Embed Size (px)
Citation preview

UNIVERZITET U NIŠU
PRIRODNO – MATEMATIĈKI FAKULTET
DEPARTMAN ZA BIOLOGIJU I EKOLOGIJU
MASTER RAD
GAJENJE AZOTOFIKSIRAJUĆIH I FOSFOSOLUBILIZIRAJUĆIH BAKTERIJA U
KONTROLISANIM USLOVIMA FERMENTORA U CILJU BIOFERTILIZACIJE
ZEMLJIŠTA
Niš, 2016.

UNIVERZITET U NIŠU
PRIRODNO – MATEMATIĈKI FAKULTET
DEPARTMAN ZA BIOLOGIJU I EKOLOGIJU
MASTER RAD
GAJENJE AZOTOFIKSIRAJUĆIH I FOSFOSOLUBILIZIRAJUĆIH BAKTERIJA U
KONTROLISANIM USLOVIMA FERMENTORA U CILJU BIOFERTILIZACIJE
ZEMLJIŠTA
Kandidat: Mentor:
Dunja Ţ. Ilić Prof. Dr Tatjana Mihajilov-Krstev
Niš, 2016.

UNIVERSITY OF NIŠ
FACULTY OF SCIENCES AND MATHEMATICS
DEPARTMENT OF BIOLOGY AND ECOLOGY
MASTER THESIS
GROWING NITROGENFIXATION AND PHOSPHORSOLUBIZING BACTERIA UNDER
CONTROLLED CONDITIONS IN FERMENTOR WITH A GOAL TO FERTILISE THE SOIL
Candidate: Mentor:
Dunja Ţ. Ilić Prof. dr Tatjana Mihajilov-Krstev
Niš, 2016.

ZAHVALNICA
Zahvaljujem svima koji su pomogli u izradi ovog master rada. Najveću zahvalnost
dugujem svom mentoru, prof. dr Tatjani Mihajilov – Krstev, kao i Jovani Jović, na
nesebičnoj podršci i pomoći. Veliku zahvalnost dugujem i firmi ,,Fertico“, kao i vlasniku
Slavoljubu Vukičeviću, koji su mi omogućili sprovođenje celokupnog istraživačkog rada
i dobijanje određenih rezultata.
Posebno zahvaljujem svojoj porodici na razumevanju, podršci i strpljenju za vreme
studiranja, a najviše svojoj mami, kojoj i posvećujem ovaj master rad.

SAŽETAK
Zemljišni mikroorganizmi imaju znaĉajnu ulogu u ekosistemu. Njihova višestruka korist se
ogleda u fiksaciji, solubilizaciji, mobilizaciji i obnavljanju nutritijenata u zemljištu. Obogaćivanje
zemljišta korisnim mikroorganizmima je biofertilizacija tj. implementacija u zemljište u formi
mikrobioloških Ċubriva.
U ovom radu je opisan proces optimizacije medijuma kao i Fed-Batch procesa gajenja
azotofiksatorskih i fosfosolubilizirajućih bakterija, sa ciljem povećanja njihove biomase, proizvodnje
mikrobioloških Ċubriva i njihove implementacije u zemljište. Bakterijske kulture Bacillus subtilis,
Bacillus megatherium i Azotobacter vinelandii su korišćene za ispitivanje procesa fermentacije u strogo
kontrolisanim uslovima bioreaktora. Rezultati naših istraţivanja su pokazali da je odgovarajućom
optimizacijom podloge i kinetikom Fed-batch procesa moguće postići što veću biomasu korisnih
mikroorganizama u fermentoru.
Kljuĉne reĉi: azotofiksirajuće i fosfosolubizirajuće bakterije, mikrobiološka Ċubriva, biofertilizacija.

ABSTRACT
Soils play the crucial role in the ecosystem. Their multiple use is shown in fixation,
solubilization, mobilization and recover of nutritients in soil. Soil fertilization with useful
microorganisms is biofertilization so called implementation of soil in form of microbiological fertilizer.
This work shows the process of optimization of medium as well as fed batch growth
nitrogenfixation and phosphorsolubizing bacteria in order to improve biomass and production of
microbiological fertilizer and their implementation in soil. Bacterias culture Bacillus subtilis, Bacillus
megatherium and Azotobacter vinelandii are used for research of process of fermentation in strictly
controlled conditions of bioreactor. The final results of the search shows that appropriate optimization
of the soil and kinetics fed batch process is possible to gain bigger biomass of useful microorganisms in
fermentor.
Key words: nitrogenfixation and phosphorsolubizing bacteria, microbiological fertilizer,
biofertilization.

SADRŽAJ
1. UVOD ................................................................................................................................................................1
2. OPŠTI DEO .............................................................................................................................................................2
2.1. OSOBINE BAKTERIJA I NJIHOVA GRAĐA ........................................................................................................2
2.2. ZEMLJIŠTE I MIKROORGANIZMI U ZEMLJIŠTU ...............................................................................................7
2.3. ODNOSI MIKROORGANIZAMA U ZEMLJIŠTU ................................................................................................9
2.4. ELEMENTI POTREBNI ZA RAZVOJ BILJAKA ................................................................................................. 11
2.5. CIKLUS AZOTA ............................................................................................................................................. 13
2.6. CIKLUS SUMPORA ....................................................................................................................................... 18
2.7. CIKLUS FOSFORA ......................................................................................................................................... 19
2. 8. ĐUBRIVA .................................................................................................................................................... 20
2.8.1. MINERALNA ĐUBRIVA ......................................................................................................................... 20
2.8.2. ORGANSKA ĐUBRIVA ........................................................................................................................... 23
2.9. MIKROBIOLOŠKA ĐUBRIVA I BIOFERTILIZACIJA.......................................................................................... 25
3. CILJ RADA ........................................................................................................................................................... 34
4. MATERIJAL I METODE ........................................................................................................................................ 35
4.1. MATERIJAL .................................................................................................................................................. 35
4.2. METODE ...................................................................................................................................................... 36
4.2.1. PRIPREMA BAKTERIJSKIH KULTURA ..................................................................................................... 36
4.2.2. PRIPREMA STARTNE KULTURE (inokuluma) ....................................................................................... 37
4.2.3. KONTROLA STERILNOSTI I ODREĐIVANJE BROJNOSTI MIKROORGANIZAMA ..................................... 39
4.2.4. PRIPREMA FERMENTORA .................................................................................................................... 40
4.2.5. METODE BIOFERMENTACIJE ................................................................................................................ 40
5. REZULTATI I DISKUSIJA....................................................................................................................................... 42
Bacillus subtilis ................................................................................................................................................... 42
Bacillus megaterium .......................................................................................................................................... 43
Azotobacter vinelandii ....................................................................................................................................... 44
6. ZAKLJUČAK ........................................................................................................................................................ 45
7. LITERATURA ....................................................................................................................................................... 46
8. BIOGRAFIJA ........................................................................................................................................................ 49

1
1. UVOD
U poljoprivrednoj proizvodnji i uzgajanju biljnih kultura je neophodno zemljištu dodavati
odreĊene asimilate, jer se oni, ili u velikoj koliĉini nalaze u vazduhu (azot), ili su prisutni u
zemljištu u formi koja je nedostupna biljkama (fosfor, azot, kalijum, gvoţĊe i dr.). Zbog toga
se zemljištu dodaju veštaĉka sintetska Ċubriva. Rezultati velikog broja istraţivanja su pokazali
da sintetsko azotno Ċubrivo to jest, dodavanje kombinovanog azota u zemljište u formi koja je
laka za asimilovanje, kroz duţi vremenski period, dovodi do potsticanja razvoja
mikroorganizama (bakterija i plesni) koje se hrane organskom materijom. Sa smanjenjem
sadrţaja organske materije, zemljište podleţe kompaktizaciji, ĉime se ograniĉava rast korena
biljaka i smanjuje sposobnost zadrţavanja vode te zemljište postaje zavisnije od iritacije i
podloţnije eroziji. Sa promenama u fiziĉko – hemijskim karakteristikama zemljišta i sa
smanjenjem raspoloţivog kiseonika usporava se rastmikroflore zemljišta, menja se struktura
prirodnih mikrobiocenoza i sloţeni sistem biološke razmene asimilativa se narušava.
Antropogeno delovanje i navedene promene u zemljištu su prisutne i na obradivim
površinama u Srbiji. Osim toga, povećanje oticanja nitrata dovodi do zagaĊenja nadzemnih
voda, dok emisija azot-dioksida u atmosferu znaĉajno doprinosi efektu staklene bašte. I
najzad, proizvodnja veštaĉkih Ċubriva troši rezerve fosilnih goriva, zagaĊuje ţivotnu sredinu i
veoma mnogo košta.
Mikroorganizmi su, kao neizostavni ĉlanovi svih ţivotnih zajednica, kljuĉni za odrţavanje
biogeohemijskih ciklusa kruţenja materija na Zemlji, i kao najznaĉajnija komponenta
biološke faze zemljišta, vaţni indikator plodnosti i procesa degradacije u njemu. Pored
azotofiksatora koji vraćaju azot iz atmosfere u zemljište, mikroorganizmi mobilišu fosfor,
kalijum, gvoţĊe, sumpor i ĉine ih dostupnim biljkama. Budućnost poljoprivredne proizvodnje
je u primeni biofertilizacije (unošenje ţivih mikroorganizama u zemljište) to jest, u upotrebi
mikrobioloških Ċubriva.
U ovom radu je vršena optimizacija uslova za gajenje azotofiksirajućih bakterija u cilju
proizvodnje velike biomase tj. mikrobioloških Ċubriva.

2
2. OPŠTI DEO
2.1. OSOBINE BAKTERIJA I NJIHOV ZNAĈAJ I GRAĐA
Mikrobiologija je nauka koja se bavi prouĉavanjem mikroorganizama, odnosno
ispitivanjem ţivota veoma sitnih organizama. Naziv mikrobiologija potiĉe od grĉkih reĉi
micros-mali, bios-ţivot, logos-nauka. Mikrobiologija se bavi prouĉavanjem svih pojava i
zakonitosti vezanih za ţivot mikroorganizama, odnosno ona ukljuĉuje morfologiju,
fiziologiju, genetiku, ekologiju i sistematiku mikroba.
Mikroorganizmi su veoma stara grupa organizama, negde oko 4 milijardi godina i
imali su ulogu u formiranju ţivota na Zemlji, a i danas uĉestvuju u njegovom odrţavanju.
Sam naziv kaţe, to su veoma mali organizmi, koji se ne mogu golim okom videti, jer su
manji od 0,1 mm. Mikrobi predstavljaju veoma heterogenu grupu u svakom pogledu, oni
obuhvataju najsitnije organizme – viruse veliĉine 0, 01 mikrometara (virusi nemaju ćelijsku
graĊu), ali obuhvataju i organizme vidljive golim okom (neke alge i gljive). Kao što je već
reĉeno, mikroorganizmi su heterogena grupa koja obuhvata viruse, bakterije, alge, gljive,
lišajeve i protozoe, pa u zavisnosti od toga, mikrobiologija obuhvata sledeće discipline:
bakteriologiju, virusologiju, algologiju, mikologiju, lihenologiju i protozologiju. Oni imaju
razliĉit oblik, veliĉinu, graĊu i fiziologiju, ali se u njihovom prouĉavanju uglavnom koriste
iste metode, dok su im zajedniĉke karakteristike: mala veliĉina tela, brz metabolizam, brzo
razmnoţavanje, rasprostranjenost po celom svetu – kosmopoliti (Petrović et al., 2007).
Mikroorganizmi koji nemaju ćelijsku graĊu su virusi, dok su mikroorganizmi sa
prokariotskim tipom ćelija (nemaju organizovano jedro) bakterije i modrozelene alge, a sa
eukariotskim tipom ćelija (imaju organizovano jedro) su protozoe, gljive, lišajevi, alge.
Uloga mikroorganizama je veoma velika, neophodni su u mnogim procesima. Oni
imaju degradacionu ulogu, to jest, ulogu razlagaĉa, ĉime se omogućava kruţenje materije i
proticanje energije u prirodi. To uglavnom obavljaju bakterije i gljive. Mikroorganizmi su i
producenti primarne organske materije i proizvoĊaĉi kiseonika, a za to su zasluţne alge. Oni
su neizostavni deo ţivota ĉoveka. Njihov uticaj moţe biti i štetan i koristan.
Štetni mikroorganizmi su izazivaĉi mnogih bolesti, pa tako ostvaruju negativan uticaj
na ĉoveka. Mikrobi su neophodni u proizvodnji hrane i pića (proizvodnja jogurta, pravljenje
hleba, turšija). Koriste se u hemijskoj, farmaceutskoj industriji, rudarstvu, agronomiji, zaštiti
šivotne sredine, medicini, genetici, dobijanju energije i drugim oblastima, što prikazuje njihov

3
pozitivan efekat po ĉoveka. Na osnovu ovoga, vidimo da je mikrobiologija veoma bitna
nauĉna disciplina, koja se još uvek razvija i koja ima veliku primenu u svim segmentima
ţivota, dok se mikroorganizmi sve više i više primenjuju u razliĉitim granama. Mikrobiologija
je doprinela razvoju molekularne biologije, genetike, biohemije, biotehnologije (Petrović et
al., 2007).
Bakterije spadaju u prokariotske mikroorganizme. One su veoma malih dimenzija,pa
se posmatraju pomoću mikroskopa. Bakterije su najsitniji organizmi celularne graĊe. Postoje
razliĉiti oblici bakterijskih ćelija, ali se kao osnovni oblici izdvajaju: loptast, štapićast, izvijen
i konĉast oblik. Loptaste bakterije su koke, koje obuhvataju više tipova: monokoke,
diplokoke, tetrade, sarcine, u zavisnosti od naĉina razdvajanja nakon deobe. Štapićaste
bakterije su bacilli koji isto obuhvataju razliĉite tipove. Bakterije koje imaju izvijen oblikili
izgled zapete su vibrioni. Konĉast oblik imaju bakterije kod kojih je jedna dimenzija
izraţenija od ostalih i one mogu biti granate i negranate. Postoje i mnogidrugi izvedeni oblici
bakterija (Petrović et al., 2007).
Bakterije u svom sastavu imaju odreĊene morfološke i funkcionalne strukture (Slika
1.) koje obuhvataju:
- površinske ćelijske strukture i vanćelijske tvorevine (spoljašnji omotaĉi, ćelijski zid
i izraštaji na ćeliji);
- citoplazmu, citoplazmatske ćelijske strukture i ćelijske inkluzije (citoplazma,
citoplazmatska membrane i intracelularne membranske tvorevine, ribozomi,
nukleoid).
Spoljašnji omotaĉi bakterijskih ćelija su:
- Glikokaliks predstavlja omotaĉ koji naleţe na ćelijski zid. On je izgraĊen od homo
ili heteropolisaharida, a reĊe od glikoproteina i polipeptida.
- Kapsula predstavlja kompaktan omotaĉ bakterijske ćelije koji je izgraĊen od
polisaharida, ĉvrstine gela. Postoje razliĉite vrste polisaharida koje grade kapsulu.
Nemaju sve bakterijske ćelije kapsulu na svojoj površini. Bakterije koje imaju
kapsulu uglavnom imaju sjajnu i glatku površinu – S tip kolonija, dok one koje je
nemaju su hrapave i mat – R tip kolonija. Osnovna funkcija kapsule je zaštita
bakterijskih ćelija. Ona je osmotska barijera, spreĉava isušivanje ćelija i štiti ćeliju
od spoljnih uticaja.
- Sluzna sara predstavlja rigidni omotaĉ proteinsko-polisaharidno-lipidne graĊe
gvoţĊevitih i manganskih bakterija.

4
- S omotaĉi su planarne kristalne strukture na samoj površini ćelija, izgraĊene od
proteina ili glikoproteina. Kod arhea su jedini omotaĉi, ali se sreću i kod ostalih
bakterija.
Slika 1. GraĊa bakterijske ćelije
Ćelijski zid bakterija je sloţene graĊe i ĉine ga polisaharidi i peptidi, koji su povezani
kovalentnom vezom. Njegova osnovna jedinica graĊe je muropeptid koji je sastavljen iz N-
acetilglukozamina povezanog β (1-4) glikozidnom vezom sa N-acetilmuraminskom
kiselinom, vezanom za tetrapeptid. Ćelijski zid se razlikuje u graĊi kod Gram pozitivnih i
Gram negativnih bakterija. Razlika je po tipu veza u mureinu, hemijskom sastavu i strukturi.
Osnovna funkcija ćelijskog zida je da daje oblik ćeliji i štiti od spoljašnjih uticaja. On ne
predstavlja osmotsku barijeru, jer je porozan. Antibiotski agensi narušavaju strukturu
ćelijskog zida, jer ometaju njegovu sintezu (Petrović et al., 2007).
Bakterije se mogu kretati pasivno i aktivno. Pasivan naĉin kretanja bakterija je rezultat
fiziĉkih sila i pokretanja ĉestica teĉnosti, dok je aktivan naĉin kretanja bakterija rezultat
metaboliĉkih procesa u samoj ćeliji, pri ĉemu se bakterije kreću klizeći ili plivajući. Bakterije
se kreću plivajući pomoću flagela. Flagele su tanki i dugaĉki izraštaji koji se sastoje iz
bazalnog tela, kuke i filamenta (u ĉijem sastavu ulaze proteinske subjedinice flagelina). Za
pokretanje flagela je neophodno prisustvo Mot proteina, bitnog za stvaranje energije. Flagele
se pokreću rotacijom, u smeru suprotnom kazaljci na satu. Broj flagela kod bakterija je
razliĉit, mogu imati jednu flagelu i to su monotrihe bakterije ili više njih i to su politrihe
bakterije.Bakterije koje nemaju flagele su atrihe. Spirohete imaju specifiĉne strukture za

5
kretanje, odnosno aksijalni filament. Ukoliko su pokreti bakterija uslovljeni nekim nadrţajem,
onda se nazivaju taksije (hemotaksije, fototaksije).
Bakterije imaju još površinskih struktura, a to su fimbrije i pili. Fimbrije nemaju ulogu
u kretanju, već omogućavaju adheziju bakterija. One su tanje od flagela i u mnogo većem
broju (Petrović et al., 2007).
Pili su sliĉni fimbrijama, graĊeni od subjedinica pilina i uglavnom se javljaju jedan do
tri. Njihova uloga je u razmeni genetiĉkog materijala, u procesu konjugacije, pa se zato i zovu
sex pili.
Citoplazma bakterija se sastoji iz vode, 70 – 80 %, u kojoj su rastvoreni razliĉiti
organski molekuli, koji su gradivne materije ćelije (aminokiseline, peptidi, proteini, lipidi,
ugljeni-hidrati, vitamini, joni). Koliĉina vode odreĊena je fiziološkim procesima u ćeliji,
starenjem ćelija, kao i spoljnim uticajima. U citoplazmi bakterijskih ćelija se ne javljaju
organele karakteristiĉne ze eukariote. Prisutni su ribozomi, koji imaju ulogu u sintezi proteina.
Broj ribozoma zavisi od metaboliĉkih aktivnosti ćelija. Bakterije kao ekvivalent jedru imaju
nukleoid, koji predstavlja deo citoplazme sa DNK, ali bez jedrovog omotaĉa (Petrović et al.,
2007).
Intracelularne membranske tvorevine – mezozomi, u širem smislu su razliĉite
morfološke invaginacije citomembrane koje predstavljaju morfološki i funkcionalno
jedinstven sistem. Ove tvorevine su, zapravo, analozi eukariotskih organela, pa tako ovde
spadaju:
- analozi mitohondrija imaju ulogu u ćelijskoj respiraciji;
- mezozomi imaju ulogu u deobi Gram pozitinih bakterija;
- kruškaste tvorevine imaju ulogu u replikaciji DNK;
- analozi glatkog, hrapavog endoplazmatiĉnog retikuluma i druge strukture.
Citoplazmatske inkluzije se mogu javljati u ćelijama i biti organskog ili neorganskog
porekla. Uglavnom predstavljaju rezervne hranljive materije (primer, glikogen i skrob, koji su
polisaharidne inkluzije) (Petrović et al., 2007).
Neki bakterijski rodovi imaju sposobnost formiranja endospora. To su zaštitne,
latentne forme ili forme za konzervaciju i uglavnom ih formiraju Gram pozitivne bakterije
(Clostridiumi Bacillus). Enzimska i metaboliĉka aktivnost endospora svedena je na minimum
ili je potpuno odsutna pa ona predstavlja dormantni stadijum. Spore nastaju tokom
sporogeneze, odnosno sporulacije koja poĉinje kada dostupnost hranljivih materija kreće da
opada u stacionarnoj fazi rasta. Spora je u stanju mirovanja sve dok ne dospe u povoljne
uslove za ţivot, što moţe trajati i nekoliko stotina godina. Aktivacija spora kreće pod uticajem

6
nekih spoljašnjih faktora. Spore imaju zaštitnu ulogu, odnosno omogućavaju preţivljavanje
nepovoljnih perioda.
Bakterije stvaraju i dodatne formacije koje su oznaĉene kao ciste (karakteristiĉne za
rod Azotobacter). Ciste su otporne na isušivanje, mehaniĉke uticaje i UV zraĉenje, ali nisu
otporne na visoke temperature i nisu potpuno dormantne. Postoje i mikrospore ćija je
otpornost mnogo manja nego li kod endospora. Neke bakterije stvaraju i egzospore, dok su za
metanogene i fotosintetske bakterije karakteristiĉne dormantne egzospore. Aktinomicete i
Gram pozitivne filamentozne razgranate bakterije formiraju konidije, koje prvenstveno sluţe
za razmnoţavanje (Petrović et al., 2007).
Kao i svi drugi organizmi, bakterije rastu i razmnoţavaju se. Za rast bakterija
neophodna je hrana koja potiĉe iz spoljašnje sredine, pri ĉemu bakterije usvajaju hranljive
materije. Hrana je neophodna za sve ţivotne procese i predstavlja izvor energije, kao i
gradivne materije. Pod rastom prokariotskih ćelija podrazumeva se povećanje koliĉine svih
hemijskih komponenti koje ih grade. U mikrobiologiji se uglavnom prati rast populacije, koja
predstavlja skupinu ogromnog broja ćelija koje vode poreklo od jedne ćelije. Ukoliko su
populacije nastale u definisanim uslovima sredine, pod kontrolisanim laboratorijskim
uslovima, oznaĉavaju se kao kulture. Kulture su ĉiste ako sadrţe samo jednu vrstu organizma,
a mešovite ako sadrţe više vrsta organizama. Rast bakterija nije beskonaĉan, jer nakon
dostizanja odreĊenih veliĉina ćelija stupa u deobu. Rast je uvek praćen deobom ćelija što
dovodi do povećanja broja ćelija. Bakterije se razmnoţavaju prostom ćelijskom deobom, koja
je binarna, popreĉna i dovodi do obrazovanja dve identiĉne ćerke ćelije. Deoba poĉinje
udvajanjem genetiĉkog materijala. Nasledni materijal bakterija je jedan, dvostruki,
cirkulatorni DNK molekul, koji je smešten u nukleoidu (Petrović et al., 2007).
Razmena genetiĉkog materijala kod prokariota, a samim tim i bakterija vrši se:
- transformacijom, usvajanjem DNK fragmenata jedne bakterije od strane druge
bakterije, direktno iz spoljašnje sredine u koju ona dospeva nakon smrti i lize
bakterijske ćelije;
- konjugacijom, uspostavljanjem bliskog kontakta dve bakterije pomoću sex pila (F
pila);
- transdukcijom, prenošenje genetskog materijala sa jednje bakterije na drugu
pomoću bakteriofaga (virusi koji inficiraju bakterije) (Petrović etal., 2007).
Na osnovu metode diferencijalnog bojenja bakterija po Gramu, koje je dobilo naziv po
danskom nauĉniku Kristian Gramu, bakterije se dele na: Gram pozitivne i Gram negativne
bakterije i zbog toga, ovaj naĉin bojenja sluţi za identifikaciju bakterijskih grupa. Ovim

7
bojenjem Gram pozitivne bakterije se boje ljubiĉastom bojom, a Gram negativne bakterije se
boje crvenom bojom. Ove razlike su uslovljene fiziĉko – hemijskim osobinama bakterija, a i
hemijskim sastavom ćelijskih zidova. Gram negativne bakterije u svom ćelijskom zidu imaju
lipopolisaharid, dok Gram pozitivne bakterije imaju peptidoglukane, koje etanol ne rastvara
pa i nakon obezbojavanja ostaju ljubiĉaste. Lipopolisaharide etanol rastvara pa se u sledećem
bojenju boje crveno (Mihajilov – Krstev, 2008).
Mikroorganizmi su kosmopoliti, odnosno oni imaju širok spektar rasprostranjenja.
Naseljavaju celokupnu biogeosferu (pedosferu, hidrosferu, atmosferu i biosferu).
Mikroorganizmi imaju znaĉajnu ulogu u stvaranju, odrţavanju i funkcionisanju razliĉitih
ekosistema i rasprostranjeni su u svim ekološkim sistemima, pri ĉemu se njihova aktivnost
odvija stalnom interakcijom abiotiĉkih i biotiĉkih komponenti (Petrović et al., 2007).
2.2. ZEMLJIŠTE I MIKROORGANIZMI U ZEMLJIŠTU
Zemljište (pedosfera) je obavezna komponenta svake biogeocenoze i predstavlja tanak
površinski sloj zemljine kore. Zemljište nastaje procesom pedogeneze, usled dugotrajnog
delovanja klimatskih, bioloških i reljefnih faktora na matiĉnu stenu. Zemljište je polidisperzni
sistem i ĉine ga tri faze: ĉvrsta, teĉna i gasovita faza. Tip i kvalitet zemljišta odreĊuju mnogi
faktori kao što su njegove fiziĉke osobine (veliĉina ĉestica), organski ostatak (humus, nastaje
degradacijom ţivotinjskih i biljnih otpadaka), voda i gasovi. Zemljište predstavlja ţivotnu
sredinu mnogih biljaka, ţivotinja i mikroorganizama, a njegova ekološka uloga se razmatra u
vidu:
- biogeocenotiĉke funkcije zemljišta i
- globalne funkcije zemljišta (Đukić et al., 2000).
Globalnim funkcijama zemljišta pripadaju:
biohemijske transformacije površinskih slojeva litosfere;
hidrološke funkcije zemljišta;
uticaj zemljišta na gasni reţim i kruţenje vode u atmosferi;
uloga zemljišta kao faktora evolucije (Đukić et al., 2000).
Biogeocenotiĉke funkcije zemljišta su u neposrednoj vezi sa ţivotnom aktivnošću
zemljišnih mikroorganizama. Ovde spadaju:
- fiziĉke funkcije (stanište i sklonište, mehaniĉki oslonac, depo semena);

8
- fiziĉko – hemijske funkcije (izvor hranljivih elemenata; stimulator i
inhibitor biohemijskih procesa; depo vlage, hranljivih materija i energije;
sorpcija materija i mikroorganizama);
- informacione funkcije (signal za niz bioloških procesa; regulator brojnosti
istrukture biogeocenoza; pogonski mehanizam nekih sukcesija; odrţavanje
istorije razvoja biogeocenoze)
- opšteekološke funkcije (akumulacija i transformacija materije i energije;
sanitarna funkcija; puferni i zaštitini biogeocenotiĉki ekran; uslovi nastanka
i evolucije organizama) (Đukić et al., 2007).
Zemljište kao ţivotna sredina pruţa najpovoljnije uslove za ţivot skoro svih grupa
mikroorganizama. Mikrobi koji se nalaze u zemljištu svojom aktivnošću utiĉu na osnovne
karakteristike zemljišta, njegovu strukturu i plodnost. Zbog svega ovoga, mikroorganizmi koji
ţive u zemljištu se smatraju posebnom grupom i predstavljaju nerazdvojni ĉinilac zemljišta.
Tako zemljište predstavlja njihov rezervoar, pri ĉemu oni dospevaju i u druge ţivotne sredine.
Odnos mikroorganizama u zemljištu je uravnoteţan i karakteristiĉan za svaki tip zemljišta.
Ukoliko se zemljište tretira pesticidima, obraĊuje i koriste Ċubriva odnos mikroorganizama se
narušava.To se manifestvuje promenom brojnosti, sastava i aktivnosti mikroorganizama. Ove
karakteristike (brojnost i aktivnost mikroorganizama) karakterišu se kao plodnost zemljišta
(Đukić et al., 2007).
U zemljištu mikroorganizmi imaju odreĊeni raspored i on moţe da bude:
- horizontalni raspored mikroorganizama;
- vertikalni raspored mikroorganizama.
Horizontalni raspored mikroba u zemljištu podrazumeva njihovo prisustvo od polova
ka ekvatoru. On je uslovljen najviše klimatskim faktorima. Na osnovu ovog rasporeda,
utvrĊeno je da su juţnija zemljišta bogatija mikoorganizmima.
Vertikalni raspored zemljišta se definiše kao raspored mikroorganizama po dubini
zemljišta. Tu je izraţena odreĊena pravilnost koja govori o odreĊenim mikrobiološkim
profililma u zemljištu, odnosno o posebnom rasporedu mikroorganizama. Ovaj mikrobiološki
profil je karakteristiĉan za svaki tip zemljišta. Kada je organska materija zastupljenija na
površini zemljišta, onda se mikroorganizmi nalaze bliţe površini i tu je njihova brojnost veća
(Petrović et al., 2007).
Mikrobno naselje u zemljištu ima veoma vaţnu i veliku ulogu. Mikroorganizmi
uĉestvuju u nastanku i stvaranju zemljišta (pedogenezi), kao i u njegovoj daljoj evoluciji.
Svojom aktivnošću oni doprinose promenama u sredini, ali i prilagoĊavaju uslove svojim

9
potrebama. Istovremeno su sposobni da se adaptiraju promenjenim uslovima. Tako utiĉu,
pored stvaranja i na odrţavanje sadašnjeg zemljišta. Oni se u zemljištu nalaze u ogromnim
koliĉinama, ali njihov precizan broj nije utvrĊen.
Na brojnost mikroba u zemljištu utiĉu mnogi faktori, kako ekološki, tako i mnogi
drugi. Najveći uticaj imaju njegove fiziĉko – hemijske osobine, tip zemljišta, karakter
vegetacije, temperatura, dostupnost vode i hranljivih materija, pH vrednosti i biotiĉki faktori.
Najveća brojnost mikroorganizama je u zemljištu koje ima visok sadrţaj organskih materija,
odnosno humusa. Nastanak humusa je uslovljen i aktivnošću mikroorganizama, pri ĉemu se
od biljnih i ţivotinjskih ostataka, procesima fermentacije, amonifikacije i saponifikacije
dobijaju CO2, H2O, NH3, mineralne soli, druga organska jedinjenja i mikrobiološki protein.
Polimerizacijom i kondenzacijom dobija se organska materija koja se razlikuje od polaznih
jedinjenja, a sadrţi visok sadrţaj humusnih kiselina pa se zato zove humus, a sam proces je
humifikacija. Ove humusne kiseline se stvaraju od fenolnih azotnih jedinjenja (Đukić et al.,
2007).
U zemljištu su zastupljene razliĉite vrste mikroorganizama i one se oznaĉavaju kao
zemljišni mikroorganizmi. Najzastupljenije su bakterije, aktinomicete, gljive, alge i na kraju
protozoe. Aktinomicete su zastupljene u manjem broju od bakterija i njihova uloga je u
razgradnji kompleksnih organskih supstrata. Odlikuje ih produkcija antibiotika. Gljive koje se
nalaze na površini zemljišta imaju ulogu u razgradnji celuloze, pektina, lignina, povezivanju
ĉestica zemljišta. Zelene i silikatne alge imaju ulogu u formiranju zemljišta. Što se bakterija
tiĉe, javljaju se i autotrofni i hetrotrofni predstavnici, aerobne i anaerobne bakterije koje mogu
biti i mezofili, termofili. Njihova uloga je ogromna, svojom metaboliĉkom aktivnošću,
celulolitskom, proteolitskom ili uzimanjem šećera tokom azotofiksacije imaju vaţnu ulogu u
obogaćivanju zemljišta azotom (Đukić et al., 2007).
2.3. ODNOSI MIKROORGANIZAMA U ZEMLJIŠTU
Mikroorganizmi u zemljištu su meĊusobno povezanim razliĉitim odnosima,ali su
takoĊe, povezani i sa drugim stanovnicima zemljišta, algama, ţivotinjama, biljkama (njihovim
korenovim sistemom). Ovi odnosi su veoma sloţeni. Njihovu osnovu ĉine kruţenje materije i
proticanje energije pri ĉemu se u toku ţivotne aktivnosti mikrobne populacije razmenjuju
izmeĊu sebe i sa drugim predstavnicima, svojim proizvodima metabolizma, biološki aktivnim
supstancama. Odnosi izmeĊu mikroorganizama su neutralizam (bez efekata populacija jedne
na drugu), amensalizam (jedna populacija suzbija drugu, ali ne trpi negativno dejstvo),
parazitizam (jedna populacija nepovoljno deluje na drugu, ali zavisi od nje), komensalizam

10
(jedna populacija ima korist iz zajednice, a druga nema ni korist, ni štetu), protokooperacija
(obe populacije imaju korist, ali njihova veza nije obligatna) i mutualizam (obe populacije
imaju korist i ne mogu jedna bez druge) (Đukić et al., 2007).
Odnosi izmeĊu mikroorganizama se dele u dve grupe i to su antagonizam i simbioza.
Prilikom borbe za hranu, pogotovo ako ona nedostaje, dominiraju konkurencija i
amensalizam. To moţe dovesti i do eliminacije jedne populacije. PrilagoĊeniji mikrobi mogu
istisnuti one manje prilagoĊene. Jedan od naĉina borbe jeste produkcija antibiotika jedne
populacije, koji ima negativni uticaj na drugu populaciju. Tako, na primer, Streptomyces
olivocinereus, koji je producent heliomicina, suzbija u nesterilnom zemljištu populaciju
Arthrobacter crystallopoietes (Đukić et al., 2007).
Simbiotski odnosi se manifestvuju razliĉito i oni se razvijaju po tipu komensalizma.
Primećena je koegzistencija izmeĊu celuloznih mikroorganizama i slobodnih azotofiksatora, u
osnovi ĉega leţi protokooperacija (Cellvibrio, Corynobacterium + Clostridium; mikromicete
+ Clostridium; Cytophaga + Azotobacter) (Đukić et al., 2007).
Pored meĊusobnih odnosa mikroorganizama znaĉajni su i njihovi odnosi sa
zemljašnim ţivotinjama, koji mogu biti pozitivni i negativni. Mikrobi ţivotinjama mogu
sluţiti kao hrana, ali oni mogu da stvaraju odreĊene antibiotiĉke materije protiv odreĊenih
zemljišnih ţivotinja (protozoe).
Znaĉajni su i odnosi izmeĊu algi i mikroba, pri ĉemu su zasnovani na protokooperaciji
i obe populacije imaju korist. Alge se primenjuju u poljoprivredi u ulozi biostimulatora
mikroba i biljaka, a i u vidu organskog Ċubriva, što omogućava manju upotrebu veštaĉkih
Ċubriva. U samom zemljištu se aktiviraju mikrobiološki procesi, smanjuju se gubici
mineralizovanog azota, zemljište se obogaćuje mikroelementima i drugim materijama (Đukić
et al., 2007).
Odnosi izmeĊu mikroorganizama i biljaka isto mogu biti pozitivni i negativni.
Negativni odnosi se odnose na produkciju jedinjenja kojia imaju inhibitorno dejstvo na rast
biljaka, ali i jedinjenja koja izazivaju razliĉita oboljenja biljaka, koja mogu zahvatati
nadzemni deo ili korenov sistem. Postoji mnogo toksiĉnih oblika bakterija i nalaze se u svim
tipovima zemljišta. Poznati su Bacillus i Pseudomonas.
Pozitivni odnosi su oni koji ĉine osnovu pedogeneze i plodnosti zemljišta. Mikrobi
zajedno sa algama pripremaju mrtve supstrate za razvoj poĉetnih biljnih zajednica. Osnovni
destruktori biljnog otpada su mikroorganizmi i time doprinose kruţenju ugljenika i drugih
elemenata u prirodi. Bakterije i drugi oblici mikroorganizama doprinose ishrani biljaka. Oni
se razvijaju aktivno u zoni korenova i njihovoj blizini transformišu organska i mineralna
jedinjenja zemljišta, snabdevajući biljke potrebnim elementima ishrane. Mikrobi stvaraju i

11
biološki aktivne materije – vitamine, aminokiseline, biljne hormone (auksine i gibereline) koji
imaju ulogu u ţivotu biljke, disanju, fotosintezi, oploĊivanju, fosfornu i azotnu razmenu
(Đukić et al., 2007).
Biljke takoĊe utiĉu na razvoj mikroba u zemljištu stvarajući im povoljne uslove za
njihov opstanak. Tokom njihove vegetacije izdvajaju se mineralna i organska jedinjenja koja
predstavljaju hranu mikrobima. One regulišu pH zemljišta i nivo vlaţnosti što stvara povoljne
uslove za rast mikroba (Đukić et al., 2007).
2.4. ELEMENTI POTREBNI ZA RAZVOJ BILJAKA
Za rast i razvoj biljaka neophodni su mnogi elementi. Biljke primaju razliĉite
mineralne soli iz zemljišta na kome ţive. Elementi bez kojih biljke ne mogu da ţive
oznaĉavaju se kao esencijalni elementi i njih ĉini grupa od 17 elemenata. To su ugljenik,
kiseonik, vodonik, azot, sumpor, fosfor, kalijum, kalcijum, magnezijum, gvoţĊe, mangan,
cink, bakar, bor, hlor, molibden, nikl. Korisni elementi su elementi bez kojih biljka moţe da
opstane,ali ipak imaju pozitivni efekat ukoliko su prisutni (silicijum je koristan za mnoge
trave, jer daje ĉvrstinu ćelijskom zidu; aluminijum utiĉe na boju nekih cvetova, a samim tim i
na oprašivanje). Na osnovu potreba biljaka, elemente moţemo da podelimo na:
- makroelemente, biljci potrebni u velikim koliĉinama (N, P, K, Ca, Mg, S );
- mikroelemente, biljci potrebni u malim koliĉinama (B, Cl, Fe, Mn, Zn, Cu, Mo, Ni)
(Kastori., 1991).
Uloga ovih elemenata je višestruka i raznovrsna. Zbog toga su oni grupisani u
nekoliko grupa:
1. Elementi koji ulaze u sastav organskih molekula. Veliki broj elemenata uĉestvuje
u izgradnji najvaţnijih organskih jedinjenja. To su ugljenik, vodonik i kiseonik.
Ugljenik, vodonik i kiseonik biljke dobijaju iz vode i ugljen-dioksida. Pored ovih
elemenata, ovde spadaju i azot, fosfor, sumpor, magnezijum, bakar i cink.
2. Kofaktori u enzimskim reakcijama. U većini enzimskih reakcija neophodno je
uĉešće nekog mineralnog elemenata. Mnogi enzimi sadrţe jone metala (kalcijum
ulazi u sastav amilaze, cink u alkoholnoj dehidrogenazi).
3. Uĉesnici u transportu elektrona. Najrasprostranjeniji su prenosioci koji sadrţe jon
gvoţĊa, bakar i molibden.
4. Slobodni joni. Neki elementi se nalaze kao slobodni joni u citosolu i tako
odrţavaju osmotski potancijal ćelije. Takvu ulogu imaju kalijum i hlor, ali mogu

12
inatrijum i kalijum da doprinesu odrţavanju osmotskog potencijala (Kastori.,
1991).
Azot je element koji uĉestvuje u izgradnji proteina, nukleinskih kiselina, u formiranju
peptidnih veza. On obezbeĊuje porast i plodonošenje voćaka, pre svega, uslovljava rast i
razvoj svih organa. U sluĉaju njegovog nedostatka biljka usporava svoj rast, zbog smanjene
sinteze organskih jedinjenja. Lišće gubi zelenu boju, sitniji su listovi, cvetovi i plodovi.
Ukoliko se nalazi u većim koliĉinama isto nepovoljno utiĉe na biljku. Stabla i listovi su bujno
razvijeni, koren je manji, smanjeni su plodovi.
Sumpor ulazi u sastav aminokiselina, koenzima i vitamina. Nedostatak sumpora je
redak u prirodi, ali dovodi do istih simptoma kao i nedostatak azota. Biljke su male,
zakrţljalog rasta, listovi su mali, uski. Prevelika koliĉina sumpora za biljku je štetna.
Fosfor je sastavni deo ATPa, nukleotida, nukleinskih kiselina, fosfolipida,
fosforilizovanih šećera, koenzima NAD, NADP, FAD. Biljka jednu istu koliĉinu fosfora moţe
stalno da koristi i to je reutilizacija fosfata, tako što fosfat prelazi iz starijih delova biljke u
mlaĊe. U sluĉaju njegovog nedostatka ili povećane koncentracije, dolazi do promena na
biljkama, odnosno nekroze biljaka.
Kalijum je jedini elemenat koji se u biljci javlja iskljuĉivo u jonskom obliku. On se
nikada ne ugraĊuje u organska jedinjenja. Kalijuma ima najviše u meristemskim tkivima u
kojima se vrše aktivne ćelijske deobe. Kalijum uĉestvuje u uspostavljanju osmotskog
potencijala (Kastori., 1991).
Kalcijum inkrustrira ćelijske zidove, odrţava integritet membrane i spreĉava njena
oštećenja i pasivnu propustljivost. Neophodan je za dejstvo mnogih enzima.
Joni magnezijuma su aktivatori mnogih enzima i uĉestvuju u graĊi molekula hlorofila.
GvoţĊe ima ulogu u razliĉitim redoks sistemima, jer lako moţe da primi i otpusti
elektron.
Mangan kao dvovalentni je aktivator mnogih enzima u ciklusu TCA, sintezi masnih
kiselina i nukleinskih kiselina.
Bakar uĉestvuje u nekim redoks sistemima, jer njegov atom moţe da menja valencu.
Sastavni je deo citohrom oksidaze u mitohondrijama i plastocijanina u hloroplastima
(Kastori., 1991).
Cink je sastavni deo alkoholne dehidrogenaze i u njegovom nedostatku biljke su
malog rasta, sa kratkim internodijama.
Bor je esencijalan samo za više biljke i lokalizovan je u ćelijskom zidu gde utiĉe na
transport kroz membranu, naroĉito na transport šećera. Molibden je sastavni deo nitratne
reduktaze i u njegovom nedostatku biljke ne mogu da koriste nitrat. Kod mikroorganizama

13
koji fiksiraju azot, molibden je neophodan i kao sastavni deo enzima nitrogenaze (Kastori.,
1991).
Hlor je kofaktor u fotosintezi pri transportu elektrona od vode do fotosistema II.
Prateći je anjon pri transportu kalijuma i ima znaĉaja za odrţavanje turgora. U sluĉaju
nedostatka svih ovih elemenata ili ukoliko se oni nalaze u povećanim koncentracijama dolazi
do promena na biljkama, koje uglavnom utiĉu na rast i boju biljaka (Kastori., 1991).
Azot i sumpor ulaze u većinu organskih jedinjenja samo u redukovanom obliku i zato
njihovi oksidovani oblici, nitrat i sulfat moraju prethodno da se redukuju. Redukciju azota i
sumpora obavljaju samo biljke. Azot je najobilniji sastojak vazduha, ali biljke ne mogu da
koriste atmosferski azot za svoje potrebe. Tu sposobnost imaju samo izvesne bakterije i
aktinomicete. One redukciju obavljaju u biljci (biljka odmah ukljuĉuje amonijum u svoje
metaboliĉke procese) ili u zemljištu, pri ĉemu ga biljka apsorbuje iz zemljišta (Kastori.,
1991).
2.5. CIKLUS AZOTA
Azot se nalazi u vazduhu i ĉini 78% atmosferskog vazduha, u obliku neorganskih jona
NO3- i NH4
- ulazi u sastav zemljišta i kao sastojak mnogih organskih jedinjenja rasprostranjen
je u celom ţivom svetu. Ovi oblici azota se neprikidno transformišu jedan u drugi i sve te
promene su povezane u ciklus azota. Više biljke dobijaju azot iskljuĉivo iz zemljišta na kome
ţive. Zemljište se stalno obogaćuje azotnim jedinjenjima koja potiĉu od ţivih ili izumrlih
ţivotinja i biljaka. Skoro 90% azota u zemljištu se nalazi u obliku organskih azotnih
jedinjenja koje bakterije i gljivice razlaţu i obrazuju amonijak (Jemcev and Đukić., 2000).
Mikrobiološka fiksacija azota je veoma bitna, pošto drugi organizmi ne mogu da
fiskiraju elementarni azot iz atmosfere. On je kao takav nedostupan za njih i zbog toga je
veoma bitno postojanje azotofiksatora. Azotofiksatori imaju sposobnost da elementarni azot iz
vazduha ukljuĉuju u svoj azotni metabolizam. Taj proces katalizuje enzimski kompleks
nitrogenaza. Za aktivnost ovog kompleksa neophodna je odreĊena koliĉina energije, ATPa.
Elementarni azot se pomoću ovog enzima redukuje do amonijaka. Amonijak se dalje koristi
za sintezu aminokiselina. Azotofiksatori su uglavnom bakterije, modrozelene alge,
aktinomicete i neke gljive. Ovi organizminastanjuju sva zemljišta i sve vodene sredine
(Jemcev and Đukić., 2000).
Postoji više grupa azotofiksatora:
- Slobodni azotofiksatori, ţive u zemljištu ili rizosferi biljaka, ali nisu vezani za
biljke i ne izazivaju morfološke promene. Slobodni azotifiksatori iz bakterijskih

14
rodova su Azotobacter, Derxia, Azomonas i Clostridium, a tu se svrstavaju i
Cianobacteria.
- Asocijativni azotofiksatori, vezani su za biljke, ali ne izazivaju morfološke
promene. Oni predstavljaju prelaz izmeĊu simbiotske i slobodne azotofiksacije.
Predstavnici ove grupe azotofiksatora su Azospirillum, Pseudomonas, Enterobacter.
- Simbiotski azotofiksatori, vezani su za biljke i izazivaju promene na biljkama. Ţive
na korenu leguminoznih biljaka i tu formiraju nodule, odnosno kvrţice. Simbioza
izmeĊu njih i biljaka je genetski odreĊena i specifiĉna. U grupu slobodnih
azotofiksatora spadaju Rhizobium i Bradyrhizobium (Jemcev and Đukić., 2000).
Od ukupnog biološkog azota, 80% proizvedu slobodni azotofiksatori. Koliĉina takvog
azota po jedinici površine je znatno manja nego što mogu da proizvedu simbiotski
azotofiksatori, ali je ipak znaĉajna, jer ĉak i bez Ċubrenja usevi mogu da daju 30% prinosa u
odnosu na poĊubrenu varijantu.
Od asocijativnih azotofiksatora najvaţniji su: Azotobacter chroooccum, Azotobacter
vinelandii, Azospirillum lipoferum, Beijerinckia derxii, Klebsiella planticola, Bacillus
subtillis, Bacillus megaterium, Pseudomonas sp., Clostridium i dr. Ovo su heterotrofni
organizmi kojih ima svuda u zemljištu, ali je vaţna njihova odlika da imaju dobar
interakcijski odnos sa biljkama zato što najbolje uslove za ţivot imaju u njihovoj rizosferi, pri
ĉemu od biljke dobijaju energiju u vidu šećera koje biljka izluĉuje na aktivnom delu
korenovog sistema, a zauzvrat joj daju fiksirani azot u obliku vezane forme NH,
hidroksilamina, aminokiselina, peptida, hormona rasta, biofungicida itd.
Osim fiksacije atmosferskog azota, većina ovih mikroorganizama je sposobna za
izvlaĉenje fosfora, kalijuma ili nekog drugog elementa iz kristalne rešetke minerala u
zemljištu i da ga stave u vodeni rastvor pristupaĉan biljkama. U okviru jedne vrste bakterija
postoji mogućnost pojave sojeva, koji su posebno prilagoĊeni na jednu biljnu vrstu ili ĉak
sortu, tako da njihovim izdvajanjem i umnoţavanjem, zatim inokulacijom semena ili
predsetvenom inkorporacijom u zemljište, moţe da se postigne povećanje prinosa 10-20%
(Jemcev and Đukić, 2000).
Kod jednogodišnjih biljaka najbolji rezultati su postignuti inokulacijom semena, pri
ĉemu bakterije prate rast mladih korenĉića, a kod povrtarskih vrsta i voća bolje je namoĉiti
korenov sistem rasada i sadnica u teĉnu kulturu azotofiksatora. Istovremeno, treba smanjiti
unos azotnih Ċubriva za 30-40%.
Pošto su ovi mikroorganizmi jako osetljivi na direktnu svetlost, taĉnije, ultravioletni
zraci su smrtonosni za njih, podrazumeva se da se tretiranje biljnog materijala vrši u senci.

15
Ima mnogo gotovih preparata sa bakterijama azotofiksatorima koji se mogu kupiti, a
moguće je vrlo lako u domaćoj radinosti napraviti mikrobiološko Ċubrivo. Obiĉno se uzima
plodna baštenska zemlja u kojoj sigurno ima Azotobactera. Sitna i prosejana zemlja se meša
sa 2-5% nekog izvora ugljenih hidrata (brašno, melasa, šećer,...), uz to se dodaje voda da
smesa doĊe u stanje lepljive plastiĉne mase, koja se u tankom sloju 1-2,5 cm razmazuje u
posudama i ostavlja na toplom. Za 3-4 dana celu masu prekrije i proţme sluzava masa od
ćelija Azotobaktera, koja se potom rastvara u vodi za nanošenje na biljni material (Radić.,
2015).
Fiksacija azota je redukcioni proces u kome se gas azota iz vazduha prevodi u
amonijak. Sve biljke imaju potrebu za azotom u obliku amonijaka ili nitrata koji im je
potreban za sintezu aminokiselina. Od tih aminokiselina one izgraĊuju svoje proteine, a biljni
proteini predstavljaju jedini izvor proteina za ţivotinje. Zato je fiksacija azota vitalan ekološki
proces. Organizmi koji mogu da obavljaju ovu vrstu redukcionih reakcija su neke vrste
bakterija i cijanobakterija (modrozelene alge). Celokupan ţivot na planeti zasnovan je na
zavisnosti od ovih organizama. (Jemcev and Đukić., 2000).
Azot koji se fiksira u obliku amonijaka ili amonijuma je osnovni materijal za
hemoautotrofne nitrifikacione bakterije.
Fiksacija azota je sloţen proces koji predstavlja seriju redukcionih reakcija. Atomi
azota su meĊusobno povezani trostrukom kovalentnom vezom unutar molekula. Zato je azot
stabilan molekul. Ovu vezu mogu da redukuju samo azotofiksirajuće bakterije i
cijanobakterije koje poseduju heterociste.
Heterociste predstavljaju nefotosintetiĉke ćelije u kojima se vrši fiksacija azota. One
koriste fotosistem i pri tome obezbeĊuju energiju koja je potrebna za fiksaciju azota. Time
gube deo procesa svetle faze fotosinteze gde dolazi do oslobaĊanja O2 (Jemcev and Đukić.,
2000.).
Na osnovu analize azotofiksatorskih fotosintetiĉkih cijanobakterija moţemo da
zakljuĉimo da su to organizmi koji su sami sebi dovoljni. Potrebni su im samo ugljen-dioksid
i neke organske materije, a sve ostalo mogu sami sebi da obezbede. Zato su ovi oblici
pionirske zajednice, jer uspevaju da kolonizuju nova staništa.
Za proces fiksacije azota neophodni su ATP i enzim nitrogenaza. Trostruka
kovalentna veza koja se javlja u atomu azota je veoma stabilna i potrebna je velika koliĉina
energije kako bi se ona raskinula. Nitrogeneza je neophodna za ovaj proces, ali se ona brzo
inaktiviše prisustvom molekularnog kiseonika i ne moţe da funkcioniše u ćeliji gde dolazi do
proizvodnje kiseonika u toku fotosinteze. Zbog toga se fiksacija azota kod cijanobakterija vrši
u heterocistama.

16
Dosta su prouĉavane bakterije koje vrše fiksaciju azota, pogotovo zbog njihovog
znaĉaja u poljoprivredi. Biljke mahunarke (pasulj, grašak, detelina) su znaĉajne kulture u
rotiranju useva, jer poboljšavaju plodnost zemljišta. To je zbog toga, jer na ĉvorićima korena
ovih biljaka ţive bakterije koje koriste šećere. Te šećere biljke proizvode u procesu
fotosinteze, a one ih zauzvrat snabdevaju amonijakom. Ovoj grupi bakterija pripada rod
Rhizobium (Ahemad et al., 2014).
Sve više i više se ovaj fenomen koristi kako bi se smanjila upotreba azotnih Ċubriva, a
poboljšao kvalitet proteina ovih kultura. Ova oblast istraţivanja ukljuĉuje selekciju,
mikrobiologiju i genetiĉki inţenjering. U daljem radu biće opisani rodovi odreĊenih vrsta
bakterija i neke od metoda koje omogućavaju fiksaciju azota, sa ciljem smanjenja upotrebe
veštaĉkih Ċubriva.
Ciklus kruţenja azota (Slika 2.) poĉinje kada azotofiksatori apsorbuju atmosferski azot
i vezuju ga u svom telu u nekim organskim jedinjenjima azota (aminokiselinama). Bakterije iz
roda Rhizobium isto vrše fiksaciju azota, u simbiozi sa korenom leguminoznih biljaka. Kada
doĊe do truljenja tela ovih azotofiksatora, njihove aminokiseline se pretvaraju u amonijak,
koji se pod uticajem drugih vrsta bakterija – nitrifikacionih bakterija, pretvara u nitrate, koji
su dostupni biljkama (Jemcev and Đukić., 2000).
Amonifikatori su mikroorganizmi koji mogu razliĉitim putevima dezaminacije
polipeptida i aminokiselina da oslobaĊaju amonijak. To je mikrobiološki proces koji dovodi
do oslobaĊanja azota iz organskih materija u formi amonijaka i oznaĉava se kao
amonifikacija. Amonifikaciju vrše razliĉiti mikroorganizmi koji mogu biti aerobi, anaerobi,
fakultativni anaerobi, sporogeni, asporogeni. U zemljištu, u kome vladaju neutralni i slabo
alkalni uslovi, glavni razlagaĉi proteina su bakterije iz roda Bacillus. To su sporogene vrste i
tu spadaju Bacillus megaterium, Bacillus subtilis, Bacillus cereus. Od asporogenih vrsta to su
Pseudomonas fluorescenes, Escherihia coli (Mihajilov – Krstev., 2008).
Ukoliko su zemljišta kisela, razlaganje proteina vrše gljive iz rodova Aspergillus,
Penicillium, Mucor, Rhizopus.
Nitrifikacija je sloţen proces, koji obuhvata dva sukcesivna procesa: nitritaciju i
nitrataciju. Bakterije koje vrše oksidaciju amonijaĉnih soli do nitrita i nitrata se oznaĉavaju
kao nitrifikatori. Nitritacija je proces oksidacije NH3+ do NO2
-i to obavljaju nitritacione
bakterije. Nitratacija je proces oksidacije NO2- do NO3
-i u tom procesu uĉestvuju nitratacione
bakterije. Procesi nitritacije i nitratacije su meĊusobno povezani. Odvijaju se istovremeno i u
zemljištu. Nitrifikatori predstavljaju obligatne aerobne mikroorganizme i pripadaju grupi
hemoautotrofa. Oksidaciju amonijaka do nitrita vrše Nitrosomonas europea,
Nitrosococusmobilis, Nitrosospira briensis i to je proces nitritacije. Proces nitratacije,

17
odnosno oksidaciju nitrita do nitrata vrši Nitrobacter winogradsky, Nitrospira gracilis
(Mihajilov – Krstev., 2008).
Heterotrofni mikroorganizmi koji u anaerobnim uslovima redukuju soli azotne kiseline
do gasovitih proizvoda (N2, N2O) su denitrifikatori. Do redukcije nitrata dolazi, jer su u
anaerobnim uslovima, akceptori soli azotne kiseline - nitrati, dok je u aerobnim uslovima,
akceptor vodonika i elektrona kiseonik. Redukcija nitrata se odvija u dva stupnja, nitrat se
redukuje u nitrit, a nitrit se zatim redukuje u amonijumov jon. Za ove procese najbitniji su
enzimi nitratna i nitritna reduktaza. Redukcija nitrata se moţe odvijati u korenu, gde se
izgraĊuju aminokiseline i amidi. Oni se dalje transportuju putem ksilema. Nitratni jon se moţe
transportovati u listove, gde se vrši sinteza i redukcija organskih jedinjenja. Tada se
aminokiseline floemom vraćaju u koren. Redukciju nitrata u nitrit vrši enzim nitratna
reduktaza u citosolu ćelija korena i lista. Redukciju nitrita u amonijum vrši enzim nitritna
reduktaza, koja je u plastidima.
Slika 2. Ciklus azota
Azot se ugraĊuje u organska jedinjenja samo u vidu amonijumovih jona. Biljka prima
amonijumov jon iz spoljašnje sredine ili on nastaje redukcijom nitrata u korenu ili listu.
Amonijum ulazi u sastav aminokiselina, a zatim i u druga jedinjenja. Biosinteza
aminokiselina je zavisna od fotosinteze koja obezbeĊuje sve ove uslove. U korenu, u
njegovim ćelijama, obavljaju se procesi sinteze aminokiselina i amida. Deo produkata se
koristi u samom korenu, a ostatak se prenosi u nadzemni deo biljke, putem ksilema (Mihajilov
– Krstev., 2008).

18
2.6. CIKLUS SUMPORA
Sumpor ima veoma veliki biološki znaĉaj. On predstavlja strukturnu komponentu u
izgradnji nekih aminokiselina (cistein, metionin), belanĉevina, kao i enzima. Sumpor se nalazi
u zemljištu u obliku mineralnih soli i organskih jedinjenja. Ulazak sumpora u ţive sisteme je
u vidu sulfata i sulfida, koje biljke apsorbuju iz zemljišta. Nakon toga, oni podleţu redukciji i
ugraĊuju se u aminokiseline, proteine (Jemcev and Đukić., 2000).
Sumpor se u atmosferi nalazi u vidu SO2, SO3 i H2S, koji nastaju sagorevanjem
fosilnih goriva. Pri sagorevanju fosilnih goriva oslobaĊaju se najveće koliĉine sumpora, u
obliku SO2. Danas je to jedan od najvećih zagaĊivaĉa vazduha. Ovi gasovi sa kišom padaju na
zemlju u vidu sulfita. Deo SO2 se transformiše do sumporne kiseline i javlja u obliku kiselih
kiša. U ciklus kruţenja (Slika 3.) sumpora ukljuĉene su i neke autotrofne, sumporne
bakterije, kao što je rod Begiatoa. Sumpor iz zemljišta se prevodi u sulfate, a zatim sintezom
proteina se pretvara u organski vezan sumpor. Najĉešći oblik sumpora koji biljke koriste je
sulfatni jon. Biološka redukcija sulfata je analogna redukciji nitrata i samo su biljke sposobne
da je obavljaju. Ipak, glavni izvor sumpornih jedinjenja u zemljištu su organske materije, koje
su poreklom od tela uginulih biljaka i ţivotinja. Usled aktivnosti razliĉitih mikoorganizama,
sumporna jedinjenja se cikliĉno oksiduju i redukuju. U kruţenju sumpora moţe doći i do
digresija putem njegove sedimentacije u obliku nerastvorljivih soli fero-sulfata (FeS), feri-
sulfata (Fe2S3) i kalcijum-sulfata (CaSO4). Tako dolazi do blokiranja gvoţĊa i ostalih
nutritivnih sastojaka koji su znaĉajni za biološke sisteme (bakar, cink,). Ukoliko doĊe do
odstupanja od normalnih koliĉina odreĊenog elementa, to se manifestvuje poremećajima u
kruţenju materije (Jemcev and Đukić., 2000).
Slika 3. Ciklus sumpora

19
2.7. CIKLUS FOSFORA
Fosfor je znaĉajan elemenat i javlja se u zemljištu u razliĉitim organskim i
neorganskim oblicima. Na osnovu dostupnosti ţivom svetu, najvaţniji oblik fosfora je
ortofosfat, koji nastaje oksidacijom u procesima mineralizacije organskih materija koje sadrţe
fosfor. Fosfor je takoĊe strukturna komponenta ţivih sistema. Uĉestvuje u graĊi ćelijskih
jedinjenja, nukleinskih kiselina, fosfolipida, jedinjenja bogata energijom (ATP, ADP, AMP).
Biljke apsorbuju fosfor u obliku ortofosfata i zatim se prenosi do konzumenata i razlagaĉa u
obliku organskog fosfora. Nakon mineralizacije, opet dolazi do biljaka (Slika 4.) (Jemcev and
Đukić., 2000).
Fosfor se u zemljištu nalazi u obliku fosfata, koji potiĉu od razliĉitih organskih
jedinjenja. Biljke najbolje primaju jednovalentni fosfat i to je pristupaĉni fosfat. Fosfat se
ugraĊuje u organska jedinjenja bez prethodne redukcije. Prvi stupanj usvajanja fosfata u
biljnim ćelijama je njegovo vezivanje ze šećer inozit pri ĉemu nastaje fitinska kiselina. Pravi
ulazak fosfora u organska jedinjenja koja imaju metaboliĉku funkciju je sinteza ATP, koja se
odvija u hloroplastima, mitohondrijama, u glikolizi. Sa ATPa se fosfat prenosi na mnoga
druga jedinjenja. Sve veze koje gradi ATP oslobaĊaju visoku energiju pri hidrolizi i one su
najvaţniji naĉin transformacije iprenosa energije u ţivoj ćeliji (Jemcev and Đukić., 2000).
Slika 4. Ciklus fosfora

20
2. 8. ĐUBRIVA
Na osnovu svega navedenog, znamo da su svi do sada opisani elementi neophodni
biljkama, a samim tim su neophodni i za nastavak ţivota, jer bi bez njih to bilo nemoguće.
Kako bi se poboljšao kvalitet zemljišta i biljkama povećala dostupnost svih hranljivih
materija, u poljoprivredi se koriste azotna Ċubriva. Azotna Ċubriva predstavljaju hranu za
biljke, odnosno ona pojaĉavaju mobilizaciju hranljivih materija u zemljištu i povećavaju
energiju ţivotnih procesa u njemu, a samim tim i menjaju svojstva zemljišta (Đukić et al.,
2007). Postoje dve osnovne grupe Ċubriva, na osnovu njihovog hemijskog sastava i to su:
- Organska Ċubriva (stajnjak, gnojivo, kokošiji izmet, treset, siderati, slama, smeće,
pleva, talog otpadnih voda, kao i bakterijska Ċubriva) i
- Mineralna Ċubriva, mogu biti jednostavna (azotna, fosforna, sumporna, kalijumova,
magnezijumova..) i kompleksna ( sloţena – Nitrofos; sloţeno-mešovita; mešovite
smese – NPK, NK).
Postoje industrijska i mesna Ċubriva, u zavisnosti od porekla i mesta njihovog
dobijanja (Đukić et al., 2007).
2.8.1. MINERALNA ĐUBRIVA
Mineralna Ċubriva koja se proizvode industrijski su zastupljena u veoma velikom
broju. Njihova široka primena u poljoprivredi uslovljena je slabom zastupljenošću zemljišta
elementima ishrane. To se posebno odnosi na zastupljenost azota. Od azotnih Ċubriva
najĉešće se koriste amonijaĉno – nitratna (amonijaĉna i kreĉno-amonijaĉna šalitra),
amonijaĉna (amonijum – sulfat, amonijum hlorid, amonijum – sulfat), nitratna (natrijumova i
kalcijumova šalitra), amidna (karbamid, urea – formaldehid). U pokušaju je dobijanje azotnih
mineralnih Ċubriva koje sporo odaju azot biljkama, odnosno Ċubriva sa povećanom
otpornošću. To su jedinjenja koja se slabo rastvaraju u vodi ili su pokrivena materijama, koje
oteţavaju brzo rastvaranje jedinjenja azota. Koliĉina dostupnog azota biljkama, u zemljištu,
je ograniĉena i odreĊena uslovima zemljišta. Njegove zalihe uglavnom ne prelaze 4%
ukupnog azota. Kako bi postale pristupaĉne biljkama, azotne materije prolaze sloţene
biohemijske transformacije, da bi se dobili amonijaĉni i nitratni azot, kojima se biljke hrane.
Azotna mineralna Ċubriva utiĉu na poboljšavanje strukture zemljišta, doprinose smanjenju
vode i slabljenju erozije. Koriste se u odreĊenim dozama, mada postoji veliki broj ĉinjenica
koje ukazuju na negativni uticaj mineralnih Ċubriva, koja se primenjuju u visokim dozama i
duţem vremenskom periodu. Kada je zemljište bogato humusom, mineralna Ċubriva

21
ubrzavaju proces mineralizacije organske materije, pa to moţe dovesti i do gubitka humusa.
Unošenjem mineralnog Ċubriva povećava se sadrţaj azota po ĉitavom zemljišnom profilu.
MeĊutim, povećanjem sadrţaja nitrata u zemlji, dolazi do zagaĊenja podezemnih voda, što
utiće na njihov ţivi svet. Mineralna Ċubriva dovode i do smanjenja pH vrednosti, pri ĉemu
slabi puferna sposobnost zemljišta. To dovodi do gubitka nekih elemenata (kalcijuma,
kalijuma, magnezijuma), aktivacije i mobilizacije gvoţĊa, mangana, vezivanja fosfora i
povećanje toksiĉnog dejstva teških metala. Nekada se smatralo da se korišćenjem mineralnih
Ċubriva povećava brojnost i biomasa zemljišnih mikroorganizama, ali danas je dokazano
suprotno. Korišćenje mineralnih azotnih Ċubriva ima negativni uticaj na zemljišne mikrobe,
gde se, pri njihovoj duţoj upotrebi smanjuje njihova brojnost i biomasa. Do toga dolazi zbog
promena aktivne reakcije zemljišnog rastvora i povećanja sadrţaja mobilnog aluminijuma.
Primena ovih Ċubriva dovodi i do poremećaja u biogeohemijskim ciklusima hranljivih
materija. Dolazi do poremećaja prirodne ravnoteţe zemljišnih mikroorganizama koji su
odgovorni za procese azotofiksacije, nitrifikacije, humifikacije. Veoma bitna pojava, koja se
javlja usled dugotrajnog unošenja azotnih mineralnih Ċubriva, jeste stvaranje ekstra – azota.
Njegova koliĉina raste još većim dodavanjem azotnog Ċubriva. Stvaranje ovog ekstra – azota
praćeno je gubitkom organskog ugljenika, pa se zato zemljištu dodaje i organik, gde se na
odreĊenu koliĉinu azotnog Ċubriva, zemljištu dodaje i odreĊena koliĉina organskog ugljenika
(5kg organskog ugljenika na 1kg unešenog azota). Primenom mineralnih Ċubriva menja se i
struktura zemljišnih organizama, dolazi do smanjenja brojnosti bakterija, ali raste broj bacila
u toj cenozi, pogotovu Bacillus mesentericus, kao posledica osiromašivanja zemljišta
organskim jedinjenjima. Specifiĉno je to da se brojnost Bacillus mycoides takoĊe smanjuje.
Primećeno je da se pod uticajem NPK povećava sadrţaj gljiva u cenozi, zbog promene pH
zemljišta. Korišćenjem velikih doza mineralnih Ċubriva smanjuje se koliĉina cijanobakterija,
pa se azotofiksatorske vrste zamenjuju drugim vrstama. Smatra se da mineralna Ċubriva
povećavaju periodiĉnu toksiĉnost zemljišta, verovatno nakupljanjem odreĊenih bakterija
(Đukić et al., 2007).
Na osnovu analize mineralnih azotnih Ċubriva (nitratnih, amonijaĉnih i amidnih)
utvrĊeno je njihovo negativno dejstvo. Oni smanjuju proces disanja u zemljištu,
azotofiksirajuću aktivnost i antibiotiĉki potencijal zemljišta, pri ĉemu stimulišu razvoj
patogena. Smatra se da su nitratna Ċubriva najmanje štetna, odnosno da su posle amidnih i
amonijaĉnih, jer najmanje utiĉu na procese azotofiksacije i denitrifikacije. MeĊutim, nitratna
Ċubriva takoĊe smanjuju brojnost odreĊenih mikoorganizama u zemljištu, što je i dokazano
odreĊenim metodama (Đukić et al., 2007).

22
Azot se u mnogim poljoprivrednim oblastima nalazi u veoma malim koliĉinama, pa
zbog toga azotna Ċubriva imaju vaţnu ulogu, jer povećavaju prinos poljoprivrednih kultura.
Neophodan uslov za njihovu primenu je kombinovanje sa fosfornim, kalijumovim i
mikroĊubrivima, kako bi se ona efikasno iskoristila. Od unete doze azotnih Ċubriva odreĊenu
koliĉinu iskorišćavaju zemljišni mikroorganizmi, dok jedan deo ĉine nepovratni gasoviti
gubici. Ti gubici su osnovni uzrok smanjenja iskorišćavanja azota od strane biljaka, što samim
tim, smanjuje i njegovu efikasnost. Nakon unošenja mineralnih Ċubriva poĉinje njihova
fiziĉka, hemijska i mikrobiološka transformacija. Gasoviti gubici azota povezani su sa
aktivnošću denitrifikacionih bakterija koje vrše redukciju nitratnog azota. Ovi gasoviti
proizvodi denitrifikacije, pored toga što smanjuju efikasnost azotnih Ċubriva, oni su i
ekološka opasnost, jer uĉestvuju u razgradnji ozonskog omotaĉa stratosfere, zbog oksdidacije
N2O i NO. Kako bi se zaštitila okolna ţivotna sredina i povećala efikasnost azotnih Ċubriva,
prouĉavani su mnogi naĉini smanjenja gubitka azota u gasovitom sadrţaju i smanjenje
sadrţaja oksida azota. Jedan od naĉina je primenom preparata, koji ograniĉavaju
razmnoţavanje denitrifikacionih bakterija, ali oni još uvek nisu razraĊeni. Zbog toga se
koriste preparati koji ograniĉavaju aktivnost nitrifikacionih bakterija. Jedan odtakvih
preparata je N-serve koji se koristi za smanjenje gubitka amonijaĉnih i amidnih oblika azotnih
Ċubriva i njegovo dejstvo se ispoljava i inhibiciji razvoja Nitrosomonas, koji je izazivaĉ prve
faze nitrifikacije. Bitna osobina ovog inhibitora je to što je male toksiĉnosti, brzo se razlaţe u
zemljištu. Dokazano je da se korišćenjem ovog inhibitora povećao rast mnogih kultura
(pšenice, kukuruza) (Đukić et al., 2007). Mnogi inhibitori koji se danas koriste utiĉu na
smanjenje razmnoţavanja nitrifikacionih bakterija, ali i denitrifikacionih bakterija. Na dejstvo
inhibitora nitrifikacije manje su osetljive amonifikacione bakterije. Njihovo razmnoţavanje je
smanjeno samo prvih 10 – 20 dana nakon unošenja preparata. Inhibicija razmnoţavanja
nitrifikacionih bakterija omogućava ĉuvanje amonijaĉnog oblika azota iz Ċubriva, pri ĉemu se
gasoviti oblici azota smanjuju. Nakon privremenog zaustavljanja razmnoţavanja zemljišnih
mikroorganizama, dolazi do fiksiranja azota Ċubriva u zemljištu, a i do jaĉanja procesa
mobilizacije azota zemljišta. Tako, pri unošenju inhibitora dolazi do povećanog usvajanja
azota od strane biljaka. Kod kultura jeĉma, ovi inhibitori nemaju uticaj na njihov prinos. Na
osnovu toga, utvrĊeno je da unošenje inhibitora ne mora dovesti do povećanja prinosa, ali ima
pozitivno dejstvo na bilans azota iz Ċubriva, na njegovo fiksiranje u zemljištu, smanjenje
gasovitih gubitaka i na mobilizaciju zemljišnog azota (Đukić et al., 2007).
Veoma je vaţno da se azotna Ċubriva unose u odreĊeno vreme, odnosno u periodu
kada biljke mogu da iskorišćavaju hranljive elemente. To povećava njihovu efikasnost
korišćenja i smanjuje gubitke. Uglavnom se azotna Ċubriva unose na jesen ili proleće.

23
Ukoliko se unose na jesen dolazi do znaĉajnih gasovitih gubitaka azota, jer su u tom periodu
usporeni mikrobiološki procesi, kao i proces denitrifikacije azota unesenih Ċubriva (Đukić et
al., 2007).
2.8.2. ORGANSKA ĐUBRIVA
U grupu organskih Ċubriva spadaju stajnjak, gnojivo, treset, izmet, komunalni otpad,
komposti i mnogi drugi. Oni predstavljaju izvore energije i mineralnih elemenata, a njihova
uloga je i u intenziviranju mikrobioloških procesa.
Stajnjak ĉini 20 – 25 % organske materije (azot, fosfor, kalijum) i ostatak predstavlja
voda. Industrijskim stoĉarenjem dobija se stajnjak koji sadrţi oko 90% vode. Postoje ĉetiri
naĉina ĉuvanja stajnjaka i to je: štalsko ĉuvanje (pod stokom), ĉuvanje na Ċubrištu, priprema
hladnog stajnjaka (ubrzanim sabijanjem) i priprema toplog stajnjaka (povremenim
rastresanjem i sabijanjem). Đubrivo viskog kvaliteta dobija se ĉuvanjem stajnjaka pod
stokom, odnosno štalskim ĉuvanjem, ali zbog antisanitarnih uslova se najmanje koristi. U
sveţem stajnjaku nalazi se veliki broj mikoorganizama, ĉiji je razvoj specifiĉan u zavisnosti
od njegovog ĉuvanja. U sazrevanju hladnog stajnjaka najveću ulogu imaju asporogene
bakterije. U sveţem stajnjaku najzastupljenije su koke predstavljene amonifikatorima koje
zapoĉinju proces truljenja. Bakterije iz roda Pseudomonas su dosta zastupljene u stajnjaku, ali
se nalaze i truleţne sporogene bakterije Bacillus subtilis, Bacillus megaterium, Bacillus
mycoides. Mnoge amonifikacione bakterije iz stajnjaka mogu da dovedu do razlaganja uree. U
njemu se nalaze i vrste koje razlaţu celulozu, zatim nitrifikatori, koji oksidišu amonijak i
razlaţu purinske baze iz stajnjaka. U nastanku stajnjaka menja se zastupljenost materija, oko
40% azota stajnjaka nalazi se u vidu hipurne i mokraćne kiseline, ali je najveći deo u vidu
uree koju hidrolizuju urobakterije i saprofitne bakterije i time se stvara amonijum – karbonat,
koji zatim disocira na amonijum i ugljendioksid (Đukić et al., 2007).
Slama se koristi kao organsko Ċubrivo i njenim unošenjem zemljište se obogaćuje
humusom, ali ona sadrţi i elemente potrebne biljkama, azot i druge. Razlaganjem slame
oslobaĊa se ugljendioksid, što povoljno utiĉe na useve. U slami je odnos ugljenika i azota
veliki, odnosno 100 : 1, pa njenim dodavanjem biljke pate od nedostatka azota, izuzetak su
leguminoze. Nedostatak azota bi se prevazišao dodavanjem azotnih Ċubriva. Smatra se da
slama sadrţi i neke toksiĉne materije za biljke, ali adekvatnom primenom i obradom zemljišta
to bi se prevazilazilo (Đukić et al., 2007).
Treset se takoĊe moţe primenjivati kao organsko Ċubrivo. On sadrţi veliku koliĉinu
vode, organskih materija, od kojih su najzastupljenija humusne supstance i supstance sliĉne

24
ligninu. Uglavnom se primenjuje nizijski treset, ali i on sadrţi malu koliĉinu azota, koji se i
sporo mineralizuje (Đukić et al., 2007).
Kompost, kako bi se dobila organska Ċubriva i time smanjilo korišćenje mineralnih
Ċubriva i hemijskih sredstava za zaštitu biljaka, primenjuje se metod kompostiranja razliĉitih
organskih otpada. Taj otpad moţe biti gradsko smeće, koje predstavlja smešu organskih i
neorganskih materija, biljni otpad (mulj). U kompostiranju se organski supstrat podvrgava
aerobnoj biodegradaciji sa mikrobima u uslovima povećane temperature i vlaţnosti. Ovim
procesom, organska komponenta trpi fiziĉke i hemijske transformacije, pri ĉemu se na kraju
stvara stabilni humikovani proizvod - kompost, koji ima veliku koliĉinu organskih jedinjenja,
biomasu uginulih mikroorganzama, odreĊenu koliĉinu ţivih organizama. Kompostiranje
predstavlja dinamiĉan mikrobiološki proces u kome uĉestvuju mikrobi (bakterije, gljive, alge,
virusi), mikrofauna (praţivotinje, protozoe), makrofauna (stonoge, krpelji, crvi) (Đukić et al.,
2007).
Ostvarivanje maksimalne produktivnosti i profita u poljoprivredi podrazumeva
intenzivnu obradu zemljišta, navodnjavanje, adekvatnu primenu Ċubriva, hemijsku kontrolu
štetoĉina, korova i bolesti, kao i maksimalnog iskorišćavanja genetskog potencijala gajenih
biljaka. Sve to dovodi do mnogih negativnih posledicana agroekosistem i kvalitet ţivota
sredine. Brz razvoj biotehnologije, zasnovan na ekološkim principima, utiĉe da tradicionalni
sistemi zemljoradnje doţive brojne promene.
Rezultati mnogih nauĉnih istraţivanja pokazali su da Ċubriva hemijskog porekla zbog
sadrţaja toksiĉnih materija, mogu da se dovedu u vezu sa neţeljenim efektima koje
ispoljavaju na zdravlje ljudi, reprodukciju, probleme sa krvlju i jetrom kao i na pojavu
hiperaktivnosti dece. Nikako se ne moţe dozvoliti da se zanemari negativan uticaj koji
prekomerna upotreba ovih Ċubriva ima na ţivotnu sredinu (Đukić et al., 2007). Razvoj
poljoprivrede je zasnovan na promenama u tehnologiji gajenja useva i oplemenjivanju
biljaka. Biljka je ţivo biće koje ţivi u dve sredine, atmosferi i pedosferi. Zemljište sluţi
biljkama za uzimanje vode, hranljivih materija i energije, koji se nalaze u njemu. Ako je
zemljište nestrukturno, bez dovoljno vode i hranljivih materija onda je broj korisnih mikroba i
zreo humus u zemljištu, osnov za planiranje dobrog i kvalitetnog prinosa. To doprinosi boljem
uspostavljanju ekološke ravnoteţe i stabilnosti prirodnih resursa. Zbog toga se uvodi novi
pojam, a samim tim i nova metoda – biofertilizacija (Singh., 2012).

25
2.9. MIKROBIOLOŠKA ĐUBRIVA I BIOFERTILIZACIJA
Biofertilizacija predstavlja unošenje ţivih mikroorganizama u zemljište sa ciljem
poboljšanja snabdevanja biljaka neophodnim nutritijentima. Tako se moţe poboljšati
snabdevanje biljaka azotom, fosforom, kalijumom, gvoţĊem, sumporom, ali i stimulisati rast
korena. Unošenjem ovih bakterija u rizosferu biljaka ubrzavaju se procesi transformacije
organske materije i biljka se snabdeva potrebnim nutritijentima (Singh., 2012).
Mikrobiološka Ċubriva predstavljaju ĉiste kulture aktivnih mikroorganizama koje se
gaje na hranljivoj podlozi. Nazivaju se i bakterijska Ċubriva. Njihova upotreba je
najzastupljenija u voćarstvu. Mikrobiološka Ċubriva svoj uticaj na biljke ispoljavaju kroz:
- ubrzanje procesa pretvaranja organske materije u forme koje su pogodne za
ishranu voćaka;
- poboljšanje snabdevanja voćaka azotom;
- oslobaĊanje elemenata iz teško pristupaĉnih formi, u forme koje voćke mogu lako
koristiti;
- smanjenje koncetracije CO2 u zemljištu;
- umanjenje mogućnosti asfikacije korena.
Mikrobiološka Ċubriva mogu se koristiti u razliĉitim oblicima: vlaţna, teĉna,
zamrznuta (granulirana) i suva (u prahu). U praksi se bakterijska Ċubriva primenjuju na
sledeće naĉine:
- potapanjem korena sadnice u suspenziji odreĊenog Ċubriva,
- zalivanjem voćaka rastvorom smeše ţivinskog i bakterocidnog Ċubriva.
Najĉešće se koriste sledeća mikrobiološka Ċubriva:
- Azotofiksatori, povećavaju koliĉinu azota u zemljištu. Đubriva mogu biti sa
slobodnim i simbiotskim azotofiksatorima, pa na osnovu toga imaju razliĉite
komercijalne nazive: Azotobakterin, Rizotrofin, Nitrogin, Azotofiksin i dr.
- Fosfobakterin-ova Ċubriva sadrţe bakterije koje mineralizuju organska jedinjenja
sa fosforom i pretvaraju ih u oblike lako dostupne korenovom sistemu biljaka.
- Mikorizin, sadrţi mikorizne gljive koje se mogu inokulirati na koren mladih
sadnica, neposredno pred sadnju. Mikorizin se sastoji od gljiva sposobnih da

26
potpomaţu ishranu voćaka (apsorpciju vode i hranljivih elemenata). Koristi se u
formi vlaţnog Ċubriva.
- Humivorin, unosi se u zemljište sa stajnjakom ili kompostom. Sastoji se od raznih
bakterija sposobnih da humus pretvaraju u mineralne materije pogodne za ishranu
biljaka.
- Silikobakterin, doprinosi oslobaĊanju kalijuma iz alumo silikata. Sojevi ovih
mikroorganizama proizvode veće kolićine ugljene kiseline i na taj naĉin rastvaraju
alumo silikate i druge minerale kalijuma.
Komponente mikroloških Ċubriva su uglavnom Azotobacter, Azospirillum, Bacillus,
Pseudomonas i druge. Pošto je azot veoma znaĉajan u ishrani biljaka, posebno mesto se
pridaje bakterijama koje snabdevaju biljke azotom. To su bakterije iz grupe azotofiksatora
koje elementarni, atmosferski azot prevode u oblike pristupaĉne biljkama. Ove bakterije se
nalaze u neposrednoj blizini korena i azot predaju neposredno biljkama.
Za rast biljaka neophodan je i fosfor, koji je prisutan u zemljištu u formama koje su
nepristupaĉne biljkama. Bakterije iz roda Bacillus i Azotobacter mogu da sintetišu organske
kiseline i fosfataze koje će nepristupaĉan fosfor prevesti u biljkama pristupaĉnu formu
(Gowathy et al., 2008).
Kalijum je u zemljištu u obliku alumosilikata. Zahvaljujući aktivnosti bakterija iz roda
Bacillus, postaje pristupaĉan biljkama. Neke bakterije zahvaljujući prisustvu siderofora
doprinose snabdevanje biljaka gvoţĊem. TakoĊe je poznato da bakterije iz roda Pseudomonas
mogu transformisati organske forme sumpora u neorganski i na taj naĉin ga uĉiniti
pristupaĉnim za biljke (Đukić et al., 2007).
Ovo ukazuje da se primenom mikrobioloških Ċubriva koja u sebi sadrţe mešane
populacije mikroorganizama moţe poboljšati snabdevanje biljaka neophodnim nutritijentima
uz istovremeno oĉuvanje ţivotne sredine i proizvodnju zdravstveno bezbedne hrane. Pored
toga ove bakterije imaju sposobnost sinteze biljnih hormona tipa giberalina, auksina, ĉime se
dodatno stimuliše biljni rast i utiĉe na otpornost biljaka. Unošenjem mikrobioloških Ċubriva u
zemljište utiĉe se na tok i usmeravanje mikrobioloških procesa u zemljištu što će uticati na
rast, razviće biljaka ali i na zemljište. Neki od mikroorganizama koji su uneti u zemljište
odlikuju se mogućnošću sinteze sluzavih materija koje igraju znaĉajnu ulogu u slepljivanju
mikroagregata što doprinosi formiranju fine strukture zemljišta. Nakon izumiranja
mikroorganizama unetih u zemljište povećava se ukupna biomasa, a efekti će se odraziti u
sledećoj vegetaciji. Povećanjem organske biomase dovešće do povećanja plodnosti zemljišta i
stvaranje biljkama neophodnih mineralnih nutritijenata. Pri tome ne dolazi do zagaĊivanja

27
zemljišta, voda i atmosfere. Azotobacter chroococum prisutan je u rizosferi mnogih biljaka, a
kod nekih biljnih vrsta pribliţava se asocijativnim azotofiksatorima. U tim uslovima
Azotobacter prevodi elementarni azot iz atmosfere u oblik koji je pristupaĉan biljkama.
Bacillus megaterium var. phosphaticus uĉestvuje u procesu amonifikacije. Fosfor na taj naĉin
dolazi u neorganski oblik, pristupaĉan za biljku. U zemljištu je prisutna silikatna bakterija
Bacillus circulans koja razlaţe alumosilikate iz kojih se oslobaĊa kalijum. Sve ovo upućuje da
primena mikrobioloških Ċubriva ima svoje mesto u savremenoj konvencionalnoj, ali i
organskoj poljoprivrednoj proizvodnji (Đukić et al., 2007).
Osnovu biofertilizatora ĉine bakterije koje direktnim ili indirektnim delovanjem
pozitivno utiĉu na rast biljke i porast useva, a vrlo se jednostavno nanose na površinu biljke,
semena ili zemlje. Reĉ je o bakterijskim vrstama koje pozitivno deluju na rast biljaka kroz
sposobnost:
- fiksacije atmosferskog azota;
- pretvaranje jedinjenja fosfora u oblike lako pristupaĉne za biljku;
- razlaţu organsku materiju;
- izluĉuju supstance antibiotskog i fungistatiĉkog delovanja i
- poseduju sposobnost produkcije hormona i drugih supstanci koji su neophodne za
rast i razviće biljaka (tzv. PGP Plant Growth Promoting bakterije). Hormoni koje
produkuju bakterije (auksini, citokinini, giberelini) su potpuno prirodni analozi
biljnim hormonima (Singh., 2012).
FOSFAT SOLUBILIZIRAJUĆI MIKROORGANIZMI (PSM)
Mikroorganizmi koji imaju sposobnost da konvertuju nedostupne u dostupne forme
jedinjenja koje korenov sistem biljaka moţe da usvoji, su solubilizatori. Fosfosolubilizatori
(PSM – Phosphorus Solubizing Microorganisms) su grupa bakterija i gljiva koje konvertuju
neorganski oblik fosfora u organski. Ovaj mehanizam se odvija zahvaljujući sekreciji
organskih kiselina (glukonska, mleĉna, limunska, oksalna, fumarna), u spoljašnju okolinu
mikroorganizama, što je rezultat razgradnje šećea. Sniţenje pH sredine dovodi do toga da
anjonska forma – kiselina, vezuje katjon neorganskog oblika fosfata i tako konvertuje
nedostupna fosfatna jedinjenja u rastvorljive forme, dostupne korenovom sistemu. Proces
zavisi od vrste mikroorganizama i kiseline koju stvara. Najefikasniji put je ekstracelularna
oksidaza glukoze, posredstvom enzima glukozna dehidrogenaza, gde se stvara glukonska
kiselina. Alifatiĉna kiselina (glukonska kiselina, 2-ketoglukonska kiselina) se pokazala
najefikasnijom (Richardson andSimpson., 2011).

28
Ovde spadaju vrste iz roda: Pseudomonas, Bacillus, Micrococcus, Penicillium,
Fusarium i dr.
FOSFAT MOBILIZIRAJUĆI ORGANIZMI (PMB)
U ovu grupu spadaju mikroorganizmi sa sposobnošću mobilizacije nedostupnih formi
jedinjenja fosfora, cinka i sumpora, ĉineći ih dostupnim za biljke. Posebno se izdvajaju
mikorizalne gljive – fosofomobilišuće gljive koje stupaju u mutualistiĉki odnos sa biljkama.
Hife povećavaju apsorpcionu površinu korena što je od velikog znaĉaja ukoliko je neki od
nutritijenata u manjku. Mikorizalne gljive se dele na:
- ektomikorize,
- endomikorize (AM – arbuskularne mikorize); Glomus, Gigaspora, Sclerocystis...
- ektendomikorize (Richardson andSimpson., 2011)
Bakterija Frateuria aurentia je poznata po sposobnosti mobilizacije kalijuma.
BILJNI HORMONI (FITOHORMONI)
Fitohormoni su organske supstance male molekulske mase, koje ne spadaju u hranljive
materije, a koje (u malim koliĉinama) stimulišu, inhibiraju ili na neki drugi naĉin modifikuju
rast i razvoj biljaka. To su prirodne supstance koje se izgraĊuju u biljnom organizmu koje
utiĉu na rastenje i razviće biljaka. Svi hormone deluju tokom celog razvića biljke na procese u
razliĉitim ćelijama i organima. Efekat jednog hormona na iste ćelije moţe biti razliĉit u
zavisnosti od stadijuma razvića ćelija. Fitohormoni obuhvataju nekoliko klasa jedinjenja,
meĊu kojima su auksini, giberelini, citokini i drugi (Kastori., 1991).
Auksini su prva grupa biljnih hormona koji su otkriveni na osnovu toga što stimulišu
rastenje biljaka. Po hemijskoj strukturi slavna grupa auksina su indolna jedinjenja, a centralno
mesto pripada indolil-3-sirćetnoj kiselini (IAA). Glavne funkcije auksina su:
- stimulativni efekat auksina na izduţivanje ćelija stabla i koleoptila (fototropska i
geotropska reakcija);
- u deobi ćelija;
- inhibicija mirujućih pupoljaka (Ortiz – Castro., 2009).

29
Giberelini su otkriveni u Japanu kao produkt gljive Ascomycetes. Biljke izgraĊuju
gibereline u tkivima koja aktivno rastu, kao što su apikalni delovi stabla, mladi listovi, koreni,
nezrela semena i plodovi. Fiziološki efekti giberelina su:
- rastenje ćelija u vegetativnim tkivima;
- izduţivanje stabla;
- razvoj listova i izduţivanje lisnih drški;
- diferencijacija floemskih elemenata;
- izduţivanje polenovih cevi;
- klijanje semena (Kastori., 1991).
Citokini su grupa biljnih hormona ĉija je osnovna funkcija da stimulišu ćelijske deobe,
ali su za to neophodni i auksini. Citokina ima najviše u vrhu korena, koji je dominantno mesto
njihove sinteze, odakle se transportuju ksilemom u ostale organe (stablo i listove). Glavne
funkcije citokina su:
- stimulacija ćelijskih deoba u prisustvu auksina;
- indukcija novih pupoljaka i stimulacija mirujućih pupoljaka;
- ukidanje apikalne dominacije;
- fascijacija i debljanje stabla;
- rastenje listova;
- diferencijacija hloroplasta;
- stimulacija cvetanja i regulacija pola cvetova;
- usmeravanje transporta i akumulacija asimilata i metabolita u organima bogatim
citokinima (Ortiz – Castro., 2011).
ANTIBIOTICI
Antibiotici su metaboliti jednih mikroorganizama, koji deluju inhibitorno ili ĉak
latentno na druge mikroorganizme. Antibiotike je prvi uoĉio Aleksandar Fleming 1929.
godine kada je primetio da gljiva Pencilium naotatum stvara zonu u kojoj ne dolazi do razvića
bakterija – zona inhibicije. Antibiotici se najĉešće dobijaju mikrobiološkim putem procesom
fermentacije, kao i polusintetskim i sintetskim putem (Kastori., 1991).
Biofertilizatori su organskog porekla i unapreĊuju primarnu poljoprivrednu
proizvodnju popravljajanjem fiziĉkih svojstava zemljišta na kome se koriste, njegovu
plodnost i produktivnost. Ĉinjenica da se njihovom upotrebom smanjuje koliĉina hemikalija
u poljoprivrednoj proizvodnji ĉini biofertilizaciju eko frendli procesom. Uticaj biofertlizatora

30
zavisi od soja bakterija, ali i od koncentracije bakterijskih ćelija u inokulatu biljne vrste,
primenjenih mera i osobina zemljišta (Singh., 2012).
Biofertilizatori kojima se pospešuje proces azotofiksacije su uglavnom bakterije iz
rodova Rhizobium, Bradyrhizobium, Sinorhizobium, Mesorhizobium, Allorhizobium,
Azorhizobium. Ovi mikroorganizmi uĉestvuju u simbiotskoj azotofiksaciji, prodiru u koren ili
stablo leguminoznih biljaka i formiraju kvrţice (nodule). Rodovi koji uĉestvuju u
asocijativnoj azotofiksaciji uglavnom ne formiraju kvrţice i koriste se pre setve na seme ili se
unose tokom vegetacije. U ovu grupu spadaju Acetobacter, Bacillus, Azrospirillum,
Pseudomonas. Sve ove vrste fiksiraju azot iz atmosfere i proizvode materije koje utiĉu na rast
biljaka (Singh., 2012).
Pored obezbeĊivanja azota, slobodni azotofiksatori produkuju i mnoga biološki
aktivna jedinjenja, organske kiseline koje razlaţu fosfate, aerišu zemljište, a i sintetišu mnoga
organska jedinjenja. Kod proizvodnje neleguminoznih biljaka i ostalih povrtarskih biljaka
uglavnom se koristi Azotobacter sp, a u proizvodnji pirinĉa se koriste razliĉite vrste
cijanobakterija (Singh., 2012).
2.9.1. ROD AZOTOBACTER
Rod Azotobacter pripada grupi Gram negativnih aerobnih/mikroaerofilnih štapića i
koka. To je heterogena grupa bakterija. Kod ovih bakterija zastupljen je respiratorni tip
metabolizma. Bakterije su pleomorfne i imaju razliĉit oblik. Mogu biti štapići ili koke.
Javljaju se pojedinaĉno, u parovima ili u nepravilnim grupama. Azotobakter su uglavnom
pokretne bakterije, kreću se peritrihama, mada su primećeni i neki nepokretni oblici. Ove
bakterije rastu mikroaerofilno i neki sojevi proizvode pigment. Za Azotobakter je
karakteristiĉna sposobnost azotofiksacije. Ove bakterije su zastupljene u zemljištu i vodi a
karakteriše ih i sposobnost formiranja asocijacija sa višim biljkama. Tipiĉan predstavnik ovog
roda je Azotobakter chroococum. Azotobacter pored sposobnosti azotofiksacije ima
sposobnost i proizvodnje hormona rasta (citokini, giberelini), vitamina (produkuje B
vitamine). On konvertuje azot u amonijak koji uzimaju biljke. Ova bakterija moţe da
proizvede i antifugalna jedinjenja, koja se koriste u zaštiti biljaka. Proizvodnja Ċubriva sa
Azotobacter vrši se na odreĊenim ĉvrstim ili teĉnim podlogama, u zavisnosti od biljne vrste
(Petrović et al., 2007).

31
Azotobacter vinelandii
Azotobacter vinelandii je rod slobodnoţivećih diazotrofnih bakterija koje fazu
odmaranja provode u vidu ciste. Nalazi se u alkalnom zemljištu, akvatiĉnoj sredini i na nekim
biljkama. To je Gram-negativna bakterija. Poseduje nekoliko metaboliĉkih mogućnosti,
ukljuĉujući i fiksaciju atmosferskog azota i njegovu konverziju do amonijaka. Njihov
jedinstveni sistem posebnih enzima nitrogenaza, omogućava ovim bakterijama korišćenje u
nauci, kako bi se bolje razumeo proces azotofiksacije i njeno korišćenje u poljoprivredi.
Azotobacter sp. ima najveću metaboliĉku stopu od drugih organizama. One su zbog
jedinstvenog naĉina metabolizma za aerobnu fiksaciju azota, privukle dosta interesovanja
nauĉnika. Ćelije sa visokom stopom respiracije omogućavaju normalnu osetljivost
nitrogenaza kada se izloţe kiseoniku. Azotobacter se moţe koristiti za produkciju proteina
koji štite nitrogenaze od iznenadnog stresa izazvanim kiseonikom. Ove bakterije sintetišu tri
nitrogenaze.
Diazotrofni organizmi, kao što je i Azotobacter, imaju kljuĉnu ulogu u svakom
ekosistemu, jer omogućavaju dostupnost azota svim organizmima. Oni procesom
azotofiksacije pretvaraju azot u amonijak, nakon ĉega se amonijak redukuje do proteina, koji
se ugraĊuju u biljke. Azotofiksacija se koristi u poljoprivredi, gde se biofertilizacijom
povećava plodnost zemljišta. Azotobacter vinelandii pored fiksacije azota, uĉestvuje u
proizvodnji nekih biljnih hormona (auksina i giberilina), ali je bitan i u dobijanju nekih
vitamina.
2.9.2. ROD BACILLUS
Rod Bacillus pripada grupi Gram pozitivnih štapića i koka koje formiraju endospore.
To su krupne, sporogene bakterije koje su aerobi ili fakultativni anaerobi. Mnoge vrste imaju
sposobnost sinteze nekih antibiotika (gramicidin, bacitracin) i nekih ekstracelularnih enzima.
Bacillus anthracis je vrsta koja je najznaĉajnija za medicinu, koja je patogena za ljude i
ţivotinje, jer izaziva antraks (crni prišt). U poslednje vreme, veoma vaţnu ulogu zauzima i
Bacillus thurigiensis, jer u toku sporulacije formira kristalni protein, odnosno parasporalno
telo. To je veoma aktivan biološki insekticid. Ovaj kristalni protein se rastvara u crevima
larvi, prelazi u krv i dovodi do smrti larve. Kako bi se zaštitila ţivotna sredina, ovaj pesticid je
veoma znaĉajan, jer se hemijski pesticidi teško uklanjaju iz zemljišta. Kao biološki agens koji
utiĉe na suzbijanje brojnosti fitopatogena, najveći znaĉaj ima Bacillus subtilis, mada se
zadnjih godina skreće paţnja i na korišćenje Bacillus mycoides i Bacillus cereus. Bakterije

32
ovog roda sintetišu veliki broj jedinjenja kojima utiĉu na svoju okolinu, povećavajući tako
pristupaĉnost potrebnih elemenata biljkama (Petrović et al., 2007).
Bacillus subtilis
Bacillus subtilis je Gram-pozitivna bakterija, koja je pokretna, štapićastog oblika.
Prvenstveno je prisutna u zemljištu, ali i u vodi, vazduhu, na biljnom materijalu. U poĉetku se
je ova bakterija zvala ,,Vibrio subtilis″, ali joj je naziv promenjen 1872. godine. Tokom
nutritivnog stresa stvara endospore koje joj omogućavaju preţivljavanje nepovoljnih uslova
(visoke temperature, zraĉenja, otrovne supstance). U preţivljavanju glavnu ulogu imajuα i β
proteini koji zasićuju molekul DNA i pri tome je štite. Ovi proteini prisutni su samo u
endospori i nema ih u vegetativnim ćelijama. Spore Bacillus subtilis su rasprostranjene svuda
u prirodi. Bacillus subtilis je mezofilna vrsta i moţe da raste na temperaturama od 200C do
2500C, dok njene spore mogu da preţive i 1000
0C 30 minuta. Tokom svoje aktivnosti stvara
veliki broj enzima (Petrović et al., 2007).
Bacillus subtilis nije patogena za ĉoveka, ali postoje podaci o infekcijama ovom
bakterijom kod osoba sa oslabljenim imunitetom. Uglavnom su to oportunistiĉke
infekcije. Bacillus subtilis je jedna od najĉešće korišćenih bakterija za produkciju enzima i
drugih produkata koji imaju široku primenu u industriji. Ova vrsta izaziva koagulaciju mleka
usled proteolitiĉke aktivnosti (slatko zgrušavanje). Bacillus subtilis razgraĊuje kompleksne
ugljenohidrate i proizvodi amilazu. Smatra se idealnim probiotikom, zbog svoje sposobnosti
preţivljavanja na niskim pH u gastrointestinalnom traktu. Stvaranjem endospora moţe se
odrţati veoma dugo. Bacillus subtilis stimuliše imuni sistem, tako da se dobije širok spektar
antitela. Koristi se za produkciju velikog broja antibiotika. Pored enzima, ova bakterija
produkuje i toksin subtilizin, koji je izazivaĉ mnogih alergijskih reakcija. Bacillus subtilis je
uzet i kao zemljišni inokulant i koristi se za dobijanje mikrobioloških Ċubriva. Ova vrsta
produkuje neke fungicidne komponente, pa se koristi i za zaštitu biljaka. Zbog lake genetiĉke
manipulacije koristi se i kao model u laboratorijama. Koriste se i za prouĉavanje kolonija
bakterija, jer dobro rastu na agru, u svim uslovima (Jemcev and Đukić, 2000).
Bacillus megaterium
Bacilllus megaterium je štapićasta, Gram-pozitivna bakterija, prisutna na razliĉitim
staništima. Ĉesto se moţe javljati u parovima ili lancima. Bacillus megaterium raste na
temperaturama od 30C pa do 45
0C, ali je optimalna temperature za njen rast 30
0C. Uglavnom
raste na biljkama i koristi se kao biokontrolni agens za zaštitu biljaka. Pre otkrića Bacillus

33
subtilis, ova bakterija je korišćena za prouĉavanje biohemijskih procesa, sporulacije i drugo.
Najveću prednost dobija razvojem biotehnologije, gde se koristi u mnoge svrhe. Veoma je
vaţan industrijski organizam. Produkuje razne penicillin amidaze koje se koriste za sintetiĉko
dobijanje penicilina, amilaze koje se koriste u pekarskoj industriji i glukozo-dehidrogenaze
koje se koriste za testove glukoze u krvi. Jedna od glavnih osobenosti Bacillus megateriuma
je sposobnost fiksacije azota, a uĉestvuje i u transformaciji fosfora, nastanku hormona rasta
biljaka i vitamina B12, kao i lekova sa fungicidnim i antiviralnim karakteristikama (Petrović
et al., 2007).

34
3. CILJ RADA
Rezultati mnogobrojnih istraţivanja su pokazali da odreĊene vrste zemljišnih
mikroorganizama imaju veliki znaĉaj u stimulaciji rasta biljaka, što se manifestuje kroz
njihovu sposobnost solubilizacije, mobilizacije i sinteze biološki aktivnih materija koje
povoljno deluju na biljni metabolizam.
Shodno tome, ciljevi ovog rada su definisani u sledećem:
1. Optimizacija medijuma za gajenje odabranih sojeva korisnih zemljišnih bakterija
sa sposobnošću sinteze aktivnih supstanci, solubilizacije fosfata i azotofiksacije, u
kontrolisanim uslovima fermentatora, radi dobijanja najveće biomase;
2. Modifikacija sastava medijuma, sa ciljem da se postigne jednostavnija i jeftinija
proizvodnja mikrobiološkog Ċubriva, dostupnog za široku, komercijalnu upotrebu.

35
4. MATERIJAL I METODE
4.1. MATERIJAL
4.1.1. BAKTERIJSKE KULTURE
U radu su korišćene bakterijske kulture Bacillus subtilis (sintetiše aktivne supstance),
Bacillus megaterium (solubilizacija fosfata) i Azotobacter vinelandii (azotofiksacija).
Navedeni mikroorganizmi su nepatogeni izolati iz zemljišta. Štok kulture bakterijskih sojeva
su ĉuvane u krioviala masa 15% glicerolom i odgovarajućom hranljivom podlogomna – 850
C.
4.1.2. HRANLJIVE PODLOGE
Podloge korišćene za zasejavanje mikroorganizama izvaĊenih iz štoka su:
- HA, hranljivi agar koji je gotova podloga (Bacillus subtilis, Bacillus megaterium)
- Pikovskaya′s agar, koristi se za fosfobakterije (Bacillus megaterium)
- Jenson′s agar, predstavlja bezazotnu podlogu za azotofiksatore (Azotobacter
vinelandii).
Tabela 1. Sastav podloga
g/l
Podloge
PSB,
Pikovskaya agar
N-fiksirajući
Јensenagar
Dekstroza 10
Glukoza 20
ekstrakt.kvasca 0.5
K2HPO4 1
MgSO4 0.1 0.5
KCl 0.2
NaCl 0.5
(NH4)2SO4 0.5
Na2MoO4 0.001
FeSO4 Tr 0.01
Ca3(PO4)2 5
CaCO3 2
Agar 15 15

36
4.2. METODE
4.2.1. PRIPREMA BAKTERIJSKIH KULTURA
Bacillus subtilis i Bacillus megaterium su presejani na petri ploĉe sa hranljivim
agarom, a Azotobacter vinelandii na (Jenson agar) bezazotnoj podlozi. Ovako presejane
kulture su inkubiranena 300
C, 3 dana Bacillus i 5 dana Azotobacter vinelandii, nakon ĉega su
se jasno uoĉavale kolonije.
Priprema bakterijske suspenzije
,,Revitalizovani″ mikroorganizmi su korišćeni za pravljenje suspenzija u 0,9 %
rastvoru NaCl (fiziološki rastvor). Turbiditet suspenzija je uz upotrebu densitometra podešen
na 0,5 McF, što odgovara 108
CFU/ml (colony formit unit).
Solubilizacija fosfata
Dokazivanje solubilizacije fosfata, od strane odabranih bakterijskih sojeva, je vršeno
kvalitativnom metodom ĉvrstog agara na Pikovskaya podlozi (SIGMA-ALDRICH), ĉiji je
sastav dat u tabeli 1. Pomenuta podloga je odmerena, sterilisana i razlivena u petri ploĉe, a
nakon hlaĊenja su na njoj ezom zasejani pomenuti sojevi. Usled prisustva nerastvornog
kalcijumfosfata, boja podloge je mleĉnobela. Solubilizacija fosfata se manifestuje tokom
inkubacije 5-7 dana na 300
C, kada se oko formiranih kolonija uoĉavaju providne zone.
Efikasnost solubilizacije se moţe odrediti na osnovu dijametra kolonije i dijametra
obezbojene zone (Karpagam and Nagalakshmi., 2014). Ovom metodom je potvrĊena
sposobnost solubilizacije od strane Bacillus megaterium.
Azotofiksacija
Sposobnost azotofiksacije soja Azotobacter vinelandii je potvrĊena zasejavanjem soja
na bezazotnoj podlozi – Jensen agar (HIMEDIA), ĉiji je sastav dat u tabeli 1. Azot je
neophodan element za rast mikroorganizama, tako da se ova podloga koristi za izolaciju i
identifikaciju mikroorganizama koji imaju sposobnost vezivanja atmosferskog azota.
37
4.2.2. PRIPREMA STARTNE KULTURE (inokuluma)
Startne kulture su pripremljene u erlenmajere i odgovarajućim sterilisanim teĉnim
podlogama za izabrane sojeve (Tabela 2.,Tabela 3. i Tabela 4.). U sterilnim uslovima
laminara (ESCO, USA) (Slika 5.) su inokulisane podloge sa po 1,5 ml odgovarajuće
bakterijske suspenzije. Ovako zasejane kulture su stavljene na rotacioni mešaĉ (200 rpm) gde
je svakog dana kontrolisana sterilnost i brojnost mikroorganizama. Mikroorganizmi su gajeni
na 300C 48h Bacillus i 72h Azotobacter vinelandii. Startne kulture su korišćene za inokulaciju
teĉnog fermentora.
Slika 5. Priprema inokuluma na rotacionom mešaĉu
Tabela 2. Sastav podloga g/l za inokulum i fermentor za Bacillus subtilis.
Bacillus subtilis Teĉna podloga
GFM modifikovana. F1 F2
Glukoza 20 10 3
Pepton 5 5
Ekstrakt kvasca 1 1 1
Mesni ekstrakt 3 3
NH4NO3 0.5 0.3
KH2PO4 5 5
MgSO4 0.5 0.5
NaCl 0.5 0.5
(NH4)2MoO4 u tragovima
Ca3(NO3)2, 1M (ml) 1 1.45
MgSO4x7H2O 0.5
(NH4)2SO4 1
PVP-40 20 20
FeCl3 0.03M (ml) 8.4 2 2
CaO 1.12
EDTA 2
Rastvor soli (ml) 10 10
NH4HPO4 2 3

38
Tabela 3. Sastav podloge g/l za inokulum i fermentor za Bacillus megaterium.
Bacillus megaterium Podloga
Glukoza 10
Ekstrakt kvasca 1
NH4NO3 0.5
KH2PO4 5
MgSO4 0.5
NaCl 0.5
Ca3(NO3)2, 1М (ml) 1
(NH4)2SO4 1
PVP-40 20
FeCl3 0.03M (ml) 2
Rastvor soli (ml) 10
NH4HPO4 5
Glicerol (ml) 15
Tabela 4. Sastav podloga g/l za inokulum i fermentor za Azotobacter vinelandii.
Azotobacter vinelandii teĉna podloga
Jenson - modif. LPM
Šećer 20 10
K2HPO4 1 0.583
MgSO4 0.5 0.098
NaCl 0.5 0.058
Na2MoO4 0.005 0.002
FeSO4 0.1 0.005
glicerol (ml) 10
CH3CO2NH4 1.2 1.2
PVP-40 20
CaSO4x2H2O
0.1
KH2PO4
0.224

39
4.2.3. KONTROLA STERILNOSTI I ODREĐIVANJE BROJNOSTI
MIKROORGANIZAMA
Svakog dana inkubacije, iz erlenmajera i fermentora (Slika 6.) je uziman uzorak kako
bi se ispitala brojnost mikroorganizama (metoda posrednog odreĊivanja brojnosti) i potvrdilo
odsustvo kontaminacije. U ove svrhe je korišćena metoda kapi ili “drop plate method”
(Somasegaran P and Hoben JH., 1994). Serija razblaţenja uzorka je pripremljena
prebacivanjem 1 ml uzorka u 9 ml sterilnog fiziološkog rastvora u pripremljenim sterilnim
epruvetama. A zatim je iz svakog razblaţenja, od 10-1
do 10-10
, uzimano po 20 mikrolitara i u
vidu kapi izruĉeno u petri ploĉe sa hranljivim agarom, koje su prethodno podeljene u
kvadrante. Svaki kvadrant je obeleţen i predstavlja odgovarajuće razblaţenje. Zasejani
mikroorganizmi su inkubirani 24h-48h, na 30ᵒ C, a brojanje je vršeno u tri ponavljanja.
Slika 6. Fermentor
Ukupan broj ćelija u 1 ml poĉetnog uzorka je raĉunat po sledećoj formuli:
N= n x 50 x10x
N –ukupan broj ćelija u 1 ml uzorka
50 – koeficijent ispravke do 1ml
10x - razblaţenje

40
4.2.4. PRIPREMA FERMENTORA
Fermentor (Slika 6.) zapremine 15L (Naprto Scientific, India) je napunjen
odgovarajućom podlogom i sterilisan na 1200 C, 20 minuta, pod pritiskom od 1,2 bara. Posle
hlaĊenja medijuma, na bioreaktoru je podešen i aktiviran sistem sledećih parametara: pH,
temperature i vazduha.
1. pH: 6,5 – 6,7 za Bacillus;
7 – 7,2 za Azotobacter vinelandii.
Regilisanje pH medijuma je postignuto uz pomoć peristaltiĉkih pumpi kojima je
kontrolisano dodavanje kiseline (1M HCL) i baze (1M NaOH).
2. Temperatura je odrţavana na 300C uz pomoć vodenog sistema (Water Chilleri).
3. Rastvoreni kiseonik je odrţavan na 30%, a ravnomerno mešanje je postignuto
magnetnim mešaĉem na 200 – 300 rpm.
4.2.5. METODE BIOFERMENTACIJE
Bacillus subtilis
Metoda 1: Startna kultura je pripremljena na već opisan naĉin u GFM modifikovanoj
podlozi (tabela 2.), a zatim je njome inokulisan fermentor u kome je takoĊe pripremljena
GFM modifikovana podloga. Proces fermentacije je praćen 72h uz stalno uzimanje uzoraka,
radi provere sterilnosti i praćenja broja ćelija (CFU/ml).
Metoda 2: Pripremljenim inokulumom je inokulisan fermentor sa podlogom f1 i f2
ĉija je receptura data u tabeli 2. Metoda fermentacije je bila po principu Fed-Batch
fermentacije -,,hranjenja″ bakterija posle 12h inkubacije, što je automatski podešeno putem
peristaltiĉkih pumpi.
Šema hranjenja:
Vreme
inkubacije
Ca(NO3)2+glukoza (NH4)2SO4+
(NH4)2HPO4
Podloga
12 – 13h
13 – 14h
14 – 15h
15 – 16h
30 ml
30 ml
60 ml
80 ml
20 ml
20 ml
20 ml
20 ml
f1
12 – 13h
13 – 14h
14 – 15h
15 – 16h
60 ml
80 ml
100 ml
120 ml
40 ml
40 ml
40 ml
40 ml
f2

41
U periodu od 12 – 16h fermentacije, peristaltiĉkom pumpom je ukupno dodato:
- za prvu podlogu f1, 200 ml rastvora glukoze + Ca(NO3)2 koncentracije 400g/l
(glukoza) , 10 g/l Ca i 80 ml (NH4)2SO4 (120 g/l) + (NH4)2HPO4 (120 g/l);
- za drugu podlogu f2, 360 ml rastvora glukoze + Ca(NO3)2 i 120 ml (NH4)2SO4 (120
g/l) + (NH4)2HPO4 (120 g/l).
Bacillus megatherium
Metode fermentacije su sliĉne opisanim metodama kod gajenja Bacillus subtilis.
Receptura ispitivane teĉne podloge u fermentoru je data u tabeli 3.
Metodom 1 je praćeno uvećanje biomase bez dodavanja hranljivih materija, dok je
metoda 2 uvećanja biomase po principu Fed-Batch fermentacije tj. hranjenja. Dodavanje
nutritijenata je vršeno posle 24h fermentacionog procesa i to: 200 ml glukoza + Ca(NO3)2 i
50 ml (NH4)2SO4 + (NH4)2HPO4 (koncentracije rastvora su identiĉne kao za metodu 2
gajenja B.subtilis).
Azotobacter vinelandii
Za azotofiksirajuću bakteriju, praćena je fementacija u dve podloge ĉiji sastav je
naveden u tabeli 4:
- Jenson′s medium (bezazotna podloga) i
- Podloga za veliku proizvodnju (LPM).
Bioreaktor (fermentor) je inokulisan startnom kulturom i svi parametri su strogo
kontrolisani u narednih 72h. U periodima posle 24, 48 i 72h fermentacije, uziman je uzorak i
vršeno zasejavanje radi odreĊivanja brojnosti mikroorganizama, metodom razreĊenja.

42
5. REZULTATII DISKUSIJA
Rezultati dobijeni ispitivanjem optimizacije medijuma za gajenje odabranih sojeva
korisnih zemljišnih bakterija u kontrolisanim uslovima fermentatora,radi dobijanja najveće
biomase, tj. jednostavnije i jeftinije proizvodnje mikrobiološkog Ċubriva, su obraĊeni i
prikazani pojedinaĉno za svaki soj.
Bacillus subtilisa
Dobijeni rezultati su pokazali da je maksimalni broj ćelija u GFM podlozi 5x108
cfu/ml posle 72h fermentacije. Sa ciljem optimizacije medijuma i dobijanja veće biomase,
metodom 2 je modifikovana podloga, a zatim je uveden reţim “hranjenja”. U tom sluĉaju je
primećen veći rast brojnosti ćelija, što se moţe videti u tabeli 5. Metodom 2, gde smo
ispitivali porast B. subtilis u podlozi f1 i f2, dobili smo sliĉne rezultate, uprkos modifikaciji
podloga i drugaĉijem reţimu ishrane. MeĊutim, u oba sluĉaja, postignut je bolji rezultat i
dobijena veća biomasa ćelija u odnosu na prvu metodu posle 72h procesa fermentacije:
2.5x109 u podlozi f1 i 2x10
9 u podlozi f2 (cfu/ml).
Tabela 5. Standardizacija rasta Bacillus subtilis
Vreme
inkubacije u
fermentoru
GFM (cfu/ml) f1 (cfu/ml) f2 (cfu/ml)
24h 1.5x10^8 9.2x10^8 9.5x10^8
48h 5x10^8 1.5x10^9 3x10^9
72h 5x10^8 2.5x10^9 2x10^9
Na osnovu datih receptura podloga (Tabela 2.), moţe se uoĉiti znaĉajna razlika u
koliĉini glukoze koja ulazi u njihov sastav. Rezultati ukazuju na neophodnost šećera kao
glavnog izvora energije, ali i da njegova koliĉina u startu nije presudna za dobijanje što veće
biomase, već da je to u korelaciji sa fazom rasta pri kultivisanju u teĉnoj podlozi. Zapravo,
hranjenjem mikroorganizama i odrţavanjem konstantnim uslove sredine u fermentoru, moţe
se produţiti eksponencijalna faza rasta, kako bi prirast biomase bio što veći (Jemcev and
Đukić., 2000). Fed-batch fermentacijom i optimizacijom procesa hranjenja bakterija,
postignut je bolji rast biomasa B.subtilis K-C3, kao i veća produkcija enzima natokinaze u
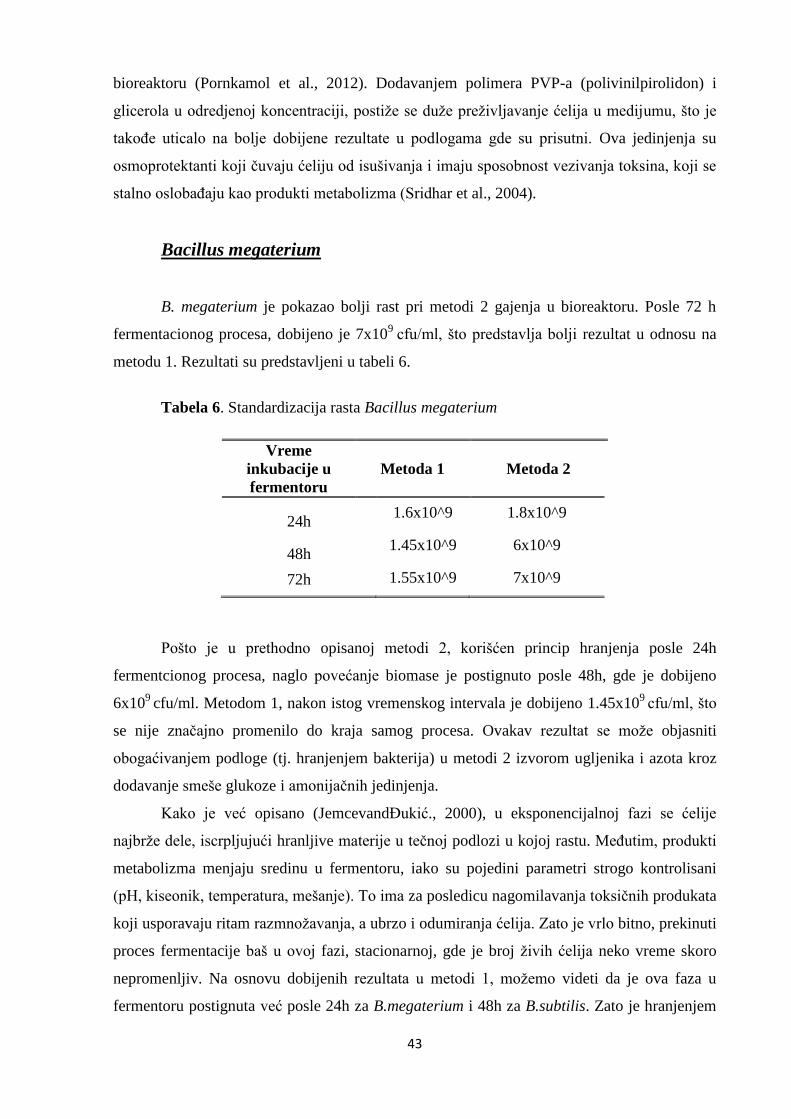
43
bioreaktoru (Pornkamol et al., 2012). Dodavanjem polimera PVP-a (polivinilpirolidon) i
glicerola u odredjenoj koncentraciji, postiţe se duţe preţivljavanje ćelija u medijumu, što je
takoĊe uticalo na bolje dobijene rezultate u podlogama gde su prisutni. Ova jedinjenja su
osmoprotektanti koji ĉuvaju ćeliju od isušivanja i imaju sposobnost vezivanja toksina, koji se
stalno oslobaĊaju kao produkti metabolizma (Sridhar et al., 2004).
Bacillus megaterium
B. megaterium je pokazao bolji rast pri metodi 2 gajenja u bioreaktoru. Posle 72 h
fermentacionog procesa, dobijeno je 7x109
cfu/ml, što predstavlja bolji rezultat u odnosu na
metodu 1. Rezultati su predstavljeni u tabeli 6.
Tabela 6. Standardizacija rasta Bacillus megaterium
Vreme
inkubacije u
fermentoru
Metoda 1 Metoda 2
24h 1.6x10^9 1.8x10^9
48h 1.45x10^9 6x10^9
72h 1.55x10^9 7x10^9
Pošto je u prethodno opisanoj metodi 2, korišćen princip hranjenja posle 24h
fermentcionog procesa, naglo povećanje biomase je postignuto posle 48h, gde je dobijeno
6x109
cfu/ml. Metodom 1, nakon istog vremenskog intervala je dobijeno 1.45x109
cfu/ml, što
se nije znaĉajno promenilo do kraja samog procesa. Ovakav rezultat se moţe objasniti
obogaćivanjem podloge (tj. hranjenjem bakterija) u metodi 2 izvorom ugljenika i azota kroz
dodavanje smeše glukoze i amonijaĉnih jedinjenja.
Kako je već opisano (JemcevandĐukić., 2000), u eksponencijalnoj fazi se ćelije
najbrţe dele, iscrpljujući hranljive materije u teĉnoj podlozi u kojoj rastu. MeĊutim, produkti
metabolizma menjaju sredinu u fermentoru, iako su pojedini parametri strogo kontrolisani
(pH, kiseonik, temperatura, mešanje). To ima za posledicu nagomilavanja toksiĉnih produkata
koji usporavaju ritam razmnoţavanja, a ubrzo i odumiranja ćelija. Zato je vrlo bitno, prekinuti
proces fermentacije baš u ovoj fazi, stacionarnoj, gde je broj ţivih ćelija neko vreme skoro
nepromenljiv. Na osnovu dobijenih rezultata u metodi 1, moţemo videti da je ova faza u
fermentoru postignuta već posle 24h za B.megaterium i 48h za B.subtilis. Zato je hranjenjem

44
bakterija posle 12h (B.subtilis) i 24h (B.megaterium) u metodi 2, eksponencijalno
razmnoţavanje produţeno, pa je shodno tome, posle 48h fermentacionog procesa i dobijen
veći broj ćelija.
Azotobacter vinelandii
Azotobacter vinelandii je pokazao brţi rast posle 24h fermentacije u bioreaktoru u
Jensons modif. podlozi, u odnosu na podlogu za veliku proizvodnju. Posle 24h postignuto je
uvećanje broja ćelija na 9.6x108
cfu/ml u prvoj podlozi, dok je u drugoj bilo znatno manje
1.8x107
cfu/ml. Ovakav rezultat se moţe objasniti većom koliĉinom šećera u prvoj podlozi,
koji predstavlja glavni izvor energije i ugljenika. Rezultati su predstavljeni u tabeli 7.
Tabela 7. Standardizacija rasta Azotobacter vinelandii
Vreme
inkubacije u
fermentoru
Teĉna podloga
Jens.modif LPM
24h 9.6x10^8 1.8x10^7
48h 4.5x10^9 1x10^9
72h 5.5x10^9 3.8x10^9
Kako je vreme kultivacije odmicalo, moţe se uoĉiti da je najveći broj ćelija u dvema
ispitivanim podlogama bio 109
cfu/ml. Iako je naglo uvećanje brojnosti ćelija uoĉeno u prvoj
podlozi posle 24h, već nakon 48h, primećuje se i u drugoj podlozi. Zapravo, posle 72h
fermentacionog procesa, moţe se videti da nema velike razlike u brojnosti kada su u pitanju
ove dve podloge: 5.5x109
i 3.8x109
cfu/ml. Moţda bi ovakav rezultat mogli da objasnimo
periodom prilagoĊavanja bakterija na uslove sredine, koji je duţi u drugoj podlozi, a samim
tim je i eksponencijalna faza odloţena. Svakako, radi se o podlozi sa manje zastupljenim
neophodnim hranljivim materijama i solima (tabela 4), što moţe uzrokovati ovakav efekat.
Samim tim, se moţe uvrstiti za komercijalnu proizvodnju siromašnija podloga, kako bi
proces i proizvod bili jeftiniji i dostupniji za komercijalnu upotrebu.

45
6. ZAKLJUĈAK
Zemljišni mikroorganizmi imaju znaĉajnu ulogu u ekosistemu. Njihova višestruka
korist se ogleda u fiksaciji, solubilizaciji, mobilizaciji i obnavljanju nutritijenata u zemljištu.
Na ovaj naĉin, korisni mikroorganizmi ĉine esencijalne mineralne elemente dostupnim
korenovom sistemu.
Alternativna tehnologija, kojom bi se izbegli svi negativni efekti hemijskog Ċubriva,
ogleda se u povećanju brojnosti korisnih mikroorganizama u zemljištu. Izolacijom ovih
sojeva, njihovom identifikacijom, umnoţavanjem kroz proces fermentacije i vraćanjem u
zemljište – BIOFERTILIZACIJA, teţi se prirodnom rešavanju problema nedostatka ili
nedostupnosti nutritijenata. Mikrobiološka Ċubriva su ekološka i isplativija, a u zavisnosti od
osobina mikroorganizama, višestruko korisna: utiĉu na rast i razvoj biljaka obavljanjem
metaboliĉkih funkcija, povećavaju biološku produktivnost zemljišta, redukuju patogene
mikroorganizme (produkcija aktivnih metabolita).
Rezultati naših istraţivanja su pokazali da je odgovarajućom optimizacijom podloge i
kinetikom Fed-batch procesa moguće postići što veću biomasu korisnih mikroorganizama u
fermentoru. Samim tim, dodavanjem najefikasnijih osmoprotektanata bi se produţio ţivotni
vek ćelija u teĉnoj formi proizvoda. U budućnosti bi trebalo pratiti variranje brojnosti ćelija u
datim formulacijama, radi taĉnog odreĊivanja efikasnosti proizvoda, gde brojnost ćelija na
osnovu deklarisanih parametara, ne bi trebalo da bude manji od 108 cfu/ml. TakoĊe,
efikasnost ispitivanih bakterija bi trebalo ispitati na biljnim kulturama, kako bi se potvrdila
mogućnost korišćenja proizvoda kao mikrobiološko Ċubrivo.

46
7. LITERATURA
1. Ahemad, M., Kibert, M. 2014: Mechanisms and applications of plant growth promoting
Rhizobacteria: Current perspective. – Journal of King Saud University – Science. 26: 1 –
20.
2. Alikhani, H. A., Saleh – Rastin, N., Antoun, H. 2006: Phosphate solubilization activity
of rhizobia native to Iranian soils. – Plant and Soil. 287: 35 – 41.
3. Chandra, K. 2005: Liquid biofertilizers – Regional centre of organic farming, India.
4. Đukić, D., Jemcev, V. 2004: Opšta i industrijska mikrobiologija, Novi Sad.
5. Đukić, D., Jemcev, V., Kuzmanova, J. 2007: Biotehnologija zemljišta. – Budućnost,
Novi Sad.
6. Glick, B. 2012: Plant Growth – Promoting Bacteria: Mechanisms and Applications. –
Scientifica. 2012, 15 pages.
7. Gomathy, M., Thangaraju, M., Gunasekaran, S., Gopal, N. O., Gopal, H. 2008: Method
and Quantity of Liquid Formulation of Phosphobacteria Required for Seed Inoculation.
– Pakistan Journal of Biological Sciences. 11(1): 86 – 91.
8. Jemcev, V., Đukić D. 2000: Mikrobiologija. – Vojnoizdavaĉki zavod, Beograd.
9. Khan, A., Jilani, G., Akhtar, S. M., Naqvi, S. M. S., Rasheed, M. 2009: Phosphorus
Solubilizing Bacteria: Occurrence, Mechanisms and their Role in Crop Production. –
Journal of Agricultural and Biological Science. 1: 48 – 58.
10. Kumaresan, G., Reetha, D. 2011: Survival of Azospirillum brasilense in liquid
formulation amended with different chemical additives. – Journal of Phytology. 3:48 -
51.
11. Leo Daniel, 2013: Effect of Polymeric Additives, Adjuvants, Surfactants on Survival,
Stability and Plant Growth Promoting Ability of Liquid Bioinoculants. – Journal of
PlantPhysiology & Pathology. 1:2.
12. Ma, X., Wang, X., Cheng, J., Nie, X., Yu, X., Zhac, Y., Wang, W. 2015:
Microencapsulation of Bacillus suptilis B99 – 2 and its biocontrol efficiency against
Rhizoctonia solani in tomato. – Biological control. 90: 34 – 41.

47
13. Mauricio, A.2013: Scale – up from shake flasks to pilot-scale production of the plant
growth – promoting bacterium Azospirillum brasilense for preparing a liquid inoculants
formulation. – Applied Microbiology and Biotechnology. 97:9665 – 9674.
14. Mihajilov – Krstev, T. 2008: Radna sveska iz mikrobiologije. – Niš: Prirodno –
matematiĉki fakultet, Odsek za biologiju sa ekologijom, Niš.
15. Monteiro, S. M. S., Clemente, J. J., Carrondo, M. J. T., Cunha, A. E. 2014: Enhanced
Spore Production of Bacillus subtilis Grown in a Chemically Defined Medium. –
Advances inMicrobiology. 4: 444 – 454.
16. Omer, A. M., 2010: Bioformulations of Bacillus Spores for using as Biofertilizer. – Life
ScienceJournal.7: 124 – 131.
17. Ortiz – Castro, R., Contreras – Cornejo, H.A., Macias – Rodriguez, L., Lopez – Bucio,
H. 2009: The role of microbial signals in plant growth and development. – Plant
Signaling and Behavior. 4: 701 – 712.
18. Petrović, O., Kneţević, P., Simeunović, J. 2007: Mikrobiologija. – Skripta za studentre,
Novi Sad.
19. Rahman, A., Sitepu, I., Tang, S.-Y., Hashidoko, Y. 2010: Salkowski′s Reagent Test as a
Primary Screening Index for Functionalities of Rhizobacteria Isolated from Wild
Dipterocarp Saplings Growing Naturally on Medium. – Strongly Acidic Tropical Peat
Soil. – Bioscience Biotechnology andBiochemistry. 74: 2202 – 2208.
20. Richardson, A., Simpson, R. 2011: Soil Microorganisms Mediating Phosphorus
Availability. – Plant physiology. 156: 989 – 996.
21. Seshadri, S., Muthukumarasamy, R., Lakshminarasimhan, C., Ignacimuthu, S. 2000:
Solubilization of inorganic phosphates by Azospirillum halopraeferans. – Current
science. 79.
22. Singh, T., Purohit, S.S. 2012: Biofertilizer Technology. – Agrobios, India.
23. Sivakumar, P. K, Parthasarthi, R., Lakshmipriya, V. P. 2014: Encapsulation of plant
growth promoting inoculant in bacterial alginate beads enriched with humic acid. –
International Journalof Current Microbiology and Applied Sciences. 3: 415 – 422.
24. Sridhar, V., Brahmaprakash, G.P., Hegde, S.V. 2004: Development of a Liquid Using
Osmoprotectants for Phosphate Solubilizing Bacterium (Bacillus megaterium). –
Kamataka. Journal of Agricultural Science. 17: 251 – 257.
25. Unrean, P., Nguyen, N., Visessanguan, W., Kitsuban, P. 2012: Improvement of
nattokinase production by Bacillus subtilis using an optimal feed strategy in fed – batch
fermentation. – Researchjournal.17: 769 – 777.

48
26. Velineni, S., Brahmaprakash, G. P. 2011: Survival and Phosphate Solubilizing Ability
of Bacillus megatherium in Liquid Inoculants under High Temperature and Desiccation
Stress. – Jornal ofAgriculture Science and Technology. 13: 795 – 802.
27. Young, C. C., Rekha, P. D., Lai, W. A., Arun, A. B. 2006: Encapsulation of Plant
Growth – Promoting Bacteria in Alginate Beads Enriched With Humic Acid. –
Biotechnology andBioengineering.95.

49
8. BIOGRAFIJA
Dunja Ilić roĊena je 26. oktobra 1990. godine u Beogradu. Osnovnu školu ,,Desanka
Maksimović”, završava 2005. godine u Grdelici i iste upisuje srednju medicinsku školu ,,Dr.
Milenko Hadzić “, u Nišu, gde završava smer medicinska sestra - vaspitaĉ. Osnovne
akademske studije na Prirodno – matematiĉkom fakultetu Univerziteta u Nišu, Departman za
biologiju i ekologiju, upisuje 2009. godine. Studije završava 2013. godine i tada upisuje
Master akademske studije, smer Biologija.

Прилог5/1
ПРИРОДНО - МАТЕМАТИЧКИФАКУЛТЕТ
НИШ
КЉУЧНА ДОКУМЕНТАЦИЈСКА ИНФОРМАЦИЈА
Редни број, РБР:
Идентификациони број, ИБР:
Тип документације, ТД: Монографска
Тип записа, ТЗ: Текстуални / графички
Врста рада, ВР: Мастер рад
Аутор, АУ: Дуња Илић
Ментор, МН: Татјана Михајилов – Крстев
Наслов рада, НР: Гајење азотофиксирајућих и фосфосолубилизирајућих
бактерија у контролисаним условима ферментора у циљу
биофертилизације земљишта
Језик публикације, ЈП: Српски
Језик извода, ЈИ: Енглески
Земља публиковања, ЗП: Р. Србија
Уже географско подручје, УГП: Р. Србија
Година, ГО: 2016
Издавач, ИЗ: Ауторски репринт
Место и адреса, МА: Ниш, Вишеградска 33.
Физички опис рада, ФО: (поглавља/страна/ цитата/табела/слика/графика/прилога)
Научна област, НО: Биологија
Научна дисциплина, НД: Микробиологија
Предметна одредница/Кључне речи, ПО: Азотофиксирајуће и фосфосолубилизирајуће бактерије, микробиолошка ђубрива, биофертилизација.
УДК 631.461.7 : 631.484 631.461.5 : 631.484
Чува се, ЧУ: Библиотека
Важна напомена, ВН:
Извод, ИЗ: Земљишни микроорганизми имају значајну улогу у екосистему. Њихова вишеструка корист се огледа у фиксацији, солубилизацији, мобилизацији, обнавлјанју нутритијената у земљишту. Обогаћиванје земљишта корисним микроорганизмима је биофертилизација, то јест имплементација у земљиште у форми микробиолошких ђубрива. У овом раду је описан процес оптимизације медијума као и Fed – Batch процеса гајења азотофиксирајућих и фосфосолубилизирајућих бактерија, са циљем повећања њихове биомасе, производње микробиолошких ђубрива и њихове имплементације у земљиште. БактеријеBacillus suptilis, Bacillus megateriumи Azotobacter vinelandiiсу коришћене за испитивање процеса ферментације у строго контролисаним условима биореактора. Резултати наших истраживања су попоказали да је одговарајућом оптималном подлогом и кинетиком Fed – Batch процеса могуће постићи што већу биомасу корисних микроорганизама у ферментору.
Датум прихватања теме, ДП:
Датум одбране, ДО:
Чланови комисије, КО: Председник:
Члан:
Члан, ментор:
ОбразацQ4.09.13- Издање

1
ПРИРОДНО - МАТЕМАТИЧКИФАКУЛТЕТ
НИШ
KEY WORDS DOCUMENTATION
Accession number, ANO: Identification number, INO: Document type, DT: Monograph
Type of record, TR: textual / graphic
Contents code, CC: Master’s thesis
Author, AU: Dunja Ilić
Mentor, MN: Tatjana Mihajilov - Krstev
Title, TI: GROWING NITROGENFIXATION AND PHOSPHORSOLUBIZING BACTERIA UNDER
CONTROLLED CONDITIONS IN FERMENTOR WITH A GOAL TO FERTILISE THE SOIL
Language of text, LT: Serbian Language of abstract, LA: English
Country of publication, CP: Republic of Serbia
Locality of publication, LP: Serbia
Publication year, PY: 2016
Publisher, PB: author’s reprint
Publication place, PP: Niš, Višegradska 33.
Physical description, PD: (chapters/pages/ref./tables/pictures/graphs/appendixes)
Scientific field, SF: Biology
Scientific discipline, SD: Microbiology
Subject/Key words, S/KW: Nitrogenfixation and phosphosolubizing bacteria, microbiological fertilizer,
biofertilization
UC 631.461.7 : 631.484 631.461.5 : 631.484
Holding data, HD: Library
Note, N: Abstract, AB: Soils play the crucial role in the ecosystem.Their multiple use is shown in fixation,solubilization,
mobilization and recover of nutritients in soil. Soil fertilization with useful microorganisms is
biofertilization so called implementation of soil in form of microbiological fertilizer. This work
shows the process of optimization of medium as well as fed batch growth nitrogenfixation and
phosphorsolubizing bacteria in order to improve biomass and production of microbiological
fertilizer and their implementation in soil. Bacterias culture Bacillus subtilis, Bacillus
megatherium and Azotobacter vinelandii are used for research of process of fermentation in
strictly controlled conditions of bioreactor. The final results of the search shows that appropriate
optimization of the soil and kinetics fed batch process is possible to gain bigger biomass of useful
microorganisms in fermentor.
Accepted by the Scientific Board on, ASB:
Defended on, DE:
Defended Board, DB: President:
Member:
Member, Mentor:

2





























