Embed Size (px)
Citation preview

ĐẠI HỌC QUỐC GIA HÀ NỘI
TRƯỜNG ĐẠI HỌC KHOA HỌC TỰ NHIÊN
---------------------
Lưu Xuân Hòa
ỨNG DỤNG KỸ THUẬT PHÂN TỬ LAI HUỲNH QUANG (FISH)
NHẰM XÁC ĐỊNH NHANH MỘT SỐ LOÀI TẢO ĐỘC HẠI
BIỂN VIỆT NAM
LUẬN VĂN THẠC SĨ KHOA HỌC
Hà Nội – Năm 2011

ĐẠI HỌC QUỐC GIA HÀ NỘI
TRƯỜNG ĐẠI HỌC KHOA HỌC TỰ NHIÊN
---------------------
Lưu Xuân Hòa
ỨNG DỤNG KỸ THUẬT PHÂN TỬ LAI HUỲNH QUANG (FISH)
NHẰM XÁC ĐỊNH NHANH MỘT SỐ LOÀI TẢO ĐỘC HẠI
BIỂN VIỆT NAM
Chuyên ngành: Di truyền học
Mã số: 60 42 70
LUẬN VĂN THẠC SĨ KHOA HỌC
NGƯỜI HƯỚNG DẪN KHOA HỌC: PGS. TS. ĐINH ĐOÀN LONG
Hà Nội – Năm 2011

i
LỜI CAM ĐOAN
Tôi xin cam đoan đây là các kết quả nghiên cứu của chính tôi. Các số liệu, kết
quả nêu trong luận văn là hoàn toàn trung thực, chính xác và chưa được ai công bố
trong bất kỳ công trình nghiên cứu nào khác. Các thông tin trong luận văn đều được
trích dẫn tường minh.
Tác giả
LƯU XUÂN HÒA

ii
LỜI CẢM ƠN
Để hoàn thành bản luận văn này, tôi đã nhận được sự giúp đỡ nhiệt tình của mọi
người. Từ tận đáy lòng, với những lời cảm ơn chân thành nhất tôi xin gửi đến:
- PGS. TS. Đinh Đoàn Long, Bộ môn Di truyền học, Khoa Sinh học, Trường Đại
học Khoa học Tự nhiên (Đại học Quốc gia Hà Nội) đã tận tình hướng dẫn tôi
hoàn thành bản luận văn này.
- Các đồng nghiệp: TS. Nguyễn Văn Nguyên, Ths. Lê Thanh Tùng, CN. Vũ Tuấn
Nam cùng các đồng nghiệp khác của Phòng nghiên cứu công nghệ sinh học
biển, Viện nghiên cứu hải sản, những người đã động viên và giúp đỡ tôi trong
suốt quá trình làm luận văn này.
- Các thầy cô Khoa sinh học, Trường Đại học Khoa học Tự nhiên (Đại học Quốc
gia Hà Nội), những người đã truyền đạt cho tôi những kiến thức quý báu giúp
tôi hoàn thiện dần tư duy nghiên cứu khoa học.
- Gia đình tôi, những người đã luôn bên tôi, giúp tôi vượt qua những khó khăn
trong thời gian qua.
Tác giả
LƯU XUÂN HÒA

iii
MỤC LỤC
MỞ ĐẦU .......................................................................................................................... 1
CHƯƠNG I. TỔNG QUAN NGHIÊN CỨU .................................................................... 2
1.1. Khái quát về kỹ thuật lai huỳnh quang tại chỗ (FISH) ............................................. 2
1.1.1. Lịch sử nghiên cứu, ứng dụng kỹ thuật FISH ................................................ 2
1.1.2. Trình tự đích và đầu dò trong phép lai huỳnh quang tại chỗ ......................... 6
1.1.3. Các loại tín hiệu đánh dấu huỳnh quang ...................................................... 9
1.1.4. Tình hình nghiên cứu ứng dụng kỹ thuật FISH trong vi tảo biển độc hại..... 13
1.2. Tình hình nghiên cứu ứng dụng kỹ thuật FISH tại Việt Nam................................. 17
1.3. Các bước cơ bản của quy trình lai huỳnh quang tại chỗ - FISH ............................. 18
1.4. Đặc điểm sinh thái của một số loài vi tảo biển độc hại nghiên cứu ........................ 19
CHƯƠNG II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU.................................... 21
2.1. Địa điểm thu mẫu và đối tượng nghiên cứu ................................................ 21
2.2. Phương pháp thu và lưu giữ mẫu ................................................................ 21
2.3. Phương pháp phân loại hình thái ................................................................ 23
2.4. Phương pháp kiểm tra phân loại bằng ADN và thiết kế đầu dò ................... 23
2.4.1. Phương pháp chuẩn bị ADN khuôn ........................................................... 23
2.4.2. Phương pháp Singel cell PCR ................................................................... 23
2.4.3. Phương pháp kiểm tra phân loại bằng ADN ............................................... 24
2.4.4. Phương pháp thiết kế đầu dò đặc hiệu........................................................ 25
2.5. Phương pháp lai huỳnh quang tại chỗ ........................................................ 25
CHƯƠNG III. KẾT QUẢ NGHIÊN CỨU VÀ THẢO LUẬN......................................... 27
3.1. Kết quả phân lập và phân loại các chủng vi tảo độc hại bằng hình thái ....... 27
3.1.1. Loài Alexandrium affine ............................................................................. 27
3.1.2. Loài A. pseudogonyaulax ........................................................................... 28
3.2. Ảnh hưởng của độ mặn tới nuôi sinh khối vi tảo tạo nguyên liệu ................ 29
3.3. Kết quả giải trình tự và phân loại bằng ADN các mẫu nghiên cứu .............. 31
3.4. Thiết kế đầu dò lai huỳnh quang tại chỗ ..................................................... 35
3.5. Tối ưu nhiệt độ lai và thử nghiệm đầu dò.................................................... 37
KẾT LUẬN..................................................................................................................... 43
KIẾN NGHỊ.................................................................................................................... 44
TÀI LIỆU THAM KHẢO ............................................................................................... 45
PHỤ LỤC ....................................................................................................................... 54

iv
DANH MỤC CHỮ VIẾT TẮT
TT Chữ viết tắt Nội dung tiếng Anh Nội dung tiếng Việt
1. ADN Deoxyribonucleic acid Axít deoxyribonucleic
2. ARN Ribonucleic acid Axít ribonucleic
3. Bp Base pair Cặp bazơ
4. FISH Fluorescent in situ hybridization Phép lai huỳnh quang tại chỗ
5. FITC Fluorescein isothiocyanate Chất phát tín hiệu huỳnh quang
6. ISH In situ hybridization Phép lai tại chỗ
7. ITS Internal transcribed spacer Vùng đệm phiên mã
8. LSU Large subunit Tiểu phần lớn
9. PCR Polymerase chain reaction Phản ứng chuỗi trùng hợp
10. PSP Paralytic shellfish poisoning Chất độc gây liệt cơ
11. rADN Ribosomal deoxyribonucleic
acid
Trình tự ADN mã hóa
ribosome
12. rARN Ribosomal ribonucleic acid ARN ribosome
13. SSU Small subunit Tiểu phần nhỏ

v
DANH MỤC CÁC BẢNG
Trang
Bảng 1.1: Một số chất huỳnh quang được dùng phổ biến trong kỹ
thuật FISH
10
Bảng 1.2: Trình tự đầu dò cho một số loài Alexandrium 16
Bảng 3.1: Một số điều kiện thích hợp cho nhân nuôi sinh khối tảo A.
affine và A. pseudogonyaulax trong phòng thí nghiệm
31

vi
DANH MỤC CÁC HÌNH
Trang
Hình 1.1. Các cách gắn nhãn tín hiệu vào đầu dò 12
Hình 1.2. Các bước cơ bản của kỹ thuật lai huỳnh quang tại chỗ FISH 19
Hình 2.1. Sơ đồ phản ứng PCR nhân vùng D1-D2 của rADN 23
Hình 3.1. Một số đặc điểm hình thái của A. affine 27
Hình 3.2. Một số đặc điểm hình thái của A. pseudogonyaulax 28
Hình 3.3. Ảnh hưởng của độ mặn tới sự sinh trưởng của tảo A. affine 30
Hình 3.4. Ảnh hưởng của độ mặn tới sự sinh trưởng của tảo A.
pseudogonyaulax
30
Hình 3.5. Kết quả điện di sản phẩm PCR lần 2 32
Hình 3.6. Cây phát sinh chủng loại của một số loài thuộc chi Alexandrium 34
Hình 3.7. So sánh sự khác biệt trình tự nucleotit của A. affine với các loài
Alexandrium
36
Hình 3.8. So sánh sự khác biệt trình tự nucleotit của A. pseudogonyaulax
với các loài Alexandrium
37
Hình 3.9. Tế bào A. affine dưới ánh sáng thường (trái) và phát huỳnh
quang dưới ánh sáng kích thích (phải) với điều kiện lai 43°C
38
Hình 3.10. Tế bào A. pseudogonyaulax dưới ánh sáng thường (trái) và
phát huỳnh quang dưới ánh sáng kích thích (phải) với điều kiện lai 43°C
38
Hình 3.11. Lai đầu dò A. affine với mẫu tự nhiên 40
Hình 3.12. Lai đầu dò A. pseudogonyaulax với mẫu tự nhiên 40

1
MỞ ĐẦU
Trong nghiên cứu phân loại sinh vật phù du biển nói chung và vi tảo biển độc
hại nói riêng, nhiều nhóm loài sinh vật rất khó hoặc hoàn toàn không thể phân loại
đến loài nếu chỉ bằng phương pháp so sánh hình thái. Sự giống nhau về nhiều đặc
điểm hình thái bên ngoài và khó nhận biết những đặc điểm khác nhau của chúng rất
dễ dẫn đến nhầm lẫn trong nghiên cứu phân loại. Cùng với số lượng loài rất lớn có
nhiều đặc điểm hình thái giống nhau, công tác nghiên cứu phân loại vi tảo biển
trong nhiều trường hợp gặp nhiều khó khăn, tốn thời gian. Những điều này đã gây
cản trở đối với công tác nghiên cứu hệ sinh thái biển, đánh giá và quản lý nguồn lợi
sinh vật biển nói chung và các nghiên cứu chuyên sâu về các nhóm loài nói riêng.
Kế thừa các đặc tính ưu việt của các kỹ thuật di lai các axit nucleic và trên nền
tảng của kỹ thuật lai tại chỗ (in situ hybridization - ISH), kỹ thuật lai huỳnh quang
tại chỗ (fluorescence in situ hydridization - FISH) đã được phát triển. Kể từ khi ra
đời, nó đã trở thành một công cụ mạnh trong nhiều nghiên cứu sinh học, không chỉ
các lĩnh vực sinh học phân tử và tế bào, mà cả trong các nghiên cứu về sinh thái
học, môi trường, trong chẩn đoán và nghiên cứu phát sinh loài. Kỹ thuật FISH ra
đời đã khắc phục được nhiều trở ngại trong nghiên cứu phân loại sinh vật. Các trở
ngại, như các vấn đề khó khăn do phân loại bằng hình thái, nhược điểm về thời gian
phân tích kéo dài, các yêu cầu đòi hỏi về lượng mẫu lớn hay các yêu cầu về môi
trường nuôi cấy đặc chủng để sàng lọc loài… có thể được khắc phục bằng kỹ thuật
FISH. Bên cạnh đó, bằng kỹ thuật này, các nhà nghiên cứu không chỉ phân tích định
tính mà còn có thể định lượng chính xác số lượng tế bào sinh vật, không gian phân
bố hay mối tương quan với môi trường của các vi sinh vật. Kỹ thuật FISH đặc biệt
hữu ích trong nghiên cứu phân loại sinh vật phù du trong môi trường biển nói chung
và các loài tảo độc hại nói riêng.
Nhằm từng bước ứng dụng kỹ thuật FISH để giải quyết những khó khăn trong
phân loại các loài vi tảo độc hại biển, chúng tôi tiến hành đề tài “Ứng dụng kỹ thuật
phân tử lai huỳnh quang (FISH) nhằm xác định nhanh một số loài tảo độc hại
biển Việt Nam”. Kết quả mong đợi của đề tài sẽ giúp cho việc nghiên cứu đánh giá,
quan trắc biến động các loài vi tảo độc hại, cảnh báo các hiện tượng ô nhiễm môi
trường biển (như thủy triều đỏ) trở nên thuận tiện, nhanh chóng và hiệu quả hơn.

2
CHƯƠNG 1. TỔNG QUAN NGHIÊN CỨU
1.1. Khái quát về kỹ thuật lai huỳnh quang tại chỗ (FISH)
1.1.1. Lịch sử nghiên cứu, ứng dụng kỹ thuật FISH
Kỹ thuật FISH được ra đời vào những năm 1980. Tuy nhiên, trước đó, khi
nghiên cứu tế bào, các nhà nghiên cứu thường sử dụng phương pháp nhuộm mô đơn
giản. Trong đó, các chất màu nhuộm tự nhiên hoặc tổng hợp đã được sử dụng để
nhuộm các cấu trúc hoặc các chất tích lũy trong tế bào. Nhưng, một nhược điểm lớn
là các chất này thường không đặc hiệu do chúng có những ái lực với những đặc tính
chung của các phần tử protein, axit nucleic, các lipit, cacbonhydrate. Chính điều này
đã cản trở cho các nghiên cứu chuyên sâu. Sau đó, các thuốc nhuộm đặc hiệu hơn
đối với các thành phần của tế bào hay các phức hợp phân tử lớn như hemosiderin,
amyloid, elastin đã được tìm thấy, phát triển và áp dụng trong việc nhuộm đặc hiệu
một số thành phần cấu trúc ở cấp độ phân tử và dưới tế bào. Dù đã có nhiều cải
thiện về tính đặc hiệu, nhưng các chất nhuộm này cũng không được áp dụng phổ
biến cho tất cả các đại phân tử sinh học được quan tâm. Khả năng phát hiện sự
giống và khác nhau của các đại phân tử đặc biệt ban đầu được dựa trên các phản
ứng kháng nguyên kháng thể. Vào những năm 1940, việc liên kết các kháng thể với
fluorochome nhưng không làm mất đi sự đặc hiệu liên kết thụ thể của chúng đã
được phát hiện. Trên cơ sở này, kỹ thuật sử dụng tín hiệu huỳnh quang phụ thuộc
kháng thể nhằm phát hiện các phân tử axit nucleic lai đã được Rudkin và Stollar
phát triển năm 1977 [58]. Tuy vậy, trước đó, kỹ thuật lai tại chỗ ISH (in situ
hybridization), tiền thân của kỹ thuật FISH, đã ra đời bởi nhóm các nhà khoa học
Pardue, Gall và John [54]. Trong phương pháp này, các thử nghiệm đã được tiến
hành với việc nhận biết được các trình tự ADN đích có trong noãn bào của chủng vi
khuẩn Xenopus bằng các đoạn đầu dò có bản chất là ADN hoặc ARN-28S đã được
đánh dấu bằng đồng vị phóng xạ. Sau đó chúng được ghi nhận bằng kỹ thuật phóng
xạ tự ghi. Kỹ thuật ISH này đã giúp các nhà khoa học có thể đưa các đoạn ngắn
ADN xâm nhập vào trong tế bào mà không làm chết, làm thay đổi hình thái của tế

3
bào hay tính toàn vẹn của các bộ phận bên trong tế bào. Trên cơ sở đó, một số cải
tiến kỹ thuật đã tiếp nối như việc ứng dụng các chất chỉ thị (reporter) dựa trên cơ sở
các enzym [33] hay sử dụng hệ thống đầu dò bằng vàng quan sát bằng kính hiển vi
điện tử [55]. Sau khi ra đời, ISH không ngừng được cải tiến nhằm nghiên cứu sự
tiến hóa của nhiễm sắc thể, phân tích các nhiễm sắc thể của các khối u, tế bào ung
thư bạch cầu và nghiên cứu di truyền học tế bào ở các loài.
Tuy nhiên, một số nhược điểm dễ nhận thấy của phương pháp ISH cũng đã
bộc lộ. Đầu tiên, rất nhiều vật liệu phóng xạ tự nhiên sử dụng cho đầu dò là kém
bền. Do các đồng vị phân rã theo thời gian nên hoạt động của đầu dò dễ bị thay đổi.
Thứ hai, mặc dù thông thường độ nhạy của phóng xạ tự ghi là cao nhưng mức độ
phân giải lại bị hạn chế. Thứ ba, để tạo ra tín hiệu có thể đo được trên film chụp X
quang thì mẫu cần phải có thời gian phơi nhiễm dài. Điều này làm chậm tiến trình
thí nghiệm, phân tích kết quả, gia tăng nguy cơ phơi nhiễm với người sử dụng.
Ngoài ra, vì kỹ thuật này đòi hỏi sử dụng nguyên liệu là các đồng vị phóng xạ nên
các đầu dò phóng xạ có giá thành cao; mặt khác đòi hỏi chúng phải được vận
chuyển, thao tác, lưu trữ và loại bỏ theo những quy trình rất nghiêm ngặt, phức tạp.
Vì những lý do đó, dù có nhiều ưu điểm nổi trội nhưng những hạn chế cơ bản trên
đã yêu cầu các nhà nghiên cứu cần phải tìm ra một biện pháp hiệu quả hơn. Có lẽ vì
vậy mà kỹ thuật ISH (với các đầu dò mang đồng vị phóng xạ) sau đó đã nhanh
chóng được thay thế bởi một kỹ thuật mới có nhiều ưu điểm hơn mang tên kỹ thuật
lai tại chỗ sử dụng các đầu dò axit nucleic có gắn tín hiệu huỳnh quang
(Fluorescence in situ hybridization - FISH). Với kỹ thuật FISH, các trở ngại về thời
gian thí nghiệm, độ phân giải và tính an toàn được giải quyết khá triệt để, mở đường
cho sự phát hiện đồng thời nhiều mục tiêu, phân tích định lượng và cho những hình
ảnh tế bào sống động.
Những nghiên cứu thử nghiệm ứng dụng tín hiệu huỳnh quang đầu tiên trong
phương pháp lai tại chỗ bắt đầu từ những năm 1980. Trong một nghiên cứu, các nhà
khoa học đã tiến hành gắn phân tử ARN trực tiếp với tín hiệu huỳnh quang ở đầu 3’
và thử nghiệm dùng nó là một đầu dò cho trình tự ADN đặc hiệu. Kết quả cho thấy

4
rằng so với các đầu dò phóng xạ, đầu dò huỳnh quang sử dụng an toàn hơn, ít gây
hại đối với môi trường và sức khỏe con người, đồng thời kết quả thu được cũng
chính xác hơn [18]. Ở những nghiên cứu khác, các đầu dò gắn biotin và các tín hiệu
streptavidin kết hợp huỳnh quang đã được dùng cho việc phát hiện các trình tự
ADN đích đặc trưng trên nhiễm sắc thể [46] và các trình tự trên mARN [60]. Năm
1989, DeLong cũng đã tiến hành thử nghiệm sử dụng các đầu dò oligonucleotit (17
- 34 nucleotit) được đánh dấu huỳnh quang để dò tìm các vi khuẩn E.coli dựa vào
trình tự đích là đoạn ARN 16s [24]. Nhiều nghiên cứu cũng chỉ ra rằng, việc sử
dụng nhiều đầu dò khác nhau cho phép xác định được các loại tế bào khác nhau
trong cùng một nhóm và thậm chí với việc phân tích cường độ huỳnh quang cũng
cho thấy được giai đoạn và tốc độ phát triển của các tế bào vi sinh vật [19, 24].
Với khả năng ứng dụng rộng rãi của kỹ thuật FISH, ngày càng nhiều nghiên
cứu được tiến hành trên các lĩnh vực khoa học. Sự phát triển ngày càng cao của
nhiều kỹ thuật phân tích đã cho thấy rằng số lượng loài có trong tự nhiên, nhất là
đối với nhóm vi khuẩn, có con số vượt xa so với số lượng các loài đã được phát
hiện và mô tả trước đó. Do đó, sự đa dạng thành phần loài vi sinh vật trong thời
gian trước đây đã từng bị đánh giá thấp. Các phương pháp cổ điển trước đây hầu
như không thể mô tả được chính xác và đầy đủ rất nhiều loài vi sinh vật. Rất nhiều
loài sau này được phát hiện mới bằng các phương pháp hiện đại và chính xác hơn,
như phương pháp FISH. FISH có khả năng đưa ra những hình ảnh chi tiết của môi
trường vi sinh mà không cần phải qua những bước làm sạch hay khuếch đại. Chính
vì vậy mà nó đã được sử dụng rộng rãi trong các lĩnh vực nghiên cứu khác nhau về
môi trường. Phương pháp này hiện nay được ứng dụng nhiều trong các nghiên cứu
về sự đa dạng sinh học của các quần thể vi sinh vật ở nhiều điều kiện địa lý khác
nhau trong tự nhiên, Llobet (1998) đã sử dụng kỹ thuật lai huỳnh quang tại chỗ như
một công cụ để các nghiên cứu về hệ vi sinh vật có trong trầm tích biển. Kết quả
cho thấy, gần 45% số tế bào vi khuẩn có trong đó thuộc nhóm ngành đã biết với
nhóm Cytophaga-Flavobacterium có trong hầu hết các lớp trầm tích [45]. Hay các
nghiên cứu sự phân bố của vi khuẩn bằng kỹ thuật FISH trong đại dương [31], trong

5
các tầng nước khác nhau tại vịnh Mariager Fjord, Đan Mạch [53], những nghiên
cứu hệ vi khuẩn ở trong sông [42], trong thủy vực trên núi cao có băng bao phủ
quanh năm [11], hay các nghiên cứu về Bacillus trong đất [26]…
Bên cạnh đó, kỹ thuật FISH cũng được ứng dụng rộng rãi trong y học.
Gersdorf (1993) đã tiến hành nghiên cứu hỗn hợp vi khuẩn phổ biến trong khoang
miệng người bằng phương pháp FISH qua đó đã tìm thấy có hơn 300 loài vi khuẩn
khác nhau, trong đó có nhiều loài rất phức tạp khi tiến hành nuôi cấy hoặc chưa
từng được nuôi cấy. Các bệnh về răng miệng như viêm răng hay viêm lợi có liên
quan đến các mạng lưới vi khuẩn đặc thù này. Trong khi một số kỹ thuật nuôi cấy
chuyên biệt chỉ thu được rất ít thành phần loài trong sự đa dạng nhóm vi sinh vật
này. Những lợi ích khi sử dụng kỹ thuật FISH đã được thể hiện rõ trên những vi
khuẩn kị khí gram âm, như Porphyromonas gingivalis, Bacterroides forsythus và
Prevotella intermedia có trong thành phần mảng bám răng của bệnh nhân viêm răng
[28, 29]. Cũng thông qua kỹ thuật FISH, các phân tích sâu hơn đã cho thấy một số
lượng lớn và đa dạng về hình thái của các loại xoắn khuẩn trên các vết bẩn của
mảng bám răng có trong khoang miệng của các bệnh nhân cùng với sự phát triển
nhanh chóng của bệnh viêm răng [21, 52]. Trong một thử nghiệm lâm sàng với các
mẫu đờm và mẫu gạc họng, một tập hợp các đầu dò oligonucleotit được thiết kế để
dò tìm đặc hiệu đối với các mầm bệnh phân lập từ tế bào xơ gan của bệnh nhân
được tiến hành. Kết quả cho thấy, các đầu dò đã phát hiện nhanh chóng và chính
xác các vi khuẩn gây cấp tính ở bệnh nhân xơ nang, bao gồm Pseudomonas
aeruginosa, Stenotrophomonas maltophilia, Pseudomonas aeruginosa,
Burkholderia cepacia, Haemophilus influenzae, Streptococcus pyogenes,
Staphylococcus aureus và Candida albicans. So với các kỹ thuật nuôi cấy thông
thường, FISH chứng tỏ là công cụ chính xác và hiệu quả [33, 34]. Ở một nghiên cứu
y học khác, khả năng ứng dụng của FISH để định danh các vi sinh vật gây bệnh
trong các mẫu máu nhiễm bệnh đã được khảo sát. Các đầu dò oligonucleotit sử
dụng đặc hiệu cho từng mầm bệnh điển hình đã cho thấy hiệu quả khi chúng dò tìm
thấy các nhóm gây bệnh bao gồm Staphylococci, Streptococci, Enterococci,

6
Enterobacteriaceae và nấm trên các bệnh nhân nhiễm bệnh. Các nhà nghiên cứu kết
luận rằng phương pháp FISH có độ nhạy và đặc hiệu tuyệt vời và kết quả thu được
chỉ trong 25 phút cho tới 2,5 giờ. Trong khi đó, việc định danh các loài bằng
phương pháp nuôi cấy truyền thống thường mất từ 24 đến 72 giờ mới hoàn thành.
Sự phát hiện nhanh chóng các nguyên nhân mầm bệnh trong máu bằng phương
pháp FISH cho phép các bác sỹ dễ dàng đưa ra được các phương pháp trị liệu thích
hợp và do đó có thể hoàn thiện việc chẩn đoán cho các bệnh nhân bị nhiễm trùng
máu [37, 41].
Cho đến nay, với các ưu thế về độ nhạy, tiết kiệm thời gian, đơn giản… kỹ
thuật FISH đã và đang là một công cụ hữu dụng trong những ứng dụng về nghiên
cứu sinh thái, đa dạng sinh học, y học, di truyền học và nhiều lĩnh vực khác.
1.1.2. Trình tự đích và đầu dò trong phép lai huỳnh quang tại chỗ
Bản chất của kỹ thuật FISH là sự lai chính xác một đoạn oligonucleotit có
gắn tín hiệu huỳnh quang (đầu dò) với một trình tự ADN đích đặc trưng. Với nhiều
mục đích khác nhau mà người ta có thể sử dụng kỹ thuật FISH nhằm dò tìm hay
phát hiện các trình tự ADN hay ARN đích thuộc các phần khác nhau của hệ gen
quan tâm. Các trình tự này có thể là một trình tự đặc trưng nằm trên nhiễm sắc thể
mã hóa cho một chức năng gen nào đó cần quan tâm. Với các đầu dò đặc hiệu cho
từng locut gen, chúng được tạo ra từ một phần của gen. Do vậy, khi lai, chúng sẽ
kết hợp bổ sung với phần đặc hiệu đó của gen trên nhiễm sắc thể. Từ đó người ta có
thể xác định được chính xác vị trí trên nhiễm sắc thể mà gen đó tồn tại. Đối với các
trình tự đích là những đoạn lặp vùng tâm động hay vùng Alphoid, đầu dò được thiết
kế sẽ cho phép chúng ta tìm thấy vùng tương ứng bổ sung tại tâm động của các
nhiễm sắc thể. Trong nhiều trường hợp, các nhiễm sắc thể được đánh dấu bởi các
màu huỳnh quang khác nhau của đầu dò. Từ đó người ta có thể xác định được cá thể
có mang đúng số lượng các nhiễm sắc thể hay không. Đối với phép lai toàn bộ
nhiễm sắc thể, tập hợp những đầu dò nhỏ được sử dụng để lai cho từng đoạn ngắn
trình tự đích dọc trên nhiễm sắc thể. Nhiễm sắc thể phát quang với các màu sắc

7
khác nhau của đầu dò giúp người ta có thể phân biệt được dễ dàng các nhiễm sắc
thể khác nhau thay vì phải căn cứ vào các vạch sáng tối trên nhiễm sắc thể theo các
phương pháp truyền thống. Mặt khác, chúng cũng giúp cho việc nhận biết dễ dàng
những bất thường trong cấu trúc của nhiễm sắc thể.
Trong một nghiên cứu của mình về bệnh bạch cầu (ABL), Michael đã tiến
hành thử nghiệm hệ thống tổ hợp các đầu dò oligonucleotit (COMBO) thông qua kỹ
thuật FISH nhằm nhận diện vùng ABL (gây bệnh bạch cầu) thuộc nhiễm sắc thể số
9 của người. Kết quả cho thấy hệ thống COMBO hoạt động rất hiệu quả và có thể
coi đây như một cách tiếp cận mới trong việc gắn nhãn đặc hiệu tại những vị trí cần
quan tâm của hệ gen người [48]. Các kết quả nghiên cứu thử nghiệm khác cũng chỉ
ra rằng để quá trình lai bổ sung được hiệu quả thì thông thường các trình tự này phải
ở giai đoạn chuẩn bị của nhiễm sắc thể trong nguyên phân hoặc trong giai đoạn gian
kỳ của tế bào [18, 19].
Mở rộng nghiên cứu, các nhà khoa học cũng đã sử dụng các tiểu phần ARN
ribosome (5s, 16s, 23s hoặc 28s) như các trình tự đích trong các thí nghiệm sử dụng
kỹ thuật FISH. Năm 1994, Trebesius cùng các đồng nghiệp của mình đã ứng dụng
phát triển kỹ thuật FISH trong việc xác định các tế bào vi khuẩn bằng việc sử dụng
các đầu dò polynucleotit cho các trình tự đích 23s ARN ribosome của vi khuẩn. Các
đầu dò này tỏ ra rất hiệu quả trong việc phát hiện các đối tượng vi khuẩn cần quan
tâm [64]. Trong một nghiên cứu khác, các trình tự thuộc các vùng V2, V4, V8 của
16s ribosome đã được các nhà khoa học sử dụng làm các trình tự đích cho các đầu
dò huỳnh quang trong phép lai tại chỗ nhằm định lượng nhóm vi khuẩn
Bifidobacterium spp. cộng sinh trong đường tiêu hóa của người [44]. Với mỗi mục
tiêu nghiên cứu, các vùng khác nhau của các tiểu phần ribosome ngày càng được
khai thác và sử dụng rộng rãi trong các ứng dụng kỹ thuật FISH khác nhau [18, 27,
36, 68]. Trong nhiều nghiên cứu về phân loại, các trình tự đích được sử dụng phổ
biến trong FISH là các rRNA 16S vì chúng ổn định về mặt di truyền, số lượng bản
sao nhiều.

8
Nhiều kết quả khác cũng đã cho thấy, việc sử dụng thành phần đích là phân
tử mARN đặc hiệu cũng có thể giúp ích trong nhiều nghiên cứu có liên quan vì
mARN là một mắt xích quan trọng trong chuỗi hoạt động sinh hóa và di truyền của
tế bào và của cơ thể [18, 25, 65].
Cùng với việc khai thác có hiệu quả và đa dạng các trình tự đích, các loại đầu
dò phục vụ trong kỹ thuật FISH cũng được nghiên cứu chế tạo và cải tiến tùy theo
từng mục tiêu. Một số nghiên cứu đã sử dụng các đầu dò là các đoạn polynucleotit.
Đây là những đoạn polynucleotit có kích thước lớn từ 200 – 300 nucleotit. Trong
một số nghiên cứu, người ta đã sử dụng chúng cho việc dò tìm các trình tự đích là
vùng siêu biến III thuộc 23s ARN ribosome của các vi khuẩn. Những đầu dò
polynucleotit này đã thể hiện tính hữu dụng trong việc phát hiện Pseudomonas
putida, Acinetobacter spp. [64] và nhóm vi khuẩn và vi khuẩn cổ phù du trong
nước biển [25]. Trong một nghiên cứu khác, bảy đầu dò polynucleotit đã xây dựng,
thiết kế nhằm nhận biết các loài Acinetobacte baumannii, A. calcoaceticus, A.
haemolyticus, A. junii, A. radioresistens, A. lwoffii, A. johnsonii cũng dễ dàng được
ứng dụng thông qua kỹ thuật FISH [39].
Bên cạnh đó, các đoạn oligonucleotit với kích thước chỉ khoảng 15 - 30
nucleotit có gắn các nhãn tín hiệu khác nhau cũng được nghiên cứu thiết kế và sử
dụng làm đầu dò tìm các thành phần đích là phân tử mARN, rARN đặc hiệu trong
nhiều nghiên cứu [25, 65]. Xét về tính hiệu quả, trong kỹ thuật lai huỳnh quang tại
chỗ nguyên vẹn tế bào, các đầu dò oligonucleotit với trình tự ngắn lại có hiệu quả
tốt hơn trong khả năng xâm nhập vào tế bào và lai đặc hiệu với trình tự đích. Đối
với các trình tự đích thuộc các tiểu phần ribosome 5s, 16s, 23s hoặc 28s của vi
khuẩn, các loài thông thường được nhận diện bằng các đầu dò oligonucleotit. Chính
vì vậy mà các nghiên cứu ứng dụng kỹ thuật FISH với đầu dò oligonucleotit cho các
mục đích xác định vi khuẩn, phân tích cấu trúc nhóm vi khuẩn, xem xét các mối
quan hệ tương tác về không gian và thời gian của các quần thể vi sinh vật trong môi
trường sống của chúng đã được nhiều nhà khoa học quan tâm trong thời gian qua.
Trên thế giới, kỹ thuật FISH cũng đã được các nhà khoa học áp dụng trên các đối

9
tượng là những sinh vật phù du nước ngọt cũng như các loài phù du sống ven biển,
ngoài khơi và cả những loài thu được từ trầm tích đáy biển. Kỹ thuật FISH đã cho
thấy được ưu thế trong khả năng xác định nhanh chóng và chính xác số lượng rất
nhỏ các tế bào vi khuẩn cũng như có khả năng phát hiện được sự đa dạng phong
phú của các quần thể sinh vật này [11, 18, 31, 46].
1.1.3. Các loại tín hiệu đánh dấu huỳnh quang
Trong lịch sử phát triển kỹ thuật FISH, nhiều biến thể của tín hiệu (thuốc
nhuộm) gắn trực tiếp hoặc gián tiếp đã được đưa ra. Điều này cho phép các nhà
nghiên cứu có thể lựa chọn, tùy theo độ đặc hiệu, độ nhạy, độ phân giải, màu sắc
phát quang của đầu dò để thiết kế các đầu dò phù hợp với các điều kiện thí nghiệm
khác nhau. Đồng thời, việc đa dạng hóa các loại tín hiệu đánh dấu (đa dạng màu
sắc) sẽ làm cho hình ảnh của một kết quả lai huỳnh quang trở nên sinh động. Hình
ảnh thu được có thể minh họa rõ ràng cho việc phân tích sự tổ hợp của hệ gen về
mặt không gian hay dễ dàng trong việc phát hiện nhiễm sắc thể và sự sai lệch của
chúng trong chu trình tế bào. Các loại thuốc nhuộm huỳnh quang thường được gắn
với đầu dò bằng các liên kết cộng hóa trị. Chúng thường có các mức năng lượng
kích thích và phát xạ tối đa khác nhau, vì vậy cho phép phát hiện đồng thời hai hoặc
nhiều vi sinh vật bằng một lần tiến hành lai phân tử duy nhất đồng thời cho ra
những hình ảnh màu sắc sinh động. Các chất nhuộm thường sử dụng cho FISH
trong phân loại vi sinh vật là các dẫn xuất của fluorescein (như Fluorescein-
Isothiocyanate (FITC), 5(6)carboxyfluorescein-N-hydroxysuccimide-ester (FluoX))
hoặc dẫn xuất của rhodamine (như Tetramethyl-Rhodamine-Isothiocyanate
(TRITC), Texas Red), ngoài ra còn có các loại thuốc nhuộm cyanine khác như là
Cy3 và Cy5… [12, 39].

10
Bảng 1.1. Một số chất huỳnh quang được dùng phổ biến trong kỹ thuật FISH [18]
Bước sóng Chất
Huỳnh quang Màu Kích thích cực đại
(nm)
Phát xạ
(nm)
Alexa488 Xanh lá cây 493 517
AMCA Xanh da trời 399 446
CY3 Đỏ 552 565
CY5 Đỏ 649 670
CY7 Tím 743 767
DAPI Xanh da trời 350 456
Fluoresscein Xanh lá cây 494 523
Rodamine Đỏ 555 580
TAMRA Đỏ 543 575
Texas Red Đỏ 590 615
TRITC Đỏ-cam 550 580
Trong kỹ thuật FISH người ta có nhiều cách để đánh dấu đầu dò. Tuy vậy,
việc đánh dấu trực tiếp chất nhuộm huỳnh quang lên đầu dò là cách thông dụng,
nhanh và rẻ nhất. Đồng thời, đầu dò được chuẩn bị theo cách này dễ dàng được phát
hiện vì không cần phải tiến hành các bước dò tìm sau quá trình lai. Trong đánh dấu
đầu dò, một hoặc nhiều phân tử thuốc nhuộm huỳnh quang được liên kết trực tiếp
với chuỗi polynucleotit/oligonucleotit trong suốt quá trình tổng hợp thông qua liên
kết amino ở đầu 5’ của đầu dò. Một cách khác, người ta cũng có thể sử dụng enzym
Terminal transferase để gắn các nucleotit đã được đánh dấu huỳnh quang vào đầu 3’
của đầu dò [52]. Ngoài ra, một số thuốc nhuộm như Flourescein-Isothiocyanate
(FITC) nếu được gắn vào đầu dò theo một dải thì có thể làm tăng cường độ tín hiệu
so với các đầu dò được đánh dấu trực tiếp [23]. Sự tăng lên về tín hiệu huỳnh
quang cũng được biểu hiện trên những đầu dò được gắn chất huỳnh quang trên cả
hai đầu 3’ và 5’ [62]. Trong nghiên cứu của mình, Trebesius cùng các đồng nghiệp
(1994) đã sử dụng các đầu dò polynucleotit cho các trình tự đích 23s ARN ribosome

11
của vi khuẩn. Các đầu dò này không gắn tín hiệu phóng xạ mà được thay bằng dẫn
xuất phát huỳnh quang của UTP (fluorescein-12-UTP, digoxigenin-11-UTP, 7-
amino-4-methyl-coumarin- 3-acetyl-6-UTP, hoặc tetramethylrhodamine-6-UTP).
Kết quả nghiên cứu cũng chỉ ra rằng, với đầu dò dạng này thì hiệu quả mang tín
hiệu huỳnh quang tăng gấp khoảng 26 lần so với đầu dò ngắn oligonucleotit [64].
Tuy vậy, ở một một số trường hợp, việc dò tìm theo cách gián tiếp tỏ ra có
hiệu quả hơn. Sự tăng độ nhạy của thí nghiệm FISH bằng cách kết hợp các đầu dò
với những chất chỉ thị như Digoxygenin (DIG) đã được thực hiện và sau đó phức
hợp này sẽ được dò tìm thông qua một chất phản huỳnh quang [69]. Ngoài ra,
người ta có thể tăng độ nhạy của FISH bằng việc sử dụng enzym nhằm khuếch đại
tín hiệu của đầu dò trong nhiều nghiên cứu định danh vi khuẩn. Ban đầu, mỗi
oligonucleotit vẫn sẽ được đánh dấu bằng digoxygenin như trên, sau đó được dò
tìm bằng một thể kháng digoxygenin, kháng thể này tiếp tục được liên kết với
enzym phosphatase kiềm. Enzym này chuyển hóa HNPP (2-hydroxy-3-naphtoic
acid-2’-phenylanilide phosphate) thành dạng Dephosphoryl, chất này tạo ra màu
huỳnh quang đỏ sáng khi kết hợp với Fast Red TR. Tín hiệu huỳnh quang thu được
theo cách này có cường độ mạnh hơn 8 lần so với tín hiệu của các oligonucleotit chỉ
gắn nhãn huỳnh quang duy nhất [40, 67].
Sau này, phương pháp khuếch đại tín hiệu bằng enzym ngày càng được cải
tiến tốt hơn nhờ vào một kỹ thuật được gọi là “hệ thống khuếch đại tín hiệu
Tyramide” (TSA). Hệ thống này làm tăng cường độ tín hiệu lên khoảng 10 – 20 lần
so với ban đầu. Tuy nhiên các nghiên cứu cũng chỉ ra rằng, do quá trình xâm nhập
của các chất cao phân tử vào trong tế bào vi khuẩn bị hạn chế hơn những chất
nhuộm thông thường nên số lượng các tế bào được lai thành công đã giảm đi một
cách rõ ràng so với việc sử dụng đầu dò thông thường được đánh dấu duy nhất một
chất huỳnh quang. Mặc dù enzym lysozyme đã giúp cải thiện tính thấm của các đầu
dò vào trong tế bào, dẫn tới cường độ tín hiệu được nâng cao, nhưng phương pháp
này dường như không áp dụng được với một số loài vi khuẩn Gram dương. Trong
một số trường hợp, việc kết hợp sử dụng các đầu dò polyribonucleotit, được đánh

12
dấu với digoxygenin, song song với việc sử dụng hệ thống TSA là một giải pháp
tốt. Sự kết hợp này đã được ứng dụng thành công khi dò tìm và phát hiện trực quan
các loài Listeria gây độc [65]. Về khía cạnh khác, các đầu dò oligonucleotit với
trình tự ngắn lại có hiệu quả tốt hơn trong khả năng xâm nhập vào tế bào và lai đặc
hiệu với trình tự đích so với các đầu dò polynucleotit.
Để tăng độ nhạy của đầu dò trong kỹ thuật FISH, việc tính toán đến số lượng
bản sao của trình tự đích cũng là một giải pháp được tính đến nhằm khuếch đại tín
hiệu để có được hình ảnh rõ nét hơn. Trong nhiều trường hợp, các trình tự đích là
các rARN hay các mARN (thường có nhiều trong tế bào) được lựa chọn để sử dụng
với các đầu dò oligonucleotit mang được ít tín hiệu đánh dấu [18].
Hình 1.1. Các cách gắn nhãn tín hiệu vào đầu dò [18].
Gắn nhãn trực tiếp (a, b) và gián tiếp (c–e) của đầu dò sử dụng 1 phân tử chỉ
thị như digoxygenin (DIG) mà nó sau đó được phát hiện bởi kháng thể huỳnh
quang, horseradish peroxidase (HRP) sử dụng phức hợp fluorescein–tyramide như
một cơ chất cho sự khuếch đại tín hiệu bằng enzym theo hệ thống TSA, hoặc sự kết
hợp sử dụng các đầu dò polyribonucleotit với hệ thống TSA.
Như vậy, bằng kỹ thuật FISH, các nhà nghiên cứu không chỉ có thể phân biệt
được các vi sinh vật quen thuộc mà còn xác định được các vi sinh vật mới, chưa biết
Gắn nhãn vào đầu 5’ (a)
Gắn nhãn vào đầu 3’ (b)
Sử dụng chất chỉ thị (c)
Sử dụng enzym (d)
Đầu dò dạng polynucleotit (e)

13
rõ về điều kiện và môi trường nuôi cấy. Đồng thời, kỹ thuật FISH có thể giúp chúng
ta phân tích được sự phân bố không gian của chúng và biết rõ về hệ thống phức tạp
của các quần thể vi sinh vật. Việc xác định số lượng, cường độ các tín hiệu phát ra
bằng các oligonucleotit của rARN đích cho phép ước tính được tỷ lệ tăng trưởng
của các tế bào riêng lẻ [38]. Cho đến nay, độ nhạy và tốc độ thực hiện là những đặc
điểm nổi bật giúp kỹ thuật lai huỳnh quang tại chỗ - FISH trở thành công cụ rất hữu
hiệu cho nhiều nghiên cứu về di truyền, sinh thái, môi trường. Qua quá trình phát
triển và cải tiến, cho đến nay chúng được xem là một trong những công cụ di truyền
phân tử ứng dụng rộng rãi trong nghiên cứu về sinh thái học, môi trường học, chẩn
đoán bệnh và phát sinh loài.
1.1.4. Tình hình nghiên cứu ứng dụng kỹ thuật FISH trong vi tảo biển độc hại
Vi tảo biển (Microalgae) là bộ phận sinh vật có thành phần rất phong phú
trong hệ sinh thái thủy sinh của biển. Trong chuỗi thức ăn, vi tảo biển là sinh vật
sản xuất sơ cấp, là một mắt xích thức ăn, nguồn cung cấp dinh dưỡng của nhiều loài
động vật phù du, ấu trùng giáp xác và nhiều loài động vật khác [7]. Bên cạnh chức
năng quan trọng đó, việc bùng phát hoặc tiết các chất gây độc của một số loài vi tảo
biển lại có thể gây nguy hại cho các loài thủy sinh, thậm chí gây nguy hiểm đến tính
mạng con người và gây ô nhiễm môi trường, phá hủy cân bằng sinh thái. Điển hình
cho những tác hại này chính là hiện tượng thủy triều đỏ hay tảo độc nở hoa, đã được
ghi nhận ở nhiều vùng biển khác nhau trên thế giới, trong đó có Việt Nam. Đây
cũng là nguyên nhân làm chết nhiều loài sinh vật biển trong tự nhiên và nuôi trồng
thủy sản.
Một số chi tảo điển hình cho hiện tượng thủy triều đỏ là Alexandrium và
Pseudo-nitzschia. Đây là những chi tảo biển có phân bố rộng, có thể bắt gặp ở nhiều
vùng biển khác nhau ở Việt Nam và trên thế giới. Chúng có khả năng sản sinh các
độc tố gây liệt cơ (PSP) và có thể gây chết người [45]. Mặt khác, rất nhiều loài vi
tảo có cấu tạo phức tạp và rất giống nhau về hình thái ngoài, do vậy thường dẫn đến
sự lẫn lộn về danh pháp giữa các loài trong cùng một chi. Điều này dẫn đến những

14
khó khăn rất lớn cho việc quan trắc, cảnh báo nguy cơ bùng phát và hạn chế những
tác hại của chúng trong tự nhiên. Tuy nhiên, kỹ thuật FISH ra đời và ngày càng
hoàn thiện đã phần nào giúp các nhà khoa học dễ dàng hơn trong việc rà soát và
phát hiện nhanh tảo độc hại và quan trắc, cảnh báo môi trường một cách hiệu quả
hơn. Phương pháp này thường được sử dụng trong các nghiên cứu quan trắc môi
trường biển cho các loài vi tảo độc hại, đặc biệt khi có những biến động bất thường
về môi trường và sự nở hoa vi tảo biển [59].
Một trong những thử nghiệm đầu tiên sử dụng các đầu dò phân tử như một
công cụ để phân loại các loài tảo độc hại được tiến hành bởi Anderson (1995). Kết
quả nghiên cứu này cho thấy độ nhạy của đầu dò lai huỳnh quang trong việc phát
hiện Pseudonitszchia pungens là rất tốt. Sự kết hợp giữa 2 loại đầu dò huỳnh quang
và đầu dò kháng thể làm tăng đáng kể hiệu quả hình ảnh thu được [15]. Năm 1996,
Adachi và đồng nghiệp đã sử dụng các kỹ thuật phân tử để phân loại hai loài A.
tamarense và A. catenella thuộc chi Alexandrium. Đầu dò ADN được thiết kế đối
với trình tự của ADN mã hóa ribosome (rADN) tại vùng ITS. Nhóm đối tượng được
tiến hành thử nghiệm bao gồm A. affine, A. insuetum, A. pseudogonyaulax, A.
lusitanicum, A. fraterculus, Gymnodinium mikimotoi, Prorocentrum micans,
Amphidinium carterae, Heterocapsa triyuetra, Heterosigma akashiwo, Chattonella
antiyua và Skeletonema costatum. Kết quả cho thấy, hai đầu dò cCAT-F1 và
cTAM-F1 chỉ bắt cặp tương ứng với các chủng khác nhau thuộc hai loài
Alexandrium catenella và A. tamarense. Ngoài ra, chúng không hề có phản ứng với
các chủng tảo độc khác hoặc xảy ra phản ứng chéo trong cùng điều kiện lai. Kết quả
này chứng tỏ, cCAT-F1 và cTAM-F1 có tính đặc hiệu cao với các loài Alexandrium
catenella, A. tamarense và có thể dùng làm đầu dò đặc hiệu để nhận dạng hai loài
vi tảo biển này [10].
Một số công bố sau đó cũng cho thấy phương pháp FISH có khả năng áp
dụng tốt trong việc phân loại các loài đặc hiệu ngay cả trong mẫu nuôi cấy và mẫu
ngoài tự nhiên [10, 59]. Cùng thời gian này, Miller và Scholin đã ứng dụng FISH
với việc sử dụng đầu dò huỳnh quang đặc hiệu rARN để phân loại chi Pseudo-

15
nitzschia. Tám đầu dò phân tử đặc hiệu (auD1, puD1, muD1, muD2, heD2-2, frD1,
deD1, và amD1) được thiết kế và sử dụng để phân loại một cách dễ dàng tám loài
khác nhau, đó là: P. australis, P. pungens, P. multiseries, P. heimii, P. fraudulenta,
P. delicatisima, P. pseudodelicatisima và P. americana. Kết quả cũng cho thấy tính
tiện dụng, chuẩn xác và tiết kiệm thời gian của phương pháp phân loại bằng các đầu
dò huỳnh quang hơn nhiều so với các phương pháp truyền thống [50, 51]. Ở một
nghiên cứu khác, kỹ thuật FISH cũng được ứng dụng thử nghiệm để nhận dạng 10
chủng tảo độc thuộc một số loài Pseudo-nitzschia được thu thập ở nhiều vùng khác
nhau trên thế giới (Nhật Bản, Mĩ và Canada) với trình tự đích là ARN. Kết quả cho
thấy, đầu dò có tín hiệu rất mạnh đối với các chủng Pseudo-nitzschia multiseries và
P. pungens, thu thập từ Mĩ và Canada. Phép lai phân tử cũng cho thấy tín hiệu
huỳnh quang giảm nhẹ khi tế bào nuôi cấy ở giai đoạn muộn chứng tỏ lượng ARN
đã giảm trong tế bào. Việc định lượng tế bào Pseudo-nitzschia bằng sử dụng kết
hợp kỹ thuật FISH cũng trở nên dễ dàng và nhanh chóng hơn [59]. Khả năng ứng
dụng kỹ thuật lai huỳnh quang tại chỗ đối với các mẫu Pseudo-nitzschia đã được cố
định bằng hóa chất bảo quản cũng đã được thử nghiệm. Các mẫu vi tảo cố đã được
cố định bằng êtanol cũng đã có phản ứng dương tính đối với thí nghiệm lai. Tuy
nhiên, độ nhạy của tín hiệu huỳnh quang thu được tùy thuộc vào thời gian cố định
và một số điều kiện bảo quản của mẫu trước khi thí nghiệm. Đối với các mẫu sau
khi lai với đầu dò huỳnh quang được lưu giữ ở 4oC sẽ duy trì được sự phát quang
trong thời gian dưới 1 tuần [49]. Trong những nghiên cứu khác, các các tiểu đơn vị
của ribosome cũng đã được lấy làm các trình tự đích cho việc quan trắc Pseudo-
nitzschia thu thập từ vịnh Chinhae Bay (Hàn Quốc). Kỹ thuật FISH với các đầu dò
đặc hiệu đã được sử dụng gồm muD1, puD1, auD1, heD2-2, frD1, deD1, amD1.
Nghiên cứu này cũng cho thấy, một số đầu dò cho tín hiệu huỳnh quang yếu hơn
các đầu dò khác. Điều này được giải thích là liên quan đến trạng thái sinh lí của tế
bào [20].
Sau này, việc sử dụng các đầu dò phân tử phát huỳnh quang trong kỹ thuật
FISH để nhận dạng và phát hiện các loài tảo độc hại ngày càng phổ biến hơn. Đầu

16
dò được thiết kế dựa trên trình tự đích thuộc vùng D1- D2 của tiểu đơn vị lớn LSU
rADN của các loài vi tảo đã được thử nghiệm trong phép lai FISH đặc hiệu trên tế
bào nguyên vẹn. Các đầu dò ADN đặc hiệu cho 6 loài Alexandrium: A. tamarense,
A. catenella, A. affine, A. fraterculus, A. insuetum và A. pseudogonyaulax đã cho kết
quả tốt và không biểu hiện ở các phản ứng lai với các loài Alexandrium khác [43].
Cũng nghiên cứu về một số loài thuộc chi Alexandrium, một số nhà khoa học khác
cũng đã thành công trong việc phát triển phương pháp lai huỳnh quang tại chỗ với
các đầu dò được thiết lập từ các trình tự đích thuộc rARN có trong tế bào A.
tamarense và A. catenella với cả mẫu nuôi cấy và mẫu tảo thu ngoài tự nhiên [63].
Một số thử nghiệm lai tại chỗ trên Heterosigma akashiwo (Raphidophyceae) với các
đầu dò thiết kế các trình tự ARN và ADN đích đã cho thấy: đối với đầu dò rARN,
kết quả là các ARN trong tế bào chất được lai với đầu dò và làm cho toàn bộ tế bào
bắt màu xanh; còn với đầu dò ADN, đầu dò lai với ADN trong nhân nên chỉ có
nhân tế bào là bắt màu xanh [21].
Khai thác trình tự vùng D1-D2 ADN mã hóa cho tiểu phần lớn của ribosome
(LSU), một số nhà khoa học đã thiết kế các đầu dò và phát triển phép lai huỳnh
quang tại chỗ với tín hiệu FITC nhằm định loại nhanh cho nhiều loài tảo độc hại
trong nhiều thủy vực [22].
Bảng 1.2. Trình tự đầu dò cho một số loài Alexandrium [22].
Tên loài Trình tự đầu dò Số hiệu trên
Genbank
A.tamiyavanichii
A. catenella
A. fraterculus
A. insuetum
A. tamarense
A. pseudogonyaulax
GTACTTAGACCACACCATA
GCAT TTGACAATTGCTCCT
GCTCTTGTTCATCAACATC
GCACTTGATGATCGATTGC
GCATTTGGCGATCAATTCT
GCACTTGATGGTTGTTTGC
AB088316
AB088280
AB088290
AB088298
AB088279
AB088302
Trong nhiều nghiên cứu về sau này, một số cải tiến của kỹ thuật FISH đã được
đưa ra nhằm hoàn thiện và tối ưu hơn cho từng mục đích hay đối tượng quan tâm

17
khác nhau [17]. Với những ưu thế về độ chính xác, độ nhạy và tiết kiệm thời gian,
kỹ thuật lai huỳnh quang tại chỗ vẫn được xem là một công cụ rất hữu hiệu cho
nhiều nghiên cứu sinh học.
1.2. Tình hình nghiên cứu ứng dụng kỹ thuật FISH tại Việt Nam
Cho đến nay, tại Việt Nam, kỹ thuật FISH chủ yếu được nghiên cứu ứng dụng
trong lĩnh vực y học. Một số nghiên cứu xây dựng quy trình chẩn đoán trước sinh
bằng kỹ thuật FISH đã được tiến hành. Các nghiên cứu này được dựa trên các trình
tự ADN đích thuộc các nhiễm sắc thể nhằm phát hiện một cách chính xác các bất
thường trên các nhiễm sắc thể (NST) tương ứng với các bệnh di truyền được quan
tâm [4]. Trong một số nghiên cứu khác, ứng dụng kỹ thuật lai tại chỗ huỳnh quang
được dùng để chẩn đoán trước sinh hội chứng Đao, hội chứng Tớc-nơ cũng đã cho
kết quả thành công. Kết quả nghiên cứu cũng cho thấy, việc sử dụng kỹ thuật lai
huỳnh quang tại chỗ đã cho kết quả rất chính xác khi so sánh với phương pháp phân
tích nhiễm sắc thể của tế bào ối [1, 2]. Bên cạnh đó, các nghiên cứu ứng dụng kỹ
thuật FISH trong việc phát hiện các lệch bội nhiễm sắc thể số 13, 18, 21, X, Y từ tế
bào dịch ối bằng kỹ thuật lai huỳnh quang tại chỗ cũng đã thành công. Trên cơ sở
phân tích các mẫu thu từ các phụ nữ mang thai giai đoạn 17 - 25 tuần có nguy cơ
cao bị dị tật, kỹ thuật FISH đã giúp phát hiện 1 trường hợp mắc hội chứng Patau
(trisomy 13), 1 trường hợp mắc hội chứng Đao (trisomy 21), 1 trường hợp bất
thường cánh dài của NST 16. Kết quả cũng cho thấy 100% mẫu kết quả FISH phù
hợp với kết quả phân tích về NST liên quan đến các NST số 13, 18, 21, X, Y. Quy
trình chuẩn nhằm chẩn đoán trước sinh bằng chọc ối làm xét nghiệm FISH an toàn
cũng đã được hoàn thiện [6]. Sử dụng kỹ thuật FISH nhằm phát hiện sự khuếch đại
gen NMYC trên các bệnh nhân u nguyên bào thần kinh đã được tiến hành tại các
bệnh viện Nhi trung ương và cho kết quả tốt đẹp. Một số biến đổi di truyền trong u
nguyên bào thần kinh có ý nghĩa quan trọng trong việc lựa chọn phác đồ điều trị và
tiên lượng bệnh, trong đó sự khuếch đại gen NMYC được xem là yếu tố quan trọng
nhất đã được tìm thấy. Kết quả thành công này có ý nghĩa quan trọng hỗ trợ các bác

18
sỹ lâm sàng trong việc phân nhóm bệnh nhân và lựa chọn phác đồ điều trị thích hợp
[5].
Các nghiên cứu ứng dụng kỹ thuật lai huỳnh quang tại chỗ FISH đối với các
loài sinh vật thủy sinh tại Việt Nam gần như rất ít. Trong một nghiên cứu gần đây
tại Viện nghiên cứu hải sản về ứng dụng kỹ thuật FISH cho việc nhận diện nhanh 2
loài tảo giáp độc hại A. tamarense và A. catenella, một số thử nghiệm đã được tiến
hành. Các trình tự đích thuộc vùng D1- D2 trên nhiễm sắc thể mã hóa cho tiều phần
lớn LSU của ribosome của loài được lấy làm cơ sở thiết kế đầu dò. Tín hiệu huỳnh
quang FITC được thiết kế gắn với đầu dò đặc hiệu. Kết quả nghiên cứu đã cho thấy
các trùng lặp, giống nhau về đặc điểm hình thái của các loài tảo giáp từng gây khó
khăn trong quan trắc tảo độc hại đã phần nào được giải quyết nhờ kỹ thuật FISH.
Đây cũng là một trong những cơ sở ban đầu cho việc ứng dụng kỹ thuật này trong
quan trắc cảnh báo về sự bùng phát các đối tượng thủy sinh gây thủy triều đỏ tại
Việt Nam [3].
1.3. Các bước cơ bản của quy trình lai huỳnh quang tại chỗ - FISH
Sự ra đời của kỹ thuật FISH dựa trên các cải tiến của kỹ thuật ISH đã ra đời
và là sự kết hợp chính xác của phép lai một đoạn oligonucleotit (đầu dò) có gắn tín
hiệu huỳnh quang với một trình tự ADN đích đặc trưng. Bằng quan sát trực quan từ
kính hiển vi huỳnh quang, các phân tử huỳnh quang phát sáng với những màu sinh
động cho phép hiển thị các đoạn ADN, nhiễm sắc thể hay tế bào đích. Nhờ đó mà
các nhà nghiên cứu có thể nhận dạng, liệt kê và định vị các tế bào sinh vật trong
mẫu môi trường tự nhiên hay trong các mô bệnh phẩm...
Quy trình này bao gồm các bước cơ bản sau:
- Bước 1: Chuẩn bị mẫu và đầu dò có trình tự nucleotit bổ sung đặc hiệu với trình
tự đặc hiệu cần nhận biết.
- Bước 2: Cố định mẫu nghiên cứu.
- Bước 3: Lai giữa đầu dò và trình tự đích đặc hiệu nằm trong tế bào của mẫu.

19
- Bước 4: Rửa mẫu để loại bỏ các đầu dò không được lai (đầu dò tự do).
- Bước 5: Hiển thị, quan sát và phân tích kết quả bằng kính hiển vi huỳnh quang.
Hình 1.2. Các bước cơ bản của kỹ thuật lai huỳnh quang tại chỗ FISH
1.4. Đặc điểm sinh thái của một số loài vi tảo biển độc hại nghiên cứu
Alexandrium affine được xác định là một trong số 31 loài tảo độc hại thuộc
phân chi Alexandrium nom. nud của chi Alexandrium (chi tảo Hai roi) trên thế
giới. Chúng được xác định có phân bố phổ biến ở vùng biển Việt Nam. Các kết quả
nghiên cứu trước đây cho thấy rằng chúng phát triển mạnh vào thời gian từ tháng 12
đến tháng 2 hàng năm và bắt gặp nhiều trong các thủy vực Thừa Thiên Huế. Loài
này cũng đã bắt gặp phát triển mạnh tại các vịnh Văn Phong, Cam Ranh (Khánh
Hòa) vào mùa khô thuộc các tháng 3 và 4. Một kết quả phân tích độc tố của loài này

20
đã cho thấy rằng, thành phần độc tố của chúng bao gồm các loại NeoSTX, STX,
GTX1-4 với nồng độ <2,28µmol/tế bào. Đây là các chất độc được sản sinh ra khi
chúng bùng phát mạnh với mật độ cao trong môi trường thủy sinh. Các chất độc này
làm chết nhiều đối tượng thủy sinh [45]. Trong các nghiên cứu vi tảo biển độc hại,
A. affine là một trong những đối tượng được quan tâm vì chúng cũng là một trong
những yếu tố chỉ thị môi trường quan trọng [7].
Một loài tảo khác là Alexandrium pseudogonyaulax đã được xác định là một
trong 15 loài tảo độc hại thuộc chi Alexandrium (phân chi Gessnerrium) có ở biển
Việt Nam. A. pseudogonyaulax được coi là loài tảo gây hại trong nhiều thủy vực
nước mặn và lợ ven biển Việt Nam. Các kết quả điều tra nghiên cứu trước đây đã
cho thấy rằng: A. pseudogonyaulax có phân bố rộng và rất dễ bùng phát và đã từng
bùng phát nhiều đợt, làm thiệt hại nghiêm trọng tới nhiều thủy vực nuôi trồng thủy
sản tại các vùng ven biển miền trung và Nha Trang [7]. Tuy không phải là nhóm vi
tảo hai roi sản sinh độc tố, nhưng do khả năng dễ tạo mật độ cao trong các thủy vực
nên A. pseudogonyaulax được đánh giá là một trong những loài tiềm tàng gây hại
cao, là nguyên nhân gây thiệt hại cho nhiều đầm nuôi thủy sản trong thời gian qua.
Chính vì vậy đây cũng là một trong những đối tượng chỉ thị môi trường quan trọng
trong nhiều thủy vực ven biển [45].
Trong nghiên cứu này, chúng tôi vì vậy đã tiến hành thử nghiệm ứng dụng
kỹ thuật lai huỳnh quang FISH nhằm nhận diện nhanh được 2 loài tảo độc hại là
Alexandrium affine và A. pseudogonyaulax thu thập ở Việt Nam.

21
CHƯƠNG 2. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Địa điểm thu mẫu và đối tượng nghiên cứu
Hai mẫu tảo độc hại được thu tại vùng biển Cát Bà – Hải Phòng, gồm có:
+ Mẫu A. sp6 (Alexandrium affine) được thu vào ngày 20/7/2010 tại khu vực
cửa biển thuộc Bến Bèo (Cát Bà, Hải Phòng) với điều kiện môi trường như sau: độ
mặn nước biển: 27‰, pH 7,72.
+ Mẫu A. sp7 (Alexandrium pseudogonyaulax) được thu vào ngày 27/4/2011
tại khu vực vịnh cảng Cát Bà (Hải Phòng) với các điều kiện môi trường nước biển
như sau: độ mặn nước biển: 29‰, pH 7,44.
2.2. Phương pháp thu và lưu giữ mẫu
Mẫu vi tảo nghiên cứu được thu theo hướng dẫn của Hallegraeff và đồng
nghiệp [32] và Anderson [15].
Mẫu được thu bằng lưới thu thực vật phù du có đường kính 20cm, kích thước
mắt lưới 20µm, thu theo phương thẳng đứng. Mẫu sau đó được bảo quản trong lọ và
giữ trong điều kiện nhiệt độ từ 4 – 15oC. Mẫu này sau đó được chuyển về phòng thí
nghiệm để phân lập.
Đối với các mẫu vi tảo thu ngoài tự nhiên để thử nghiệm đầu dò, 2 phương
pháp bảo quản, xử lý mẫu được tiến hành:
- Xử lý mẫu thu được từ lưới kéo như trên và sau đó mẫu này được sử
dụng cho các thí nghiệm lai như trình bày ở phần sau.
- Các mẫu thu từ ngoài tự nhiên không kịp tiến hành thí nghiệm lai với đầu
dò ngay trong ngày thì được cố định bằng etanol 70% hoặc formandehyt
10%. Các mẫu này sau đó được đưa vào thí nghiệm lai như mô tả ở phần
sau (Shoko và cộng sự [60]).

22
2.2.1 Phân lập các chủng vi tảo (theo phương pháp thu tế bào đơn lẻ bằng
micropipet của Richmon [57] và Andersen [13]:
Mẫu tảo sau khi chuyển về phòng thí nghiệm, được phân lập bằng pipet mao
quản (pipet thủy tinh kéo dài). Sau đó, tế bào được rửa sạch bằng cách hút chuyển
từng tế bào cần phân lập vào môi trường nuôi tảo vài lần có cùng độ mặn, cho tới
khi mẫu sạch hoàn toàn. Tiếp theo, mỗi tế bào được chuyển sang một giếng nuôi đã
chứa sẵn môi trường đã được pha sẵn với độ mặn tương ứng ngoài tự nhiên.
Môi trường nuôi được sử dụng là môi trường IMK (Daigo IMK – mua từ
hãng Nihon Pharmaceutical Co., Ltd.) dành cho nhóm tảo giáp (Alexandrium).
2.2.2 Phương pháp nuôi giữ các chủng vi tảo thu được (theo Shoko và cộng sự
[60], Nguyễn Văn Nguyên và cộng sự [3]):
Các mẫu tảo sau khi thu thập được nuôi giữ như sau:
+ Bước 1 - Nuôi trong giếng: Mẫu sau khi phân lập được rửa sạch, mỗi tế
bào (hoặc chuỗi tế bào) được chuyển vào một giếng. Giếng nuôi có đường kính
16mm, thể tích khoảng 5ml (loại 24 giếng/khay của hãng Corning - Mỹ). Sau 3 đến
5 ngày nuôi và theo dõi, nếu tảo đã thuần và phát triển tốt, tiến hành cấy truyền sang
lọ nuôi bằng nhựa (flask). Đối với các mẫu chưa thuần, phân lập được lặp lại cho tới
khi thuần hoàn toàn.
+ Bước 2 - Nhân nuôi sinh khối trong lọ nhựa hoặc bình tam giác: Mẫu tảo
phát triển tốt được tiếp tục nuôi tạo sinh khối trong lọ nhựa hoặc bình tam giác
50ml làm nguyên liệu cho các thí nghiệm tiếp theo.
Mẫu tảo độc được nuôi trong các điều kiện nhân tạo được kiểm soát về các
thông số môi trường, bao gồm:
Cường độ chiếu sáng: 2.500 lux, bằng hệ thống ánh sáng trắng (đèn neon);
Chế độ chiếu sáng: 13h sáng liên tục/11h tối liên tục, được điều khiển bởi
đồng hồ đóng ngắt tự động;
Nhiệt độ nuôi: ổn định ở 24oC liên tục trong buồng có điều hòa nhiệt độ;
Môi trường nuôi: IMK (hãng Nihon Pharmaceutical Co., Ltd., Nhật Bản).

23
2.3. Phương pháp phân loại hình thái
Việc phân loại các mẫu tảo giáp được thực hiện chủ yếu dựa vào công thức
tấm vỏ tế bào đã được nhuộm Calco - fluor, quan sát dưới kính hiển vi quang học và
có sự hỗ trợ của chuyên gia phân loại hình thái. Các tài liệu chính để phân loại là:
Balech (1995), Abe (1967a,b), Larsen và cộng sự (2004) [8, 9, 16, 45].
2.4. Phương pháp kiểm tra phân loại bằng ADN và thiết kế đầu dò
2.4.1. Phương pháp chuẩn bị ADN khuôn (Sebastián và cộng sự (2001) [62]):
Chuẩn bị ADN khuôn: Một tế bào được phân lập từ mẫu nuôi bằng pipet
pasteur thủy tinh kéo dài. Tế bào được rửa 3 lần bằng nước biển lọc có độ mặn giảm
dần từ 27‰ đến 20‰ và 15‰. Cuối cùng, tế bào được rửa 3 lần bằng nước cất
trước khi đưa vào trong ống PCR chứa sẵn 10µl nước khử ion đã được tiệt trùng. Tế
bào được giữ ở -20oC và sau đó được đưa vào sốc nhiệt để phá vỡ tế bào, chuẩn bị
cho phản ứng PCR.
Phương pháp sốc nhiệt : Các ống mẫu chứa tế bào được chuyển đột ngột qua
lại 3 lần giữa hai trạng thái lạnh (-20oC ) và nóng (90oC) trong thời gian mỗi 20 giây
ở mỗi điều kiện, nhằm làm vỡ tế bào và dung giải ADN ra ngoài. Các mẫu được lắc
rung (votex) và ly tâm trở lại (5000vòng/phút) chuẩn bị cho phản ứng PCR.
2.4.2. Phương pháp Singel cell PCR (Sebastián và cộng sự (2001) [62], Shoko và cộng sự (2005) [60])
Phản ứng PCR được tiến hành để nhân vùng D1 – D2 của gen mã hóa tiểu
đơn vị lớn 28s ARN ribosome. Tuy nhiên, do lượng ADN khuôn ban đầu ít (từ 1 tế
bào) nên quy trình PCR được thực hiện thông qua 2 phản ứng theo sơ đồ hình 2.1).
Sản phẩm phản ứng PCR 2 (vùng ADN mục tiêu)
Pr28-
PrS1R
PrD2
PrD1
SSU ITS1 5S ITS2
D1 D2 D3 D4 D5 D6 D7 D8 ……
SSU ITS1 5S ITS2 D1 D2 D3 D4 D5
ADN mã hóa Ribosom
Cặp mồi đặc hiệu cho phản ứng PCR1
Sản phẩm phản ứng PCR1
PCR1
PCR2
D1 D2
Cặp mồi đặc hiệu cho phản ứng PCR2
Hình 2.1. Sơ đồ phản ứng PCR nhân vùng D1-D2 của rADN

24
Phản ứng PCR lần 1: Nhân đoạn ADN dài khoảng 5.000 bp, mã hóa cho rARN của
Alexandrium từ vùng tiểu phần nhỏ SSU tới vùng D6 của tiểu phần lớn LSU. Cặp
mồi sử dụng cho phản ứng PCR lần 1: S1R (5’-TACCTGGTTGATCCTGCCAG-
3’) và 28-1483R (5’-GCTACTACCACCAAGATCTGC-3’). Chu trình nhiệt: 950C
5phút, (950C 1phút, 550C 1phút, 720C 5phút 30 giây) x 35 chu kỳ, 720C 10 phút .
Thể tích phản ứng PCR là 25µl có chứa các thành phần như sau:
- MgCl2: 20mM (1x đệm Takara Ex Taq)
- dNTPs: 200µM (Takara Ex)
- Taq: 1U (Takara Ex Taq)
- Mồi: 1µM mỗi mồi (TOS-Nhật Bản)
- ADN khuôn: 5µl
Phản ứng PCR lần 2: Nhân đoạn D1-D2 của tiểu phần LSU, với độ dài
khoảng 700bp. Thể tích phản ứng PCR lần 2 là 25µl có thành phần tương tự như
trên, sử dụng ADN khuôn là 5µl sản phẩm PCR lần 1. Cặp mồi sử dụng là: D1R
(5’-ACCCGCTGAATTTAAGCATA-3’) và D2C (5’-
CCTTGGTCCGTTTCAAGA-3’). Chu trình nhiệt: 95oC 5phút, (95oC 30 giây, 55oC
30 giây, 72oC 1phút 50 giây) x 35 chu kỳ, 72oC 10 phút.
Các phản ứng PCR được thực hiện trên máy PCR Thermal Perkin Elmer 9600.
Kiểm tra sản phẩm PCR: Sản phẩm PCR lần 2 được điện di trên gel agarose
1,5%, nhuộm trong ethidium bromide, soi trên đèn UV có bước sóng 365nm để
kiểm tra và đánh giá kết quả.
2.4.3. Phương pháp kiểm tra phân loại bằng ADN
Giải trình tự ADN vùng D1-D2: Sản phẩm thu được của PCR lần 2 được tinh
sạch bằng cột tinh sạch của hãng Promega và gửi đi giải trình tự trên máy Applied
Biosystems 3130 Series (tại phòng thí nghiệm của Viện vi sinh vật và Công nghệ
sinh học - Đại học Quốc gia Hà Nội).

25
Kiểm tra phân loại bằng ADN: Dựa trên trình tự vùng D1 - D2 thu được,
chúng tôi xây dựng cây phát sinh chủng loại theo phương pháp kết nối lân cận
(neighbor joining) sử dụng thuật toán Jukes-Cantor với độ lặp lại 1000 lần, sử dụng
trình tự số hiệu AF260387 trên ngân hàng gen (genbank) làm loài ngoài nhóm để so
sánh. Cây phát sinh chủng loại được xây dựng trên phần mềm Mega phiên bản 5.03.
2.4.4. Phương pháp thiết kế đầu dò đặc hiệu
Dựa trên trình tự ADN thu được, sử dụng công cụ ClustalW, phần mềm
BioEdit để sắp xếp, so sánh các trình tự nucleotit thu được với trình tự của những
loài gần gũi về mặt phân loại. Trên cơ sở đó, lựa chọn đoạn ADN đặc trưng nhất
cho loài để thiết kế đầu dò. Đầu dò được thiết kế và được đặt hàng tại công ty TOS
(Nhật Bản) để chế tạo, với yêu cầu gắn thêm chất phát huỳnh quang fluorescein
isothiocyanate (FITC) ở đầu 5’ để phục vụ cho việc hiển thị sự hiện diện của trình
tự đặc trưng.
Trong nghiên cứu này, kỹ thuật lai huỳnh quang tại chỗ giữ nguyên vẹn tế
bào được áp dụng. Các đầu dò huỳnh quang gắn tín hiệu FITC được thiết kế để gắn
đặc hiệu với trình tự đích trên tiểu phần lớn của ribosome của tế bào vi tảo. Với số
lượng lớn các ribosome trong mỗi tế bào, kết quả lai tốt sẽ cho lượng lớn các đầu dò
có gắn tín hiệu huỳnh quang được gắn lên ribosome trong mỗi tế bào của loài. Dưới
sự tác động của chùm sáng 490nm, tín hiệu FITC được kích thích và phát sáng
(xanh lam) đặc trưng sẽ làm toàn bộ tế bào phát sáng xanh lam. Do đó tế bào dễ
dàng được phát hiện bằng kính hiển vi huỳnh quang.
2.5. Phương pháp lai huỳnh quang tại chỗ (dựa theo phương pháp của Shoko
và cộng sự (2005) [60])
Mẫu (5000 - 7000 tế bào/ml) được đưa vào ly tâm thu tế bào (3000vòng/phút
trong 1 phút ở 4oC).
Cố định tế bào bằng 1ml dung dịch PBS (5x) có chứa paraformandehyde
(4%), để mẫu trên đá lạnh trong 5 phút.
Ly tâm mẫu 3000vòng/phút trong 1 phút (4oC).
Loại nước trong mẫu theo 2 bước:

26
- Thêm 1ml dung dịch ethanol 50% vào mẫu, ủ trên đá trong 5 phút. Ly tâm
thu tế bào (3.000vòng/phút trong 1 phút, 4oC).
- Thêm 1ml dung dịch ethanol 80% vào mẫu, ủ trên đá trong 5 phút. Ly tâm
thu tế bào (3.000vòng/phút trong 1 phút, 4oC).
Thêm 500µl dung dịch lai (gồm: Formamide (40%), SSC (5x), mẫu dò có
gắn huỳnh quang (50 pmol)).
Lai tế bào ở các thang nhiệt độ khác nhau: 37°C, 40°C, 43°C, 46°C, 49°C,
52°C trong 5 phút.
Sau khi lai, mẫu được rửa 2 lần:
- Thêm 1ml SSC (5x) vào mẫu, ủ mẫu ở 50oC trong 5 phút. Ly tâm thu tế bào
(3000vòng/phút trong 1 phút, 4oC).
- Thêm 1ml SSC (5x) vào mẫu, ủ mẫu ở 50oC trong 5 phút. Ly tâm thu tế bào
(3000vòng/phút trong 1 phút, 4oC).
Sản phẩm lai huỳnh quang sau đó được quan sát trên kính hiển vi huỳnh quang
(ECLIPSE E800, Nikon, Tokyo, Japan) với kính lọc sắc B-2A, bước sóng kích thích
tín hiệu FITC 490 nm, hiển thị và chụp ảnh tế bào bằng camera DS Nikon.
Chi tiết thành phần các dung dịch được thể hiện trong phụ lục 1.

27
CHƯƠNG 3. KẾT QUẢ NGHIÊN CỨU VÀ THẢO LUẬN
3.1. Kết quả phân lập và phân loại các chủng vi tảo độc hại bằng hình thái
Trên cơ sở các mẫu thu được từ vùng biển Cát Bà, 2 chủng vi tảo độc hại
đã được phân lập, nuôi giữ và phân loại sơ bộ dựa trên các đặc điểm về hình thái.
Hai loài tảo giáp được xác định sơ bộ là thuộc chi Alexandrium bao gồm:
3.1.1. Loài Alexandrium affine
Một số đặc điểm hình thái của tế bào được ghi nhận qua công thức tấm vỏ
của loài đặc trưng như sau: Kích thước tế bào khoảng 38 - 45µm, có dạng bất đối
xứng. Tấm 1' liên kết trực tiếp với tổ hợp lỗ đỉnh và có một lỗ bụng nhỏ ở vị trí
trung tâm trên mép phải tấm 1' nơi tiếp xúc với tấm 4'. Tấm S.a có dạng hình
thang và không có phần phụ đi kèm. Một số hình ảnh loài được ghi nhận qua
hình 3.1.
Hình 3.1. Một số đặc điểm hình thái của A. affine.
Hình dạng ngoài của tế bào (A); Tấm 1' với lỗ bụng và tổ hợp lỗ đỉnh (B, C);
Tấm S.a hình thang, không có phần phụ (D).
28
So sánh các đặc điểm này cho thấy có sự trùng hợp với các đặc điểm phân
loại được mô tả đặc trưng cho loài này trong các nghiên cứu trước đây. Tuy
nhiên, đây cũng là những đặc điểm khó nhận biết và dễ nhầm lẫn trong phân tích
công thức tấm vỏ của loài. Do đó việc kết luận loài là A. affine chỉ được khẳng
định lại một cách chắc chắn từ kết quả phân loại bằng ADN được trình bày ở
phần sau của luận văn.
3.1.2. Loài A. pseudogonyaulax
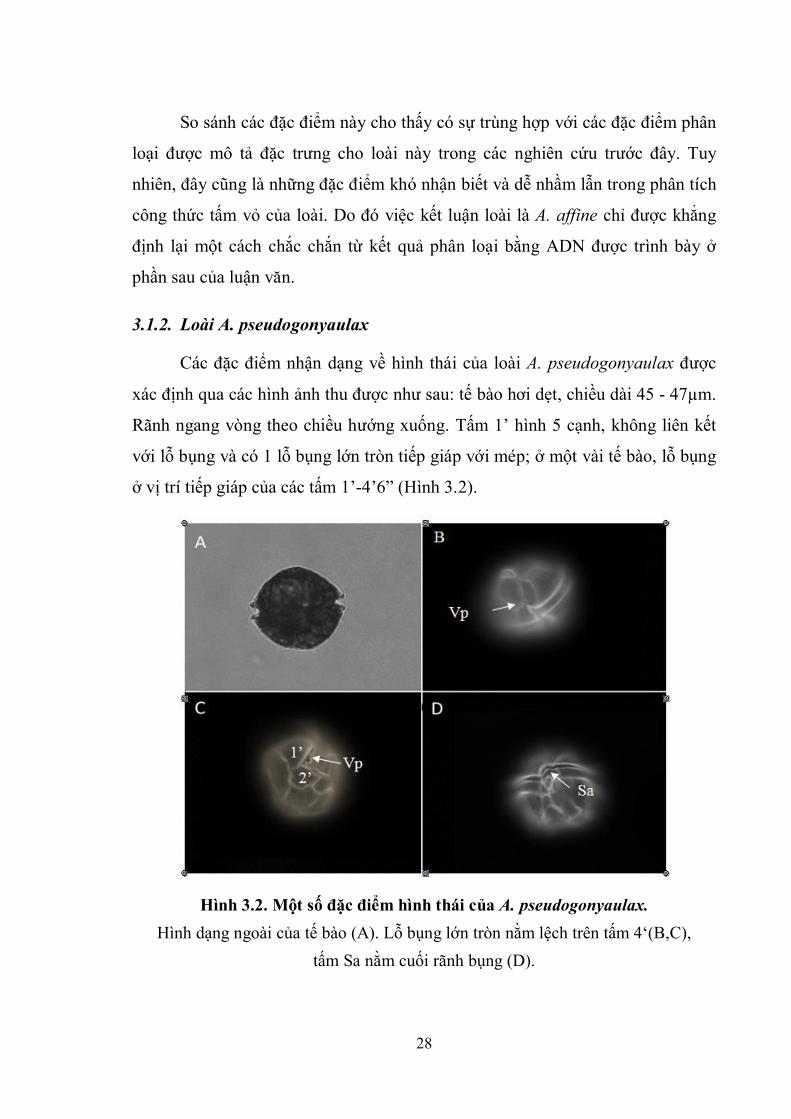
Các đặc điểm nhận dạng về hình thái của loài A. pseudogonyaulax được
xác định qua các hình ảnh thu được như sau: tế bào hơi dẹt, chiều dài 45 - 47µm.
Rãnh ngang vòng theo chiều hướng xuống. Tấm 1’ hình 5 cạnh, không liên kết
với lỗ bụng và có 1 lỗ bụng lớn tròn tiếp giáp với mép; ở một vài tế bào, lỗ bụng
ở vị trí tiếp giáp của các tấm 1’-4’6” (Hình 3.2).
Hình 3.2. Một số đặc điểm hình thái của A. pseudogonyaulax.
Hình dạng ngoài của tế bào (A). Lỗ bụng lớn tròn nằm lệch trên tấm 4‘(B,C),
tấm Sa nằm cuối rãnh bụng (D).

29
Cũng giống như đối với loài A. affine, kết quả phân loại dựa trên hình thái
đối với loài A. pseudogonyaulax không thật sự rõ ràng. Mặc dù các đặc điểm thu
được có nhiều tương đồng so với mô tả của Larsen [45]. Do vậy, việc xác định
chính xác tên loài được kết hợp với những kết quả so sánh trình tự ADN vùng
D1 - D2 của mẫu thu được với các trình tự mẫu trên Genbank sẽ được mô tả ở
phần sau của luận văn.
3.2. Ảnh hưởng của độ mặn tới nuôi sinh khối vi tảo tạo nguyên liệu
Do trong điều kiện thí nghiệm này đã sử dụng môi trường nuôi dưỡng
IMK dành riêng cho tảo giáp (hãng Nihon Pharmaceutical Co., Ltd., Nhật Bản)
và đã tham khảo một số điều kiện chiếu sáng tối ưu trong các nghiên cứu trước
đây về tảo giáp [7], nên tác giả chỉ tập trung vào tìm hiểu ảnh hưởng của độ mặn
lên sự sinh trưởng của tảo trong phòng thí nghiệm, nhằm thu được sinh khối tốt
nhất cho các thí nghiệm tiếp theo.
Độ mặn được xem là một trong những yếu tố quyết định cho việc sinh
trưởng của các loài vi tảo biển. Hai mẫu vi tảo biển độc hại thu được được đưa
vào nhân nuôi trong phòng thí nghiệm với các điều kiện đã nêu ở trên. Kết quả
thí nghiệm nuôi cấy trong các điều kiện độ mặn khác nhau cho thấy, ảnh hưởng
của độ mặn lên sự sinh trưởng của từng loài là khác nhau (Hình 3.3 và 3.4).
Trong điều kiện nuôi có độ mặn 30‰ (cường độ chiếu sáng 2500lux, chế độ
chiếu sáng 13 giờ sáng: 11 giờ tối) loài A. affine phát triển tốt nhất trong các chế
độ nuôi cấy. Kết quả này cũng thống nhất với kết quả nghiên cứu trước đây là độ
mặn thích hợp nhất cho sự phát triển của loài này là 20 – 35‰ với chủng tảo thu
được ở vùng biển miền trung Việt Nam [7, 45].
Một kết quả nghiên cứu khác của Larsen (2004) đối với loài này với
chủng thu được trên vùng biển Autralia cho thấy chúng có ngưỡng chịu độ mặn
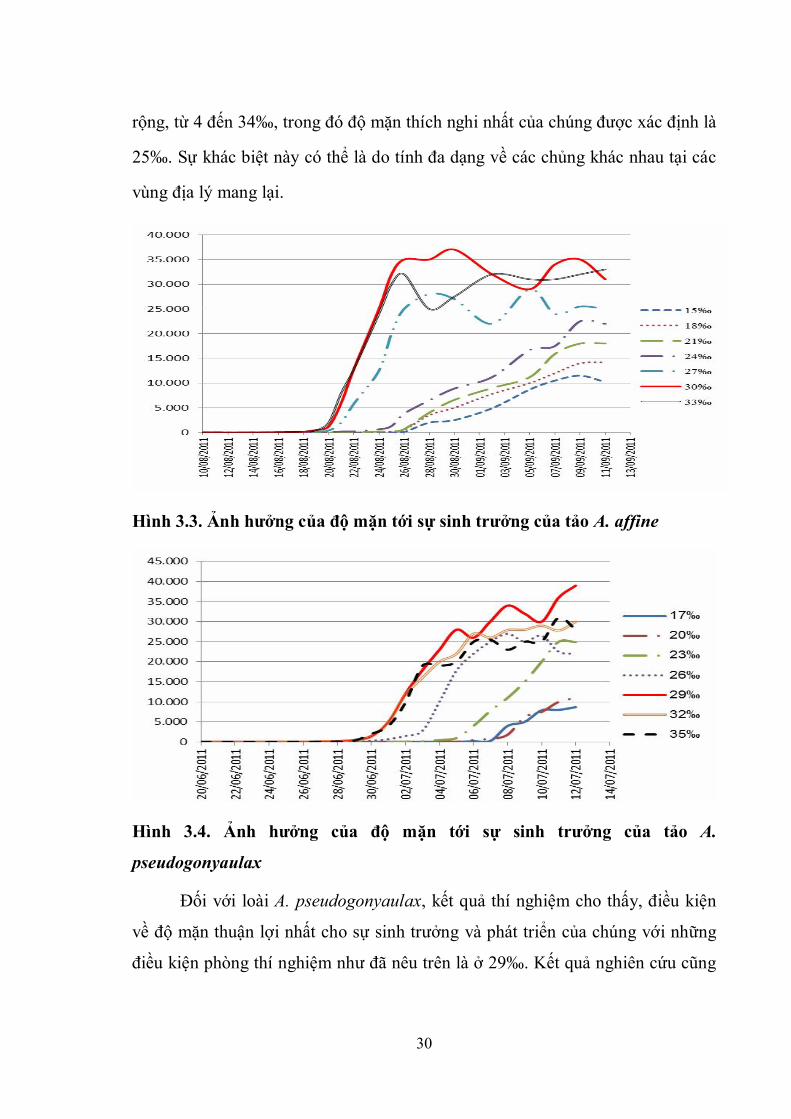
30
rộng, từ 4 đến 34‰, trong đó độ mặn thích nghi nhất của chúng được xác định là
25‰. Sự khác biệt này có thể là do tính đa dạng về các chủng khác nhau tại các
vùng địa lý mang lại.
Hình 3.3. Ảnh hưởng của độ mặn tới sự sinh trưởng của tảo A. affine
Hình 3.4. Ảnh hưởng của độ mặn tới sự sinh trưởng của tảo A.
pseudogonyaulax
Đối với loài A. pseudogonyaulax, kết quả thí nghiệm cho thấy, điều kiện
về độ mặn thuận lợi nhất cho sự sinh trưởng và phát triển của chúng với những
điều kiện phòng thí nghiệm như đã nêu trên là ở 29‰. Kết quả nghiên cứu cũng

31
chỉ ra rằng, với những điều kiện độ mặn từ 23 - 35‰ là khoảng thích hợp cho
nuôi sinh khối loài này trong phòng thí nghiệm. Một kết quả nghiên cứu trước
đây của Chu Văn Thuộc (2006) cũng cho kết quả tương tự [7].
Như vậy, một số điều kiện tốt nhất cho nuôi nhân tạo 2 loài tảo trên trong
điều kiện phòng thí nghiệm được chúng tôi rút ra như được nêu trên bảng 3.1.
Bảng 3.1. Một số điều kiện thích hợp cho nhân nuôi sinh khối tảo A. affine
và A. pseudogonyaulax trong phòng thí nghiệm
Loài Các yếu tố
A. affine A. pseudogonyaulax
Môi trường IMK (g/L) 2,52 2,52
Độ mặn (‰) 30 29
Chế độ chiếu sáng (Sáng/Tối) 13/11 13/11
Cường độ chiếu sáng (lux) 2500 2500
Các kết quả nghiên cứu trên đã cho thấy được điều kiện thích hợp nhất
cho việc nuôi lưu giữ cũng như nhân sinh khối hai loài này một cách tốt nhất để
làm nguyên liệu cho các nghiên cứu tiếp theo.
3.3. Kết quả giải trình tự và phân loại bằng ADN các mẫu nghiên cứu
Trên cơ sở ADN khuôn thu được từ sản phẩm PCR lần 1 (nhân đoạn ADN
mã hóa cho rARN của tảo Alexandrium, từ vùng tiểu phần nhỏ SSU tới vùng D6
của tiểu phần lớn LSU), sản phẩm PCR lần 2 nhằm nhân đoạn ADN mã hóa
vùng D1 - D2 của ribosome đã có kết quả. Ở mỗi mẫu thí nghiệm, đoạn mồi
D1R, D2C đã nhân được một đoạn ADN duy nhất có kích thước tương ứng với
đoạn dự kiến, khoảng gần 700bp, trong khi sản phẩm PCR của mẫu đối chứng
âm không cho băng nào. Điều này chứng tỏ rằng phản ứng PCR rất đặc hiệu. Kết
quả được thể hiện ở sản phẩm điện di trên gel agarose 1,5% (hình 3.5).

32
Hình 3.5. Kết quả điện di sản phẩm PCR lần 2
(M1: Marker5 (ФX174/HincII), M2: Wide Range DNA Ladder 50-10.000bp. L1-3: A.affine, L4-6: A.pseudogonyaulax; L7: Đối chứng âm)
Sản phẩm PCR lần 2 (L3 và L4) của 2 loài nói trên đã được tiến hành giải
trình tự. Kết quả cho thấy, vùng D1-D2 của mẫu L3 có kích thước gồm 651bp
và có trình tự nucletotit như sau:
CCTTGGTCCGTGTTTCAAGACGGGTCAAGCAGAAGCATTTTGCCAGC
AATTGTGTGCCAGGGCTTGTAAGCTCTAGTAGGGTAGGCCCAGCTTC
AAGAAATGAGCAATTACAAAAGCACCAGACACATGTATCAAAACAC
AAATCATCACAAACACATGCAATCCAAACCCCGCAGGAAAATTACCA
TTCATGTGCAAGGGTAATCAAATGTACAAATAGAAACTGGCAAGCAA
AGCAAGAATCATCACACCCACAATCAAAACCTATTGTCAAGCAAGCA
CCACAATCTCACCAAGCAACATCAAATCTGTTTATTTTCTGTTCAGCA
AATACAAACTCATTTAATTCTCTTTTCAAAGTCCTTTTCATATTTCCCT
CATGGTACTTGTGTGCTATCAGTCTCAATCCTATATTTACCTTTACAT
GAAACTTACCACCCACTTTGCATTCCAATGCCAAGGAGTGTGACTCA
GCAAACATACACCGTGCACAGAGGATTGCACATGACAAACAGGATTC
TCACCCTCATTGACGCTTTCTTTCAATAGGCTGACACCTGCACCTCTG
M2
1000bp 750bp
500bp
M1
1057bp 770bp 612bp

33
TTGGTAATACATTTCAAAAATACAAGTCGAAGCCAGAGGCTCCAATT
GTCAAGCTAAGCATGTCCTTGTTCATTCGCATAC
Đối với mẫu L4, vùng D1-D2 có trình tự ADN thu được bao gồm 650 bp
và có trật tự các nucleotit như sau:
CCCGCTGAATTTAAGCATATAAGTAAGCGGTGGAAAATGAACCAAAT
GGGATTCCTTTAGTAATTGCGAATGAACAAGGATGTGCTCAGCTTGA
CAAATGGAGTTTTTGAGCTTTGAATTGTAATTTTGCAATGCATCACCA
GCGGAGGTACAGTTGCAAGCCTTTTGAAAGACAGCGCCATTGAGGGT
GAAATTCCTTTTTGTCTTCTGCAACCTTTGTGCACGGTATGTATTTGGT
GAATCACATTCCTCGGCATTGGAATGCAAATTGGGTGGTAAATTTCA
CGCAAGGGAAAATATATTATTGAGTCTGATAATGAACAAGTACCATG
AGGGAAATATGCAAAGGACTTTGAAAAGAAAATTAAAAGAGCTTGC
ATTTGCTAAACAGAAAACTAGCAGAAGTGCTTTATCTTGGTAAGATT
GCTGCGCATGGGTTTACTGTGTAAATTGTATGAGCATTGGTTCTTACT
TTGTGTGTCAATTCCATTGTCAATTTTAATATCTTTGCTCATGAATTGT
GAAATGCTTGCGGGTGTTAGATTGCATGTGCATTCATTAATTTGGACT
TGGTGCAGTGCTTGGCAACAGCTTGCATGTAGAGTGTGCGTGCAATG
TACAATTGCGATTGTCAGCTGTTGCCAACCACTTTTGTTGGCAATGTG
ATTCTGCTTGACCCGTCTTGAAACACGGACCAAGG
34
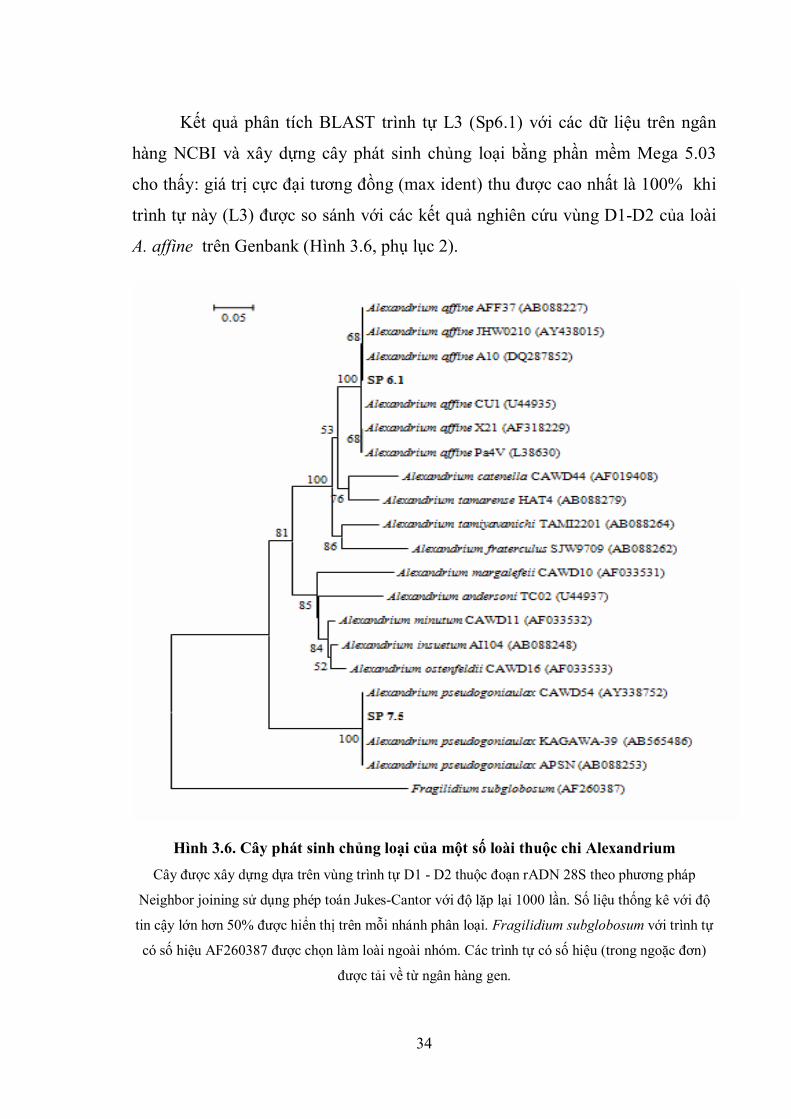
Kết quả phân tích BLAST trình tự L3 (Sp6.1) với các dữ liệu trên ngân
hàng NCBI và xây dựng cây phát sinh chủng loại bằng phần mềm Mega 5.03
cho thấy: giá trị cực đại tương đồng (max ident) thu được cao nhất là 100% khi
trình tự này (L3) được so sánh với các kết quả nghiên cứu vùng D1-D2 của loài
A. affine trên Genbank (Hình 3.6, phụ lục 2).
Hình 3.6. Cây phát sinh chủng loại của một số loài thuộc chi Alexandrium
Cây được xây dựng dựa trên vùng trình tự D1 - D2 thuộc đoạn rADN 28S theo phương pháp
Neighbor joining sử dụng phép toán Jukes-Cantor với độ lặp lại 1000 lần. Số liệu thống kê với độ
tin cậy lớn hơn 50% được hiển thị trên mỗi nhánh phân loại. Fragilidium subglobosum với trình tự
có số hiệu AF260387 được chọn làm loài ngoài nhóm. Các trình tự có số hiệu (trong ngoặc đơn)
được tải về từ ngân hàng gen.

35
Chủng SP 6.1 có trình tự vùng D1-D2 tương đồng với các chủng thuộc
loài Alexandrium affine thu thập tại các vùng bờ Biển Nhật Bản (AFF37), Hàn
Quốc (JHW0210), Hong Kong (A10), Thái Lan (CU1), Pháp (X21) và Tây Ban
Nha (Pa4V). Như vậy có thể thấy trình tự này rất tương đồng và trùng với các
kết quả nghiên cứu trước đây. Điều này cũng là cơ sở khẳng định đây chính là
loài A. affine và những kết quả phân loại loài qua hình thái đã nói ở trên là phù
hợp.
Đối với trình tự L4 (Sp7.5), các kết quả so sánh độ tương đồng và xây
dựng cây phát sinh chủng loại cũng đã chỉ ra rằng, L3 có trình tự phù hợp với
các trình tự đoạn D1 – D2 của loài A. pseudogonyaulax có trên nguồn dữ liệu
của Genbank (NCBI). Chủng SP 7.5 có trình tự vùng D1 - D2 tương đồng với
các chủng thuộc loài Alexandrium pseudogoniaulax thu thập tại các vùng bờ
Biển Newzeland (CAWD54) và Nhật Bản (KAGAWA-39, APSN). Điều này
giúp khẳng định loài Sp7.5 thu được là A. pseudogonyaulax (Hình 3.6, phụ lục
3).
3.4. Thiết kế đầu dò lai huỳnh quang tại chỗ
Trên cơ sở 2 trình tự thu được từ vùng D1 - D2 của hai mẫu tảo độc hại
trên, phần mềm Bioedit được sử dụng để sắp xếp, so sánh với các trình tự tham
khảo (từ NBCI) tại vùng tương đồng D1 - D2 của các loài thuộc chi
Alexandrium. Những khác biệt từ các trình tự bảo thủ này so với các loài khác là
cơ sở cho việc chọn lọc trình tự phù hợp để thiết kế đầu dò đặc hiệu cho từng
loài. Kết quả sắp xếp và so sánh đối với loài A. affine được thể hiện trong hình
3.7 và phụ lục 4. Theo đó, có 2 đoạn polynucleotit có trình tự khác biệt nhiều với
các trình tự tương ứng của các loài khác thuộc chi Alexandrium là trình tự
nucleotit từ vị trí 630 đến 649 và 657 đến 676.
Dựa trên các yêu cầu về thiết kế đầu dò ADN có kích thước khoảng 20
nucleotit, trình tự có tỷ lệ GC cao được lựa chọn làm khuôn để thiết kế đầu dò.

36
Trong nghiên cứu này, đoạn trình tự nucleotit từ 657 đến 676 được lựa chọn làm
mẫu xây dựng trình tự đầu dò. Trình tự đầu dò cho loài A. affine được xác định
là: 5’-TGTAAGCTCTAGTAGGGTAG-3’ có trình tự bổ sung với trình tự đoạn
ADN trên. Đoạn trình tự đầu dò này đã được kiểm tra lại bằng BLAST trên
NBCI và không thấy có trình tự nào bổ sung tương đồng. Kết quả này khẳng
định rằng trình tự này phù hợp cho việc thiết kế đầu dò đặc hiệu cho loài A.
affine.
Hình 3.7. So sánh sự khác biệt trình tự nucleotit của A. affine
với các loài Alexandrium
Tương tự như vậy, các kết quả so sánh đối với trình tự đoạn D1 – D2 của
loài A. pseudogonyaulax (hình 3.8 và phụ lục 5) cũng cho thấy: sự khác biệt
nhiều trong trình tự các nucleotit so với các loài khác trong cùng chi xuất hiện
trong đoạn trình tự 774 đến 794. Đây có thể được coi là một đoạn bảo thủ, đặc
trưng của loài. Dựa trên kết quả này, trình tự đầu dò được xác định là: 5’-
ACAGCTGACAATCGCAATTG-3’. Kết quả kiểm tra lại bằng công cụ BLAST
cho thấy đoạn trình tự này phù hợp cho đầu dò cần thiết kế do chúng không tạo
liên kết bổ sung với đoạn ADN nào khác trong NCBI. Đồng thời, trình tự đầu dò

37
này khác với các trình tự đầu dò đã được nghiên cứu đối với loài này của Chong
và các đồng nghiệp [22].
Hình 3.8. So sánh sự khác biệt trình tự nucleotit của A. pseudogonyaulax
với các loài Alexandrium
Các trình tự này sau đó được đặt sản xuất bởi công ty TOS (Nhật Bản) với
điều kiện có gắn tín hiệu Fluorescein 5-isothiocyanate (FITC) vào đầu 5’ để
phục vụ cho các thí nghiệm lai huỳnh quang tiếp theo.
3.5. Tối ưu nhiệt độ lai và thử nghiệm đầu dò
Trên cơ sở đầu dò thiết kế được, các thí nghiệm lai huỳnh quang tại chỗ
kiểm tra cho 2 loài A. affine và A. pseudogonyaulax đã được tiến hành. Tuy
nhiên do có sự khác biệt về đối tượng cũng như trình tự đầu dò nên yếu tố nhiệt
độ lai cho từng đối tượng và đầu dò tương ứng đã được thử nghiệm.
Trên cơ sở các nghiên cứu trước đây, nhiệt độ lai trong kỹ thuật FISH
thông thường chỉ từ 40 đến 50°C [18]. Mặt khác, việc bổ sung formamide nồng
độ cao trong dung dịch đệm lai cũng sẽ giúp hạ nhiệt độ lai xuống thấp [18], nên
các thử nghiệm về nhiệt độ lai trong nghiên cứu này được tiến hành trong dải
nhiệt độ gồm 37°C, 40°C, 43°C, 46°C, 49°C và 52°C.

38
Kết quả thử nghiệm cho thấy, có sự giống nhau về kết quả lai đối với cả 2
đầu dò trên hai đối tượng tảo khác nhau. Đối với loài A. affine, nhiệt độ lai phù
hợp là 40°C và 43°C. Trong đó, tín hiệu huỳnh quang thu được rõ nét nhất ở tại
43°C (Hình 3.9). Độ nhạy của tín hiệu huỳnh quang tại nhiệt độ lai này lớn hơn
rất nhiều so với tại nhiệt độ là 40°C. Trong khi đó, các mẫu có nhiệt độ lai khác
đều cho tín hiệu huỳnh quang rất kém hoặc không có tín hiệu (phụ lục 6), mẫu
đối chứng âm có sử dụng đầu dò A. affine cho việc nhận biết các loài gần gũi là
A. tamarense và A. catenella đều không có kết quả (không có tín hiệu). Như vậy,
chúng tôi kết luận nhiệt độ lai thích hợp nhất đối với đầu dò được thiết kế cho
loài A. affine là khoảng 43°C.
Hình 3.9. Tế bào A. affine dưới ánh sáng thường (trái) và phát huỳnh
quang dưới ánh sáng kích thích (phải) với nhiệt độ lai 43°C
Hình 3.10. Tế bào A. pseudogonyaulax dưới ánh sáng thường (trái) và
phát huỳnh quang dưới ánh sáng kích thích (phải) với nhiệt độ lai 43°C

39
Tương tự đối với loài A. pseudogonyaulax, kết quả thử nghiệm cũng đã
cho thấy, đối với mẫu được lai ở nhiệt độ 43°C cho kết quả tốt nhất, hình ảnh sắc
nét. Trong khi các mẫu còn lại, thử nghiệm ở các nhiệt độ 37, 40, 46, 49 và 52°C
đều không thu được tín hiệu huỳnh quang khi hiển thị trên kính hiển vi. Điều này
cho thấy với nhiệt độ không phù hợp, các phân tử đầu dò rất khó hoặc không lai
được với trình tự đích (hình 3.10). Từ các hình ảnh thu được cũng cho thấy, với
bước rửa cuối, việc loại bỏ những đầu dò dư thừa hay những trình tự gắn kết
không hoàn toàn là thích hợp để thu được chỉ những đầu dò liên kết vững chắc
với các trình tự đích và từ đó có thể có được những hình ảnh huỳnh quang tốt
nhất. Như vậy nhiệt độ lai phù hợp trong thí nghiệm này thấp hơn nhiệt độ nóng
chảy của đầu dò theo tính toán thông thường. Kết quả này cũng trùng hợp với
các nghiên cứu trước đây đã chỉ ra rằng, việc bổ sung formamide vào trong dung
dịch lai sẽ góp phần làm hạ thấp nhiệt độ lai trong lai tại chỗ FISH [18].
Như vậy với đầu dò được thiết kế, phép lai huỳnh quang tại chỗ đã được
tiến hành thành công với hai loài vi tảo A.affine và A.pseudogonyaulax nuôi
trong phòng thí nghiệm.
40
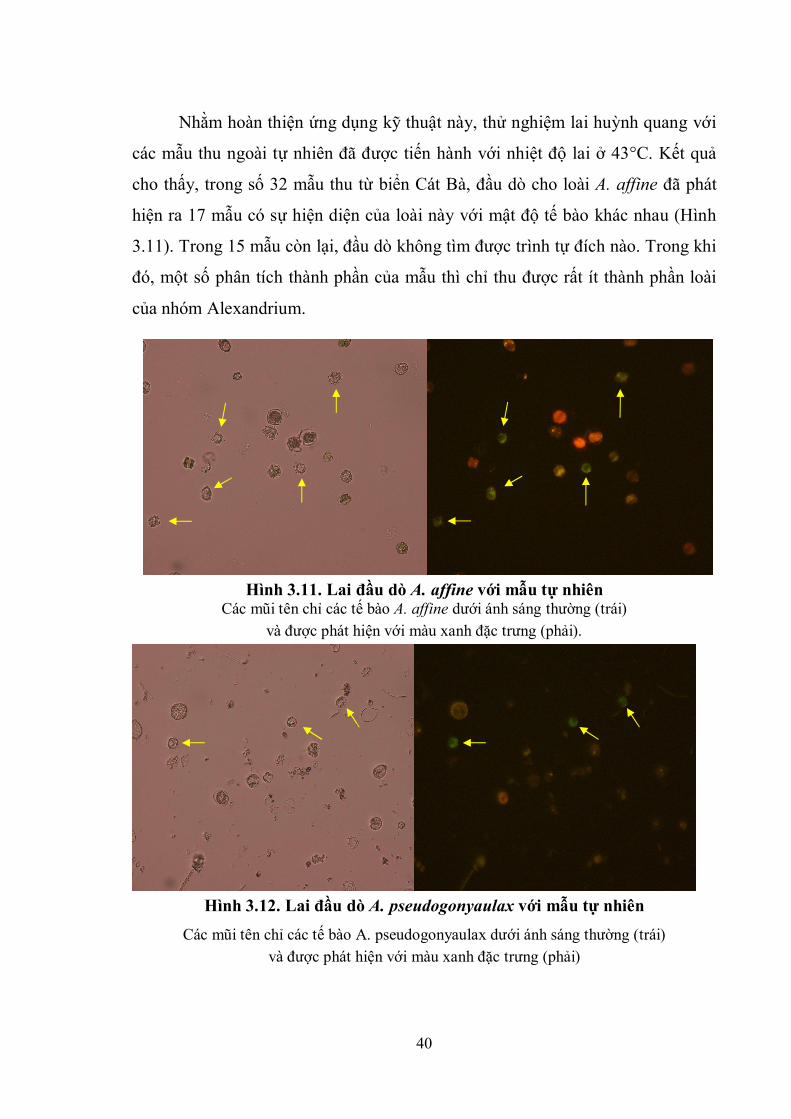
Nhằm hoàn thiện ứng dụng kỹ thuật này, thử nghiệm lai huỳnh quang với
các mẫu thu ngoài tự nhiên đã được tiến hành với nhiệt độ lai ở 43°C. Kết quả
cho thấy, trong số 32 mẫu thu từ biển Cát Bà, đầu dò cho loài A. affine đã phát
hiện ra 17 mẫu có sự hiện diện của loài này với mật độ tế bào khác nhau (Hình
3.11). Trong 15 mẫu còn lại, đầu dò không tìm được trình tự đích nào. Trong khi
đó, một số phân tích thành phần của mẫu thì chỉ thu được rất ít thành phần loài
của nhóm Alexandrium.
Hình 3.11. Lai đầu dò A. affine với mẫu tự nhiên Các mũi tên chỉ các tế bào A. affine dưới ánh sáng thường (trái)
và được phát hiện với màu xanh đặc trưng (phải).
Hình 3.12. Lai đầu dò A. pseudogonyaulax với mẫu tự nhiên
Các mũi tên chỉ các tế bào A. pseudogonyaulax dưới ánh sáng thường (trái)
và được phát hiện với màu xanh đặc trưng (phải)

41
Tương tự, với đầu dò A. pseudogonyalax đã phát hiện 9 mẫu có trình tự đích
cần tìm (hình 3.12). Trong các mẫu còn lại, đầu dò cũng không tìm thấy trình tự
đích nào.
Từ các kết quả thử nghiệm cho thấy, đầu dò lai huỳnh quang của cả hai loài
đều hoạt động tốt. Kỹ thuật lai huỳnh quang có thể được dùng để xác định nhanh
hai loài tảo độc hại A. affine và A. pseudogonyaulax.
Quy trình lai huỳnh quang tại chỗ (FISH) sử dụng đầu dò rARN có gắn tín
hiệu FITC đối với hai loài A. affine và A. pseudogonyaulax được tổng hợp lại và
đề nghị như sau:
Mẫu (5000-7000 tế bào/ml) được đưa vào ly tâm thu tế bào
(3000vòng/phút trong 1 phút, 4oC).
Cố định tế bào bằng 1ml dung dịch PBS (5x) có chứa paraformandehyde
(4%), để mẫu trên đá lạnh trong 5 phút.
Ly tâm mẫu 3000vòng/phút trong 1 phút (4oC).
Loại nước trong mẫu theo 2 bước:
- Thêm 1ml dung dịch ethanol 50% vào mẫu, ủ trên đá trong 5 phút. Ly tâm
thu tế bào (3000vòng/phút trong 1 phút, 4oC).
- Thêm 1ml dung dịch ethanol 80% vào mẫu, ủ trên đá trong 5 phút. Ly
tâm thu tế bào (3000vòng/phút trong 1 phút, 4oC).
Thêm 500µl dung dịch lai (gồm: Formamide (40%), SSC (5x), mẫu dò có
gắn huỳnh quang (50 pmol)).
Lai tế bào ở 43°C trong 5 phút.
Sau khi lai, mẫu được rửa 2 lần:
- Thêm 1ml SSC (5x) vào mẫu, ủ mẫu ở 50oC trong 5 phút. Ly tâm thu tế
bào (3000vòng/phút trong 1 phút, 4oC).
- Thêm 1ml SSC (5x) vào mẫu, ủ mẫu ở 50oC trong 5 phút. Ly tâm thu tế
bào (3000vòng/phút trong 1 phút, 4oC).
Chi tiết thành phần các dung dịch được thể hiện trong phụ lục 1.

42
Sản phẩm lai huỳnh quang sau đó được quan sát trên kính hiển vi huỳnh
quang (ECLIPSE E800, Nikon, Tokyo, Japan) với kính lọc sắc B-2A, bước sóng
kích thích tín hiệu FITC 490 nm, hiển thị và chụp ảnh tế bào bằng camera DS
Nikon. Tổng thời gian thao tác cho quá trình này đã được tính toán là khoảng 2,5
giờ. Điều này cho phép các thí nghiệm nghiên cứu phát hiện các loài vi tảo biển
độc hại chính xác, nhanh hơn nhiều so với các phương pháp phân tích truyền
thống bằng hình thái trước đây.

43
KẾT LUẬN
Chúng tôi đã rút ra được một số kết luận chính sau từ nghiên cứu này:
1. Đã xác định trình tự gen vùng mã hóa cho đoạn D1 – D2 của tiểu phần lớn
(LSU) thuộc ribosome của hai loài tảo giáp độc hại A. affine và A.
pseudogonyaulax biển Việt Nam.
2. Đã thiết kế và thử nghiệm thành công hai đầu dò phân tử gắn tín hiệu
huỳnh quang FITC phục vụ cho ứng dụng kỹ thuật lai huỳnh quang tại
chỗ đối với hai loài tảo độc A. affine và A. pseudogonyaulax biển Việt
Nam. Các đầu dò hoạt động tốt trên cả mẫu nuôi cấy và mẫu thu ngoài tự
nhiên và các đầu dò đảm bảo tính đặc hiệu của loài.
3. Đã ứng dụng thành công kỹ thuật FISH trong việc xác định nhanh chóng
và chính xác 2 loài tảo độc hại biển Việt Nam nói trên, làm cơ sở cho việc
ứng dụng phương pháp quan trắc, đánh giá nhanh môi trường biển, nghiên
cứu dự đoán diễn biến thủy triều đỏ tại các vùng ven biển Việt Nam sau
này.

44
KIẾN NGHỊ
Cần nghiên cứu và thiết kế thêm các đầu dò nhằm ứng dụng kỹ thuật lai
huỳnh quang để nhận diện nhanh, nhiều loài tảo biển độc hại khác trong các
vùng biển và ven biển Việt Nam.
Cần có các nghiên cứu, ứng dụng các kỹ thuật di truyền trong việc quan
trắc, phân loại nhanh đối với các loài vi tảo biển Việt Nam nhằm có thể theo dõi
thường xuyên, đánh giá nhanh những biến đổi môi trường gây thủy triều đỏ do
tảo biển độc hại gây ra, hạn chế thiệt hại trong nuôi trồng thủy sản ven biển.

45
TÀI LIỆU THAM KHẢO
Tiếng Việt
1. Trần Thị Thu Hương, Hoàng Thu Lan, Trần Thị Ngọc Lan, Trịnh Đức
Cường (2005), “Ứng dụng kỹ thuật lai tại chỗ huỳnh quang để chẩn đoán
trước sinh hội chứng down”, Tạp chí y học thực hành, 11, tr. 9-11.
2. Trần Thị Thu Hương, Hoàng Thu Lan, Trần Thị Ngọc Lan, Trịnh Đức
Cường, Nguyễn Thị Quỳnh Thơ (2006), “Chẩn đoán trước sinh hội chứng
Down, hội chứng Turner bằng kỹ thuật lai tại chỗ huỳnh quang kết hợp phân
tích nhiễm sắc thể của tế bào ối”, Tạp chí nghiên cứu y học, 1, tr. 4-8.
3. Nguyễn Văn Nguyên, Lê Thanh Tùng, Lưu Xuân Hòa, Vũ Tuấn Nam, Vũ
Minh Hào, Đinh Thái Bình (2011), “Thử nghiệm kỹ thuật lai huỳnh quang tại
chỗ F.I.S.H nhận diện nhanh và chuẩn xác một số loài tảo giáp độc hại”,
Tuyển tập báo cáo Hội nghị khoa học và công nghệ biển toàn quốc lần thứ V,
Quyển 4 - Sinh học và nguồn lợi biển, tr. 261-268.
4. Đặng Thị Hồng Nhung, Nguyễn Duy Ngọc (2008), “Áp dụng kỹ thuật huỳnh
quang tại chỗ trong phát hiện bất thường cấu trúc nhiễm sắc thể tại Bệnh viện
Nhi Trung ương”, Tạp chí nghiên cứu y học, 57, tr. 57-62.
5. Vũ Đình Quang, Ngô Diễm Ngọc, Nguyễn Thị Phương Mai, Đinh Thị Hồng
Nhung, Phùng Tuyết Lan, Trần Đức Hậu, Trần Ngọc Sơn, Nguyễn Thanh
Liêm (2010), “Đánh giá sự khuếch đại gen NMYC trên bệnh nhân
Neuroblastoma”, Tạp chí Nhi khoa, 3 (3), tr. 358-363.
6. Nguyễn Thị Tân Sinh, Ngô Diễm Ngọc, An Thuỳ Lan, Đinh Thị Hồng
Nhung, Lê Thị Liễu, Nguyễn Thanh Liêm (2011), “Ứng dụng kỹ thuật lai
huỳnh quang tại chỗ trong chẩn đoán trước sinh các lệch bội nhiễm sắc thể
thường gặp”, Tạp chí Y học lâm sàng, số đặc biệt, tháng 2, tr. 253-258.

46
7. Chu Văn Thuộc (2002), “Nghiên cứu thành phần loài, phân bố và thăm dò
khả năng gây hại của một số loài tảo độc hại (harmful algae) thuộc ngành tảo
giáp (Dinophyta) ở vùng cửa sông, ven biển miền Bắc Việt Nam”, Luận án
tiến sĩ ngành sinh học, Viện nghiên cứu hải sản.
Tiếng Anh
8. Abé, T. H. (1967a), “The armoured Dinoflagellata: II. Prorocentridae and
Dinophysidae (A)”, Publ. Seto Mar. Biol., 14(5), pp. 369 - 389.
9. Abé, T. H. (1967b), “The armoured Dinoflagellata: II. Prorocentridae and
Dinophysidae (B), Dinophysis and its allied genera”, Publ. Seto Mar. Biol.,
15(1), pp. 37 - 78.
10. Adachi M., Sako Y., Ishida Y. (1996), “Analysis of Alexandrium
(Dinophyceae) species using sequences of the 5,8S ribosomal DNA and
internal transcribed spacer regions”. J. Phycol, 32, pp. 424–432.
11. Alfreider A., Pernthaler J., Amann R., Sattler B., Glöckner F. O., Wille A.,
and Psenner R. (1996), “Community analysis of the bacterial assemblages in
the winter cover and pelagic layers of a high mountain lake by in situ
hybridization”, Appl. Environ. Microbiol, 62, pp: 2138 - 2144.
12. Amann R., Fuchs B. M., Beherens S. (2001), “The identification of
microorganisms by fluorescence in situ hybridization”, Curr. Opin.
Biotechnol., 12, pp. 231 – 236.
13. Andersen, G. (2005), “Traditional Microalgae Isolation Techniques, Algal
cuturing techniques”, Phycologycal society of America, 578p.
14. Anderson, P. (1996), “Design and implementation of some harmful agal
monitoring system”, IOC Technical Series, 44, UNESCO.

47
15. Anderson, D.M. (1995), “Identification of harmful algal species using
molecular probes, an emerging technology”, Harmful Marine Algal Blooms,
Lavoiser Science Publishers, pp. 3–13.
16. Balech, I. (1995), “The genus Alexandrium Halim (Dinoflagellata)”, Trans.
Am. Microscop. Soc., 113, pp. 216 - 220.
17. Baoyu Z., Guofu C., Guangce W., Douding L. (2010), “Identification of two
Skeletonema costatum-like diatoms by fluorescence in situ hybridization”.
Chinese Journal of Oceanology and Limnology, 28(2), pp. 310 - 314.
18. Benedetta B., Danilo E., Monica G., Erasmo N. (2006), “Application of
FISH technology for microbiological analysis: current state and prospects”,
Appl. Microbiol. Biotechnol., 73, pp. 485 – 494.
19. Bouvier T., Del G. (2003), “Factors influencing the detection of bacterial
cells using fluorescence in situ hybridization (FISH)”, FEMS Microbiol.
Ecol., 44, pp. 3 – 15.
20. Cho E. S., Hur H. J., Byun H. S., Lee S. G., Rhodes L. L., Jeong C. S. and
Park J. G. (2002), “Monthly monitoring of domoic acid producer Pseudo-
nitzschia multiseries (Hasle) Hasle using species-specific DNA probes and
WGA lectins and abundance of Pseudo-nitzschia species (Bacillariophyceae)
from Chinhae Bay”, Korea. Bot. Mar., 45, pp. 364 - 372.
21. Choi, B. K. (1994), “Diversity of cultivable and uncultivable oral spirochetes
from a patient with severe destructive periodontitis”. Infect. Immun., 62, pp.
1889 – 1895.
22. Choong J. K., Yoshihiko S. (2005), “Molecular identification of toxic
Alexandrium tamiyavanichii (Dinophyceae) using two DNA probes”,
Harmful Algae, 4, pp. 984 - 991.

48
23. Deere, D. (1998), “Evaluation of fluorochromes for flow cytometric
detection of Cryptosporidium parvum oocysts labelled by fluorescence in
situ hybridization”, Lett. Appl. Microbiol., 27, pp. 352 – 356.
24. DeLong, E. F. (1989), “Phylogenetic stains: ribosomal RNA-based probes
for the identification of single microbial cells”, Science, 243, pp. 1360 –
1363.
25. DeLong E. F., Taylor L. T., Marsh T. L. and Preston C. M. (1999),
“Visualization and enumeration of mMarine planktonic archaea and bacteria
by using polyribonucleotide probes and fluorescent in situ hybridization”,
Applied and Environmental Microbiology, 65, pp. 5554 - 5563.
26. Felske, A. (1998), “In situ detection of an uncultured predominant Bacillus
in dutch grassland soils”, Appl. Environ. Microbiol., 64, pp. 4588 – 4590.
27. Fuchs B. M., Syutsubo K., Ludwig W., Amann R. (2000), “In situ
accessibility of Escherichia coli 23S rRNA to fluorescently labeled
oligonucleotide probes”, Appl. Environ. Microbiol., 67, pp. 961 – 968.
28. Gersdorf H. (1993a), “Identification of bacteroides forsythus in subgingival
plaque from patients with advanced periodontitis”, J. Clin. Microbiol., 31,
pp. 941 – 946.
29. Gersdorf, H. (1993b), “Fluorescence in situ hybridization for direct
visualization of Gram-negative an aerobes in subgingival plaque samples”,
FEMS Immunol. Med. Microbiol., 6, pp. 109 – 114.
30. Glockner, F. O. (1996), “An in situ hybridization protocol for detection and
identification of planctonic bacteria”, Syst. Appl. Microbiol., 19, pp. 403 –
406.
31. Glöckner F. O., Fuchs B. M. and Amann R. (1999), “Bacterioplankton
compositions of lakes and oceans: a first comparison based on fluorescence
in situ hybridization”, Appl. Environ. Microbiol., 65, pp. 3721 - 3726.

49
32. Hallegraeff G. M., Anderson D. M., Cembella A. D., Enevoldsen H. O.
(1995), “Manual on harmful marine microalgae, IOC Manual and Guides”,
UNESCO, 33, 794p.
33. Hogardt M., (2000), “Specific and rapid detection by fluorescent in situ
hybridization of bacteria in clinical samples obtained from cystic fibrosis
patients”, J. Clin. Microbiol., 38, pp. 818 – 825.
34. Hogardt M., (2000), “Specific and rapid detection by fluorescent in situ
hybridization of bacteria in clinical samples obtained from cystic fibrosis
patients”, J. Clin. Microbiol. 38, pp. 818 – 825.
35. Hougaard D. M., Hansen H. and Larsson L. I. (1997), “Non-radioactive in
situ hybridization for mRNA with emphasis on the use of
oligodeoxynucleotide probes”, Histochem. Cell Biol., 108, pp. 335 - 344.
36. Inacio J., Beherens S., Fuchs B. M., Fonseca A., Spencer M. I., Amann R.
(2003), “In situ accessibility of Saccharomyces cerevisiae 26S rRNA to Cy3-
labeled oligonucleotide probes comprising the D1 and D2 domains”, Appl.
Environ. Microbiol., 69, pp. 2899 – 2905.
37. Jansen, G. J. (2000), “Rapid identification of bacteria in blood cultures by
using fluorescently labeled oligonucleotide probes”, J. Clin. Microbiol., 38,
pp. 814 – 817.
38. Jeffrey M. L. and Robert H. S. (2003), “Fluorescence in situ hybridization:
past, present and future”, Journal of Cell Science, 116, pp. 2833 – 2838.
39. Johannes Z., Wolfgang L., Karl H. S. (2001), “DNA polynucleotide probes
generated from representatives of the genus acinetobacter and their
application in fluorescence in situ hybridization of environmental samples”,
System. Appl. Microbiol., 24, pp. 238 – 244.
40. Kagiyama, N. (1993), “A novel fluorescent method for in situ hybridization”,
Acta Histochem. Cytochem., 26, pp. 441 – 445.

50
41. Kempf, V. A. (2000), “Fluorescent in situ hybridization allows rapid
identification of microorganisms in blood cultures”, J. Clin. Microbiol., 38,
pp. 830 – 838.
42. Kenzaka, T. (1998), “rRNA targeted fluorescent in situ hybridization
analysis of bacterial community structure in river water”, Microbiology, 144,
pp. 2085 – 2093.
43. Kim C. J., Kim C. H., Sako Y. (2005), “Development of molecular
identification method for genus Alexandrium (Dinophyceae) using whole-
cell FISH”, Marine Biotechnology, 7, pp. 215 – 222.
44. Langendijk P. S, Schut F., Jansen G. J., Raangs G. C., Kamphuis G. R.,
Wilkinson M. H. F., Welling G. W. (1995), “Quantitative fluorescence in situ
hybridization of Bifidobacterium spp. with genus-specific 16S rRNA-
targeted probes and its application in faecal samples”, Appl. Environ.
Microbiol., 61, pp. 3069 – 3075.
45. Larsen J. and Nguyen N. L. (2004), “Potentially toxic microalgae of
Vietnamese waters”, Opera Botanica, 140, 216p.
46. Llobet B., Rosselló M., and Amann R. (1998), “Microbial community
composition of wadden sea sediments as revealed by fluorescence in situ
hybridization”, Appl. Environ. Microbiol., 64, pp. 2691 - 2696.
47. Manuelidis L., Langer P. R. and Ward D. C. (1982), “High-resolution
mapping of satellite DNA using biotin-labeled DNA probes”, J. Cell Biol.,
95, pp. 619 - 625.
48. Michael H., Ralph W., Georg H., Jutta F., Andrea W., Alexander R.,
Eberhard S., Siegfried J., Christoph C. (2003), “COMBO-FISH: specific
labeling of nondenatured chromatin targets by computer-selected DNA
oligonucleotide probe combinations”, BioTechniques, 35(3), pp. 564 – 577.

51
49. Miller P. E., Scholin C. A. (2000), “On detection of Pseudo-nitzschia
(Bacillariophyceae) species using whole cell hybridization: sample fixation
and stability”, J. Phycol., 36, pp. 238 – 250.
50. Miller P. E., Scholin C. A. (1996), “Identification of cultured
Pseudonitzschia (Bacillariophyceae) using species-specific LSU
rRNAtargeted fluorescent probes”, J. Phycol., 32, pp. 646 – 655.
51. Miller P. E., Scholin C. A. (1998), “Identification and enumeration of
cultured and wild Pseudo-nitzschia (Bacillariophyceae) using species-
specific LSU rRNA-targeted fluorescent probes and filter-based whole cell
hybridization”, J. Phycol., 34, pp. 371 – 382.
52. Moter, A. (1998), “Molecular epidemiology of oral treponemes associated
with periodontal disease”, J. Clin. Microbiol., 36, pp. 1399 – 1403.
53. Niels B. R., Henrik F., Timothy G. F., Finn A., Bo T. (1996), “Distribution
of bacterial populations in a stratified fjord (Mariager Fjord, Denmark)
quantified by in situ hybridization and related to chemical gradients in the
water column”, Appl. Environ. Microbiol., 62, pp. 1391 – 1404.
54. Pardue, M. L. (1969), “Molecular hybridization of radioactive DNA to the
DNA of cytological preparations”, Proc. Natl. Acad. Sci., 64, pp. 600 –604.
55. Pernthaler J., Glöckner F. O., Schönhuber W., and Amann. R. (2001),
“Fluorescence in situ hybridization (FISH) with rRNA-targeted
oligonucleotide probes”, Methods in Microbiology: Marine Microbiology,
30, pp. 207 – 210.
56. Puvion D., and Puvion E., (1996), “Non-isotopic electron microscope in situ
hybridization for studying the functional sub-compartmentalization of the
cell nucleus”, Histochem. Cell Biol., 106, pp. 59 - 78.
57. Richmon, A. (2004), “Handbook of Microalgal culture: Biotechnology and
Applied Phycology”, Blackwell Science, 577p.

52
58. Rudkin G. T., and Stollar B. D. (1977), “High resolution detection of DNA-
RNA hybrids in situ by indirect immunofluorescence”, Nature, 265, pp. 472-
473.
59. Scholin C., Miller P. E., Buck K., Chavez F., Harris P., Haydock P., Howard
J. (1997), “Detection and quantification of Pseudo-nitzschia australis in
cultured and natural populations using LSU rRNA-targeted probes”, Limnol.
Oceanogr. 42(5), pp. 1265 –1272.
60. Shoko H. T., Sako Y. (2005), “Rapid detection of natural cells of
Alexandrium tamarense and A. catenella (Dinophyceae) by fluorescence in
situ hybridization”, Harmful Algae, 4, pp. 319 – 328.
61. Singer R. H., and Ward D. C. (1982), “Actin gene expression visualized in
chicken muscle tissue culture by using in situ hybridization with a biotinated
nucleotide analog”, Proc. Natl. Acad. Sci., 79, pp. 7331 - 7335.
62. Sebatián C. R., and Ryan C. O’. (2001), “Single-cell sequencing of
dinoflagellate (Dinophyceae) nuclear ribosomal genes”, Molecular Ecology
Resources, 1(4), pp. 329-331.
63. Tanabe S. H., Sako Y. (2006), “Development and application of fluorescence
in situ hybridization (FISH) method for simple and rapid identification of the
toxic dinoflagellates Alexandrium tamarense and Alexandrium catenella in
cultured and natural seawater”, Fishersies science, 72(1), pp. 77 - 82.
64. Trebesius K., Amann R., Ludwig W., Mühlegger K., and Schleifer K.
(1994),“ Identification of whole fixed bacterial cells with nonradioactive 23S
rRNA-targeted polynucleotide probes”, Appl. Environ. Microbiol. 60, pp.
3228 - 3235.
65. Wagner M., Schmid S. J., Trebesius K., Bubert A., Goebel W., and Schleifer
K. (1998), “In situ detection of a virulence factor mRNA and 16s rRNA in
Listeria monocytogenes”, FEMS Microbiology Letters, 160, pp. 159 - 168.

53
66. Werner M., Rudolf A., Wolfgang L., Marc V., and Karl H. S. (1996),
“Application of a suite of 16s rRNA-specific oligonucleotide probes
designed to investigate bacteria of the phylum cytophaga-flavobacter-
bacteroides in the natural environment”, Microbiology, 142, pp. 1097 – 1106.
67. Yamaguchi, N. (1996). “Detection of specific bacterial cells with 2-hydroxy-
3-naphthoic acid-29-phenylanilide phosphate and Fast Red TR in situ
hybridization”, Appl. Environ. Microbiol., 62, pp. 275–278.
68. Ylmaz L. S., Hatice E. O., Noguera D. R. (2005), “Making all parts of the
16S rRNA of Escherichia coli accessible in situ to single DNA
oligonucleotides”, Appl. Environ. Microbiol., 72, pp. 733 – 744.
69. Zarda, B. (1991), “Identification of single bacterial cells using
digoxigeninlabelled, rRNA-targeted oligonucleotides”, J. Gen. Microbiol.,
137, pp. 2823 – 2830.

54
PHỤ LỤC
Phụ lục 1. Thành phần các dung dịch sử dụng cho phép lai huỳnh quang tại chỗ
Dung dịch Thành phần
PBS NaCl (684mM)
KCl (14mM)
Na2H2PO4.12H2O (405mM)
KH2PO4 (7,5mM)
pH 7,5
Đệm lai SSC 5X NaCl (83mM)
Sodium Citrate (83mM)
pH 7,0

55
Phụ lục 2. Kết quả so sánh trình tự đoạn L3 nhân vùng D1 – D2 của mẫu A. affine thu được từ vùng biển Cát Bà với chủng AFF37 của loài Alexandrium affine vùng biển Nhật Bản (Số hiệu Genbank: AB088227):
gb|AB088227| Alexandrium affine strain AFF37 large subunit ribosomal RNA gene, partial sequence Length=711 Score = 1201 bits (650), Expect = 0.0 Identities = 650/650 (100%), Gaps = 0/650 (0%) Strand=Plus/Plus Query 1 AAGCATATAAGTAAGTGGTGGAAATTAAACCAAATGGGATTCCTTAAGTAATTGCGAATG 60 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 14 AAGCATATAAGTAAGTGGTGGAAATTAAACCAAATGGGATTCCTTAAGTAATTGCGAATG 73 Query 61 AACAAGGACATGCTTAGCTTGACAATTGGAGCCTCTGGCTTCGACTTGTATTTTTGAAAT 120 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 74 AACAAGGACATGCTTAGCTTGACAATTGGAGCCTCTGGCTTCGACTTGTATTTTTGAAAT 133 Query 121 GTATTACCAACAGAGGTGCAGGTGTCAGCCTATTGAAAGAAAGCGTCAATGAGGGTGAGA 180 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 134 GTATTACCAACAGAGGTGCAGGTGTCAGCCTATTGAAAGAAAGCGTCAATGAGGGTGAGA 193 Query 181 ATCCTGTTTGTCATGTGCAATCCTCTGTGCACGGTGTATGTTTGCTGAGTCACACTCCTT 240 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 194 ATCCTGTTTGTCATGTGCAATCCTCTGTGCACGGTGTATGTTTGCTGAGTCACACTCCTT 253 Query 241 GGCATTGGAATGCAAAGTGGGTGGTAAGTTTCATGTAAAGGTAAATATAGGATTGAGACT 300 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 254 GGCATTGGAATGCAAAGTGGGTGGTAAGTTTCATGTAAAGGTAAATATAGGATTGAGACT 313 Query 301 GATAGCACACAAGTACCATGAGGGAAATATGAAAAGGACTTTGAAAAGAGAATTAAATGA 360 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 314 GATAGCACACAAGTACCATGAGGGAAATATGAAAAGGACTTTGAAAAGAGAATTAAATGA 373 Query 361 GTTTGTATTTGCTGAACAGAAAATAAACAGATTTGATGTTGCTTGGTGAGATTGTGGTGC 420 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 374 GTTTGTATTTGCTGAACAGAAAATAAACAGATTTGATGTTGCTTGGTGAGATTGTGGTGC 433 Query 421 TTGCTTGACAATAGGTTTTGATTGTGGGTGTGATGATTCTTGCTTTGCTTGCCAGTTTCT 480 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 434 TTGCTTGACAATAGGTTTTGATTGTGGGTGTGATGATTCTTGCTTTGCTTGCCAGTTTCT 493 Query 481 ATTTGTACATTTGATTACCCTTGCACATGAATGGTAATTTTCCTGCGGGGTTTGGATTGC 540 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 494 ATTTGTACATTTGATTACCCTTGCACATGAATGGTAATTTTCCTGCGGGGTTTGGATTGC 553 Query 541 ATGTGTTTGTGATGATTTGTGTTTTGATACATGTGTCTGGTGCTTTTGTAATTGCTCATT 600 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 554 ATGTGTTTGTGATGATTTGTGTTTTGATACATGTGTCTGGTGCTTTTGTAATTGCTCATT 613 Query 601 TCTTGAAGCTGGGCCTACCCTACTAGAGCTTACAAGCCCTGGCACACAAT 650 |||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 614 TCTTGAAGCTGGGCCTACCCTACTAGAGCTTACAAGCCCTGGCACACAAT 663

56
Phụ lục 3. Kết quả so sánh trình tự đoạn L4 nhân vùng D1 – D2 của mẫu A. pseudogonyaulax thu được từ vùng biển Cát Bà với chủng AFF37 của loài Alexandrium affine vùng biển Nhật Bản (Số hiệu Genbank: AB565486):
dbj|AB565486.1| Alexandrium pseudogoniaulax gene for large subunit rRNA, partial sequence Length=701 Score = 1293 bits (651), Expect = 0.0 Identities = 651/651 (100%), Gaps = 0/651 (0%) Strand=Plus/Plus Query 1 CCCGCTGAATTTAAGCATATAAGTAAGCGGTGGAAAATGAACCAAATGGGATTCCTTTAG 60 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 2 CCCGCTGAATTTAAGCATATAAGTAAGCGGTGGAAAATGAACCAAATGGGATTCCTTTAG 61 Query 61 TAATTGCGAATGAACAAGGATGTGCTCAGCTTGACAAATGGAGTTTTTGAGCTTTGAATT 120 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 62 TAATTGCGAATGAACAAGGATGTGCTCAGCTTGACAAATGGAGTTTTTGAGCTTTGAATT 121 Query 121 GTAATTTTGCAATGCATCACCAGCGGAGGTACAGTTGCAAGCCTTTTGAAAGACAGCGCC 180 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 122 GTAATTTTGCAATGCATCACCAGCGGAGGTACAGTTGCAAGCCTTTTGAAAGACAGCGCC 181 Query 181 ATTGAGGGTGAAATTCCTTTTTGTCTTCTGCAACCTTTGTGCACGGTATGTATTTGGTGA 240 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 182 ATTGAGGGTGAAATTCCTTTTTGTCTTCTGCAACCTTTGTGCACGGTATGTATTTGGTGA 241 Query 241 ATCACATTCCTCGGCATTGGAATGCAAATTGGGTGGTAAATTTCACGCAAGGGAAAATAT 300 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 242 ATCACATTCCTCGGCATTGGAATGCAAATTGGGTGGTAAATTTCACGCAAGGGAAAATAT 301 Query 301 ATTATTGAGTCTGATAATGAACAAGTACCATGAGGGAAATATGCAAAGGACTTTGAAAAG 360 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 302 ATTATTGAGTCTGATAATGAACAAGTACCATGAGGGAAATATGCAAAGGACTTTGAAAAG 361 Query 361 AAAATTAAAAGAGCTTGCATTTGCTAAACAGAAAACTAGCAGAAGTGCTTTATCTTGGTA 420 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 362 AAAATTAAAAGAGCTTGCATTTGCTAAACAGAAAACTAGCAGAAGTGCTTTATCTTGGTA 421 Query 421 AGATTGCTGCGCATGGGTTTACTGTGTAAATTGTATGAGCATTGGTTCTTACTTTGTGTG 480 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 422 AGATTGCTGCGCATGGGTTTACTGTGTAAATTGTATGAGCATTGGTTCTTACTTTGTGTG 481 Query 481 TCAATTCCATTGTCAATTTTAATATCTTTGCTCATGAATTGTGAAATGCTTGCGGGTGTT 540 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 482 TCAATTCCATTGTCAATTTTAATATCTTTGCTCATGAATTGTGAAATGCTTGCGGGTGTT 541 Query 541 AGATTGCATGTGCATTCATTAATTTGGACTTGGTGCAGTGCTTGGCAACAGCTTGCATGT 600 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 542 AGATTGCATGTGCATTCATTAATTTGGACTTGGTGCAGTGCTTGGCAACAGCTTGCATGT 601 Query 601 AGAGTGTGCGTGCAATGTACAATTGCGATTGTCAGCTGTTGCCAACCACTT 651 ||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 602 AGAGTGTGCGTGCAATGTACAATTGCGATTGTCAGCTGTTGCCAACCACTT 652

57
Phụ lục 4. So sánh trình tự vùng D1- D2 một số loài tảo độc thuộc chi Alexandrium với trình tự của A. affine

58
Phụ lục 5. So sánh trình tự vùng D1- D2 một số loài tảo độc thuộc chi
Alexandrium với trình tự của A. peudogonyaulax

59
Phụ lục 6. Kết quả thử nghiệm đầu dò A. affine trong ở các nhiệt độ lai khác nhau Tế bào A.afine dưới ánh sáng thường (trái) và ánh sáng huỳnh quang (phải)
(A, B: 37°C; C, D: 40°C; E, F: 43°C; G, H: 46°C; I, J: 49°C; K, L: 52°C)
A B
C D
F E
G
I
K L
J
H

60
Phụ lục 7. Kết quả thử nghiệm đầu dò A. pseudonyaulax trong ở các nhiệt độ lai khác nhau Tế bào A. pseudonyaulax dưới ánh sáng thường (trái) và ánh sáng huỳnh quang (phải)
(A, B: 37°C; C, D: 40°C; E, F: 43°C; G, H: 46°C; I, J: 49°C; K, L: 52°C)
A B
C D
E
G
I
K L
J
H
F





























