjena koja je ranije opisana pri terenskoj primjeni intermedijarnih sojeva (Olesen i sur., 2018.). Kako literaturni podatci upućuju na to da je ipak potrebno više od jedne mutacije za značajniju promjenu tropizma IBDV-a (Qi i sur., 2009.), vjerojatnije je da je prividno veća virulentnost pasažiranog virusa uzrokovana aplikacijom virusa u količini većoj od potrebne. Naime, promjena H253Q uzrokuje promjenu preferencije virusa za tip stanica u kojima preferencijalno raste, i to s ne-limfoidnih na limfoidne stanice (Tamás, 2009.). Kako u vrijeme provođenja testa ovo nije bilo poznato, titar pasa-žiranog materijala procjenjivan je temeljem titracije na neodgovarajućem tipu stanica te je on značajno podcijenjen. Prema procjeni, ovo je dovelo do inokulacije značajno veće količine virusa od potrebne, što je moglo rezultirati i većim oštećenjem burze nego kod matičnog soja VMG91. Tijekom atenuacije divljeg virusa, pasažiranjem u staničnim kulturama ili embrioniranim jajima stvara se evolucijski pri-tisak kojim se umjetno favoriziraju manje patogene podvrste ishodnog soja. Suprotno se događa pri terenskoj primjeni tako atenuiranog cjepiva. Replikacijom in vivo dolazi do spontanih i nasumičnih mutacija koje stvaraju genetski pri-tisak i favoriziraju podvarijante koje su virulentnije i koje se brže repliciraju (Olesen i sur., 2018.). Učinak može biti i pojačan ako su jedinke već procijepljene cjepivom protiv tog ili antigenski sličnog patogena. Mutacije koje dovode do imunoevazije (izbjegavanja domaćinove aktivne imunosti protiv sličnog patogena) bit će favorizirane, jer će takvi mutanti imati veće izglede za propagaciju u domaćinu (Jackwood i Sommer-Wagner, 2011.). Vjerojatnost da će doći do povrata virulencije atenuiranog cjepnog soja u terenskim uvjetima raste s brojem generacija kroz koje se virus može umnožiti. Stoga se pravilnim cijep-ljenjem nastoji istodobno imunizirati što veći dio jata kako bi broj naivnih jedinki podložnih sekundarnoj infekciji bio što manji te se time minimizirala vjerojatnost stvaranja neželjenih mutacija. U slučaju virusa zarazne bolesti burze ovo je naročito relevantno, jer je poznato da se horizontalni prijenos virusa može uspješno spriječiti istodobnim cijeplje-njem cijelog jata (Olesen i sur., 2018.). Razlog tome je taj što burza koja je već inficirana IBDV-om postaje manje prijemčiva za re-infekciju jednakim ili drugim IBDV-om. Posljedica je to snažnog anti-virusnog odgovora na inici-jalnu infekciju, koji se očituje u stvaranju proupalnih ci-tokina, čime se učinkovito sprječava vjerojatnost ponovne infekcije burze istim ili sličnim patogenom (Yasmin i sur., 2016.). Ovo je potvrđeno i našim rezultatima, gdje je poka-zano da se u burzama cijepljenih, a potom divljim virusom izazvanih pilića nalazi isključivo cjepni virus koji je prvi populirao burzu, a u kontrolnih (ne cijepljenih) pilića nalazi se, očekivano, izazivački IBD virus.
S obzirom na to da IBD virus preferencijalno napada Fabricijevu burzu koja je važan imunosni organ, infekcija IBDV-om može imati i imunosupresivne učinke. Stoga je regulatorni zahtjev da se živa atenuirana cjepiva testiraju testom imunosupresije koji mora pokazati da cijepljenje IBD cjepivom neće imati imunosupresivni učinak na cijepljenje protiv ND (Ph.Eur. 2013). Farmakopejskim testom poka-zano je da odgovor na cjepivo protiv ND u pilićima pret-hodno imuniziranima visokom dozom soja VMG91 nije statistički značajno različit od odgovora kontrolne skupine koja je primila samo cjepivo protiv ND; obje skupine bile su 100% zaštićene od izazivačke infekcije virulentnim sojem ND-a. Treba naglasiti da je test dizajniran tako da se cjepivo protiv ND aplicira 7. dan nakon cijepljenja IBDV cjepivom, dakle, u razdoblju u kojem se očekuje najveće oštećenje burze uzrokovano IBD virusom te se time simulira poten-cijalno najgori mogući scenarij koji se može očekivati u terenskoj primjeni. Intermedijarni cjepni sojevi aplicirani u dozama višestruko većim od preporučenih mogu izazvati prolaznu depleciju limfocita u burzi. Imunosupresivnost korelira s virulent-nošću cjepnog soja te se očekuje da će cjepiva s „vrućim“ cjepnim sojevima uzrokovati veći imunosupresivni učinak od intermedijarnih sojeva, ali i vjerojatno pružiti bolju zaštitu protiv IBD-a (Eterradossi i Saif, 2008.).
Studije učinkovitosti cjepiva
Učinkovitost cjepiva provjerava se na ciljnim vrstama živo-tinja, i to najprije u laboratorijskim pokusima, a potom i u terenskim studijama. Kod cjepiva za perad učinkovitost je potrebno najprije pokazati na SPF životinjama. S obzirom na to da prisustvo materinskih protutijela (engl. maternal derived antibodies, MDA) nastalih cijepljenjem roditelja protiv ciljnih bolesti može interferirati s cijepljenjem poto-maka, učinkovitost je potrebno potvrditi i na komercijalnim jedinkama. Farmakopeja nalaže da se učinkovitost na SPF pilićima potvrdi apliciranjem minimalne preporučene doze cjepiva te potom izazivačkom infekcijom relevantnim divljim virusom (Ph.Eur. 2013). Najvažniji rezultati laboratorijskih testiranja početka i tra-janja imunosti sažeti su u tablici 1. Rezultati pokazuju da cjepivo učinkovito štiti piliće 14. dan od cijepljenja te da aktivna imunost traje najmanje do 28. dana od cijepljenja. Aktivna imunost vjerojatno se počinje razvijati i značajno ranije, no novije farmakopeje kao kriterij imunosti propisuju zaštitu protiv izazivačke infekcije, što nije realno očekivati prije 14. dana od cijepljenja. Slično, trajanje aktivne imu-nosti vjerojatno je i duže od testiranog razdoblja, ali kako je pokus ciljao na zaštitu brojlera, odabran je vremenski raspon relevantan za prosječan proizvodni ciklus brojlera. U testiranjima učinkovitosti cjepiva na komercijalnim broj-lerima cjepivo je uspješno zaštitilo cijepljene skupine protiv izazivačke infekcije u oba testa (test početka i test trajanja imunosti), usprkos činjenici da je prosječni titar protutijela u oba pokusa bio značajno veći od ciljanog (236 i 608 ELISA jedinica), kako pokazuju rezultati sažeti u tablici 2.

PROVJERA SIGURNOSTI I UČINKOVITOSTI CJEPIVA PROTIV ZARAZNE BOLESTI BURZE PREMA ZAHTJEVIMA EUROPSKE FARMAKOPEJE
117
Tablica 1. Učinkovitost cjepnog soja VMG91 na SPF pilićima cijepljenima minimalnom dozom te izazvanih vrlo virulentnim sojem IBDV 14. odnosno 28. dana od cijepljenja
Table 1. Efficacy of the vaccine strain VMG91 on SPF chickens vaccinated with a minimum recommended dose and challenged using very virulent strain of IBDV on the 14th and 28th day post vaccination
Grupa* / Group*
Prosječan ELISA titar prije izazivačke infekcije
Average ELISA titer prior to challenge infection
Prosječno oštećenje burze nakon izazivačke
infekcije** / Average damage to bursa after challenge infection**
Pobol** Morbidity**
Smrtnost** Mortality**
Test početka imunosti (14 dana od cijepljenja) Test of immunity initiation (14 days of vaccination)
Cijepljeni / Vaccinated 2347A 0.90 0%A 0%A
Kontrolni (necijepljeni) Control (nonvaccinated) 41B n.d.*** 90%B 100%B
Test trajanja imunosti (28 dana od cijepljenja) Test of immunity duration (28 days of vaccination)
Cijepljeni / Vaccinated 5258A 1,2 A 0%A 0%A
Kontrolni (necijepljeni) Control (nonvaccinated) 66B 5,0 B 90%B 100%B
*20 cijepljenih pilića, 10 kontrolnih pilića **Rezultati s različitim slovom u eksponentu označuju statistički značajno različite rezultate unutar pokusa (p>0,05) ***Analiza burzi nije rađena na uginulim niti na pilićima s teškim znakovima zarazne bolesti burze *20 vaccinated broilers, 10 control broilers **Figures marked with different letters in superscript denote statistically significantly different results within the test (p>0.05) ***Analysis of bursae was not performed on dead broilers and those with severe signs of infectious bursal disease Tablica 2. Učinkovitost cjepnog soja VMG91 na komercijalnim pilićima cijepljenim minimalnom dozom te izazvanih vrlo
virulentnim sojem IBD 14. odnosno 28. dana od cijepljenja. Dani cijepljenja određeni su prema deventerskoj formuli (De Wit 2001) koristeći parametre za intermedijerna cjepiva
Table 2. Efficacy of the vaccine strain VMG91 on commercial chickens vaccinated with a minimum recommended dose and challenged using very virulent strain of IBDV on the 14th and 28th day post vaccination. Vaccination days were determined using Deventer formula (De Wit 2001) using parameters characteristic for intermediate vaccines
Skupina* / Group*
Prosječan ELISA titar / Average ELISA titer Prosječno oštećenje burze nakon izazivačke
infekcije** / Mean bursal damage following infection
challenge**
Nakon valjenja Post hatching
Na dan cijepljenja**
On the day of vaccination**
Na dan izazivačke infekcije**
On the day of infection challenge**
Test početka imunosti (14 dana od cijepljenja) Test of immunity initiation (14 days of vaccination)
Cijepljeni (starost: 38 dana) / Vaccinated (age: 38 days)
4353 236A 6784 1.8A
Kontrolni (necijepljeni) / Control (nonvaccinated) 349A n.d. 4.8B
Test trajanja imunosti (28 dana od cijepljenja) Test of immunity duration (28 days of vaccination)
Cijepljeni (starost: 13 dana) Vaccinated (age: 13 days)
1383 608 2628 A 2.0A
Kontrolni (necijepljeni) Control (nonvaccinated) 147 B 4.9B
*20 cijepljenih pilića, 10 kontrolnih pilića **Rezultati s različitim slovom u eksponentu označuju statistički značajno različite rezultate unutar pokusa (p>0.05) *20 vaccinated broilers, 10 control broilers **Figures marked with different letters in superscript denote statistically significantly different results within the test (p>0.05) Serokonverzija je bila značajna kod cijepljenih pilića koji su na dan izazivačke infekcije, dakle 14. odnosno 28. dana od cijepljenja, imali prosječni titar od 6784 odnosno 2628 ELISA jedinica. Oštećenja burze bila su umjerena/niska kod
cijepljenih životinja, dok je kod kontrolnih došlo do gotovo potpune deplecije limfocita i gubitka folikularne strukture. Analizom uzoraka burzi pokazano je da se u burzama cijepljenih pilića nakon izazivačke infekcije nalazi samo

PROVJERA SIGURNOSTI I UČINKOVITOSTI CJEPIVA PROTIV ZARAZNE BOLESTI BURZE PREMA ZAHTJEVIMA EUROPSKE FARMAKOPEJE
118
cjepni virus, što pokazuje da izazivački IBDV nije probio zaštitu i populirao burzu. Očekivano, kod svih kontrolnih pilića potvrđen je izazivački virus u uzorcima burzi. Ovaj nalaz jasno pokazuje koliko je važno jednoliko i istodobno imunizirati cijelo jato kako bi se spriječilo širenje virusa kroz nezaštićene jedinke.
Terenske studije
U terenskoj studiji na brojlerima (linija Hybrid Ross 308) pilići su na 18. dan starosti primili po jednu uobičajenu dozu cjepiva (cjepivo s VMG91 sojem ili komparatorsko cjepivo) putem vode za piće. Usprkos činjenici da je na dan ci-jepljenja (18. dan) prosječni titar anti-IBDV protutijela bio 615 ELISA jedinica, serokonverzija je bila dobra te je na 40. dan ciklusa izmjeren prosječni titar 3407 ELISA jedinica. Proizvodni parametri bili su u okviru uobičajenih vrijed-nosti: klanje je provedeno na 37. dan s prosječnom težinom od 2.28 kg, faktorom konverzije 1.81 te proizvodnim indeksom od 325. Nije bilo statistički značajne razlike iz-među skupina niti je uočeno prisustvo divljeg IBD virusa. Jata komercijalnih nesilica (linija Lohmann brown) cijep-ljena su dvaput, jer je takva praksa uobičajena na pokusnoj farmi. Prosječni titar materinskih protutijela na dan prvog cijepljenja bio je 413 ELISA jedinica. Serokonverzija na cje-pivo bila je vrlo dobra: tri tjedna nakon prve doze prosječni titar protutijela na IBDV bio je 9138 ELISA jedinica, a četiri tjedna nakon druge 10512 ELISA jedinica. Kontrolno jato koje je primilo komparatorsko cjepivo pokazalo je rezultate koji nisu bili statistički značajno različiti od pokusne sku-pine. Smrtnost, tjelesna težina i proizvodni parametri bili su u skladu s uobičajenim vrijednostima za pokusnu farmu i nisu se statistički značajno razlikovali između skupina. Za nesilice nije neuobičajeno aplicirati živo IBDV cjepivo dvaput. Budući da bi eventualno odgađanje da titar mate-rinskih protutijela u cijelom jatu padne ispod titra proboja cjepiva nepotrebno izložilo cijelo jato riziku od proboja divljeg IBDV-a, dobra praksa je uvesti do-cjepljivanje jata nekoliko tjedana nakon prve doze. Drugom dozom će se procijepiti jedinke jata koje nisu optimalno primile prvu dozu, bilo zbog previsokog titra protutijela, bilo zbog drugih razloga. Na taj način druga doza doprinosi imunosti jata i uniformnosti titra zaštitnih protutijela (OIE 2016).
Zaključak
Prikazan je dio rezultata in vitro i in vivo karakterizacije cjepnog soja VMG91 provedenog prema zahtjevima Ph. Eur. i EMA-e za registraciju novih cjepiva. Sigurnost i učinkovitost cjepiva potvrđena je u laboratorijskim i teren-skim uvjetima na SPF pilićima kao i na komercijalnim brojlerima i nesilicama. Test genetske stabilnosti ukazuje na potrebu da se cijepljenje provede kvalitetno i prema upu-tama struke kako bi se postigla visoka istodobna proci-jepljenost jata i spriječio horizontalni prijenos virusa.
Literatura
Basic Local Alignment Search Tool. Accessed 7.1.2019. https://blast.ncbi.nlm.nih.gov/Blast.cgi.
BERG, T. P. V. D. (2000). Acute infectious bursal disease in poultry: a review, Avian Pathology, 29, 175-94.
CAO, Y., YEUNG, W., LAW, M., BI, Y., LEUNG, F., and LIM, B. (1998). Molecular characterization of seven Chinese isolates of infectious bursal disease virus: classical, very virulent, and variant strains, Avian diseases, 42, 340-51.
DE WIT, J. (2001): Gumboro disease: estimation of optimal time of vaccination by the Deventer formula. In: Proceedings of the 3rd meeting of working group, Vol. 3. 21-28.
EMA (1999): European Medicines Agency. Committee for medicinal products for veterinary use. Note for guidance: Duration of protection achieved by veterinary vaccines. EMEA/CVMP/682/99.
EMEA (2001): The European agency for the evaluation of medicinal products. Committee for veterinary medicinal products. Note for guidance: Field trials with veterinary vaccines. EMEA/CVMP/852/99.
EMEA (2005): European Medicines Agency. Veterinary Medicines and Inspections. Guideline on Target Animal Safety for Veterinary Live and Inactived Vaccines. EMEA/CVMP/ /VICH/359665/2005.
EMEA (2000): The European Agency for the Evaluation of Medicinal Products. Veterinary Medicines and Information Technology Unit. Guideline on Good Clinical Practices. CVMP/VICH/595/98. (2000).
ETERRADOSSI, N., and SAIF, Y. (2008). Infectious bursal disease, Diseases of poultry, 12th ed.: 185-208.
JACKWOOD, D. J. (2017). Advances in vaccine research against economically important viral diseases of food animals: Infectious bursal disease virus, Vet. Microbiol., 206. 121-25.
JACKWOOD, D. J., and SOMMER-WAGNER, S. E. (2011). Amino acids contributing to antigenic drift in the infectious bursal disease Birnavirus (IBDV), Virology, 409: 33-37.
OIE (2016). OIE Terrestrial Manual. 2.3.12. Infectious Bursal Disease (Gumboro disease), 655.
OLESEN, L., DIJKMAN, R., KOOPMAN, R., VAN LEEUWEN, R., GARDIN, Y., DWARS, R., DE BRUIJN, N., BOELM, G., ELATTRACHE, J., and DE WIT, J. (2018). Field and labo-ratory findings following the large scale use of intermediate type infectious bursal disease vaccines in Denmark, Avian Pathology, 47, 595-606.
PH. EUR. (2013): Council of Europe. European Pharmacopoeia. 7th ed. Avian infectious bursal disease vaccine (live). 04/2013:0587.
PH. EUR. (2013.): Council of Europe. European Pharmacopoeia. 7th ed. 5.2.6. Evaluation of safety of veterinary vaccines and immunosera.
PH. EUR. (2017): Council of Europe. European Pharmacopoeia. 9th ed. Vaccines for veterinary use.
QI, X., GAO, H., GAO, Y., QIN, L., WANG, Y., GAO, L., and WANG, X. (2009). Naturally occurring mutations at residues 253 and 284 in VP2 contribute to the cell tropism and virulence of very virulent infectious bursal disease virus, Antiviral Res., 84, 225-33.
TAMÁS, M. 2009. 'Genetic diversity of infectious bursal disease virus strains and the molecular basis for change in virulence',

PROVJERA SIGURNOSTI I UČINKOVITOSTI CJEPIVA PROTIV ZARAZNE BOLESTI BURZE PREMA ZAHTJEVIMA EUROPSKE FARMAKOPEJE
119
Doctoral thesis, Veterinary Medical Research Institute of the Hungarian Academy of Sciences.
YASMIN, A., YEAP, S., HAIR-BEJO, M., and OMAR, A. (2016). Characterization of chicken splenic-derived dendritic cells following vaccine and very virulent strains of infectious bursal disease virus infection, Avian diseases, 60, 739-51.
ZORMAN ROJS, O., KRAPEŽ, U., SLAVEC, B., MANKOČ, S., JURIŠIČ-CIZERL, R., and BARLIČ-MAGANJA, D. (2008). Molecular characterisation of infectious bursal disease viruses isolated in recent acute outbreaks in Solvenia, Acta Veterinaria Hungarica, 56, 255-64.
SAFETY AND EFFICACY ASSESSMENT OF THE VACCINE AGAINST INFECTIOUS BURSAL DISEASE ACCORDING TO THE EUROPEAN PHARMACOPOEIA REQUIREMENTS
Summary
Vaccination of chickens using live attenuated vaccines is the method of choice for the prevention and control of the infectious bursal disease. New vaccines intended for use in the European Union have to comply with the requirements of the European Pharmacopoeia (Ph. Eur.), which prescribes in detail in vitro and in vivo control of safety and efficacy of the master seeds, as well as of each individual production batch. Here we present results of in vitro characterization of the master seed and results of in vivo assessment of the safety and efficacy of the live attenuated vaccine against infectious bursal disease based on intermediate VMG91 strain. General safety was evaluated using SPF layers according to the Ph. Eur. protocol. A hundredfold minimum recommended vaccine dose (106.0 TCID50) did not cause any clinical signs in SPF chickens during the period of 21 days after vaccination. Efficacy of the vaccine against Newcastle disease in chickens which previously received VMG91-based vaccine showed that vaccine was not immunosuppressive according to the Ph. Eur. criteria: protection against challenge with virulent Newcastle disease virus was not significantly affected by vaccination with VMG91. The test of genetic stability revealed a single point mutation at H253Q on the VP2 gene after in vivo passaging, a phenomenon previously described for intermediate vaccine strains. Laboratory efficacy studies on SPF and commercial birds immunized with a minimum recommended dose demonstrated 100% efficacy in the protection against challenge. Results of the field trials on layers and broilers confirmed the safety and efficacy under the production conditions. Overall, study results demonstrated the suitability of the vaccine strain to be used for protection against infectious bursal disease, according to the current requirements of the EU. Keywords: infectious bursal disease; live attenuated vaccines; European Pharmacopoeia

PERADARSKI DANI 2019. 120
DEVELOPMENT OF LIVE ATTENUATED VACCINE AGAINST INFECTIOUS BRONCHITIS VIRUS ACCORDING TO CURRENT EUROPEAN REGULATIVE
Lana Ljuma Skupnjak1, Gert Jan Boelm2, Anto Vrdoljak1
1Genera d.d. (Genera d.d. part of Dechra Pharmaceuticals PLC Group), Rakov Potok, Croatia 2GD Animal Health, Deventer, The Netherlands
Summary
Infectious bronchitis of fowls is a highly contagious disease of respiratory and urogenital tract of birds. Causative agent of the disease is infectious bronchitis virus (IBV). Due to significant losses the disease causes in poultry production worldwide, strict biosecurity and prophylactic measures are undertaken. There are many different live attenuated and inactivated vaccines against IBV present on the market nowadays, based on several different vaccine strains. The most widely used live attenuated vaccine against IBV contains H120 strain, which belongs to the Massachusetts serotype and is present on the market for more than 50 years. European Pharmacopoeia together with other relevant EU regulative strictly prescribes safety and efficacy tests and criteria which a vaccine needs to comply to be granted a marketing authorization across the EU. We have developed a live attenuated vaccine against IBV based on H120 vaccine strain. Safety tests were performed in SPF chickens where it was shown that even after 6 passages in chickens, our vaccine strain did not revert to virulence. Furthermore, in the dissemination and spreading test IB H120 was able to spread at least 3 times between groups of IB naïve SPF chickens without inducing morbidity and mortality. H120 was detected in trachea, spleen, kidney, lung, cecal tonsil and jejunum without causing pathological changes in these organs. H120 vaccine is most frequently administered to chicks during their first week of life. At this age, the level of maternally derived antibodies (MDA) is still very high, which may hamper development of active immunity. In our research, the efficacy of IB H120 vaccine was assessed in broiler chickens with and without MDA to IB virus. Chicks were vaccinated by spray, eye-nose-drop, or orally, or were left unvaccinated. At three different time points after vaccination, protection against M41 challenge was determined. The results showed the onset of immunity to start already 10 days after vaccination. Judged by the protection level achieved, there was no influence of MDA on the vaccination by any of the administration methods tested. Safety and efficacy of the vaccine was confirmed also in field conditions, on a broiler farm in France and a layer farm in Slovenia. Production results were comparable to those achieved by already marketed IBV vaccine with the same indication, showing that our IB H120 vaccine was safe and efficacious also in field conditions.
Keywords: infectious bronchitis virus, vaccination, maternally derived antibodies
Introduction Infectious bronchitis is a highly contagious disease of poultry which causes remarkable economic losses all over the world (Toro, van Santen, and Jackwood, 2012). The causative agent of the disease is infectious bronchitis virus (IBV), a single-stranded positive-sense RNA virus, belonging to the order Nidovirales, family Coronaviridae genus Coronavirus (Franzo et al., 2017). IBV causes mainly upper respiratory tract disease, with a morbidity of typically 100% and mortality up to 50% in the case of nephro-
pathogenic strains or when accompanied by opportunistic infections. In addition, reproductive tract of layers and breeders can be affected causing decreased egg production and altered egg quality (Jackwood, 2012). Both live attenuated and inactivated vaccines are used for vaccination of chickens against IBV. The most widely used IBV vaccine strain is H120, which has been used all over the world for more than 50 years (Bijlenga et al., 2004). When developing a vaccine, strict precautions must be taken to ensure that the vaccine strain does not revert to virulence after in vivo passaging in susceptible birds. European
Lana Ljuma Skupnjak, mag.ing. biotechn., Genera Inc., Svetonedeljska cesta 2, Kalinovica. HR-10436 Rakov Potok, Croatia; Tel. +385-1-3388636; Email: [email protected]

DEVELOPMENT OF LIVE ATTENUATED VACCINE AGAINST INFECTIOUS BRONCHITIS VIRUS ACCORDING TO CURRENT EUROPEAN REGULATIVE
121
Pharmacopoeia (Ph. Eur.) together with Directive 2001/82/EC describes the tests and defines criteria that a vaccine has to comply to be granted marketing authorization in the EU. Furthermore, safety of the vaccine should be demonstrated for all the administration routes proposed. Dissemination of the vaccine strain in the body of vaccinated animal and spread between vaccinated and unvaccinated group of chicks have to be investigated. Depending on the properties of the vaccine strain, special precaution measures should be implemented into the Summary of Product Characteristics (SPC). Since vaccination of 1-day old chicks with H120 vaccine is widely performed, the vaccine virus may be neutralized by circulating maternally derived antibodies (MDA), which are still at a very high level at this time point. While some authors report a negative effect of high levels of MDA on vaccine strain (Pattani et al., 2012; Mondal and Naqi, 2001), others did not detect lower efficacy of a vaccine when given to day-old chicks with high MDA levels (Darbyshire and Peters, 1985; Davelaar and Kouwenhoven, 1977). In this work, we have assessed the safety and efficacy of IB H120 vaccine according to requirements of Ph. Eur. and Directive 2001/82/EC both in laboratory and field con-ditions. Furthermore, the onset and duration of immunity in broilers with MDA were determined and the influence of MDA on development of active immunity was evaluated.
Materials and Methods
Viruses The infectious bronchitis virus H120 vaccine used in this study (Avishield IB H120, Genera Inc., part of Dechra Pharmaceuticals PLC Group) is produced in compliance with European Pharmacopoeia and Directive 2004/28/EC concerning the quality, safety and efficacy of live poultry vaccines. For the efficacy testing, vaccine virus was reconstituted in an appropriate volume of diluent, depending on the inoculation route to provide 3.5 log10 50% embryo infectious dose (EID50) per bird, which is the minimal dose recommended for vaccination as required by Ph. Eur. monograph 04/2013:0442 and the specifications of the manufacturer. Challenge of chickens in vaccination challenge studies was conducted using the virulent IBV strain M41. For the safety testing, vaccine virus was reconstituted in an appropriate volume of diluent, to provide either maximum dose (4.5 log10 EID50 per bird) or 10-fold maximum dose (5.5 log10 EID50 per bird), depending on the requirement by Ph. Eur. monograph 04/2013:0442.
Ciliary activity The ciliary activity of tracheal explants from all challenged chickens was examined 5 days after challenge. Shortly after euthanizing the chickens, the trachea of each chicken was extirpated by removing the skin of the neck. Subsequently, the trachea was disconnected from the surrounding tissues, and cut just proximal of the larynx and just proximal of the
thorax and put in Minimum Essential Medium with 4-2-hydroxyethyl-1-piperazineethanesulfonic acid buffer (MEM-H medium of 37 °C). Transverse sections of 0.5-2 mm of the trachea were made using a scalpel. The transverse sections of the trachea were placed in MEM-H medium (37 °C) on a 20-well plate and examined under the microscope (40-100x). Ciliary activity of three sections of the upper part, four sections of the middle part and three sections of the lower part of the trachea were examined. All tracheal explants were examined within 2 hours after sampling. The activity of the cilia in each tracheal section were scored according to Ph. Eur. monograph 04/2013:0442, section 2-4-3 Immunogenicity, as follows: score 0: >50% of the tracheal section shows cilia movement, and score 1: <50% of the tracheal section shows cilia movement. A tracheal section was considered normal when at least 50% of the internal ring showed vigorous ciliary movement. A chicken was considered protected if at least 9 out of 10 tracheal rings showed normal ciliary activity. PCR detection in organs and swabs For the detection of IB H120 in collected samples and swabs, a real-time PCR procedure developed by Callison et al. (Callison et al., 2006) was used. Briefly, IBV specific primers (IBV-F: GCT-TTT-GAG-CCT-AGC-GTT and IBV-R: G-CCA-TGT-TGT-CAC-TGT-CTA-TTG) amplify a 143-bp fragment of the gene of the IBV 5’-UTR. The isolated RNA was translated into cDNA by the Reverse Transcriptase enzyme. The PCR protocol started with the Reverse Transcriptase (RT) incubation step of 30 min at 45 °C, followed by a heating step at 95 °C. The PCR cycle in this PCR method consisted of two steps. In the first step, the double-stranded DNA was denaturized at 95 °C and two single-stranded DNA molecules were created. In the second step, the primers were attached to single stranded DNA molecules at a temperature of 60 °C. At this temperature, a (Taqman) probe was also bound. The Taq-polymerase enzyme was responsible for the extension of the primers and new target molecules were formed. During the extension of the target DNA, the bound probes were removed and the fluorochrome was released into the solution. The fluorescence signal of the fluorochrome was measured.
Animal studies A set of laboratory and field studies required for registration of live IBV vaccine in EU countries was performed in animals to assess vaccine safety and efficacy. Studies were designed and conducted in accordance with guidelines outlined in relevant Ph. Eur. monographs and EMA chapters. Details on the design of each specific study may be found in the documents cited in Table 1, with specific details briefly described in the Results section where applicable. Animal experiments were conducted following national and European Union regulations regarding the use and pro-tection of animals used for scientific purposes.

DEVELOPMENT OF LIVE ATTENUATED VACCINE AGAINST INFECTIOUS BRONCHITIS VIRUS ACCORDING TO CURRENT EUROPEAN REGULATIVE
122
Table 1. Summary of in vivo studies performed using H120 vaccine strain Tablica 1. Sažetak in vivo studija provedenih sa cjepnim sojem H120
Study type / Vrsta studije Laboratory or field study
Laboratorijska ili terenska studija
Chicken breed Vrsta peradi
Design according to Dizajn studije u skladu s
Safety: dissemination and spreading / Neškodljivost: diseminacija i širenje
Laboratory / Laboratorijska
SPF layers / SPF nesilice
Ph. Eur. 04/2013:50206
Safety: increase in virulence / Neškodljivost: porast virulencije
Ph. Eur. 04/2013:0442 Ph. Eur. 04/2013:50206
Safety for respiratory tract and kidneys / Neškodljivost za respiratorni trakt i bubrege
Ph. Eur. 04/2013:0442
Safety for reproductive system / Neškodljivost za reproduktivni sustav
Ph. Eur. 04/2013:0442 Ph. Eur. 04/2013:50206 EMEA/CVMP/VICH/359665/2005
Efficacy in SPF chickens / Učinkovitost u SPF pilićima
Ph. Eur. 04/2013:0442 Ph. Eur. 04/2008:50207
Efficacy in MDA-positive chickens / Učinkovitost u MDA-pozitivnim pilićima
Commercial broilers / Komercijalni brojleri
Ph. Eur. 04/2013:0442 Ph. Eur. 04/2008:50207
Safety and efficacy in field conditions / Sigurnost i učinkovitost u terenskim uvjetima
Field / Terenska
Commercial broilers / Komercijalni brojleri EMEA/CVMP/852/99
Ph. Eur. 04/2008:50207 EMEA/CVMP/VICH/359665/2005 Commercial layers /
Komercijalne nesilice Results and Discussion
Dissemination and spreading In this study, dissemination of IBV strain H120 was assessed by inoculating 15 1-day-old SPF-chickens with 104.5 EID50 of H120 vaccine strain. Five days after inoculation, 5 chickens were euthanized to demonstrate the dissemination of vaccine virus within the body and 10 chickens were introduced to non-inoculated hatch mates in a clean isolator. Cloacal and oropharyngeal swabs were taken at 5 and 10 days post exposure to the vaccine virus to demonstrate spread of the vaccine virus from inoculated to non-inoculated chickens. This procedure was repeated 3 times by introducing infected chickens five days after contact with inoculated or infected group into a clean isolator containing non-inoculated hatch mates, resulting in four groups of animals in total (groups A, B, C, D; Tables 2 and 3). The IB H120 could be detected in oropharyngeal, tracheal and cloacal swabs 5 and 10 days after exposure to the vaccine virus in any group (Table 3). This shows that both respiratory and cloacal excretions are important transmis-sion routes of this vaccine virus. The uptake of IB H120 by subsequent IB naïve groups did not result in morbidity or mortality. Dissemination of IB H120 to the trachea, spleen, kidney, lung, cecal tonsil and jejunum occurred after
exposure to the vaccine virus, without inducing patholo-gical changes to these organs, however, in the last group IB H120 was not detected in the jejunum (Table 2). The presence of IBV in organs other than respiratory tract was not unexpected considering the fact that IBV replicates in many epithelial surfaces, which contributes to broad tissue tropism. It is well known that besides respi-ratory tissues (nose, trachea, lungs and air sacs) IBV replicates in kidney, oviduct, testes and many parts of alimentary tract (esophagus, proventriculus, duodendum, jejunum, cecal tonsils, bursa of Fabricius, rectum and cloaca (Cavanagh, 2007; Bande et al., 2016; Zhao et al., 2015). Based on our results, it can be concluded that our vaccine containing IB H120 is safe for use in chickens. Spreading between chickens by respiratory and cloacal excretion of vaccine virus ensures that chickens which were missed during vaccination will receive vaccine virus from chickens which took the vaccine, which supports the efficacy of this vaccine at flock level (Matthijs et al., 2008). It must be emphasized that even though horizontal spread of attenuated vaccine virus increases the probability of mutations and may lead to reversion to a more virulent state, we did not observe any indication of increase in virulence of our vaccine strain IB H120 after three passages in chicks.

DEVELOPMENT OF LIVE ATTENUATED VACCINE AGAINST INFECTIOUS BRONCHITIS VIRUS ACCORDING TO CURRENT EUROPEAN REGULATIVE
123
Table 2. Results of dissemination and spread test. Detection of IB H120 in different organs by PCR Tablica 2. Rezultati studije diseminacije i širenje virusa. Detekcija IB H120 u različitim organima pomoću PCR-a
Day of the study / Dan studije
Organ / Organ D5 D10 D15 D20 D25
Group A Skupina A
Group A Skupina A
Group B Skupina B
Group C Skupina C
Group D Skupina D
Days after exposure / Dani nakon izlaganja 5 10 10 10 10 Trachea / Traheja +* +/+** +/+ +/+ +/+ Spleen / Slezena + +/+ +/+ +/+ +/- Kidney / Bubreg + +/+ +/+ +/+ -/+ Lung / Pluća + +/+ +/+ +/+ +/+ Cecal tonsil / Cekalne tonzile + +/+ +/+ +/+ +/+ Jejunum / Jejunum + +/+ +/+ -/+ -/-
+ = IB H120 detected; - = IB H120 not detected; *1 pool of 5 samples; **2 pools of 5 samples + = IB H120 detektiran; - = IB H120 nije detektiran; *1 skupni uzorak iz 5 pojedinačnih uzoraka; **2 skupna uzorka od 5 pojedinačnih uzoraka Table 3. Results of dissemination and spread test. Detection of IB H120 in oropharyngeal, tracheal and cloacal swabs Tablica 3. Rezultati studije diseminacijei širenje virusa. Detekcija H120 u brisevima orofarinska, traheje i kloake
Swab / Bris D5 D10 D15 D20 D25
Group A Skupina A
Group A Skupina A
Group B Skupina B
Group B Skupina B
Group C Skupina C
Group C Skupina C
Group D Skupina D
Group D Skupina D
Days after exposure Dani nakon izlaganja 5 10 5 10 5 10 5 10
Oropharynx / Orofarinks +/+ +/+ +/+ Trachea / Traheja +/+ +/+ +/+ +/+ +/+ Cloaca / Kloaka +/+ +/+ +/+ +/+ +/+ +/+ +/+ +/+
+ = IB H120 detected; - = IB H120 not detected; + = IB H120 detektiran; - = IB H120 nije detektiran Increase in virulence In this study, IB H120 was passaged six times over 14-day-old SPF chickens without inducing clinical signs of IB. First group of chickens was inoculated with 104.5 EID50 of H120 vaccine strain by eye drop. Four days after inoculation, the chickens were euthanized and a suspension of trachea was made. The presence of IBV was determined by PCR. For subsequent groups, 0.05 mL of the suspension of trachea was administered to five 14-day-old SPF chickens by eye drop. This procedure was repeated five times. Reversion of virulence of IB H120 was determined in subsequent studies in which the safety for respiratory tract and kidneys, as well as safety for the reproductive tract were compared between H120 vaccine strain and H120 passaged six times over chickens. Safety for the respiratory tract and kidneys Two groups of 20 one-day-old SPF-chickens were inoculated with a tenfold dose of either IB H120 vaccine
strain or IB H120 passaged six times over SPF-chickens. On each of days 5, 7 and 10 after inoculation, ciliary activity of tracheal explants was determined and a sample of the kidney was taken. No clinical signs or mortality due to IB H120 was observed. In 1 out of 20 chickens inoculated with IB H120 passaged six times over SPF-chickens, slightly increased ventilation due to accumulation of mucus in the trachea was observed, which was considered secondary to infection with IB H120. The mean ciliary activity scores were comparable between chickens vaccinated with IB H120 and IB H120 passaged six times over SPF-chickens, indicating that there is no increase of virulence of IB H120. The mean kidney lesion scores after inoculation with IB H120 were low for both groups. At most, moderate nephritis was observed in 3 out of 15 chickens inoculated with IB H120 and in 3 out of 15 chickens inoculated with IB H120 passaged over SPF-chickens. The results show that the used strain IB H120 has a low nephropathogenic tendency and that there is no evidence of an increase of virulence.

DEVELOPMENT OF LIVE ATTENUATED VACCINE AGAINST INFECTIOUS BRONCHITIS VIRUS ACCORDING TO CURRENT EUROPEAN REGULATIVE
124
It is concluded that IB H120 complies with the safety test for the respiratory tract and kidneys as stated in EP monograph 04/2013:0442 without an indication of an increase of virulence. Safety for reproductive system Infectious bronchitis virus is an epitheliotropic virus able to cause macroscopic abnormalities in organs and tissues that are lined with susceptible epithelial cells. Different pathotypes of IB virus exist, which is reflected by the diversity in clinical signs that can be found. IB strains which affect the oviducts can cause macroscopic abnormalities of the oviducts, especially when chickens are infected in their first week of life. Up to 50% of the hens can be affected, hence economic loss can be severe. Two groups of 46 one-day-old female SPF-chickens were inoculated by eye-nose drop with either vaccine strain IB H120 or IB H120 passaged six times over SPF-chickens in a dose of 104.5 EID50 per chicken. The dose given was equivalent to the maximum virus titer likely to be contained in one dose of the vaccine. All the inoculated chickens were observed for a period of ten weeks after inoculation. Subsequently, the chickens were euthanized and the oviducts were examined macroscopically for abnormalities. No abnormalities of the oviducts were observed either in the chickens inoculated with vaccine strain IB H120 or in the chickens inoculated with maximally passaged IB H120 virus. Therefore, it can be concluded that vaccine strain IB H120 is safe for the reproductive tract of chickens even if given at one day of age. Efficacy in SPF chickens Efficacy of IBV H120 vaccine was first demonstrated in SPF chickens by vaccination with minimum recommended dose, by all three recommended administration routes, following Ph. Eur. requirements. Chickens were vaccinated at day-old by coarse spray and eye-nose drop method or at 7 days old by drinking water administration. Two groups of control chickens were included in the study and left unvaccinated. One group was challenged at 21-day-old (control group for coarse spray and eye-nose drop method), while second group was challenged at 28-day-old (control group for drinking water administration). Three weeks after vaccination, antibody titers to IBV were determined using commercial ELISA kit (Idexx). Subsequently, each chicken was challenged with IBV strain M41 by eye drop in a dose of 104 EID50 per chicken. The mean ELISA antibody titers to IBV of 312, 421 and 357 were found for the group of chickens vaccinated by coarse spray, eye-nose drop and oral administration, respectively. No clinical signs indicating disease were observed during the study. After challenge, protection rates of 100% (0 out of 20 affected), 100% (0 out of 20 affected) and 95% (1 out of 20 affected) were found for the group of chickens vaccinated by coarse spray, eye-nose drop and oral administration,
respectively. All non-vaccinated challenged chickens were affected and complete ciliostasis was observed in each chicken. For each administration route, not each vaccinated chicken responded serologically to IB (titer >396). For each vaccine group, the mean ELISA antibody titer to IBV remained low during the 3-week period after vaccination. It is not uncommon for live attenuated vaccines that non-responders are observed during the first few weeks after vaccination as at this time point local immunity on mucosal surfaces (e.g., trachea) is set on (Lancellotti, Montassier et al., 2014; Cook, Jackwood and Jones, 2012; Cavanagh, 2007). Due to IgA, which does not become systemic and is not detected by the serological test used, the onset of local immunity in the trachea caused by IB H120 vaccine, using ciliary activity of tracheal explants as read out, was clearly demonstrated. The lack of correlation between circulating antibodies and protection against challenge was reported many times before (Gelb Jr, Nix and Gellman, 1998; Darbyshire and Peters, 1985; Cavanagh, 2007; Toro and Fernandez, 1994).
Efficacy in MDA-positive chickens Protection against IB is normally provided in young chicks by passive maternally acquired immunity and active immunity induced by early vaccination. IBV vaccines based on strain H120 are usually administered to chicks in the first week of their life. At this age, the potential interference of MDA with vaccines may hamper the development of active immunity. MDA are passively transmitted from the hen to the offspring to provide protection of young birds against pathogens during the first week of life (Hamal et al., 2006). Since vaccination of 1-day-old commercial broilers against IBV is widely performed, the vaccine virus may be partially or fully neutralized by circulating MDA and prevent development of active immunity. Several groups have reported a negative effect of high levels of MDA against the vaccine strain when it is applied on the day of hatch (Mondal and Naqi, 2001; Pensaert and Lambrechts, 1994), whereas others did not detect lower efficacy of the day-old vaccination in the presence of maternal antibodies (Darbyshire and Peters, 1985; Davelaar and Kouwenhoven, 1977). In the present study, we assessed the efficacy of Avishield IB H120 in young birds with and without MDA. The vaccine was applied on 1-day-old broiler chicks by spray and oculonasal route and on 7-day-old broiler chicks via drinking water. Each vaccinated group consisted of 20 chickens, while control groups consisted of 5 chickens per group. At different time points (days 10, 21 and 35) after vaccination, protection against homologous IBV challenge was tested to assess the onset and duration of the immunity achieved by the vaccine. The onset of protection was observed already at 10 days after vaccination, reaching 50%-85% protection depending on the route of administration (Table 4). Ten-day-old MDA broilers which were left unvaccinated were completely

DEVELOPMENT OF LIVE ATTENUATED VACCINE AGAINST INFECTIOUS BRONCHITIS VIRUS ACCORDING TO CURRENT EUROPEAN REGULATIVE
125
unprotected already at this time point, regardless of high MDA levels at D0 (Table 4). This correlates well with previous findings where it was demonstrated that although MDA protect chickens when challenged at day old, the protection diminishes rapidly within the first weeks of life (Mockett, Cook and Huggins, 1987; Mondal and Naqi, 2001). Differences in the levels of protection between the MDA positive and MDA negative groups within each application method were not significant (p=0.0931 for eye-nose drop; p=0.3373 for spray and p=0.2560 for oral administration). Protection reached 100% by day 21 post vaccination in all vaccinated groups. It cannot be excluded that complete protection was achieved even sooner. Immunity developed apparently faster than reported in previous comparable experiments (al-Tarcha, Kojnok and Varro, 1991; Davelaar and Kouwenhoven, 1977; Andrade, Villegas and Fletcher, 1983). Duration of immunity was tested at 35 days after
vaccination, when all vaccinated groups showed complete protection. It may be expected that immunity would sustain even longer than the last challenge point at 35 days post vaccination (Darbyshire and Peters, 1984; Gough and Alexander, 1979). In all of the non-vaccinated groups, complete ciliostasis was observed in tracheal explants after challenge with IB M41 (Table 4). From our results, it can be concluded that MDA had no detectable impact on vaccination of day-old broilers via spray/eye-nose drop route or on the vaccination of 7-day-old broilers via drinking water. From previous studies, authors also report successful vaccination of 1-,15- and 20- day-old MDA+ chicks, whereas lower protection and more clinical signs were observed after vaccination of 6- and 10-day-old chicks (Davelaar and Kouwenhoven, 1977). Contrary, in another experiment, vaccination of day-old chicks was ineffective in MDA+ chicks (Mondal and Naqi, 2001).
Table 4. Protection of chickens vaccinated with live IBV H120 vaccine against challenge with 4 log10 EID50 of IB M41 based on
ciliostasis in tracheal explants Tablica 4. Zaštita pilića cijepljenih živim cjepivom IBV H120 protiv izazivačke infekcije s 4 log10 EID50 IB M41 na temelju
određivanja ciliostaze trahealnih prstenova
Administration route of vaccine
Put primjene cjepiva MDAC n
Protection after challenge / Zaštita nakon izazivačke infekcije
10 days after vaccination 10 dana nakon cijepljenja
21 days after vaccination 21 dan nakon cijepljenja
35 days after vaccination 35 dana nakon cijepljenja
Eye-nose drop Okulo-nazalno
Positive / Pozitivni 20 60%A 100% 100%
Negative / Negativni 20 84%A** - 100%
Spray / Sprej Positive / Pozitivni 20 65%B 100% 100%
Negative / Negativni 20 50%B - 100%
Not vaccinated Necijepljeni
Positive / Pozitivni 5 0% 0% 0%
Negative / Negativni 5 0% 0% 0%
Oral / Oralno Positive / Pozitivni 20 85%C 100% 100%
Negative / Negativni 20 70%C - 100%
Not vaccinated Necijepljeni
Positive / Pozitivni 5 0% 0% 0%
Negative / Negativni 5 0% 0% 0%
Cmaternally derived antibodies n= number of chicks per group, - not tested *For each administration route and each time point, pairwise comparisons were made between broilers with (=positive) MDA and broilers without (=negative) MDA using Fisher exact test (p<0.05). Different superscript letters designate significantly different groups **n=19 Croditeljska antitijela, n= broj pilića po skupini, - nije ispitano. *za svaki put primjene i svaku točku ispitivanja usporedba stupnja zaštite između brojlera s roditeljskim antitijelima (MDA pozitivni) i brojlera bez roditeljskih antitijela (MDA negativni) je provedena koristeći Fisher exact test (p<0.05). Različita slova u eksponentu označavaju skupine kod kojih je bilo statistički značajne razlike **n=19
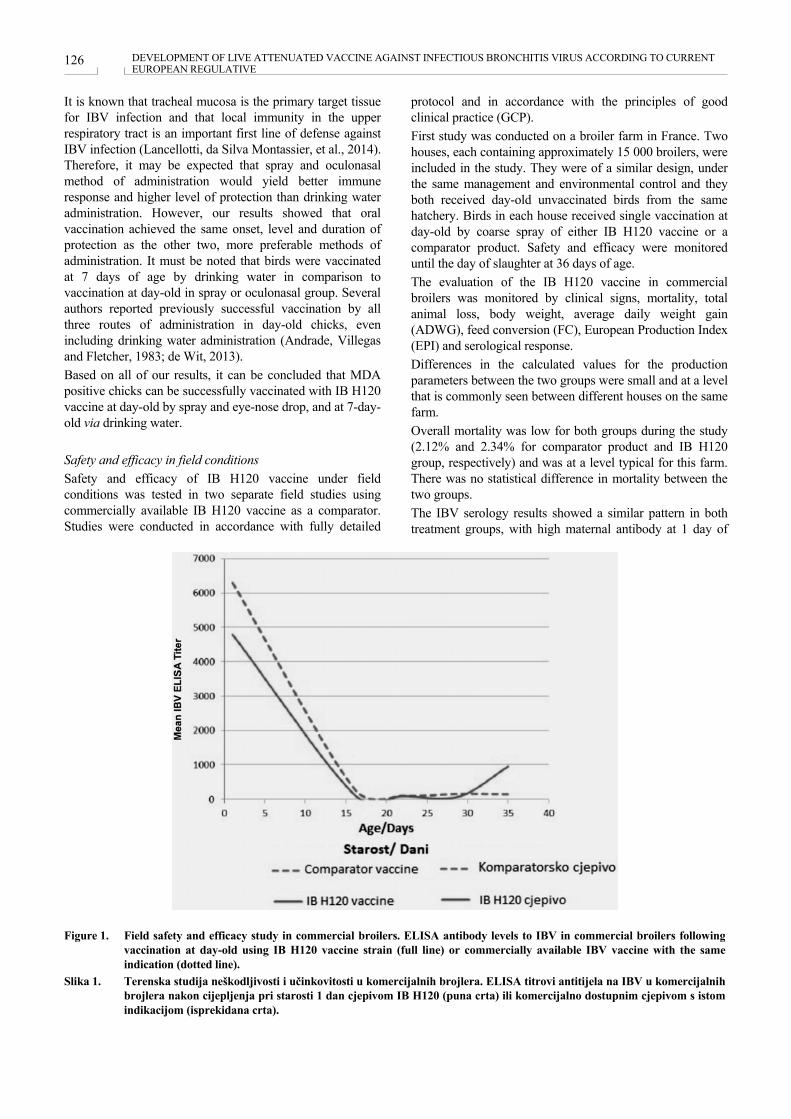
DEVELOPMENT OF LIVE ATTENUATED VACCINE AGAINST INFECTIOUS BRONCHITIS VIRUS ACCORDING TO CURRENT EUROPEAN REGULATIVE
126
It is known that tracheal mucosa is the primary target tissue for IBV infection and that local immunity in the upper respiratory tract is an important first line of defense against IBV infection (Lancellotti, da Silva Montassier, et al., 2014). Therefore, it may be expected that spray and oculonasal method of administration would yield better immune response and higher level of protection than drinking water administration. However, our results showed that oral vaccination achieved the same onset, level and duration of protection as the other two, more preferable methods of administration. It must be noted that birds were vaccinated at 7 days of age by drinking water in comparison to vaccination at day-old in spray or oculonasal group. Several authors reported previously successful vaccination by all three routes of administration in day-old chicks, even including drinking water administration (Andrade, Villegas and Fletcher, 1983; de Wit, 2013). Based on all of our results, it can be concluded that MDA positive chicks can be successfully vaccinated with IB H120 vaccine at day-old by spray and eye-nose drop, and at 7-day-old via drinking water. Safety and efficacy in field conditions Safety and efficacy of IB H120 vaccine under field conditions was tested in two separate field studies using commercially available IB H120 vaccine as a comparator. Studies were conducted in accordance with fully detailed
protocol and in accordance with the principles of good clinical practice (GCP). First study was conducted on a broiler farm in France. Two houses, each containing approximately 15 000 broilers, were included in the study. They were of a similar design, under the same management and environmental control and they both received day-old unvaccinated birds from the same hatchery. Birds in each house received single vaccination at day-old by coarse spray of either IB H120 vaccine or a comparator product. Safety and efficacy were monitored until the day of slaughter at 36 days of age. The evaluation of the IB H120 vaccine in commercial broilers was monitored by clinical signs, mortality, total animal loss, body weight, average daily weight gain (ADWG), feed conversion (FC), European Production Index (EPI) and serological response. Differences in the calculated values for the production parameters between the two groups were small and at a level that is commonly seen between different houses on the same farm. Overall mortality was low for both groups during the study (2.12% and 2.34% for comparator product and IB H120 group, respectively) and was at a level typical for this farm. There was no statistical difference in mortality between the two groups. The IBV serology results showed a similar pattern in both treatment groups, with high maternal antibody at 1 day of
Figure 1. Field safety and efficacy study in commercial broilers. ELISA antibody levels to IBV in commercial broilers following
vaccination at day-old using IB H120 vaccine strain (full line) or commercially available IBV vaccine with the sameindication (dotted line).
Slika 1. Terenska studija neškodljivosti i učinkovitosti u komercijalnih brojlera. ELISA titrovi antitijela na IBV u komercijalnihbrojlera nakon cijepljenja pri starosti 1 dan cjepivom IB H120 (puna crta) ili komercijalno dostupnim cjepivom s istomindikacijom (isprekidana crta).

DEVELOPMENT OF LIVE ATTENUATED VACCINE AGAINST INFECTIOUS BRONCHITIS VIRUS ACCORDING TO CURRENT EUROPEAN REGULATIVE
127
age, which declined to very low levels in both groups by two weeks after vaccination. Serology results showed no significant differences between the two groups until five weeks after vaccination (Figure 1), where the mean titer was significantly higher in the IB H120 vaccinated group compared with the comparator vaccine group (p<0.001) and the number of positive birds was much higher in the H120 vaccinated group (14/20) compared with the comparator group (3/20). There were no clinical signs of IB or significant increase in mortality accompanying this rise in titer and the growth curve showed that growth rates were not affected during this time. PCR analysis showed that out of 10 cloacal swabs taken 35 days after vaccination in the group vaccinated with IB H120, 3 samples were positive for the presence of IBV. Sequence analysis showed 99% homology with H120 for all three positive samples. These results indicated that there was no high level of active IB virus circulating in the birds and no variant strains of IB were identified. In the absence of any clinical signs to indicate disease, it can be concluded that an increase in IB titer over this kind of range (9-3564) is likely to be due to seroconversion following the IB vaccination at day-old rather than infection. PCR analysis performed on 10 cloaca
swab samples taken in comparator group did not detect the presence of IBV in this group 35 days after vaccination. Safety and efficacy of IB H120 vaccine in layers under field conditions was tested on two different sites in Slovenia. Two houses of a similar structure and environmental control, as well as a similar number of birds (approximately 10 000 birds) were enrolled in the study. Birds in both houses derived from the same breeder flock. The vaccine was administered to day-old chicks (as spray administration), then again at 4, 8 and 12 weeks of age (orally via drinking water). Birds in one house were vaccinated with IB H120 vaccine and in the other house with comparator product. Safety and efficacy were monitored for 16 weeks (4 weeks after last re-vaccination). The evaluation of the tested vaccine was conducted by serological response, clinical signs, mortality, total animal loss, body weight and food consumption. Overall mortality was low in both groups during the study and was at a level that was typical of the two farms. The IBV serology results showed that both treatment groups started with high MDA after hatch (Figure 2). Titers declined over the first 2 months, with both groups
Figure 2. Field safety and efficacy study in commercial layers. ELISA antibody levels to IBV in commercial layers followingvaccination at 1 day-old (spray administration) and again at 4-, 8- and 12-week-old (drinking water administration) using IB H120 vaccine strain (full line) or commercially available IBV vaccine with the same indication (dotted line).
Slika 2. Terenska studija neškodljivosti i učinkovitosti u komercijalnih nesilica. ELISA titrovi antitijela na IBV ukomercijalnim nesilicama nakon cijepljenja pri starosti 1 dan (primjena sprejem) te ponovno pri starosti 4, 8 i 12tjedana (primjena u vodi za piće) cjepivom IB H120 (puna crta) ili komercijalno dostupnim cjepivom s istomindikacijom (isprekidana crta).

DEVELOPMENT OF LIVE ATTENUATED VACCINE AGAINST INFECTIOUS BRONCHITIS VIRUS ACCORDING TO CURRENT EUROPEAN REGULATIVE
128
having a titer below 750 eight weeks after hatch. After this time point, the two groups diverged in their serological pattern. At 12 and 16 weeks of age, the mean titer was significantly higher in the IB H120 group compared with the comparator group accompanied by the higher number of positive birds. There were no clinical signs of IB or significant increase in mortality during this period. On the contrary, the mean body weights were higher in this group at the end of the study. Additional SN serology tests were performed 12 weeks after hatch in order to identify the presence of antibodies to strains of IB that may have caused the increase in the ELISA antibody titer at this age. SN results showed high titers of antibody to M41 (SN titer groups 9-11), as well as evidence of antibody to variant strains D388 and 4/91 (SN titer groups 7-9). These results indicated that field strains were circulating in the house where IB H120 vaccinated group was settled and that natural IB challenge occurred during this study. The fact that antibody titers (ELISA) at 16 weeks of age were not as high as at 12 weeks of age indicated that the vaccinal program was effective in preventing circulation of the IB virus field challenge further within the house. The birds in the comparator vaccine group did not show similar seroconversion at 12 weeks of age and the titers for birds in this treatment group remained below 850 at all testing points other than 1 day of age. The PCR on cloacal swabs taken at 16 weeks of age did not identify any circulating IB virus in either of the treatment groups. The absence of IB virus at this time point showed that the vaccinal program was effective in preventing recirculation of the live field challenge virus in the group vaccinated with IB H120 vaccine. In conclusion, the parameters such as mortality rates, clinical signs, feed intake and body weights for the IB H120 group were broadly similar to a licensed comparator product, when both vaccines were administered to day-old chicks with high levels of MDA (by spray administration) and then again at 4, 8 and 12 weeks of age (orally via drinking water). Based on all of our results, it can be concluded that our vaccine against IB H120 completely satisfies a compre-hensive set of laboratory and field studies required by the European Pharmacopoeia and EMA guidelines in respect to efficacy and safety of live attenuated veterinary vaccines.
References
AL-TARCHA, B., J. KOJNOK, C. S. VARRO (1991): Immuni-zation of day-old chicks having maternally derived antibodies against infectious bronchitis: degree of protection as monitored by ciliary activity after intratracheal challenge. Acta Vet. Hung., 39, 83-93.
ANDRADE, L. F., P. VILLEGAS, O. J. FLETCHER (1983): Vaccination of day-old broilers against infectious bronchitis: effect of vaccine strain and route of administration. Avian Dis. 27, 178-187.
BANDE, F., S. S. ARSHAD, A. R. OMAR, M. H. BEJO, M. S. ABUBAKAR, Y. ABBA (2016): Pathogenesis and diagnos-tic approaches of avian infectious bronchitis. Adv. Vir. 1-11.
BIJLENGA, G., J. K. COOK, J. GELB, JR., J. J. DE WIT (2004): Development and use of the H strain of avian infectious bronchitis virus from the Netherlands as a vaccine: a review. Avian Pathol. 33, 550-557.
CALLISON, S. A., D. A. HILT, T. O. BOYNTON, B. F. SAMPLE, R. ROBISON, D. E. SWAYNE, M. W. JACKWOOD (2006): Development and evaluation of a real-time Taqman RT-PCR assay for the detection of infectious bronchitis virus from infected chickens. J.Virol. Meth. 138, 60-65.
CAVANAGH, D. (2007): Coronavirus avian infectious bronchitis virus. Vet. Res. 38, 281-297.
COOK, J. K. A., M. JACKWOOD, R. C. JONES (2012): The long view: 40 years of infectious bronchitis research. Avian Pathol. 41, 239-250.
Council of Europe. European Pharmacopoeia. 7th ed. Avian infectious bronchitis vaccine (live). 04/2013:0442.
Council of Europe. European Pharmacopoeia. 7th ed. 5.2.6. Evaluation of safety of veterinary vaccines and immunosera. 04/2013:50206
Council of Europe. European Pharmacopoeia. 7th ed. 5.2.7. Evaluation of efficacy of veterinary vaccines and immunosera. 04/2013:50207
DARBYSHIRE, J. H., R. W. PETERS (1985): Humoral antibody response and assessment of protection following primary vaccination of chicks with maternally derived antibody against avian infectious bronchitis virus. Res. Vet. Sci. 38, 14-21.
DARBYSHIRE, J. H., R. W. PETERS (1984): Sequential development of humoral immunity and assessment of pro-tection in chickens following vaccination and challenge with avian infectious bronchitis virus. Res. Vet. Sci. 37, 77-86.
DAVELAAR, F. G., B. KOUWENHOVEN (1977): Influence of maternal antibodies on vaccination of chicks of different ages against infectious bronchitis. Avian Pathol. 6, 41-50.
DE WIT, J. J. (2013): Underestimation of the difficulties of vaccination against viral respiratory diseases by mass application methods, XVIIIth International Congress of the World Veterinary Poultry Association. 19.-23.8.2013., Nantes, France, 63-67.
European Medicines Agency. Veterinary Medicines and Inspections. GUIDELINE ON TARGET ANIMAL SAFETY FOR VETERINARY LIVE AND INACTIVED VACCINES. EMEA/CVMP/VICH/359665/2005.
FRANZO, G., P. MASSI, C. M. TUCCIARONE, I. BARBIERI, G. TOSI, L. FIORENTINI, M. CICCOZZI, A. LAVAZZA, M. CECCHINATO, A. MORENO (2017): Think globally, act locally: Phylodynamic reconstruction of infectious bronchitis virus (IBV) QX genotype (GI-19 lineage) reveals different population dynamics and spreading patterns when evaluated on different epidemiological scales. PLoS One. 12, e0184401.
GELB JR, J., W. A. NIX, N. D. GELLMAN (1998): Infectious bronchitis virus antibodies in tears and their relationship to immunity. Avian Dis. 364-374.
GOUGH, R. E., D. J. ALEXANDER (1979): Comparison of duration of immunity in chickens infected with a live infectious bronchitis vaccine by three different routes. Res. Vet. Sci. 26: 329-332.

DEVELOPMENT OF LIVE ATTENUATED VACCINE AGAINST INFECTIOUS BRONCHITIS VIRUS ACCORDING TO CURRENT EUROPEAN REGULATIVE
129
HAMAL, K. R., S. C. BURGESS, I. Y. PEVZNER, G. F. ERF (2006): Maternal antibody transfer from dams to their egg yolks, egg whites, and chicks in meat lines of chickens. Poultry Sci. 85, 1364-1372.
JACKWOOD, M. W. (2012): Review of infectious bronchitis virus around the world. Avian Dis. 56, 634-641.
LANCELLOTTI, M., M. F. DA SILVA MONTASSIER, F. S. FERNANDO, K. R. SILVA, C. H. OKINO, I. LEONARDO DOS SANTOS, R. MOREIRA DOS SANTOS, M. MONEZI BORZI, A. C. ALESSI, G. T. PEREIRA (2014): Relationship Between Mucosal Antibodies and Immunity Against Avian Infectious Bronchitis Virus. Int. J. Poult. Sci. 13, 138-144.
MATTHIJS, M. G., A. BOUMA, F. C. VELKERS, J. H. VAN ECK, J. A. STEGEMAN (2008): Transmissibility of infectious bronchitis virus H120 vaccine strain among broilers under experimental conditions. Avian Dis. 52, 461-466.
MOCKETT, A. P. A., J. K. A. COOK, M. B. HUGGINS (1987): Maternally-derived antibody to infectious bronchitis virus: Its detection in chick trachea and serum and its role in protection. Avian Pathol. 16, 407-416.
MONDAL, S. P., S. A. NAQI (2001): Maternal antibody to infectious bronchitis virus: its role in protection against infection and development of active immunity to vaccine. Vet. Immunol. Immunopathol. 79, 31-40.
PENSAERT, M., C. LAMBRECHTS (1994): Vaccination of chickens against a Belgian nephropathogenic strain of infectious bronchitis virus B1648 using attenuated homo-logous and heterologous strains. Avian Pathol. 23, 631-641.
The European agency for the evaluation of medicinal products. Committee for veterinary medicinal products. Note for guidance: Field trials with veterinary vaccines. EMEA/CVMP/852/99. (2001).
TORO, H., I. FERNANDEZ (1994): Avian infectious bronchitis: specific lachrymal IgA level and resistance against challenge. J.Vet. Med. B. 41, 467-472.
TORO, H., V. L. VAN SANTEN, M. W. JACKWOOD (2012): Genetic diversity and selection regulates evolution of infectious bronchitis virus. Avian Dis. 56, 449-455.
ZHAO, Y., J. L. CHENG, X. Y. LIU, J. ZHAO, Y. X. HU, G. Z. ZHANG (2015): Safety and efficacy of an attenuated Chinese QX-like infectious bronchitis virus strain as a candidate vaccine. Vet. Microbiol. 180, 49-58.
RAZVOJ ŽIVOG ATENUIRANOG CJEPIVA PROTIV PTIČJEG ZARAZNOG BRONHITISA U SKLADU S EUROPSKOM REGULATIVOM
Sažetak
Ptičji zarazni bronhitis je vrlo zarazna bolest respiratornog i urogenitalnog sustava u ptica, a uzrokovana je virusom ptičjeg zaraznog bronhitisa (engl. infectious bronchitis virus, IBV). Zbog velikih gubitaka koje ova bolest uzrokuje u peradarskoj proizvodnji važno je primjenjivati stroge biosigurnosne i profilaktičke mjere. Na tržištu je prisutan velik broj živih, atenuiranih i inaktiviranih cjepiva protiv IBV-a koja se temelje na različitim cjepnim sojevima. Najčešće korišteno živo atenuirano cjepivo protiv IBV-a sadrži soj H120 koji pripada Massachusetts serotipu i prisutno je na tržištu više od 50 godina. Europska farmakopeja (Ph. Eur.) i druge europske regulative strogo propisuju koje testove sigurnosti i učinkovitosti treba zadovoljiti cjepivo da bi bilo uspješno registrirano u zemljama Europske Unije. U našem laboratoriju razvili smo živo, atenuirano cjepivo protiv IBV-a temeljeno na cjepnom soju H120. Studije neškodlijvosti su provedene na SPF pilićima gdje je dokazano da čak nakon 6 uzastopnih pasaža na pilićima naš cjepni soj ne postaje virulentniji. Nadalje, u testu širenja i diseminacije IB H120 se proširio 3 puta između skupina SPF pilića, a da nije uzrokovao pobol ili smrtnost. H120 je otkriven u trahejama, slezeni, bubrezima, plućima, cekalnim tonzilama i jejunumu pilića bez da je uzrokovao patološke promjene na tim organima. Cjepivo temeljeno na H120 soju virusa najčešće se primjenjuje u prvom tjednu života pilića kada je razina roditeljskih antitijela vrlo visoka. Takva primjena predstavlja određen rizik da će roditeljska antitijela neutralizirati dio cjepnog virusa i time negativno utjecati na razvoj imunosti. U našem istraživanju proveli smo ispitivanje učinkovitosti cjepiva baziranog na soju H120 na tovnim pilićima s roditeljskim antitijelima i bez njih. Pilići su cijepljeni okulo-nazalno, sprejanjem, putem vode za piće ili su ostali necijepljeni. U različitim vremenskim intervalima nakon cijepljenja određivana je zaštita od izazivačke infekcije patogenim sojem M41. Rezultati su pokazali da već nakon 10 dana dolazi do značajnog razvoja imunosti. U ovom ispitivanju nije dokazan negativan utjecaj roditeljskih antitijela na učinkovitost cijepljenja. Nakon provedenih laboratorijskih studija učinkovitosti i sigurnosti, kvaliteta cjepiva dokazana je i u terenskim studijama koje su provedene na komercijalnim farmama u Francuskoj i Sloveniji. Proizvodni rezultati su pokazali da je cjepivo sigurno i učinkovito za primjenu u terenskim uvjetima i usporedivo sa sličnim postojećim cjepivima protiv IBV-a. Ključne riječi: virus zaraznog bronhitisa, cijepljenje, protutijela majčina podrijetla

PERADARSKI DANI 2019. 130
UTJECAJ VISINE STELJE NA JAČINU KONTAKTNOG DERMATITISA NA JASTUČIĆIMA NOGU I TARZALNIM ZGLOBOVIMA TOVNIH PILIĆA
Ivana Sabolek1, Srebrenka Nejedli2, Ivan Vlahek3, Kristina Matković1, Sven Menčik3, Aneta Piplica3, Željko Pavičić1, Mario Ostović1
1 Sveučilište u Zagrebu, Veterinarski fakultet Zagreb, Zavod za higijenu, ponašanje i dobrobit životinja, Zagreb, Hrvatska 2 Sveučilište u Zagrebu, Veterinarski fakultet Zagreb, Zavod za anatomiju, histologiju i embriologiju, Zagreb, Hrvatska 3 Sveučilište u Zagrebu, Veterinarski fakultet Zagreb, Zavod za uzgoj životinja i stočarsku proizvodnju, Zagreb, Hrvatska
Sažetak
Stelja je čimbenik o kojem ovise dobrobit, proizvodnost i kvaliteta mesa peradi. Cilj ovoga rada bio je istražiti utječe li i u kojoj mjeri visina stelje na jačinu kontaktnog dermatitisa na nogama pilića u tovu. Četiri skupine pilića (n=30 svaka) držane su na stelji od sjeckane slame različite visine (2 cm, 5 cm, 10 cm i 15 cm) tijekom šestotjednog tova, uz gustoću naseljenosti od 30 kg/m2. Jačina kontaktnog dermatitisa na jastučićima nogu i tarzalnim zglobovima procijenjena je početkom svakog tjedna te na liniji klanja, na temelju protokolu Welfare Quality®. Analiza podataka provedena je referentnim programom Statistica v. 13.4. Rezultati istraživanja pokazali su da je kontaktni dermatitis na jastučićima nogu i tarzalnim zglobovima, procijenjen na farmi i na liniji klanja, bio u prosjeku značajno izraženiji (p<0,05) u pilića držanih na stelji visine 2 cm u usporedbi s 5 cm, 10 cm i 15 cm. Procjena na liniji klanja također je pokazala da su lezije na tarzalnim zglobovima bile značajno izraženije (p<0,05) u pilića držanih na visini stelje od 5 cm i 10 cm naspram 15 cm. S obzirom na istraživane visine stelje i utvrđene jačine kontaktnog dermatitisa na jastučićima nogu i tarzalnim zglobovima može se zaključiti da bi visina stelja u tovu pilića trebala iznositi najmanje 5 cm. Uz to, u svrhu smanjenja jačine kontaktnog dermatitisa na tarzalnim zglobovima trebalo bi dodavati nove količine stelje nakon petog tjedna tova. Ključne riječi: tovni pilići, stelja, kontaktni dermatitis, dobrobit životinja Uvod
Kontaktni dermatitis na jastučićima nogu i tarzalnim zglo-bovima tovnih pilića važan je pokazatelj, a ujedno i jedan od problema njihove dobrobiti. U početnom stadiju očituje se diskoloracijom kože, hiperkeratozom i nekrozom epidermi-sa, dok u uznapredovalim slučajevima dolazi do ulceracije i upalnih procesa u potkožnom tkivu (Greene i sur., 1985.; Bessei, 2006.; Žužul i sur., 2017.). Najčešće zahvaća obje noge, a ulceracije su često prekrivene krastama i fecesom (Martland, 1985.; Berg, 2004.). Česte su i sekundarne infekcije koje pogoršavaju stanje (Bessei, 2006.). Prema rezultatima istraživanja Berga (1998.) i de Jonga i sur. (2012.a) prosječna učestalost kontaktnog dermatitisa na jastučićima nogu pilića u tovu iznosi gotovo 40%, a na tarzalnim zglobovima 12% (Hepworth i sur., 2011.).
Kontaktni dermatitis uzrokuje poremetnje u hodu pilića, narušavajući ne samo njihovu dobrobit, već i proizvodnost. Promjene na nogama su bolne, pilići se manje kreću, slabije se hrane i imaju manji prirast. U najtežim slučajevima ugi-baju od izgladnjelosti (Berg, 2004.; Avdalovic i sur., 2017.). Kontaktni dermatitis na nogama pilića važan je i s ekonomskog gledišta. Kako navode Shepherd i Fairchild (2010.), pileće su nogice treći najisplativiji dio pileta, iza prsa i krila. Brojna istraživanja pokazala su da je kontaktni dermatitis uzrokovan ponajprije vlagom i kemijskim supstancama u stelji koje djeluju nadražujuće (Martland, 1985.; Berg, 1998.; de Jong i sur., 2014.; Taira i sur., 2014.; Žužul i sur., 2017.), uz mnoštvo čimbenika koji pogoduju njegovu razvoju, kao što su vrsta, tekstura i veličina čestica stelje (Berk, 2009.; Bilgili i sur., 2009.; Cengiz i sur., 2011.;
Ivana Sabolek, dr. med. vet., asistent, Veterinarski fakultet Sveučilišta u Zagrebu, Zavod za higijenu, ponašanje i dobrobit životinja, Heinzelova 55, 10000 Zagreb, Hrvatska, Tel. +385 (0)1 2390-291, Fax: +385 (0)1 2441-390, e-mail: [email protected]

UTJECAJ VISINE STELJE NA JAČINU KONTAKTNOG DERMATITISA NA JASTUČIĆIMA NOGU I TARZALNIM ZGLOBOVIMA TOVNIH PILIĆA
131
Avdalovic i sur., 2017.), genotip (Shepherd i Fairchild, 2010.), hranidba, elastičnost kože, tjelesna masa i spol pi-lića, razdoblje godine (Mayne, 2005.), gustoća naseljenosti (Sørensen i sur., 2000.; Dozier i sur., 2005.; 2006.; Buijs i sur., 2009.; Spindler i Hartung, 2011.), vrsta pojilica (Ekstrand i sur., 1997.; Berg, 1998.), režim osvijetljenosti (Škrbić i sur., 2015.) i dr. S obzirom na utjecaj visine stelje na pojavnost kontaktnog dermatitisa, u različitim su istraživanjima utvrđeni različiti rezultati. Ekstrand i sur. (1997.) ustanovili su da su lezije češće u pilića držanih na višoj stelji (>5 cm) u usporedbi s onom manje visine (<5 cm), kao i Berg (1998.). Tucker i Walker (1999.) utvrdili su pak da je pojavnost kontaktnog dermatitisa na nogama pilića veća na nižoj (2,5 cm i 5 cm), nego na višoj (10 cm) stelji. Do takvih spoznaja došli su i Haslam i sur. (2007.), Meluzzi i sur. (2008.) i Shao i sur. (2015.). Ispitujući utjecaj visine stelje na pojavnost kontakt-nog dermatitisa na jastučićima nogu, Shepherd i sur. (2017.) zaključili su da bi visina stelje u tovu pilića trebala iznositi najmanje 7,6 cm. Stephenson i sur. (1960.) zaključili su da visina stelje nema utjecaja na pojavnost prsnih kvrga, još jednog oblika kontaktnog dermatitisa u tovnih pilića. Cilj ovoga istraživanja bio je doprinijeti razjašnjenju dosa-dašnjih oprečnih spoznaja o utjecaju visine stelje na po-javnost kontaktnog dermatitisa u tovnih pilića.
Materijal i metode
Istraživanje je provedeno u nastambi za tov pilića na obiteljskom poljoprivrednom gospodarstvu, u kontroliranim uvjetima proizvodnje. Istraživanje je obuhvatilo ukupno 120 pilića hibrida Ross 308 tijekom šestotjednog tova u ljetnom razdoblju. Pilići su bili podijeljeni u četiri jednake skupine (n=30) ograđene žičanom mrežom, od kojih je svaka držana na stelji od sjeckane slame različite visine (2 cm, 5 cm, 10 cm i 15 cm), uz gustoću naseljenosti od 30 kg/m2. Stelja ni u jednoj od skupina pilića nije preokretana niti su dodavane nove količine stelje tijekom trajanja tova. U svakom boksu bila je jedna hranilica i 5 nipl pojilica na visini koja se prilagođavala rastu pilića. Pilići su se hranili kompletnim krmnim smjesama. Jačina kontaktnog dermatitisa na jastučićima nogu i tarzal-nim zglobovima pilića pratila se u tjednim razmacima (1., 8., 15., 22., 29. i 36. dan), na temelju protokola Welfare Quality® i bodovne ljestvice 0-4 (Welfare Quality® konzorcij, 2009.). Ako su na nogama pileta utvrđene razli-čite jačine lezija, u obzir su se uzele izraženije promjene. Jačina kontaktnog dermatitisa na jastučićima nogu i tarzal-nim zglobovima procijenjena je i na liniji klanja (42. dan). Tijekom razdoblja istraživanja (28. dan) uginulo je pile u skupini držanoj na stelji visine 10 cm. Analiza podataka provedena je referentnim računalnim programom Statistica v. 13.4. (Tibco Software Inc., 2017.). Podatci su obrađeni uobičajenim testovima deskriptivne sta-tistike, a normalnost njihove raspodjele provjerena je Kol-mogorov-Smirnovljevim testom. Razlike u jačini kontakt-nog dermatitisa između tjedana unutar pojedine skupine te-
stirane su Friedmanovom analizom varijance i Wilcoxono-vim testom, a između skupina unutar pojedinog tjedna Krus-kal-Wallisovom analizom varijance. Statistički značajnom smatrala se razlika na razini p<0,05. Istraživanje je odobrilo Povjerenstvo za etiku u veterinarstvu Veterinarskog fakulteta Sveučilišta u Zagrebu.
Rezultati i rasprava
Brzi rast i povećanje tjelesne mase rezultiraju smanjenom aktivnošću te produljenim sjedenjem i odmorom tovnih pilića, zbog čega je stelja jedan od najvažnijih čimbenika koji utječu na njihovo zdravlje i dobrobit, odnosno prirast i kvalitetu mesa (Matković i sur., 2015.; Žužul i sur., 2017.). U ovom radu istraživana je jačina kontaktnog dermatitisa na jastučićima nogu i tarzalnim zglobovima pilića držanih na različitim visinama stelje. Lezije na nogama pilića mogu se razviti već nakon nekoliko dana, no najčešće se razvijaju u drugom do trećem tjednu starosti (Greene i sur., 1985.; Ekstrand i Algers, 1997.; Kjaer i sur., 2006.; Taira i sur., 2014.; Avdalovic i sur., 2017.). Prve lezije na jastučićima nogu u ovom istraživanju pojavile su se u pilića držanih na stelji visine 2 cm i 5 cm u drugom tjednu tova, a u pilića na 10 cm i 15 cm visine stelje u trećem tjednu (tablica 1.). U trećem tjednu uočene su i prve lezije na tarzalnim zglobovima u pilića na visini stelje od 2 cm, dok je u skupinama na 5 cm, 10 cm i 15 cm to bio slučaj u četvrtom tjednu (tablica 2.). Promjene na jastučićima nogu u pravilu su postajale izraženije u svim skupinama sa sva-kim tjednom tova, a takvu tendenciju pokazivale su i promjene na tarzalnim zglobovima u pilića držanih na visini stelje od 2 cm, koje su se u ostalim skupinama, na većim visinama stelje, sporije razvijale (tablice 1. i 2.). Greene i sur. (1985.) i Škrbić i sur. (2015.) također su ustanovili da se kontaktni dermatitis na tarzalnim zglobovima razvija sporije u usporedbi s lezijama na jastučićima nogu. U trećem tjednu tova utvrđene su prve značajne razlike (p<0,05) u jačini kontaktnog dermatitisa na jastučićima nogu između skupina, pri čemu su promjene bile izraženije u pilića držanih na visini stelje od 2 cm u usporedbi s 10 cm i 15 cm. Do kraja tova razlike u jačini lezija na jastučićima nogu utvrđene su jedino između skupine pilića držane na visini stelje od 2 cm i ostalih pojedinih skupina (tablica 1.). Razlike u jačini kontaktnog dermatitisa na tarzalnim zglo-bovima između skupina po tjednima tova pratile su sličan trend (tablica 2.). Promjene na jastučićima nogu, kao i na tarzalnim zglo-bovima u prosjeku su bile značajno izraženije (p<0,05) tijekom tova u skupini pilića držanih na stelji visine 2 cm nego na 5 cm, 10 cm i 15 cm (tablice 1. i 2.). Dobiveni rezultati u suglasju su s rezultatima istraživanja Tuckera i Walkera (1999.), Haslama i sur. (2007.), Meluzzija i sur. (2008.), Shaoa i sur. (2015.) i Shepherda i sur. (2017.), a mogu se objasniti čeprkanjem pilića, pri čemu su razmi-canjem najmanje količine stelje bili izloženiji betonskom podu koji je doprinio ozljedama kože za koje je poznato da pogoduju razvoju kontaktnog dermatitisa (de Jong i sur.,
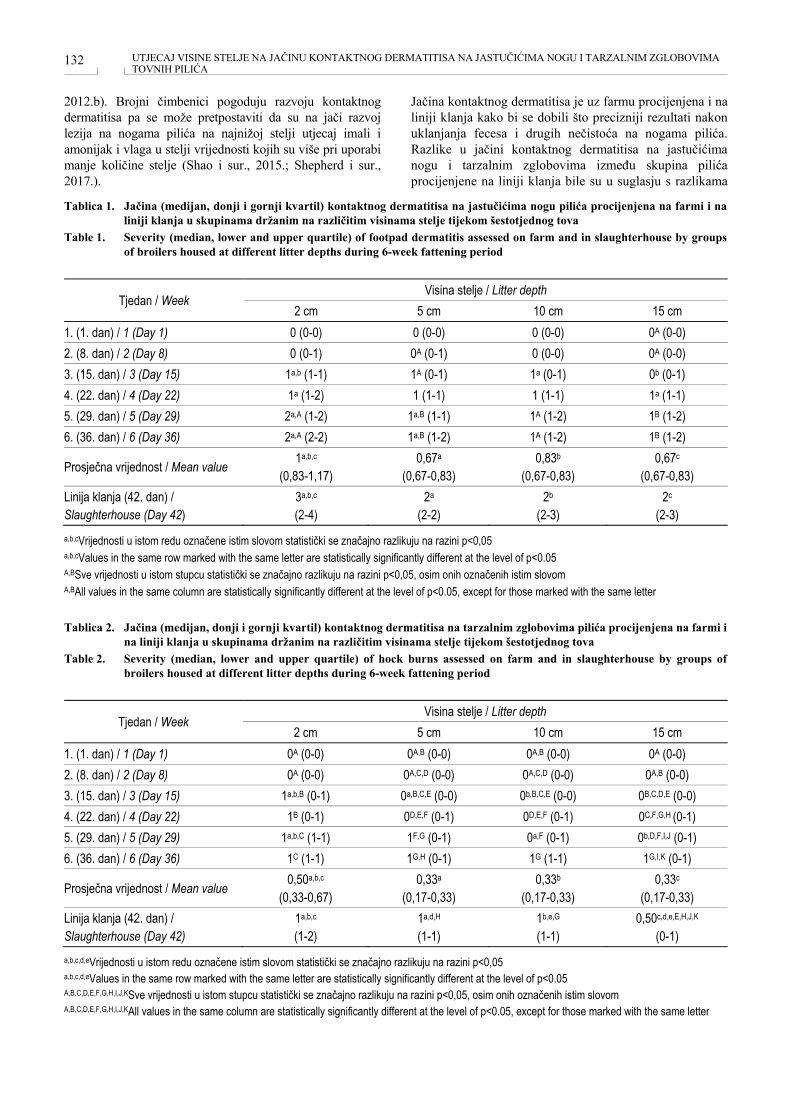
UTJECAJ VISINE STELJE NA JAČINU KONTAKTNOG DERMATITISA NA JASTUČIĆIMA NOGU I TARZALNIM ZGLOBOVIMA TOVNIH PILIĆA
132
2012.b). Brojni čimbenici pogoduju razvoju kontaktnog dermatitisa pa se može pretpostaviti da su na jači razvoj lezija na nogama pilića na najnižoj stelji utjecaj imali i amonijak i vlaga u stelji vrijednosti kojih su više pri uporabi manje količine stelje (Shao i sur., 2015.; Shepherd i sur., 2017.).
Jačina kontaktnog dermatitisa je uz farmu procijenjena i na liniji klanja kako bi se dobili što precizniji rezultati nakon uklanjanja fecesa i drugih nečistoća na nogama pilića. Razlike u jačini kontaktnog dermatitisa na jastučićima nogu i tarzalnim zglobovima između skupina pilića procijenjene na liniji klanja bile su u suglasju s razlikama
Tablica 1. Jačina (medijan, donji i gornji kvartil) kontaktnog dermatitisa na jastučićima nogu pilića procijenjena na farmi i na liniji klanja u skupinama držanim na različitim visinama stelje tijekom šestotjednog tova
Table 1. Severity (median, lower and upper quartile) of footpad dermatitis assessed on farm and in slaughterhouse by groups of broilers housed at different litter depths during 6-week fattening period
Tjedan / Week Visina stelje / Litter depth
2 cm 5 cm 10 cm 15 cm 1. (1. dan) / 1 (Day 1) 0 (0-0) 0 (0-0) 0 (0-0) 0A (0-0) 2. (8. dan) / 2 (Day 8) 0 (0-1) 0A (0-1) 0 (0-0) 0A (0-0) 3. (15. dan) / 3 (Day 15) 1a,b (1-1) 1A (0-1) 1a (0-1) 0b (0-1) 4. (22. dan) / 4 (Day 22) 1a (1-2) 1 (1-1) 1 (1-1) 1a (1-1) 5. (29. dan) / 5 (Day 29) 2a,A (1-2) 1a,B (1-1) 1A (1-2) 1B (1-2) 6. (36. dan) / 6 (Day 36) 2a,A (2-2) 1a,B (1-2) 1A (1-2) 1B (1-2)
Prosječna vrijednost / Mean value 1a,b,c (0,83-1,17)
0,67a
(0,67-0,83) 0,83b
(0,67-0,83) 0,67c
(0,67-0,83) Linija klanja (42. dan) / Slaughterhouse (Day 42)
3a,b,c (2-4)
2a (2-2)
2b
(2-3) 2c
(2-3) a,b,cVrijednosti u istom redu označene istim slovom statistički se značajno razlikuju na razini p<0,05 a,b,cValues in the same row marked with the same letter are statistically significantly different at the level of p<0.05 A,BSve vrijednosti u istom stupcu statistički se značajno razlikuju na razini p<0,05, osim onih označenih istim slovom A,BAll values in the same column are statistically significantly different at the level of p<0.05, except for those marked with the same letter Tablica 2. Jačina (medijan, donji i gornji kvartil) kontaktnog dermatitisa na tarzalnim zglobovima pilića procijenjena na farmi i
na liniji klanja u skupinama držanim na različitim visinama stelje tijekom šestotjednog tova Table 2. Severity (median, lower and upper quartile) of hock burns assessed on farm and in slaughterhouse by groups of
broilers housed at different litter depths during 6-week fattening period
Tjedan / Week Visina stelje / Litter depth
2 cm 5 cm 10 cm 15 cm 1. (1. dan) / 1 (Day 1) 0A (0-0) 0A,B (0-0) 0A,B (0-0) 0A (0-0) 2. (8. dan) / 2 (Day 8) 0A (0-0) 0A,C,D (0-0) 0A,C,D (0-0) 0A,B (0-0) 3. (15. dan) / 3 (Day 15) 1a,b,B (0-1) 0a,B,C,E (0-0) 0b,B,C,E (0-0) 0B,C,D,E (0-0) 4. (22. dan) / 4 (Day 22) 1B (0-1) 0D,E,F (0-1) 0D,E,F (0-1) 0C,F,G,H (0-1) 5. (29. dan) / 5 (Day 29) 1a,b,C (1-1) 1F,G (0-1) 0a,F (0-1) 0b,D,F,I,J (0-1) 6. (36. dan) / 6 (Day 36) 1C (1-1) 1G,H (0-1) 1G (1-1) 1G,I,K (0-1)
Prosječna vrijednost / Mean value 0,50a,b,c
(0,33-0,67) 0,33a
(0,17-0,33) 0,33b
(0,17-0,33) 0,33c
(0,17-0,33) Linija klanja (42. dan) / Slaughterhouse (Day 42)
1a,b,c (1-2)
1a,d,H (1-1)
1b,e,G (1-1)
0,50c,d,e,E,H,J,K (0-1)
a,b,c,d,eVrijednosti u istom redu označene istim slovom statistički se značajno razlikuju na razini p<0,05 a,b,c,d,eValues in the same row marked with the same letter are statistically significantly different at the level of p<0.05 A,B,C,D,E,F,G,H,I,J,KSve vrijednosti u istom stupcu statistički se značajno razlikuju na razini p<0,05, osim onih označenih istim slovom A,B,C,D,E,F,G,H,I,J,KAll values in the same column are statistically significantly different at the level of p<0.05, except for those marked with the same letter

UTJECAJ VISINE STELJE NA JAČINU KONTAKTNOG DERMATITISA NA JASTUČIĆIMA NOGU I TARZALNIM ZGLOBOVIMA TOVNIH PILIĆA
133
utvrđenim na farmi, usto što su na liniji klanja značajno jače (p<0,05) lezije na tarzalnim zglobovima utvrđene i u pilića držanih na stelji visine 5 cm i 10 cm u odnosu na 15 cm (tablice 1. i 2.). Ovi nalazi mogu se protumačiti tanjom i osjetljivijom kožom na tarzalnim zglobovima u usporedbi s jastučićima nogu te kasnijom pojavom lezija na tarzalnim zglobovima u skupinama pilića držanih na 5 cm, 10 cm i 15 cm visine stelje. Tako su promjene na tarzalnim zglo-bovima postale najizraženije na kraju tova kada pilići najviše vremena provode sjedeći uslijed povećanja tjelesne mase, uz istodobno pogoršanje kvalitete stelje (Bessei, 2006.). To je rezultiralo jačim razvojem lezija na tarzalnim zglobovima u svih skupina pilića držanih na visini stelje manje od 15 cm.
Zaključak
Iz rezultata istraživanja može se zaključiti da bi visina stelje u tovu trebala biti minimalno 5 cm s obzirom na po-javnost kontaktnog dermatitisa na jastučićima nogu i tar-zalnim zglobovima pilića. Također, u svrhu smanjenja jači-ne kontaktnog dermatitisa na tarzalnim zglobovima uputno je dodavati svježu stelju nakon petog tjedna tova.
Literatura
AVDALOVIC, V., M. VUCINIC, R. RESANOVIC, J. AVDALOVIC, D. MASLIC-STRIZAK, M. VUCICEVIC (2017): Effect of pelleted and chopped wheat straw on the footpad dermatitis in broilers. Pak. J. Zool. 49, 1639-1646.
BERG, C. (2004): Pododermatitis and hock burn in broiler chickens. In: Measuring and Auditing Broiler Welfare. (C. A. Weeks, A. Butterworth, Eds.), CABI Publishing, Wallingford, UK, 37-49.
BERG, C. C. (1998): Foot-pad dermatitis in broilers and turkeys – prevalence, risk factors and prevention. PhD Thesis. Swedish University of Agricultural Sciences, Uppsala, Sweden. [https://pub.epsilon.slu.se/1514/1/Lotta_Berg_Avhandling.pdf, (28.12.2018.)]
BERK, J. (2009): Effect of litter type on prevalence and severity of pododermatitis in male broilers. Berl. Munch. Tierarztl. Wochenschr. 122, 257-263.
BESSEI, W. (2006): Welfare of broilers: a review. Worlds Poult. Sci. J. 62, 455-466.
BILGILI, S. F., J. B. HESS, J. P. BLAKE, K. S. MACKLIN, B. SAENMAHAYAK, J. L. SIBLEY (2009): Influence of bedding material on footpad dermatitis in broiler chickens. J. Appl. Poult. Res. 18, 583-589.
BUIJS, S., L. KEELING, S. RETTENBACHER, E. VAN POUCKE, F. A. M. TUYTTENS (2009): Stocking density effects on broiler welfare: identifying sensitive ranges for different indicators. Poult. Sci. 88, 1536-1543.
CENGIZ, Ö., J. B. HESS, S. F. BILGILI (2011): Effect of bedding type and transient wetness on footpad dermatitis in broiler chickens. J. Appl. Poult. Res. 20, 554-560.
DE JONG, I., C. BERG, A. BUTTERWORTH, I. ESTEVÉZ (2012b): Scientific report updating the EFSA opinions on the welfare of broilers and broiler breeders. Supporting Publi-cations 2012:EN-295. [https://www.efsa.europa.eu/en/ /supporting/pub/en-295, (25.01.2019.)]
DE JONG, I. C., H. GUNNINK, J. VAN HARN (2014): Wet litter not only induces footpad dermatitis but also reduces overall welfare, technical performance, and carcass yield in broiler chickens. J. Appl. Poult. Res. 23, 51-58.
DE JONG, I. C., J. VAN HARN, H. GUNNINK, V. A. HINDLE, A. LOURENS (2012a): Footpad dermatitis in Dutch broiler flocks: prevalence and factors of influence. Poult. Sci. 91, 1569-1574.
DOZIER, W. A., 3rd, J. P. THAXTON, J. L. PURSWELL, H. A. OLANREWAJU, S. L. BRANTON, W. B. ROUSH (2006): Stocking density effects on male broilers grown to 1.8 kilograms of body weight. Poult. Sci. 85, 344-351.
DOZIER, W. A., 3rd, J. P. THAXTON, S. L. BRANTON, G. W. MORGAN, D. M. MILES, W. B. ROUSH, B. D. LOTT, Y. VIZZIER-THAXTON (2005): Stocking density effects on growth performance and processing yields of heavy broilers. Poult. Sci. 84, 1332-1338.
EKSTRAND, C., B. ALGERS (1997): The effect of litter mois-ture on the development of footpad dermatitis in broilers. 11th International Congress of the World Veterinary Poultry Association. 18.-22.08.1997., Budapest, Hungary, 370.
EKSTRAND, C., B. ALGERS, J. SVEDBERG (1997): Rearing conditions and foot-pad dermatitis in Swedish broiler chickens. Prev. Vet. Med. 31, 167-174.
GREENE, J. A., R. M. McCRACKEN, R. T. EVANS (1985): A contact dermatitis of broilers – clinical and pathological findings. Avian Pathol. 14, 23-38.
HASLAM, S. M., T. G. KNOWLES, S. N. BROWN, L. J. WILKINS, S. C. KESTIN, P. D. WARRISS, C. J. NICOL (2007): Factors affecting the prevalence of foot pad dermatitis, hock burn and breast burn in broiler chicken. Br. Poult. Sci. 48, 264-275.
HEPWORTH, P. J., A. V. NEFEDOV, I. B. MUCHNIK, K. L. MORGAN (2011): Hock burn: an indicator of broiler flock health. Vet. Rec. 168, 303.
KJAER, J. B., G. SU, B. L. NIELSEN, P. SØRENSEN (2006): Foot pad dermatitis and hock burn in broiler chickens and degree of inheritance. Poult. Sci. 85, 1342-1348.
MARTLAND, M. F. (1985): Ulcerative dermatitis in broiler chickens: the effects of wet litter. Avian Pathol. 14, 353-364.
MATKOVIĆ, K., D. MARUŠIĆ, Ž. PAVIČIĆ, N. POLJIČAK MILAS, M. OSTOVIĆ, H. LUCIĆ (2015): Pregled morfo-loških svojstava autopodija pilića u tovu kao pokazatelja kvalitete smještaja i dobrobiti. XI. simpozij Peradarski dani 2015. s međunarodnim sudjelovanjem. 13.-16.05.2015., Ši-benik, Hrvatska, 129-132.
MAYNE, R. K. (2005): A review of the aetiology and possible causative factors of foot pad dermatitis in growing turkeys and broilers. Worlds Poult. Sci. J. 61, 256-267.
MELUZZI, A., C. FABBRI, E. FOLEGATTI, F. SIRRI (2008): Effect of less intensive rearing conditions on litter characteristics, growth performance, carcass injures and meat quality of broilers. Br. Poult. Sci. 49, 509-515.
SHAO, D., J. HE, J. LU, Q. WANG, L. CHANG, S. R. SHI, T. H. BING (2015): Effects of sawdust thickness on the growth performance, environmental condition, and welfare quality of yellow broilers. Poult. Sci. 94, 1-6.
SHEPHERD, E. M., B. D. FAIRCHILD (2010): Footpad dermatitis in poultry. Poult. Sci. 89, 2043-2051.
SHEPHERD, E. M., B. D. FAIRCHILD, C. W. RITZ (2017): Alternative bedding materials and litter depth impact litter

UTJECAJ VISINE STELJE NA JAČINU KONTAKTNOG DERMATITISA NA JASTUČIĆIMA NOGU I TARZALNIM ZGLOBOVIMA TOVNIH PILIĆA
134
moisture and footpad dermatitis. J. Appl. Poult. Res. 26, 518-528.
SØRENSEN, P., G. SU, S. C. KESTIN (2000): Effects of age and stocking density on leg weakness in broiler chickens. Poult. Sci. 79, 864-870.
SPINDLER, B., J. HARTUNG (2011): Prevalence of podo-dermatitis in broiler chickens kept according to Directive 2007/43/EC stocking densities. XVth International Congress of the International Society for Animal Hygiene (ISAH). Volume 1. 03.-07.07.2011., Vienna, Austria, 39-42.
STEPHENSON, E. L., J. M. BEZANSON, C. F. HALL (1960): Factors affecting the incidence and severity of a breast blister condition in broilers. Poult. Sci. 39, 1520-1524.
ŠKRBIĆ, Z., Z. PAVLOVSKI, M. LUKIĆ, V. PETRIČEVIĆ (2015): Incidence of footpad dermatitis and hock burns in
broilers as affected by genotype, lighting program and litter type. Ann. Anim. Sci. 15, 433-445.
TAIRA, K., T. NAGAI, T. OBI, K. TAKASE (2014): Effect of litter moisture on the development of footpad dermatitis in broiler chickens. J. Vet. Med. Sci. 76, 583-586.
TUCKER, S. A., A. W. WALKER (1999): Hock burn in broilers. In: Recent Developments in Poultry Nutrition. (J. Wiseman, P. C. Garnsworthy, Eds.), Nottingham University Press, Thrumpton, UK, 33-50.
WELFARE QUALITY® (2009): Welfare Quality® assessment protocol for poultry. Welfare Quality® Consortium, Lelystad, The Netherlands.
ŽUŽUL, S., M. OSTOVIĆ, K. MATKOVIĆ, Ž. PAVIČIĆ, J. TRŠAN, M. MIKULIĆ, I. RAVIĆ (2017): Uloga duboke stelje u tovu pilića i svinja. Meso 19, 223-228.
EFFECT OF LITTER DEPTH ON SEVERITY OF FOOTPAD DERMATITIS AND HOCK BURNS IN BROILERS
Summary
Litter is a factor influencing poultry welfare, performance and meat quality. The objective of this study was to investigate whether and to what extent the litter depth affects the severity of contact dermatitis on broiler legs. Four groups of broilers (n=30 each) were housed on chopped straw at different litter depths (2 cm, 5 cm, 10 cm and 15 cm) during 6-week fattening period, with stocking density of 30 kg/m2. The severity of footpad dermatitis and hock burns was assessed based on the Welfare Quality® protocol at the beginning of each week at a farm and in a slaughterhouse. Data analysis was performed using the respective Statistica v. 13.4 software. Study results showed the average severity of footpad dermatitis and hock burns assessed at the farm and in the slaughterhouse to be significantly higher (p<0.05) in broilers housed at 2 cm litter depth compared to 5 cm, 10 cm and 15 cm. Also, hock burns assessed in the slaughterhouse were significantly more severe (p<0.05) in broilers housed at 5 cm and 10 cm litter depth compared to 15 cm. Considering the investigated litter depths and determined severities of footpad dermatitis and hock burns, it can be concluded that litter depth for broilers should be at least 5 cm. In addition, new amounts of litter should be provided from fifth week of fattening period onwards in order to reduce the severity of hock burns. Key words: broilers, litter, contact dermatitis, animal welfare

PERADARSKI DANI 2019. 135
UČINAK GUSTOĆE NASELJENOSTI NA KONFORMACIJU TOVNIH PILIĆA
Almira Softić1, Aida Kavazović1, Aida Kustura1, Velija Katica1, Behija Dukić2, Amina Hrković-Porobija3, Maja Varatanović1, Ajla Ališah1
1 Univerzitet u Sarajevu, Veterinarski fakultet, Zavod za zootehniku i peradarstvo, Sarajevo, Bosna i Hercegovina 2 Univerzitet u Sarajevu, Veterinarski fakultet, Zavod za epizootiologiju, Sarajevo, Bosna i Hercegovina 3 Univerzitet u Sarajevu, Veterinarski fakultet, Zavod za morfologiju, Sarajevo, Bosna i Hercegovina
Sažetak
Cilj istraživanja je bio utvrditi učinak različite gustoće naseljenosti na konformaciju i kvalitetu trupova tovnih pilića linije Cobb. U pokusu je korišteno ukupno 120 jednodnevnih pilića podijeljenih u tri skupine (A, B i K) u trajanju do šest tjedana. Skupina A je bila s manjom gustoćom naseljenosti (12 pilića/m2), skupina B s većom gustoćom naseljenosti (18 pilića/m2) i kontrolna K skupina (15 pilića/m2) u kojoj je naseljenost bila u skladu s tehnološkim normativima ove linije pilića. Na kraju prvog tjedna tova metodom slučajnog uzorka uzeto je i obilježeno po 10 pilića iz svake skupine. Tijekom pokusa kod obilježenih pilića tjedno su praćeni i mjereni opseg prsa, opseg batka, duljina kobilice, dubina prsa i prsni kut te je mjerena živa tjelesna masa. Na kraju tova sva mjerenja provedena na živim jedinkama provedena su i na njihovim trupovima, kao i prosječna masa trupa, randman te udjeli jestivih dijelova, težine jetre i abdominalne masti u odnosu na masu trupa. Najbolje vrijednosti konformacije živih pilića te njihovih trupova imali su pilići kontrolne skupine. Utvrđene razlike ovih pokazatelja u odnosu na pokusne skupine nisu bile statistički značajne osim kod trupova pilića za opseg prsa, prsni kut, duljinu kobilice i dubinu prsa (p<0,05). Uz to, veći ali ne i statistički značajno veći udio ispitivanih jestivih dijelova u odnosu na završnu masu trupa utvrđen je kod pilića kontrolne skupine naseljenost kojih je bila u skladu s tehnološkim normativima. Povoljniju konformaciju iskazano kroz indekse imali su također pilići kontrolne skupine (p<0,05), osim za opseg batka. Ključne riječi: gustoća naseljenosti, tovni pilići, kvaliteta trupa, konformacija Uvod U peradarskoj proizvodnji gustoća naseljenosti podrazu-mijeva količinu završne tjelesne mase ili broj jedinki po kvadratnom metru prostora od početka do kraja proizvodnog ciklusa. Ona ovisi o brojnim čimbenicima, a najvažniji je način držanja. Minimalni standardi u odnosu na dobrobit brojlera usmjereni su na prostor za njihovo hodanje, što je glavni preduvjet za razvoj lokomotornog aparata i demon-straciju osnovnih oblika ponašanja. Optimalni uvjeti smje-štaja sprječavaju gubitke u proizvodnji, a postiže se i za-dovoljavajuća ekonomska dobit. Gustoća naseljenosti utječe na veličinu proizvodnih pokazatelja i dobrobit pilića (Škrbić i sur. 2011., Giersberg, 2017.), imunološkog i fiziološkog statusa (Tong i sur., 2012., Qaid i sur., 2016., Adeyemo i sur., 2016.), ekonomičnosti (Škrbić i sur., 2008., Salihbašić i sur. 2014.) i kvalitetu trupova (Škrbić i sur. 2009). Kvaliteta trupova tovnih pilića u velikoj mjeri određuje mesnatost i ima genetsko (Ojedapo i sur. 2008.), a samim tim i
ekonomsko značenje (Pavlovski i sur., 2006.), što se pro-cjenjuje randmanom i kvalitetom jestivih dijelova trupa (Nikolova i Pavlovski, 2009.). Određivanje konformacije na živim pilićima je značajno u uzgojno-selekcijskom radu. Na osnovi procjene konformacije dobiva se opći utisak o mesnatosti pilića, što određuje mogućnost plasmana, posti-zanje određene klase i odgovarajuće cijene mesa. Cilj istraživanja je bio utvrditi učinak različite gustoće naseljenosti na konformaciju živih i kvalitetu trupova tovnih pilića linije Cobb.
Materijal i metode rada
U pokusu je korišteno ukupno 120 jednodnevnih pilića po-dijeljenih u tri skupine (A, B i K) u trajanju od šest tjedana. Skupina A je bila skupina pilića s manjom gustoćom naseljenosti (12 pilića/m2), skupina B je skupina s većom gustoćom naseljenosti (18 pilića/m2) i kontrolna skupina K (15 pilića/m2) u skladu s tehnološkim normativima date
Dr. sc Almira Softić, izvanredni profesor, Veterinarski fakultet Univerziteta u Sarajevu, Zmaja od Bosne 90, 71000 Sarajevo, Bosna i Hercegovina, Tel.: 00387 33 729 119, e-mail: [email protected]

UČINAK GUSTOĆE NASELJENOSTI NA KONFORMACIJU TOVNIH PILIĆA 136
linije. Na kraju prvog tjedna tova metodom slučajnog uzorka uzeto je i obilježeno po 10 pilića iz svake skupine. Jedinke iz sve tri skupine su cijepljene prema utvrđenom programu imunoprofilakse. Smještaj pilića te oprema za hranjenje i na-pajanje, provjetravanje i rasvjetu bili su primjereni podnom sustavu držanja usklađeni prema tehnologiji, ali prilagođeni pokusnim uvjetima držanja. Za ishranu su korištene kompletne krmne smjese (starter, grover i finišer) u skladu s normativno-nutritivnim potrebama za tovne piliće linije Cobb. Pilići su hranu i vodu dobivali ad libitum. Tijekom pokusa kod obilježenih pilića tjedno su praćeni i mjereni opseg prsa, opseg batka, duljina kobilice, dubina prsa i prsni kut te je mjerena živa tjelesna masa. Na kraju tova sva mjerenja provedena na živim jedinkama provedena su i na njihovim trupovima, kao i prosječna masa trupa, randman te udjeli jestivih dijelova, težine jetre i abdominalne masti u odnosu na masu trupa. Na osnovi dobivenih rezultata izvršena je statistička obrada podataka analizom varijance (ANOVA). Ako se vrijednost F pokazala značajnom (p<0,05), razlike srednjih vrijednosti između skupina testirane su Duncanovim testom.
Rezultati i rasprava
Najbolje vrijednosti konformacije živih pilića te njihovih trupova imali su pilići kontrolne skupine (tablica 1.). Utvr-đene razlike ovih pokazatelja u odnosu na pokusne skupine
nisu bile statistički značajne osim kod trupova za opseg prsa, duljinu kobilice, dubinu prsa i prsni kut (p<0,05). Komfor-macija predstavlja tjelesni oblik, skeletni sklop ili građu, odnosno u širem smislu, tjelesnu formu koja obuhvaća i mesnatost te neznatnu količinu i raspored masnog, naročito potkožnog tkiva. Kod živih jedinki najvažniji utjecaj ima u uzgojno-selekcijskom radu, a na trupovima u ocjenjivanju kvalitete mesa (Pavlovski i sur., 2006.). Najveće vrijednosti za opseg prsa, opseg batka i duljinu kobilice, dubinu prsa i prsni kut u skupini od 15 pilića/m2 utvrdili su na živim pilićima (Softić i sur., 2010.), što je u skladu s rezultatima dobivenim tijekom naših istraživanja. Ojedapo i sur. (2016.) izvršili su evaluaciju parametara potrebnih za opis rasta kod dvije komercijalne linije tovnih pilića i ustanovili da su pilići linije Cobb postigli bolju tjelesnu konformaciju u odnosu na liniju Marshall. Utvrđene vrijednosti za opseg prsa i duljinu kobilice u našem pokusu bile su veće, a za opseg batka iste. Moramo naglasiti da su spomenuti autori imali različit pokusni dizajn u odnosu na naš. Povoljniju konformaciju iskazanu kroz indekse imali su također pilići kontrolne skupine (p<0,05) osim za opseg batka i nešto veće vrijednosti za dubinu prsa i duljinu kobilice od rezultata koje su objavili Pavlovski i sur. (2006.), s tim što su spomenuti autori ispitivali utjecaj nekoliko genotipova u različitim vremenskim razdobljima i utvrdili značajan doprinos u povećanju tjelesne mase pilića i udjela prsa, a samim tim i većem prsnom kutu.
Tablica 1. Konformacija tovnih pilića Table 1. Body conformation of broilers
Pokazatelj - Parameter Živi pilići - Live chicken
Skupina A - A group Skupina B - B group Skupina K - K group Opseg prsa (mm) - Breast circumference (mm) 322,60 315,40 324,10 Duljina kobilice (mm) - Keel length (mm) 111,20 108,10 113,00 Dubina prsa (mm) - Breast depth (mm) 102,40 101,40 104,00 Opseg batka (mm) - Drumstick circumference (mm) 139,40 137,00 141,30 Prsni kut (stupnjevi) - Breast angle (degrees) 102,80 105,80 107,00
Pokazatelj - Parameter Trupovi pilića - Chicken carcasses
Skupina A - A group Skupina B - B group Skupina K - K group Opseg prsa (mm) - Breast circumference (mm) 310,40a 301,50b 313,20a Duljina kobilice (mm) - Keel length (mm) 108,30a 109,10b 115,00a Indeks (g/mm) - Index (g/mm) 21,54a 21,23ab 20,40b Dubina prsa (mm) - Breast depth (mm) 108,00a 103,00ab 100,30b Indeks (g/mm) - Index (g/mm) 21,66a 22,75ab 23,54b Opseg batka (mm) - Drumstick circumference (mm) 143,00 145,30 146,80 Indeks (g/mm) - Index (g/mm) 16,31 15,96 16,35 Prsni kut (stupnjevi) - Breast angle (degrees) 109,60a 117,60b 125,70b
a,bvrijednosti u istom redu s različitim superskriptom razlikuju se značajno a,bvalues in the same row with different superscripts differed significantly

UČINAK GUSTOĆE NASELJENOSTI NA KONFORMACIJU TOVNIH PILIĆA 137
Na kraju tova 42. dana pilići kontrolne skupine ostvarili su bolju prosječnu tjelesnu masu, masu trupa te težinu jetre i abdominalne masti (tablica 2.). Prosječna tjelesna masa pilića kontrolne skupine iznosila je 2.345,45 g, što je za 0,60% odnosno 1,21% više u odnosu na piliće pokusnih skupina. Veće vrijednosti randmana mesa (slika 1.) utvrđene su kod pilića kontrolne skupine. S obzirom na to da su pilići kontrolne skupine ostvarili nešto veću, ali ne i značajno veću prosječnu tjelesnu masu na kraju tova od 42 dana u skladu je s rezultatima Softić i sur. (2013.) koji su ispitivali utjecaj gustoće naseljenosti na kvalitetu trupova tovnih pilića linije Cobb, kao i Petričević i sur. (2011.) ispitujući utjecaj genotipa na proizvodne i klaoničke karakteristike brojlerskih pilića. Uspoređujući utjecaj gustoće naseljenosti i genotipa na proizvodne pokazatelje muških i ženskih brojlerskih jedinki Siaga i sur. (2017.) utrvrdili su tjelesnu masu od 2.346,0 g pri gustoći naseljenosti od 30 kg/m2, što je u skladu s našim rezultatima kontrolne skupine (15 pilića/m2).
Iz slike 1. vidljivo je da je utvrđeni randman mesa također veći kod pilića kontrolne skupine u odnosu na pokusne skupine, što je suglasno s rezultatima Softić i sur. (2013.), a manji u odnosu na rezultate koje su objavili Pasca i sur. 2008. (88,9%), Beg i sur. 2011. (74,17%), Adeyemo i sur. 2016. (78,53%), Siaga i sur. 2017. (82,70%) i Henrique i sur. 2017. (74,56%). Veći, ali ne i statistički značajno veći udio ispitivanih jesti-vih dijelova u odnosu na završnu masu trupa utvrđen je kod pilića kontrolne skupine (tablica 3.). Udio prsa, batka s karabatkom i abdominalne masti u odnosu na prosječnu masu trupa se nisu značajno razlikovali među tovnim pilićima u odnosu na gustoću naseljenosti i u skladu su s dobivenim rezultatima Softić i sur. (2013.) za kontrolnu skupinu pilića. U našem pokusu dobivene vrijednosti udjela prsa su bile veće u odnosu na rezultate koje su objavili Siaga i sur. (2017.), a za udio vrata i leđa od rezultata Zenunovića i sur. (2018.).
Tablica 2. Završna tjelesna masa, masa trupa, težina jetre i abdominalna mast kod ispitivanih tovnih pilića Table 2. Final body weight, carcass weight, liver weight and abdominal fat in study broilers
Pokazatelj - Parameter Skupina A - A group Skupina B - B group Skupina K - K group Završna tjelesna masa (g) - Final body weight (g) 2.331,96 2.317,06 2.345,45 Masa trupa (g) - Carcass weight (g) 1.659,80 1.628,28 1.697,69 Težina jetre (g) - Liver weight (g) 53,12 52,91 55,23 Težina abdominalne masti (g) - Weight of abdominal fat (g) 17,68 16,57 19,16
Figure 1. Randman mesa (%) kod pilića nakon 42 dana tova. Slika 1. Dressing percentage (%) of chickens after 42 days of fattening. Tablica 3. Udio jestivih dijelova, jetre i abdominalne masti u odnosu na masu trupa Table 3. Share of edible portions, liver and abdominal fat relative to carcass mass
Pokazatelj - Parameter A skupina - A group B skupina - B group K skupina - K group Prsa (%) - Breast (%) 30,13 31,19 30,43 Batak sa zabatkom (%) - Drumstick with thigh (%) 22,41 22,15 22,61 Krila (%) - Wings (%) 9,91 10,30 10,26 Leđa (%) - Back (%) 16,89 17,49 17,21 Vrat (%) - Neck (%) 5,79 6,01 5,91 Jetra (%) - Liver (%) 3,22 3,25 3,26 Abdominalna mast (%) - Abdominal fat (%) 1,08 1,03 1,13

UČINAK GUSTOĆE NASELJENOSTI NA KONFORMACIJU TOVNIH PILIĆA 138
Madilindi i sur. (2018.) u svom istraživanju pri gustoći naseljenosti od 30 kg/m2 su utvrdili niže vrijednosti udjela prsa, leđa, vrata i krila osim abdominalne masti u usporedbi s našim rezultatima, ali moramo naglasiti da su spomenuti autori uz gustoću naseljenosti u suptropskim uvjetima ispitivali i utjecaj spola. Ostvareni udio jetre u našem istraživanju je bio veći u odnosu na rezultate koje navode Madilindi i sur. (2018.), Adeyemo i sur. (2016.), a u skladu su s rezultatima koje navode Siaga i sur. (2017).
Zaključci
Na osnovu rezultata istraživanja konformacije tovnih pilića linije Cobb u ovisnosti od gustoće naseljenosti mogu se izvesti sljedeći zaključci: - Kod trupova kontrolne skupine (naseljenost 15 pilića/m2)
utvrđeni su statistički značajno veći opseg prsa, prsni kut, duljina kobilice i dubina prsa (p<0,05) u odnosu na pokusne skupine. Povoljniju konformaciju iskazanu kroz indekse osim za opseg batka ostvarili su pilići kontrolne skupine (p<0,05).
- Jedinke kontrolne skupine ostvarile su bolje rezultate završne tjelesne mase, mase trupa i randmana u odnosu na pokusne skupine pilića bez statističke značajnosti razlika.
Literatura
ADEYEMO G. O., O. O. FASHOLA, T. I. ADEMULEGUN (2016): Effect of Stocking Density on the Performance, Carcass Yield and Meat Composition of Broiler Chickens. British Biotechnology Journal, 14 (1), 1-7.
BEG M. A. H., M. A. BAQUI, N. R. SARKER, M. M. HOSSAIN (2011): Effect of Stocking Density and Feeding Regime on Performance of Broiler Chicken in Summer Season. International Journal of Poultry Science, 10 (5), 365-375.
HENRIQUE CLEONEIDE DOS SANTOS, ANDRÉIA FRÓES GALUCI OLIVEIRA, THALES SILVA FERREIRA, EDUARDO SANTOS SILVA, BRUNA FINOTTI FONSECA REIS DE MELLO, AGNER DE FREITAS ANDRADE, VINÍCIUS DA SILVA FREITAS MARTINS, FERNANDO OLIVEIRA DE PAULA, ELIS REGINA DE MORAES GARCIA, LUÍS DANIEL GIUSTI BRUNO (2017): Effect of stocking density on performance, carcass yield, productivity, and bone development in broiler chickens Cobb 500®. Semina: Ciencias Agraris, Londrina, 38 (4), 1, 2705-2718.
MADILINDI M. A., A. MOKOBANE, P. B. LETWABA, T. S. TSHILATE, C. B. BANGA, M. D. RAMBAU, E. BHEBHE, K. BENYI (2018): Effects of sex and stocking density on the performance of broiler chickens in a sub-tropical environment. South African Journal of Animal Science, 48 (3), 459-468.
NIKOLOVA N., Z. PAVLOVSKI (2009): Major carcass parts of broiler chicken from different genotype, sex, age and nutrition
system. Biotechnology in Animal Husbandry, 25, 5-6, 1045-1054.
OJEDAPO, L. O., O. A. KINOKUN, T. A. ADEDEJI, T. B. OLAYENI, S. A. AMEEN, S. R. AMAO (2008): Effect of strain and sex on carcass characteristics of those commercial broilers reared in deep litter system in the derived savannah area of Nigeria. World Journal of Agricultural Sciences, 4 (4), 487-491.
PASCA I., T. OROIAN, R. OROIAN, DANA PUSTA, R. MORAR, S. PASCALĂU, M. BALLA, I. DALEA, C. BAGITA (2008): The phenotipical performances of Ross 308 and Cobb 500, exploited in the north western part of Romania. Bulletin UASVM, Veterinary Medicine, 65 (1), 340-345.
PAVLOVSKI Z, M. LUKIĆ, R. CMILJANIĆ, Z. ŠKRBIĆ (2006): Konformacija trupova pilića. Biotechnology in Animal Husbandry, 22 (3-4), 83-96.
QAID M. I., H. I. ALBATSHAN, T. I. SHAFEY, E. I. HUSSEIN, A. M. ABUDABOS (2016): Effect of Stocking Density on the Performance and Immunity of 1- to 14-d- Old Broiler Chicks. Brazilian Journal of Poultry Science Revista Brasileira de Ciência Avícola. 18 (4), 683-692.
SALIHBAŠIĆ E., M. BAŠIĆ, A. ZENUNOVIĆ (2014): Eko nomičnost tova pilića u zavisnosti od gustoće naseljenosti economics of chickens fattening depending on stocking density. Tranzicija / Transition, XVI (33), 95-105.
SIAGA R., J. J. BALOYI, M. D. RAMBAU, K. BENYI (2017): Effects of Stocking Density and Genotype on the Growth Performance of Male and Female Broiler Chickens. Asian J Poult.Sci., 11 (2), 96-104.
SOFTIĆ A., A. GAGIĆ, V. KATICA, A. KAVAZOVIĆ, V. ŠA-KIĆ, R. ĆUTUK, M. VARATANOVIĆ (2010): Utjecaj gusto-će naseljenosti kao faktora tehnologije proizvodnje na tjelesnu konformaciju brojlerskih pilića. Vetrinaria, 59 (1-4), 11-19.
SOFTIĆ A., A. KAVAZOVIĆ, V. KATICA, V. ŠAKIĆ, A. KATICA, N. MLAĆO, M. VARATANOVIĆ (2013): Influence of stocking density on carcass quality of broilers. Proceedings of the 10th International Symposium Modern Trends in Livestock Production. October 2-4, 2013., Belgrade, Serbia, 761-768.
ŠKRBIĆ Z, Z. PAVLOVSKI, M. LUKIĆ (2008): Efekat gustine naseljenosti na pojedine klanične osobine brojlera genotipa Cobb. Biotechnology in Animal Husbandry, 24 (1-2), 51-58.
ŠKRBIĆ, Z, Z. PAVLOVSKI, M. LUKIĆ, V. PETRIČEVIĆ, M. DUKIĆ-STOJČIĆ, D. ŽIKIĆ (2011): The effect of stocking density on individual broiler welfare parameters: 2. Different broiler stocking densities. Biotechnol. Anim. Husb., 27, 17-24.
TONG H. B., J. LU, J. M. ZOU, Q. WANG, S. R. SHI (2012): Effects of stocking density on growth performance, carcass yield, and immune status of a local chicken breed. Poultry Science, 91 (3), 667–673.
ZENUNOVIĆ A., M. GLAVIĆ (2018): Effect of stocking density and gender of chickens in the share of certain parts of body and kalo cut of fattening chicken. International Journal of Advances in Agricultural Science and Technology, 5 (8), 83-94.

UČINAK GUSTOĆE NASELJENOSTI NA KONFORMACIJU TOVNIH PILIĆA 139
EFFECT OF STOCKING DENSITY ON BODY CONFORMATION OF BROILERS
Summary
The aim of the research was to determine the effect of different population density on the conformation and quality of carcasses of fattening Cobb chickens. In the experiment, a total of 120 one-day chickens were divided into three groups (A, B and K) for up to six weeks. Group A was a group of chickens with a low population density (12 chickens/m2), group B was a group with a higher population density (18 chickens/m2) and control group K (15 chickens/m2) in accordance with technological standards for the given broiler line. At the end of the first week, a random sample was taken and 10 chickens were marked. During experiments with the marked chicks, the measured circumference breast, thigh girth, keel length, breast depth and breast angle were measured weekly, and body mass was measured. At the end of the test, all measurements carried out on live chickens were carried out on their carcasses, along with average weight of carcass, meat yield, and the proportion of edible parts, liver weight and abdominal fat compared to the weight of the carcass. The best values for the conformation of live chickens and their carcasses were recorded in control group. Differences in these indicators in relation to the experimental groups were not statistically significant, except for the carcasses of chickens for breast circumference, breast angle, keel length and breast depth (p<0.05). In addition, a higher but not statistically significantly higher proportion of examined edible parts in final carcass weight was found in the control group of chickens with stocking density that was in accordance with the technological recommendations. Favorable conformation expressed through indices was also recorded in control group chickens (p<0.05) except for thigh girth. Key words: stocking density, fattening chickens, carcass quality, conformation

PERADARSKI DANI 2019. 140
PRISUTNOST RODA CAMPYLOBACTER U KOŽAMA VRATOVA TOVNIH PILIĆA TIJEKOM LJETNOG RAZDOBLJA 2018. GODINE
Sanja Furmeg, Vesna Jaki Tkalec, Maja Kiš, Jadranka Sokolović, Darko Majnarić, Željko Cvetnić
Hrvatski veterinarski institut, Veterinarski zavod Križevci, Križevci, Hrvatska
Sažetak
Bakterije roda Campylobacter uzrokuju bolest kampilobakteriozu, zoonozu koja se na čovjeka prenosi sa životinja i proizvoda životinjskog podrijetla. Najčešće infekcije kod ljudi izazivaju Campylobacter jejuni i Campylobacter coli. Campylobacter spp. je široko rasprostranjen, možemo ga naći u svim vrstama mesa i proizvodima od mesa, nepasteriziranom mlijeku i proizvodima od nepasteriziranog mlijeka, rjeđe u jajima, ali i u ostaloj hrani ako je došlo do križne kontaminacije te u kontaminiranoj vodi. U posljednjih desetak godina Campylobacter spp. je najčešći uzročnik bakterijskog trovanja hranom kod ljudi u Europskoj Uniji i u SAD-u. Kampilobakterioza je učestalija ljeti, jer više temperature pogoduju rastu uzročnika ove bolesti. Tijekom ljetnog razdoblja 2018. godine kroz 4 tjedna sakupljeno je 60 uzoraka kože vratova tovnih pilića iz tri klaonice koji su pretraživani na bakterije iz roda Campylobacter. Dobiveni rezultati pokazuju da od ukupnog broja pretraženih uzoraka, 31 (52%) uzorak je bio nesukladan prema Uredbi Komisije (EU) 2017/1495 (MDK <1000 cfu/g). Uzorci su obrađeni prema standardnoj metodi za dokazivanje prisutnosti i određivanje broja Campylobacter spp. (HRN EN ISO 10272-2:2017). Zbog osiguranja zdravstvene ispravnosti hrane, a s obzirom na patogenost bakterija iz roda Campylobacter, od iznimnog su značenja termička obrada hrane, dezinfekcija vode, sprječavanje križne kontaminacije te higijena prostora u kojem se hrana priprema. Ključne riječi: Campylobacter spp., kože vratova tovnih pilića, kontaminacija, dobra higijenska praksa Uvod
Bakterije roda Campylobacter uzročnici su kampilobak-terioze, zoonoze koja se na čovjeka prenosi sa životinja i proizvoda životinjskog podrijetla. To je bolest koja predstavlja najčešće prijavljivanu bolest uzrokovanu hranom u Europskoj Uniji te predstavlja značajan problem javnog zdravstva širom svijeta (EFSA i ECDC, 2018.). Uzročnici ove bolesti su kampilobakteri, zakrivljeni gram negativni mikroorganizmi dužine 2-5 µm, promjera 0,3 µm, zavijeni poput zareza, nalik galebovim krilima. Ne stvaraju spore, posjeduju po jedan bič na jednom ili oba pola, što im omogućuje veliku pokretljivost (Cvetnić i sur., 2013.). Kampilobakteri su mikroaerofilne bakterije, što znači da im za rast pogoduju uvjeti sa smanjenom količinom kisika, odnosno atmosferski uvjeti s otprilike 10% CO2, 5% O2 i 85% N2, što ih razlikuje od ostalih uzročnika koji se prenose hranom (Mikulić i sur., 2017.). S obzirom na termofilnost
najbolje rastu na temperaturi inkubacije od 42 °C, što je vjerojatno posljedica prilagodbe njihovom normalnom sta-ništu, probavnom sustavu ptica (Vučković i Abram, 2009.). Najčešćim uzročnicima kampilobakterioze u ljudi smatraju se pripadnici termotolerantnih kampilobaktera i to Cam-pylobacter jejuni, Campylobacter coli te Campylobacter lari i Campylobacter upsaliensis (Mikulić i sur., 2016.). Brojna istraživanja pokazala su da je C. jejuni vodeći uzročnik bakterijskih crijevnih infekcija. U posljednjih nekoliko godina kampilobakterioza zauzima vodeće mjesto bakterijskog trovanja hranom (EFSA i ECDC, 2018.). Kampilobakteri su sveprisutni mikroorganizmi te ih nala-zimo u mesu peradi i domaćih životinja, mesnim i mliječnim prerađevinama, sirovom mlijeku, pitkoj vodi. Također, možemo ih naći u potocima, rijekama i vodama stajaćicama, što je pokazatelj onečišćenja izmetom divljih ptica ili stoke. Ipak, najvažnijim izvorom ovog patogena smatra se meso peradi, nedovoljno termički obrađeno meso te gotova hrana
Sanja Furmeg, dipl.sanit.ing, Hrvatski veterinarski institut, Veterinarski zavod Križevci, Laboratorij za mikrobiologiju hrane i hrane za životinje, Ivana Zakmardija Dijankovečkog 10, 48260 Križevci; Tel: +385 48 279 269; Fax: + 385 48 279 255; e-mail: [email protected]

PRISUTNOST RODA CAMPYLOBACTER U KOŽAMA VRATOVA TOVNIH PILIĆA TIJEKOM LJETNOG RAZDOBLJA 2018. GODINE
141
koja je bila u kontaktu sa sirovim mesom peradi. Kam-pilobakteri se nalaze u crijevu peradi, a prilikom klaoničke obrade onečiste pileće meso, preživljavaju tijekom lanca proizvodnje te tako postaju rizik za ljudsko zdravlje (Levak, 2015., Mikulić i sur., 2017.). Kampilobakterioza se kod ljudi manifestira proljevom s jakim bolovima u trbuhu. Bolest traje sedam do deset dana, a može biti popraćena povišenom tjelesnom temperaturom, grčevima u crijevima, mučninama, a ponekad i povra-ćanjem. Teža klinička slika može se javiti kod dojenčadi, male djece, starijih te odraslih osoba koje boluju od teških kroničnih bolesti, kao i u osoba s oslabljenim imunitetom. Moguće su pojave komplikacija kao što su bakterijemija (prodor bakterija u krv), hepatitis (upala jetre), reaktivni artritis, te Guillian-Barreov sindrom. Iako svatko može biti zaražen Campylobacter spp., podaci o oboljelima upućuju na to da najviše obolijevaju djeca mlađa od pet godina te mladi u dobi od 15 do 29 godina (Marinculić i sur., 2009.). Kampilobakterioza se može javiti u pojedinačnim slu-čajevima ili manjim obiteljskim epidemijama. Glavnu opasnost u domaćinstvu predstavlja križno onečišćenje svježe i hrane spremne za potrošnju kako izravno tako i neizravno preko ruku, radnih površina, kuhinjska pomagala, kao i nedovoljno toplinski obrađena onečišćena svježa hrana (Mikulić i sur., 2016a.). Također, kampilobakterioza je učestalija ljeti, jer više temperature pogoduju rastu uzročinka ove bolesti (Patrick i sur., 2004.). S obzirom na prilagođenost bakterija roda Campylobacter, one u probavnom traktu životinja iznimno rijetko uzrokuju bolest, ali zato otežano preživljavaju u hrani te se u njoj ne mogu umnažati (Joshua i sur., 2006.). Preživljavanje C. jejuni u aerobnim uvjetima pripisuje se metaboličkom komenzalizmu s bakterijama roda Pseudomonas i stvaranju zaštitnog biofilima (Hilbert i sur., 2010.). Campylobacter spp. pokazuje osjetljivost na temperature kuhanja, pasteriza-cije, sušenja, niski pH i koncentracije soli. Također, osjetljiv je na uobičajene dezinficijense, kloriranje vode te postupak smrzavanja koji može umanjiti njegovu koncentraciju u određenoj namirnici (Marinculić i sur., 2009.). S obzirom na provođenje postupaka klanja i obrade mesa peradi, velika mogućnost kontaminacije javlja se u kasnijim fazama proizvodnje kao što je čupanje perja, evisceracija, pranje, pohranjivanje, rashlađivanje i zamrzavanje te kon-taminacija putem ruku radnika, opreme ili same okoline, stoga je od iznimnog značenja provedba higijensko-sani-tarnih mjera te pravilno čišćenje i dezinfekcija objekata (Levak, 2015.). Koža peradi predstavlja jedan od glavnih izvora ovog patogena (Musgrove i sur., 1997.), a ako je koža konta-minirana, vjerojatnost da je i meso kontaminirano je 35 puta veća u odnosu na kožu koja nije kontaminirana (Jeffrey i sur., 2001.). S obzirom na to da je kampilobakterioza vodeća zoonoza, a koža vratova peradi je zbog načina klaoničke obrade potencijalno najveći izvor Campylobacter spp., cilj ovog istraživanja bio je odrediti učestalost roda Campylo-bacter u kožama vratova tovnih pilića.
Materijali i metode
Tijekom ljetnog razdoblja 2018. godine kroz 4 tjedna sa-kupljeno je 60 uzoraka koža vratova tovnih pilića iz tri klaonice. Kože vratova tovnih pilića uzimane su slučajnim odabirom s linije klanja nakon faze evisceracije i rashla-đivanja. Uzorci su uzorkovani u različite radne dane, istog su dana dostavljeni u laboratorij u prijenosnom hladnjaku u kontroliranim uvjetima uz održavanje hladnog lanca. Uzorci su istog dana stavljeni u obradu. Uzorkovanje kože vratova tovnih pilića provedeno je prema standardnom postupku uzimanja uzoraka s trupova (HRN EN ISO 17604:2015). Za izdvajanje bakterija korištena je standardna metoda za dokazivanje prisutnosti i određivanje broja Campylobacter spp. (HRN EN ISO 10272-2:2017). Za rast kampilobaktera korištena je selektivna podloga Campylobacter blood – free selective agar base (mCCDa, Oxoid LtD, Basingstoke, UK) na koju je inokuliran 0,1 mL predobogaćenog bujona s uzorkom. Tako nacijepljene ploče inkubirane su u mikro-aerofilnim uvjetima pri 41,5 °C tijekom 48 h. Tipične kolonije na mCCD agaru su sivkaste, ponekad metalnog sjaja, plosnate i vlažne, često konfluirajuće, no prepo-znavanje kolonija Campylobacter spp. u velikoj mjeri je stvar iskustva, jer njihov izgled može varirati. Porasle sumnjive kolonije nacijepljene su na krvni agar koji je inkubiran u aerobnim uvjetima na temperaturi od 25 °C tijekom 48 h. Izostanak rasta kulture Campylobacter spp. na krvnom agaru jedan je od potvrdnih pokazatelja prisutnosti Campylobacter spp. u uzorku. Zbog brzog propadanja Campylobacter spp. u aerobnim uvjetima postupci konfir-macije moraju se izvršiti u što kraćem vremenu. Tijekom potvrde korišteni su testovi oksidaze i katalaze te mikro-skopski pregled nativnog preparata s pokrovnicom na kojem se jasno vidi pokretljivost bakterija roda Campylobacter.
Rezultati i rasprava
Dobiveni rezultati potvrđuju prisutnost Campylobacter spp. u kožama vratova tovnih pilića. Od ukupno 60 pretraženih uzoraka kože vratova tovnih pilića u klaonicama peradi njih 31 (52%) nije bilo sukladno prema Uredbi Komisije (EU) 2017/1495 gdje je maksimalno dozvoljena koncentracija (MDK) <1000 cfu/g. Campylobacter spp. određen je na temelju izgleda rasta na selektivnom agaru, pozitivnih te-stova oksidaze i katalaze, mikroskopske pretrage te izo-stanka rasta na krvnom agaru u aerobnim uvjetima na 25 °C. Dobiveni rezultati u ovom istraživanju veoma su slični rezultatima Hadžiabdića i sur. (2013.) koji su prisutnost bakterija roda Campylobacter spp. utvrdili u 58,1% pilećih trupova i koža na liniji klanja u Bosni i Hercegovini. Koža peradi predstavlja jedan od glavnih izvora Campylobacter spp. upravo zbog slijevanja sadržaja s površine trupova te se pretpostavlja da je to područje trupa peradi najčešće kontaminirano bakterijama. Kao mjera smanjenja broja bakterija u klaonicama se redovito koriste šurionici u kojima temperatura vode iznosi 72 °C (Zglavnik i Sokolović, 2009.). Nakon šurenja trupovi prolaze kroz aparat za linijsko čupanje perja te potom slijedi

PRISUTNOST RODA CAMPYLOBACTER U KOŽAMA VRATOVA TOVNIH PILIĆA TIJEKOM LJETNOG RAZDOBLJA 2018. GODINE
142
evisceracija prilikom koje može doći do puknuća crijeva. Prilikom puknuća crijeva najčešće dolazi do kontaminacije trupova peradi i kože. U klaoničkim objektima od iznimne je važnosti pridržavanje higijensko-sanitarnih uvjeta, redovito čišćenje i dezinfekcija opreme i pribora u cilju smanjenja pojavljivanja bakterija iz roda Campylobacter na mesu peradi (Zglavnik i Sokolović, 2009., Levak, 2015.). Na slici 1. prikazan je raspon vrijednosti kod pozitivnih uzoraka koji se kretao od 1090 cfu/g sve do 5273 cfu/g. Također, u 11 uzoraka utvrđena je prisutnost Campylobacter spp., no vrijednosti su bile manje od dozvoljenih prema Uredbi 2017/1495. Raspon dobivenih rezultata kretao se od 182 cfu/g do 909 cfu/g. Prema istraživanjima Robinsona (1981.), infektivna doza za C. jejuni iznosi 500 cfu/g, dok su Black i sur. (1988.) na zdravim dobrovoljcima potvrdili infektivnu dozu od 800 cfu/g. C. jejuni je najčešće izolirana vrsta iz mesa peradi. Dobivene vrijednosti sukladnih uzoraka nisu zanemarive uzimajući u obzir infektivnu dozu koja također ovisi o fizičkom statusu i dobi bolesnika. U ovom istraživanju u 18 uzoraka nije potvrđena prisutnost Campylobacter spp.
Prema najnovijim podacima Europske agencije za sigurnost hrane (EFSA) i Europskog centra za sprječavanje i kontrolu bolesti (ECDC), humana alimentarna kampilobakterioza je najzastupljenija zoonoza još od 2005. godine. Broj prijav-ljenih slučajeva humane kampilobakterioze na području Europe u 2017. iznosio je 246.158. U posljednjih pet godina ipak se bilježi usporavanje rasta broja oboljelih te se u 2017. godini bilježi pad u broju oboljelih od kampilobakterioze (64,8 slučajeva na 100.000 stanovnika), što je neznatno smanjenje u odnosu na 2016. (66,3 slučajeva na 100.000 stanovnika). Najviše stope izvještavanja za 2017. godinu zabilježene su u Češkoj (230,0 slučajeva na 100.000), Slovačkoj (127,8), Švedskoj (106,1) i Luksemburgu (103,8). Najniže stope u 2017. zabilježene su u Bugarskoj, Cipru, Latviji, Poljskoj, Portugalu i Rumunjskoj (≤5,8 na 100.000). Od ukupnog broja potvrđenih kampilobakterioza, C. jejuni je dokazan u 84,4% slučajeva, C. coli u 9,2%, C. lari u 0,1%, a C. upsaliensis u 0,1%, slučajeva. U Hrvatskoj je ukupno zabilježeno 1.686 slučajeva oboljelih od kampilo-bakterioze, čime se bilježi lagani porast broja oboljelih iz godine u godinu (EFSA i ECDC, 2018.).
Slika 1. Prikaz vrijednosti nesukladnih uzoraka na broj Campylobacter spp. prema Uredbi Komisije (EU) 2017/1495 u pogledu
roda Campylobacter u trupovima brojlera (MDK <1000 cfu/g). Figure 1. Results of unsuitable samples on the number of Campylobacter spp. according to Commission Regulation (EU)
2017/1495 as regards Campylobacter in broiler carcasses (limit of 1000 cfu/g).

PRISUTNOST RODA CAMPYLOBACTER U KOŽAMA VRATOVA TOVNIH PILIĆA TIJEKOM LJETNOG RAZDOBLJA 2018. GODINE
143
U kategoriji hrane, najveća prisutnost zabilježena je u svježem mesu iz brojlera (37,4%), a slijedi ga svježe meso purana (31,5%). Španjolska je jedina zemlja koja je dostavila podatke za kriterij higijene procesa prema Uredbi 2017/1495. U Španjolskoj je ukupno pretraženo 150 uzoraka kože vratova iz ohlađenih trupova broilera, od kojih je 66 (44%) bilo nesukladno, 53 (84%) u rasponu između 1000 i 10.000 cfu/g, dok je 13 uzoraka imalo više od 10.000 cfu/g (EFSA i ECDC, 2018.). Rezultati našeg istraživanja prema broju nesukladnih uzoraka koža vratova vrlo su slični rezultatima istraživanja u Španjolskoj. U ovom istraživanju nije zabilježen nijedan slučaj u kojem je vrijednost bila iznad 10.000 cfu/g.
Zaključak
Kampilobakterioza predstavlja važan javnozdravstveni problem, a zauzima vodeće mjesto prema broju oboljelih u Europskoj Uniji kao najčešća bolest koja se prenosi hranom. Dosadašnja istraživanja ukazuju na moguću visoku pojavnost bakterija iz roda Campylobacter tijekom različitih faza proizvodnje mesa peradi. Rezultati ovog istraživanja potvrđuju prisutnost Campylo-bacter spp. u 52% uzoraka kože vratova tovnih pilića. Dobiveni rezultati se slažu s prethodno provedenim istra-živanjima na području Europe i pokazuju visoku pojavnost Campylobacter spp. S obzirom na to da je broj slučajeva kampilobakterioze u Hrvatskoj u stalnom porastu, od iz-nimne je važnosti kontrola prisutnosti Campylobacter spp. u peradarskoj proizvodnji. Provođenje biosigurnosnih mjera i praćenje prevalencije kampilobaktera nužni su u klao-ničkim objektima kako bi se smanjila učestalost pojave ove zoonoze.
Literatura
BLACK, R. E., M. M. LEVINE, M. L. CLEMENTS, T. P. HUGHES, M. J. BLASER (1988): Experimental Campylobacter jejuni infection in humans. J. Infect. Dis., 157, 472-479.
CVETNIĆ, Ž., A. KVESIĆ (2013): Specijalna bakteriologija. U: Mikrobiologija i parazitologija., Mostar, 33-98.
EFSA i ECDC (2018): The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA Journal 2018, 16 (12), 5500, 10-22.
HILBERT, F., M. SCHERWITZEL, P. PAULSEN, M. P. SZOSTAK (2010): Survival of Campylobacter jejuni under conditions of atmospheric oxygen tension with the support of Pseudomonas spp. Appl. Environ. Microbiol., 76, 5911-5917.
HRN EN ISO 10272 – 2:2017 – Mikrobiologija u lancu hrane – Horizontalna metoda za dokazivanje prisutnosti i određivanje broja Campylobacter spp. – 2. Dio: Postupak određivanja broja kolonija (izvornik: International Organization for Standardization, ISO 10272 – 2:2017; EN ISO 10272 – 2:2017)
HRN EN ISO 17604 – 2015 – Mikrobiologija lanca hrane - Uzorkovanje životinjskih trupova za mikrobiološku analizu (ISO 17604:2015; EN ISO 17604:2015)
JEFFREY, J. S., K. H. TONOOKA, J. LOZANO (2001): Prevalence of Campylobacter spp. from skin, crop and intestine of commercial broiler chicken carcasses at processing. Poultry Sci., 80, 1390-1392.
JOSHUA, G. W. P., C. GUTHRIE – IRONS, A. V. KARLYSHEV, B. W. WREN (2006): Biofilm formation in Campylobacter jejuni. Microbiology, 152, 387-396.
LEVAK STEFANI (2015): Campylobacter spp. u mesu peradi. Meso XVII (6), 558-565.
MARINCULIĆ, A., B. HABRUN, LJ. BARBIĆ, R. BECK (2009): Biološke opasnosti u hrani. Hrvatska agencija za hranu, Osijek.
MIKULIĆ MARINA, ANDREA HUMSKI, B. NJARI, DORA STOJEVIĆ, L. JURINOVIĆ, S. ŠPIČIĆ, SANJA DUVNJAK, Ž. CVETNIĆ (2017): Metode izdvajanja i dokazivanja bakterija roda Campylobacter – klasične i molekularne metode (I. dio). Veterinarska stanica, 48 (4), 297 – 303.
MIKULIĆ, MARINA, ANDREA HUMSKI, B. NJARI, M. OSTOVIĆ, Ž. CVETNIĆ (2016): Termotolerantni Campylobacter spp. – uzročnici kampilobakterioze (I. dio). Veterinarska stanica, 47 (4), 327-334.
MIKULIĆ, MARINA, ANDREA HUMSKI, B. NJARI, M. OSTOVIĆ, Ž. CVETNIĆ (2016a): Termotolerantni Campylobacter spp. – uzročnici kampilobakterioze (II. dio). Veterinarska stanica, 47 (5), 447-454.
MUSGROVE, M. T., J. A. CASON, D. L. FLETCHER, N. J. STERN, N. A. COX, J. BAILEY (1997): Effect of cloacal plugging on microbial recovery from Partially processed broilers. Poultry Sci., 76, 530-533.
PATRICK, M. E., L. E. CHRISTIANSEN, M. WAINO, S. ETHELBERG, H. MADSEN, H. C. WEGENER (2004): Effects of climate on incidence of Campylobacter spp. in humans and prevalence in broiler flocks in Denmark. Appl. Environ. Microbiol., 70, 7474-7480.
ROBINSON, D. A. (1981): Infective dose of Campylobacter jejuni in milk. Br. Med. J., 282, 1584.
Uredba komisije (EU) 2017/1495 o izmjeni Uredbe (EZ) br. 2073/2005 u pogledu Campylobactera u trupovima brojlera. https://eur-lex.europa.eu/legal-content/HR/TXT/PDF/?uri=CELEX:32017R1495&from=EN. Pristupljeno 15.01.2019.
VUČKOVIĆ DARINKA, MAJA ABRAM (2009): Kampilobakteri – najčešći uzročnik bakterijskog proljeva u ljudi širom svijeta. Medicina, 45 (4), 344-350.
ZGLAVNIK T., MARIJANA SOKOLOVIĆ (2009): Pojavnost Campylobacter spp. u fecesu, brisevima s pilećih trupova i otpadne vode na liniji klanja. VIII Simpozij "Peradarski dani 2009." s međunarodnim sudjelovanjem, Zbornik "Peradarski dani 2009.", 25.-28. ožujka 2009. Poreč, Hrvatska, 184-190.

PRISUTNOST RODA CAMPYLOBACTER U KOŽAMA VRATOVA TOVNIH PILIĆA TIJEKOM LJETNOG RAZDOBLJA 2018. GODINE
144
PRESENCE OF THE GENUS CAMPYLOBACTER IN BROILER NECK SKIN DURING THE SUMMER PERIOD OF 2018
Summary
Bacteria of the genus Campylobacter cause the infection campylobacteriosis, a zoonotic disease transmitted to humans from animals and animal products. In humans, the disease is mainly caused by Campylobacter jejuni and Campylobacter coli. Campylobacter spp. is a widespread pathogen, found in all types of meat and meat products, raw milk and products from raw milk, less frequently in eggs. It could also be found in different foods as the result of cross-contamination, as well as in contaminated water. Over the past decade, Campylobacter spp. is the most common cause of bacterial food-borne illness in the European Union and United States. Campylobacteriosis is more prevalent during the summer months, due to high temperature that favor the bacterial growth. During the four weeks in the summer period of 2018, 60 samples of broiler neck skin were collected from three slaughterhouses and tested for the presence of bacteria of the genus Campylobacter. Results showed that out of all tested samples, 31 (52%) samples were unsuitable according to the Commission Regulation (EU) 2017/1495 (limit of 1000 cfu/g). Samples were processed according to the standard method for detection and determination of Campylobacter spp. count (HRN EN ISO 10272-2:2017). Because of the pathogenicity of the bacteria of the genus Campylobacter and in order to ensure food safety, implementation of measures of water disinfection, thermal processing of food, prevention of cross-contamination and good sanitation at food processing facilities are of utmost importance. Keywords: Campylobacter spp., broiler neck skin, contamination, good hygiene practices

PERADARSKI DANI 2019. 145
UTJECAJ GODIŠNJEG RAZDOBLJA NA POJAVNOST GLJIVICA U HRANI ZA TOVNE PILIĆE
Ivica Ravić1, Mario Ostović2, Matija Kovačić3, Anamaria Ekert Kabalin2, Kristina Matković2, Ivana Sabolek2, Željko Pavičić2, Ivona Žura Žaja2, Hrvoje Valpotić2, Danijela Horvatek Tomić2
1 Veterinarski zavod, Mostar, Bosna i Hercegovina 2 Sveučilište u Zagrebu, Veterinarski fakultet, Zagreb, Hrvatska 3 Poljoprivredna proizvodnja Kovačić, Šopron, Kalnik, Hrvatska
Sažetak
Cilj istraživanja bio je utvrditi broj i kvalitativni sastav gljivica u hrani za tovne piliće u različitim razdobljima godine, s naglaskom na vrste iz roda Aspergillus kao jednima od najpatogenijih gljivica za perad. Uzorci hrane (n=5) uzimani su izravno iz hranilica jedanput na tjedan tijekom 36-dnevnog tova u ljetnom i zimskom razdoblju godine, u komercijalnim uvjetima proizvodnje. Pilići su hranjeni ad libitum kompletnim krmnim smjesama. Podatci su analizirani referentnim programom Statistica v. 13.4. Rezultati istraživanja pokazali su da je broj gljivica u hrani bio najmanji u prvom tjednu tova, značajno veći u trećem, a najveći u posljednjem, petom tjednu tijekom oba razdoblja (p<0,05 sve). Broj gljivica bio je značajno veći u prvom i četvrtom tjednu tova tijekom ljetnog razdoblja u usporedbi sa zimskim, ali i u prosjeku tijekom tova ljeti (p<0,05 sve). Tijekom oba razdoblja iz hrane je u prosjeku izdvojeno najviše kvasaca, no značajno više ljeti (p<0,05). Godišnje razdoblje nije imalo utjecaja na prosječan broj plijesni A. niger, A. terreus, Cladosporium sp., Fusarium sp. i Rhizopus sp., dok je A. flavus bio značajno češće izdvojen iz hrane u toplom, a Mucor sp. i Penicillium sp. u hladnom razdoblju godine (p<0,05 sve). Može se zaključiti da učestalost i zastupljenost pojedinih vrsta gljivica u hrani za piliće u tovu ovise o razdoblju godine. Više pozornosti higijeni hrane za tovne piliće trebalo bi pridati tijekom toplijih mjeseci u godini. Ključne riječi: tovni pilići, gljivice, hrana, godišnje razdoblje Uvod
Današnji linijski hibridi peradi koji se uzgajaju za pro-izvodnju mesa i jaja balansiraju između zdravog i bolesnog stanja, a njihov genetski potencijal očitovat će se ako im se, među ostalima, osiguraju optimalni uzgojni uvjeti (Vu-čemilo, 2008.). Većina bolesti koje se pojavljuju na farmama upravo su uvjetne ili uzgojne bolesti. Samo zdrave životinje mogu odgovoriti na postavljene zahtjeve i uspješno proizvoditi, a zdravlje se može definirati kao rezultat djelovanja nekoliko čimbenika, kao što su genetski potencijal, kvaliteta hrane, živi agensi, uvjeti okoliša i tehnologija proizvodnje (Vučemilo i sur., 2006.). Gljivice su heterotrofni eukariotski mikroorganizmi u koje spadaju plijesni i kvasci, a bolest mogu uzrokovati izravnim zaražavanjem i oštećenjem tkiva i organa ili neizravno stvaranjem mikotoksina koji uobičajeno oralnim putem
dospijevaju u organizam te uzrokuju intoksikaciju i imunosupresiju, narušavajući proizvodnost peradi (Jand i sur., 2005.; Dhama i sur., 2013.). Gljivične bolesti relativno su rijetke u životinja, ali kada se pojave uglavnom su vrlo razorne za domaćina te uzrokuju velike ekonomske gubitke. Pogodovni čimbenici su dugotrajna izloženost velikim količinama spora gljivica, stres, imunosupresija, prisutnost drugih bolesti, neprikladna hranidba, slabo prozračivanje, loši higijenski uvjeti, vlažna hrana i stelja i dr. (Biđin, 2008.; Beernaert i sur., 2010.; Kapetanov i sur., 2011.; Sokolović i sur., 2015.). Gljivične bolesti peradi dobivaju sve više na važnosti diljem svijeta i s obzirom na često prekomjernu uporabu antibiotika koji eliminirajući prirodnu bakterijsku mikrofloru otvaraju ulazna vrata za oportunističke infekcije (Swayne i sur., 2013.). Uz to, bitan je i zoonotski potencijal gljivičnih bolesti (Girma i sur., 2016.; Ostović i sur., 2017.a).
doc. dr. sc. Mario Ostović, Veterinarski fakultet Sveučilišta u Zagrebu, Zavod za higijenu, ponašanje i dobrobit životinja, Heinzelova 55, 10000 Zagreb, Hrvatska, Tel. +385 (0)1 2390-294, Fax: +385 (0)1 2441-390, e-mail: [email protected]

UTJECAJ GODIŠNJEG RAZDOBLJA NA POJAVNOST GLJIVICA U HRANI ZA TOVNE PILIĆE 146
Hrana je jedan od najvažnijih izvora gljivica u peradarskoj proizvodnji (Arné i sur., 2011.). Broj gljivica u hrani za perad kreće se u prosjeku od 102 do 106 CFU/g (Magnoli i sur., 1998.; Marković i sur., 2005.; Krnjaja i sur., 2008.; 2014.), pri čemu su najčešće izdvojene vrste iz rodova Aspergillus, Fusarium, Penicillium, Rhizopus i Mucor (Dalcero i sur., 1998.; Marković i sur., 2005.; Krnjaja i sur., 2008.; Saleemi i sur., 2010.; Cegielska-Radziejewska i sur., 2013.; Rahman i sur., 2015.). Gljivične bolesti, među kojima je najznačajnija aspergiloza s obzirom na učestalost, zdravstvenu i ekonomsku važnost (Beernaert i sur., 2010.; Swayne i sur., 2013.), pojavljuju se tijekom čitave godine u uzgojima peradi. Međutim, rezultati dosadašnjih istraživanja s jedne strane upućuju na veću pojavu bolesti i gljivica u zraku peradarskih nastambi tijekom toplog razdoblja u vlažnim uvjetima, a s druge strane tijekom hladnijih mjeseci i uz nižu vlagu zraka (Debey i sur., 1995.; Sajid i sur., 2006.; Wójcik i sur., 2010.; Sowiak i sur., 2012.; Matković i sur., 2013.; Sultana i sur., 2015.; Ostović i sur., 2017.b). Cilj ovog istraživanja bio je utvrditi kako toplo odnosno hladno razdoblje godine utječe na pojavnost gljivica u hrani za perad, s naglaskom na vrste iz roda Aspergillus.
Materijal i metode
Istraživanje je provedeno u Koprivničko-križevačkoj žu-paniji u kontroliranim, komercijalnim uvjetima proizvodnje tijekom 36-dnevnog tova pilića u ljetnom (toplom) i zimskom (hladnom) razdoblju godine: srpanj-kolovoz 2016. godine i prosinac 2016.-siječanj 2017. godine. Tijekom svakog turnusa u nastambi je bilo 18 000 pilića hibrida Ross držanih na stelji od sjeckane slame i piljevine s gustoćom naseljenosti do 33 kg/m2. Hranjenje i napajanje pilića bili su ad libitum, a hranili su se kompletnim krmnim smjesama (starter, grower ~10 dan i finisher ~ 20 dan, Biodar, Varaždin). Hrana se dopremala iz mješaonice i skladištila u silosima koji su se punili ovisno o kapacitetu svakih nekoliko dana. Jedanput na tjedan tijekom svakog turnusa (1., 8., 15., 22. i 35. dan) određivale su se učestalost i zastupljenost pojedinih vrsta gljivica u hrani. Uzorci su prikupljeni izravno iz hranilica (n=5), nakon raspodjele hrane te je 10 g uzorka deseterostruko razrijeđeno sterilnom destiliranom vodom, od čega je 100 µL nasađeno razmazivanjem na Sabouraud agar (Sabouraud dextrose agar, Biolife, Italija) i inkubirano pri 22 °C kroz 5 dana. Identifikacija gljivica temeljila se na makroskopskom izgledu izraslih kolonija te mikroskopskoj
pretrazi spora bojanjem laktofenolom (De Hoog i sur., 2000.; Samanta, 2015.). Rezultati su izraženi kao broj gljivica po gramu (CFU/g) hrane. U tjednim razmacima mjerene su i temperatura i relativna vlaga zraka izvan nastambe uz silos prenosivim digitalnim uređajem (Testo, Njemačka). Podatci su analizirani referentnim računalnim programom Statistica v. 13.4 (TIBCO Software Inc., 2017.). Normalnost raspodjele podataka provjerena je Kolmogorov-Smir-novljevim testom. Za testiranje značajnosti razlika u broju gljivica između razdoblja godine (ljeto-zima) po pojedinim tjednima tova, kao i prosječnih razlika korišten je Mann-Whitneyjev U-test, a za testiranje razlika unutar razdoblja između pojedinih tjedana tova Friedmanova analiza varijance i Wilcoxonov test. Za testiranje značajnosti razlika u prosječnim vrijednostima temperature i relativne vlage zraka između razdoblja godine korišten je Studentov t-test. Statistički značajnom smatrana je razlika na razini p<0,05.
Rezultati i rasprava
U ovom radu istraživani su broj i kvalitativni sastav gljivica u hrani za piliće tijekom tova u toplom i hladnom razdoblju godine. Kretanje broja gljivica po tjednima tova u različitim razdobljima prikazano je na slici 1., a prosječan broj i sastav mikoflore po razdobljima tova u tablicama 1. i 2. Broj gljivica u svim tjednima tova tijekom oba razdoblja (slika 1.) odnosno njihov prosječan broj (tablica 1.) bio je unutar vrijednosti utvrđenih u istraživanjima Magnoli i sur. (1998.), Marković i sur. (2005.) i Krnjaja i sur. (2008.; 2014.), s time da u prethodnim istraživanjima uzorci hrane nisu uzimani izravno iz hranilica, već iz skladišta na farmama ili tvornica hrane. No, značajno manji broj gljivica (p<0,05) ustanovljen je u prvom i četvrtom tjednu tova tijekom zimskog razdoblja (slika 1.), ali i u prosjeku zimi (tablica 1.). Ovaj nalaz može se protumačiti slabijim preživljavanjem i razmnožavanjem gljivica u hladnom razdoblju. Vrijednosti temperature zraka mjerene izvan nastambe kretale su se od 14,00 °C do 29,00 °C tijekom tova u toplom razdoblju godine, a u hladnom od -6,70 °C do 4,60 °C, pri čemu je značajno niža prosječna temperatura zraka (p<0,05) utvrđena u hladnom razdoblju (-0,02 °C prema 24,90 °C). Vrijednosti relativne vlage zraka izvan nastambe kretale su se od 37,90% do 84,10% tijekom tova u toplom razdoblju, a u hladnom od 40,20% do 87,20%, u kojem je utvrđena značajno viša prosječna relativna vlaga zraka (p<0,05) (66,80% prema 55,99%).
Tablica 1. Prosječan broj gljivica u hrani za piliće tijekom tova u ljetnom i zimskom razdoblju godine Table 1. Average fungal count in broiler feed during fattening cycle in summer and winter seasons
Pokazatelj / Parameter Ljeto / Summer Zima / Winter
Gljivice / Fungi (CFU/g) Medijan / Median 4,30×103* 1,80×103* min. / Min 1,00×103 1,00×102 maks. / Max 1,72×104 4,57×104
*p<0,05
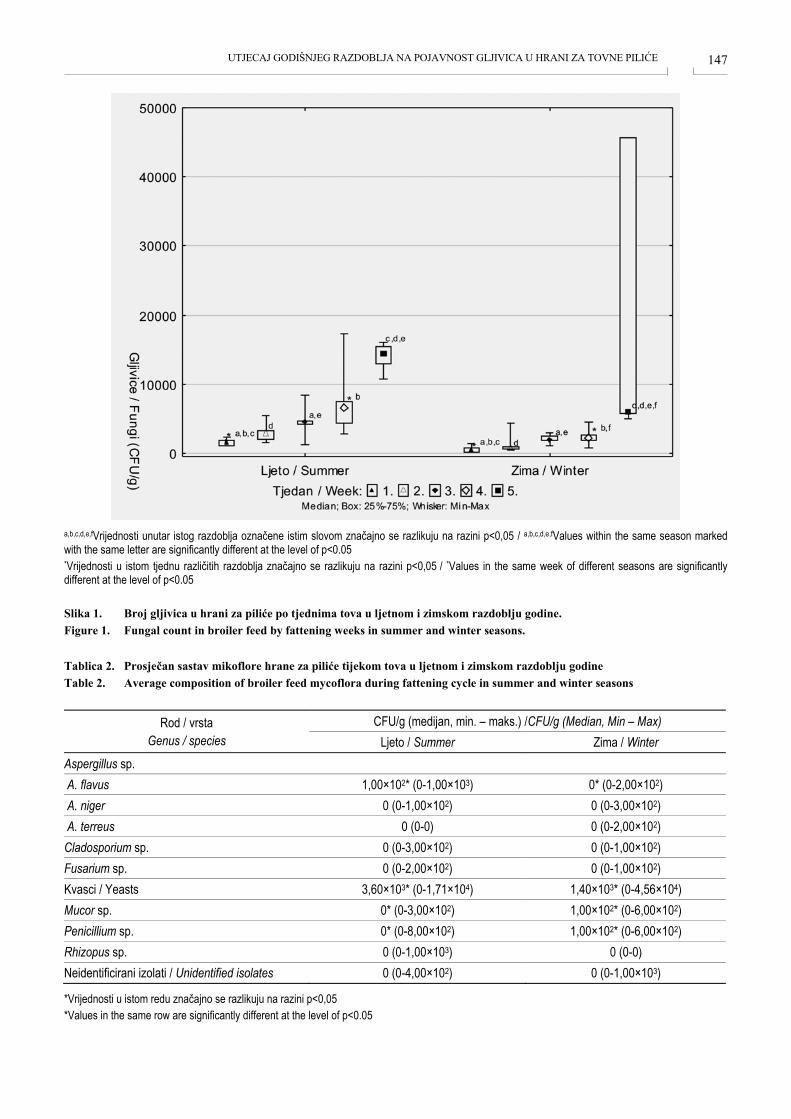
UTJECAJ GODIŠNJEG RAZDOBLJA NA POJAVNOST GLJIVICA U HRANI ZA TOVNE PILIĆE 147
a,b,c,d,e,fVrijednosti unutar istog razdoblja označene istim slovom značajno se razlikuju na razini p<0,05 / a,b,c,d,e,fValues within the same season marked with the same letter are significantly different at the level of p<0.05 *Vrijednosti u istom tjednu različitih razdoblja značajno se razlikuju na razini p<0,05 / *Values in the same week of different seasons are significantly different at the level of p<0.05 Slika 1. Broj gljivica u hrani za piliće po tjednima tova u ljetnom i zimskom razdoblju godine. Figure 1. Fungal count in broiler feed by fattening weeks in summer and winter seasons.
Tablica 2. Prosječan sastav mikoflore hrane za piliće tijekom tova u ljetnom i zimskom razdoblju godine Table 2. Average composition of broiler feed mycoflora during fattening cycle in summer and winter seasons
Rod / vrsta Genus / species
CFU/g (medijan, min. – maks.) /CFU/g (Median, Min – Max) Ljeto / Summer Zima / Winter
Aspergillus sp. A. flavus 1,00×102* (0-1,00×103) 0* (0-2,00×102) A. niger 0 (0-1,00×102) 0 (0-3,00×102) A. terreus 0 (0-0) 0 (0-2,00×102) Cladosporium sp. 0 (0-3,00×102) 0 (0-1,00×102) Fusarium sp. 0 (0-2,00×102) 0 (0-1,00×102) Kvasci / Yeasts 3,60×103* (0-1,71×104) 1,40×103* (0-4,56×104) Mucor sp. 0* (0-3,00×102) 1,00×102* (0-6,00×102) Penicillium sp. 0* (0-8,00×102) 1,00×102* (0-6,00×102) Rhizopus sp. 0 (0-1,00×103) 0 (0-0) Neidentificirani izolati / Unidentified isolates 0 (0-4,00×102) 0 (0-1,00×103)
*Vrijednosti u istom redu značajno se razlikuju na razini p<0,05 *Values in the same row are significantly different at the level of p<0.05

UTJECAJ GODIŠNJEG RAZDOBLJA NA POJAVNOST GLJIVICA U HRANI ZA TOVNE PILIĆE 148
Broj gljivica bio je najmanji početkom tova, značajno veći sredinom, a najveći na kraju tova tijekom oba razdoblja (p<0,05 sve) (slika 1.), što se može objasniti time da hrana za starije piliće zbog svog sastava sadrži više gljivica (Cegielska-Radziejewska i sur., 2013.). S obzirom na vrste gljivica utvrđene u ovom istraživanju rezultati su u suglasju s rezultatima drugih istraživanja (Dalcero i sur., 1998.; Marković i sur., 2005.; Krnjaja i sur., 2008.; Saleemi i sur., 2010.; Cegielska-Radziejewska i sur., 2013.; Rahman i sur., 2015.). Iz hrane su izdvojene sljedeće gljivice: A. flavus, A. niger, A. terreus, Cladosporium sp., Fusarium sp., kvasci, Mucor sp., Penicillium sp. i Rhizopus sp. Najučestalije gljivice u prosjeku su bile kvasci tije- kom oba razdoblja, u značajno većem broju (p<0,05) ljeti (tablica 2.). Rast gljivica na žitaricama može smanjiti njihovu hranid-benu vrijednost, a kontaminirati se mogu prije ili nakon žetve, tijekom skladištenja. Vrste iz rodova Aspergillus, Fusarium i Penicillium najčešće su plijesni koje razgrađuju žitarice, odnosno kontaminiraju ih svojim sekundarnim me-tabolitima, mikotoksinima (Valpotić i Šerman, 2006.; Krnja-ja i sur., 2017.; Ekwomadu i sur., 2018.). Razvoj plijesni i proizvodnja mikotoksina ovise o mnogim čimbenicima, kao što su vrsta hranjivog supstrata, sadržaj vlage, pH vrijednost, temperatura i relativna vlažnost zraka, oštećenje zrna žitarica i dr. (Perši i sur., 2011.). Rezultati ovog istraživanja pokazali su da je hrana za piliće ljeti sadržavala u prosjeku značajno više (p<0,05) plijesni vrste A. flavus, a zimi Mucor sp. i Penicillium sp. Prosječan broj ostalih plijesni utvrđenih u istraživanju nije se značajno razlikovao između razdoblja godine. A. terreus nije izdvojen iz hrane u ljetnom, a Rhizopus sp. u zimskom razdoblju godine (tablica 2.). Habib i sur. (2015.) ispitivali su pojavnost Aspergillus sp. u hrani za perad. A. fumigatus bio je najzastupljeniji izolat, zatim A. parasiticus, A. flavus, A. niger i A. terreus. U istraživanjima Azarakhsh i sur. (2011.) i Aliyu i sur. (2016.) hrana je također najčešće bila kontaminirana vrstom A. fumigatus, dok u drugim istraživanjima (Magnoli i sur., 1998.; Rahman i sur., 2015.) to nije bio slučaj. U ovom istraživanju A. fumigatus kao najpatogenija gljivica iz ovog roda (Fulleringer i sur., 2006.; Arné i sur., 2011.) nije utvrđen ni u toplom niti u hladnom razdoblju godine.
Zaključak
Iz dobivenih rezultata može se zaključiti da godišnje raz-doblje utječe na učestalost i zastupljenost pojedinih vrsta gljivica u hrani za tovne piliće. Broj gljivica u hrani veći je tijekom toplog, nego hladnog razdoblja, stoga bi tijekom toplijih mjeseci u godini više pozornosti trebalo posvetiti njezinoj higijeni, odnosno kvaliteti.
Literatura
ALIYU, R. M, M. B ABUBAKAR, Y. YAKUBU, A. B. KASARAWA, N. LAWAL, M. B. BELLO, A. Y. FARDAMI (2016): Prevalence of potential toxigenic Aspergillus species
isolated from poultry feeds in Sokoto metropolis. Sokoto J. Vet. Sci., 14, 39-44.
ARNÉ, P., S. THIERRY, D. WANG, M. DEVILLE, G. LE LOC’H, A. DESOUTTER, F. FÉMÉNIA, A. NIEGUITSILA, W. HUANG, R. CHERMETTE, J. GUILLOT (2011): Aspergillus fumigatus in poultry. Int. J. Microbiol., Volume 2011, Article ID 746356, 14 p.
AZARAKHSH, Y., A. SABOKBAR, M. BAYAT (2011): Incidence of the most common toxigenic Aspergillus species in broiler feeds in Kermanshah Province, West of Iran. Glob. Vet., 6, 73-77.
BEERNAERT, L. A., F. PASMANS, L. VAN WAEYEN-BERGHE, F. HAESEBROUCK, A. MARTEL (2010): Asper-gillus infections in birds: a review. Avian Pathol., 39, 325-331.
BIĐIN, Z. (2008): Bolesti peradi. Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska.
CEGIELSKA-RADZIEJEWSKA, R., K. STUPER-SZABLEW-SKA, T. SZABLEWSKI (2013): Microflora and mycotoxin contamination in poultry feed mixtures from western Poland. Ann. Agric. Environ. Med., 20, 30-35.
DALCERO, A., C. MAGNOLI, M. LUNA, G. ANCASI, M. M. REYNOSO, S. CHIACCHIERA, R. MIAZZO, G. PALACIO (1998): Mycoflora and naturally occurring mycotoxins in poultry feeds in Argentina. Mycopathologia, 141, 37-43.
DEBEY, M. C., D. W. TRAMPEL, J. L. RICHARD, D. S. BUNDY, L. J. HOFFMAN, V. M. MEYER, D. F. COX (1995): Effect of environmental variables in turkey confinement houses on airborne Aspergillus and mycoflora composition. Poult. Sci., 74, 463-471.
DE HOOG, G. S., J. GUARRO, J. GENE, M. J. FIGUERAS (2000): Atlas of Clinical Fungi, 2nd ed. Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands.
DHAMA, K., S. CHAKRABORTY, A. K. VERMA, R. TIWARI, R. BARATHIDASAN, A. KUMAR, S. D. SINGH (2013): Fungal/mycotic diseases of poultry – diagnosis, treatment and control: a review. Pak. J. Biol. Sci., 16, 1626-1640.
EKWOMADU, T. I., R. E. GOPANE, M. MWANZA (2018): Occurrence of filamentous fungi in maize destined for human consumption in South Africa. Food Sci. Nutr., 6, 884-890.
FULLERINGER, S. L., D. SEGUIN, S. WARIN, A. BEZILLE, C. DESTERQUE, P. ARNÉ, R. CHERMETTE, S. BRETAGNE, J. GUILLOT (2006): Evolution of the environmental contamination by thermophilic fungi in a turkey confinement house in France. Poult. Sci., 85, 1875-1880.
GIRMA, G., M. ABEBAW, M. ZEMENE, Y. MAMUYE, G. GETANEH (2016): A review on Aspergillosis in poultry. J. Vet. Sci. Technol., 7, 1-5.
HABIB, M. A., P. ABDU, C. N. KWANASHIE, J. KABIR, A. NEGEDU (2015): Isolation and identification of Aspergillus species from poultry feeds in Kaduna State, Nigeria. Microbiol. Res. Int., 3, 27-32.
JAND, S. K., P. KAUR, N. S. SHARMA (2005): Mycoses and mycotoxicosis in poultry: a review. Ind. J. Anim. Sci., 75, 465-476.
KAPETANOV, M. C., D. V. POTKONJAK, D. S. MILANOV, I. M. STOJANOV, M. M. ŽIVKOV BALOŠ, B. Z. PRUNIĆ (2011): Investigation of dissemination of aspergillosis in poultry and possible control measures. Proc. Nat. Sci., Matica Srpska Novi Sad, 120, 269-278.
KRNJAJA, V., A. STANOJKOVIĆ, S. STANKOVIĆ, M. LUKIĆ, Z. BIJELIĆ, V. MANDIĆ, N. MIĆIĆ (2017): Fungal contamination of maize grain samples with a special focus on toxigenic genera. Biotechnol. Anim. Husb., 33, 233-241.

UTJECAJ GODIŠNJEG RAZDOBLJA NA POJAVNOST GLJIVICA U HRANI ZA TOVNE PILIĆE 149
KRNJAJA, V., LJ. STOJANOVIĆ, R. CMILJANIĆ, S. TRENKOVSKI, D. TOMAŠEVIĆ (2008): The presence of potentially toxigenic fungi in poultry feed. Biotechnol. Anim. Husb., 24, 87-93.
KRNJAJA, V., Z. PAVLOVSKI, M. LUKIĆ, Z. ŠKRBIĆ, LJ. STOJANOVIĆ, Z. BIJELIĆ, V. MANDIĆ (2014): Fungal contamination and natural occurrence of ochratoxin A (OTA) in poultry feed. Biotechnol. Anim. Husb., 30, 481-488.
MAGNOLI, C., A. M. DALCERO, S. M. CHIACCHIERA, R. MIAZZO, M. A. SAENZ (1998): Enumeration and identification of Aspergillus group and Penicillium species in poultry feeds from Argentina. Mycopathologia, 142, 27-32.
MARKOVIĆ, R. V., N. D. JOVANOVIĆ, D. S. ŠEFER, Z. J. SINOVEC (2005): Mould and mycotoxin contamination of pig and poultry feed. Proc. Nat. Sci., Matica Srpska Novi Sad, 109, 89-95.
MATKOVIĆ, K., M. VUČEMILO, I. ŠTOKOVIĆ, R. ŠIMIĆ, D. MARUŠIĆ, B. VINKOVIĆ, S. MATKOVIĆ (2013): Concentrations of airborne bacteria and fungi in a livestock building with caged laying hens. Vet. arhiv, 83, 413-424.
OSTOVIĆ, M., I. RAVIĆ, M. KOVAČIĆ, S. ŽUŽUL, K. MATKOVIĆ, Ž. PAVIČIĆ, A. EKERT KABALIN, S. MENČIK, D. HORVATEK TOMIĆ (2017a): Pojavnost gljivica na izloženim površinama u nastambi za tov pilića. XII. simpozij „Peradarski dani 2017. s međunarodnim sudje-lovanjem“. 10.-13.05.2017., Šibenik, Hrvatska, 122-126.
OSTOVIĆ, M., S. MENČIK, I. RAVIĆ, S. ŽUŽUL, Ž. PAVIČIĆ, K. MATKOVIĆ, B. ANTUNOVIĆ, D. HORVATEK TOMIĆ, A. EKERT KABALIN (2017b): Relation between microclimate and air quality in the extensively reared turkey house. Mac. Vet. Rev., 40, 83-90.
PERŠI, N., J. PLEADIN, A. VULIĆ, M. ZADRAVEC, M. MITAK (2011): Mikotoksini u žitaricama i hrani životinjskog podrijetla. Vet. stn., 42, 335-345.
RAHMAN, M. M., M. S. ISLAM, M. Z. ALAM, M. ASHRAFUZZAMAN (2015): An investigation into the
microbial infestation of poultry feeds. J. Bangladesh Agric. Univ., 13, 79-86.
SAJID, M. A., I. A. KHAN, U. RAUF (2006): Aspergillus fumigatus in commercial poultry flocks, a serious threat to poultry industry in Pakistan. J. Anim. Plant Sci., 16, 79-81.
SALEEMI, M. K., M. Z. KHAN, A. KHAN, I. JAVED (2010): Mycoflora of poultry feeds and mycotoxins producing potential of Aspergillus species. Pak. J. Bot., 42, 427-434.
SAMANTA, I. (2015): Veterinary Mycology. Springer, India. SOKOLOVIĆ, M., B. ŠIMPRAGA, F. KRSTULOVIĆ, M. BE-
RENDIKA (2015): Značaj patogenih plijesni u peradarstvu. XI. simpozij „Peradarski dani 2015. s međunarodnim sudjelovanjem“. 13.-16.05.2015., Šibenik, Hrvatska, 32-38.
SOWIAK, M., K. BRÓDKA, A. KOZAJDA, A. BUCZYŃSKA, I. SZADKOWSKA-STAŃCZYK (2012): Fungal aerosol in the process of poultry breeding – quantitative and qualitative analysis. Med. Pr., 63, 1-10.
SULTANA, S., S. M. H. RASHID, M. N. ISLAM, M. H. ALI, M. M. ISLAM, M. G. AZAM (2015): Pathological investigation of avian Aspergillosis in commercial broiler chicken at Chittagong district. Int. J. Innov. Appl. Stud., 10, 366-376.
SWAYNE, D. E., J. R. GLISSON, L. R. McDOUGALD, L. K. NOLAN, D. L. SUAREZ, V. NAIR, Eds. (2013): Diseases of Poultry, 13th ed. Wiley-Blackwell, John Wiley & Sons, Inc., Ames, Iowa.
VALPOTIĆ, H., V. ŠERMAN (2006): Utjecaj mikotoksina na zdravlje i proizvodnost svinja. Krmiva, 48, 33-42.
VUČEMILO, M. (2008): Higijena i bioekologija u peradarstvu. Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska.
VUČEMILO, M., B. VINKOVIĆ, K. MATKOVIĆ (2006): Smještaj i dobrobit životinja na farmama, te uvjeti kojima moraju udovoljavati farme u svjetlu novog Pravilnika. Krmiva, 48, 43-47.
WÓJCIK, A., Ł. CHORĄŻY, T. MITUNIEWICZ, D. WITKOW-SKA, K. IWAŃCZUK-CZERNIK, J. SOWIŃSKA (2010): Microbial air contamination in poultry houses in the summer and winter. Pol. J. Environ. Stud., 19, 1045-1050.
SEASONAL IMPACT ON THE OCCURRENCE OF FUNGI IN BROILER FEED
Summary
The aim of the study was to determine the count and qualitative composition of fungi in broiler feed in different seasons, with emphasis on Aspergillus species as one of the most pathogenic fungi for poultry. Feed was sampled (n=5) weekly directly from feeders during 36-day fattening cycle in summer and winter seasons, in commercial production conditions. The broilers were fed ad libitum with complete feed mixtures. Data were analyzed using the respective Statistica v. 13.4 software. Study results showed the lowest fungal count in feed in the first fattening week, a significantly higher count in the third week, and the highest count in the last, fifth, week during both seasons (p<0.05 all). The fungal count was significantly higher in the first and fourth fattening week of the summer season compared to winter season, as well as average fungal count recorded in the summer fattening season (p<0.05 all). During both seasons, yeasts were most frequently isolated; yet, significantly more in the summer (p<0.05). The season had no impact on the average counts of moulds A. niger, A. terreus, Cladosporium sp., Fusarium sp. and Rhizopus sp., while A. flavus was significantly more often isolated in the warm season, and Mucor sp. and Penicillium sp. in the cold season (p<0.05 all). It can be concluded that frequency and diversity of fungi in broiler feed depend on the season. More attention should be paid to the broiler feed hygiene during warmer months of the year. Key words: broilers, fungi, feed, season

PERADARSKI DANI 2019. 150
PREVALENCE OF GALLIBACTERIUM ANATIS AND ESCHERICHIA COLI ON BROILER BREEDER FARMS IN CROATIA
Liča Lozica, Danijela Horvatek Tomić, Željko Gottstein
University of Zagreb, Faculty of Veterinary Medicine, Zagreb, Croatia
Summary
Gallibacterium (G.) anatis and Escherichia (E.) coli are some of the most common causative agents of peritonitis, folliculitis and salpingitis, as well as upper respiratory tract disorders in poultry. They can act as primary pathogens or appear in coinfection with other microorganisms such as Mycoplasma spp., infectious bronchitis virus (IBV), etc. Less prominent clinical symptoms and pathomorphological lesions usually follow Gallibacterium infections, while colibacillosis is recognized by more severe lesions. The infection can be subclinical, but usually causes great production decrease and high mortality rates. In this study, we wanted to determine the prevalence of G. anatis and E. coli on two broiler breeder farms to assess the epizootiologic situation in Croatia. During this epizootiologic survey, poultry carcasses and swabs from live animals were brought from two broiler breeder farms to the Department of Poultry Diseases with Clinic where necropsy was done. During examination, 200 swab samples were taken from the pharynx, peritoneum, liver, oviduct and bone marrow of 128 animals in total. Samples were then plated on 5% sheep blood agar and incubated aerobically at 37 °C for 24 hours. Identification was done by Gram staining, biochemical tests and MALDI-TOF spectrometry method. Afterwards, the isolates were stored in BHI at -80 °C for further analyses. Altogether 15.5% of the samples from both farms were positive for G. anatis. Of these, 93.5% of isolates originated from the pharynx, while only 6.5% originated from the peritoneum. In contrast, 38% of the samples from both farms were positive for E. coli and even 62.5% of the isolates originated from the oviduct. Co-infection with G. anatis and E. coli was present only in two cases, but the flocks were previously positive for Mycoplasma synoviae and 793B IBV strains were detected. These data confirmed the assumption on G. anatis being an opportunistic pathogen and demonstrated the higher prevalence of E. coli on the examined broiler breeder farms. Since the isolates are usually farm specific and often multidrug resistant, therapy is not always successful and the use of autogenous vaccines with adjustment of the immunoprophylaxis program for every flock individually is recommended. Keywords: Gallibacterium anatis, Escherichia coli, broiler breeder, autogenous vaccine, poultry Introduction
Gallibacterium (G.) anatis is part of the normal upper respiratory and lower reproductive tract flora of poultry and wild birds. However, it can cause various lesions including hepatitis, pericarditis, salpingitis, oophoritis, peritonitis, septicemia, enteritis and respiratory tract lesions, which lead to production decrease and high mortality rates (Neubauer et al., 2009; Bojesen et al., 2003a; Christensen et al., 2003; Bisgaard et al., 1993; Bisgaard et al., 1977). In the past two decades, the name of the bacterium has changed several times from Pasteurella haemolytica, Actinobacillus salpin-gitidis, Pasteurella anatis to Gallibacterium anatis. In 1993, Bisgaard proposed that Gallibacterium should be established
as a genus within the family Pasteurellaceae, which was later confirmed based on 16S rRNA gene sequences (Christensen et al., 2003). Escherichia (E.) coli is also one of the most frequent causative agents of various disorders in poultry. The infection can be localized, but often develops into a systemic infection and leads to septicemia. The most common lesions associated with colibacillosis are peri-hepatitis, airsacculitis, pericarditis, salpingitis, fibrinous peritonitis and septicemia (Landman et al., 2013; Mellata, 2013; Ozaki and Murase, 2009; Dziva and Stevens, 2008). Gallibacterium infection is similarly characterized by very unspecific clinical signs and pathologic lesions that occur in many different diseases besides colibacillosis, such as infectious bronchitis, infectious coryza and fowl cholera.
Liča Lozica, DVM, University of Zagreb, Faculty of Veterinary Medicine, Department of Poultry Diseases with Clinic, Heinzelova 55, 10000 Zagreb, Croatia; Tel: +385 (0)1 239 0281; fax: 01/2390410; e-mail: [email protected]

PREVALENCE OF GALLIBACTERIUM ANATIS AND ESCHERICHIA COLI ON BROILER BREEDER FARMS IN CROATIA 151
Depression, diarrhea and egg drop syndrome due to folli-cular atrophy, folliculitis or salpingitis can be observed, as well as variation in egg size, egg deformities, and high mortality rates caused by peritonitis. Although there are examples of acute infection which led to septicemia and sudden mortality, more frequently, the infection is chronic. The most common lesions are peritonitis, hepatitis, sal-pingitis and respiratory syndrome (Bojesen et al., 2004; Kristensen et al., 2011; Paudel et al., 2013; Persson and Bojesen, 2015; Proctor et al., 2006; Singh et al., 2016). Like E. coli, it is considered one of the most common causes of respiratory and reproductive tract disorders in layers (Proctor et al., 2006; Bojesen et al., 2007; Kristensen et al., 2011; Singh et al., 2016). Both bacteria can be transmitted horizontally and vertically (Poulsen et al., 2017; Persson and Bojesen, 2015; Paudel et al., 2014). In layers, reproductive disorders, which lead to decreased egg production and economic losses, are dominant, whereas in broilers the infection usually causes high mortality rates. In broiler breeders, G. anatis causes mostly systemic infections that are recognized by high mortality, egg drop syndrome, peritonitis and airsacculitis (Jones et al., 2013). It can be a primary pathogen, but also part of co-infection with different microorganisms such as E. coli, Mycoplasma spp., Avi-bacterium paragallinarum and infectious bronchitis virus (IBV) (Paudel et al., 2017; Bojesen et al., 2003b). Since colibacillosis is more easily recognized and better known in poultry production, Gallibacterium infection is often underdiagnosed or misdiagnosed, which has led to
inaccurate therapy of the poultry flocks and development of the multi-drug resistant bacteria, without resolving the cause of the problem. Due to great financial losses that are caused by E. coli and G. anatis infection, we wanted to conduct an epizootiological survey to evaluate the prevalence of both bacteria on two broiler breeder farms as individual causative agents and as part of co-infection.
Material and Methods
Epizootiological survey was conducted from December 2017 until December 2018. Poultry carcasses and swabs from live animals were brought from two broiler breeder farms (Ross 308) to the Department of Poultry Diseases with Clinic, where necropsies were done. Altogether 128 animals were examined (Table 1). During examinations, 200 swab samples in total were taken from the pharynx, peri-toneum, liver, oviduct and bone marrow. Samples were then plated on Columbia agar (Oxoid, UK) enriched with 5% sheep blood (Biognost, Croatia) and incubated aerobically at 37 °C for 24 hours. Colonies that showed ß-hemolysis zones on blood agar and morphologically matched the G. anatis description were sub-cultured. Colonies that morpho-logically matched E. coli were subcultered on Brilliant Green agar (Oxoid, UK) in order to confirm the identifi-cation. Afterwards, final identification was done by Gram staining, biochemical tests and MALDI-TOF spectrometry method (Alispahić et al., 2012). The isolates were stored in BHI broth (Oxoid, UK) at -80 °C for further analyses.
Table 1. Number of live animals and carcasses sampled per farm Tablica 1. Broj živih i uginulih pretraženih životinja s obzirom na farmu podrijetla
Total number of sampled animals Ukupan broj uzorkovanih životinja Live animals / Žive životinje (%) Carcasses / Trupovi (%)
Farm 1 / Farma 1 83 45 (54.2) 38 (45.8)
Farm 2 / Farma 2 45 30 (66.7) 15 (33.3) Table 2. Prevalence of G. anatis and E. coli from pharynx, peritoneum, liver, oviduct and bone marrow from both farms Tablica 2. Učestalost bakterija G. anatis i E. coli u uzorcima ždrijela, peritoneuma, jetre, jajovoda i koštane srži s obiju farmi
G. anatis positive samples Uzorci pozitivni na G. anatis (%)
E. coli positive samples (%) Uzorci pozitivni na E. coli
Total number of samples per organ Ukupan broj uzoraka po organu
Pharynx / Ždrijelo 29 (34.9) 16 (19.28) 83
Peritoneum / Peritoneum 2 (9.1) 11 (50.0) 22
Liver / Jetra 0 17 (47.22) 36
Oviduct / Jajovod 0 20 (62.5) 32
Bone marrow / Koštana srž 0 12 (44.4) 27
Total / Ukupno 31 (15.5) 76 (38.0) 200
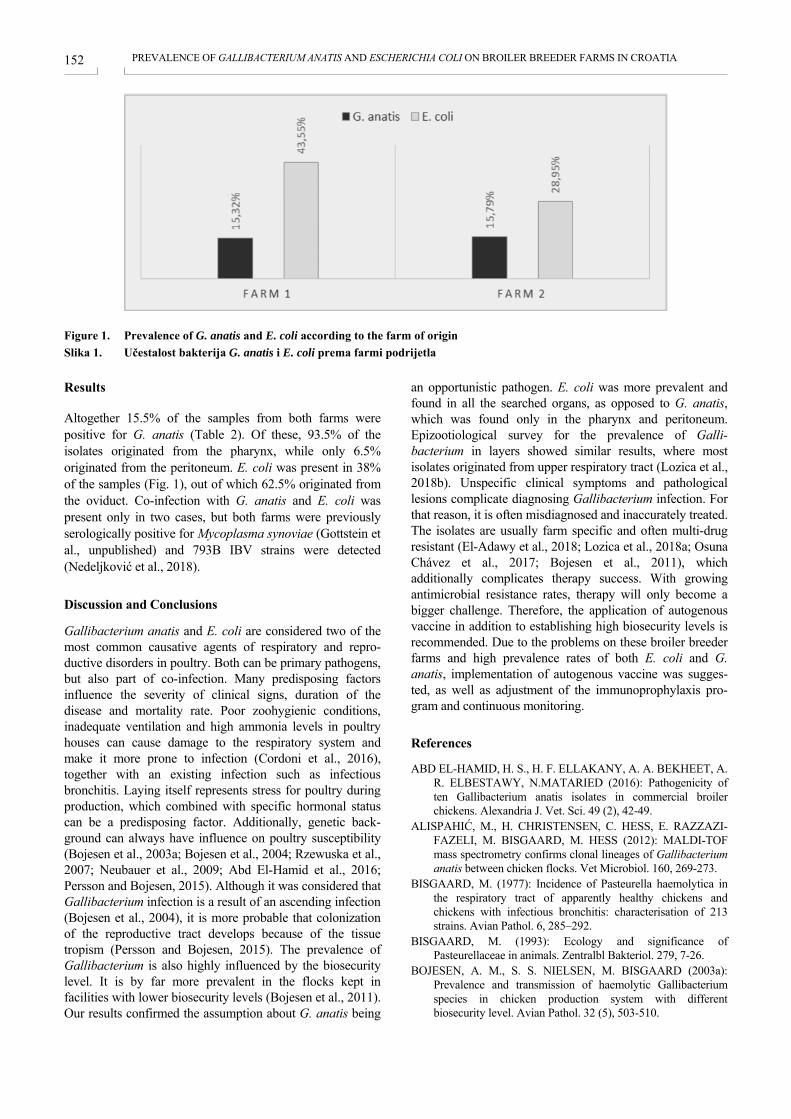
PREVALENCE OF GALLIBACTERIUM ANATIS AND ESCHERICHIA COLI ON BROILER BREEDER FARMS IN CROATIA 152
Figure 1. Prevalence of G. anatis and E. coli according to the farm of origin Slika 1. Učestalost bakterija G. anatis i E. coli prema farmi podrijetla Results
Altogether 15.5% of the samples from both farms were positive for G. anatis (Table 2). Of these, 93.5% of the isolates originated from the pharynx, while only 6.5% originated from the peritoneum. E. coli was present in 38% of the samples (Fig. 1), out of which 62.5% originated from the oviduct. Co-infection with G. anatis and E. coli was present only in two cases, but both farms were previously serologically positive for Mycoplasma synoviae (Gottstein et al., unpublished) and 793B IBV strains were detected (Nedeljković et al., 2018).
Discussion and Conclusions
Gallibacterium anatis and E. coli are considered two of the most common causative agents of respiratory and repro-ductive disorders in poultry. Both can be primary pathogens, but also part of co-infection. Many predisposing factors influence the severity of clinical signs, duration of the disease and mortality rate. Poor zoohygienic conditions, inadequate ventilation and high ammonia levels in poultry houses can cause damage to the respiratory system and make it more prone to infection (Cordoni et al., 2016), together with an existing infection such as infectious bronchitis. Laying itself represents stress for poultry during production, which combined with specific hormonal status can be a predisposing factor. Additionally, genetic back-ground can always have influence on poultry susceptibility (Bojesen et al., 2003a; Bojesen et al., 2004; Rzewuska et al., 2007; Neubauer et al., 2009; Abd El-Hamid et al., 2016; Persson and Bojesen, 2015). Although it was considered that Gallibacterium infection is a result of an ascending infection (Bojesen et al., 2004), it is more probable that colonization of the reproductive tract develops because of the tissue tropism (Persson and Bojesen, 2015). The prevalence of Gallibacterium is also highly influenced by the biosecurity level. It is by far more prevalent in the flocks kept in facilities with lower biosecurity levels (Bojesen et al., 2011). Our results confirmed the assumption about G. anatis being
an opportunistic pathogen. E. coli was more prevalent and found in all the searched organs, as opposed to G. anatis, which was found only in the pharynx and peritoneum. Epizootiological survey for the prevalence of Galli-bacterium in layers showed similar results, where most isolates originated from upper respiratory tract (Lozica et al., 2018b). Unspecific clinical symptoms and pathological lesions complicate diagnosing Gallibacterium infection. For that reason, it is often misdiagnosed and inaccurately treated. The isolates are usually farm specific and often multi-drug resistant (El-Adawy et al., 2018; Lozica et al., 2018a; Osuna Chávez et al., 2017; Bojesen et al., 2011), which additionally complicates therapy success. With growing antimicrobial resistance rates, therapy will only become a bigger challenge. Therefore, the application of autogenous vaccine in addition to establishing high biosecurity levels is recommended. Due to the problems on these broiler breeder farms and high prevalence rates of both E. coli and G. anatis, implementation of autogenous vaccine was sugges-ted, as well as adjustment of the immunoprophylaxis pro-gram and continuous monitoring.
References
ABD EL-HAMID, H. S., H. F. ELLAKANY, A. A. BEKHEET, A. R. ELBESTAWY, N.MATARIED (2016): Pathogenicity of ten Gallibacterium anatis isolates in commercial broiler chickens. Alexandria J. Vet. Sci. 49 (2), 42-49.
ALISPAHIĆ, M., H. CHRISTENSEN, C. HESS, E. RAZZAZI-FAZELI, M. BISGAARD, M. HESS (2012): MALDI-TOF mass spectrometry confirms clonal lineages of Gallibacterium anatis between chicken flocks. Vet Microbiol. 160, 269-273.
BISGAARD, M. (1977): Incidence of Pasteurella haemolytica in the respiratory tract of apparently healthy chickens and chickens with infectious bronchitis: characterisation of 213 strains. Avian Pathol. 6, 285–292.
BISGAARD, M. (1993): Ecology and significance of Pasteurellaceae in animals. Zentralbl Bakteriol. 279, 7-26.
BOJESEN, A. M., S. S. NIELSEN, M. BISGAARD (2003a): Prevalence and transmission of haemolytic Gallibacterium species in chicken production system with different biosecurity level. Avian Pathol. 32 (5), 503-510.

PREVALENCE OF GALLIBACTERIUM ANATIS AND ESCHERICHIA COLI ON BROILER BREEDER FARMS IN CROATIA 153
BOJESEN, A. M., H. CHRISTENSEN, O. L. NIELSEN, J. E. OLSEN, M. BISGAARD (2003b): Detection of Galli-bacterium spp. in chickens by fluorescent 16S rRNA in situ hybridization. J. Clin. Microbiol. 41 (11), 5167-5172.
BOJESEN, A . M., O. L. NIELSEN, J. P. CHRISTENSEN, M. BISGAARD (2004): In-vivo studies of Gallibacterium anatis infection in chicken. Avian Pathol. 33 (2), 145-152.
BOJESEN, A. M, M. E. VAZQUEZ, F. ROBLES, C. GONZALEZ, E. V. SORIANO (2007): Specific identifi-cation of Gallibacterium by a PCR using primers targeting the 16S rRNAand 23S rRNA genes. Vet. Microbiol. 123 (1-3), 262-268.
BOJESEN, A. M., M. E. VAZQUEZ, F. J. BAGER, D. IFRAH, C. GONZALEZ, F. M. AARESTRUP (2011): Antimicrobial susceptibility and tetracycline resistance determinant geno-typing of Gallibacterium anatis. Vet. Microbiol. 148, 105-110.
CHRISTENSEN, H., M. BISGAARD, A. M. BOJESEN, R. MUTTERS, J. E. OLSEN (2003): Genetic relationships among avian isolates classified as Pasteurella haemolytica, “Actinobacillus salpingitidis” or Pasteurella anatis with proposal of Gallibacterium anatis gen. nov., comb. nov. and description of additional genomospecies within Gallibacterium gen. nov. Int. J. Syst. Evol. Microbiol. 53, 275–287.
CORDONI, G., M. J. WOODWARD, H. WU, M. ALANAZI, T. WALLIS, R. M. LA RAGIONE (2016): Comparative genomics of European avian pathogenic E. coli (APEC). BMC Genomics. 17, 960-981.
DZIVA, F., M. P. STEVENS (2008): Colibacillosis in poultry: unravelling the molcular basis of virulence of avian pahogenic Escherichia coli in their natural hosts. Avian Pathol. 37 (4), 355-366.
EL-ADAWY, H., H. BOCKLISCH, H. NEUBAUER, H. M. HAFEZ, H. MOTZEL (2018): Identification, differentitation and antibiotic susceptibility of Gallibacterium isolates from diseased poultry. Ir. Vet. J. 71(5), 1-10.
JONES, K. H., J. K. THORNTON, Y. ZHANG, M. J. MAUEL (2013): A 5-year retrospective report of Gallibacterium anatis and Pasteurella multocida isolates from chickens in Mississippi. Poul. Sci. 92 (12), 3166-3171.
KRISTENSEN, B.M., D. FREES, A. M. BOJESEN (2011): Expression and secretion of the RTX-toxin GtxA among members of the genus Gallibacterium. Vet. Microbiol. 153, 116-123.
LANDMAN, W. J. M., A. HEUVELINK, J. H. H. VAN ECK (2013): Reproduction of the Escherichia coli peritonitis syndrome in laying hens. Avian Pathol. 42 (2), 157-162.
LOZICA, L., D. HORVATEK TOMIĆ, E. PRUKNER-RADOV-ČIĆ, Ž. GOTTSTEIN (2018a): Antimicrobial resistance of Gallibacterium anatis isolated from layer poultry farms in Croatia. 6th International Veterinary Poultry Congress, 27.02.-1.3.2018., Tehran, Iran, 155.
LOZICA, L., D. HORVATEK TOMIĆ, E. PRUKNER-RADOV-ČIĆ, Ž. GOTTSTEIN (2018b): Prevalence of Gallibacterium anatis isolated from layer poultry farms in Croatia. XVth European Poultry Conference, 17.-21.09.2018., Dubrovnik, Croatia, 197.
MELLATA, M. (2013): Human and avian extraintestinal patho-genic Escherichia coli: Infections, zoonotic risks, and anti-biotic resistance trends. Foodborne Pathog. Dis. 10, 916-932.
NEDELJKOVIĆ, G., L. LOZICA, A. GAVRILOVIĆ, Ž. GOTTSTEIN (2018): Infectious bronchitis virus- neglected (persistent) troublemaker in poultry. Proceedings of 11th International Congress for Veterinary Virology- ESVV 2018. 27.-30.8.2018.,Vienna, Austria.
NEUBAUER, C., M. DE SOUZA-PILZ, A. M. BOJESEN, M. BISGAARD, M. HESS (2009): Tissue distribution of haemolytic Gallibacterium anatis isolates in laying birds with reproductive disorders. Avian Pathol. 38 (1), 1-7.
OSUNA CHÁVEZ, R. F., R. M. MOLINA BARRIOS, J. F. HERNÁNDEZ CHÁVEZ, J. ROBLES MASCAREÑO, J. G. A. ICEDO ESCALANTE, M. ACUÑA YANES (2017): First report of biovar 6 in birds immunized against Gallibacterium anatis in poultry farms located in Sonora, Mexico. Veterinaria México OA. 4 (3), 1-8.
OZAKI, H., T. MURASE (2009): Multiple routes of entry for Escherichia coli causing colibacillosis in commercial layer chickens. J. Vet. Med. Sci. 71 (12), 1685-1689.
PAUDEL, S., M. ALISPAHIC, D. LIEBHART, M. HESS, C. HESS (2013): Assessing pathogenicity of Gallibacterium anatis in a natural infection model: the respiratory and reproductive tracts of chickens are targets for bacterial colonization. Avian Pathol. 42 (6), 527-535.
PAUDEL, S., D. LIEBHART, C. AURICH, M. HESS, C. HESS (2014): Pathogenesis of Gallibacterium anatis ina natural infection model fulfils Koch's postulates: 2. epididymitis and decreased semen quality are the predominant effects in spe-cific pathogen free cockerels. Avian Pathol. 43 (6), 529-534.
PAUDEL, S., M. HESS, C. HESS (2017): Coinfection of Avibacterium paragallinarum and Gallibacterium anatis in specific-pathogen-free chickens complicates clinical signs of infectious coryza which can be prevented by vaccination. Avian Dis. 61, 55-63.
PERSSON, G., A. M. BOJESEN ( 2015): Bacterial determinants of importance in the virulence of Gallibacterium anatis in poultry. Vet. Res. 46 (1), 57.
POULSEN, L. L., I. THØFNER, M. BISGAARD, J. P. CHRISTENSEN, R. HEIDEMANN OLSEN, H. CHRISTENSEN (2017): Longitudinal study of transmission of Escherichia coli from broiler breeders to broilers. Vet. Microbiol. 207, 13-18.
PROCTOR, R. A., C. VAN EIFF, B. C. KAHL, K. BECKER, P. MCNAMARA, M. HERMANN, G. PETERS (2006): Small colony variants: a pathogenic form of bacteria that facilitates persistent and recurrent infections. Nat. Rev. Microbiol. 4 (4), 295-305.
RZEWUSKA, M., E. KARPIŃSKA, P. SZELESZCZUK, M. BINEK (2007): Isolation of Gallibacterium spp. from peacocks with respiratory tract infections. Med. Weter. 63 (11), 1431-1433.
SINGH, S.V., B. R. SINGH, D. K. SINHA, V. KUMAR OR, P. A. ADHANA, M. BHARDWAJ, S. DUBEY (2016): Gallibacterium anatis: An Emerging Pathogen of Poultry Birds and Domiciled Birds. J. Vet. Sci.Techno. 7 (3), 324.

PREVALENCE OF GALLIBACTERIUM ANATIS AND ESCHERICHIA COLI ON BROILER BREEDER FARMS IN CROATIA 154
PROŠIRENOST BAKTERIJA GALLIBACTERIUM ANATIS I ESCHERICHIA COLI NA FARMAMA RODITELJSKIH JATA TEŠKIH LINIJA KOKOŠI U HRVATSKOJ
Sažetak
Gallibacterium (G.) anatis i Escherichia (E.) coli neki su od najčešćih uzročnika peritonitisa, folikulitisa i salpingitisa, kao i poremećaja gornjeg dišnog sustava u peradi. Mogu biti primarni patogeni ili djelovati u koinfekciji s drugim mikroorganizmima kao što su Mycoplasma spp., virus zaraznog bronhitisa (IBV) ili međusobno. Slabije izraženi klinički znakovi i patomorfološke promjene obično prate infekciju bakterijom G. anatis, dok su u slučaju kolibaciloze izraženije promjene. Infekcija može biti kliničkog ili supkliničkog tijeka, ali u oba slučaja uzrokuje velike gubitke u proizvodnji i visoke stope smrtnosti. U ovom istraživanju htjeli smo odrediti učestalost obiju bakterija na dvjema farmama roditeljskih jata teških linija s ciljem procjene epizootiološke situacije u Hrvatskoj. Tijekom istraživanja lešine kokoši i obrisci podrijetlom od živih životinja doneseni su s farmi na Zavod za bolesti peradi s klinikom, gdje je napravljena patomorfološka pretraga. Pretraženo je ukupno 200 obrisaka ždrijela, peritoneuma, jetre, jajovoda i koštane srži podrijetlom od 128 životinja. Obrisci su nasađeni na 5%-tni krvni agar, nakon čega su inkubirani aerobno, na 37 °C tijekom 24 sata. Identifikacija je napravljena na temelju morfoloških karakteristika, biokemijskih testova te metodom MALDI-TOF spektrometrije. Nakon identifikacije uzorci su spremljeni u BHI bujon na -80 °C do daljnjih pretraga. Sveukupno 15,5% obrisaka s obiju farmi bilo je pozitivno na G. anatis. Od toga je 93,5% pozitivnih uzoraka bilo podrijetlom iz ždrijela, dok je ostalih 6,5% podrijetlom iz peritoneuma; 38% uzoraka s obiju farmi bilo je pozitivno na E. coli, a od toga je čak 62,5% uzoraka bilo podrijetlom iz jajovoda. Koinfekcija bakterijom E. coli je dokazana samo u dva slučaja, iako je na obje farme prethodno serološki dokazana Mycoplasma synoviae te su dokazani 793B sojevi IBV. Ovi podaci potvrđuju pretpostavku da je G. anatis oportunistički patogen i prikazuju visoku učestalost E. coli na pretraženim farmama. S obzirom na to da su izolati najčešće specifični za farmu i nerijetko multirezistentni, terapija nije uvijek uspješna. Iz tog razloga se preporuča primjena autogenog cjepiva uz obveznu prilagodbu programa cijepljenja individualnom jatu. Ključne riječi: Gallibacterium anatis, Escherichia coli, roditeljska jata teških linija, autogeno cjepivo, perad

PERADARSKI DANI 2019. 155
ISPITIVANJE PRISUTNOSTI SALMONELLA SPP. U UZORCIMA SVJEŽEG PORCIONIRANOG PILEĆEG MESA U MALOPRODAJI
Zdenko Mlinar, Ivančica Kovaček, Ana Vasiljević, Vedran Prahin
Nastavni zavod za javno zdravstvo „Dr. Andrija Štampar“, Zagreb, Hrvatska
Sažetak
Republika Hrvatska je pristupanjem Europskoj Uniji (EU) postala dio jedinstvenog unutarnjeg tržišta EU, tzv. single-market tržišta. Osnovna ideja takvog jedinstvenog tržišta jest slobodno kretanje ljudi, roba, usluge i novca. Bitnu ulogu u smanjenju i konačnom uklanjanju prepreka slobodnoj trgovini ima uzajamno priznavanje zasnovano na standardizaciji kvalitete u najširem smislu. U tom procesu su uklonjeni mnogi tehnički, zakonski i birokratski zahtjevi. Brojnim tvrtkama je to omogućilo bolju konkurentnost, uslijed čega je došlo i do značajnog pada cijena uz povećanje ponude. Taj proces prati i zajednička legislativa koja za ulogu ima osiguranje načela pravednosti, zaštitu potrošača i ekološku održivost. Iz svega navedenog je jasno da sigurnost hrane unutar svake zemlje članice predstavlja znatno veći izazov. Unatoč želji da jedinstveno tržište EU ima jednake standarde sigurnosti i kvalitete hrane, to se u praksi nije pokazalo lako dostižnim ciljem. Neujednačenost u standardima kvalitete i sigurnosti osobito je izražena pri proizvodnji i u prometu hrane. S obzirom na takvo stanje potreban je povećan oprez koji zahtijeva i sustavni pristup odnosno sustavnu kontrolu. U službenom monitoringu Ministarstva zdravstva na maloprodajnom tržištu sjeverozapadne Hrvatske provedenom 2015. godine prikupljeni su i ispitani uzorci svježeg porcioniranog pilećeg mesa. Primarni cilj ispitivanja je bilo utvrđivanje prisutnosti Salmonella spp, a u tu svrhu je prikupljeno i ispitano 80 uzoraka, od čega je u 2 uzorka dokazana Salmonella spp. Serološkom tipizacijom je utvrđeno da se radilo o S. enterica subsp. enterica serovar Typhimurium i S. enterica subsp. enterica serovar Infantis, što je dodatno potvrđeno u Nacionalnom referentnom laboratoriju za serološku tipizaciju Salmonella spp. izdvojenih iz hrane. Ključne riječi: Salmonella spp., mikrobiološko ispitivanje, monitoring, pileće meso Uvod Republika Hrvatska je pristupanjem Europskoj Uniji postala dio jedinstvenog single-market tržišta EU, što spada u najbolje primjere europske ekonomske integracije. Na tom jedinstvenom tržištu ljudi, roba, usluge i novac se mogu kretati na području EU slobodno kao unutar jedne države. Bitnu ulogu u smanjenju i konačnom uklanjanju prepreka trgovini ima uzajamno priznavanje, stoga su u tom procesu kreiranja jedinstvenog tržišta uklonjeni mnogobrojni tehnički, zakonski i birokratski zahtjevi. Tvrtkama je to omogućilo proširenje poslovanja uz povećanje konku-rentnosti, uslijed čega je došlo i do značajnog pada cijena uz povećanje ponude. Sve to prati legislativa koja za ulogu ima osiguranje načela pravednosti, zaštitu potrošača i ekološku održivost. Unatoč težnji o jedinstvenom tržištu EU koje ima jednake standarde sigurnosti i kvalitete hrane, to je u praksi teško provedivo zbog različitih praktičnih pristupa (unatoč standardizaciji), ali i kulturoloških razlika u zemljama
članicama. Iz navedenog je jasno da sigurnost hrane unutar svake zemlje članice predstavlja znatno veći izazov. U prometu robom, u ovom slučaju hranom, potreban je povećan oprez koji zahtijeva sustavni pristup odnosno sustavnu kontrolu. Takva kontrola je regulirana nacio-nalnom regulativom usuglašenom s europskom legislativom. U monitoringu Ministarstva zdravstva na maloprodajnom tržištu sjeverozapadne Hrvatske provedenom 2015. godine prikupljeni su i ispitani uzorci svježeg porcioniranog pilećeg mesa. Meso peradi je zbog relativno visokog udjela vode u mesu, čak do 75,9% (Medić i sur., 2009.) i nešto višeg pH 5.7-5.96 (Hassanin i sur., 2017.) iznimno povoljan medij za rast salmonela. Sojevi Salmonella su relativno osjetljivi na niske vrijednosti aktiviteta vode (aw <0,94), a ne toleriraju niti veće koncentracije soli, u pravilu <9% (Vlaemynck, 1996.). Cilj ispitivanja je bilo utvrđivanje prisustva Salmonella spp. u pilećem mesu u maloprodaji. Nomenklatura Salmonellae je kompleksna i u osnovnoj podjeli razlikuje dvije vrste, S.
Zdenko Mlinar, univ. spec. techn. aliment.; Mirogojska cesta 16, 10000 Zagreb, Hrvatska; +385 (0)1 4696 246, e-mail: [email protected]

ISPITIVANJE PRISUTNOSTI SALMONELLA SPP. U UZORCIMA SVJEŽEG PORCIONIRANOG PILEĆEG MESA U MALOPRODAJI 156
bongori i S. enterica, koje ukupno broje preko 2541 serovara (Porwollik i sur., 2004.). S. enterica je podijeljena na sljedeće podvrste: S. enterica subsp. enterica, S. enterica subsp. salamae, S. enterica subsp. arizonae, S. enterica subsp. diarizonae, S. enterica subsp. houtenae and S. enterica subsp. indica (Grimont i Weill, 2007.). Salmonellae spadaju u porodicu Enterobacteriaceae, a morfološki ih karakteriziraju mali, pravilni bojenjem po Gramu negativni (-) štapići (0,7-1,5 x 2-5 μm) koji ne stvaraju spore (Holt i sur., 1994.). Većina salmonela je pokretna, dok se jedan serovar ističe kao nepokretan – S. Pullorum (Vlaemynck, 1996.). Što se ostalih bitnih obilježja tiče, Salmonellae su glukoza pozitivne (+), saharoza negativne (-), oksidaza negative (-), katalaza pozitivne (+), indol negativne (-), citrat pozitivne (+), lizin pozitivne (+), urea negativne (-) te su većinom H2S pozitivne (+).
Materijali i metode
Opis uzoraka Ispitivanje je provedeno na uzorcima pilećeg mesa por-cioniranog i pakiranog prema različitim dijelovima (batak, krila, batak sa zabatakom, zabatak, prsa, hrbat). Podrijetlo uzoraka Svi uzorci koji su bili iz uvoza bili su uvezeni iz Italije od različitih proizvođača. Većina ispitanih uzoraka (n=42) je bila uzorkovana u subjektima u poslovanju s hranom koji se bave samo prodajom mesa i mesnih prerađevina, dok je manji dio uzoraka izdvojen iz maloprodajnih trgovina mješovitom robom (n=38). Oba nesukladna uzorka su podrijetlom iz hrvatskih klaonica; jedan uzorak je uzorkovan iz maloprodajne trgovine mje-šovitom robom, dok je drugi uzorak uzorkovan u spe-cijaliziranoj trgovini tvrtke registrirane za proizvodnju, preradu, pakiranje i distribuciju purećeg i pilećeg mesa. Prisutnost Salmonella spp. ispitivala se metodom kultivacije iz uzoraka i to prema normi HRN EN ISO 6579:2003, Mikrobiologija hrane i stočne hrane - Horizontalna metoda
za otkrivanje Salmonella spp. (ISO 6579:2002; EN ISO 6579:2002) (HRN EN ISO 6579, 2013.). Ispitivanje je provedeno uz postupke neselektivnog i selektivnog prednamnažanja, nacjepljivanja na selektivne i neselektivne hranjive podloge te potvrdne biokemijske i serološke testove prema propisanom protokolu prikazanom u slici 1. Za neselektivno prednamnažanje je korištena puferirana peptonska voda (PPV), a za selektivno namnažanje su korišteni Rappaport-Vassiliadis soja bujon (RVS) i Muller-Kauffmann tetrathionat-novobiocin bujon (MKTTn). Za nacjepljivanje su korišteni Xylose Lysine Deoxycholate agar (XLD) i Salmonella Shigella agar (SS). Biokemijski testovi koji su ključni u identifikaciji Sal-monella spp. su prikazani u tablici 1., a sastavni su dio testova API 20E i ID 32E (bioMérieux) korištenih u ovom ispitivanju. Tablica 1. Biokemijski testovi koji su ključni u identifikaciji
Salmonella spp. Table 1. Main biochemical tests for Salmonella spp.
identification
Glukoza / Glucose + Saharoza / Saccharose - Urea / Urea - L-Lizin / L-Lysine + Serološkom tipizacijom je utvrđeno da se radilo o S. enterica subsp. enterica serovar Typhimurium i S. enterica subsp. enterica serovar Infantis, što je dodatno potvrđeno u Nacionalnom referentnom laboratoriju za serološku tipizaciju Salmonella spp. izdvojenih iz hrane.
Rezultati
Od prikupljenih i ispitanih 80 uzoraka porcioniranog i pakiranog pilećeg mesa, u 78 (97,5%) uzoraka nije utvrđena
Slika 1. Protokol za dokazivanje prisutnosti Slamonella spp. prema HRN EN ISO 6579:2017. Figure 1. Protocol for verification of the presence of Slamonella spp. according to HRN EN ISO 6579:2017.

ISPITIVANJE PRISUTNOSTI SALMONELLA SPP. U UZORCIMA SVJEŽEG PORCIONIRANOG PILEĆEG MESA U MALOPRODAJI 157
prisutnost Salmonella spp., odnosno ti uzorci su bili su-kladni. U 2 (2,5%) ispitana uzorka utvrđena je prisutnost Salmonella spp. Izolirani sojevi su podvrgnuti daljnjim sero-loškim testovima te je utvrđeno da se radilo o Salmonella enterica serovar Typhimurium i Salmonella enterica subsp. enterica serovar Infantis.
Rasprava i zaključci
Ispitivanje je provedeno na 80 uzoraka porcioniranog i pakiranog pilećeg mesa. Od ukupnog broja ispitanih uzo-raka, 75 (93,75%) uzoraka je bilo domaćeg podrijetla, dok je 5 (6,25%) uzoraka bilo iz uvoza s tržišta EU. Rezultati ispitivanja pokazali su prisutnost Salmonella spp. tek u 2,5% ispitanih uzoraka. Slična ispitivanja provedena su u drugim zemljama te su pokazala drugačije rezultate. Primjerice, prema rezultatima ispitivanja provedenih u Ruskoj Federaciji, učestalost iznosi čak 31,5% (n=698) na ukupan broj ispitanih uzoraka (Alali i sur., 2012.). Rezultati ispitivanja provedenog u Pakistanu pokazali su sličnu situaciju kao u Rusiji s obzirom na to da je opća učestalost iznosila 33% (n=200), tj. 30% (n=100) u smrznutom pilećem mesu i 36% (n=100) u svježem pilećem mesu (Samad i sur., 2018.). Iz ostalih dostupnih rezultata istraživanja može se vidjeti da postoje ozbiljno ugrožena tržišta s iznimno visokom stopom učestalosti Salmonella spp. u pilećem mesu, Kolumbija 37% (Donado-Godoy i sur., 2014.), Kina 52,2% (n=1152) (Yang i sur., 2011.) i Tajland 57% (n=72) (Padungtod i Kaneene, 2006.). Situacija u zemljama članicama EU pruža jednako zanimljive rezultate; od iznimno visokih stopa učestalosti se ističu Portugal 60% (n=60) (Antunes i sur., 2003.), Belgija 36% (n=772) (Uyttendaele i sur., 1999.) i Španjolska 35,8% (n=198) (Pointon i sur., 2008.). Nešto umjerenije stope gdje se ukla-paju rezultati ovog ispitivanja zabilježene su na Novom Zelandu 3% (n=232) (Wong i sur., 20017.), u SAD-u 4,2% (n=212) (Zhao i sur., 2001.), a od zemalja EU u Ujedi-njenom Kraljevstvu 4% (n=877) (Meldrum i Wilson, 2007.), Irskoj 5,1% (Madden i sur., 2011.) i Poljskoj 6,5% (n=200) (Radkowski i Zdrodowska, 2016.). Iz prikazanih rezultata, izuzevši rezultate zemalja izvan EU, nedvojbeno je jasno da postoji velika i nesrazmjerna razlika u standardima pojedinih članica EU, a samim time je pot-vrđena teza kako je teško ostvariva ideja o jedinstvenim standardima za sve članice Unije s obzirom na single-market tržište. Jedan od načina za zadovoljavanje visoko zadanih kriterija je ujednačeno provođenje preventivnih mjera te nadzor provedbe istih. Preventivne mjere je nužno provoditi u cijelom lancu od primarne proizvodnje do prerade i distribucije svježeg porcioniranog mesa. Tek takvim standardiziranim pristupom u osiguranju kvalitete i sigurnosti u svim fazama procesa se može ostvariti stvarna prevencija pojave većeg broja ne-sukladnih uzoraka. Apsolutnu odsutnost Salmonella spp. ne treba očekivati niti je to realan zahtjev. Prikladan cilj bi bilo postavljanje zahtjeva u smjeru zadržavanja relativno niskog postotka pojavnosti Salmonella spp., naročito izrazito
patogenih sojeva koji mogu ugroziti zdravstvenu ispravnost proizvoda, ali što je još opasnije, stvoriti realnu opasnost za zdravlje krajnjih potrošača.
Literatura
ALALI, W. Q., R. GAYDASHOV, E. PETROVA, A. PANIN, O. TUGARINOV, A. KULIKOVSKII, DZ. MAMLEEVA, I. WALLS, M. DOYLE (2012): Prevalence of Salmonella on Retail Chicken Meat in Russian Federation. Journal of food protection. 75. 1469-73. 10.4315/0362-028X.JFP-12-080.
ANTUNES, P., C. REU, J. C. SOUSA, L. PEIXE, N. PESTANA (2003): Incidence of Salmonella from poultry products and their suscepti-bility to antimicrobial agents. Int. J. Food Microbiol., 82, 97–103.
DONADO-GODOY P., V. CLAVIJO, M. LEÓN, A. AREVALO, R. CASTELLANOS, J. BERNAL, M. A. TAFUR, M. V. OVALLE, W. Q. ALALI, M. HUME, J. J. ROMERO-ZUÑIGA, I. WALLS, M.P. DOYLE (2014): Counts, serovars, and antimicrobial resistance phenotypes of Salmonellaon raw chicken meat at retail in Colombia. J. Food Prot., 77 (2), 227-235.
GRIMONT P. A. D., WEILL, F. X. (2007): Antigenic formulae of the Salmonella serovars, WHO Collaborating Centre for Reference and Research on Salmonella, Institut Pasteur, Paris, France
HASSANIN S. F., MOHAMED, HASSAN, A., SHALTOUT, F., SHAWQY, N. A., ABD-ELHAMEED, A. G. (2017): Chemical criteria of chicken meat., 33. 457-464.
HOLT, J. G., N. R. KRIEG, P. H. A. SNEATH, J. T. STALEY, S. T. WILLIAMS (1994): Bergey`s Manual of Determinative Bacteriology, 9th edition, Williams & Wilkins, Baltimore, Maryland
HRN EN ISO 6579:2003 - Mikrobiologija hrane i stočne hrane -- Horizontalna metoda za otkrivanje Salmonella spp (ISO 6579:2002; EN ISO 6579:2002)
MADDEN R. H., L. MORAN, P. SCATES, J. McBRIDE, C. KELLY (2011): Prevalence of Campylobacter and Salmonella in raw chicken on retail sale in the republic of Ireland. J. Food Prot., 74 (11), 1912-1916.
MEDIĆ H., S. VIDAČEK, K. SEDLAR, V. ŠATOVIĆ, T. PETRAK (2009): Utjecaj vrste i spola peradi te tehnološkog procesa hlađenja na kvalitetu mesa. Meso, XI ( 4), 222-231.
MELDRUM, R. J., I. G. WILSON (2007): Salmonella and Campylobacter in United Kingdom retail raw chicken in 2005. J. Food Prot., 70, 1937-1939.
PADUNGTOD, P., J. B. KANEENE (2006): Salmonella in food animals and humans in northern Thailand. Int. J. Food Microbiol., 108, 346-354.
POINTON, A., M. SEXTON, P. DOWSETT, T. SAPUTRA, A. KIERMEIER, M. LORIMER, G. HOLDS, G. ARNOLD, D. DAVOS, B. COMBS, S. FABIANSSON, G. RAVEN, H. MCKENZIE, A. CHAPMAN, J. SUMNER (2008): Abaseline survey of the microbiological quality of chicken portions andcarcasses at retail in two Australian states (2005 to 2006). J. FoodProt., 71,1123-1134.
PORWOLLIK S., E. F. BOYD, C. CHOY, P. CHENG, L. FLOREA, E. PROCTOR, M. MCCLELLAND (2004): Cha-racterization of Salmonella enterica Subspecies I Genovars by Use of Microarrays. J Bacteriol., 186 (17), 5883-5898.
RADKOWSKI M., B. ZDRODOWSKA (2016): Prevalence of Salmonella spp. In broiler chicken meat. Medycyna Wetery-naryjna., 72. 516-519. 10.21521/mw.5542.

ISPITIVANJE PRISUTNOSTI SALMONELLA SPP. U UZORCIMA SVJEŽEG PORCIONIRANOG PILEĆEG MESA U MALOPRODAJI 158
SAMAD, A., A. FERHAT, T. ZUNERA, A. ZAFAR, A. IRSHAD, G. P. SIMON, N. NIGHAT, A. TAUSEEF, A. RAZIQ, I. S. SHEIKH, M. N. BARECH, O. POKRYSHKO, M. Z. MUSTAFA (2018): Prevalence of Salmonella spp. in chicken meat from Quetta retail outlets and typing through multiplex PCR. Romanian Biotechnological Letters. 10.26327/RBL2018.210.
UYTTENDAELE, M., P. DE TROY, J. DEBEVERE (1999): Incidence of Salmonella, Campylobacter jejuni, Campylo-bacter coli and Listeria monocytogenes in poultry carcasses and different types of poultry products for sale on the Belgian retail market. J. Food Prot., 62, 735–740.
VLAEMYNCK G. (1996): Značaj patogenih mikroorganizama u sirovom mlijeku. Mljekarstvo, 46 (3), 216-231
WONG, T. L., C. NICOL, R. COOK, S. MACDIARMID (2007): Salmonellain uncooked retail meats in New Zealand. J. Food Prot., 70, 1360-1365.
YANG, B., M. XI, X. WANG, S. CUI, T. YUE, H. HAO, Y. WANG, Y. CUI,W. Q. ALALI, J. MENG, I. WALLS, D. M. WONG, M. P. DOYLE (2011): Prevalence of Salmonellaon raw poultry at retail markets in China. J. Food Prot., 74, 1724-1728.
ZHAO, C., B. GE, J. DE VILLENA, R. SUDLER, E. YEH, S. ZHAO, D. G.WHITE, D. WAGNER, AND J. MENG (2001): Prevalence of Campylobacter spp., Escherichia coli, and Salmonella serovars in retail chicken, turkey, pork, and beef from the Greater Washington, D.C. area. Appl.Environ. Microbiol., 67, 5431-5436.
TESTING FOR SALMONELLA SPP. PRESENCE IN RETAIL FRESH PORTIONED CHICKEN MEAT SAMPLES
Summary
Upon accession to the European Union (EU), Republic of Croatia has become part of the united internal EU market, so-called single market. The main idea of such an integral market is free movement of persons, goods, services and capital. Mutual recognition based on quality standardization in a broad sense is of utmost importance for reduction and eventual elimination of obstacles to free trade. In this process, many technical, legal and bureaucratic requirements have been eliminated, thus enabling higher competitiveness to numerous companies, which in turn has resulted in considerable price drop along with an increase in supply. The process has been accompanied by common legislative, which has to ensure the principles of equity, consumer protection and ecologic sustainability. All this clearly shows that food safety within each EU Member Country is a much greater challenge. In spite of the equal food safety and quality standards desired on the integral EU market, it has proven to be hardly achievable in practice. Lack of uniformity in quality and safety standards is especially pronounced in food production and turnover. Therefore, great caution is warranted, which requires systematic approach and surveillance. During official monitoring launched by the Ministry of Health on retail market in north-east Croatia in 2015, samples of fresh portioned chicken meat were collected and examined. The primary goal of testing was to determine the presence of Salmonella spp., which was verified in 2 of 80 samples collected. Serotyping revealed the presence of S. enterica subsp. enterica serovar Typhimurium and S. enterica subsp. enterica serovar Infantis, additionally verified at the National Reference Laboratory for serotyping of Salmonella spp. isolated from food. Key words: Salmonella spp., microbiological examination, monitoring, chicken meat

PERADARSKI DANI 2019. 159
MONITORING REZIDUA VETERINARSKO-MEDICINSKIH PROIZVODA I OSTALIH TVARI U PERADI I JAJIMA U EUROPSKOJ UNIJI U RAZDOBLJU 2009. - 2016. GODINE
Damir Pavliček, Marija Denžić Lugomer
Hrvatski veterinarski institut, Veterinarski zavod Križevci, Križevci, Hrvatska
Sažetak
Prisutnost zabranjenih tvari, rezidua veterinarsko-medicinskih proizvoda ili kemijskih kontaminanata u hrani predstavlja opasnost za zdravlje ljudi. Zakonodavni okvir Europske unije propisuje najveće dozvoljene koncentracije u hrani i programe monitoringa za kontrolu ovih supstanci u hranidbenom lancu. U ovom radu daje se detaljan pregled rezultata monitoringa uzoraka peradi i jaja na prisutnost tvari iz skupina A (tvari koje imaju anabolički učinak i zabranjene tvari) i B (veterinarsko-medicinski proizvodi i kontaminanti) u Europskoj uniji u razdoblju od 2009. do 2016. godine. Obrađeni podaci odnose se na uzorke prikupljene kroz ciljano uzorkovanje prema Direktivi Vijeća 96/23/EZ, dok su rezultati uzoraka dobivenih drugim strategijama uzorkovanja (sumnjivi, uvozni ili “ostalo”) koje ne prate plan monitoringa prikazani odvojeno. Najveći broj analiziranih uzoraka peradi i jaja zabilježen je u 2014. godini, od čega je najviše pretraga provedeno na rezidue skupine B1 (antibakterijske tvari - antibiotici, sulfonamidi, kinolini) te A6 (zabranjene tvari: nitrofurani, nitroimidazoli i dr.) i B2b (kokcidiostatici) podgrupe tvari, a najmanji broj uzoraka peradi je bio 2010. odnosno jaja 2011. godine. U 2009. godini prijavljen je najveći broj nesukladnih uzoraka, većinom na prisutnost kokcidiostatika, a otada je primijećen pad u učestalosti nesukladnih uzoraka, što je vrlo vjerojatno rezultat mjera koje su uslijedile uvođenjem Direktive Komisije 2009/8/EZ. U Hrvatskoj kao članici Europske unije od 2013. godine također se provodi godišnji monitoring rezidua u hrani životinjskog podrijetla i u ovom radu će se prikazati dosad izvješteni podaci za perad i jaja.
Ključne riječi: monitoring, rezidue, perad, jaja, Europska unija, Hrvatska Uvod
Definicija rezidua u legislativi Europske Unije (EU) uklju-čuje farmakološki aktivne tvari, njihove metabolite i druge tvari prenesene u životinjske proizvode, koje predstavljaju rizik za ljudsko zdravlje (Serratosa i sur., 2006.). Prisutnost rezidua u dozvoljenim količinama odnosno njihov potpun izostanak u hrani životinjskog podrijetla rezultat je uspješne suradnje svih strana (poljoprivrednici, veterinari i dr.) uključenih u hranidbeni lanac "od polja do stola". Njihova primarna zadaća je osiguravanje kvalitete i sigurnosti hrane na tržištu upotrebom provjerenih farmakološki aktivnih tvari u poljoprivrednoj proizvodnji. Također, vrlo je bitno pri-državanje propisanih doza i poštivanje karence kod životinja čija se tkiva i/ili proizvodi upotrebljavaju za prehranu ljudi. Ako za neki veterinarsko-medicinski proizvod (VMP) nije određena karenca tada on sadrži takvu farmakološki djelatnu tvar za koju je dokazano da u bilo kojoj količini nije štetna za ljudsko zdravlje ili upravo suprotno, da je i u najmanjoj količini štetna za zdravlje ljudi. U tom slučaju u potpunosti je zabranjena za primjenu na životinjama koje se ili čiji se
proizvodi koriste u prehrani ljudi. Razlike u zahtjevima sigurnosti hrane između raznih zemalja uvelike su utjecale na trgovinsku razmjenu hrane životinjskog podrijetla pa su stoga donesene nove uredbe kojima se potrošačima u EU jamči sigurnost hrane na tržištu, bez obzira je li proizvedena u EU ili trećim zemljama. Direktivom Vijeća 96/23/EZ utvrđene su mjere za sustavno praćenje određenih tvari i nji-hovih rezidua u živim životinjama i proizvodima životinj-skog podrijetla (EC, 1996.). Dodatno, Odlukom Vijeća 97/747/EZ su određene razine i učestalosti uzorkovanja propisanih Direktivom Vijeća 96/23/EZ (EC, 1997.). Ured-bom Komisije (EZ) br. 37/2010 propisane su najviše dopu-štene koncentracije rezidue VMP u životinjama za ljudsku prehranu i njihovim proizvodima (EC, 2010.). Uredba Ko-misije (EZ) 1881/2006 utvrđuje najviše dopuštene količine određenih kontaminanata u hrani životinjskog podrijetla (EC, 2006.). Nadležna tijela u svakoj državi članici EU izrađuju programe monitoringa u kojima su navedene sve obuhvaćene životinjske vrste i njihovi proizvodi te broj i na-čin odabira uzoraka koji se uzimaju za kontrolu na prisutnost tvari iz skupina A i B prema tablici 1. (EC, 1996.).
Damir Pavliček, mag.chem., Hrvatski veterinarski institut, Veterinarski zavod Križevci, Laboratorij za analitičku kemiju i rezidue, Ivana Zakmardija Dijankovečkog 10, 48260 Križevci; Tel: +385 (0)48 279 268; Fax: + 385 (0)48 279 255; e-mail: [email protected]

MONITORING REZIDUA VETERINARSKO-MEDICINSKIH PROIZVODA I OSTALIH TVARI U PERADI I JAJIMA U EUROPSKOJ UNIJI U RAZDOBLJU 2009. - 2016. GODINE
160
Tablica 1. Popis tvari koje se pretražuju u sklopu programa monitoringa rezidua Table 1. List of substances included in the residue monitoring program
Skupina A – tvari koje imaju anabolički učinak i zabranjene tvari Group A – substances having anabolic effect and unauthorized substances
A1 Stilbeni, derivati stilbena, soli i esteri stilbena Stilbenes, stilbene derivatives, and their salts and esters A4 Laktoni rezorcilne kiseline, uključujući zeranol
Resorcylic acid lactones, including zeranol
A2 Antitireoidne tvari / Antithyroid agents A5 Beta-agonisti / Beta-agonists
A3 Steroidi / Steroids A6 Zabranjene tvari: nitrofurani, nitroimidazoli i dr. Unauthorized substances: nitrofurans, nitroimidazoles, etc.
Skupina B – VMP i kontaminanti / Group B – Veterinary drugs and contaminants
B1 Antibakterijske tvari, uključujući sulfonamide i kinolone Antibacterial substances, including sulfonamides, quinolones
B2 Drugi veterinarski lijekovi (VMP) / Other veterinary drugs B3 Druge tvari i kontaminanti Other substances and environmental contaminants
B2a Antiparazitici / Anthelmintics B3a Organoklorni spojevi, uključujući PCB-ove Organochlorine compounds including PCBs
B2b Kokcidiostatici, uključujući nitroimidazole Anticoccidials, including nitroimidazoles B3b Organofosforni spojevi
Organophosphorus compounds
B2c Karbamati i piretroidi Carbamates and pyrethroids B3c Kemijski elementi (teški metali)
Chemical elements (heavy metals) B2d Sedativi / Sedatives B3d Mikotoksini / Mycotoxins
B2e Nesteroidni protuupalni lijekovi Non-steroidal anti-inflammatory drugs (NSAIDs) B3e Bojila / Dyes
B2f Druge farmakološki aktivne tvari (kortikosteroidi) Other pharmacologically active substances (corticosteroids) B3f Drugo / Others
Državni programi monitoringa rezidua (DPMR) moraju biti pripremljeni na način da se zadovolje minimalni kriteriji prilikom uzorkovanja (vrsta, spol, starost i dr.), a uz ciljane mogu uključivati i sumnjive uzorke kod kojih je već u sklopu prethodnih kontrola utvrđena nesukladnost ili postoji sumnja i dokazi o korištenju zabranjenih tvari odnosno nepoštivanju vremena karence za propisane VMP (EFSA, 2011.). Zahtjevi za analitičke metode kojima se pretražuju službeni uzorci i tumače rezultati određeni su Odlukom Komisije 2002/657/EZ (EC, 2002.). Dobiveni rezultati državnih monitoringa rezidua pohranjuju se u bazu podataka Glavne uprave za zdravlje i sigurnost hrane (DG SANTE), čija je zadaća cjelokupna koordincija prikupljanja podataka za rezidue od zemalja članica EU te provjera jesu li oni u skladu s donesenim planovima monitoringa. U ovom radu prikazat će se stanje rezidua u peradi i jajima u EU u razdoblju 2009.-2016. godine. Napravljen je osvrt na broj analiziranih uzoraka prikupljenih putem držav- nih programa monitoringa te udio nesukladnih uzoraka i njihova podjela prema skupinama tvari kojima pripadaju spojevi detektirani u previsokim koncentracijama. Prikazani su i statistički podaci za Hrvatsku od ulaska u EU 2013. godine.
Materijali i metode
U okviru članka 31. Uredbe (EZ) br. 178/2002 Europska Zajednica je zatražila Europsku agenciju za sigurnost hrane (EFSA) da objedini rezultate nacionalnih monitoringa u EU dobivenih na temelju uputa Direktive Vijeća 96/23/EZ i objavljuje ih u obliku godišnjih izvješća (EFSA, 2018.). Iz-vješća EFSA-e o rezultatima monitoringa rezidua veteri-narsko-medicinskih proizvoda i drugih tvari u životinjama i životinjskim proizvodima poslužila su za izradu ovog rada. Statistički su obrađeni dostupni podaci o monitoringu uzo-raka peradi i jaja u EU od 2009. do 2016. godine pri-kupljenih kroz ciljano uzorkovanje. Uspoređen je broj analiziranih i nesukladnih uzoraka za odabrano razdoblje te njihova podjela prema skupinama tvari na koje su izvršene pretrage. Uz ciljane uzorke prikazani su i podaci o uzorcima prikupljenih drugim strategijama uzorkovanja (sumnjivi, uvozni i ostali), koji iz različitih razloga nisu reprezentativni za procjenu prisutnosti rezidua u hrani životinjskog podri-jetla, u zemljama članicama EU. Važno je napomenuti kako broj nesukladnih uzoraka ne odgovara nužno broju nesukladnih rezultata s obzirom na to da se u jednom uzorku može detektirati i više tvari u razi-nama iznad najveće dopuštene količine (NDK). Također nije dostupna informacija o točnoj vrsti matriksa pojedinog uzorka peradi, što onemogućava detaljniju statističku obradu i identifikaciju uzoraka nesukladnih na više tvari.

MONITORING REZIDUA VETERINARSKO-MEDICINSKIH PROIZVODA I OSTALIH TVARI U PERADI I JAJIMA U EUROPSKOJ UNIJI U RAZDOBLJU 2009. - 2016. GODINE
161
Rezultati
Perad U sklopu nacionalnih monitoring programa, prema Direktivi Vijeća 96/23/EZ, minimalan broj uzoraka za svaku kate-goriju peradi mora odgovarati 1 na 200 t godišnje proiz-vodnje (EC, 1996.). Ako godišnja proizvodnja premašuje 5000 t, tada je minimum 100 uzoraka za svaku skupinu tvari. Iz tablice 2. vidljivo je kako je u razdoblju 2009.-2016. godine na razini ukupnog broja zemalja EU postavljeni kriterij zadovoljen (EFSA, 2018.).
Uzorci peradi se najviše kontroliraju na prisutnost zabra-njenih tvari podskupine A6 (~29%), antibakterijskih tvari koje pripadaju podskupini B1 (~27%) te kokcidiostatika iz podskupine B2b (~12%). Najveći broj nesukladnih uzoraka peradi zabilježen je 2009. godine kada se od njih 185 čak 131 odnosilo na skupinu tvari B2b, što je činilo 2,05% svih analiziranih uzoraka iz te skupine odnosno 70,8% svih nesukladnih uzoraka u toj godini (EFSA, 2011.; 2012.; 2013.; 2014.; 2015.; 2016.; 2017.; 2018.). Ako se gleda pre-ma skupinama tvari može se zaključiti kako je kloramfeni-kol najzastupljenija supstanca iz podskupine A6 u nesuklad-nim uzorcima kroz promatrano razdoblje. Godine 2011. u Nizozemskoj je detektiran Estradiol-17-Beta iz podskupine tvari A3 u šest uzoraka od ukupno 88 koliko ih je ana-lizirano na tu skupinu tvari, što predstavlja izolirani slučaj s obzirom na to da su u ostalih sedam godina u EU sve skupa prijavljena tek dva steroidna spoja u ciljanim uzorcima peradi. Doksiciklin i enrofloksacin su glavni predstavnici podskupine B1 tvari što se tiče prisutnosti njihovih rezidua u peradi. Kokcidioza predstavlja najznačajniju bolest inten-zivne proizvodnje peradi, koja nije moguća bez kontrole kokcidija kokcidiostaticima (Varenina i sur., 2017.) pa iz tog razloga tvari iz ove podskupine spojeva u uzorcima peradi tijekom nacionalnog monitoring programa bivaju utvrđene u najvećem broju. Nikarbazin je kokcidiostatik koji je 2009. i 2010. godine u peradi prijavljen čak 106 odnosno 46 puta u koncentracijama iznad dopuštenih, u čemu su prednjačili
uzorci s područja Ujedinjenog Kraljevstva gdje je najveća proizvodnja peradi, a samim time i najveći broj uzoraka uključenih u monitoring (EFSA, 2011.; 2012.). Zgodno je spomenuti, također, kako je 2009. godine u Portugalu 11% uzoraka pregledanih na podskupinu B2b bilo nesukladno zbog rezidua nikarbazina, a u Francuskoj čak i više, 21%. U idućim godinama ta brojka je drastično pala i prijavljena su sve skupa tek dva slučaja u EU. Od spojeva B2b valja još izdvojiti lasalocid i maduramicin koji se pronađu u peradi gotovo redovito svake godine u malo većem broju od osta-lih, a posljednjih godina tu spada i salinomicin (EFSA, 2011.; 2012.; 2013.; 2014.; 2015.; 2016.; 2017.; 2018.).
Slika 1. Raspodjela nesukladnih uzoraka peradi prema
podskupinama tvari kojima pripadaju rezidue detektirane u višim koncentracijama od dozvo-ljenih u Europskoj Uniji od 2009. do 2016. godine
Figure 1. Distrubution of non-compliant poultry samples according to substance subgroups to which residues detected in higher concentrations than permitted in the European Union belonged during the 2009-2016 period
Tablica 2. Proizvodnja peradi i broj ciljanih i nesukladnih uzoraka u razdoblju 2009.-2016. Table 2. Production of poultry and number of targeted and non-compliant samples during the 2009 -2016 period
Godina (broj zemalja EU) Year (Member States number)
Proizvodnja (t) Production (t)
Ciljani uzorci Targeted samples
Analizirani uzorci / 200 t Samples tested / 200 ta
Nesukladni uzorci Non-compliant samples
2009 (EU 27) 11,383,434 61,989 1.00 185 2010 (EU 27) 11,804,262 61,259 1.08 105 2011 (EU 27) 12,417,108 65,942 1.12 52 2012 (EU 27) 12,845,333 68,770 1.11 54 2013 (EU 28) 12,930,555 71,186 1.11 58 2014 (EU 28) 12,909,837 72,486 1.12 69 2015 (EU 28) 13,394,013 71,223 1.10 49 2016 (EU 27b) 12,239,495 64,501 1.10 48
(a) izračunato prema proizvodnji iz prethodne godine / (a) in relation to the production in the previous year (b) 2016. rezultati iz Francuske nisu dostupni. / (b) 2016 results from France not available

MONITORING REZIDUA VETERINARSKO-MEDICINSKIH PROIZVODA I OSTALIH TVARI U PERADI I JAJIMA U EUROPSKOJ UNIJI U RAZDOBLJU 2009. - 2016. GODINE
162
Uzorkovanje i analiza sumnjivih uzoraka peradi u razdoblju 2009.-2016. godine pokazala se potpuno opravdana ako se promotri postotak nesukladnih uzoraka. Recimo, 2011. Go-dine, metronidazol (A6) je detektiran u Njemačkoj 54 puta (17,3%), a 2012. i 2013. godine u Grčkoj su AMOZ (A6) i AOZ (A6) prijavljeni u velikom postotku uzoraka (EFSA, 2013.; 2014.; 2015.). Iz podskupine tvari B1 ponovno su najviše puta detektirani spojevi doksiciklin i enrofloksacin, kao i kod ciljanog uzorkovanja, uz oksitetraciklin koji se ponajviše pojavljivao u prvim godinama praćenja (EFSA, 2011.; 2012.; 2013.; 2014.; 2015.; 2016.; 2017.; 2018.). Salinomicin je bio najzastupljeniji kokcidiostatik u sumnji-vim uzorcima peradi. Što se tiče uzoraka iz uvoza i onih prikupljenih drugim programima monitoringa dostupno je
manje podataka. Ono što valja istaknuti je pojavnost spoja klopidola (B2b) u uvoznim uzorcima peradi koji je 2012. godine zabilježen u tri zemlje EU (Cipar, Njemačka i Irska) u otprilike 30% pregledanih uzoraka. Također, u razdoblju od 2010. do 2013. godine u Njemačkoj je primijećen porast broja uzoraka peradi iz uvoza koji sadrže živu (3%-13%). Jaja Broj uzoraka koje je potrebno uzorkovati svake godine u sklopu nacionalnih monitoring programa mora biti barem 1 na 1000 t godišnje proizvodnje jaja, a pritom je minimum barem 200 uzoraka (EC, 1996.). Iz tablice 3. vidljivo je kako je u razdoblju 2009.-2016. godine na razini ukupnog broja zemalja EU zadovoljen postavljeni kriterij (EFSA, 2018.). Uzorci jaja se najviše kontroliraju na prisutnost zabranjenih tvari podskupine A6 (~28%), antibakterijskih tvari koje pri-padaju podskupini B1 (~35%), kokcidiostatika iz podsku-pine B2b (~30%) te organoklornih spojeva podskupine B3a (~15%). Najveći broj nesukladnih uzoraka jaja zabilježen je 2009. godine kada se 43 od ukupno 50 odnosilo na skupinu kokcidiostatika, što je činilo 1,19% svih analiziranih uzoraka iz te skupine odnosno 86% svih nesukladnih uzoraka u toj godini (EFSA, 2011.; 2012.; 2013.; 2014.; 2015.; 2016.; 2017.; 2018.). Iako je tijekom godina uočen pad u broju
nesukladnih uzoraka jaja, 2015. i 2016. godine se taj broj ponovno približio maksimumu iz 2009., ali su pritom oni više raspodijeljeni po skupinama tvari. Godine 2015. su kokcidiostatici tako činili 57%, antibakterijske tvari 18%, a organoklorni spojevi 20% svih nesukladnih uzoraka jaja. U razdoblju 2009.-2016. godine u ciljanim uzorcima jaja re-zidue tvari iz skupine A6 su prijavljene tek četiri puta, i to kloramfenikol 2014. (dva uzorka) i 2015. godine (dva uzorka) u Latviji. Doksiciklin i enrofloksacin su najčešće antibakterijske tvari prisutne u jajima, a iz podskupine tvari B2b to su lasalocid, maduramicin, narazin i salinomicin. Dioksini i posebice zbroj dioksina i dioksinima sličnih PCB-a (WHO-PCDD/F-PCB-TEQ) detektirani su također u cilja-nim uzorcima jaja tijekom promatranog razdoblja.
Slika 2. Raspodjela nesukladnih uzoraka jaja prema pod-
skupinama tvari kojima pripadaju rezidue detek-tirane u višim koncentracijama od dozvoljenih u Europskoj Uniji od 2009. do 2016. godine.
Figure 2. Distrubution of non-complaint egg samples accor-ding to substance subgroups to which residues detected in higher concentrations than permitted in the European Union belonged during the 2009-2016 period.
Tablica 3. Proizvodnja jaja i broj ciljanih i nesukladnih uzoraka u razdoblju 2009.-2016. Table 3. Production of eggs and number of targeted and non-compliant samples during the 2009-2016 period
Godina (broj zemalja EU) Year (Member States number)
Proizvodnja (t) Production (t)
Ciljani uzorci Targeted samples
Analizirani uzorci / 1000 t / Samples tested / 1000 ta
Nesukladni uzorci Non-compliant samples
2009 (EU 27) 6,137,732 13,031 2.2 50 2010 (EU 27) 6,101,039 12,715 2.1 20 2011 (EU 27) 6,136,691 12,248 2.0 38 2012 (EU 27) 6,070,174 12,596 2.1 23 2013 (EU 28) 6,070,334 13,323 2.2 26 2014 (EU 28) 6,271,679 13,391 2.2 29 2015 (EU 28) 6,255,410 13,158 2.1 40 2016 (EU 27b) 5,424,380 12,700 2.4 44
(a) izračunato prema proizvodnji iz prethodne godine. / (a) in relation to the previous year production. (b) 2016. rezultati iz Francuske nisu dostupni. / (b) 2016 results from France not available

MONITORING REZIDUA VETERINARSKO-MEDICINSKIH PROIZVODA I OSTALIH TVARI U PERADI I JAJIMA U EUROPSKOJ UNIJI U RAZDOBLJU 2009. - 2016. GODINE
163
Kao i kod peradi, analizirani sumnjivi uzorci jaja pokazali su u određenim slučajevima visok postotak nesukladnosti, pri-mjerice na kloramfenikol 2015. u Latviji (67%), doksiciklin 2014. u Poljskoj (86%) i/ili dioksine 2009. u Italiji (43%). Rezidue dioksina su utvrđene i u uzorcima jaja u Italiji uzorkovanih drugim monitoring programima tijekom raz-doblja 2010.-2013. godine, gdje se u prosjeku oko 9% takvih uzoraka pokazalo nesukladnim.
Monitoring peradi i jaja u Hrvatskoj Državni plan monitoringa rezidua (DPMR) u Republici Hrvatskoj prvi puta je izrađen 1998. godine, a od 2009. Go-dine provodi se prema zahtjevima za provedbu monitoringa sukladno onima kod zemalja članica EU (Bosnić, 2011.). Pristupanjem Hrvatske u EU 2013. godine rezultati DPMR-a su svake godine detaljno obrađeni i uključeni u opsežne iz-vještaje o prisutnosti rezidua VMP-a i ostalih kontaminanata u hrani životinjskog podrijetla od strane EFSA-e. U tablici 4. prikazani su podaci o monitoringu rezidua u peradi i jajima u Hrvatskoj od 2013. do 2016. godine, iz kojih je vidljivo kako su zadovoljeni propisani kriteriji prema Direktivi Vijeća 96/23/EZ (EFSA 2015.; 2016.;
2017.; 2018.). Iako nije dostupan podatak o količini pro-izvedenih jaja 2011. ili 2012. godine na temelju kojeg se izrađuje plan monitoringa za 2013. godinu, može se za-ključiti je kako je i tada analiziran dovoljan broj uzoraka na 1000 t godišnje proizvodnje jaja s obzirom na to da je uzorkovano više uzoraka nego u idućim godinama. Gledano po godinama uočava se kako je kod uzoraka peradi najveći broj nesukladnosti prijavljen 2014. i 2015. godine (slika 3.), ponajviše zbog prisutnosti mikotoksina zeara-lenona i njegovih derivata α-zearalenola i β-zearalenola (podskupina tvari B3d) u velikom postotku analiziranih uzoraka. Kod uzoraka jaja broj nesukladnih uzoraka kroz godine je u blagom porastu, pričem se najveći udio odnosi na kokcidiostatike. Uspoređujući stanje rezidua u Hrvatskoj i EU (slika 4.) uočava se veći postotak nesukladnih uzoraka u odnosu na ukupnu situaciju u EU, osobito kod peradi. Što se tiče sumnjivih uzoraka, 2014. godine detektiran je toltrazuril (B2b) u 40% uzoraka peradi, a 2015. godine diklazuril (B2b) u jedinom analiziranom uzorku jaja. Za uzorke iz uvoza i one prikupljene drugim načinima uzorko-vanja dostupno je vrlo malo podataka.
Tablica 4. Proizvodnja peradi i jaja te broj ciljanih uzoraka u Hrvatskoj u razdoblju 2013.-2016. Table 4. Production of poultry and eggs and number of targeted samples in Croatia during the 2013-2016 period
Godina Year
Perad / Poultry Jaja / Eggs Proizvodnja (t) Production (t)(a)
Broj uzoraka Number of samples
Analizirani uzorci / 200 t Samples tested / 200 t
Proizvodnja (t) Production (t)(a)
Broj uzoraka Number of samples
Analizirani uzorci / 1000 t Samples tested / 1000 t
2013. 7,1745 459 1.3 0 736 NA 2014. 70,463 549 1.6 32,173 721 22.4 2015. 56,505 365 1.3 33,305 382 11.5 2016. 57,075 402 1.4 34,320 571 16.6
(a) podaci o proizvodnji uzeti za izradu plana monitoringa rezidua odnose se na prethodnu godinu (a) Production data used for development of the Residue Control Plan refer to the previous year
Slika 3. Broj nesukladnih uzoraka peradi i jaja u Hrvatskoj prikazan po godinama za razdoblje 2013.-2016. i njihova
raspodjela prema podskupinama tvari kojima pripadaju rezidue detektirane u višim koncentracijama od dozvoljenih. Figure 3. Number of non-compliant samples of poultry and eggs in Croatia according to years for the 2013-2016 period and
their distribution according to substance subgroups to which residues detected in higher concentrations than permitted belonged.

MONITORING REZIDUA VETERINARSKO-MEDICINSKIH PROIZVODA I OSTALIH TVARI U PERADI I JAJIMA U EUROPSKOJ UNIJI U RAZDOBLJU 2009. - 2016. GODINE
164
Slika 4. Postotak nesukladnih uzoraka peradi i jaja prikupljenih kroz ciljano uzorkovanje u Hrvatskoj i Europskoj Uniji
prikazan po godinama za razdoblje 2013.-2016. odnosno 2009.-2016. Figure 4. Percentage of non-compliant samples of poultry and eggs collected through target sampling in Croatia and European
Union by years in the 2013-2016 and 2009-2016 periods. Rasprava i zaključci Planovi uzorkovanja, gdje su navedene sve specifikacije oko odabira uzoraka, nisu nužno jednaki svake godine i stoga analiza podataka na razini EU ne odražava u potpunosti pravu sliku oko stanja rezidua u svakoj pojedinoj zemlji članici. U ciljanim uzorcima peradi, u sklopu nacionalnih monitoring programa u EU, najviše analiza se provodi na zabranjene tvari (A6), antibakterijske tvari (B1) i kokcidio-statike (B2b). Na temelju objavljenih rezultata vidljivo je kako je naveći broj nesukladnih uzoraka peradi zabilježen 2009. godine, a otada je primijećen pad u njihovoj učesta-losti, prvenstveno za podskupinu kokcidiostatika. To je vrlo vjerojatno rezultat mjera koje su uslijedile uvođenjem Direktive Komisije 2009/8/EZ kojom su propisane najveće dopuštene količine neizbježnog onečišćenja hrane za živo-tinje kokcidiostaticima ili histomonostaticima za one vrste i/ili kategorije životinja za koje njihovo dodavanje u hrani nije namijenjeno. Na ciljanim uzorcima jaja kontrola se u najvećem postotku provodi na iste vrste tvari kao i kod peradi (A6, B1 i B2b), uz iznimku organoklornih spojeva podskupine B3a koji se zbog svoje pojavnosti u jajima tije-kom godina također smatraju skupinom spojeva na koju je potrebno povećavati broj pretraga. Nesukladni rezultati za uzorke jaja prijavljeni su u najvećem broju 2009. godine, od čega većinom zbog prisutnosti kokcidiostatika u količinama višim od dozvoljenih. Ulaskom Republike Hrvatske u EU 2013. rezultati DPMR-a su također obrađeni od strane EFSA-e i uključeni u godišnje izvještaje s rezultatima svih ostalih zemalja članica. Postotak nesukladnih uzoraka peradi i jaja u Hrvatskoj tijekom razdoblja 2013.-2016. godine je bio viši u odnosu na prosjek EU, kao što je prikazano na slici 4. To se prvenstveno odnosi na uzorke peradi 2014. i 2015. godine kada su de-tektirani zearalenon i njegovi derivati u čak 18 odnosno 14 uzoraka, što je dovelo do ukupne vrijednosti od 3,83% odnosno 5,21% nesukladnih uzoraka. S obzirom na to da su ti rezultati činili otprilike 3/4 svih nesukladnih rezultata za
2014. godinu, a 2015. čak i preko 85%, zaključuje se kako se radi o iznimnom slučaju te bi se postotak nesukladnih uzoraka peradi u Hrvatskoj, uz kontinuirano provođenje i daljnje unaprjeđenje DPMR-a, trebao kretati oko 0,5%-1,0% svih pretraženih uzoraka. Kod uzoraka jaja najveću prijetnju za zdravlje potrošača predstavljaju kokcidiostatici prisutnost kojih se u koncentracijama višim od NDK posljednjih godina bilježi u sve većem broju, što je donekle i očekivano uzimajući u obzir golemu upotrebu ovih lijekova u borbi protiv kokcidija. Broj nesukladnih uzoraka iz uzorkovanja sumnjivih uzoraka prijavljenih od strane zemalja članica EU ne predstavlja pravu sliku o stanju rezidua u tim zemljama. Uzeti su kao mjera opreza nakon prethodno uočene ne-sukladnosti u ciljanim uzorcima ili dokaza o posjedovanju i upotrebi zabranjenih tvari. Procedura uzorkovanja u slučaju sumnje se može razlikovati između zemalja članica EU. Primjerice, u klaonicama u Belgiji zajedno s uzorkom mi-šića mora se uzorkovati i svako mjesto na kojem se aplicira lijek u životinju i sve zajedno se prosljeđuje u laboratorij na multirezidualnu analizu. Ovakav pristup rezultira većom mogućnošću pojave nesukladnog rezultata za sumnjivi uzorak na više od jedne tvari. Relativno malen broj rezultata je objavljen za uzorke iz uvoza od strane zemalja članica, s obzirom na to da su te kontrole više vezane uz monitoring trećih zemalja nego monitoring rezidua u EU. Proizvodnja zdravstveno ispravne i prehrambeno vrijedne hrane od strateškog je interesa za sve zemlje članice EU. Po-java rezidua VMP-a i ostalih kemijskih kontaminanata u namirnicama životinjskog podrijetla postala je realnost, a s obzirom na činjenicu da je zbog toga ugrožen okoliš te zdravlje životinja i ljudi neophodno je praćenje stup-nja kontaminiranosti hrane kao što su perad i jaja. S tim ciljem su u ovom radu objedinjeni rezultati nacionalnih mo-nitoring programa provedenih na uzorcima peradi i jaja u EU tijekom 2009.-2016. godine. Također, raspravljeni su i rezultati analiza ciljanih uzoraka peradi i jaja u Hrvatskoj od 2013. do 2016. godine te uspoređeni s onima na razini cijele EU u istom razdoblju kako bi se dobila slika o stanju rezidua u prvim godinama članstva.

MONITORING REZIDUA VETERINARSKO-MEDICINSKIH PROIZVODA I OSTALIH TVARI U PERADI I JAJIMA U EUROPSKOJ UNIJI U RAZDOBLJU 2009. - 2016. GODINE
165
Literatura
BOSNIĆ, M. (2011): Sadržaj i ciljevi Državnog programa monitoringa rezidua [http://hvk.hr/hrv/skupovi/vdani11/ /prezentacije/pdf/bosnic-sadrzaj_i_ciljevi_drzavnog_progra-ma_monitoringa_rezidua.pdf, (26.10.2011.)]
EC (1996): Council Directive 96/23/EC of 29 April 1996 on measures to monitor certain substances and residues thereof in live animals and animal products and repealing Directives 85/358/EEC and 86/469/EEC and Decisions 89/187/EEC and 91/664/EEC. Off. J. Eur. Commun. L125, 10-32.
EC (1997): Commission Decision of 27 October 1997 fixing the levels and frequencies of sampling provided for by Council Directive 96/23/EC for the monitoring of certain substances and residues thereof in certain animal products. Off. J. Eur. Commun. L303, 12-15.
EC (2002): Commission Decision of 12 August 2002 implementing Council Directive 96/23/EC concerning the performance of analytical methods and the interpretation of results. Off. J. Eur. Commun. L221, 8-36.
EC (2006): Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Un. L364, 5-24.
EC (2010): Council Regulation (EU) No 37/2010 of 22 December 2009 on pharmacologically active substances and their classification regarding maximum residue limits in foodstuffs of animal origin. Off. J. Eur. Commun. L15, 1-72.
EFSA (2011): Report for 2009 on the results from the monitoring of veterinary medicinal product residues and other substances in live animals and animal products. Supporting Publications 158, 1-70.
EFSA (2012): Report for 2010 on the results from the monitoring of veterinary medicinal product residues and other substances in live animals and animal products. Supporting Publications EN-212, 1-65.
EFSA (2013): Report for 2011 on the results from the monitoring of veterinary medicinal product residues and other substances in live animals and animal products. Supporting Publications 158, 1-70.
EFSA (2014): Report for 2012 on the results from the monitoring of veterinary medicinal product residues and other substances in live animals and animal products. Supporting Publications EN-540, 1-65.
EFSA (2015): Report for 2013 on the results from the monitoring of veterinary medicinal product residues and other substances in live animals and animal products. Supporting Publications EN-723, 1-69
EFSA (2016): Report for 2014 on the results from the monitoring of veterinary medicinal product residues and other substances in live animals and animal products. Supporting Publications EN-923, 1-70.
EFSA (2017): Report for 2015 on the results from the monitoring of veterinary medicinal product residues and other substances in live animals and animal products. Supporting Publications 14(11):EN-1150, 1-69
EFSA (2018): Report for 2016 on the results from the monitoring of veterinary medicinal product residues and other substances in live animals and animal products. Supporting Publications EN-1358, 1-75.
SERRATOSA J., A. BLASS, B. RIGAU, B. MONGRELL, T. RIGAU, M. TORTADÈS, E. TOLOSA, C. AQUILAR, O. RIBÓ, J. BALAGUÉ (2006): Residues from veterinary medicinal products, growth promoters and performance enhancers in food-producing animals: a European Union perspective. Rev. sci. tech. Off. int. Epiz., 25 (2), 637-653.
VARENINA I., N. BILANDŽIĆ, Đ. BOŽIĆ LUBURIĆ, B. SOLOMUN KOLANOVIĆ, I. VARGA, B. KOS, R. BECK (2017): Zaostajanje kokcidiostatika u hrani kao posljedica prevencije i liječenja zaraze peradi Eimeria sp.Veterinarska stanica, 48, 451 - 463.
MONITORING OF VETERINARY MEDICINAL PRODUCT RESIDUES AND OTHER SUBSTANCES IN POULTRY AND EGGS IN THE EUROPEAN UNION DURING THE 2009-2016 PERIOD
Summary
The presence of unauthorized substances, residues of veterinary medicinal products or chemical contaminants in food may pose a risk factor for public health. The European Union (EU) legislative framework defines maximum limits permitted in food and monitoring programs for the control of the presence of these substances in the food chain. This report gives a comprehensive review of the results of monitoring of substances from groups A (substances having anabolic effect and unauthorized substances) and B (veterinary medicinal products and contaminants) present in poultry and egg samples in the EU during the 2009-2016 period. Data analysis presented in this paper was focused on targeted samples reported under Council Directive 96/23/EC. Samples collected through other sampling strategies (suspect, import or ‘other’) did not follow the designed monitoring plan; therefore, results on those samples were reported separately from the results on targeted samples. The largest number of analyzed samples of poultry and eggs was reported in 2014. Substances from B1 (antibacterial substances - antibiotics, sulfonamides, quinolones), A6 (unauthorized substances: nitrofurans, nitroimidazoles, etc.) and B2b (anticoccidials) subgroups were analyzed more frequently than others. The least number of analyzed poultry samples was in 2010, and of eggs in 2011. The largest number of non-complaint samples was recorded in 2009, mostly referring to anticoccidials. Since then, a decrease has been observed in the rate of non-compliant samples for anticoccidials (B2b), most likely as a result of the awareness and measures that followed implementation of the Commission Directive 2009/8/EC. Also, this paper presents up-to-date data on poultry and eggs obtained from annual residue monitoring program in animal-derived food in Croatia, the EU Member Country since 2013. Keywords: monitoring, residues, poultry, eggs, European Union, Croatia

PERADARSKI DANI 2019. 166
MIKROBIOLOŠKA KVALITETA HRANE ZA ŽIVOTINJE U REPUBLICI HRVATSKOJ
Marijana Sokolović, Marija Berendika, Borka Šimpraga, Fani Krstulović
Hrvatski veterinarski institut, Centar za peradarstvo, Zagreb, Hrvatska
Sažetak
Mikrobiološka ispravnost hrane za životinje kao i odgovarajuća hranidbena vrijednost neophodni su u uzgoju životinja i proizvodnji hrane animalnog podrijetla. Sirovine, a često i gotova hrana mogu biti izložene različitim okolišnim uvjetima. Navedeno olakšava kontaminaciju različitim mikroorganizmima koji ne samo da mogu štetiti životinjama, već mogu predstavljati i potencijalnu opasnost za ljude. Prema posljednjim izvješćima Europske agencije za sigurnost hrane i Europskog centra za kontrolu bolesti o trendovima i izvorima zoonoza, zoonotskih uzročnika i pojavi obolijevanja u ljudi uzrokovanih hranom u zemljama Europske Unije (EU), otrovanja hranom uzrokovana patogenim mikroorganizama još uvijek su najznačajnije zoonoze uzrokovane proizvodima animalnog podrijetla. U većini slučajeva otrovanja hranom u ljudi dokazana je i poveznica između kontaminirane hrane za životinje i neispravne hrane za ljude. Navedeno ukazuje na potrebu redovite kontrole kvalitete sirovina i hrane u svim fazama proizvodnje. U većini zemalja svijeta, pa tako i u zemljama EU, proizvodnja, prerada, transport, skladištenje i korištenje hrane regulirani su zakonom. Sirovine i hrana koja udovoljava navedenim kriterijima u pravilu neće predstavljati opasnost za zdravlje životinja, ljudi i okoliša. Cilj ovog rada je prikazati rezultate mikrobioloških analiza uzoraka sirovina i hrane za životinje tijekom dvogodišnjeg razdoblja (2017.-2018.) dostavljenih u Laboratorij za analitiku stočne hrane Centra za peradarstvo Hrvatskoga veterinarskog instituta u Zagrebu.
Ključne riječi: mikrobiološka kvaliteta, bakterije, salmonella, plijesni, hrana za životinje
Uvod
Procjena mikrobiološke ispravnosti hrane neophodna je u osiguranju kvalitetne hranidbe životinja. Cilj uzgoja živo-tinja je osigurati sve potrebne zdravstvene i tehnološke uvjete koji će omogućiti postizanje željenih proizvodnih rezultata i proizvodnju zdravstveno ispravnih proizvoda animalnog podrijetla. Kontinuirana pojava obolijevanja i trovanja ljudi hranom animalnog podrijetla ukazuje na to da je sigurnost hrane značajan problem (veterinarskog) javnog zdravstva u cijelome svijetu. Prema posljednjim izvješćima Europske agencije za sigurnost hrane (EFSA-e) i Europskog centra za kontrolu bolesti (ECDC) o trendovima i izvorima zoonoza, zoonotskih uzročnika i pojavi obolijevanja u ljudi uzrokovanih hranom u 2017. godini, kampilobakterioza, salmoneloza i bolesti uzrokovane patogenim sojevima bakterije Escherichia (E.) coli još uvijek su najznačajnije zoonoze uzrokovane proizvodima animalnog podrijetla (EFSA-ECDC, 2018.). Nedavna pojava bolesti u ljudi u Republici Hrvatskoj vezana uz nalaze salmonela i pesticida u hrani izazvala je burnu reakciju javnosti i potakla brojna pitanja o sigurnosti hrane na našem tržištu (HAH, 2018.).
Zbog navedenog, brojne zemlje zakonskim propisima (za-koni, uredbe, pravilnici, vodiči) nastoje regulirati čitav hranidbeni lanac uključujući proizvodnju žitarica i gotove hrane, uzgoj životinja te proizvode biljnog i animalnog pod-rijetla namijenjene za prehranu ljudi. Pritom najznačajnija je Uredba (EZ) br. 178/2002 Europskog parlamenta i Vijeća od 28. siječnja 2002. godine o utvrđivanju općih načela i uvjeta zakona o hrani, osnivanju Europske agencije za sigurnost hrane te utvrđivanju postupaka u područjima sigurnosti hrane. Ova Uredba propisuje da se na tržište smije stavljati samo ona hrana koja je "sigurna", odnosno da ista nije štetna za zdravlje ljudi ili neprikladna za prehranu ljudi. Prema navedenoj Uredbi, glavnu odgovornost za sigurnost hrane imaju subjekti u poslovanju s hranom u svim fazama proizvodnje, prerade, skladištenja i distribucije koje su pod njihovom kontrolom. U uzgoju životinja i proizvodnji hrane za životinje zakonski propisi EU također propisuju da sub-jekti u poslovanju s hranom koriste hranu koja je u skladu s propisanim kriterijima (183/2005/EZ). Nadalje, zakonski je regulirana uporaba dodataka hrani (1831/2003/EZ), genetski modificirana hrana (1829/2003/EZ; 2001/18/EZ), uporaba sirovina (2002/96/25/EZ), prisutnost neželjenih tvari u hrani
Dr. sc. Marijana Sokolović, dr.med.vet, Hrvatski veterinarski institut, Centar za peradarstvo, Laboratorij za analitiku stočne hrane, Henizelova 55, 10000 Zagreb, Hrvatska; Tel.: +385 (0)1 2440 214; Fax: +385 (0)1 2441 396; e-mail:[email protected]

MIKROBIOLOŠKA KVALITETA HRANE ZA ŽIVOTINJE U REPUBLICI HRVATSKOJ 167
za životinje (2002/32/EZ), hrana za posebne hranidbene namjene (93/74/EEZ) te određeni proizvodi koji se koriste u hranidbi životinja (82/471/EZ). Poštivanje navedenih zakonskih propisa i primjena pravila dobre proizvođačke prakse uvelike doprinose postizanju željenih ciljeva. Usprkos svim kontrolama i postupcima koje provode inspekcijska tijela ili subjekti u poslovanju s hranom, u praksi ipak dolazi do problema u uzgoju životinja u obliku slabijih proizvodnih rezultata, pojave simptoma bolesti, a ponekad i do uginuća životinja. U analizi uzroka često se tada otkrije kako hrana ili nije bila hranidbeno prikladna ili ista sadrži potencijalno patogene mikroorganizme ili mikroorganizme kvarenja koji mogu prouzročiti bolesti i životinja i ljudi. Također se često otkrije kako kontrola hranidbenih i mikrobioloških parametara nije bila provedena kako treba te se uklanjanjem neodgovarajuće hrane za životinje (ako je isto ekonomski i praktično izvedivo) dje-lomice ili u potpunosti može umanjiti nastala šteta. Stoga uz službene kontrole sigurnosti i kvalitete hrane, samokontrole (u okviru npr. HACCP, engl. Hazard Analysis and Critical Control Point, hrv. Analiza opasnosti i kritične kontrole točke) koje proizvode subjekti u poslovanju s hranom ključni su element osiguranja kvalitete i sigurnosti hrane (FAO, 2010.; Sokolovic i Ruk, 2011.). Pri proizvodnji hrane potrebno je osigurati odgovarajuće ekološke uvjete u tvornici i njezinom okolišu. Isto se postiže kontrolom abiotičkih (vlaga, zrak, oprema) i biotičkih (glo-davci, ptice, insekti, mikroorganizmi) čimbenika. Međutim, u procesu proizvodnje hrane za životinje sirovine su ipak primarni izvor potencijalnog onečišćenja gotove hrane (EFSA, 2008.). Stoga je prije uporabe sirovine nju potrebno analizirati, ali i definirati kriterije prema kojima se ona nabavlja. Budući da su sirovine redovito izložene različitim okolišnim uvjetima koji olakšavaju kontaminaciju različitim mikroorganizmima, a i zbog globalizacije tržišta i velikih tržišnih zahtjeva, u praksi je nabava takvih sirovina pravi izazov. Cilj ovog rada je prikazati mikrobiološku ispravnost uzoraka sirovina i hrane za životinje tijekom dvogodišnjeg razdoblja (2017.-2018.) analiziranih u Laboratoriju za analitiku stočne hrane Centra za peradarstvo Hrvatskoga veterinarskog insti-tuta u Zagrebu.
Materijal i metode
Za detekciju bakterija iz roda Salmonella korištena je stan-dardna akreditirana metoda (HRN EN ISO 6579-1:2017). Prisutnost bakterija roda Clostridium ispitivana je stan-dardnom metodom HRN EN ISO 7937:2005, bakterije roda Staphylococcus metodom HRN EN ISO 6888-1:2004, enterobakterije metodom HRN EN ISO 21528-1:2017, listerije metodom HRN EN ISO 11290-2:2017, dok je ukupan broj bakterija utvrđen metodom HRN EN ISO 4833:2013, a ukupan broj kvasaca i plijesni metodama HRN EN ISO 21527-1:2008 i HRN EN ISO 21527-2:2008. Sukladnost dobivenih rezultata ispitana je u skladu sa zahtjevima Pravilnika o sigurnosti hrane za životinje (NN 102/2016.). Navedeni Pravilnik određuje nepoželjne tvari u proizvodima namijenjenim za hranu za životinje i utvrđuje kriterije za pojedine mikroorganizme za hranu za životinje neživotinjskog podrijetla (tablica 1.). Pravilnik također pro-pisuje kategorizaciju sirovina i gotove hrane za životinje određivanjem razreda kvalitete (I.-IV.) na temelju utvrđi-vanja ukupnog broja indikatorskih mikroorganizama (sa-profitskih bakterija, kvasaca i plijesni) mikrobiološkom metodom Udruženja njemačkih agronomskih, analitičkih i istraživačkih instituta (njem. Verband Deutscher Landwirt-schaftlicher Untersuchungs und Forschungsanstalten, VDLUFA) (NN 102/2016 (tablica 2.). Ispitani uzorci dostavljeni su u okviru kontrola tvornica stočne hrane izravno od proizvođača životinja ili su bili analizirani kao dio službenih kontrola.
Rezultati i rasprava
Subjekti u poslovanju s hranom za životinje koji obavljaju poslove proizvodnje, prerade, skladištenja, prijevoza i distribucije hrane za životinje, uključujući uvoz hrane za životinje iz trećih zemalja i izvoz hrane za životinje u treće zemlje, obvezni su primjenjivati odredbe Pravilnika o sigurnosti hrane za životinje (NN 102/2016.). Uz ostale uvjete propisane odredbama navedenog Pravilnika, sirovine i hrana za životinje moraju udovoljavati mikrobiološkim kriterijima utvrđenim u Prilogu III. kako bi se mogla koristiti i/ili stavljati na tržište. Takvi se proizvodi smatraju "sigur-nima" i u skladu s člankom 15. Uredbe (EZ) br. 178/2002 te Uredbom (EZ) br. 882/2004.
Tablica 1. Mikrobiološki kriteriji za patogene bakterije (Prilog III., Odjeljak I., NN 102/2006.) Table 1. Microbiological criteria for pathogenic microorganisms (Prilog III., Odjeljak I., NN 102/2006)
Vrsta mikroorganizama / Microorganism Vrsta hrane za životinje / Feed type Broj mikroorganizama*/
Microorganism number*
Salmonella spp. Krmiva i krmne smjese za sve vrste životinja Raw materials and compoud feed for all animal species
0/25 g
Clostridium perfringens 100/ g
Listeria spp. Silaže i krmiva s visokim postotkom vlage Silage and raw materials with high moisture percentage 0/25 g
*cfu/g - broj koloniformnih jedinica u gramu / *cfu - colony forming unit per gram of feed

MIKROBIOLOŠKA KVALITETA HRANE ZA ŽIVOTINJE U REPUBLICI HRVATSKOJ 168
Tablica 2. Kategorizacija sirovina i hrane za životinje u razrede kvalitete prema mikrobiološkoj metodi VDLUFA (Prilog III., Odjeljak III., NN 102/2016.)
Table 2. Categorization of raw materials and animal feedstuffs in quality categories according to VDLUFA microbiological method (Prilog III., Odjeljak III., NN 102/2006)
Razred kvalitete / Quality category Kriterij / Criterion I. Poželjna kvaliteta / I Desirable quality
Broj IM O number
je manji od / is less than
OV II. Smanjena kvaliteta / II Lower quality do 5 puta veći od / Up to 5 times bigger than III. Loša kvaliteta / III Poor quality 5-10 puta veći od / 5-10 bigger than IV. Nije prihvatljivo za hranidbu životinja IV Not acceptable for animal feeding 10 puta veći od / 10 times bigger than
IM - indikatorski mikroorganizmi, OV - orijentacijske vrijednosti IM - indicator microorganisms, OV - orientation values Tijekom dvogodišnjeg razdoblja u Laboratoriju za analitiku stočne hrane analizirano je ukupno 999 uzoraka sirovina i gotove hrane za životinje (tablica 3.). Pritom su na prisutnost bakterija iz roda Salmonella ispitana 263 uzorka, na prisut-nost drugih bakterija ispitano je 600 uzoraka (ukupan broj bakterija, bakterije rodova Clostridium, Staphylococcus, E. coli i druge), dok je na prisutnost plijesni ispitano 136 uzoraka. U određenom broju uzoraka izdvojen je veći broj bakterija i plijesni, ali su ti uzorci s obzirom na propisanu kate-gorizaciju bili prihvatljivi za hranidbu životinja te time i u skladu sa zahtjevima Pravilnika (Prilog III, Odjeljak II. I Odjeljak III, NN 102/2016). S obzirom na vrijednosti sa-profitskih mikroorganizama i indikatora onečišćenja, većina analiziranih uzoraka pripadala je prvom i drugom razredu kvalitete, dok je svega 5% uzoraka bilo neprikladno za hranidbu životinja (IV. razred kvalitete). Navedeni Pravilnik kategorizira sirovine i krmne smjese prema utvrđenom broj bakterija, plijesni i kvasaca s obzirom na propisane ori-jentacijske vrijednosti indikatorskih mikroorganizama. Pritom su mezofilne aerobne bakterije žuto pigmentirane bakterije (bakterije karakteristične za proizvod, IM I), bacili, mikrokoki, koagulaza negativni stafilokoki (bakterije indika-tori onečišćenja, IM II) i stroptomicete (bakterije indikatori onečišćenja, IM III). Plijesni su podijeljenje u plijesni polja
(plijesni karakteristične za proizvod, IM IV), skladišne plijesni (indikatori onečišćenja, IM V) te plijesni iz roda Mucorales (indikatori onečišćenja, IM VI), a kvasci su kao indikatori onečišćenja navedeni u skupini IM VII. Budući da Pravilnik ne sadrži mikrobiološke kriterije za hranu za pojedine vrste životinja (npr. purane, patke, guske, koze, ovce i drugo), kao ni za pojedine dobne skupine životinja, hrana za te životinje ne može se kategorizirati u razrede kvalitete. Uz navedeno, plijesni kao indikatorski mikroorganizmi prema ovom Pravilniku podijeljene su u ka-tegorije IV. i V. kroz nejasnu podjelu na plijesni polja i skladišne plijesni. Kako bi se odredila sukladnost ovom Pravilniku potrebno je potražiti pojašnjenje navedenih indi-katorskih mikroorganizama u izvornim metodama opisanim u knjizi VDLUFA (VDLUFA, 2007a., 2007b., 2007c., 2007d.). Kategorizacija sirovina i hrane za životinje opisana u knjizi VDLUFA omogućuje precizniju procjenu higijen-skog statusa hrane za životinje. Prednost navedenih metoda je kategorizacija u skladu s mikrobiološkom kvalitetom, ali ovisno i o načinu proizvodnje gotove hrane za životinje. Po-znato je da određeni postupci obrade hrane mogu smanjiti broj potencijalno štetnih mikroorganizama, stoga metoda drugačije klasificira sipku i peletiranu hranu (Gafner, 2012.; Wolf i sur., 2014.).
Tablica 3. Broj mikrobioloških analiza uzoraka hrane za životinje u razdoblju 2017.-2018. godine Table 3. Number of microbiological tests of animal feed during the 2017-2018 period
Vrsta analize / Analysis Vrsta uzorka / Sample type
Ukupno / Total Hrana za životinje / Animal feed Sirovine / Raw materials Ostalo* / Other*
2017 2018 2017 2018 2017 2018 2017 2018 Bakteriološke* / Bacteriologic* 241 247 151 204 7 13 399 464 Mikološke analize* / Mycologic* 39 36 25 34 1 1 65 71 Ukupno / Total 280 283 176 238 8 14 464 535
*Bakteriološke analize: Salmonella spp., Clostridium spp., Listeria spp., Enterobakterije, E. coli O157; Mikološke analize: analize na patogene plijesni. Ostalo: premiksi / *Bacteriological analysis - detection of bacteria: Salmonella spp., Clostridium spp., Listeria spp., Enterobacteria, E. coli O157; Mycologic analysis: analysis for detection of pathogenic moulds. Other: premixtures.

MIKROBIOLOŠKA KVALITETA HRANE ZA ŽIVOTINJE U REPUBLICI HRVATSKOJ 169
Tablica 4. Učestalost bakterija iz roda Salmonella u analiziranim uzorcima sirovina i gotove hrane za životinje tijekom 2017. i 2018. godine
Table 4. Incidence of Salmonella in analyzed samples of raw materials and compound feed during the 2017-2018 period
Broj uzoraka / Number of samples Vrsta uzorka / Sample type
Ukupno / Total Hrana za životinje Animal feed
Sirovine Raw materials Ostalo* / Other*
2017 2018 2017 2018 2017 2018 2017 2018 Ukupno analiza / Total analysis number 110 62 50 34 3 4 163 100 Ukupno pozitivnih / Positive (total number) 0 7 2 5 0 0 2 12 Postotak pozitivnih* (%) / Positives* (%) 0 11,3 4,0 14,7 0 0 1,2 12,0
*Postotak pozitivnih uzoraka s obzirom na broj analiziranih uzoraka. Ostalo - premiksi *Percentage of positive samples in total number of analyzed samples.Other: premixtures Od patogenih mikroorganizama propisanih Pravilnikom dokazana je prisutnost bakterija iz roda Salmonella, zbog čega su ispitivani uzorci bili nesukladni zahtjevima Pra-vilnika za kriterij patogenih mikroorganizama (tablica 4.). Od izdvojenih bakterija roda Salmonella, serološkom tipi-zacijom i kemijskom karakterizacijom dokazane su vrste S. Senftenberg, S. Montevideo, S. Mbandaka, S. Tennessee, S. Enteritidis te jedan netipizirani soj. Od ostalih patogenih mikroorganizama ni u jednom ispitanom uzorku nije doka-zana prisutnost bakterija iz roda Listeria, kao ni bakterija Clostridium perfringens. Kao i prethodnih godina, bakterije iz roda Salmonella predstavljaju problem u uzgoju životinja i proizvodnji hrane animalnog podrijetla. Prisutnost ovih patogenih mikroorga-nizama može imati štetne posljedice ne samo za zdravlje životinja, nego i za zdravlje ljudi. U našem istraživanju salmonele su u 2017. godini utvrđene u svega 1,2% ispitanih uzoraka, dok je u 2018. godini, usprkos manjem broj ispi-tanih uzoraka, 12% bilo pozitivno. Prema izvješću EFSA-e i ECDC-a za 2017. godinu, pri-sutnost bakterija iz roda Salmonella ispitana je u 24 zemlje članice na ukupno 21868 uzoraka te je dokazana prisutnost salmonela u 1,32% ispitanih uzoraka (EFSA-ECDC, 2018.). Navedena relativno niska učestalost salmonela u hrani za životinje u skladu je s rezultatima dobivenim u našem Labo-ratoriju. Porast koji je zabilježen tijekom 2018. godine ne može se usporediti s rezultatima na razini EU budući da navedeno izvješće nije završeno. Potrebno je naglasiti kako monitoring sirovina i hrane za životinje nije ujednačen u svim zemljama članicama EU, čime je i teže procijeniti do-bivene rezultate. Također treba istaknuti kako analiza relativno malog broja uzoraka ne može biti pravi pokazatelj stvarne kontaminacije sirovina i hrane za životinje. Uspo-redbom podataka iz Sustava brzog uzbunjivanja za hranu i hranu za životinje (RASFF, engl. Rapid Alert System for Food and Feed) nije uočen porast u broju pozitivnih uzoraka hrane za životinje. Tijekom 2017. godine bilo je 114 oba-vijesti o neispravnim uzorcima sirovina i hrane za životinje, dok su tijekom 2018. godine zabilježene 94 obavijesti. Pritom većina obavijesti odnosila se je na nalaz salmonela, dok je u manjem broj uzoraka tijekom 2017. dokazana
prisutnost enterobakterija, a 2018. godine i bakterija iz roda Brucella. Zahtjev Pravilnika za kontrolu sirovina i hrane za životinje na odabrane patogene mikroorganizme (Clostridium perfrin-gens i Listeria spp.) ima cilj prevencije obolijevanja živo-tinja, a rezultat je epidemioloških istraživanja koja povezuju nastanak obolijevanja u ljudi s prisutnošću navedenih mikroorganizmima u hrani životinjskog podrijetla (Boelaert i sur., 2016.; Buchanan i sur., 2018.; EFSA-ECDC, 2018.). Učestalost bakterija iz roda Clostridium analizirana u skupnom izvješću EU (EFSA-ECDE, 2018.) ispitana je također u relativno malom broju uzoraka hrane za životinje u samo dvije zemlje članice EU (85 uzoraka podrijetlom iz Bugarske i Litve), pričem nije dokazan niti jedan pozitivan uzorak. Nedovoljna učestalost kontrole sirovina i gotove hrane ne utječe samo na potencijalni nastanak bolesti i sla-bijih proizvodnih rezultata u uzgoju životinja, nego poslije-dično može otežati i epidemiološka istraživanja nastanka bolesti uzrokovanih hranom u ljudi. Tako je u 2017. godini u zemljama EU usprkos sumnji na trovanje hranom u čak 33% prijavljenih obolijevanja nedostajao podatak o uzročniku trovanja. Prema dobivenim rezultatima analiza mikrobiološke isprav-nosti sirovina i gotove hrane za životinje u našem Labo-ratoriju može se zaključiti kako je prisutnost salmonela i drugih patogenih mikroorganizama redovit problem u pro-izvodnji hrane za životinje. Usprkos svih mjera higijene te primjene ostalih načela dobre proizvođačke prakse, kontrola sirovina i hrane u svim fazama proizvodnje nezaobilazan je zadatak svih subjekata u poslovanju s hranom. Takvo kon-tinuirano praćenje mikrobiološke ispravnosti sirovina i hrane pomaže u postizanju dobrih proizvodnih rezultata, a uz to omogućava i procjenu rizika temeljem koje će se izraditi plan uzorkovanja i kontrole kako na razini države, tako i samokontrole subjekata u poslovanju s hranom.
Zaključci
Zbog potencijalne prisutnosti salmonela, patogenih sojeva bakterije E. coli, plijesni te drugih mikroorganizama u hrani za životinje i proizvodima animalnog podrijetla mikro-

MIKROBIOLOŠKA KVALITETA HRANE ZA ŽIVOTINJE U REPUBLICI HRVATSKOJ 170
biološka ispravnost hrane važan je problem veterinarskog javnog zdravstva. Kako bi se proizvela hrana odgovarajuće kvalitete potrebno je primijeniti sve dostupne preventivne mjere te pravila dobre proizvođačke prakse uključujući i redovitu kontrolu sirovina i gotove hrane za životinje. Cilj proizvodnje hrane nije proizvodnja sterilne hrane, nego hrane koja ne sadrži potencijalno patogene mikroorganizme, dok ostale indikatorske mikroorganizme sadrži u "sigurnoj" količini. Integrirani pristup kontrole duž cijelog prehram-benog lanca preduvjet je za postizanje visoke razine sigurnosti sirovina, hrane i hrane za životinje u svim fazama proizvodnje. Iako zakonska regulativa propisuje mikrobio-loške kriterije za većinu sirovina i gotove hrane za životinje, zbog propusta u definiranju orijentacijskih vrijednosti in-dikatorskih mikroorganizama za pojedine vrste hrane nije moguća kategorizacija hrane za sve vrste životinja, kao ni za životinje različite dobi. Stoga bi se u izradi nove verzije Pravilnika koji regulira mikrobiološku ispravnost hrane za životinje trebale definirati orijentacijske vrijednosti indika-torskih mikroorganizma za hranu drugih kategorija životinja, ali i nedvosmisleno opisati podjelu patogenih plijesni.
Literatura
BOELAERT, F., G. AMORE, Y. VAN DER STEDE, M. HUGAS (2016): EU-wide monitoring of biological hazards along the food chain: achievements, challenges and EFSA vision for the future. Current Opinion in Food Science., 12, 52–62.
BUCHANAN, R. L., L. G. M. GORRIS, M. M. HAYMAN, T. C. JACKSON, R. C. WHITING (2018): A review of Listeria monocytogenes: An update on outbreaks, virulence, dose–response, ecology, and risk assessments Food Control, 88, 236–236.
Council Directive 82/471/EEC of 30 June 1982 concerning certain products used in animal nutrition.
Council Directive 93/74/EEC of 13 September 1993 on feedings-tuffs intended for particular nutritional purposes.
Direktiva 2001/18/EZ Europskog parlamenta i Vijeća od 12. ožujka 2001. o namjernom uvođenju u okoliš genetski modificiranih organizama i o stavljanju izvan snage Direktive Vijeća 90/220/EEZ.
Direktiva 2002/32/EZ Europskog parlamenta i Vijeća od 7. svibnja 2002. o nepoželjnim tvarima u hrani za životinje 2002/96/25/EZ
EFSA (2008): Microbiological risk assessment in feedingstuffs for food-producing animals. Scientific Opinionof the Panel on Biological Hazards. The Efsa Journal, 720, 1-84.
EFSA-ECDC (2018): The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA Journal, 16 (12), 5500.
FAO (2010): Manual of Good Practices for the Feed Industry. Poveznica za originalni dokument: http://www.fao.org/3/ /i1379e/i1379e00.htm [30.03.2019]
GAFNER, J. L. (2012): Microbiologische qualität von Futter-mitteln. Agrarforschung Sweiz 3. 252-257.
HAH - Hrvatska agencija za hranu (2018): pregled dostupnih rezultata mikrobiološke pretrage hrane koju uzorkuju subjekti u poslovanju hranom u okviru svojih planova samokontrole, stručno izvješće. Izvor: https://www.hah.hr [30.03.2019]
HRN EN ISO 11290-1:2017. Mikrobiologija u lancu hrane -- Horizontalna metoda za dokazivanje prisutnosti i određivanje broja Listeria monocytogenes i drugih Listeria spp. --2. dio: Metoda određivanja broja (ISO 11290-2:2017; EN ISO 11290-2:2017).
HRN EN ISO 21528-1:2017. Mikrobiologija u lancu hrane -- Horizontalna metoda za dokazivanje prisutnosti i određivanje broja Enterobacteriaceae -- 1. dio: Dokazivanje prisutnosti Enterobacteriaceae (ISO 21528-1:2017; EN ISO 21528-1:2017).
HRN EN ISO 4833-2:2013/Ispr.1:2014. Mikrobiologija lanca hrane -- Horizontalna metoda za određivanje broja mikroorganizama -- 2. dio: Određivanje broja kolonija pri 30 °C tehnikom nasađivanja na površinu podloge -- Tehnički ispravak 1 (ISO 4833-2:2013/Cor 1:2014; EN ISO 4833-2:2013/AC:2014).
HRN EN ISO 6579-1:2017. Mikrobiologija u lancu hrane -- Horizontalna metoda za dokazivanje prisutnosti, određivanje broja i serotipizaciju Salmonella -- 1. dio: Dokazivanje prisutnosti Salmonella spp. (ISO 6579-1:2017; EN ISO 6579-1:2017).
HRN EN ISO 6888-1:2004. Mikrobiologija hrane i stočne hrane -- Horizontalni postupak brojenja koagulaza-pozitivnih stafilokoka (Staphylococcus aureus i druge vrste) -- 1. dio: Postupak primjene Baird-Parkerove hranjive podloge na agaru (ISO 6888-1:1999+Amd 1:2003; EN ISO 6888-1:1999-+A1:2003).
HRN EN ISO 7937:2005. Mikrobiologija hrane i stočne hrane -- Horizontalna metoda za brojenje Clostridium perfringens -- Tehnika brojenja kolonija (ISO 7937:2004; EN ISO 7937:2004).
HRN ISO 21527-1:2012. Mikrobiologija hrane i hrane za životinje -- Horizontalna metoda za brojenje kvasaca i plijesni -- 1. dio: Tehnika brojenja kolonija u proizvodima s aktivitetom vode većim od 0,95 (ISO 21527-1:2008).
HRN ISO 21527-2:2012. MIkrobiologija hrane i hrane za životinje -- Horizontalna metoda za brojenje kvasaca i plijesni -- 2. dio: Tehnika brojenja kolonija u proizvodima s aktivitetom vode manjim ili jednakim 0,95 (ISO 21527-2:2008).
Pravilnik o sigurnosti hrane za životinje. Narodne Novine 102/2016.
SOKOLOVIĆ, M., I. RUK (2011): Sigurnost hrane za životinje i primjena HACCP-a u tvornicama hrane za životinje. Sto-čarstvo, 65 (3), 219-235.
Uredba (EZ) br. 178/2002 Europskog parlamenta i Vijeća od 28. siječnja 2002. godine o utvrđivanju općih načela i uvjeta zakona o hrani, osnivanju Europske agencije za sigurnost hrane te utvrđivanju postupaka u područjima sigurnosti hrane.
Uredba (EZ) br. 1829/2003 Europskog parlamenta i Vijeća od 22. rujna 2003. o genetski modificiranoj hrani i hrani za životinje Tekst značajan za EGP.
Uredba (EZ) br. 183/2005 Europskog parlamenta i Vijeća od 12. siječnja 2005. o utvrđivanju zahtjeva u pogledu higijene hrane za životinje Tekst značajan za EGP.
Uredba (EZ) br. 1831/2003 Europskog parlamenta i Vijeća od 22. rujna 2003. o dodacima hrani za životinje Tekst značajan za EGP.
Uredba (EZ) br. 882/2004 Europskog parlamenta i Vijeća od 29. travnja 2004. o službenim kontrolama koje se provode radi provjeravanja poštivanja propisa o hrani i hrani za životinje te propisa o zdravlju i dobrobiti životinja.

MIKROBIOLOŠKA KVALITETA HRANE ZA ŽIVOTINJE U REPUBLICI HRVATSKOJ 171
VDLUFA (2007a): Standard operating procedure for the enumeration of microorganisms using solid culture media. In VDLUFA Method Book III, Supl. 7 (28.1.1.). Darmstadt: VDLUFA.
VDLUFA (2007b): Standard operation procedure to enumerate bacteria, yeasts, moulds, and Dematiaceae. In VDLUFA Method Book III, Supl. 7 (28.1.2.). Darmstadt: VDLUFA.
VDLUFA (2007c): Standard operating procedure for identifying bacteria, yeasts, moulds, and Dematiacae as product-typical or
spoilage indicating microorganisms. In VDLUFA Method Book III, Supl. 7 (28.1.3.). Darmstadt: VDLUFA.
VDLUFA (2007d): Standard operating procedure for micro-biological quality assessment. In VDLUFA Method Book III, Supl. 7 (28.1.4.). Darmstadt: VDLUFA.
WOLF, P., U. SIESENOP, J. VERSPOHL, J. KAMPHUES (2014): Hygienic quality of feedstuffs for small mammals sent to the consultation service. Tierarztl Prax Ausg K Kleintiere Heimtiere. 42(2):101-106.
MICROBIOLOGICAL QUALITY OF ANIMAL FEED IN THE REPUBLIC OF CROATIA
Summary
Microbiological quality of feed along with appropriate nutritive value is extremely important in the production of animals and food of animal origin. Raw materials, as well as compound feed are exposed to different environmental conditions making contamination with different microorganism ease to acquire. Such contamination can have harmful consequences for animal health and be a potential risk for human health. According to the latest European Union (EU) summary report on the trend and sources of zoonoses, zoonotic agents and food-borne outbreaks, food poisoning caused by pathogenic microorganisms still represents the most important zoonosis caused by food of animal origin. In most cases of food poisoning, the link between contaminated feed and food has been found, indicating the importance of regular quality control of raw materials, food and feed at all production stages. In many countries worldwide, including EU Member States, a number of regulative measures exist and cover production, processing, transport, storage and use of food and feed. Raw materials, food and feed that comply with regulations should not represent a threat to animal and human health, or the environment. The aim of this work is to present the results of microbiological analysis of raw materials and animal feed analyzed during a two-year period (2017-2018) at the Feed Analysis Laboratory, Poultry Centre, Croatian Veterinary Institute in Zagreb. Key words: microbiological quality, bacteria, Salmonella, moulds, feed

PERADARSKI DANI 2019. 172
UTJECAJ DODATKA ETERIČNIH ULJA LIMUNA I KOMORAČA U HRANU NA TOVNA I KLAONIČKA SVOJSTVA TE SENZORSKA SVOJSTVA MESA TOVNIH PILIĆA
Brigita Hengl1, Gordana Kralik2, Slobodan Lilić3, Estella Prukner Radovčić4, Mislav Đidara2, Marcela Šperanda2
1 Hrvatska agencija za poljoprivredu i hranu, Osijek, Hrvatska 2 Sveučilište Josipa Jurja Strossmayera, Fakultet agrobiotehničkih znanosti, Osijek, Hrvatska 3 Institut za higijenu i tehnologiju mesa, Beograd, Srbija 4 Sveučilište u Zagrebu, Veterinarski fakultet, Zagreb, Hrvatska
Sažetak
Eterična ulja i njihove komponente uvelike se koriste u tovu pilića. Zbog njihovih antimikrobnih i antioksidativnih svojstava te djelovanja na poboljšanu probavljivost hrane može se očekivati njihov pozitivan utjecaj na zdravstveni status životinja, a time i bolje krajnje rezultate tova. Pokus je proveden na 96 pilića hibrida Ross u kojemu je pokusnoj skupini dodan u hranu pripravak AromaCorm® (Ireks Aroma, Hrvatska) u koncentraciji od 0,4%. U ovom istraživanju utvrđeno je da su pilići pokusne skupine imali značajno (p<0,05) veću tjelesnu masu 25. dana pokusa, dok kasnije te razlike nisu bile značajne. Konverzija hrane bila je podjednaka, što je rezultiralo većom masom trupa i podjednakim randmanom. Veća masa krila, batka sa zabatkom i leđa utvrđena je u skupini hranjenoj s dodatkom eteričnih ulja limuna i komorača, ali bez značajnih razlika. Sve praćene senzorske karakteristike mesa prsa i batka sa zabatkom bile su bolje u mesu pokusne skupine, a značajno (p<0,05) bolje bile su boja, tekstura, sočnost, mekoća i prihvatljivost mirisa u mesu prsa, a u mesu batka sa zabatkom tekstura, sočnost i mekoća. Ključne riječi: Eterična ulja, limun (Citrus limon), komorač (Foeniculum vulgare), tovni pilići Uvod Eteričnim uljima pripisuju se mnogobrojna svojstva među kojima valja istaknuti njihovo svojstvo antioksidativnog djelovanja i poboljšanje funkcija probave, poboljšanje okusa hrane, povećanja imunosti organizma i antimikrobna svojstva, zbog čega ih se sve više pokušava prikazati kao prirodne promotore rasta i uspješnu zamjenu za anti-mi-krobne promotore rasta (Hippenstiel i sur., 2011.; Karas-kova i sur., 2015.). Pozitivan učinak eteričnih ulja i njihovih komponenti na osobine rasta i zdravlje kod životinja posljedica su njihovih antimikrobnih svojstava i sposobnosti poticanja imunosti (Yang i sur., 2009.; Hippenstiel i sur., 2011.). Uz to, moguće je i kombinirati različita eterična ulja ili njihove komponente kako bi se postigao sinergistički učinak i na taj način još više poboljšala njihova djelotvornost.
Do 2006. godine u zemljama Europske Unije u krmne smjese dodavali su se antibiotici koji su imali ulogu pro-motora rasta, jer bi u posebno stresnim situacijama za piliće, kao što je promjena hrane, visoka temperatura okoliša, nepovoljni vremenski uvjeti utjecali na djelovanje patogenih mikroorganizama. Dodavanje antibiotika zabra-njeno je u zemljama EU, ali i u nekim drugim zemljama u svijetu, dok je u našoj zemlji zabrana stupila na snagu 2007. godine (Pravilnik o dodacima hrani za životinje, 2007.). Posljednjih godina zanimanje za fitogene kao za-mjenu za antibiotske promotore rasta sve se više povećava. Smatra se da je primarni način njihovog djelovanja anti-mikrobna aktivnost, dok je još i veća korist od toga što kroz kontrolu mogućih patogena utječu na ekosistem gastrointestinalne mikropopulacije. Na taj način životinje su manje izložene bakterijskim toksinima i drugim ne-željenim metabolitima koji potječu od mikroorganizama (npr. amonijak i biogeni amini), što omogućuje povećanu
Dr. sc. Brigita Hengl, Hrvatska agencija za poljoprivredu i hranu, Centar za sigurnost hrane, I. Gundulića 36b, Osijek, Hrvatska; Tel.: +385 (0)31 227 662; Email: [email protected]

UTJECAJ DODATKA ETERIČNIH ULJA LIMUNA I KOMORAČA U HRANU NA TOVNA I KLAONIČKA SVOJSTVA TE SENZORSKA SVOJSTVA MESA TOVNIH PILIĆA
173
crijevnu apsorpciju esencijalnih nutrijenata te bolji rast životinje unutar svog genetskog potencijala. Kao poslije-dica njihove antimikrobne aktivnosti javlja se i smanji-vanje imunog obrambenog stresa u kritičnim fazama proiz-vodnje životinja (Windisch i sur., 2007.). Postoje brojna iskustva u upotrebi eteričnih ulja u tovu brojlera. Komorač (Foeniculum vulgare) je biljka koja sadrži zna-čajan udio masnih kiselina, linolne i stearinske, te 16,81% trans anetola plus i 47,2% estragola. Takva kombinacija ukazuje na mogućnost utjecaja na povećanja tjelesne mase uz istodobno poboljšanje konverzije hrane (Mohammed i Abbas, 2009.). Komorač dodan u hranu za brojlere u omjeru od 0,05% nije dao statistički značajno povećanje u prirastu i konverziji hrane u odnosu na kontrolnu skupinu, iako je došlo do povećanja završne mase u tovu kao i bolje konverzije hrane (El-Deek i sur., 2003.). Isti autori navode kako je došlo do značajnog povećanja količine masti u mesu kod pilića hranjenih s dodatkom 0,1% komorača u odnosu na kontrolnu skupinu, što može biti posljedica veće količine masti u komoraču. Mohammed i Abbas (2009.) dobili su statistički značajno povećanje tjelesne mase brojlera hranjenih s dodatkom komorača od 1, 2 i 3 g/kg, ali ne i u potrošnji hrane. Saki i sur. (2014.) navode poboljšane karakteristike tova (dnevni unos hrane, dnevni prirast i konverzija hrane) kod skupine u čiju je hranu dodan komorač u odnosu na kontrolnu, što je u suprotnosti s navodima Saleha i sur. (2018.) koji su utvrdili da se u skupinama u kojima se u hrani nalazio dodatak sjemenki komorača u praškastom obliku značajno smanjio unos hrane u odnosu na kontrolu, a do kraja tova je došlo do značajne razlike u konverziji hrane koja je bila najmanja u pokusnoj pokusnoj grupi s najvećom količinom dodatka komorača. Tjelesna masa, prirast, unos i konverzija hrane za tovne piliće u čiju je hranu dodana citrusna pulpa u različitim postotcima (5% ili 10%) bili su značajno manji u odnosu na kontrolnu skupinu, a osobito uz dodatak od 10%. Moguće je kako pilići nisu mogli zadovoljiti potrebu za energijom potrebnom za normalan rast i pohranu masti. Ovakav energetski deficit u pilića hranjenih s dodatkom citrusne pulpe još se više pogoršao prilikom dodatka 10%, što je zasigurno posljedica primijećenog povećanja probavne cijevi. Već je otprije poznato kako je povećanje veličine probavne cijevi usko povezano s povećanjem energetskih potreba (Jorgensen i sur., 1996.). Slične rezultate postigli su Erhana i Bölükbaşıja (2017.) kod kojih se ukupan unos hrane značajno smanjio kod pokusnih skupina u čiju je hranu dodano eterično ulje limuna (0,1%, 0,2%, 0,3%), a s druge strane značajno se poboljšala konverzija hrane. Na ukupan prirast i masu dijelova trupa nije bilo utjecaja. Razlog tomu može biti taj što razine aktivnih sastojaka u eteričnom ulju limuna mogu varirati te se može dodavati u različitim koncentracijama u hranu, a na razinu utječu i različite sorte limuna. I u istraživanju Mouraoa i sur. (2008.) dodatak citrusne pulpe u hranu značajno je smanjio konverziju hrane, povećao unos hrane i smanjio prirast.
Glavne odlike kvalitete pilećeg mesa obuhvaćaju izgled, teksturu, sočnost, okus i funkcionalnost (Cross i sur., 1986.; Petracci i Baeza, 2011.), ali kod potrošača na prvom mjestu su izgled i tekstura, dok se sočnost i okus više pri-pisuju načinu priređivanja nego samom proizvodu (Fletcher, 2002.). Različiti rezultati koji se navode u istraživanjima otežavaju masovnije korištenje eteričnih ulja te se optimalne doze kao i kombinacije tek trebaju utvrditi (Hippemstiel i sur., 2011.). Cilj ovoga istraživanja bio je utvrditi utjecaj kombinacije eteričnih ulja limuna (Citrus limon) i komorača (Foenicu-lum vulgare) na proizvodne karakteristike tovnih pilića, na randman hrane te na senzorska svojstva pilećeg mesa.
Materijali i metode
Ukupno 96 pilića hibrida Ross 308 podijeljeno je u kon-trolnu (K) i pokusnu skupinu (P) te je svaka skupina imala 3 ponavljanja od po 16 pilića, ukupno 48 pilića po skupini s jednako zastupljena oba spola. Pilići su bili smješteni u čvrsto zidanom objektu s dvije potpuno odvojene etaže, betonskim podom, automatskom regulacijom ventilacije, temperature, vlage i osvjetljenja u objektu. Hranjenje i napajanje bilo je ad libitum. Pilići su hranjeni s tri kategorije hrane: starterom do 15. dana, finišerom 1 do 24. dana i finišerom 2 do kraja tova (tablica 1.). U sve tri kategorije hrane za skupinu P dodano je 0,04% kombinacije eteričnih ulja limuna i komorača u obliku proizvoda pod nazivom Aroma Korm® (Ireks Aroma, Hrvatska). Klanje pilića obavljeno je u odobrenom klaoničkom objektu na liniji klanja za piliće s električnim omamlji-vanjem. Deset sati prije klanja pilićima je uskraćena hrana. Automatsko rasijecanje na krila, prsa, leđa sa zdjelicom i batka sa zabatkom provedeno je prema Kralik i sur. (2008.). Osnovni dijelovi izvagani su pojedinačno za svaku skupinu. Senzorske osobine mesa određivale su se na mesu prsa i batka sa zabatkom. Metodologija ocjenjivanja pilećeg me-sa napravljena je u skladu s preporukama Svjetske udruge znanosti o peradi (World's Poultry Science Association) o standardiziranim metodama senzornih analiza brojlera (WPSA, 1987.). Za ocjenjivanje su korištene kvantita-tivno-deskriptivne ljestvice s mogućim rasponom vrijed-nosti od 1 (najlošija prihvatljivost) do najviše 8 (najbolja prihvatljivost). Rezultati istraživanja obrađeni su pomoću statističkog programa TIBCO Software Inc. (2018.), značajnosti raz-lika testirane su Studentovim t-testom. Rezultati senzor-skih analiza obrađeni su post hoc Tukey-Kramerovim HSD testom (Lowry, 2008.). Tijekom pokusa sa životinjama se postupalo u skladu sa Zakonom o zaštiti životinja (NN RH 102/2017.) i Pra-vilnikom o zaštiti životinja koje se koriste u pokusima ili u druge znanstvene svrhe (NN RH 55/2013.).

UTJECAJ DODATKA ETERIČNIH ULJA LIMUNA I KOMORAČA U HRANU NA TOVNA I KLAONIČKA SVOJSTVA TE SENZORSKA SVOJSTVA MESA TOVNIH PILIĆA
174
Tablica 1. Sastav i analiza startera, finišera 1 i finišera 2
STARTER FINIŠER 1 FINIŠER 2 SIROVINA % % % KUKURUZ, KUKURUZNI GLUTEN 47,05 47,06 54,00 PŠ. STOČNO BRAŠNO 2,00 3,00 11,00 PŠENICA 8,30 9,00 3,50 SOJA SAČMA, SOJINO ULJE 37,89 36,63 27,06 LIZIN 0,29 0,14 0,21 TREONIN 0,08 - 0,06 KOLIN KLORID 60% 0,09 0,06 0,05 SOL, VAPNENAC 2,30 2,07 2,05 RODIMET - 0,23 0,29 MCP-F 1,75 1,26 1,23 FYSAL - 0,30 0,30 PX BRO-S PIPO 0,25%* 0,25 - - PX BRO-F1 PIPO 0,25%* - 0,25 - PX BRO-F2 PIPO 0,25%* - - 0,25 UKUPNO 100 100 100 ANALIZA SIROVI PROTEINI 22,00 20,50 17,50 UKUPNA MAST 4,47 5,62 4,00 SIROVA VLAKNINA 3,15 3,09 2,87 AMINOKISELINE (bez lizina) 4,22 3,83 3,47 LIZIN 1,42 1,24 1,09 Ca 1,05 0,90 0,88 UKUPNI P 0,74 0,63 0,61 ISKORISTIVI P 0,41 0,33 0,33 Na 0,14 0,14 0,14 K 0,90 0,86 0,74 Cl 0,22 0,19 0,21 LINOLNA KISELINA 2,31 2,88 2,14 PEPEO 6,49 5,76 5,43 ME (Kcal/kg) 2.950 3.050 3.020
* originalna receptura Pipo Čakovec Hrvatska Table 1. Composition and analysis of starter, finisher 1 and finisher 2
STARTERR FINISHER 1 FINISHER 2 RAW MATERIAL % % % CORN, CORN GLUTEN 47.05 47.06 54.00 WEAT LIVESTOCK FLOUR 2.00 3.00 11.00 WEAT 8.30 9.00 3.50 SOYBEAN MEAL, SOYBEAN OIL 37.89 36.63 27.06 LYSINE 0.29 0.14 0.21

UTJECAJ DODATKA ETERIČNIH ULJA LIMUNA I KOMORAČA U HRANU NA TOVNA I KLAONIČKA SVOJSTVA TE SENZORSKA SVOJSTVA MESA TOVNIH PILIĆA
175
STARTERR FINISHER 1 FINISHER 2 THREONINE 0.08 - 0.06 CHOLINE CHLORIDE 60% 0.09 0.06 0.05 SALT, LIMESTONE 2.30 2.07 2.05 RHODIMET - 0.23 0.29 MCP-F 1.75 1.26 1.23 FYSAL - 0.30 0.30 PX BRO-S PIPO 0,25%* 0.25 - - PX BRO-F1 PIPO 0,25%* - 0.25 - PX BRO-F2 PIPO 0,25%* - - 0.25 TOTAL 100 100 100 ANALYSIS CRUDE PROTEINS 22.00 20.50 17.50 TOTAL FAT 4.47 5.62 4.00 CRUDE FIBER 3.15 3.09 2.87 AMINO ACIDS (one lysine) 4.22 3.83 3.47 LYSINE 1.42 1.24 1.09 Ca 1.05 0.90 0.88 TOTAL P 0.74 0.63 0.61 USABLE FORM P 0.41 0.33 0.33 Na 0.14 0.14 0.14 K 0.90 0.86 0.74 Cl 0.22 0.19 0.21 LINOLEIC ACID 2.31 2.88 2.14 ASH 6.49 5.76 5.43 ME, Kcal/kg 2.950 3.050 3.020
* original recipe by Pipo Čakovec Hrvatska
Rezultati
Rezultati istraživanja prikazani su u tablicama 2.-6.
Rasprava i zaključci
Tjelesna masa pilića 25. dana tova bila je značajno (p<0,05) veća u pokusnoj skupini u odnosu na kontrolnu. To je u skladu s rezultatima Alçiçeka i sur. (2003.) koji su u hranu dodali mješavinu eteričnih ulja koja je sadržavala ulje komorača (Foeniculum vulgare), mravinca (Origanum spp.) i citrusa (Citrus spp.), a koji potvrđuju značajno veću tjelesnu masu pilića do 21. dana tova. Do kraja tova veća tjelesna masa zadržana je u pokusnoj skupini, ali bez statističkih značajnosti (tablica 2.). Također i rezultati Alçiçeka i sur. (2003) koji su u hranu dodali mješavinu eteričnih ulja koja je sadržavala i ulje komorača (Foeniculum vulgare), mravinca (Origanum spp.) i citrusa (Citrus spp.) potvrđuju veću tjelesnu masu pilića 42. dana tova, ali ne značajno. Povećanje završne tjelesne mase pilića
u skupini u čiju je hranu dodan limun i komorač (tablica 2.) nije u skladu s rezultatima Mourao i sur. (2008.) kod kojih je dodatak citrusne pulpe u hranu značajno smanjio konverziju hrane, povećao unos hrane i smanjio prirast, a u skladu je s rezultatima El-Deek i sur. (2003.) kod kojih je komorač dodan u hranu za tovne piliće u omjeru od 0,05% doveo do povećanja završne mase u tovu, ali bez statističkih značajnosti, dok su Mohammed i Abbas (2009.) dobili statistički značajno povećanje tjelesne mase brojlera hranjenih s dodatkom komorača od 1, 2 i 3 g/kg. Moguće je kako utjecaj na povećanje tjelesne mase ima eterično ulje komorača, ali ne i citrusa te u njihovoj kombinaciji utjecaj citrusa prevladava. Konverzija hrane bila je lošija kod dodavanja hrani citrusne pulpe i komorača (Mourao i sur., 2008., El-Deek i sur., 2003.), dok se u našem istraživanju konverzija hrane (tablica 3.) neznatno razlikovala među skupinama (1,86-1,87), tako da se može zaključiti kako nije bilo niti pozitivnog niti negativnog utjecaja tretmana na konverziju hrane.

UTJECAJ DODATKA ETERIČNIH ULJA LIMUNA I KOMORAČA U HRANU NA TOVNA I KLAONIČKA SVOJSTVA TE SENZORSKA SVOJSTVA MESA TOVNIH PILIĆA
176
Tablica 2. Masa pilića 0., 25. i 42. dan tova i prirast
Masa pilića 0. dan (g) Masa pilića 25. dan (g) Masa pilića 42. dan (g) Prirast (g) Skupina x ± x ± x ± x ± K 44,31±1,34 1407,92a±135,55 2110,42±234,31 2066,10±234,14 P 43,98±1,51 1491,04b±132,61 2170,00±228,61 2126,02±228,62 p-vrijednost 0,255 0,003 0,210 0,207 K = kontrolna; P = pokusna a, b - različita slova označavaju značajne razlike (p < 0,05) x =aritmetička sredina; = standardna devijacija
Table 2. Broiler mass and growth on day 0, 25 and 42 of fattening period
Broiler mass day 0 (g) Broiler mass day 25 (g) Broiler mass day 42 (g) Growth (g)
Group x ± x ± x ± x ±
K 44.31±1.34 1407.92a±135.55 2110.42±234.31 2066.10±234.14 P 43.98±1.51 1491.04b±132.61 2170.00±228.61 2126.02±228.62 p- value 0.255 0.003 0.210 0.207
K = control; P = experimental a, b – different letters denote significant differences (p < 0.05) x =arithmetic mean; = standard deviation
Tablica 3. Utrošak hrane do 15., 25. i 42. dana tova, masa trupova i randman
Oznaka skupine K P
Utrošak hrane do 15. dana u kg (Starter) x 9,81 9,76
0,457 0,475
Utrošak hrane od 15. do 25. dana u kg (Finišer 1) x 15,00 15,00
0,000 0,000
Utrošak hrane od 25. do 42. dana u kg (Finišer 2) x 36,90 38,95
0,740 0,480
Ukupno utrošeno hrane u kg x 61,71 63,71
1,129 0,411
Razlika utrošene hrane u odnosu na kontrolnu skupinu u % +3,24
Konverzija hrane x 1,86 1,87
0,032 0,010
Masa trupa (g) x 1564,58 1623,23/*0,09
172,268 172,104
Randman (%) x 74 75/*0,155
0,017 0,032
K = kontrolna; P = pokusna *p-vrijednost x =aritmetička sredina; = standardna devijacija

UTJECAJ DODATKA ETERIČNIH ULJA LIMUNA I KOMORAČA U HRANU NA TOVNA I KLAONIČKA SVOJSTVA TE SENZORSKA SVOJSTVA MESA TOVNIH PILIĆA
177
Table 3. Feed conversion on day 15, 25 and 42, carcas mass and yield
Group K P Feed consumption on day 15 - kg (Starter) x 9.810 9.760 0.457 0.475 Feed consumption from day 15 to day 25 - kg (Finisher 1) x 15.000 15.000 0.000 0.000 Feed consumption from day 25 to day 42 - kg (Finisher 2) x 36.900 38.950 0.740 0.480 Total feed amount (kg) x 61.710 63.710 1.129 0.411 Difference in feed consumption versus control group (%) +3.24 Feed conversion ratio x 1.860 1.870 0.032 0.010 Carcas mass (g) x 1564.580 1623.23/*0.09 172.268 172.104 Yield (%) x 74 75/*0.155 0.017 0.032
K = control; P = experimental *p-value x =arithmetic mean; = standard deviation
Tablica 4. Masa batka sa zabatkom, prsa, krila i leđa
Skupina Batak sa zabatkom Prsa Krila Leđa
x ± x ± x ± x ±
K 266,00 ±25,60 626,50 ±51,43 94,41 ±10,78 146,32 ±15,32 P 271,00 ±19,05 623,00 ±51,31 98,75 ±9,66 152,00 ±12,44 p-vrijednost 0,325 0,830 0,072 0,075
K = kontrolna; P = pokusna x =aritmetička sredina; = standardna devijacija
Table 4. Mass of drumstick with thigh, breast, wings and back
Group Drumstick with thigh Breast Wings Back x ± x ± x ± x ± K 266.00 ±25.60 626.50 ±51.43 94.41 ±10.78 146.32 ±15.32 P 271.00 ±19.05 623.00 ±51.31 98.75 ±9.66 152.00 ±12.44 p- value 0.325 0.830 0.072 0.075
K = control; P = experimental x =arithmetic mean; = standard deviation
Različiti rezultati u literaturi navode se za utrošak hrane, koji je u našem istraživanju bio veći u pokusnoj skupini (tablica 3.) te je u suprotnosti s rezultatima dodatka citrusne pulpe u
hranu za tovne piliće koje su prikazali Mourao i sur. (2008.), te Mohammed i Abbas (2009.) opisujući smanjenje unosa hrane kod dodatka komorača u hranu.

UTJECAJ DODATKA ETERIČNIH ULJA LIMUNA I KOMORAČA U HRANU NA TOVNA I KLAONIČKA SVOJSTVA TE SENZORSKA SVOJSTVA MESA TOVNIH PILIĆA
178
Masa trupa pilića bila je lošija u kontrolnoj skupini, dok su dijelovi trupa, osim prsa, imali veću masu u pokusnoj skupini (p=0,09; tablica 4.). U skladu s tim su i rezultati Alçiçeka i sur. (2003.) koji su u hranu dodali mješavinu eteričnih ulja koja je sadržavala i ulje komorača (Foeniculum vulgare), mravinca (Origanum spp.) i citrusa (Citrus spp.), a koja je utjecala na veću masu trupova nakon evisceracije i hlađenja. Mourao i sur. (2008.) zaključili su kako dodatak citrusne pulpe u hranu značajno smanjuje konverziju hrane, povećava unos hrane i smanjuje prirast, jer pilići ne mogli zadovoljiti potrebu za energijom potrebnom za normalan rast i pohranu masti. Senzorske analize mesa prsa i mesa batka sa zabatkom napravljene su kako bi se moglo procijeniti ima li promjena u boji, teksturi, sočnosti, mekoći, prihvatljivosti mirisa i okusa u mesu, koje su nastale kao posljedica dodanih
eteričnih ulja u hranu za piliće (tablice 5. i 6.). U literaturi pronalazimo vrlo malo podataka o utjecaju eteričnih ulja ili njihovih komponenti dodavanih u hranu brojlera na senzorska svojstva mesa. Pretpostavlja se kako zbog njihovog brzog izlučivanja iz organizma njihov utjecaj na senzorsku kakvoću mesa peradi, osobito na sočnost, miris i okus, ne bi trebao imati neprihvatljive osobine za potrošača (Lee, 2002.). U našem pokusu zabilježili smo pozitivan utjecaj eteričnih ulja dodanih u hranu na senzorske karakteristike pilećeg mesa, prsa i batka sa zabatkom. Značajno bolja tekstura, sočnost i mekoća mesa prsa i mesa batka sa zabatkom bila je u skupini u čiju je hranu dodan limun i komorač u odnosu na kontrolnu skupinu. Dodatak eteričnih ulja utjecao je boju, prihvatljivost mirisa, značajno samo u mesu prsa. Bez značajnosti, bolja je bila i prihvatljivost okusa.
Tablica 5. Prikaz rezultata senzorskih osobina mesa prsa pilića
Skupina Boja Tekstura Sočnost Mekoća Prihvatljivost mirisa Prihvatljivost okusa K 4,50 3,50 5,58 5,50 5,66 6,33 P 3,50 4,66 6,33 6,50 6,58 6,41 p vrijednost <0,05 <0,05 <0,05 <0,05 <0,05 n.s. n.s. = nije značajno K = kontrolna; P = pokusna Table 5. Mean values of sensory characteristics of broiler breast meat
Group Color Texture Juiciness Softness Odor acceptability Taste acceptabilitiy K 4.50 3.50 5.58 5.50 5.66 6.33 P 3.50 4.66 6.33 6.50 6.58 6.41 p value <0.05 <0.05 <0.05 <0.05 <0.05 n.s.
n.s. = nonsignificant K = control; P = experimental Tablica 6. Prikaz senzorskih osobina mesa batka sa zabatkom
Skupina Boja Tekstura Sočnost Mekoća Prihvatljivost mirisa Prihvatljivost okusa K 3,67 3,58 5,50 5,17 6,25 6,42 P 3,67 4,67 6,33 6,42 6,67 6,50 p vrijednost n.s. <0,05 <0,05 <0,05 n.s. n.s.
n.s. = nije značajno K = kontrolna; P = pokusna Table 6 Mean values of sensory characteristics of broiler drumstick with thigh
Group Color Texture Juiciness Softness Odor acceptability Taste acceptabilitiy K 3.67 3.58 5.50 5.17 6.25 6.42 P 3.67 4.67 6.33 6.42 6.67 6.50 p value n.s. <0.05 <0.05 <0.05 n.s. n.s. n.s. = nonsignificant K = control; P = experimental

UTJECAJ DODATKA ETERIČNIH ULJA LIMUNA I KOMORAČA U HRANU NA TOVNA I KLAONIČKA SVOJSTVA TE SENZORSKA SVOJSTVA MESA TOVNIH PILIĆA
179
Bolje ocjene za sve promatrane pokazatelje senzornih kvaliteta mesa prsa i batka sa zabatkom značajan su rezultat koji svakako ide u prilog mnogim dugim osobinama ete-ričnih ulja i njihovih komponenti, čime bi njihovo korištenje kao dodatak hrani za piliće imalo još veće opravdanje. Može se zaključiti kako je hrana s dodatkom eteričnih ulja limuna i komorača doprinijela rastu pilića do 25. dana tova. Također pozitivno, iako bez statističke značajnosti, utjecala je na veličinu krila, batka sa zabatkom i leđa pilića, a zna-čajno je utjecala na poboljšanje praćenih senzornih karak-teristika mesa.
Literatura
ALÇIÇEK, A., M. BOZKURT, M. ÇABUK (2003): The effect of an essential oil combination derived from selected herbs growing wild in Turkey on broiler performance. S. Afr. J. Anim. Sci., 33, 89-94.
CROSS, H. R., P. R. DURLAND, S. C. SEIDEMAN (1986): Sensory qualities of meat. In: Muscle As Food. (P.J. Bechtel, Ed.), Academic Press, New York, NY, 279-320.
EL-DEEK, A. A., Y. A. ATTIA, M. M. HANNFY (2003): Effect of anise (Pimpinella anisium), ginger (Zingiber officinale roscoe) and Fennel (Foeniculum vulgare) and their mixture of performance of broilers. Arch. Geflugelkd., 67, 92-96.
ERHAN, M. K. S. C. BÖLÜKBAŞI (2017): Citrus peel oils supplementation in broiler diet: effects on performance, jejunum microflora and jejunum morphology. Braz. J. Poultry Sci. Special Issue Nutrition., Jan-Mar, 15-22.
FLETCHER, D.L. (2002): Poultry meat quality. Worlds Poult. Sci. J., 58, 131-145.
HIPPENSTIEL, F., A. A. A. ABDEL-WARETH, S. KEHRAUS, K. H. SÜDEKUM (2011): Effects of selected herbs and essential oils, and their active components on feed intake and performance of broilers – a review. Arch. Geflugelkd., 75, 226-234.
JORGENSEN, H., X. ZHAO, B. EGGUM (1996): The influence of dietary fiber source and level on the development of the gastrointestinal tract, digestibility, degree of fermentation in the hind-gut and energy metabolism in pigs. Br. J. Nutr., 75, 365–378.
KARÁSKOVÁ, K., P. SUCHY, E. STRAKOVA (2015): Curent use of Phytogenic feed additives in animal nurtition: a review. Czech. J. Anim. Sci., 60, 521-530.
KRALIK, G., E. HAS-SCHÖN, D. KRALIK, M. ŠPERANDA (2008): Peradarstvo, biološki i zootehnički principi. Poljo-privredni fakultet Sveučilišta J. J.Strossmayer u Osijeku, Osijek, Hrvatska.
LEE, K. W. (2002): Essential oils in broiler nutrition, Utreht, The Netherlands.
LOWRY, R. (2008): One Way ANOVA–Independent Samples. Vassar.edu. Retrieved on December 4th, 2008. http://vassarstats.net/textbook/ch14pt2.html. Zadnja posjeta stranici 11.09.2012.
MOHAMMED, A. A., R. J. ABBAS (2009): The effect of using fennel seeds (Foeniculum vulgare L.) on productive perfor-mance of broiler chickens. Int. J. Poult. Sci., 8, 642-644.
MOURAO, J. L., V. M. PINHEIRO, J. A. M. PRATES, R. J. B. BESSA, L. M. A. FERREIRA, C. M. G. A. FONTES, P. I. P. PONTE (2008): Effect of dietary dehydrated pasture and citrus pulp on the performance and meat quality of broiler chickens. Poult. Sci., 87, 733-743.
PETRACCI, M., E. BAEZA (2011): Harmonization of methodologies for the assessment of poultry meat quality features. Worlds Poult. Sci. J., 67, 137-151.
Pravilnik o dodacima hrani za životinje (2007). Narodne novine 9/07.
SAKI, A. A., M. KALNTAR, E. RAHMETNEJAD, F. MIRZAAGHATABAR (2014): Health Characteristics and preformance of broiler chicks in response to tigonella foenum gracum and Feoniculum vulgare. Iran J. Appl. Anim. Sci., 4, 387-391.
SALEH, L., P. S. RAM, A. KAMIN, W. R. PRAMOD, NEERAJ (2018): Efficacy of Foeniculum vulgare Seeds Powder on Feed Intake and Feed Conversation Ratio in Broilers. International Journal of Excellence Innovation and Development., 1, 14-17.
TIBCO Software Inc. (2018). Statistica (data analysis software system), version 13. http://tibco.com.
WINDISCH, W., K. SCHEDLE, C. PLITZNER, A. KROISMAYR (2007): Use of phytogenic products as feed additives for swine and poultry. J. Anim. Sci., 86, 140-148.
WPSA (1987) Working Group No. 5: Recommendation for a standardized method of sensory analysis for broilers. Worlds Poult. Sci. J., 43, 64-68.
YANG, Y., P. A. IJI, M. CHOCT (2009): Dietary modulation of gut microflora in broiler chickens: a review of the role of six kinds of alternatives to in-feed antibiotics. Worlds Poult. Sci. J., 65, 97-114.
ADDITION OF LEMON AND FENNEL ESSENTIAL OIL TO FEED INFLUENCES BROILER PERFORMANCE, CARCASS COMPOSITION AND SENSORY MEAT QUALITY
Summary
Essential oils and their components have great possibilities for use in broiler fattening. Because of their antimicrobial and antioxidant properties, and their effects on improved digestibility of feed, their positive effect on animal health status can be expected, and thus better final fattening results. The study included 96 Ross hybrid broilers divided into two groups: experimental group was fed with dietary addition of AromaCorm® (Ireks Aroma, Croatia) in a dose of 0.4%. Study results revealed the experimental group chickens to have a significantly (p<0.05) higher body mass on day 25 of the experiment, whereas later these differences were not significant. Increased body mass and growth were observed in the experimental group,

UTJECAJ DODATKA ETERIČNIH ULJA LIMUNA I KOMORAČA U HRANU NA TOVNA I KLAONIČKA SVOJSTVA TE SENZORSKA SVOJSTVA MESA TOVNIH PILIĆA
180
while the feed conversion ratio was similar, resulting in a larger body mass and an equal dressing percentage. The larger mass of wings, drumstick with thigh and back was found in the group fed with the addition of essential oils of lemon and fennel, but without statistical significance. In the experimental group, all sensory characteristics of the breast meat and drumstick with thigh meat were better, while (p<0.05) the color, texture, juiciness, softness and acceptability of odor in the breast meat, and juiciness, softness and texture in drumstick with thigh meat were significanly better. Key words: Essential oils, citrus (Citrus limon), fennel (Foeniculum vulgare), broilers





























