Embed Size (px)
Citation preview

Proc. Indian Acad. Sci. (Plant Sci.), Vol. 96, No. 6, December 1986, pp. 475-486. �9 Printed in India.
Ontogeny of palmately compound leaves in angiosperms: 3. Ar… Spp.
K P E R I A S A M Y and E A M U R U G A N A T H A N * Department of Botany, Bharathidasan University, Tiruchirapalli 620 023, India
* Present Address: Department of Botany, G'overnment Arts College, Ootacamund 648 001, India
MS received 8 July 1986; revised 90ctober 1986
Abstract. Arisaema is mono or bifoliar, exhibiting sympodial growth with annual conversion of the shoot apex into the floral apex. The shoot apex shows pendulum symmetry during successive plastoehrons.
Leaf initiation is hypodermal. The primordium quickly acquires the configuration of a 5 layered laminar plate meristem and its base extends around the shoot apex as the sheath. Following this, apical growth ceases in the primordium. Its apex beeomes a hood like lamina wing and the sheath develops a median adaxial meristem which leads to its thickening. The lamina wing expends and becomes plicately folded at right angles to its surface. Each fold becomes a leaflet by the elongation of the abaxial edges and suppression of the adaxial edges of the folds. Early abortion of the lamina wing results in a scale leal which comprises the sheath portion alone.
An adult leaf has 300-400 veins which run independently from the corro to the leaflets through the petiole. The dorsal median strand develops first and goes to the midrib of the median leaflet. Subsequent strands arise laterally on either side of this in ~ se¡ of tangential rows. Those of the tirst row bifurcate at the tip of the petiole to enter two neighbouring leaflets. The strands run parallel in the leaflet midrib and diverge at different levels into the lamina as lateral veins.
Keywords. Arisaema; palmately compound leaf; leaf ontogeny; monocot leal'; leaf venation.
1. Introduction
Leaf on togeny in Tabebuia pentaphylla (dicot) and Dioscorea pentaphylla (monocot) has been dealt with in the first two papers of this series (Periasamy and M u r u g a n a t h a n 1985a, b). Araceae is a monoco ty ledonous family in which the leaf venat ion is reticulate and also certain members have c o m p o u n d lamina. Since bo th
these features are rare in monocotyledons , a s tudy of c o m p o u n d leaf on togeny in this family was under t aken and this paper deals with Arisaema.
2. Materials and methods
Arisaema tortuosum Schoff. was collected from Yercaud hills and A. leschenaultii B1.
from Kolli hills and O o t a c a m u n d of Tamil N a d u State. The shoot apices and leaves at various stages of development were fixed in FAA in the field itself. Most of the scale leaves were carefully removed before fixing. Cus tomary alcohol-xylol series dehydra t ion and paraffŸ embedding were done. Serial micro tome sections at 8-10q were stained with tannic acid-ferric chloride, Heidenhains i ron a lum
haematoxyl in and erythrosin following the procedure of e l iminat ing the e thanol
475

476 K Periasamy and E A Muruganathan
series (Periasamy 1967). Clearing was done with 5-10% NaOH solution and also a mixture of chloral hydrate, lactic acid, phenol, clove oil and xylene in the proportion of 2 : 2 : 2 : 2 : 1 by weight (Herr 1971). SEM of FAA fixed material was made at the Department of Botany, University of Massachusetts, USA, by Otto L-Stein.
3. Observations
3.1 Leaf morphology
Arisaema, a genus of the Araceae is widely distributed in Kodaikanal, Yercaud, Ootacamund and Kolli hills of Tamil Nadu and is commonly referred to as Cobra flower. The compound leaves of two species of this genus A. tortuosum and A. lesche- naultii differ in their leaflet arrangement in addition to floral characters.
A. leschenaultii possesses only one mature foliage leaf at a time. The leaflets may vary from 3-18 and radiate from the tip of the rachis (figure 1).
A. tortuosum possesses two mature foliage leaves at a time. The leaves are compound with 3-18 leafiets. All the leaflets do not radiate directly from the rachis tip as in A. leschenaultii but form 2 groups: (i) a single median leaflet which is directly attached te the rachis, (ii) two lateral groups of 7-9 leaflets each attached to short secondary rachises that arise from the primary rachis (figure 2).
Even though A. ieschenaultii bears one mature foliage leaf and A. tortuosum two leaves at a time, a seedling or vegetative bud produces many leaves one after the other before it flowers for the first time. Thereafter every year only two ~oliage leaves are produced foUowed by the inflorescence. In both the species, the first leaf of the seedling is simple but the first leaf that grows from adventious buds is trifoliate. The successive leaves produce increased number of leaflets.
3.2 Apical organisation
The shoot apex is convex in longitudinal section (figure 3). During the maximal phase, the apex measures 180-200/~m in diameter and 80-100 #m in height. A minimal apex measures 140-160 #m in diameter and 30-40 #m in height.
The shoot apex exhibits two anticlinally dividing surface layers (figure 3). Pe¡ dJvisions occur in the hypodermal layer at the time of leaf initiation.
Cytohistologically, the apex exhibits 3 zones, central zone, peripheral zone and ¡ meristem zone (figure 3). The central zone is characte¡ by large vacuolated ceUs in which cell division is less frequent. In the pe¡ zone, located at the flanks of the apex, the cells are smaller with dense cytoplasm and they divide more frequently in all planes. Predominantly transverse divisions at the base of the central zone results in the formation of a rib meristem.
Leaf primordia arise alternately from the opposite sides of the apical meristem and the divergence angle is nearly 180 o. Leaf initiation occurs very close to the summit of the maximal shoot apex and uses up its major po~tion leading to the minimal phase (figure 6). During the growth of the apex to the maximal phase, it enlarges towards the side opposite to that of the youngest primordium. Thus during successive plastochrons the apex becomes alternately shifted by 180 ~ and exhibits a pendulum symmetry.

L e a f ontogeny in Arisaema Spp. 477
i �91
Figures 1~. 1. A. leschenaultii. The leaflets of compound lamina radiating from the rachis tip ( x 'A). 2. A. tortuosum. Leaflets arranged in 3 groups ( x V4). 3-6. A. leschenaultii. 3. LS of shoot apex (• 285). 4. TLS of leaf primordium tip through the lamina wing which is seen as two strips at the apex due to its adaxial folding; note 5 layered nature at the base of the strips ( x 190). 5. TS of the sheath portion of leaf primordium showing adaxial meristem ( x 285). 6. LS of apical bud showing the initiation of a leaf primordium and the adaxial growth of the base of the next older primordium ( x 135). (A, Adaxial meristem; LI, L2, youngest and next older leaf primordia respectively; LW, lamina wing; S, shoot apex).

478 K Periasamy and E A Muruganathan
Figures 7-10. A. leschenaultii. 7. TS of young leaflet lamina, note periclinal division of submarginal initial ( x 430). 8. TS of aborting lamina of scale leal" ( x 210). 9. TLS of leaf p¡ showing the upright terminal leaflet and drooping lateral leallets (x 60). 10. Cleared tip of leaf primordium showing the dorsal median strand and two lateral bundles on either side; note bifurcation of lateral bundles before entering the leaflet ( • 50). (DL, Degenerating lamina wing; DM, dorsal median strand; LL, lateral leaflet; LS, lateral strand; TL, terminal leatlet).
3.3 Leaf initiation
Leaf initiation commences with the periclinal division of a few adjacently placed hypodermal cells. This is soon followed by random divisions of subjacent corpus cells to f o r m a crescent shaped foliar primordium which is characteristically 5 layers thick (figure 6). The initial growth of the primordium is apical and the 5 layered nature continues to be maintained at its apex (figure 6). As its apex grows verticaUy, the base of the primordium spreads around the flanks of the shoot apex (figure 11) to form the sheath, delimiting a lower sheath region and an upper lamina-petiole region. When the primordium reaches a height of 50 #ra, its apical growth slows down. Further vertical growth is predominantly intercalary.
Simultaneously with the early apical growth, conspicuous adaxial growth commences in the periole region above the sheath. The adaxial meristem is characte¡ by repeated periclinal divisions of the hypodermal layers and radial orientation of the derivatives along the adaxial face of the primordium (figure 5).
Leaf ontogeny in Arisaema Spp. 479
3.4 Leaflet formation
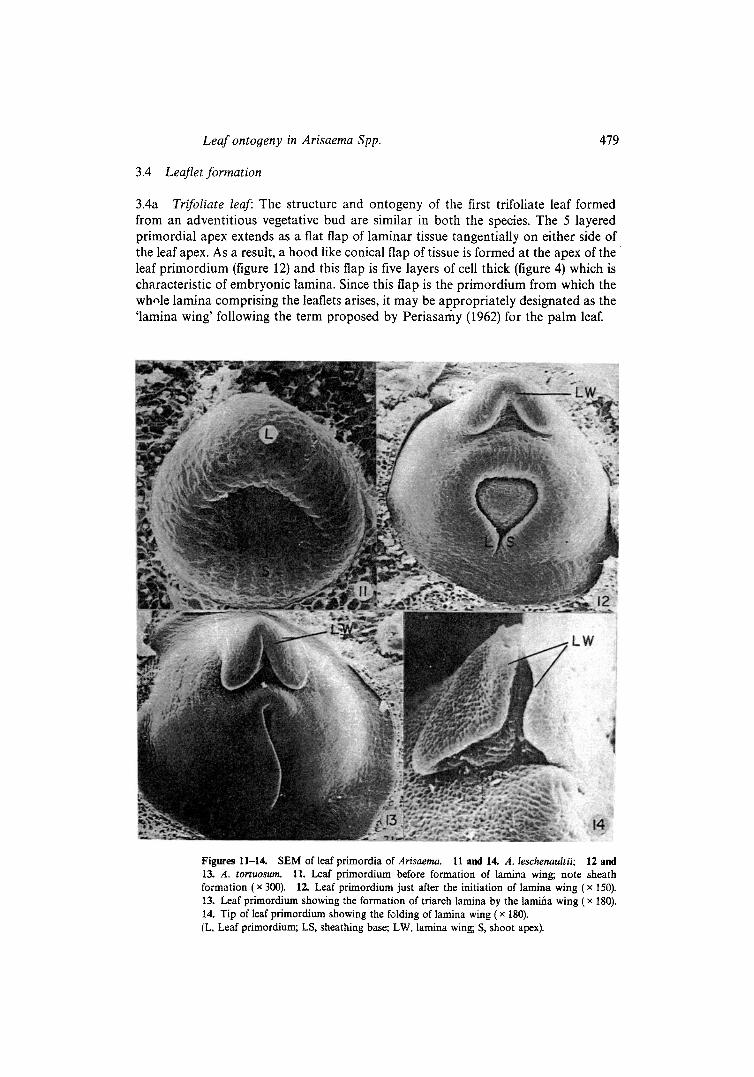
3.4a Trifoliate leaf: The structure and ontogeny of the first trifoliate leaf formed from an adventitious vegetative bud are similar in both the species. The 5 layered primordial apex extends a s a flat flap of laminar tissue tangentially on either side of the leaf apex. A s a result, a hood like conical fiap of tissue is formed at the apex of the leaf pr imordium (figure 12) and this flap is five layers of cell thick (figure 4) which is characteristic of embryonic lamina. Since this flap is the pr imordium from which the wh,qe lamina comprising the leaflets arises, it may be appropriately designated as the ' lamina wing' following the term proposed by Periasar¡ (1962) for the palm leaf.
Figures 11-14. SEM of leaf primordia of Arisaema. 11 and 14. A. leschenaultii; 12 and 13. A. tortuosum. 11. Leaf primordium before formation of lamina wing; note sheath formation ( x 300). 12. Leal primordium just after the initiation of lamina wing ( x 150). 13. Leaf primordium showing the formation of t¡ lamina by the lami¡ wing (x 180). 14. Tip of leaf primordium showing the f~lding of lamina wing ( x 180). (L, Leaf primordium; LS, sheathing base; LW, lamina wing; S, shoot apr
480 K Periasamy and E A Muru#anathan
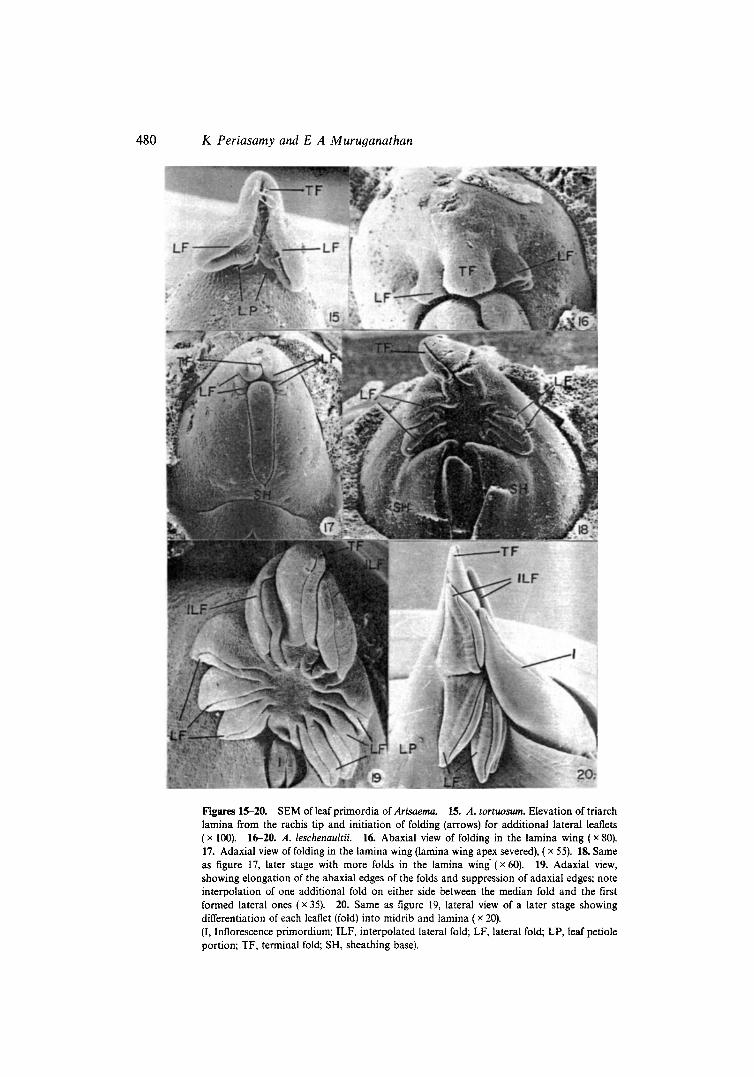
Figures 15-20. SEM of leal primordia of Arisaema. 15. A. tortuosum. Etevation of triarch lamina from the rachis tip and initiation of folding (arrows) for additional lateral leaflets ( x 100). 16-20. A. leschenaultii. 16. Abaxial view of folding in the lamina wing ( x 80). 17. Adaxial view of folding in the lamina wing (lamina wing apex severed), ( x 55). 18. Same as figure 17, later stage with more folds in the lamina wing-(x 60). 19. Adaxial view, showing elongation of the abaxial edges of the folds and suppression of adaxial edges; note interpolation of one additional fold on either side between the median fold and the first formed lateral ones ( x 35). 20. Same as figure 19, lateral view of a later stage showing differentiation of each leaflet (fold) into midrib and lamina ( x 20). (I, Inflorescence primordium; ILF, interpolated lateral fold; LF, lateral fold; LP, leaf petiole portion; TF, terminal fold; SH, sheathing base).

Leaf ontogeny in Arisaema Spp. 481
The apical part of the lamina wing grows vertically and folds adaxially (figures 13, 14, 15). The two sides expand downwards forming a triangular flat laminar flap on either side (figures 14, 15). The free margins of the flaps fold inward (adaxially) so that the lamina at this stage is folded in a triarch manner with one terminal and two lateral folds (figures 14, 15). All the folds are formed mechanically and no differential growth is involved. The terminal fold ultimately develops into the median and the lateral folds into the two lateral leaflets. Of the 3 folds, the terminal one is directed upwards, while the two laterals are directed downwards (figure 15).
3.4b Multifoliate leafl In A. tortuosum, after the lamina wing forms the triarch fold, it is elevated from the sheath by intercalary growth at the tip of the petiole portion (figure 15). Additional folds are then initiated at the morphological base of the two lateral folds (figure 16) such that the additional folds have no direct connection with the rachis (figure 15).
In A. leschenaultii, after the formation of the first 3 folds, the lamina is not elevated from the sheath. Additional downwardly directed foldings are formed basipetally in continuation with the lateral folds of the triarch lamina (figures 16-18). The additional folds are directly attached to the tip of the petiole region of the leaf primordium (figures 16-18). Sometimes no folds are i~itiated between the median vertical fold and the lateral d.ooping .folds of the primary triarch lamina (figures 9, 18). But often, an extra vertical fold becomes interpolated on either side between the median and first lateral folds (figures 19, 20) before additional folds are formed. In any case, till as many number of folds as there are leaflets in an adult'leaf are formed, the lamina remains essentially simple in both the species.
Further development in both the species is similar. The foldings deepen by expansion brought about by the activity of the plate meristem of each fold. However, during this process, only the free outer (abaxial) edges of the folds elongate at right angles to the deepening while the inner (adaxial) edges fail to elongate and this leads to the separation of each fold asa leaflet (figures 18-20). The elongated outer edges of the folds differentiate as the leaflet midribs.
3.5 Lamina development
The widening of the leaflet lamina takes place initially by the activity of a marginal meristem which represents the continuation of the meristematic activity of the lamina wing (figure 7). In the cells of the protoderm at the margins of the folds, definite marginal initials and derivatives are not distinguishable and they all undergo anticlinal divisions to add to the protoderm. The submarginal initials by their oblique anticlinal divisions form the abaxiaI and adaxial layers. Usually the abaxial layer divides periclinally to form the middle layer. However, the middle layer may also be directly formed by the periclinal division of the submarginal initial (fgure 7).
The activity of the 5 layered plate meristem increases the length and breadth of the lamina (figure 9). At the beginning, the lamina vernation is convolute (figure 9) but su bsequently the outer margin also rolls inwards to result in an involute vernation. The veins develop from the middle layer. The cells of middle layer not involved in the formation of veins, and the cells of the abaxial layer, divide periclinally and contribute to the mesophyll. The mature lamina has about 8 layers of cells consisting

482 K Periasamy and E A Muru9anathan
of the upper and lower epidermal layers, a single layer of palisade parenchyma and 5 layers of spongy mesophyll.
3.6 Scale leaf
The 3 or 4 scale leaf primordia, formed before each foliage leaf, elongate along with the latter. While the foliage leaf attains a height of 100-130cm, the outermost (oldest) scale leaf alone grows to about 25-30 cm, the inner ones remaining much shorter.
The development of a scale leaf is similar to that of a foliage leaf up to the formation of the triarch folding of the lamina wing. Subsequently, however, the lamina wing aborts and shrivels up (figure 8) leaving a black spot at the tip. After this the sheath grows markedly and the lamina petiole region is completely suppressed.
3.7 Leaf venation
Each leaf of both the species, contains 300-400 independently running veins from the corm to the leaflets. The leaflet venation is camptodromous. Each leaflet has 10-15 lateral veins which unite inside the margin to f o r m a prominent intramarginal vein. The median vein runs from the base to the tip of the leaflet, while the lateral veins numbering 20-30 run parallel to the median until they bend at right angles and deviate as lateral veins. The lateral veins and the median veins of the leaflets con- stitute the primary veins since they are all derived directly from the corŸ (figure 21).
3.8 Development of venation
When the leaf primordium is aboat 120 pm in height, the first median procambial strand appears at the place of insertion of the primordium and develops acropetally. This median dorsal strand becomes the median vein of the median leaflet without branching (figure 10). The subsequent strands arise laterally on either side of this median strand in a marginally directed sequence to fo rma tangential row of bundles and develop acropetaUy. Each lateral strand of the first row bifurcates before entering the lamina. One branch enters a leaflet and becomes its median vein (figures 10, 21) while the other enters the immediate older leaflet and becomes a lateral vein. The first formed tangential row of bundles forro mainly the median veins of leaflets (figure 21). The strands of the rows other than the first do not bifurcate before entering the leaflet midrib.
The second tangential row of bundles ate laid down adaxial to and alternating with the first row of bundles. The two median strands of this second tangential row enter the first formed leaflet and run parallel to the median strand (figure 21) and become lateral veins. Each of the remaining strands of this tangential row also enters a later formed leaflet and becomes one of its lateral veins. Subsequently more and more additional adaxial tangential rows are formed in a basipetal sequence and each strand enters a leaflet to become a lateral vein. In the scale leaves, usually after the formation of the first tangential row of strands r,o more additional rows ate laid down.

Leaf ontogeny in Arisaema Spp. 483
Figure 21. A. leschenaultii. Cleared leal primordium showing the venation pattern. (DM, Dorsal median strand: LI. lateral strands of the first tangential row that bifurcate before entering the leaflet" L2, lateral strands of the ser tangential row that do not bifurcate before entering the leaflet).
3.9 Structure and development of stomata
The leaves are hypos tomat ic and the s tomata are paracyt ic in bo th species. In ,4. les- chenaultii the s tomata l index and frequency ate respectively 14.8 and 82"1 and in A. tortuosum 17-8 and 123.2. Dur ing development, a small t r iangular meris temoid cut off by a polygonal protodermal cell functions directly as the guard mother cell and gives rise to the two guard cells.
4. Discussion
4.1 Lea.['succession
Arisaema possess only one of two mature foliage leaf a t a time and the next foliage leaf does not develop until the existing one dies. This is a unique feature in the

484 K Periasamy and E A Muruganathan
angiosperms and these plants may be called 'mono or bifoliar' in contrast to the usually multifoliar condition. Every foliage leaf is usually followed by scale leaves whose early ontogeny is similar to that of the foliage leal'. Subsequently the lamina wing aborts in the scale leaf followed by conspicuous development of the sheath part, whereas in the foliage leaf, the sheath is suppressed and the lamina-petiole part develops conspicuously. This indicates that from the time of its initiation, the lamina has an inhibitory effect on the sheath portion. A similar inhibition by the mature and physiologically active lamina may be instrumental in arresting lamina development in the primordia that follow to reduce them to scale leaves.
While the inhibition of axillary buds by apical dominance is well known in angiosperms, the inhibition of the sheath by the lamina and the lamina by the immediately older foliage leal appears to be a special phenomenon. In this regard it is worth mentioning that among pteridophytes, there are monofoliar forms like Ophioglossam, Botrychium and Helminthostachys, the last two resembling Arisaema superficially. This feature assumes some significance in the light of the monocot o¡ of angiosperms through pteridophyte ancestors put forward by Burger (1981).
4.2 Leaf initiation
In Arisaerna, the successive leaf p�98 diverge 180 ~ resulting in distichous phyUotaxy. The shoot apex, du¡ its growth from the minimal to maximal phase, grows more towards the side opposite to the youngest leaf primordium and becomes displaced a s a whole by 180 ~ during suciessive plastochrons. This behaviour of the shoot apex has been termed as pendulum symmetry (Goebel 1928; K• 1973). A meaningful interpretation of this would be to assume that the shoot apr regains its maximal phase by the recouperation of exactly the same portion set apart or lost during leal" initiation without much alteration or addition to the other portions. Such a concept would also account for the spiral displacement or circumnutation of the shoot apex.
From what is stated above, leaf initiation may be viewed as the fractionation or setting apart of a part of the indeterminate shoot apex as a determinate leaf primordial meristem. Ir the shoot apex is considr as a continuing me¡ residue (Newman 1965) the portion set apart for the leaf primordium can be termed as a terminating meristematic residue whose capacity for perpetuating growth (Periasamy 1965) and production of derivatives persists only fo r a short period in contrast to the shoot apex.
In this context, the two p¡ types of growth, monopodiat and sympodial are perhaps the result of the relative proportion of the shoot apex that is fractionated for indeterminate and determinate growths. Where the indeterminate fraction is relatively larger, the growth woutd be ctearly monopodial, but tend to be sympodial where the indeterminate fraction becomes smaller and atso remains inactive f o r a long time.
4.3 Leaf differentiation
One important aspect of the leaf primordium of Arisaema is that the character of the 5 layered laminar meristem exhibited immediately after its initiation is maintained at

Leaf ontogeny in Arisaema Spp. 485
the apex of the primordium and in the lamina wing until the initiation folds in it and the formation of the plate meristem. In the microphotographs of leaf ontogeny in Acorus presented by Kaplan (1970a) also, figure 7 shows a clear 5 layered laminar plate meristem character of the primordium as ir begins to protrude above the level of the shoot apex. So it appears that in these plants the lamina is initiated first and this perpetuates itself at the tip of the primordium and later becomes the lamina wing, while the basal derivatives give rise to the petiole and midrib by basipetal growth and maturation. Such a perpetuation growth has been reported in the palm leaf primordium by Periasamy (1965).
Previous workers on the compound leaf (Foster 1936; Denne 1966; Hagemann 1970; Kaplan 1970b; Coleman and Greyson 1976; Merril 1979) have reported that leaflets are initiated by the fractionation of an initial marginal meristem in both pinnately and palmately compound forms. But Periasamy and Muruganathan (1985a, b) have shown that in the palmately compound leaves of Tabebuia pentaphylla and Dioscorea pentaphylla, the leaflets are initiated even before the establishment of the marginal meristem. In Arisaema, leaflet formation is unique since they are formed by the folding of the lamina wing and this has some resemblance to what happens in palms.
In the palm leaf (Periasamy 1962) the lamina wing gives rise to submarginal plications by a characteristic growth pattern leaving the margin straight; the submarginal plications elongate both along the adaxial and abaxial ridges while the intercalary meristem that connects them expands into the lamina. But in Arisaema, the folds are merely mechanical and the outer edges of the folds corresponding to the abaxial ridges of the palm leaf plications alone elongate and later become the leaflet midribs while the inner edges corresponding to the adaxial ridges of iAications are suppressed, resulting in the isolation of each fold into an individual leaflet early in ontogeny. In palms the lamina breaks up into leaflets or gets dissected only when the leaf emerges from the crown and unfolds (Periasamy 1965).
4.4 Leaf venation
Leaf venation of Arisaema is reticulate comptodromous unlike the parallel venation of palms but the vascular framework and its ontogeny in the sheath and petiole resemble those of palms (Periasamy 1966). In both, the median dorsal bundle of the petiole arises first, followed by additional tangential lateral series, and aU the bundles remain independent until they reach the lamina. In palms, there is no diversion of bundles into the lamina from the vascular complex of the leaflet midrib. But in Arisaema, the early formed lateral bundles traverse along the middle of leaflets to form the first bundle of their midrib, and the later formed bundles run parallel to the first to varying distances in the midrib and diverge into the lamina as lateral veins. The bifurcation of the early formed primary strands of the petiole in Arisaema before entering the lamina, resembles what is found in Phoenix in which the strands bifurcate twice before entering the haut formed by the fusion of the adaxial ridges of the plications (Periasamy 1966).
In Populus (Isebrands and Larson 1980) and Acorus (Kaplan 1970a) the ontogeny and course of venation is essentially similar to that of Arisaema. This type appears to be different from the one where the midrib has a single composite vasculature from

486 K Periasamy and E A Muru9anathan
which the secondary veins branch off. The latter condition is found in dicotyledons in general (Avery 1933; Merril 1979) but is seen in the monocotyledonous Dioscorea also (Periasamy and Muruganathan 1985b). The tendency for the union of periolar bundles is seen in the dicotyledonous Tabebuia (Periasamy and Muruganathan 1985a) in which the strands either unite among themselves or with the p¡ midrib strands at various levels in the rachis.
Acknowledgement
We thank Otto L Stein of the University of Massachussetts, USA, for making the SEM of leaf primordia which was supported by NSF grant BMS 75-02883 and faeulty research grants from the University of Massachussetts.
References
Avery G S Jr. 1933 Structure and development of the tobacco leaf; Ara. J. Bot. 20 565-592 Burger W C 1981 Heresy revived; The monocot theory of Angiosperm origin; Evol. Theory 5 189-225 Coleman W K and Greyson R I 1976 The growth and development of the leaf in tomato (Lycopersicon
esculentum) II. Leaf ontogeny; Can. J. Bot. 54 2704-2717 Denne M P 1966 Leaf development in Trifolium repens; Bot. Gaz. 127 202-210 Foster A S 1936 Leaf differentiation in angiosperms; Bot. Rey. 2 349-372 Goebel K 1728 Organographie der Pflanzen Teil. I Allgemeine organo9raphie; Gustar Fischer, Jena Hagemann W 1970 Studien zur Entwicklungsgeschichte der Angiospermenblatter; 8ot. Jb. 90 297-413 Herr J M Jr. 1971 A new clearing squash technique for the study of ovule development in angiosperms;
Aro. J. Bot. 58 785-790 Isebrands J G and Larson P R 1980 Ontogeny of major veins in the lamina ofPopulus deltoides Bartr;, Aro.
J. Bot. 67 23-33 Kaplan D R 1970a Comparative foliar histogenesis in Acorus calamus and its bea¡ on the Phyllode
theory of monocotyledonous leaves; Aro. d. 8ot. 57 331-361 Kaplan D R 1970b Comparative development and morphological interpretation of "rachis leaves" in
Umbelliferae; New Res. Plato Anat. 63 101-125 (Suppl.) Kaplan D R 1973 Comparative developmental analysis of the heteroblastic leal series of axillary shoots of
Acorus calamus L. (Araceae); La Cellule. 69 253-290 Merrill E K 1979 Comparison of ontogeny of three types of leaf architecture in Sorbus L.; Bot. Gaz. 140
328-337 Newmann I V 1956 Pattern in meristems of vascular plants. I. Cell partition in living apices and in the
cambial zone in relation to the concepts of initial cells and apical ceUs; Phytomorphology 6 1-19 Periasamy K 1962 Morphological and ontogenetic studies in palms. I. Development of the plicate
condition in the palm leaf; Phytomorphology 12 54-64 Pe¡ K 1965 Morphotogical and ontogenetic studies in palms. II. Growth pattern of the leaves of
Cocos nacifera and Borassusflabellifer after the initiation of plication; Aust. J. Bot. 13 225-234 Periasamy K 1966 Morphological and ontogenetic studies in palms. IV. Ontogeny of the vascular pattern
in four genera of palms; Aust. J. Bot. 14 277-91 Periasamy K 1967 A technique of staining sections of paraftin-embedded plant materials without
employing a graded ethanol se¡ J. R. Micross. Soc. 87 109-112 Periasamy K and Muruganathan E A 1985a Ontogeny of palmately compound leaves in angiosperms.
1. Tabebuia pentaphylla Hense; Proc. Indian Acad. Sci. (Plant Sci.) 95 429--436 Periasamy K and Muruganathan E A 1985b Ontogeny of palmatety compound leaves in angiosperms.
2. Dioscorea pentaphyUa L; Indian Botanical Cono'actor 2 75-84





























