Embed Size (px)
DESCRIPTION
Ontogeny
Citation preview

Museum fuÈ r Naturkunde, Institut fuÈ r Systematische Zoologie, Berlin, Germany
Early ontogeny and placentation of the grey short-tailed opossum, Monodelphisdomestica (Didelphidae: Marsupialia): contribution to the reconstructionof the marsupial morphotype
U. ZELLER and C. FREYER
AbstractThis study provides new ®ndings on the placenta of Monodelphis domestica and a reconstruction of the marsupial morphotype. To achieve this,early ontogeny and placentation of the grey short-tailed opossum, M. domestica, from 3 h after copulation until birth (day 15), were studied andcompared with other mammals. Both the ultrastructure and histochemistry of egg membranes, foetal membranes, oviduct and uterus wereexamined. The results of this study provide the ®rst detailed ultrastructural description of a trophoblastic syncytium in a marsupial. In addition,this is the ®rst original documentation of an invasive trophectoderm and an in¯ammatory reaction at parturition in M. domestica. These ®ndingswere compared with literature data and included into the reconstruction of the marsupial morphotype. Based on marsupial phylogeny asproposed by Luckett (J. Mammal. Evol. 2, 255±283, 1994), characters that are consistent at least within didelphids and dasyurids were determinedto be characters of the marsupial morphotype. These characters are a central yolk separated from the peripheral yolk-poor cytoplasm in theunfertilized oocyte, the presence of a zona pellucida, a mucoid coat and a shell coat, the absence of a corona radiata, oviductal mucoid secretion,no shell secretion distal to the isthmus of the oviduct, uterine shell secretion, a short tubal passage (1 day at maximum), the apposition ofblastomeres to the zona pellucida prior to intercellular association, the absence of a morula stage, the polarity of the zygotic yolk, the localizedsegmentation of deutoplasm (yolk) during the ®rst cleavage and subsequent extrusion of yolk vesicles during the ®rst two cleavage stages. Withregard to the marsupial morphotype, the non-polarized yolk distribution in the zygote [Hartman (J. Morphol. 27, 1±84, 1916); McCrady (Am.Anat. Mem. 16, 1±233, 1938)] is a derived character of Didelphis virginiana. Didelphis virginiana [Hartman (J. Morphol. 27, 1±84, 1916); Hartman(J. Morphol. 32, 1±139, 1919); McCrady (Am. Anat. Mem. 16, 1±233, 1938)] and Didelphis marsupialis (Hill, Q. J. Micr. Sci. 63, 91±139, 1918)share the synapomorphous reduction of deutoplasmolysis to a generalized extrusion of vesicles. The absence of separated yolk and consequently acleavage without yolk extrusion (Renfree and Lewis, Reprod. Fert. Dev. 8, 725±742, 1996) are apomorphies of macropodids. This is possiblycorrelated with the association of blastomeres in early cleavage stages (Renfree and Lewis, Reprod. Fert. Dev. 8, 725±742, 1996). A yolk sacplacenta and a vascularized allantochorion can be assumed for part of the ontogeny in the marsupial morphotype, irrespective of the formation ofan allantoic placenta at near term stages. The character polarization of the mode of placentation and parturition needs further investigation.
Key words: Monodelphis domestica ± Marsupialia ± early ontogeny ± evolution ± morphotype ± placentation ± phylogeny ± reproduction
Abbreviations: bc, blastocyst cavity; bm, blastomeres; CRL, crown±rump length; DAB, diaminobenzidine; dpc, days post copulation; em,endometrium; EM, electron microscopy; ER, endoplasmatic reticulum; ESI, electron spectroscopic imaging; fbc, foetal blood cell; fc, formativecytoplasm; HC, histochemistry; hpc, hours post copulation; ht, histiotrophe; ic, intercellular capillary; l, lipid droplet; LM, light microscopy; mc,mucoid coat; me, mesenchyme; mt, mitochondrion; n, nucleus; ne, nucleated erythrocytes; PAS, Periodic Acid Schi�'s Reagent; pp, pseudopodia-like process; pr, propria; r, ribosomes; rer, rER, rough endoplasmatic reticulum; S.E.M., Standard Error of Mean; SEM, scanning electronmicroscopy; sl, syncytial knot; sm, shell coat; sp, secretory process; st, sperm tail; te, tubal epithelium; TEM, transmission electron microscopy; tr,trophoblast; trc, trophoblast cell; trs, trophoblastic syncytium; ue, uterine epithelium; ug, uterine gland; ul, uterine lumen; uv, uterine vessel; vv,vitelline vessel; y, yolk; yp, yolky cytoplasm; zp, zona pellucida
Introduction
This study is a contribution to the reconstruction of themorphotype of recent marsupials by providing current ®nd-ings, a character polarization and a phylogenetic interpret-ation.
Marsupialia (Metatheria) and Placentalia (Eutheria) formthe monophyletic Theria, which are the sistergroup of theMonotremata (Novacek et al. 1988; Shoshani and McKenna
1998; Zeller 1999b). Although marsupials and placentals areboth viviparous (unlike the egg-laying monotremes) theirmodes of reproduction di�er considerably (see Table 1). The
polarization of most of these characters, as well as theirevolutionary signi®cance are still unresolved. For the recon-struction of the evolution of reproductive strategies in both
groups, the morphotypes, which are the sum of characterspresent in the last common ancestors (cf. Hennig 1950;KoÈ nigsmann 1975), of Theria, Marsupialia, Placentalia andinternal groups must be reconstructed.
The morphotype reconstruction is based on marsupialsystematics as proposed by Luckett (1994). According to
Luckett (1994),marsupials can be divided into themonophyleticsister-taxa Australidelphia and Ameridelphia. Ameridelphianscomprise all American marsupials except Dromiciops australis,australidelphians consist of all Australasian marsupials inclu-
ding the South American Dromiciops australis. The monophylyof these groups is supported by the sperm pairing in theepididymis of ameridelphians (Temple-Smith 1987) and the
fusion of two facets on the calcaneus (con¯uent lower ankle jointpattern, Szalay 1993) in australidelphians. This basal dichotomyof marsupials is essential for the morphotype reconstruction,
based on character optimization and outgroup comparison. Forcharacter optimization and outgroup comparison the furtherdivision into syndactyls and diprotodonts is useful. The phylo-
geny of marsupials is represented by the cladogram in Fig. 1.The di�erences in reproductive biology could have had
important in¯uence on the di�erences in extinction rates,radiation and migration between marsupials and placentals at
the Cretaceous±Tertiary boundary (Archibald 1982; Case andWoodburne 1986; Caroll 1997). We regard the reconstructionof the marsupial morphotype as a ®rst step toward an
U.S. Copyright Clearance Center Code Statement: 0947±5745/01/3903±0137$15.00/0 www.blackwell.de/synergy
J. Zool. Syst. Evol. Research 39 (2001) 137±158Ó 2001 Blackwell Wissenschafts-Verlag, BerlinISSN 0947±5745
Received on 15 February 2000

Table
1.Charactersofreproductionin
placentalsandmarsupials
Charactersof
reproduction
Marsupials
Placentals
neonate
Extrem
elyaltricial,activemovem
entto
nipple,®xed
tonipple
for14±127days1
,2Precocialrelativeto
marsupialneonates,never
®xed
tonipple2
Pregnancy
Alwaysshort
intrauterinedevelopment2:9.5
3±49.7
4days(includingsperm
storage,
Longintrauterinedevelopmentpossible2:165±6686days
possible
arrestofconceptusdevelopment)
Oocyte
Largeatovulation,manyyolk
bodies7
,8Small,less
yolk
bodies7
,8
Zonapellucida
Thinner
thanin
placentals2,9:110±8.6
11
lm10±30
lm12,13
Cleavage
Includingdeutoplasm
olysisthatreleasesmaterialasmem
brane-bound
Withoutdeutoplasm
olysisthatreleasesmaterialasmem
branebound
vesicles7
,14±18
vesicles7
,14,15
Start
ofcleavage
Intheuterus2
,14±16,18
Inmost
casesin
theoviduct;exception:Elephantulusmyurus1
9
Morula
stage
Absent(directblastocyst
form
ation)2,16,18,20
Inmost
casespresent(w
hen
enteringtheuterus)14,exception:threegenera
ofinsectivora
21
Entypic
condition
Absent10,14,16,18,22±24
Present14,32,(inner
cellmass
form
slaterin
threegeneraofinsectivora
21)
oftheblastocyst
Eggcoats
Shellcoat,mucoid
coat9,25
Absence
ofshellcoat,absence
ofmucoid
25;exceptionin
lagomorphs2
5
andhorse9
,26
Placenta
Seldom
involvingtheallantois2,15,27±29,never
mesodermalvilli
Alwaysinvolvingtheallantois15,26,27,29,30,31Mesodermalvilliattheside
atthesideoftheallantochorion31
oftheallantochorion30,32
Lactationphase
Alwayslonger
thanintrauterinedevelopment:50days(M
onodelphis,
Canbeshorter
thanintrauterinedevelopment2:5days3
3±4.5
years
34
Cercartetus)±540days(M
acropusgiganteus,M.fuliginosus)2
Oestrouscycle
Since
longer
thanpregnancy
nota�ectedbypregnancy
±exception:Wallabia
bicolor,
Usuallya�ectedbypregnancy
37
Macropusgiganteus3
5,36a�ectedbylactation2,37
1Renfree
(1994);
2Tyndale-BiscoeandRenfree
(1987);
3Sminthopsismacroura,SelwoodandWoolley
(1991);
4Dasykaluta
rosamondae,
Woolley
(1991);
5Mesocricetusauratus,
Lochbrunner
(1956);
6Elephasmaxim
us,Nowak(1999);
7SelwoodandSathananthan(1988);
8Renfree
andLew
is(1996);
9Selwood(2000);
10Hartman(1916);
11Selwoodet
al.(1997);
12Austin
(1961);
13Bedford
(1996);
14Hill
(1910);
15Cruzand
Pedersen
(1991);
16Selwood
(1992);
17Selwood
(1980);
18Selwood
and
Young(1983);
19van
der
Horst(1942)cited
inTyndale-Biscoeand
Renfree
(1987);
20Selwood
(1996);
21Elephantulusmyurous,vander
Horst(1942);Hem
icentetes:BluÈntschli(1937);Goetz(1938);Eremitalpa,Gabie
1959allcitedin
Mossman(1987);
22MacC
rady(1938);
23Selwood(1986a);
24Selwood
(1986b);
25Hughes
(1977);
26Herrler
etal.(1997);
27Amoroso
(1952);
28Morris(1975);
29Padykula
andTaylor(1976);
30Luckett(1977);
31WoodingandFlint(1994);
32Mossman(1987);
33Cavia,Nowak
(1999);
34Pantroglodytes,Nowak(1999);
35Tyndale-Biscoe(1973),p.53;36Renfree
(1980);
37HeapandFlint(1984).
138 ZELLER and FREYER

evolutionary scenario that explains the mammalian evolutionat the Cretaceous±Tertiary boundary.
Materials and methodsMonodelphis domestica was bred in a colony established at the Instituteof Anatomy of the University of GoÈ ttingen. This colony was latermoved to the Institute of Zoology of the University of Mainz and thenon to the Institute of Systematic Zoology at the Museum of NaturalHistory, Berlin, where it is presently located. Estrus of femaleMonodelphis is induced by the presence of male pheromones (Fadem1985; Hinds et al. 1992). Copulation takes place within the estrus(Baggott et al. 1987; Fadem and Rayve 1985). Ovulation occurs 20(Mate et al. 1994), 22.5 (n� 1, own observation) to 24 h (Baggott andMoore 1990) after copulation. Fertilization occurs 22 (Mate et al. 1994),22.5 (n� 1, own observation) to 24 h after copulation (Mate et al. 1994).Mating was monitored by direct observation. All embryos and stagesmentioned in this study are dated according to the time of copulation.Thus, the ®rst day of pregnancy coincides with the day of fertilization,that is, about 24 h aftermating. Parturition occurs between the 14th and15th day after copulation (Fadem et al. 1982; personal observation)after a 13- to13.5-day period of gestation (Mate et al. 1994).
The consistency between our ®ndings and descriptions of Mate et al.(1994) con®rmed the normality of embryos described in this study.Abnormal stages were not included in the description. In addition, thechronology of development in M. domestica as described here matchesthe timetable given by Mate et al. (1994).
Twenty-seven adult female M. domestica were killed with chloro-form at di�erent times of pregnancy, ranging between a few hours and15 days after copulation. They were ®xed by perfusion through thevascular system immediately after their death. Gravid uteri, embryosand pieces of yolk-sac placenta were prepared for histochemistry, lightand electron microscopy (EM) according to the following procedures.
Light microscopy
Specimens were either (1) ®xed with Bouin¢ s and embedded inparaplast or (2) ®xed with 4% formol and embedded in methylmeth-
acrylate (Kulzer). Specimens embedded in paraplast were sectioned at7 lm and stained with hematoxylin and eosin (HE), azan or trichrome(Mason-Goldner). Specimens embedded in methylmethacrylate weresectioned at 2 lm and stained with HE.Frozen sections were obtained from tissues shock frozen in liquid N2
and later sectioned at 4±6 lm with a Kryostat (Leica1 Frigocut, LeicaInstruments GmbH, Heidelbergerstr. 17±19, 69226 Nuûloch, Ger-many) at )25 °C.
Histochemistry
Four methacrylate sections of specimen 82 [29 h post copulation (hpc)]and one frozen section of specimen 135 [7 days post copulation (dpc)]were stained with Kongo red (Romeis 1968). One frozen section ofspecimen 135 was also stained for keratin by Martinotti (Romeis 1968)to demonstrate the shell coat. Numerous frozen sections of specimen135 were stained with Sudan red.Twenty-four methacrylate sections of specimens 45 (22.5 hpc), four
of specimen 85 (29 hpc) and eight of specimen 95 (4dpc) were stainedwith PAS. Six para�n sections of specimen 45 (22.5 hpc) were stainedwith peroxidase-conjugated Ulex-europaeus-lectin (substrate: DAB).Four frozen sections of specimen 135 were ®xed in acetone (6 °C) for
10 min and treated with antibodies against keratin, laminin, collagentype IV and ®bronectin (Table 2); the immunoreaction was visualizedby the usage of biotinylated secondary antibodies and the biotin±avidin±peroxidase method according to Dako22 LSABÒ-Kit (DAKODiagnostika GmbH, Hamburg, Germany). In addition, sections weretreated with collagenase (Sigma no. C-0773; Sigma-Aldrich ChemieGmbH, Taufkirchen, Germany33 ). Antibodies against transferrin wereapplied to two frozen sections of specimen 132 and to two para�nsections of specimen 28 (12 dpc), followed by the application of DakoLSABÒ-Kit.Control: sections treated with all chemicals used in immunohisto-
chemistry without the primary antibody did not stain.
Electron microscopy
For transmission electron microscopy (TEM), specimens were ®xed byperfusion through the maternal vascular system with a mixture of1.5% glutaraldehyde and 1.5% freshly prepared paraformaldehyde in0.1 M cacodylate bu�er (pH 7.35; total osmolality 658 mosmol/l).Specimens of yolk-sac placenta (endometrium with adjacent yolk-sacwall) prepared for TEM were cut into small pieces and further ®xed for1 h with glutaraldehyde/paraformaldehyde in cacodylate bu�er. Tissueblocks were rinsed for 1 h in cacodylate bu�er, post-®xed for 2 h with1% osmium tetroxide by immersion (specimen 194 (14 dpc) with 1%tannic acid) and embedded in araldite after dehydration with ethanol.Ultrathin sections (60±80 nm) were stained with lead citrate and uranylacetate and examined with a Zeiss44 CEM 902 electron microscope (CarlZeiss Oberkochen, Germany) operating at 80 kV with an integratedimaging electron energy spectrometer and a Phillips55 CM 10 (PhillipsElectron Optics, Chatswood, NSW, Australia) operating at 60 kV.Electron spectroscopic imaging (ESI) without elemental analysis wasperformed at energy losses of either 0 eV (`elastic bright ®eld') or 250eV (with the carbon signal at minimum). For scanning electronmicroscopy (SEM), embryos or pieces of endometrium were driedaccording to the critical point method, sputtered with gold palladiumand examined with the Zeiss DSM 960 scanning electron microscope.
Photography
Whole embryos were photographed with a Wild66 macroscope M 400(Leica), and a Zeiss Axioplan was used for microphotography.The specimens and procedures adopted are summarized in Table 2.
Taxonomy
Species names are used according to the nomenclature of Wilson andReeder (1993). If authors cited in this paper used synonyms, the correctname is given in square brackets. Classi®cation of species into highertaxonomic units also follows the taxonomy of Wilson and Reeder(1993). The species included in both the discussion and the morpho-type reconstruction belong to the higher marsupial taxa as summarizedin Table 3.
Fig. 1. Cladogram representing marsupial systematics as proposed byLuckett (1994). 1, Ameridelphia; 2, Australidelphia; 3, Syndactyla;4, Dasyuroidea; 5, Diprotodontia; 6, most phalangeroids; 7, Vombati-formes; 8, Macropodoidea
Early ontogeny and placentation of the grey short-tailed opossum 139

Phylogenetic reconstruction
Reconstruction of morphotypes follows the algorithm for reconstruct-ing ancestral states using parsimony as described by Cunningham et al.(1998). As pointed out by Cunningham et al. (1998), `the algorithm usesa ``downpass'' and ``uppass'' traversal to optimize ancestral states usingtwo rules: Rule 1: if descendant nodes share any states in common,assign the set of shared states to the ancestor; Rule 2: if no states are
shared in descendant nodes, assign the union of descendant's states toancestor.' (p. 362, Box 1). According to the algorithm, the downpassoptimization proceeds `down' the tree towards the root, optimizingeach ancestral node. The uppass optimization proceeds `up' the treeaway from the root, optimizing each ancestral node. In a ®naloptimization, the sets of downpass and uppass reconstruction areoptimized to the state that has the greatest number in both reconstruc-
Table 2. Female Monodelphis domestica investigated in this study
Stage Specimen no. Methods
3 hpc 68 LM, SEM4 hpc 81 LM22.5 hpc 45 HC (PAS, Ulex-europaeus-lectin (DAB)), LM24 hpc 36 LM, SEM29 hpc 85 HC (Kongo red, PAS), LM31 hpc 118 LM, immunohistochemistry: Dako EPOS: mouse-antihuman antibodies against keratin,
clone MNF 116, Code No. U7022, against keratin 10, 17, 18 (45±56.5 kDa), DAKO EPOS CK10,DE-K10, Lot 043011, against keratin 10
4 dpc 95 LM5 dpc 128 LM6 dpc 102, 145 LM7 dpc 135 left uterus: LM
right uterus: LM, HC: Kongo red, Sudan red, Keratin (Martinotti), collagenase (Sigma no. C-0733);immunohistochemistry:
(1) tissuGnost (Merck): mouse-anti-human antibodies against intermediate keratin ®laments(54 kDa), LSAB-Kit (Dako)
(2) Dako EPOS: mouse-anti-human antibodies against keratin, clone MNF 116, Code No. U7022,against keratin 10, 17, 18 (45±56.5 kDa)
(3) Dako Collagen Type IV (CIV 22): mouse-anti-human antibodies against collagen(Code No. M 785. Lot no. 102, 1 : 100), LSAB-kit
(4) Dako Laminin (4C7): mouse-anti-human antibodies against laminin (Code No. M 638,Lot no. 121, 1 : 25), LSAB-kit
(5) Dako Fibronectin: rabbit-anti-human antibodies against ®bronectin (Code No. A 245,Lot. no. 117, 1 : 400), LSAB-kit
8 dpc 146 LM10 dpc 101 LM
156 TEM11 dpc 187, 231 LM
8, 46 LM, TEM132 Left uterus: HC
Immunohistochemistry: Dako anti-transferrin, rabbit-anti-human antibodies against transferrin(Code No. A 0621 02, Lot. no. 040 D, 1 : 800), LSAB-kit.
12 dpc 28, 44 LM, TEMHCImmunohistochemistry: Dako anti-transferrin (same as in 11 dpc)
13 dpc 12 LM, TEM14 dpc 6, 194, 429 LM, TEM
677 LM15 dpc 122 LM
HC, histochemistry; hpc, hours post copulation; dpc, days post copulation; LM, light microscopy; SEM, scanning electron microscopy; TEM,transmission electron microscopy.
Taxon Species
Didelphidae Monodelphis domestica, Didelphis marsupialis, Didelphis virginiana, Philander opossumDasyuridae Dasyurus viverrinus, Sminthopsis crassicaudata, Sminthopsis macroura, Antechinus
stuartii, Phascogale sp.Peramelidae Isoodon obesulus, Isoodon macrourus, Perameles nasuta, Perameles gunnii, Echymipera
rufescens, Echymipera kalubuVombatidae Vombatus ursinusPhascolarctidae Phascolarctos cinereusPseudocheiridae Petauroides volans, Pseudocheirus peregrinusPhalangeridae Trichosurus vulpeculaAcrobatidae Acrobates pygmeus, Distoechurus pennatusMacropodidae Macropus eugenii, Petrogale sp., Macropus robustus, Macropus giganteus,
Macropus rufogriseus, Setonix brachyurusPotoroidae Potorous tridactylus, Bettongia cuniculus
Table 3. Species mentioned in thisstudy and their relation to highermarsupial taxa
140 ZELLER and FREYER

tions. This procedure implies that there is no constant outgroup.Instead, the outgroup changes as does the respective ingroup during theprocess of optimizing within the group of investigation.
This procedure is conducted using the cladogram by Luckett (1994).We favour this hypothesis of marsupial phylogeny, because it is basedon total evidence (molecular and morphological data). Since ontoge-netic data are often restricted to didelphids and dasyurids, which arerepresentatives of ameridelphians and australidelphians, respectively,consistent data within both groups are assumed to be characters of themarsupial morphotype. Certain of these morphotype hypotheses canbe supported by ®ndings among additional australidelphians. Incon-sistent data within ameridelphians or australidelphians were polarizedusing the mentioned method for morphotype reconstruction.
Due to insu�cient and inconsistent data concerning marsupialplacentation, reconstruction of the allantochorion in the marsupialmorphotype was separately conducted by optimizing with the sister-group (Placentalia) of marsupials and the outgroup of the Theria(Monotremata). In some cases, the state of marsupial morphotypecharacters (e.g. zona pellucida, mucoid coat) is determined byoutgroup comparison with monotremes.
Results
By 3 hpc, 11 tertiary follicles were present in the right ovary
investigated. The oocytes contain large yolk vacuoles and aresurrounded by follicular epithelial cells. Paired spermatozoacan be seen in the oviduct on SEM images. One hour later, the
follicle epithelium degenerates.By 22.5 hpc, the corpora lutea have developed. The eight
oocytes passing each oviduct lack a corona radiata and are
merely surrounded by the zona pellucida, which is PAS-reactive. They measure 170 lm in diameter. The oocytenucleus is located in the peripheral, formative cytoplasm,which contains minimal yolk (Fig. 2a). The formative, yolk-
free cytoplasm encloses the yolky deuteroplasm (Fig. 2a),which in turn encloses a small central zone that contains littleyolk. Paired spermatozoa are visible inside the oviduct by light
microscopy (LM) and SEM (24 hpc). Fertilization occurs inthe oviduct (Fig. 2a). At this time, the second polar body isseparated o�. The surface of the zygote is in close association
with the tubal epithelium (Fig. 2a) and binds the lectin Ulexeuropaeus agglutinin (UEA) like oviduct cells, but unlike theuterine epithelium. Mucoproteid layers covering the surface ofthe conceptus are secreted by non-ciliated oviductal cells.
Corpora lutea are already present in the ovary.By 29 hpc, the zygotes (n� 5) have entered the uterus in the
pronuclear stage. Thus, a maximum of 7 h is needed for the
passage down the oviduct. Each zygote is covered by aconcentric, lamellar mucoproteid layer (mucoid coat)(Fig. 2b), in which spermatozoa are trapped. A secondary
yolk polarity is achieved by polar accumulation of theformative cytoplasm around the pronuclei (Fig. 2b), whichfuse at 31 hpc. The uterine epithelium secretes the proteina-
ceous shell coat, which is Kongo red reactive but does not bindantibodies against keratin.
At the fourth day after copulation, the 16-cell stage (n� 1)has been achieved. The yolk, which is not a�ected by cleavage,
forms a compact mass opposite the blastomeres (Fig. 2c). Thisis due to deutoplasmolysis and apocrine yolk extrusion duringthe ®rst and second cleavage at 42±50 hpc and 58±68 hpc,
respectively (Fromm and Zeller 1997). The diameter of theconceptus measures 180 lm inside the zona pellucida and itsentire diameter measures 440 lm. The mucoid coat surround-
ing the zona pellucida consists of two layers. The thinner outerone is strong PAS reactive, whereas the thicker inner one is less
reactive to PAS. Pseudopodia-shaped processes of mucoidcoat and shell coat project into the niches of the endometrium(Fig. 2d). The shell coat cannot be stained with PAS.
By 5 dpc, the blastomeres of the conceptus (n� 7) containlarge yolk vacuoles. One day later, the zona pellucida is partlybroken down (n� 9). The blastomeres containing yolky lipid
droplets in vacuoles form the early unilaminar blastocyst. Themucoid coat is still visible.By 7 dpc (bilaminar blastocyst, n� 10), the mucoid coat and
yolk have been incorporated into the blastocyst and the
blastomeres are apposed to the peripheral shell coat (Fig. 2e).At this time, the diameter of the blastocyst measures 340 lmand almost doubles within one day (670 lm by 8 dpc). At this
stage the pseudopodia-shaped projections are absent. Theembryonic area partly consists of a thickened ectoderm withunderlying endoderm, and it does not form an inner compact
cell mass (Fig. 2e).The shell coat surrounding the blastocyst cannot be stained
with PAS or Alcian blue and does not bind antibodies against
cytokeratin, laminin or ®bronectin. It is resistant to collagen-ase, but can be stained for keratin by Martinotti, and Kongored. Keratin was not detectable by immunohistochemistry.By 10 dpc (embryo, ®ve to six somites, n� 6), the vesicle has
formed a bulging sphere 5 mm in diameter. The embryo has aneural groove, neural crest with a developing mesenchyme, aprimitive streak, a paraxial mesoderm with ®ve to six somites,
a lateral plate mesoderm with pericardium, extraembryonicmesoderm and a yolk sac cavity.One-third of the yolk sac lying towards the embryonic pole
forms the trilaminar omphalopleure, the abembryonic part isbilaminar. Ultrastructurally, the trophendoderm (yolk sacendoderm) consists of ¯at cells connected by cell projections
and tight junctions. Wing-shaped processes of trophendodermcells partly appose to the bases of trophectoderm cells inbilaminar omphalopleure and project with short processes intothe basal folding, whereas extended parts of trophectoderm
(ectoderm of the yolk sac) cells remain uncovered. The cellshave abundant polyribosomes, mitochondria, endoplasmaticreticula (ER) and lipid droplets.
The trophectoderm (outer wall of the yolk sac) is formed ofcolumnar to ¯attened distinct cells leaving small intercellularcapillaries for transport in between. Apices are united by
junctional complexes. Exocytotic vesicles secrete into wideintercellular gaps (about 15 lm in width) between trophendo-derm and trophectoderm. The apical surface is densely coveredwith microvilli directed to the endometrium. The cells contain
numerous mitochondria, smooth and rough ER, polyribo-somes, vacuoles and lipid droplets. Endocytotic complexes andtransport vesicles are present. The cell bases are folded. All this
indicates active transepithelial transport of material.The cells of the one-layered, high-columnar uterine epithe-
lium are densely covered with microvilli on their luminal
surface, their bases border a capillary net in the propria. Abasal lamina could not be found. The propria contains only fewcells. The endometrial glands consist of a thick pad of coiled
tubular glands, which in some areas extend for 3 mm. They aredevoid of glandular ducts and secrete histiotrophes directly intothe uterine lumen. The myometrium does not exceed 0.2 mm.The shell coat is still intact and measures 1.8 lm.
By 11 dpc (4 mm total length), the di�erentiation of thefrontal half of the embryo is highly advanced following rapiddevelopment. It shows a strong curvature of head and neck, a
dorsally concave lumbar curvature, a foregut with visceral
Early ontogeny and placentation of the grey short-tailed opossum 141

142 ZELLER and FREYER

clefts and visceral arches, brain plate with eye anlagen, earplacodes, a heart anlage, and crest-shaped anlagen of upperextremities. The vitelline duct is still wide open. The neuraltube, somites, notochord, aortae, coelom and nephrogenic
crest can be seen on transverse sections through the embryo.The cranial and caudal ends of the neural tube are still openand form neuropores, respectively. Two-thirds of the anterior
end of the embryo are sunk into the yolk sac and covered withthe two-layered proamnion (ecto- and endoderm). The caudalthird of the embryo is still part of the surface of the conceptus.
A small extra-embryonic coelom can be seen between thetrilaminar yolk sac and proamnion.
The yolk sac is trilaminar for over one-third of its extension.
It measures 5±6 mm in diameter and is greatly expanded withelaborate folds increasing the surface area (cf. Fig. 2f). It isclosely apposed to the endometrium (Fig. 3a). The endodermalcells of the bilaminar omphalopleure have long projections
enclosing large intercellular spaces between trophendodermand trophectoderm. In the trilaminar yolk sac wall theendodermal cells are ¯attened and closely connected to each
other. Blood vessels with endothelium and haemocytoblastsare developed within the mesenchyme of the trilaminar yolksac. The sinus terminalis appears at the border between the
bi-and trilaminar omphalopleure.The trophectoderm mainly consists of distinct cells united by
desmosomes with intercellular transport capillaries in between.In particular, the cells of the trophectoderm of the bilaminar
omphalopleure are ¯at and contain large vacuoles exceedingthe diameter of the cell nucleus. Ultrastructurally, the troph-ectoderm is formed in most parts by distinct cells, which are
apically united by junctional complexes (Figs 3b, 4a) withintercellular transport capillaries in between (Fig. 3b). Sometrophectodermal cells of the trilaminar omphalopleure have
already fused and intercellular borders can not be seen. Thenuclei of those cells are close to each other. The trophectodermis densely covered with microvilli (Figs 3a, 4a), and pinocytotic
vesicles are present on its surface. The cells contain numeroustransport vesicles, mitochondria, lipid droplets, rough andsmooth ER, free ribosomes, and big euchromatic nuclei with amaximum diameter of 21 lm (Figs 3b, 4a). This indicates the
contribution of the trophectoderm to both transport andsynthesis.
The cells of the single-layered, high-columnar uterine
epithelium contain often more than one large euchromaticnucleus (up to 15 lm in diameter) with prominent nucleoli
(Fig. 3a). They are connected by apical junctional complexes,and are covered with a dense apical border of microvilli of3 lm length (Fig. 3a). Secretory canaliculi are located
between the cells of the apocrine secreting epithelium. Thecells contain mitochondria, rER, polyribosomes, secretorygranules and abundant lipid droplets (Fig. 4a). The uterine
epithelium mainly serves as a pathway for materials from thematernal blood to the embryo. The basal plasmalemma isfolded and borders on a capillary net in the propria, which issituated 0.5±2 lm beneath the epithelium. A basal lamina
is lacking. The capillary endothelium is not fenestrated. Twolayers of endometrial glands can be distinguished: Theendometrial glands near the myometrium consist of a single-
layered secreting epithelium with basal nuclei and cytoplasmthat is comb shaped due to abundant inclusions. Theendometrial glands near the uterine epithelium have mostly
homogeneous cytoplasm and central nuclei, with inclusionsand a comb-shaped cytoplasm that is restricted to their bases.The cells of the uterine glands contain ergastoplasm, dictyo-
somes, mitochondria, lipid droplets and secretory granules.The surface is covered partly with branching microvilli andpartly with kinocilia. The uterine glands are the sites ofsynthesis and secretion of histiotrophes. The propria contains
more free cells (histiocytes, plasmacytes, lymphocytes, poly-morphonuclear granulocytes) on its subepithelial side than onits basal side.
The shell coat, which measures 2.3 lm, is unruptured at thisstage (Figs 3a, 4a). It consists of narrow meshes, through whichall material for the exchange between mother and embryo has
to pass. Shell coats of adjacent embryos do not fuse.By 12 dpc [5 mm crown±rump length (CRL)], the di�eren-
tiation of the embryo is more advanced, especially in the facial
and branchial regions as well as the upper extremities. Thelarge heart supports the yolk sac circulation. The lumbarcurvature is ¯attened out, the caudal amnion expanded and theembryo has completely sunk into the yolk sac. The exocoelom
is expanded.Only the yolk sac, which is now vascularized over two-thirds
of its extension, contributes to the placenta (Fig. 2f). The
endodermal cells of the vascular part are ¯at and in closecontact with each other. The yolk sac vessels are the sites ofblood formation by mitotic multiplication of proerythroblasts
and myeloblasts. Immunohistochemistry demonstrates trans-ferrin within the maternal epithelium as well as in thetrophectoderm and the underlaying vascular layer of theomphalochorion, indicating transferrin-rich histiotrophe pass-
ing the meshes of the shell coat (Zeller 1999a).The trophectoderm still consists in most parts of distinct,
laterally interconnected cells, that are united by apical junc-
tional complexes (compare Fig. 4a). The length of themicrovilli is reduced. The cells contain large vacuoles and bignuclei mainly arranged in a regular pattern. At some areas of
the trilaminar omphalopleure the nuclei of the trophectodermare situated close to each other and between those areas thetrophectoderm free of nuclei is ¯attened. The extent of folding
of the endometrium is increased (Fig. 2f). The length ofmicrovilli of the uterine epithelium is reduced (1.7 lm).Macro- and microapocrine secretion occurs. The endometrialglands secrete histiotrophes (Fig. 3c). The thickness of the
glandular pad is reduced to about 1.5 mm. The former two-layered condition of the glandular pad has been lost. The shellcoat, remnants of which can be seen between trophoblast and
endometrium (Fig. 3c), is ruptured.
Fig. 2. Monodelphis domestica, photomicrographs of early develop-ment from tubal zygote to placentation. (a) 22.5 hpc, tubal zygote. Theformative cytoplasm (fc) surrounds the central yolky cytoplasm (yp).The surface of the zygote is in close association with the tubalepithelium (te). �, sperm tails, scale bar� 10 lm. (b) 29 hpc, uterinezygote. The yolky cytoplasm (yp) is polarized. Pronuclei (arrows) arelocated in the formative cytoplasm (fc). arrows, pronuclei; *, shell coat;circle, artefact; scale bar� 20 lm. (c) 4 dpc, 16-cell stage. After yolkextrusion, the yolk (y) forms a compact mass opposite the blastomeres(�). scale bar� 50 lm. (d) 6 dpc. Pseudopodia-shaped processes (pp) ofthe mucoid coat (mc) and the shell membrane (asterisk) project intothe niches of the uterine epithelium (ue). bm, blastomeres; s, artefact;scale bar� 50 lm. (e) 7 dpc, bilaminar blastocyst. The embryoconsisting of a thickened ectoderm and underlying endoderm (arrows)does not form an inner compact cell mass. bc, blastocoel; ul, uterinelumen; scale bar� 50 lm. (f) 12 dpc. The yolk-sac placenta is greatlyexpanded with elaborate folds increasing the surface area. em,endometrium; arrows, yolk sac; �, embryo; scale bar� 500 lm
Early ontogeny and placentation of the grey short-tailed opossum 143

144 ZELLER and FREYER

By 13 dpc (8.5 mm CRL), the neural tube is closed,peripheral nerves and ganglia, the eye cup, the labyrinth
vesicle, an epithelial nasal sac, a muscular tongue, a chond-ri®ed endoskeleton with occipital pillar, an axial skeleton,Meckel's cartilage and the anterior extremity, a gut canal,
liver, heart, lung and mesonephros are di�erentiated. Thefrontal acropodium is pentarch, whereas the back one is stillpaddle-shaped.
The shell coat is completely broken down. Most of the
trophectodermal cells of the trilaminar omphalopleure havefused to form a trophoblastic syncytium (see below).
By 14 dpc (10 mm CRL), the embryo has a two-layered
head amnion lined by high microvilli on the side of the yolksac. This indicates a transport of amniotic ¯uid between yolksac and amnion. The exocoelom, which never covers the whole
embryo, is located ventrolaterally to the embryo, from its navelto the caudal end. The allantois is small.
The trophendoderm of the trilaminar omphalopleure
consists of ¯at, expanded cells united by junctional complexes.It is covered with short microvilli on the side of the yolk saclumen. The yolk sac vessels around the navel are embedded inlarge, bulging cells. The trophendodermal cells of the bilami-
nar omphalopleure are also ¯attened and close to the base ofthe trophectodermal cells (Fig. 5d).
Most cells of the trophectoderm of the trilaminar ompha-
lopleure have fused to form a syncytium (Figs 4b, 5a). Cellborders are rare. The thin basal lamina underlying the singlelayered trophectoderm measures about 150 nm. The syncytium
is covered with microvilli and shows pinocytosis, intracellularvesicles and ®laments (Fig. 3d), indicating active transepithelialtransport of materials. It is rich in rER, polyribosomes andlipid droplets (Fig. 3d) and multivesicular bodies. Nuclei are
located in syncytial knots (2±5 nuclei per knot) (Fig. 4b).Between the syncytial knots are thin lamellae of syncytialepithelium (Fig. 4b). Occasionally, cell borders were found
within the syncytial lamellae. Lipid droplets and accumulationsof glycogen are mainly located in the knots, cell organelles are
rare. The trophectoderm is in close association with the uterineepithelium with an intimate interdigitation of trophoblastic andendometrial microvilli (Fig. 3d). The height of the microvilli
does not exceed 1.6 lm. The invasive trophoblast penetratesthe maternal epithelium at regular intervals, and reaches thematernal endothelium (Figs 3e, 5a). To the side of the invasive
areas, the trophoblast is intimately interconnected with theuterine epithelium by newly formed cellular adhesions(Fig. 5a). The invasive trophoblast displaces the uterineepithelium at either small (Fig. 3e) or expanded areas (Fig. 5a).
Thus, a mixed epithelio-endothelio-chorial yolk sac placenta isformed. At the invasive areas, the maternal endothelium, thetrophoblast (with thin basal lamina underneath), interconnect-
ed cells of the connective tissue of the yolk sac, the extracellularmatrix and the foetal endothelium form the layers, throughwhich the foeto-maternal exchange of material and gases takes
place. Owing to the thinning of foetal layers, the barrierbetween foetal and maternal blood does in some areas notexceed 2.5 lm. In these areas, the uterine epithelium is close to
the cells of the foetal connective tissue near the vitelline vessels(Fig. 4b). Bilaminar parts of the yolk sac are rare, probably dueto fusion of yolk sacs of adjacent embryos and subsequent lossof these areas (Freyer in prep.). In the bilaminar yolk sac wall,
fusion of trophectodermal cells could not be found (Fig. 5b±d).Thinning of the trophoblast and accumulations of lipiddroplets are similar to the trophoblastic syncytium (Fig. 5b±
d). The trophoblast of the bilaminar yolk sac walls were closelyattached to the surface of the maternal epithelium. However,invasive areas could not be found.
The endometrium is strongly folded. The largest folds formchambers for single embryos and their foetal appendices. Aspointed out above, the height of the microvilli is reduced. The
thickness of the endometrial glandular pad is reduced to0.2±0.4 mm. The thickness of the myometrium remainsunchanged at 0.1 mm.At the 15th day after copulation, the propria is highly
invaded by polymorphonuclear granulocytes, macrophagesand lymphocytes (Fig. 3f). Granulocytes are also found in theglandular tubes. This is followed by the parturition of the
placenta.
Discussion
In the present study of the grey short-tailed opossum,M. domestica, the early ontogeny and placentation fromovulation to parturition are described. To con®rm and
complement previously published detailed studies and time-tables of early embryonic development (Harder et al. 1993;Mate et al. 1994; Selwood et al. 1997), the development of the
oocyte and zygote, the embryonic development, as well aschanges of the embryonic membranes and the endometriumduring ontogeny are considered. Special emphasis is placed on
the ultrastructural changes of the placenta, and the develop-ment of structures is interpreted functionally, where possible,by correlating data obtained from LM, EM, histochemistry
and immunohistochemistry. The following discussion is aimedat reconstructing the marsupial morphotype based on thecharacter distribution of own ®ndings and literature data.
Tubal passage
The entrance of the zygote into the uterus at 29 hpc inM. domestica relates well to the times given by Mate et al.
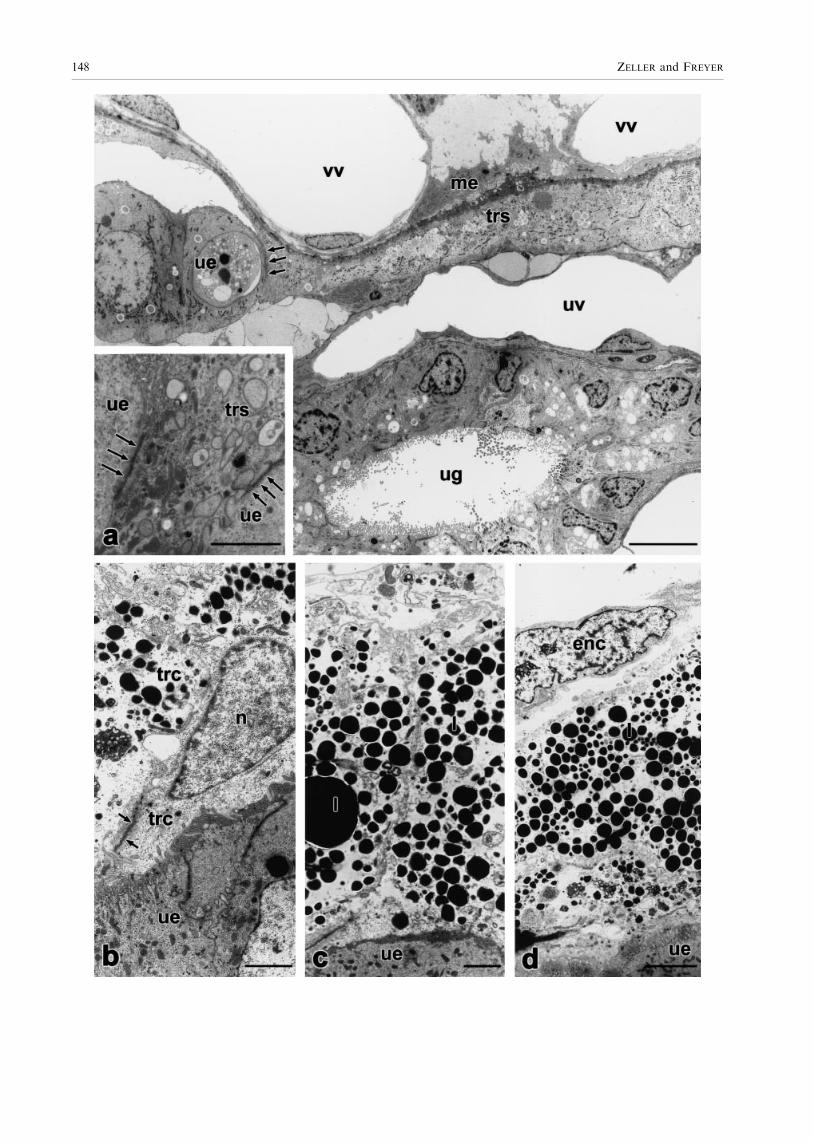
Fig. 3. Monodelphis domestica, (a±e) transmission electron micro-graphs of the yolk-sac placenta from 11 to 14 dpc. (f) photomicrographof post-partum endometrium. (a) 11 dpc. The shell coat (sm) separatesuterine epithelium (ue) and trophoblast, which consists of individualcells (trc) covered with numerous micovilli. l, lipid droplets; me,mesenchyme; scale bar� 10 lm. (b) 11 dpc. The trophoblast consistsof single cells united by junctional complexes (arrows) that areinterspersed with intercellular capillaries (ic). Numerous mitochondria(mt), rER (rer), ribosomes (r) and lipid droplets (l) are present; scalebar� 10 lm. (c) 12 dpc. Remnants of the shell coat (arrows) laybetween trophoblast (tr) and uterine epithelium (ue). The uterineepithelium releases secretory processes (sp), which form histiotrophes(ht); scale bar� 10 lm. (d) 14 dpc, non-invasive area of the trophob-lastic syncytium (trs). Trophoblast and uterine epithelium are denselypacked with microvilli (thin arrows). The trophoblastic syncytium isrich in rER (thick arrows), polyribosomes, ®laments (arrow heads) andlipid droplets. *, mitochondrium; ue, uterine epithelium; scalebar� 10 lm. (e) 14 dpc, invasive area of the trophoblastic syncytium(trs). The trophoblast erodes the uterine epithelium (ue) and reachesuterine vessels (uv). The barrier for foeto-maternal exchange betweenvitelline vessels (vv) containing nucleated erythrocytes (ne) and uterinevessels (emptied by perfusion) is diminished; scale bar� 10 lm. (f) 15dpc, post-partum uterus. The propria (pr) is invaded by leucocytes(thin arrows), i.e. granulocytes, lymphocytes, monocytes and others.Indentation (thick arrows) in the uterine epithelium (ue) remain frominvasive areas of the trophoblastic syncytium. ug, uterine gland; scalebar� 30 lm
Early ontogeny and placentation of the grey short-tailed opossum 145

146 ZELLER and FREYER

(1994) and Selwood et al. (1997) (24 hpc and 30 hpc,respectively). This short time for passing the oviduct (about7 h) is equivalent to that found in Macropus eugenii (Macro-podidae) with 7±13 h (Renfree and Lewis 1996) and Antechinus
stuartii (Dasyuridae), in which the zygote enters the uterus12 h after fertilization (Selwood 1980). In Didelphis virginiana(Didelphidae), the zygote enters the uterus between 15 and
24 h after ovulation (Hartman 1924; Rodger and Bedford1982). Thus, a short tubal passage lasting 1 day at maximumcan be regarded as a character of the marsupial morphotype.
Egg coats
Zona pellucida and corona radiataThe oocyte of M. domestica is surrounded by the zona
pellucida (Phillips and Fadem 1987; Baggott and Moore 1990;Selwood et al. 1997; personal observations). Phillips andFadem (1987) describe the zona pellucida as 2±3 lm thickand ®lamentous. The zona pellucida, which is also found in
monotremes (Hughes 1984) and eutherians, is present in allmarsupial taxa so far examined (cf. Tyndale-Biscoe andRenfree 1987: Didelphidae, Dasyuridae, Peramelidae, Phalan-
geridae, Macropodidae; Ward and Renfree 1988: Acrobatidae)and is, consequently, a plesiomorphic character of the marsu-pial morphotype. As already summarized by Tyndale-Biscoe
and Renfree (1987), the thickness of the zona pellucida rangesfrom 1 lm in Didelphis marsupialis (McCrady 1938) to6.3 � 1.4 lm in M. eugenii (Hughes and Hall 1984). Thewidth of the zona pellucida, like that of the mucoid coat and
shell coat, may change by pressures set up by the extrusion ofthe yolk mass and the ®rst two cleavages as observed inA. stuartii (Selwood and Young 1983). The disappearance of
the zona pellucida by expansion of the blastocyst aswe observed in M. domestica (5 dpc) is also shown forD. virginiana (Hartman 1928) and Acrobates pygmaeus (Ward
and Renfree 1988). Bancroft (1973) also reports the absence ofthe zona pellucida at the blastocyst stage of Petauroides volans.Ovulated eggs of M. domestica (Phillips and Fadem 1987;
Baggott and Moore 1990; this article), D. virginiana (Krause1998), Dasyurus viverrinus (Hill 1910) and Sminthopsis crassi-caudata (Breed and Leigh 1990) lack a corona radiata. Thus,the absence of the corona radiata can be assumed for the
marsupial morphotype.
Mucoid coat
The mucoid coat of M. domestica consists of concentric layers(Phillips and Fadem 1987; this article). Spermatozoa areentrapped in the mucoid coat (Phillips and Fadem 1987;
Baggott and Moore 1990; Selwood and Vandeberg 1992; thisarticle). This is also described for other marsupial taxa(D. viverrinus: Hill 1910; M. eugenii: Renfree and Shaw 1996;
Schoinobates [Petauroides] volans: Bancroft 1973; A. pygmaeus:Ward and Renfree 1988), which suggests that it may preventpolyspermy in addition to the zona reaction (Selwood 1982;
Baggott and Moore 1990). In addition, Hartman (1919)assumes a nutritive function of the mucoid coat inD. virginiana. Furthermore, Selwood (2000) suggests that the
mucoid coat as well as the molecules that are trapped in it aretaken up by the trophoblast and may thus be a nutritionalsource for the embryo. The mucoid coat is also suggested toact as an osmotic stabiliser (Selwood 2000). The secretion of
the mucoid coat by oviductal cells as described for severalother marsupial taxa as well as for monotremes (monotremes:Hughes and Carrick 1978; Trichosurus vulpecula: Hughes 1974;
S. crassicaudata: Roberts et al. 1994), is also observed inM. domestica (Phillips and Fadem 1987; Baggott and Moore1990; this article). Thus, mucoid coat secretion by oviductal
cells is a plesiomorphic character of the marsupial morpho-type. Accordingly, the oviduct consists of ciliated and non-ciliated secretory cells (Phillips and Fadem 1987; Baggott and
Moore 1990; this article). The secretion of mucoid coatprecursors in S. crassicaudata is shown to be restricted to theoviduct (Roberts et al. 1994). Similar to D. virginiana (Hart-man 1928), T. vulpecula (Hughes 1974), D. viverrinus (Hill
1910), Bettongia cuniculus [B. gaimardi] (Kerr 1935) andA. pygmeus (Ward and Renfree 1988), the mucoid coat inM. domestica disappears, with the zona pellucida, when the
blastocyst expands.
Shell coat
Although the marsupial shell coat is suggested to consist ofkeratin, namely ovokeratin (Hughes 1974; Krause 1998), it wasimpossible to verify the presence of keratin by immunohisto-
chemistry in M. domestica. This could be due to the absence ofkeratin, a change of epitopes during condensation or amasking e�ect.The shell coat is secreted within the uterus in all marsupial
taxa so far examined (M. domestica: Baggott and Moore 199077 ,this article; T. vulpecula: Hughes 1974, 1977, 1984; Hughes andHall 1984; S. crassicaudata: Roberts and Breed 1996;
A. stuartii: Selwood 1982; personal comm.; D. virginiana:Hartman 1916; McCrady 1938; Dasyurus, Perameles, Tri-chosurus, Macropus, Petrogale, Phascogale, Acrobates, Phas-
colarctos, Bettongia: Hill 1910; M. eugenii: Tyndale-Biscoe andRenfree 1987). Thus, at least uterine shell coat secretion can beassumed to be a character of the marsupial morphotype.Inconsistent data exist concerning a possible oviductal shell
coat secretion in addition to the uterine shell coat secretion.Whereas Hill (1910) reports that the shell coat is laid down inthe oviduct in Dasyurus, Perameles, Trichosurus, Macropus,
Petrogale, Phascologale [Phascogale], Acrobates, Phascolarctosand Bettongia, the shell coat of M. domestica (this article,Baggott and Moore 1996) and T. vulpecula (Hughes 1974,
1977, 1984; Hughes and Hall 1984) was found to be secreted byuterine cells only. As Hughes (1974) points out, althoughoocytes recovered from the oviduct of T. vulpeculamay possess
a shell coat, shell glands are restricted to the uterus, suggestinga retrograde ¯ow of uterine shell secretions. However,Tyndale-Biscoe and Renfree (1987) argue that such a retro-grade ¯ow is impossible in M. eugenii and other macropodids.
In S. crassicaudata, as in M. domestica, only uterine embryoshave a shell coat, whereas those recovered from the oviducthave no shell coat (Roberts and Breed 1996). Immunohisto-
chemically, shell coat precursors of S. crassicaudata are found
Fig. 4. Monodelphis domestica, transmission electron micrographs ofthe yolk-sac placenta. (a) 11 dpc, cytotrophoblast. The trophoblastconsists of individual cells (trc) connected by tight junctions (arrows).Trophoblast and uterine epithelium (ue) are separated by the shell coat(sm). fbc, foetal blood cell; me, mesenchyme; n, nucleus of trophoblastcell; scale bar� 10 lm (b) 14 dpc, syncytiotrophoblast. The tropho-blast cells fused to form a syncytium (trs). Nuclei (n) are located insyncytial knots (sl). Only thin lamellae of syncytial epithelium separateuterine epithelium and vitelline vessels (vv). The trophoblastic syncy-tium penetrates the uterine epithelium (ue). me, mesenchyme; ne,nucleated erythrocyte; scale bar� 10 lm
Early ontogeny and placentation of the grey short-tailed opossum 147
148 ZELLER and FREYER

to be restricted to the anterior region of the uterus, but also tothe utero-tubal junction and adjacent glands (Roberts et al.1994). In D. virginiana (Hartman 1916; McCrady 1938) andA. stuartii (Selwood 1982; personal comm.), zygotes recovered
from the `lower part of the oviduct' were covered with shellcoat material. In the `lower oviduct' of D. virginiana, there arenumerous mucosal glands (Andersen 1928); the posibility that
they are shell glands cannot be excluded as the function ofthese is unknown (Tyndale-Biscoe and Renfree 1987). It can beassumed, that the part described as `lower oviduct' in
D. virginiana and A. stuartii corresponds to the utero-tubaljunction that was found to secrete shell coat precursors inS. crassicaudata. Although Hill (1910) does not mention which
part of the oviduct secretes the shell coat, tubal shell secretioncan be excluded distal to the isthmus of the oviduct indidelphids (this study; Baggott and Moore 1996), phalangerids(Hughes 1974, 1977, 1984; Hughes and Hall 1984) and
dasyurids (Roberts and Breed 1996). Based on this distribu-tion, shell secretion distal to the isthmus of the oviduct can beexcluded for the marsupial morphotype. Since the shell coat is
secreted by cells of the utero-tubal junction in dasyurids(Roberts et al. 1994) and didelphids (McCrady 1938; Hartman191688 ), this can be assumed for the marsupial morphotype as
well. In conclusion, the shell is secreted by the utero-tubaljunction and the uterus in the marsupial morphotype.
The monotreme and marsupial shell coats have beensuggested to be partly homologous (Hughes 1977). The
monotreme shell coat was found in eggs recovered from the`lower oviduct', but it is also found to be secreted in the uterus(Caldwell 1887). The presence of shell coat glands in the lower
third of the Fallopian tube and within the uterus (Hughes1977) shows that this is due to both tubal and uterine shell coatsecretion. Thus, shell secretion by cells of both uterus and the
proximal oviduct (but not distal to the isthmus) may be aplesiomorphic character of the marsupial morphotype.
Yolk arrangement and cleavage
Yolk arrangementOur observations agree with that of Baggott and Moore (1990)in that the peripheral yolk-poor cytoplasm, which is theformative cytoplasm, surrounds the central yolky cytoplasm
(deutoplasm) in the unfertilized oocyte of M. domestica. Asthis arrangement is also found in unfertilized oocytes ofD. virginiana (Krause 1998), S. crassicaudata (Breed and Leigh
1990) and A. stuartii (Selwood and Young 1983), this can beconcluded to be a character of the marsupial morphotype. InM. domestica, we additionally observed a central portion of
yolk-poor cytoplasm enclosed in the deutoplasm. The yolkbodies of compound yolk and lipid yolk in D. virginiana, asKrause (1998) points out, move toward the peripheral cyto-
plasm. This probably leads to the three-layer arrangement ofcentral and peripheral yolk-free cytoplasm and subperipheralyolky cytoplasm (Hartman 1916; McCrady 1938).
In the fertilized oocyte (zygote), the yolky cytoplasm is polar-oriented in M. domestica (Baggott and Moore 1990; Selwoodand Vandeberg 1992; Breed et al. 1994; Selwood et al. 1997;
Zeller 1999a), D. viverrinus (Hill 1910), S. crassicaudata (Breedand Leigh 1990; Selwood and VandeBerg 1992), A. stuartii(Selwood 1982; Selwood and Sathananthan 1988), Phascolarc-tos cinereus (Caldwell 1887) and T. vulpecula (Frankenberg and
Selwood 1998). Thus, polarity of the zygotic yolk is concludedto be a character of the marsupial morphotype. The polarizedfeatures in the zygote are suggested to identify the future
embryonic and abembryonic poles (Selwood 1994); the side ofthe pronuclei is suggested to be the embryonic hemisphere,whereas the yolky cytoplasm is located in the abembryonic pole
(Frankenberg and Selwood 1998). Although the yolk massbecomes eccentric prior to ovulation in dasyurids (D. viverrinus:Hill 1910; S. crassicaudata: Breed and Leigh 1990) an
P. cinereus (Caldwell 1887), fertilization enhances the priorpolarity (cf. Merry et al. 1995). The zygote of D. virginianashows no yolk polarity (McCrady 1938; Hartman 1916), butlipid droplets can be located more on one pole (Hartman 1916;
McCrady 1938). However, Hartman (1916) and Hill (1918)agree that `an inherent, if nonvisible, polarity probably doesexist in the unsegmented ovum' (Hill 1918; p. 105). Although the
pronuclei are ®rst eccentrically located (Hartman 1916), theylater migrate to the centre (Hartman 1916;McCrady 1938). Thenon-polarized yolk arrangement of the zygote is a derived
character ofD. virginianawith regard to the proposedmarsupialmorphotype.
CleavageIn all marsupials so far examined cleavage starts in the uterus(Selwood 2000), probably due to the shortness of the tubalpassage. This can also be assumed for the marsupial
morphotype.In M. domestica, the ®rst cleavage is meridional and
accompanied by segmentation of most of the yolk and subse-
quent extrusion of smaller yolk vesicles (Baggott and Moore1990; Fromm and Zeller 1997; Selwood et al. 1997). Thesubsequent two-cell stage (46±53 hpc: Selwood et al. 1997) is
characterized by apocrine extrusion of further yolk vesicles(Fromm and Zeller 1997). The second cleavage is asynchro-nous, meridional (Selwood et al. 1997; Fromm and Zellerunpublished) and sometimes latitudinal (Baggott and Moore
1990; Selwood and Vandeberg 1992; Selwood et al. 1997)depending on the amount of yolk. This cleavage is accompan-ied by further yolk extrusion (Baggott and Moore 1990;
Selwood and Vandeberg 1992; Selwood et al. 1997; Fromm andZeller unpublished). Separated yolk and deutoplasmolysis arealso found in D. virginiana (Hartman 1916, 1928), dasyurids
(D. viverrinus: Hill 1910; A. stuartii: Selwood and Young 1983;S. crassicaudata: Selwood 1987), phalangerids (T. vulpecula:Frankenberg and Selwood 1998) and acrobatids (A. pygmaeus:
Ward and Renfree 1988). Consequently, a cleavage includingdeutoplasmolysis is a character of the marsupial morphotype.The absence of any separated yolk in the zygote of macropod-ids (grey kangaroos, M. eugenii: Renfree and Lewis 1996)
and the correlated cleavage without deutoplasmolysis is aderived character of this group and may be a synapomorphyof macropodids. The mode of deutoplasmolysis is the same
in M. domestica (Didelphidae) and dasyurids (Sminthopsis
Fig. 5. Monodelphis domestica, transmission electron micrographs ofthe yolk-sac placenta,14 dpc. (a) The trophoblastic syncytium (trs)largely invades the maternal mucosa and reaches up to uterine vessels(uv). Between invasive trophoblast and uterine epithelium (ue) cellularadhesions are present (arrows, inset).vv, vitelline vessels; me, mesen-chyme; ug, uterine gland; scale bar (inset)� 2 lm, scale bar� 10 lm.(b±d) Bilaminar yolk sac wall. Trophoblastic cells (trc) are clearlyseparated from each other and apically united by junctional complexes(arrows). n, nucleus; l, lipid droplets; ue, uterine epithelium; enc,trophendodermal cell. (b) scale bar� 2.5 lm; (c) scale bar� 2.5 lm;(d) scale bar� 5 lm
Early ontogeny and placentation of the grey short-tailed opossum 149

macroura, S. crassicaudata, A. stuartii). In these taxa, most ofthe deutoplasm (yolk) is separated o� at one pole during the®rst cleavage, lesser quantities are subsequently extruded as
numerous vesicles during the ®rst two cleavages (Selwood 1982;Selwood 1987; Baggott and Moore 1990; Selwood and Vande-Berg 1992; Fromm and Zeller unpublished). En bloc segmen-
tation of deutoplasm (yolk) during the ®rst cleavage is alsodescribed for D. viverrinus (Hill 1910). Based on characterdistribution, this mode is a character of the marsupialmorphotype. Didelphis virginiana (Hartman 1916, 1919;
McCrady 1938) and D. marsupialis (Hill 1918) show nolocalized en bloc segmentation of deutoplasm (yolk). Instead,deutoplasm is separated o� by generalized extrusion of vesicles.
This is a synapomorphy of D. virginiana and D. marsupialiswith regard to the proposed marsupial morphotype.As pointed out by Selwood (198299 ), yolk quantity, yolk
polarization and the mode of deutoplasmolysis may becorrelated. This correlation is based on the assumption thatoocyte dimension is determined by the amount of yolk. Then,
large oocytes such as those of D. viverrinus (Hill 1910),S. crassicaudata and S. macroura (Selwood 1987) with 240,254 and 343 lm diameter, respectively, contain great yolkquantities. Medium-sized oocytes of A. stuartii (Selwood and
Young 1983) and M. domestica (this investigation) with 158and 170 lm contain medium amounts of yolk. Small eggs, asfound in D. virginiana (Hartman 1916) and D. marsupialis (Hill
1918) with 135±165 and 144 lm, respectively, are poor in yolk.Early yolk polarization, i.e. before fertilization, was found inspecies with yolk-rich oocytes, namely Dasyurus and Sminth-
opsis. In those species, the morphotype mode of deutoplasmo-lysis was found. The same applies to species with oocytes with amedium amount of yolk (A. stuartii,M. domestica). Both types
di�er in the time of yolk polarization. In yolk-rich oocytes,polarization ®rst occurs in the ovary oocyte (Hill 1910; Breedand Leigh 1990), whereas in oocytes with a medium amount ofyolk, polarization is only found at the pronuclear stage (this
paper; Selwood 1982, 1992). The yolk-poor oocytes ofD. virginiana and D. marsupialis do not exhibit yolk polarityand the deutoplasmolysis is reduced to a generalized extru-
sion of vesicles. The smallest oocytes, 126 lm in diameter(M. eugenii: Tyndale-Biscoe and Renfree 1987), are foundamong macropodids which lack a separated deutoplasm (yolk).
In M. domestica, the asynchronous, third (Baggott andMoore 1990; Selwood and Vandeberg 1992; Selwood et al.1997; Fromm and Zeller unpublished) and fourth cleavage(Baggott and Moore 1990; Selwood and Vandeberg 1992; this
paper) are latitudinal or meridional, the ®fth cleavage wasfound to be latitudinal (Selwood and Vandeberg 1992). Thesequence of meridional and latitudinal cleavages leads to a
blastocyst at 6 dpc (this paper). At this stage, the blastocoelcontains little yolk (this paper; Fromm and Zeller unpub-lished). We assume that the yolk has been taken up by
blastomeres.In M. domestica, the cells are closely apposed to the zona
pellucida at the two-cell stage (Fromm and Zeller unpublished)
and eight-cell stage (Baggott and Moore 1990; Selwood et al.1997; Fromm and Zeller unpublished), whereas no cell contactis established. Baggott and Moore (1990) report ®rst intercel-lular junctions between blastomeres at the 16-cell stage in
M. domestica. Apposition of blastomeres to the zona pellucidaprior to intercellular association and connection by tightjunctions was also found in A. stuartii (third + fourth
cleavage, Selwood and Young 1983), S. crassicaudata
(second cleavage, Selwood et al. 1997) and T. vulpecula(second cleavage, Frankenberg and Selwood 1998). InM. eugenii, the blastomeres are in close association with one
another during the ®rst cleavages, but no tight junctions arepresent at least until the eight-cell stage is reached (Renfreeand Lewis 1996). This is probably a derived character
of macropodids. This could be correlated with the absence ofseparated yolk in the zygote (see above). The apposition ofblastomeres to the zona pellucida prior to intercellularassociation, and the consequent absence of a morula stage
(Selwood 1996) is a character of the marsupial morphotype,since this is present in ameridelphian didelphids, as well as inaustralidelphian dasyurids and phalangerids.
Placentation
Our observations agree with that of Harder et al. (1993)concerning both the close apposition of the trophectoderm to
the uterus at 13 and 14 dpc and the microvillous surface of theendo- and trophectoderm in M. domestica. Our observedincrease of foldings at 12 dpc is con®rmed by the increase of
the index of epithelial surface area from 1.66 at 9 dpc to 6.9 at13 dpc measured by Harder et al. (1993). We can furthercon®rm, that M. domestica possesses a choriovitelline placenta(Harder et al. 1993; Roberts and Breed 1994a) during the
entire development from implantation to birth. The observeddrop in uterine gland density inM. domestica between 9 and 14dpc (Harder et al. 1993) can be correlated with the decreasing
thickness of the uterine gland pad from 3 mm at 10 dpc,1.5 mm at 12 dpc to 0.2±0.4 mm at 14 dpc. This indicates theshift from histiotrophic to haemotrophic nutrition of the
embryo.
Invasiveness of the placenta
Occurrence and degree of invasion of uterine epithelium by thetrophectoderm of the bilaminar and trilaminar yolk sac variesamong marsupials as summarized in Table 4.
Contrary to earlier reports given by Harder et al. (1993) and
Roberts and Breed (1994a), who report the trophectoderm ofM. domestica to be non-invasive, our electron-microscopicobservations clearly demonstrate that the trophoblastic syncy-
tium of the trilaminar yolk sac penetrates the uterine epithe-lium and is thus invasive (Fig. 3e, Fig. 4b). An invasivetrophectoderm in a didelphid was only known for Philander
opossum (Enders and Enders 1969).The variation between invasive and non-invasive trophec-
toderm among closely related species may be explained byeither interspeci®c variation or a lack of information. The
latter suggestion is supported by the ®ndings of an invasivetrophectoderm in M. domestica, which was formerly describedas possessing a non-invasive trophectoderm. Thus, this char-
acter needs to be re-examined during the entire ontogeny fromimplantation to birth, and by EM.
Formation of an allantochorion and yolk sac placentaThe di�erent conditions of the allantois relative to the chorionfound in di�erent marsupial taxa were often mentioned and
grouped into four placental types (Sharman 1959; Hughes1984; Tyndale-Biscoe and Renfree 1987), which are sum-marized in Table 5.
However, it must be kept in mind that this classi®cation is
an arti®cial grouping of di�erent characters, namely thecontact between allantois and chorion, the size of allantois
150 ZELLER and FREYER

and yolk sac, their vascularization and the function of bothallantois and yolk sac. This is the reason why a characterpolarization based on these types has failed. For a phyloge-netic interpretation of these ®ndings, a polarization of
homologous characters is needed. To date, the insu�cientand inconsistent data concerning the allantochorion inmarsupials can only be polarized by comparison with the
sistergroup (eutherians) of marsupials and the outgroup ofthe Theria (monotremes). Since a fusion between chorion andallantois was found in Dasyuridae, Vombatiformes and
Perameloidea, and this is a character also found in eutherians
(Wooding and Flint 1994) as well as in monotremes (Caldwell1887), this may be a plesiomorphic character of the marsupialmorphotype. Thus, the absence of a chorioallantois indidelphids on one side and australidelphian taxa (acrobatids,
macropodids, phalangerids) on the other side is a convergentreduction. The parsimony of this assumption, however, isweakened by the absence of an allantochorion in the dasyurid
S. crassicaudata (Roberts and Breed 1994a). In a morphotypereconstruction using parsimony (see Cunningham et al.1998), the downpass reconstructed marsupial morphotype
remains ambiguous, but the uppass reconstruction suggests an
Table 4. Distribution of invasive bilaminar and trilaminar yolk sac areas among marsupials
Taxon Species Bilaminar (avascular) yolk sac Trilaminar (vascular) yolk sac
Didelphidae Monodelphis domestica non-invasive1 invasive2
Philander opossum non-invasive3 invasive3
Didelphis virginiana non-invasive1,4,5 non-invasive1,4,5
Didelphis marsupialis non-invasive1,4,5 non-invasive1,4,5
Dasyuridae Dasyurus viverrinus invasive6 non-invasive6
Sminthopsis crassicaudata invasive7,8 non-invasive8
Peramelidae Isoodon obesulus non-invasive9,10,11 non-invasive9,10,11
Perameles gunnii non-invasive11 non-invasive11
Perameles nasuta non-invasive7,12 non-invasive7,12
Echymipera rufescens non-invasive7 non-invasive7
Echymipera kalubu non-invasive13 non-invasive13
Phalangeridae Trichosurus vulpecula non-invasive7,12,15 non-invasive7,12,15
Pseudocheiridae Pseudocheirus peregrinus non-invasive12,16 non-invasive12,15,16
Petauroides volans non-invasive14 invasive14
Acrobatidae Distoechurus pennatus invasive17 non-invasive17
Macropodoidea Potorous tridactylus non-invasive12,15 non-invasive12,15
Macropus robustus non-invasive7 non-invasive7
Macropus giganteus non-invasive18 non-invasive18
Macropus rufogrisea non-invasive15 non-invasive15
Setonix brachyurus non-invasive15,19 non-invasive15,19
Macropus eugenii non-invasive20 non-invasive20
Bettongia cuniculus non-invasive21 invasive21
Vombatiformes Phascolarctos cinereus invasive7,12,22 non-invasive7,22
Vombatus ursinus invasive7 non-invasive7
Source: 1Harder et al. (1993); 2 this paper; 3 Enders and Enders (1969); 4Hartman (1923); 5McCrady (1938); 6Hill (1900a); 7Hughes (1974);8Roberts and Breed (1994a); 9Hill (1895); 10Hill (1898); 11 Flynn (1923); 12Hughes and McNally (1968); 13Hughes et al. (1990); 14 Bancroft(1973); 15 Sharman (1961); 16Hughes et al. (1965); 17Hughes et al. (1987); 18 Chapman (1882); 19 Sharman (1959); 20 Tyndale-Biscoe and Renfree(1987); 21 Flynn (1930); 22 Caldwell (1884).
Table 5. Types of marsupial placentation according to Tyndale-Biscoe and Renfree (1987)
Type Description Taxon
Type I Allantois remains small and does not fuse with the chorion, only the well vascularizedyolk sac contributes to the placenta
Didelphidae1±4, Phalangeridae1,
Macropodidae1, Acrobatidae5
Type II Allantois reaches the chorion and seems partly to fuse with the chorion, but allantoicvessels degenerate, later in ontogeny the allantois retreats and degenerates;assumption: the allantochorion does not form a placenta
Dasyurus viverrinus6
Type III Allantois fuses with chorion, neither mesodermal nor ectodermal chorionic villi at theside of the allantochorion, neither invasion of uterine epithelium nor fusion of theuterine epithelium with the allantochorion, no replacement of the yolk sac placenta;assumption: yolk sac is the main organ of nutritive absorption, allantochorionfunctions principally as a respiratory organ
Phascolarctos cinereus1,7±10
Vombatus ursinus10
Type IV Allantois fuses with chorion, no mesodermal villi at the side of the allantochorion,ectodermal villi at the side of the allantochorion, fusion of uterine epithelium andtrophoblast of the allantochorion, no replacement of yolk sac placenta; assumption:allantochorion forms a placenta additionally to the yolk sac placenta
Peramelidae1,11±16
Source: 1 Tyndale-Biscoe and Renfree (1987); 2 Harder et al. (1993); 3 Roberts and Breed (1994a); 4 this paper; 5 Hughes et al. (1987); 6 Hill(1900a); 7 Caldwell (1884); 8 Semon (1894); 9 Hughes (1974); 10 Hughes (1984); 11 Hill (1895); 12 Hill (1898); 13 Hill (1900b); 14 Flynn (1923);15 Padykula and Taylor (1976); 16 Padykula and Taylor (1982).
Early ontogeny and placentation of the grey short-tailed opossum 151

allantochorion to be present in the marsupial morphotype(Fig. 6). The absence of an allantochorion in phalangeroids
(phalangerids, acrobatids) and macropodids may thus besynapomorphic. Since the allantochorion is vascularized inmonotremes (Caldwell 1887), placentals (Wooding and Flint
1994), peramelids and vombatiforms, it is to date parsimo-nious to assume a reduction of this vascularization inD. viverrinus derived from a vascularized allantochorion inthe marsupial morphotype. This preliminary polarization,
however, demands a re-examination of recent ®ndings, theexamination of the ontogeny from implantation until birth infurther species and the re-evaluation of marsupial systematics,
which are in progress (Freyer, Zeller and Renfree in prep.).We are aware that our hypothesis appears to contradict withthe interpretation of Luckett (1977; p. 490), who assumes a
`retardation in expansion of the exocoelom and allantois' inthe marsupial morphotype.Although the chorioallantoic placenta in marsupials never
forms mesodermal villi and never replaces the yolk sacplacenta as that of eutherians (Luckett 1977), it can beassumed that the allantochorion in vombatiforms and per-amelids serves at least respiratory function (Luckett 1977;
Semon 1894; Hughes 1984; Tyndale-Biscoe and Renfree 1987).A respiratory function of the vascularized allantochorion inthe marsupial stem species seems likely. However, it is
unknown whether this allantochorion formed a placentawhich also allowed the exchange of material.All marsupials so far examined possess a yolk sac placenta
for most of the time after implantation (cf. Tyndale-Biscoe andRenfree 1987). Thus, a yolk sac placenta can be assumed forpart of the ontogeny in the marsupial morphotype (Luckett1977), irrespective of the formation of an allantois placenta at
near-term stages.
Formation of a trophoblastic syncytiumZeller (1999a) provides the ®rst evidence of a trophoblastic
syncytium ever observed in a marsupial by EM. Here, we givea detailed description of the ultrastructure of this syncytium bypresenting original electron micrographs. We assume that the
aggregations of nuclei in plasma accumulations are syncytialknots rather than plasmodia, i.e. nuclear division. Thisassumption is based on the constant nuclear sizes from 11 to14 dpc (Fig. 4a versus 4b), the absence of evidence on nuclear
divisions at any stage, and the great extension of the syncytialparts relative to former cell sizes and regular cell arrangementswith tight junctions between single cells (Fig. 4a versus 4b).
Hill (1900) reported a trophoblastic syncytium in D. viverrinusby LM. As the cell boundaries can hardly be seen by LM, thisdescription needed to be veri®ed by EM. However, the electron
microscopic ®ndings in M. domestica presented herein make asyncytium in D. viverrinus highly probable. There is no reportof fusions between cells of the trophoblast or extended parts of
it in any other marsupial species. If further examinationscon®rmed a trophoblastic syncytium to be present inD. viverrinus, a trophoblastic syncytium could be assumedfor the marsupial morphotype.
The description of trophoblastic giant cells inM.domestica byRoberts and Breed (1994a) may represent syncytial knots of thetrophoblastic syncytium not identi®ed by the authors. The
question whether the trophoblastic giant cells as observed inother marsupial species, e.g. S. crassicaudata (Roberts andBreed 1994a), are syncytical knots can only be answered
through further examination.
Formation of a feto-maternal syncytiumUntil now, there has been no evidence for a foeto-maternal
syncytium in M. domestica. A foeto-maternal syncytium is
Fig. 6. Reconstruction of theallantochorion in the marsupialmorphotype using parsimony asdescribed by Cunningham et al.(1998)
152 ZELLER and FREYER

known for the bilaminar yolk sac of Dasyurus sp. andP. cinereus (Flynn 1923). In S. crassicaudata, cells of thetrilaminar omphalopleure partly fuse with uterine epithelial
cells (Roberts and Breed 1994b). In peramelids (Isoodonmacrourus), the trophoblast of the chorioallantois `mayhave fused with maternal homokaryons to create placental
Table 6. Characters of the marsupial morphotype (0) and apomorphic characters (1)
Characterno.
Character state 0(morphotype) Taxon
Character state 1(apomorphy) Taxon
Character notapplicable [?] Taxon
1 Central yolky cytoplasmand peripheral yolk-poor cytoplasm in theunfertilized egg
Didelphidae;Dasyuridae
No separatedyolk in oocyte
Macropodidae
2 Zona pellucida Didelphidae;Dasyuridae;Macropodidae;Peramelidae;Phalangeridae;Acrobatidae
3 Absence of a coronaradiata
Didelphidae;Dasyuridae
4 Cell apposition to zonapellucida prior tointercellular connection
Didelphidae;Dasyuridae;Phalangeridae
Intercellular associationof blastomeres atearly cleavage stages
Macropus eugenii
5 Mucoid coat Didelphidae;Dasyuridae;Macropodiae;Pseudocheiridae;Acrobatidae;Phalangeridae
6 Spermatozoa trapped inthe mucoid coat
Didelphidae;Dasyuridae;Acrobatidae;Pseudocheiridae
7 Oviductal mucoidsecretion
Didelphidae;Dasyuridae;Phalangeridae
8 Shell coat Didelphidae;Dasyuridae;Phalangeridae;Peramelidae;Macropodidae;Acrobatidae
9a Uterine shell coatsecretion
Didelphidae;Dasyuridae;Phalangeridae;Peramelidae;Macropodidae;Acrobatidae
9b No oviductal shell coatsecretion distal to theisthmus
Didelphidae;Dasyuridae;Phalangeridae
10 Short tubal passage(3±7 h)
Didelphidae;Dasyuridae;Macropodidae;
11 Separated yolk in zygote Didelphidae;Dasyuridae;Acrobatidae;
No separated yolk inzygote
Macropodidae
12 Yolk polarity in thezygote
Didelphidae;Dasyuridae;Phascolarctidae;Phalangeridae
Non-polarized yolkarrangement in thezygote
Didelphisvirginiana
No separatedyolk in zygote
Macropodidae
13 No morula stage Didelphidae;Dasyuridae;Macropodidae
14 Localized segmentation ofdeutoplasm at the ®rstcleavage and subsequentextrusion of yolk vesiclesduring the ®rst twocleavage stages
Monodelphisdomestica;
Dasyuridae;Acrobatidae
No localizedsegmentation ofdeutoplasm;generalized extrusionof yolk vesicles
Didelphisvirginiana;Didelphismarsupialis
No separatedyolk in zygote
Macropodidae
Early ontogeny and placentation of the grey short-tailed opossum 153

heterokaryons' (Padykula and Taylor 1982; p.95). This database is insu�cient for a morphotype reconstruction.
Parturition
The timing of parturition in M. domestica determined to be 15dpc in this study equals that of 15.2 � 0.3 (n� 16) found byHarder et al. (1993) and almost matches that of 13.5±14 days
after fertilization found by Moore (1992).At the time of parturition, the propria of M. domestica was
found to be highly invadedbypolymorphonuclear granulocytes,
macrophages and lymphocytes (Fig. 3f). Granulocytes are alsofound in the glandular tubes. In a single specimen of S. crassi-caudata, which had given birth 3±7 h before the time of death,`the nuclei of the trophoblast giant cells had become pyknotic
and there was a massive in®ltration of granular leucocytes andlymphocytes into the endometrial stroma' (Roberts and Breed1994a10 , p.109). In late pregnancy (CRL 2.9±3.5 mm), scattered
neutrophils, lymphocytes and macrophages were found in theendometrial stroma of S. crassicaudata, `particularly below theinvasive trophoblast giant cells' (Roberts and Breed 1994a;
p. 108). Although Cruz and Selwood (1993) report thatlymphocytes are clustered in a dense mat in the uterine stromaadjacent to the basal lamina of the endometrial epithelium inboth pregnant and nonpregnant specimens of A. stuartii, it is
striking that the number of lymphocytes was found to besigni®cantly greater in pregnant than non-pregnant animals onthe ®rst and 15th day after ovulation. In addition, the lympho-
cyte density decreases in non-pregnant animals, but increases inpregnant animals from the fourth to the eighth day after
ovulation.An in®ltration of the placenta by polymorphonuclearleucocytes is also reported for two specimens of Peramelesobesula [Isoodon obesulus] (Flynn 1923). Whether this in¯am-
matory reactions is in any way related to birth is still unknown.
Conclusions
This study is a contribution to the reconstruction of themorphotype of recent marsupials by providing current ®nd-ings, a character polarization and a phylogenetic interpret-
ation. This is based on marsupial systematics as proposed byLuckett (1994).
Our data support most of the previous examinations on
the early ontogeny and placentation of M. domestica. Wedescribe both the ultrastructure of a trophoblastic syncytiumand an invasive trophoblast for the ®rst time in M.
domestica by presenting original electron micrographs.Table 6 summarizes characters that are concluded to bepresent in the marsupial morphotype and those that are
derived, based on the character distributions within marsu-pials (Fig. 7).
To date, polarization of most characters of the marsupialplacenta is impossible due to insu�cient and incongruent data.
Obviously, a yolk sac placenta is established at early stages ofplacentation in the marsupial morphotype. In a preliminaryparsimony analysis, a vascularized allantochorion can be
assumed for the marsupial morphotype. The contribution ofthis allantochorion to placentation in the marsupial morpho-type has not yet been established.
Fig. 7. Reconstruction of the earlyontogeny in the marsupial mor-photype using parsimony as des-cribed by Cunningham et al.(1998). For character coding seeTable 6. Numbers above rectangleare character numbers; the num-bers below rectangle are the char-acter states
154 ZELLER and FREYER

The early ontogeny and placentation in the stem species ofrecent marsupials may have occurred as follows: Whenreleased from the oviduct, the oocyte was covered by the zona
pellucida and a corona radiata was lacking. The yolkycytoplasm was centrally located and surrounded by theformative, yolk-poor cytoplasm. Fertilization occurred in the
oviduct. Prior to or shortly after fertilization, the yolkycytoplasm migrated to one pole (abembryonic pole) oppositethe formative cytoplasm (embryonic pole). During the passagedown the oviduct, the oocyte became surrounded by the
mucoid coat, which was secreted by non-ciliated oviductalcells. The remaining spermatozoa were entrapped into themucoid coat, preventing polyspermy. The mucoid coat also
prevented direct contact between the oocyte and the tubalepithelium and may also have functioned as an osmoticstabilizer. With the assistance of ciliated oviductal cells, the
conceptus reached the uterus within 1 day after ovulation.There was no cleavage during the tubal passage. Cells of boththe utero-tubal junction and uterus secreted shell coat precur-
sors, which polymerized into the shell coat on the mucoid coat.The shell coat may have consisted of keratin which di�ered inits chemical characteristics to known human keratins. Themeshes of the shell coat could be passed by transferrin,
nutrients and gases, but it also prevented direct contact of theconceptus with the uterine epithelium. The cleavage of thezygotes that contained moderate to large amounts of yolk
started in the uterus. Most yolk was separated o� at one poleduring the ®rst cleavage. Lesser quantities were subsequentlyextruded during the ®rst two cleavages. A sequence of
equatorial and latitudinal cleavages lead to the unilaminarblastocyst. Blastomere±zona adhesions preceded blastomere±blastomere connections. Thus, a morula stage was omitted.
Subsequently, the yolk was reabsorbed by the blastomeresand the blastocyst expanded. The zona pellucida and themucoid coat disappeared during this process. The uptake ofmucoid coat and entrapped molecules may have served as a
nutritional source for the embryo. Before implantation theembryo was nourished by secretions (histiotrophe) of theuterine glands. Within the last third of pregnancy, the shell
coat broke down and the trophectoderm apposed to theuterine epithelium. In certain areas the trophoblast may havepenetrated the uterine epithelium. Early at implantation, the
placenta was solely formed by the yolk sac. Histiotrophicnutrition may have been shifted toward an exchange ofmaterial and gases between foetal and maternal vessels(haemotrophic nutrition). Later in ontogeny, a vascularized
allantochorion was formed, but did not replace the yolk sacplacenta. It is unknown, whether this allantochorion servedmaterial transfer beside its respiratory function.
We regard the reconstruction of the marsupial morphotypeas a ®rst step toward an evolutionary (holistic) scenario thatexplains mammalian evolution arround the Cretaceous±
Tertiary boundary.
AcknowledgementsWe thank Dr H.-J. Kuhn for his support in maintaining the breedingcolony of Monodelphis domestica at the Institute of Anatomy,University of GoÈ ttingen, until 1994 and for providing researchfacilities. We are indebted to the University of Mainz, Institute ofZoology, for providing research facilities during Dr Zeller's stay therefrom 1994 to 1996. We are also grateful to the Humboldt-UniversityBerlin for the substantial support in building new research facilitiesincluding animal housing at the Museum fuÈ r Naturkunde since 1996.
We thank Mrs. J. Zeller for technical assistance. We are indebted to DrM. Ade, Dr S. Frahnert and Dr A. Mess for critical comments andfruitful discussions. We especially thank Dipl. Biol. P. Giere for hisassistance with linguistic corrections. We thank Dr P. Luckett, Dr L.Selwood, Dr M. B. Renfree and Dr G. Shaw for critical comments andfruitful discussions. We especially thank Dr M. B. Renfree for her kindpermission to use her research facilities during Dr Zeller' s stay therefrom 3±16 October 2000.
ZusammenfassungFruÈ he Ontogenie und Plazentation der grauen Hausspitzmausbeutelratte,Monodelphis domestica (Didelphidae: Marsupialia): Ein Beitrag zurRekonstruktion des Grundplans der Marsupialia
Die vorliegende Arbeit beschreibt die fruÈ he Ontogenese und Plazen-tation von 3 Stunden nach der Kopulation bis zur Geburt derBeutelratte Monodelphis domestica. Es wird die Ultrastruktur undHistochemie der EihaÈ ute, der Fetalmembranen, des Oviductes und desUterus beschrieben. Erstmalig wird die Ultrastruktur eines tropho-blastischen Syncytiums bei einem Beuteltier beschrieben. Weiterhinwird ein invasives Trophektoderm und eine EntzuÈ ndungsreaktion zumZeitpunkt der Geburt bei M. domestica festgestellt. Die Befunde dieserStudie und Literaturdaten werden verglichen und in eine Grun-dplanrekonstruktion integriert. Merkmale, die mindestens zwischenVertretern der Didelphidae und Dasyuridae uÈ bereinstimmen, werdenbasierend auf dem phylogenetischen System der Marsupialia nachLuckett, J. Mammal. Evol. 2, 255±283, 1994, fuÈ r den Grundplan derMarsupialia angenommen. Diese Merkmale sind zentral separierterDotter und peripheres dotterarmes Zytoplasma in der unbefruchtetenEizelle, das Vorhandensein von Zona pellucida, Mucoidschicht undSchalenhaut, das Fehlen einer Corona radiata, die Mucoidsekretiondurch den Oviduct, die Schalensekretion durch den Uterus und nichtdistal der Isthmusregion des Oviductes, eine kurze Tubenwanderung(maximal einen Tag), die Anlagerung der Blastomeren an die Zonapellucida vor der interzellulaÈ ren Verbindung, das Fehlen einesMorulastadiums, die DotterpolaritaÈ t in der Zygote, die lokale Dotte-rabtrennung bei der ersten Teilung und die anschlieûende Dotterex-trusion waÈ hrend der ersten beiden Teilungen. In Bezug auf denGrundplan der Marsupialia ist die unpolare Dotterverteilung in derZygote ein abgeleitetes Merkmal von Didelphis virginiana. Didelphisvirginiana und Didelphis marsupialis teilen als Synapomorphie dieReduktion der Deutoplasmolyse auf eine generelle Vesikelextrusion.Das Fehlen separierten Dotters in der Oocyte und die resultierendeFurchung ohne Dotterextrusion [Renfree and Lewis, Reprod. Fert.Dev. 8, 725±742, 1996] ist eine Apomorphie der Macropodidae.Hiermit haÈ ngt moÈ glicherweise die fruÈ he Zusammenlagerung derBlastomeren zusammen [Renfree and Lewis, Reprod. Fert. Dev. 8,725±742, 1996]. Ein vaskularisiertes Allantochorion und eine Dotter-sackplazenta koÈ nnen fuÈ r einen Teil der Ontogenese im Grundplan derMarsupialia angenommen werden. Ob das Allantochorion neben derRespiration auch dem Sto�austausch diente ist unklar. Die Lesrich-tung fuÈ r den Modus der Plazentation und der Geburt bedarf weitererUntersuchungen.
ReferencesAmoroso, E. C., 1952: Placentation. In: Parkes, A. S. (ed.), Marshall'sPhysiology of Reproduction, 3rd edn. Vol. 2, London, UK:Longmans Green, pp. 127±311.
Andersen, D. H., 1928: Comparative anatomy of the uterotubaljunction. Histology and physiology in the sow. Am. J. Anat. 42,255±305.
Archibald, J. D., 1982: A Study of Mammalia and Geologyacross the Cretaceous±Tertiary Boundary in Gar®eld County,Montana. Geological Sciences 122. University of California Press,Berkeley1111 .
Austin, C. R., 1961: The Mammalian Egg. Oxford, UK: Blackwell.Baggott, L. M.; Moore, H. D. M., 1990: Early embryonic developmentof the grey short-tailed opossumMonodelphis domestica, in vivo andin vitro. J. Zool. Lond. 222, 623±639.
Early ontogeny and placentation of the grey short-tailed opossum 155

Baggott, L. M.; Davis-Butler, S.; Moore, H. D. M., 1987: Character-ization of oestrus and timed collection of oocytes in the greyshort-tailed opossum, Monodelphis domestica. J. Reprod. Fert. 79,105±114.
Bancroft, J., 1973: Embryology of Schoinobates volans (Kerr) (Mar-supialia: Petauridae). Aust. J. Zool. 21, 33±52.
Bedford, J. M., 1996: What marsupial gametes disclose about gametefunction in eutherian mammals. Reprod Fertil Dev 8, 569±580.
Breed, W. G.; Leigh, C. M., 1990: Morphological changes in theoocyte and its surrounding vestments during in vivo fertilization inthe dasyurid marsupial Sminthopsis crassicaudata. J. Morphol. 204,177±196.
Breed, W. G.; Simerly, C.; Navara, C. S.; VandeBerg, J. L.; Schatten,G., 1994: Microtubule con®gurations in oocytes, zygotes, and earlyembryos of a marsupial, Monodelphis domestica. Dev. Biol. 164,230±240.
Caldwell, M. A., 1887: The embryology of Monotremata andMarsupialia I. Phil. Trans. Roy. Soc. London 178, 463±486.
Caroll, R. L., 1997: Early Cenozoic mammals. In: Caroll, R. L. (ed.),Patterns and Processes of Vertebrate Evolution. Cambridge, UK:Cambridge University Press, pp. 352±358.
Case, J. A.; Woodburne, M. O., 1986: South American marsupials: asuccessful crossing of the Cretaceous±Tertiary Boundary. Palaios 1,413±416.
Chapman, H. C., 1882: On a foetal kangaroo and its membranes. Proc.Acad. Nat. Sci. Philadelphia 1881, 468±471.
Cruz, Y. P.; Pedersen, R. A., 1991: Origin of embryonic andextraembryonic cell lineages in mammalian embryos. In: Pedersen,R. A.; McLaren, A.; First, N. L. (eds), Animal Applications ofResearch in Mammalian Development. Cold Spring Harbor NY,USA: Cold Spring Harbor Laboratory Press, pp. 147±204.
Cruz, Y. P.; Selwood, L., 1993: Uterine histology of the dasyuridmarsupial, Antechinus stuartii: relationship with di�erentiation ofthe embryo. J. Reprod. Fertil. 99, 237±242.
Cunningham, C. W.; Omland, K. E.; Oakley, T. H., 1998: Recon-structing ancestral character states: a critical reappraisal. TREE 13,361±366.
Enders, A. C.; Enders, R. K., 1969: The placenta of the four-eyedopossum (Philander opossum). Anat. Rec. 165, 431±450.
Fadem, B. H., 1985: Evidence for the activation of female reproduc-tion by males in a marsupial, gray short-tailed opossum (Monodel-phis domestica). Biol. Reprod. 33, 112±116.
Fadem, B. H.; Rayve, R. S., 1985: Characteristics of the oestrous cycleand in¯uence of social factors in grey short-tailed opossums(Monodelphis domestica). J. Reprod. Fertil. 73, 337±342.1313
Fadem, B. H.; Trupin, G. L.; Maliniak, E.; VandeBerg, J. L.; Hayssen,V., 1982: Care and breeding of the gray short-tailed opossum(Monodelphis domestica). Lab. Anim. Sci. 32, 405±409.
Flynn, T. T., 1923: The yolk-sac and allantoic placenta in Perameles.Q. J. Micr. Sci. 67, 123±182.
Flynn, T. T., 1930: The uterine cycle of pregnancy and pseudopre-gnancy as it is in the diprotodont marsupial Bettongia cuniculus withnotes on other reproductive phenomena in this marsupial. Proc.Linn. Soc. NSWales 55, 506±531.
Frankenberg, S.; Selwood, L., 1998: An ultrastructural study of therole of an extracellular matrix during normal cleavage in amarsupial, the brushtail possum. Mol Reprod Development 50,420±433.
Fromm, K.; Zeller, U., 1997: FruÈ he Embryonalentwicklung vonMonodelphis domestica (Didelphidae: Marsupialia). Verh. Dt. Zool.Ges. 90, 74.
Harder, J. D.; Stonerook, M. J.; Pondy, J., 1993: Gestation andplacentation in two new world opossums: Didelphis virginiana andMonodelphis domestica. J. Exp. Zool. 266, 463±479.
Hartman, C. G., 1916: Studies in the development of the opossum,Didelphis virginiana L. I. History of the early cleavage, II. Formationof the blastocyst. J. Morphol. 27, 1±84.
Hartman, C. G., 1919: Studies in the development of the opossumDidelphys virginiana. III. Description of new material on matur-ation, cleavage and endoderm formation. IV. The bilaminarblastocyst. J. Morphol. 32, 1±139.
Hartman, C. G., 1923: Breeding habits, development and birth of theopossum. Smithson. Inst. Annu. Report 1921, 347±363.
Hartman, C. G., 1924: Observations on the motility of the opossumgenital tract and the vaginal plug. Anat. Rec. 27, 293±303.
Hartman, C. G., 1928: The breeding season of the opossum (Didelphisvirginiana) and the rate of intrauterine and postnatal development.J. Morphol. Physiol. 46, 143±215.
Heap, R. B.; Flint, A. P. F., 1984: Pregnancy. In: Austin, C. R.; Short,R. V. (eds), Reproduction in Mammals, Book 3, Hormonal Controlof Reproduction. Cambridge, UK: Cambridge University Press,pp. 153±194.
Hennig, W., 1950: GrundzuÈ ge einer Theorie der PhylogenetischenSystematik. Berlin, Germany: Deutscher Zentralverlag.
Herrler, A.; Pell, J. M.; Beier, H. M.; Allen, W. R.; Stewart, F., 1997:Detection of insulin-like growth factor binding protein 3 (IGFBP3)in the coats of preimplantation rabbit and horse embryos. J. Reprod.Fert. Abstr. Ser. 19, 11±12.
Hill, J. P., 1895: Preliminary note on the occurrence of a placentalconnection in Perameles obesula and on the foetal membranes ofcertain macropodids. Proc. Linn. Soc. NSWales 10, 578±581.
Hill, J. P., 1898: The placentation of Perameles (contributions to theembryology of the marsupialia-I.). Q. J. Micr. Sci. 40, 385±446.
Hill, J. P., 1900a:On the foetalmembranes, placentation and parturitionof the native cat (Dasyurus viverrinus). Anat. Anz. 18, 364±373.
Hill, J. P., 1900b: Contributions to the embryology of the Marsupialia.2. On a further stage of placentation of Perameles. 3. On the foetalmembranes of Macropus parma. Q. J. Micr. Sci. 43, 1±22.1414
Hill, J. P., 1910: Contributions to the embryology of the Marsupialia.4. The early development of the Marsupialia with special referenceto the native cat (Dasyurus viverrinus). Q. J. Micr. Sci. 56, 1±134.
Hill, J. P., 1918: Some observations on the early development ofDidelphys aurita. Q. J. Micr. Sci. 63, 91±139.
Hinds, L. A.; Reader, M.; Wernberg-Moller, S.; Saunders, N. R., 1992:Hormonal evidence for induced ovulation inMonodelphis domestica.J. Reprod. Fertil. 95, 303±312.
Hughes, R. L., 1974: Morphological studies on implantation inmarsupials. J. Reprod. Fertil. 39, 173±186.
Hughes, R. L., 1977: Egg membranes and ovarian function duringpregnancy in monotremes and marsupials. In: Calaby, J. H.;Tyndale-Biscoe, C. H. (eds), Reproduction and Evolution. Can-berra, Australia: Australian Academy of Science, pp. 281±291.
Hughes, R. L., 1984: Structural adaptations of the eggs and the fetalmembranes of monotremes and marsupials for respiration andmetabolic exchange. In: Seymour, R. S. (ed.), Respiration andMetabolism of Embryonic Vertebrates. Doordrecht; Boston; Lan-caster: Dr W. Junk Publishers, pp. 389±421.
Hughes, R. L.; Carrick, F. N., 1978: Reproduction in femalemonotremes. In: Augee, M. L. (ed.), Monotreme Biology, Vol. 20,Aust. Zool. Sydney, NSW, Australia: Royal Zoological Society ofNSW, pp. 233±253.
Hughes, R. L.; Hall, L. S., 1984: Embryonic development in thecommon brushtail possum Trichosurus vulpecula. In: Smith, A. P.;Hume, I. D. (eds), Possums and Gliders. Sydney, NSW, Australia:Australian Mammal Society, pp. 197±212.
Hughes, R. L.; McNally, J., 1968: Marsupial foetal membranes withparticular reference to placentation. J. Anat. 103, 211.
Hughes, R. L.; Hall, L. S.; Aplin, K. P.; Archer, M., 1987:Organogenesis and fetal membranes in the new guinea pen-tailedpossum, Distoechurus pennatus (Acrobatidae: Marsupialia).In: Archer, M. (ed.), Possums and Opossums: Studies in Evolution.Sydney, NSW, Australia: Surrey Beattey and Sons and the RoyalZoological Society of New South Wales, pp. 715±724.
Hughes, R. L.; Hall, L. S.; Archer, M.; Aplin, K., 1990: Observationson placentation and development in Echymipera kalubu. In: Seebeck,J. H.; Brown, P. R.; Wallis, R. L.; Kemper, C. M. (eds), Bandicootsand Bilbies. Sydney, NSW, Australia: Surrey Beatty and Sons,pp. 259±270.
Hughes, R. L.; Thomson, J. A.; Owen, W. H., 1965: Reproduction innatural populations of the australian ringtail possum, Pseudocheirusperegrinus (Marsupialia: Phalangeridae), in Victoria. Aust. J. Zool.13, 383±406.
156 ZELLER and FREYER

Kerr, T., 1935: Blastocysts of Potorous tridactylus and Bettongiacuniculus. Q. J. Micr. Sci. 77, 305±315.
KoÈ nigsmann, E., 1975: Termini der phylogenetischen Systematik.Biologische Rundschau 13, 99±115.
Krause, W. J., 1998: A review of histogenesis/ organogenesis in thedeveloping North American opossum (Didelphis virginiana). Adv.Anat. Embryol. Cell. Biol. 143, 1±120.
Lochbrunner, A., 1956: BeitraÈ ge zur Biologie des Syrischen Goldham-sters Mesocricetus auratus (Nehring). Zool. Jb., Abt. All. Zool. U.Physiol. D. Tiere 66 (2/3), 389±428.
Luckett, P., 1994: Suprafamilial relationships within Marsupialia:resolution and discordance from multidisciplinary data. J. Mammal.Evol. 2, 255±283.
Luckett, P., 1977: Ontogeny of amniote fetal membranes and theirapplication to phylogeny. In: Hecht, M. K.; Goody, P. C.;Hecht, B. M. (eds), Major Patterns in Vertebrate Evolution.New York; London: Plenum Publishing Corporation, pp.439±516.
Mate, K. E.; Robinson, E. S.; Vandeberg, J. L.; Pedersen, R. A.,1994: Timetable of in vivo embryonic development in the greyshort-tailed opossum (Monodelphis domestica). Mol. Reprod. Dev.39, 365±374.
McCrady, E., 1938: The embryology of the opossum. Am. Anat. Mem.16, 1±233.
Merry, N. E.; Johnson, M. H.; Gehring, C. A.; Selwood, L., 1995:Cytoskeletal organization in the oocyte, zygote, and early cleavingembryo of the stripe-faced dunnart (Sminthopsis macroura). Mol.Repr. Dev. 41, 212±224.
Moore, H. D. M., 1992: Reproduction in the gray short-tailedopossum, Monodelphis domestica. In: Hamlett, W. C. (ed.), Repro-ductive Biology of South American Vertebrates, Vol. I-XVII. NewYork; Berlin etc: Springer Verlag, pp. 229±241.
Morris, G., 1975: Placental evolution and embryonic nutrition. In:Steven, D. H. (ed.), Comparative Placentation: Essays in Structureand Function. New York: Academic Press,1616 pp. 87±107.
Mossman, H. W., 1987: Vertebrate fetal membranes. New Brunswick,NJ, USA: Rutgers University Press.
Novacek, M. J.; Wyss, A. R.; McKenna, M. C., 1988: The majorgroups of eutherian mammals. In: Benton, M. J. (ed.), ThePhylogeny and Classi®cation of the Tetrapods, Vol. 2: Mammals.Systematic Association Special Volume No. 35B. Oxford, UK:Clarendon Press, pp. 31±71.
Nowak, R. M., 1999: Walker's Mammals of the World, Vol. I-II.Baltimore; London: The John Hopkins University Press, pp. 1278±1282.
Padykula, H. A.; Taylor, J. M., 1976: Ultrastructural evidence for lossof the trophoblastic layer in the chorioallantoic placenta ofAustralian bandicoots (Marsupialia: Peramelidae). Anat. Rec. 186,357±386.
Padykula, H. A.; Taylor, J. M., 1982: Marsupial placentation and itsevolutionary signi®cance. J. Reprod. Fertil. Suppl. 31, 95±104.
Phillips, D. M.; Fadem, B. H., 1987: The oocyte of a new worldmarsupial, Monodelphis domestica: structure, formation, and func-tion of the enveloping mucoid. J. Exp. Zool. 242, 363±371.
Renfree, M. B., 1980: Endocrine activity in marsupial placentation. In:Cumming, I. A.; Funder, J. W.; Mendelsohn, F. A. O. (eds),Endocrinology 1980. Canberra, Australia: Australian Academy ofScience, pp. 83±86.
Renfree, M. B., 1993: Diapause, pregnancy, and parturition inAustralian marsupials. J. Exp. Zool. 266, 450±462.
Renfree, M. B.; Lewis, A. McD., 1996: Cleavage in vivo and in vitro inthe Marsupial Macropus eugenii. Reprod. Fert. Dev. 8, 725±742.
Renfree, M. B.; Shaw, G., 1996: Reproduction of a marsupial: Fromuterus to pouch. Anim. Reprod. Sci. 42, 393±403.
Roberts, C. T.; Breed, W. G., 1994a: Placentation in the dasyuridmarsupial, Sminthopsis crassicaudata, the fat-tailed dunnart, andnotes on placentation of the didelphid Monodelphis domestica.J. Reprod. Fertil. 100, 105±113.
Roberts, C. T.; Breed, W. G., 1994b: Embryonic-maternal cell-interactions at implantation in the fat-tailed dunnart, a dasyuridmarsupial. Anat. Rec. 240, 59±76.
Roberts, C. T.; Breed, W. G., 1996: The marsupial shell coat ± anultrastructural and immunogold localization study. Cell Tissue Res.284, 99±110.
Roberts, C. T.; Breed, W. G.; Mayrhofer, G., 1994: Origin of theoocyte shell coat of a dasyurid marsupial ± an immunohistochemicalstudy. J. Exp Zool. 270, 321±331.
Rodger, J. C.; Bedford, J. M., 1982: Induction of oestrus, recovery ofgametes, and the timing of fertilization events in the opossum,Didelphis virginiana. J. Reprod. Fertil. 64, 159±169.
Romeis, B., 1968: Mikroskopische Technik. 16th edn. MuÈ nchen; Wien:R. Oldenbourg-Verlag.
Selwood, L., 1980: A timetable of embryonic development of thedasyurid marsupial Antechinus stuartii (MacLey). Aust. J. Zool. 28,649±668.
Selwood, L., 1982: A review of maturation and fertilization inmarsupials with special reference to the dasyurid: Antechinusstuartii. In: Archer, M. (ed.), Carnivorous Marsupials. Sydney,NSW, Australia: Royal Zoological Society New South Wales,pp. 65±76.
Selwood, L., 1986a: The marsupial blastocyst ± a study of theblastocyst in the Hill Collection. Aust. J. Zool. 34, 177±187.
Selwood, L., 1986b: Cleavage in vitro following destruction of someblastomeres in the marsupial Antechinus stuartii (McLeay).J. Embryol. Exp. Morphol. 92, 71±84.
Selwood, L., 1987: Embryonic development in culture of two dasyuridmarsupials, Sminthopsis crassicaudata (Gould) and Sminthopsismacroura (Spencer) during cleavage and blastocyst formation.Gamete Res. 16, 355±370.
Selwood, L., 1992: Mechanisms underlying the development ofpatterns in marsupial embryos. Curr. Top Dev Biol. 27, 175±233.
Selwood, L., 1994: Development of early cell lineages in marsupialembryos: an overview. Reprod. Fertil. Dev. 6, 507±527.
Selwood, L., 1996: The blastocyst epithelium is not a protoderm indasyurid marsupials: a review of the evidence. Reprod. Fertil. Dev.8, 711±723.
Selwood, L., 2000: Marsupial egg and embryo coats. Cell Tiss Org 166,208±219.
Selwood, L.; Sathananthan, A. H., 1988: Ultrastructure of earlycleavage and yolk extrusion in the marsupial Antechinus stuartii.J. Morph. 195, 327±344.
Selwood, L.; VandeBerg, J. L., 1992: The in¯uence of incubationtemperature on oocyte maturation, parthenogenetic and embryonicdevelopment in vitro of the marsupial Monodelphis domestica. AnimReprod Sci. 29, 99±116.
Selwood, L.; Woolley, P. A., 1991: A timetable of embryonicdevelopment, and ovarian and uterine changes during pregnancy,in the stripe-faced dunnart, Sminthopsis macroura (Marsupialia:Dasyuridae). J. Reprod. Fert. 91, 213±227.
Selwood, L.; Young, G. J., 1983: Cleavage in vivo and in culture in thedasyurid marsupial Antechinus stuartii. J. Morphol. 176, 43±60.
Selwood, L.; Robinson, E. S.; Pedersen, R. A.; Vandeberg, J. L., 1997:Development in vitro of marsupials: a comparative review of speciesand a timetable of cleavage and early blastocyst stages of develop-ment in Monodelphis domestica. J. Dev. Biol. 41, 397±410.
Semon, R., 1894: Beobachtungen uÈ ber die Lebensweise undFortp¯anzung der Monotremen nebst Notizen uÈ ber ihre KoÈ rper-temperatur. Die EmbryonalhuÈ llen der Monotremen undMarsupialier. Zur Entwicklungsgeschichte der Monotremen.In: Semon, R. (ed.), Zoologische Forschungsreisen in Australienund Dem Malayischen Archipel. Jena, Germany: Verlag VonGustav Fischer, pp. 1±74.
Sharman, G. B., 1959: Marsupial reproduction. In: Keast, A.; Crocker,R. L.; Christian, C. S. (eds), Biogeography and Ecology in Australia.Den-Haag, The Netherlands: Dr W. Junk, pp. 332±367.
Sharman, G. B., 1961: The embryonic membranes and placentation in®ve genera of diprotodont marsupials. Proc. Zool. Soc. London 137,197±220.
Shoshani, J.; McKenna, M. C., 1998: Higher taxonomic relationshipsamong extant mammals based on morphology, with selectedcomparisons of results from molecular data. Mol. Phyl. Evol. 9,572±584.
Early ontogeny and placentation of the grey short-tailed opossum 157

Szalay, F. S., 1993: Metatherian taxon phylogeny: evidence andinterpretation from the cranioskeletal system. In: Szalay, F. S.;Novacek, M. J.; McKenna, M. C. (eds), Mammal Phylogeny. NewYork, USA: Springer Verlag, pp. 216±242.
Temple-Smith, P. D., 1987: Sperm structure and marsupial phylogeny.In: Archer, E. M. (ed.), Possums and Opossums: Studies inEvolution. Sydney, NSW, Australia: Surrey Beatty and the RoyalZoological Society of New South Wales, pp. 171±193.
Tyndale-Biscoe, C. H., 1973: Life of Marspials. London, UK: Arnold.Tyndale-Biscoe, C. H.; Renfree, M. B., 1987: Reproductive Physiology
of Marsupials (Monographs on Marsupial Biology). Cambridge,UK: Cambridge University Press.
Ward, S. J.; Renfree, M. B., 1988: Reproduction in females of thefeathertail glider Acrobates pygmaeus (Marsupialia). J. Zool., Lond.216, 225±239.
Wilson, D. E.; Reeder, D. M., 1993: Mammal Species of the World.A Taxonomic and Geographic Reference. Washington; London:Smithsonian Institution Press.
Wooding, F. B. P.; Flint, A. P. F., 1994: Ovulation and early pregnancy.In: Lamming, G. E. (ed.), Marshall's Physiology of Reproduction,4th edn, Vol. 3: Pregnancy and Lactation, Part 1, Chapter 4,Placentation. London, UK: Chapman and Hall, pp. 233±460.
Woolley, P. A., 1991: Reproduction in Dasykaluta rosamondae(Marsupialia: Dasyuridae): ®eld and laboratory observations. Aust.J. Zool. 39, 549±568.
Zeller, U., 1999a: Mammalian reproduction: origin and evolutionarytransformations. Zool. Anz. 238, 117±130.
Zeller, U., 1999b: Phylogeny and systematic relations of the Mono-tremata: why we need an integrative approach. Cour. Forsch.-Inst.Senckenberg 215, 227±232.
Authors' addresses: Professor Dr Ulrich Zeller (for correspondence),Museum fuÈ r Naturkunde, Institut fuÈ r Systematische Zoologie, Inval-idenstraûe 43, 10115 Berlin, Germany; E-mail: [email protected]; Claudia Freyer, Department of Zoology, The Universityof Melbourne, Parkville, Victoria 3052, Australia a
158 ZELLER and FREYER





























