Embed Size (px)
DESCRIPTION
Struktura e membranes qelizore
Citation preview

KAPITULLI DY 2
Struktura e Membranës
SHTRESA E DYFISHTË LIPIDIKE
PROTEINAT E MEMBRANËS
Një qelizë e gjallë është një sistem vetë-riprodhues molekulash të mbyllura brenda një membrane
plazmatike. Kjo membranë është një shtresë aq e hollë dhe transparente lipidike saqë nuk mund të
shihet drejtpërdrejt në mikroskop me dritë. Çdo qelizë në Tokë përdor një membranë për të ndarë dhe
mbrojtur përbërësit e saj kimik nga ambienti i jashtëm. Pa këto membrana, nuk do të kishte qeliza,
dhe si rrjedhojë, nuk do të kishte jetë.
Membrana plazmatike ka formë të thjeshtë: struktura e saj bazohet në dy shtresa molekulare rreth
5nm—ose e trashë sa 50 atome. Vetitë e saj, megjithatë, nuk ngjasojnë me asnjë shtresë të materialit
që ne njohim nga jeta e përditshme. Edhe pse membrana plazmatike shërben për mbrojtjen e
përmbajtjes së qelizës nga ambienti rrethues (Figura 11-1 A), ajo bën shumë më tepër se aq. Që qeliza
të mbijetojë dhe të rritet, materiet ushqyese (nutricientët) duhet të hyjë nëpërmjet membranës, ndërsa
produktet e panevojshme të dalin jashtë saj. Për lehtësimin e këtij shkëmbimi, membrana penetrohet
nga kanale dhe pompa –molekula proteinike tepër përzgjedhëse të cilat lejojnë importin dhe eksportin
e disa substancave specifike.
Figura 11-1 Membranat qelizore shërbejnë si mburoja përzgjedhëse. (A) Membrana plazmatike ndan
qelizën nga ambienti i jashtëm , dhe në rastet e qelizave bakteriale, kjo është e vetmja membranë. Ajo mundëson
që përbërja molekulare e qelizës të ndryshojë nga përbërja e ambientit. (B) Qelizat eukariote kanë membrana të
brendshme që mbështjellin organelet individuale. Në të dyja rastet, membrana pengon përzierjen e molekulave
të njërës anë me ato të anës tjetër.
Proteinat e tjera në membranë shërbejnë si sensorë që i mundësojnë qelizës të marrë informacion në
lidhje me ndryshimet e ambientit dhe të reagojë ndaj tyre (Figura 11-2). Vetitë mekanike të
membranës janë gjithashtu mbreslënëse. Kur një qelizë rritet apo ndryshon formën, membrana e bën
të njëjtën gjë: ajo e zgjeron sipërfaqen e saj duke shtuar pjesë të reja të membranës, pa e humbur
vazhdimësinë, dhe mund të deformohet pa u shkëputur. Nëse membrana shpohet, ajo nuk pëlcet si një
balonë e as nuk mbetet e çarë, por ajo ngjitet përsëri me shpejtësi.
Bakteret më të thjeshta kanë vetëm një membranë – membranën plazmatike. Qelizat eukariote, nga
ana tjetër, kanë shumë membrana të brendshme të cilat mbështjellin hapësirat brendaqelizore për të
formuar organelet e ndryshme, përfshirë këtu dhe rrjetin endoplazmatik, aparatin e Golxhi-t dhe
mitokondriet (Figura 11-3). Këto membrana të brendshme janë ndërtuar me të njëjtat parime si
membrana plazmatike, dhe ato gjithashtu shërbejnë si bariera përzgjedhëse në mes hapësirave që
përmbajnë molekula të ndryshme (shih Figurën 11-1B). Diferencat e lehta në përbërjen e këtyre

membranave, veçanërisht në proteinat e tyre përkatëse, janë ato që i japin secilit organel karakterin e
tij dallues.
Figura 11-2 Membrana plazmatike është e përfshirë në komunikimin e qelizës, importin dhe eksportin e
molekulave, rritjen e qelizës dhe lëvizshmërinë. (1) Proteinat receptore në membranën plazmatike i
mundësojnë qelizës marrjen e sinjaleve nga ambienti; (2) proteinat transportuese në membranë mundësojnë
importin dhe eksportin e molekulave të vogla; (3) fleksibiliteti i membranës dhe kapaciteti i saj për zgjerim
mundëson rritjen dhe lëvizshmërinë e qelizës.
Pavarësisht vendndodhjes së tyre, të gjitha membranat qelizore përbëhen nga lipidet dhe proteinat,
dhe kanë të njëjtën strukturë të përgjithshme (Figura 11-4). Lipidet janë të organizuara në dy shtresa
të afërta me njëra-tjetrën, që formojnë shtresën e dyfishtë lipidike (Shih Figurën 11-4B dhe C). Kjo
shtresë e dyfishtë lipidike i jep membranës srukturën e saj themelorë dhe shërben si një barierë
përzgjedhëse për shumicën e molekulave që treten në ujë. Proteinat kryejnë shumicën e funksioneve
të tjera të membranës dhe i japin membranave të ndryshme karakteristikat e tyre të veçanta.
Figura 11-3
Membranat
formojnë hapësira
të ndryshme të
ndara në një qelizë
eukariote. Këtu janë
paraqitur organelet e
mbështjella me
membranë, në një
qelizë tipike të
kafshëve. Të keni
parasysh që bërthama
dhe mitokondriet janë
secila nga këto të
mbështjellura me dy
membrana.
Në këtë kapitull shqyrtojmë strukturën e membranave biologjike dhe organizimin e dy përbërësve
kryesor të tyre: lipidet dhe proteinat. Edhe pse ne përqëndrohemi kryesisht në membranën plazmatike,
shumica e koncepteve që ne shqyrtojmë vlejnë edhe për membranat e brendshme. Funksionet e

membranave qelizore, përfshirë edhe rolin e tyre në komunikimin e qelizave, transportin e molekulave
të vogla dhe prodhimin e energjisë, do të shqyrtohen në kapitujt pasues.
Figura 11-4 Një membranë qelizore mund të egzaminohet në disa mënyra. (A) Një mikrografi elektronike e
membranës plazmatike të një qelize të kuqe të gjakut të njeriut e parë në prerje tërthore. (B dhe C) Paraqitjet
skematike tregojnë pamje dy- dhe tre-dimensionale të një membrane qelizore. (E mundësuar nga Daniel S.
Friend.)
SHTRESA E DYFISHTË LIPIDIKE
Shtresa e dyfishtë lipidike konsideerohet baza universale e strukturës së membranës, dhe vetitë e saj
janë përgjegjëse për vetitë kryesore të të gjitha membranave qelizore. Për shkak se qelizat janë të
mbushura – dhe të rrethuara – nga tretësira ujore, ne e fillojmë këtë sektor duke egzaminuar se si
struktura e membranave qelizore është një pasojë e sjelljes së membranave lipidike në ambient ujorë.
Lipidet e membranës formojnë shtresa të dyfishta në ujë
Lipidet në membranat qelizore kombinojnë dy veti shumë të ndryshme në një molekulë të vetme: çdo
lipid ka një kokë hidrofile ("që pranon ujin") dhe një ose dy bishta hidrokarbonik hidrofob (që nuk e
pranojnë ujin) (Figura 11-5). Pjesën më të madhe të lipideve në membranat qelizore e përbëjnë
fosfolipidet, molekula në të cilat koka hidrofile lidhet me pjesën tjetër të lipidit nëpërmjet një grupi
fosfat. Lloji më i zakonshëm i fosfolipidit në shumicën e membranave qelizore është fosfatidilkolina,
e cila përmban molekulën e vogël kolinë të lidhur me një fosfat si një kokë hidrofile, dhe dy zinxhirë
hidrokarbonik si bishta të saj hidrofob (Figura 11-6).
Figura 11-5 Një molekulë lipidike e membranës ka një kokë hidrofile
dhe bishta hidrofob.
Molekulat me veti hidrofile dhe hidrofobe quhen amfipatike. Kjo veti kimike gjendet gjithashtu në dy
lloje të tjera të pipideve membranore, përfshirë këtu sterolet (si kolesteroli që gjendet në membranat e
qelizave shtazore) dhe glikolipidet, të cilat kanë sheqerna si pjesë të kokës së tyre hidrofile (Figura 11-
7). Prezenca e përbashkët e pjesëve hidrofile dhe hidrofobe luan një rol kryesor në orientimin e
molekulave lipidike që të grumbullohen në shtresa të dyfishta në një ambient ujorë.
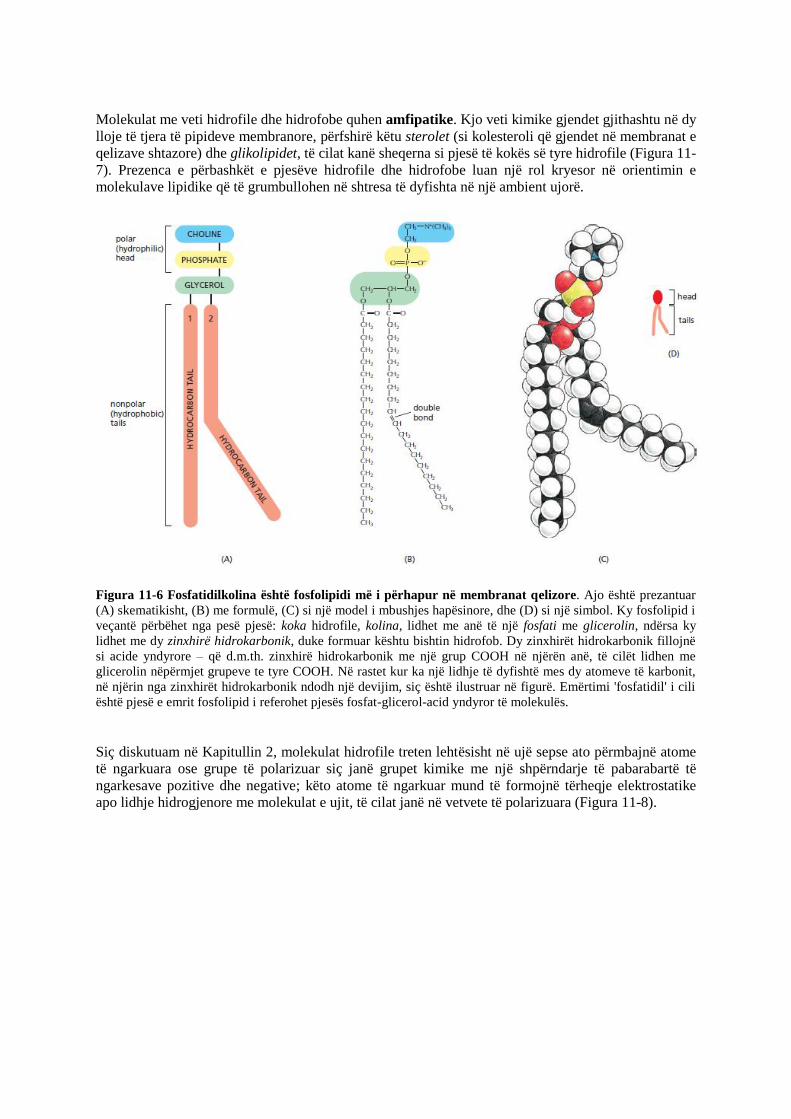
Figura 11-6 Fosfatidilkolina është fosfolipidi më i përhapur në membranat qelizore. Ajo është prezantuar
(A) skematikisht, (B) me formulë, (C) si një model i mbushjes hapësinore, dhe (D) si një simbol. Ky fosfolipid i
veçantë përbëhet nga pesë pjesë: koka hidrofile, kolina, lidhet me anë të një fosfati me glicerolin, ndërsa ky
lidhet me dy zinxhirë hidrokarbonik, duke formuar kështu bishtin hidrofob. Dy zinxhirët hidrokarbonik fillojnë
si acide yndyrore – që d.m.th. zinxhirë hidrokarbonik me një grup COOH në njërën anë, të cilët lidhen me
glicerolin nëpërmjet grupeve te tyre COOH. Në rastet kur ka një lidhje të dyfishtë mes dy atomeve të karbonit,
në njërin nga zinxhirët hidrokarbonik ndodh një devijim, siç është ilustruar në figurë. Emërtimi 'fosfatidil' i cili
është pjesë e emrit fosfolipid i referohet pjesës fosfat-glicerol-acid yndyror të molekulës.
Siç diskutuam në Kapitullin 2, molekulat hidrofile treten lehtësisht në ujë sepse ato përmbajnë atome
të ngarkuara ose grupe të polarizuar siç janë grupet kimike me një shpërndarje të pabarabartë të
ngarkesave pozitive dhe negative; këto atome të ngarkuar mund të formojnë tërheqje elektrostatike
apo lidhje hidrogjenore me molekulat e ujit, të cilat janë në vetvete të polarizuara (Figura 11-8).
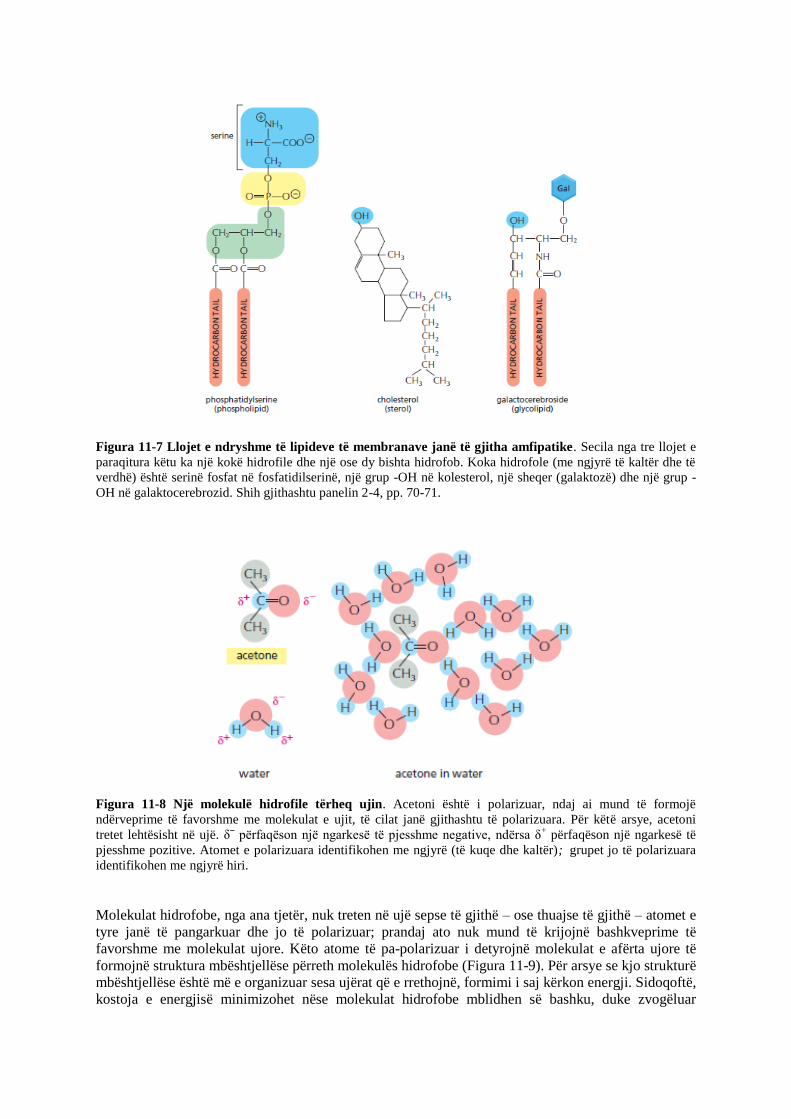
Figura 11-7 Llojet e ndryshme të lipideve të membranave janë të gjitha amfipatike. Secila nga tre llojet e
paraqitura këtu ka një kokë hidrofile dhe një ose dy bishta hidrofob. Koka hidrofole (me ngjyrë të kaltër dhe të
verdhë) është serinë fosfat në fosfatidilserinë, një grup -OH në kolesterol, një sheqer (galaktozë) dhe një grup -
OH në galaktocerebrozid. Shih gjithashtu panelin 2-4, pp. 70-71.
Figura 11-8 Një molekulë hidrofile tërheq ujin. Acetoni është i polarizuar, ndaj ai mund të formojë
ndërveprime të favorshme me molekulat e ujit, të cilat janë gjithashtu të polarizuara. Për këtë arsye, acetoni
tretet lehtësisht në ujë. δˉ përfaqëson një ngarkesë të pjesshme negative, ndërsa δ+ përfaqëson një ngarkesë të
pjesshme pozitive. Atomet e polarizuara identifikohen me ngjyrë (të kuqe dhe kaltër); grupet jo të polarizuara
identifikohen me ngjyrë hiri.
Molekulat hidrofobe, nga ana tjetër, nuk treten në ujë sepse të gjithë – ose thuajse të gjithë – atomet e
tyre janë të pangarkuar dhe jo të polarizuar; prandaj ato nuk mund të krijojnë bashkveprime të
favorshme me molekulat ujore. Këto atome të pa-polarizuar i detyrojnë molekulat e afërta ujore të
formojnë struktura mbështjellëse përreth molekulës hidrofobe (Figura 11-9). Për arsye se kjo strukturë
mbështjellëse është më e organizuar sesa ujërat që e rrethojnë, formimi i saj kërkon energji. Sidoqoftë,
kostoja e energjisë minimizohet nëse molekulat hidrofobe mblidhen së bashku, duke zvogëluar

kontaktin e tyre me ujin në minimumin e mundshëm. Si rrjedhojë, molekulat tërësisht hidrofobe, si
yndyrnat që gjenden në qelizat yndyrore të shtazëve dhe vajrat që gjenden në farat e bimëve (Figura
11-10A), mblidhen së bashku në një pikë të vetme kur hidhen në ujë.
Figura 11-9 Një molekulë hidrofobe ka tendencën ta shmangë ujin. Për arsye se molekula 2-metilpropan
është krejtësisht hidrofobe, ajo nuk mund të formojë ndërveprime të favorshme me ujin, dhe i detyron molekulat
e ujit përreth të riorganizohen në formë rrethore përreth saj.
Në ndryshim me të parat, molekulat amfipatike, si fosfolipidet (Figura 1l-10B), janë subjekt i dy
forcave të kundërta: koka hidrofile tërhiqet nga uji, ndërsa bishti hidrofob e largon ujin dhe tenton të
lidhet me molekula të tjera hidrofobe. Ky konflikt zgjidhet më së miri nga krijimi i shtresës së
dyfishtë lipidike – një marrrëdhënie që i kënaq të gjitha palër dhe ështe më e favorshmja nga aspekti
energjitik. Kokat hidrofile i qasen ujit nga të dy anët e bishtrsës; bishtat hidrofob janë të mbrojtur nga
uji ndërsa ata qëndrojnë afër njëri tjetrit në brendësi, si mbushja e një senduiçi (Figura 11-11).
Figura 11-10 Molekulat
yndyrore janë hidrofobe, ndërsa
fosfolipidet janë amfipatike. (A)
Triacilglicerolet, të cilët janë
përbërësit kryesor të yndyrave
shtazore dhe vajrave bimor, janë
tërësisht hidrofob. (B) Fosfolipidet
si psh. fosfatidiletanolamina janë
amfipatike dhe përmbajnë si pjesë
hidrofobe, ashtu dhe hidrofile.
Pjesët hidrofobe janë me ngjyrë të
kuqe, ndërsa pjesët hidrofile me
ngjyrë të kaltër dhe të verdhë.
(Bishti i tretë hidrofob i molekulës
së triacilglicerolit është paraqitur
me drejtim nga lart, për tu
krahasuar me fosfolipidin, edhe
pse normalisht ai paraqitet me
drejtim për poshtë.)
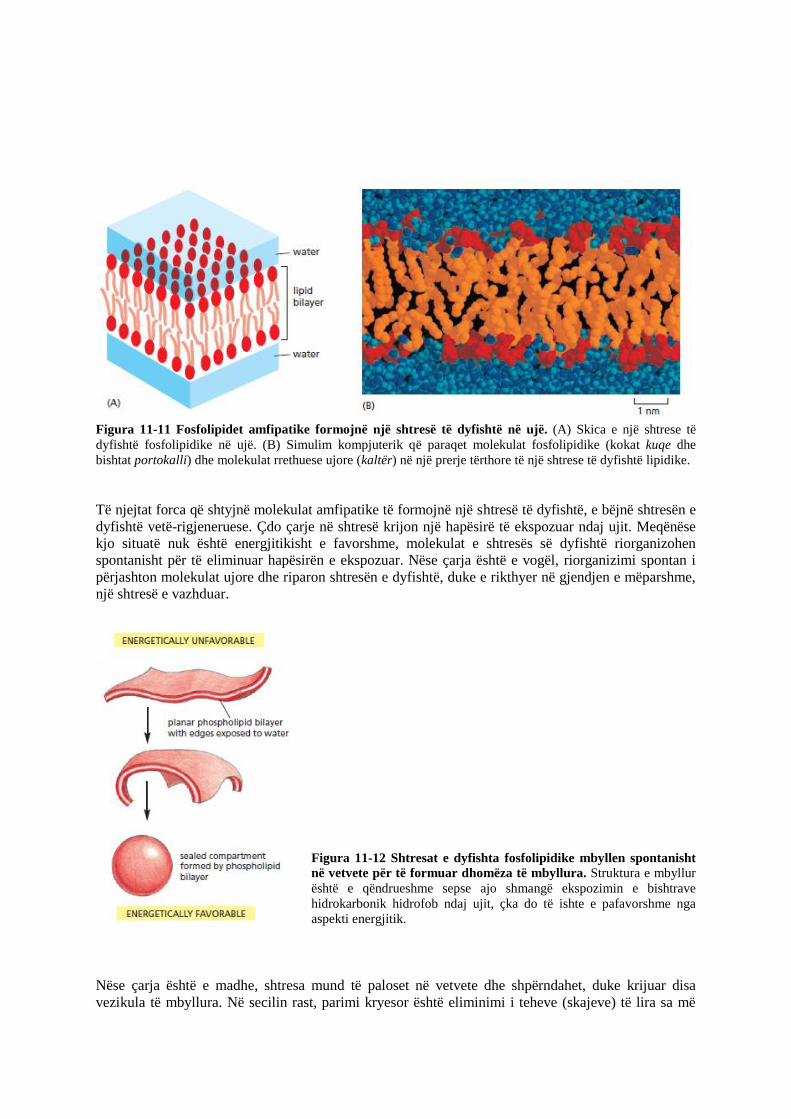
Figura 11-11 Fosfolipidet amfipatike formojnë një shtresë të dyfishtë në ujë. (A) Skica e një shtrese të
dyfishtë fosfolipidike në ujë. (B) Simulim kompjuterik që paraqet molekulat fosfolipidike (kokat kuqe dhe
bishtat portokalli) dhe molekulat rrethuese ujore (kaltër) në një prerje tërthore të një shtrese të dyfishtë lipidike.
Të njejtat forca që shtyjnë molekulat amfipatike të formojnë një shtresë të dyfishtë, e bëjnë shtresën e
dyfishtë vetë-rigjeneruese. Çdo çarje në shtresë krijon një hapësirë të ekspozuar ndaj ujit. Meqënëse
kjo situatë nuk është energjitikisht e favorshme, molekulat e shtresës së dyfishtë riorganizohen
spontanisht për të eliminuar hapësirën e ekspozuar. Nëse çarja është e vogël, riorganizimi spontan i
përjashton molekulat ujore dhe riparon shtresën e dyfishtë, duke e rikthyer në gjendjen e mëparshme,
një shtresë e vazhduar.
Figura 11-12 Shtresat e dyfishta fosfolipidike mbyllen spontanisht
në vetvete për të formuar dhomëza të mbyllura. Struktura e mbyllur
është e qëndrueshme sepse ajo shmangë ekspozimin e bishtrave
hidrokarbonik hidrofob ndaj ujit, çka do të ishte e pafavorshme nga
aspekti energjitik.
Nëse çarja është e madhe, shtresa mund të paloset në vetvete dhe shpërndahet, duke krijuar disa
vezikula të mbyllura. Në secilin rast, parimi kryesor është eliminimi i teheve (skajeve) të lira sa më

shpejt. Parandalimi i hapësirave të lira ka një rrjedhojë domethënëse: e vetmja mënyrë që një shtresë
të shmangë hapësira të lira është që ajo shtresë të paloset dhe ngjitet, duke formuar kështu një kufi
rreth një hapësire të mbyllur (Figura 11-12). Rrjedhimisht, molekulat amfipatike si fosfolipidet
mblidhen detyrimisht duke krijuar dhomëza të mbyllura. Kjo sjellje mbresëlënëse, esenciale për
krijimin e një qelize të gjallë, është në thelb rezultati i vetisë që çdo molekulë është hidrofile në një
anë, dhe hidrofobe në anën tjetër.
Shtresa e dyfishtë lipidike është një lëng dy-dimensional
Ambienti ujor brenda dhe jashtë qelizës i pengon lipidet e membranës për të dalë nga shtresa e
dyfishtë, por asgjë nuk i ndalon këto molekula të levizin rrotull dhe të ndërrojnë vendet brenda
hapësirës së shtresës së dyfishtë. Si rrjedhojë, membrana sillet si një lëng dy-dimensional, i cili është
thelbësor për funksionimin dhe integritetin e membranës (Filmimi 11.1). Kjo veti dallon nga
fleksibiliteti (lakueshmëria), që është aftësia e membranës të lakohet. Fleksibiliteti i membranës është
gjithashtu i rëndësishëm, dhe ai përcakton një kufi më të ulët prej rreth 25 nm në madhësinë e
vezikulës, që membranat qelizore mund të formojnë.
Fluiditeti i shtresave të dyfishta lipidike mund të studiohet duke përdorur shtresa të dyfishta lipidike
sintetike, të cilat mund të prodhohen lehtësisht nga grumbullimi spontan i molekulave amfipatike
lipidike në ujë. Janë dy lloje të shtresave të dyfishta lipidike sintetike që përdoren gjerësisht në
eksperimente. Vezikulat e mbyllura sferike, të quajtura lipozome, formohen nëse fosfolipidet e pastra i
shtohen ujit; diametri i tyre varion nga 25 nm deri në 1 mm (Figura 11-13). Si alternativë, shtresat e
dyfishta të rrafshta fosfolipidike mund të formohen përgjatë një vrime në një ndarje në mes dy
hapësirave ujore (Figura 11-14).
Figura 11-13 Fosfolipidet e pastra mund të formojnë lipozome të mbyllura sferike. (A) Një mikrografi
elektronike e vezikulave fosfolipidike që tregon strukturën e shtresës së dyfishtë të membranës. (B) Një vizatim
i lipozomeve të vogla sferike në prerje tërthore.
Këto shtresa të dyfishta të thjeshta artificiale lejojnë matjen e lëvizjes së molekulave lipidike, duke
treguar që disa lëvizje janë shumë të rralla ndërsa disa të tjera shumë të shpeshta dhe të shpejta. Si
rrjedhojë, në shtresat e dyfishta lipidike sintetike, molekulat fosfolipidike rrallëherë kalojnë nga njëra
gjysëm e shtresës së dyfishtë, apo shtresës së njëfishtë, te gjysma tjetër. Në mungesë të proteinave për
ta lehtësuar këtë proces dhe në kushte të ngjashme me ato të qelizës, vlerësohet që ky rast, i quajtur
“flip-flop”, ndodh më rrallë se një herë në muaj për secilën molekulë lipidike individuale. Nga ana
tjetër, si rezultat i veprimeve termike, molekulat lipidike brenda njërës shtresë vazhdimisht ndërrojnë
vendet me njëra-tjetrën (Figura 11-15). Ky shkëmbim shkakton përhapjen e shpejtë të molekulave
lipidike në rrafshin e membranës, në mënyrë që, për shembull, një lipid në një shtresë të dyfishtë
artificiale mund të zgjerohet në madhësinë e një qelize të madhe bakteriale (~2 µm) brenda një

sekonde. Nëse temperatura ulet, rënia e energjisë termale e ulë ritmin e lëvizjes lipidike, duke e bërë
shtresën e dyfishtë më pak fluide.
Figura 11-14 Një shtresë e dyfishtë fosfolipidike sintetike mund të formohet përmes një vrime të vogël
(me diametër rreth 1mm) në ndarjen në mes dy hapësirave ujore. Për ta bërë shtresën e dyfishtë të rrafshët,
pjesëza shytet në një solucion ujor dhe një solucion fosfolipidik (në një tretës jo-ujor) stoliset përmes vrimës me
një furçë.
Rezultate të ngjashme arrihen kur egzaminohen membrana qelizore dhe qeliza të plota të izoluara, çka
tregon që shtresa e dyfishtë lipidike e një membrane qelizore sillet gjithashtu si një lëng dy-
dimensional brenda të cilit molekulat përbërëse lipidike janë të lira të lëvizin brenda shtresës së tyre
në çdo drejtim të hapësirës së membranës. Këto studime gjithashtu tregojnë që zinxhirët lipidik
hidrokarbonik janë elastik dhe që molekulat individuale lipidike brenda njërës shtresë rrotullohen me
shumë shpejtësi rreth akseve të tyre të gjatë—disa arrijnë shpejtësi prej 30,000 rrotullime në minut
(shih Figurën 11 -15). Në qeliza si dhe në shtresa e dyfishtë lipidike sintetike, molekulat individuale
fosfolipidike normalisht lëvizin brenda hapësirës së njërës shtresë të tyre dhe nuk “kërcejnë”
spontanisht.
Figura 11-15 Fosfolipidet mund të lëvizin
brenda planit të membranës. Skica tregon
tipet e lëvizjes së mundshme të molekulave të
fosfolipideve në një shtresë të dyfishtë të
fosfolipideve.
Fluiditeti i një shtrese të dyfishtë lipidike lipidike varet nga përbërja e saj
Fluiditeti i një membranë qelizore – lehtësia me të cilën molekulat e saj lipidike lëvizin në hapësirën e
shtresës së dyfishtë lipidike – është i rëndësishëm për funksionimin e membranës dhe duhet të
përmbahet brenda disa kufijve. Fluiditeti i një shtrese të dyfishtë lipidike lipidike në një temperaturë
të caktuar varet nga përbërja e saj fosfolipidike dhe, në veçanti, nga natyra e bishtave hidrokarbonik:
sa më e afërt dhe më e rregullt vendosja e këtyre bishtave, aq më viskoze dhe më pak fluide do jetë
shtresa e dyfishtë lipidike. Janë veti kryesore të bishtave hidrokarbonik që ndikojnë se sa afër ata
paketohen në shtresën e dyfishtë lipidike: gjatësia e tyre dhe numri i lidhjeve të dyfishta që ata
përmbajnë.
Një gjatësi më e shkurtër e zinxhirit e ulë tendencën e bishtave hidrokarbonik për të ndërvepruar me
njëra-tjetrën, dhe ndërkohë rritë fluiditetin e shtresës së dyfishtë lipidike. Gjatësia e bishtave
hidrokarbonik të fosfolipideve të membranës ndryshon në mes 14 dhe 24 atome karboni, ndërsa
gjatësia më e zakonshme është 18-20 atome karboni. Shumica e fosfolipideve përmbajnë një bisht
hidrokarbonik që ka një ose më shumë lidhje të dyfishta në mes atomeve të afërta të karbonit, dhe një
bisht të dytë që ka vetëm lidhje të njëfishta (shih Figurën 11-6). Zinxhiri që përmban një lidhje të
dyfishtë nuk përmban numrin maksimal të atomeve të hidrogjenit që mund, në parim, të bashkohen
me strukturën e tij karbonike; në këtë rast thuhet që zinxhiri është i pangopur me hidrogjen. Bishti
me acid yndyror pa lidhje të dyfishta ka numrin e plotë të atomeve të hidrogjenit; ky quhet i ngopur.
Secila lidhje e dyfishtë në një bisht të pangopur krijon një devijim të vogël në bishtin hidrokarbonik
(shih Figurën 11-6), çka e bën më të vështirë për bishtat të ngjeshen me njëri-tjetrin. Për këtë arsye,
shtresat e dyfishta lipidike që përmbajnë shumë bishta hidrokarbonik të pangopur janë më fluid sesa
ato me një numër më të vogël të tyre.
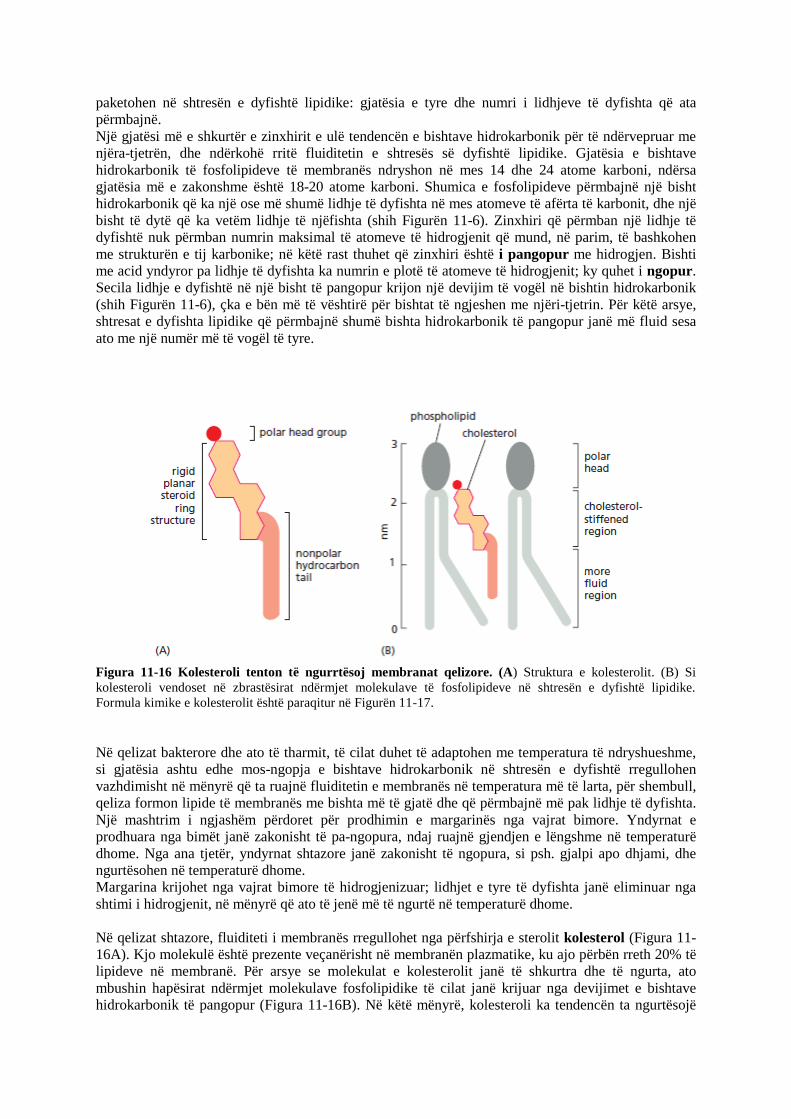
Figura 11-16 Kolesteroli tenton të ngurrtësoj membranat qelizore. (A) Struktura e kolesterolit. (B) Si
kolesteroli vendoset në zbrastësirat ndërmjet molekulave të fosfolipideve në shtresën e dyfishtë lipidike.
Formula kimike e kolesterolit është paraqitur në Figurën 11-17.
Në qelizat bakterore dhe ato të tharmit, të cilat duhet të adaptohen me temperatura të ndryshueshme,
si gjatësia ashtu edhe mos-ngopja e bishtave hidrokarbonik në shtresën e dyfishtë rregullohen
vazhdimisht në mënyrë që ta ruajnë fluiditetin e membranës në temperatura më të larta, për shembull,
qeliza formon lipide të membranës me bishta më të gjatë dhe që përmbajnë më pak lidhje të dyfishta.
Një mashtrim i ngjashëm përdoret për prodhimin e margarinës nga vajrat bimore. Yndyrnat e
prodhuara nga bimët janë zakonisht të pa-ngopura, ndaj ruajnë gjendjen e lëngshme në temperaturë
dhome. Nga ana tjetër, yndyrnat shtazore janë zakonisht të ngopura, si psh. gjalpi apo dhjami, dhe
ngurtësohen në temperaturë dhome.
Margarina krijohet nga vajrat bimore të hidrogjenizuar; lidhjet e tyre të dyfishta janë eliminuar nga
shtimi i hidrogjenit, në mënyrë që ato të jenë më të ngurtë në temperaturë dhome.
Në qelizat shtazore, fluiditeti i membranës rregullohet nga përfshirja e sterolit kolesterol (Figura 11-
16A). Kjo molekulë është prezente veçanërisht në membranën plazmatike, ku ajo përbën rreth 20% të
lipideve në membranë. Për arsye se molekulat e kolesterolit janë të shkurtra dhe të ngurta, ato
mbushin hapësirat ndërmjet molekulave fosfolipidike të cilat janë krijuar nga devijimet e bishtave
hidrokarbonik të pangopur (Figura 11-16B). Në këtë mënyrë, kolesteroli ka tendencën ta ngurtësojë

shtresën e dyfishtë, duke e bërë atë më pak të depërtueshme. Vetitë kimike të lipideve të membranës –
dhe si ato e ndikojnë fluiditetin e membranës – shqyrtohen në Filmimin 11.2.
Fluditeti i membranës është i rëndësishëm për gjithë qelizat, për shumë arsye. Kjo i lejon proteinat e
membranës të përhapen me shpejtësi në hapësirën e shtresës së dyfishtë dhe të ndërveprojnë me njëra-
tjetrën, çka është tepër e rëndësishme sidomos për sinjalizimin e qelizave (diskutohet në Kapitullin
16). Fluiditeti u lejon lipideve dhe proteinave të membranës të përhapen në shtresën e dyfishtë pas
sintezës së tyre në zona të tjera të qelizës. Ai u mundëson membranave të bashkohen me njëra-tjetrën
dhe të bashkojnë molekulat e tyre, dhe siguron që molekulat e membranës shpërndahen në mënyrë të
barabartë mës qelizave bija pasi një qelizë ndahet. Nëse membranat biologjike nuk do ishin fluide, do
ishte shumë e vështirë të imagjinoje si një qelizë do të jetonte, të rritej dhe të shumohej.
Shtresa e dyfishtë lipidike është asimetrike
Membranat qelizore janë në përgjithësi asimetrike; ato kanë një pamje krejt tjetër kah brendia e
qelizës apo organelit krahasuar me atë në anën e jashtme. Dy gjysmat e shtresës së dyfishtë shpesh
përmbajnë grupe krejtësisht të ndryshme fosfolipidesh dhe glikolipidesh (Figura 11-17). Për më tepër,
proteinat e membranës janë të integruara në shtresën e dyfishtë me një orientim specifik, çka është
tepër e rëndësishme për funksionin e tyre.
Figura 11-17 Fosfolipidet dhe glikolipidet shpërndahen në mënyrë asimetrike në membranën plazmatike
të shtresës së dyfishtë lipidike. Pesë llojet e molekulave fosfolipidike paraqiten me ngjyra të ndryshme:
fosfatidilkolina (e kuqe), sfingomelina (kafe), fosfatidilserina (e gjelbër e qelët), fosfatifilnozitol (e gjelbër e
errët), dhe fosfatidiletanolamina (e verdhë). Glikolipidet janë vizatuar me grupe kokash heksagonale të kaltra,
për të përfaqësuar sheqernat. Të gjitha molekulat glikolipidike janë në shtresën e njëfishtë të jashtme të
membranës, ndërsa kolesteroli (hiri) është shpërndarë thuajsë njëtrajtësisht në dy shtresat e njëfishta.
Fosfatidilnozitoli (nuk paraqitet këtu) është një lipid i vogël që ndodhet gjithmonë në shtresën e njëfishtë në
anën citosolike të membranës plazmatike, ku shërben për sinjalizim në qelizë. Përderisa grupi i kokës së tij është
një sheqer inozitol ai është një përjashtim, meqë ndodhet diku tjetër për dallim nga ajo se ku janë të gjitha
glikolipidet e tjera.
Asimetria lipidike krijohet dhe ruhet derisa membrana rritet. Në qelizat eukariote, fosfolipidet e reja
prodhohen nga enzimat e lidhura me pjesën e membranës së rrjetit endoplazmatik, me drejtim nga
citosoli. Këto enzima, të cilat përdorin acidet e lira yndyrore si substrat (shih Panelin 2-4, pp. 70-71),
depozitojnë gjithë fosfolipidet e sapo prodhuara në gjysmën citosolike të shtresës së dyfishtë. Për të
mundësuar rritjen uniforme të gjithë membranës, gjysma e molekulave të reja fosfolipidike duhet të
transferohen te shtresa tjetër e njëfishtë. Ky transferim përshpejtohet nga enzimat e quajtura flipaza
(Figura 11-18). Në membranën plazmatike, flipazat transferojnë fosfolipidet specifike në mënyrë të
përzgjedhur, në mënyrë që lloje të ndryshme të përqëndrohen në çdo shtresë të njëfishtë.

Figura 11-18 Flipazat luajnë rol në sintezën e shtresës së dyfishtë lipidike. Molekulat fosfolipidike të sapo
sintetizuara i shtohen që të gjitha anës citosolike të membranës së REP. Flipazat më pas transferojnë disa nga
këto molekula në shtresën tjetër të njëfishtë, në mënyrë që zgjerohet e tërë shtresa e dyfishtë.
Sidoqoftë, përdorimi i flipaseve selektive nuk është menyra e vetme për të prodhuar asimetri në
shtresat e dyfishta lipidike. Një mekanizëm tjetër funksionon për glikolipidet – lipidet që shfaqin
asimetrinë më të theksuar dhe të vazhdueshme në qelizat shtazore (shih Figurën 11-17). Për të
shpjeguar shpërndarjen e tyre, është i nevojshëm shqyrtimi i mënyrës si membrana e re prodhohet në
qelizat eukariote.
Asimetria lipidike ruhet gjatë transportimit të membranës
Thuajse gjithë sintezat e reja të membranave në qelizat eukariote ndodhin në membranën e një
hapësire brendaqelizore – retikulumin endoplazmatik (REP) (shih Figurën 11-3). Membrana e re e
ndërtuar aty eksportohet te membranat e reja të qelizës nëpërmjet një cikli krijimi dhe bashkimi
membranash: copëza të shtresës së dyfishtë shkëputen nga REP dhe formojnë sfera të vogla të
quajtura vezikula, të cilat më pas inkorporohen në një membranë tjetër, si membrana plazmatike, duke
u ngjitur me të (Figura 11-19). Orientimi i shtresës së dyfishtë në lidhje me citosolin ruhet gjatë
formimit dhe ngjitjes së vezikulës. Kjo ruajtje e orientimit tregon që gjithë membranat qelizore, qoftë
membrana e jashtme plazmatike ashtu dhe membrana ndërqelizore përreth ornganelit, kanë anë të
ndryshme të brendshme dhe të jashtme, të cilat përcaktohen në kohën e sintezës së membranës: ana
citosolike është gjithmonë pranë citosolit, ndërsa ana jo-citosolike është e ekspozuar si ndaj pjesës së
jashtme të qelizës, ashtu dhe ndaj pjesës së brendshme të organelit (shih Figurën 11-19).

Figura 11-19 Membranat e ruajnë orientimin
e tyre edhe pasi trasnferohen nga njëra anë e
qelizës tek tjetra. Membranat transportohen
nëpërmjet procesit të krijimit dhe ngjitjes së
vezikulave. Këtu, një vezikul paraqitet duke u
krijuar nga një organel i mbështjellë nga
membrana (si rrjeti endoplazmik ose Golxhi)
dhe duke u bashkuar me membranën plazmatike.
Kini parasysh që orientimi i membranës ruhet
gjatë procesit: sipërfaqja origjinale citosolike (e
kuqe) mbetet e kthyer kah citosoli dhe sipërfaqja
jo-citosolike (portokalli) vazhdon të qëndrojë e
kthyer në anën e kundërt me citosolin, në drejtim
të lumenit të organelit apo vezikulës, ose drejtë
lëngut jashtëqelizor.
Glikolipidet ndodhen kryesisht në membranën plazmatike, edhe ata gjenden vetëm në gjysmën jo-
citosolike të shtresës së dyfishtë. Për këtë arsye, grupet e tyre të sheqerit janë të ekspozuar ndaj pjesës
së jashtme të qelizës (shih Figurën 11-17), ku ato formojnë pjesën e një mbulese mbrojtëse të
vazhdueshme të karbohidrateve e cila mbështjell shumicën e qelizave shtazore. Molekulat
glikolipidike i përfitojnë grupet e tyre të sheqerit në aparatin e Golxhit, organeli ku mblidhen
zakonisht proteinat dhe membranat e prodhuara në REP (studiohet në kapitullin 15). Enzimat që
shtojnë grupet e sheqerit qëndrojnë brenda aparatit të Golxhit, në mënyrë që sheqernat ti shtohen
vetëm molekulave lipidike që ndodhen në gjysmën jo-citosolike të shtresës së dyfishtë lipidike. Pasi
molekula glikolipidike të jetë krijuar në këtë mënyrë, ajo mbetet në këtë anë të shtresës së dyfishtë,
sepse nuk ka flipaza që ti trasnferojnë glikolipidet te shtresa në anën citosolike. Pra, kur një molekulë
glikolipidike transportohet te membrana plazmatike, ajo nuk drejtohet nga citosoli dhe ekspozon
sheqerin te pjesa e jashtme e qelizës (shih Figurën 11-19).
Molekula të tjera lipidike tregojnë lloje të tjera të shpërndarjeve asimetrike, të lidhura me funksione të
tjera. Për shembull, inozitolet fosfolipidike janë përbërës të vegjël të membranës plazmatike, por ata
luajnë një rol të veçantë në përçimin e sinjaleve nga sipërfaqja e qelizës te përbërësit ndërqelizor të
cilët reagojnë ndaj këtyre sinjaleve (studiohet ne Kapitullin 16). Ata veprojnë vetëm pasi sinjali është
transmetuar përmes membranës plazmatike; pra ata janë të përqëndruar te gjysma citosolike e shtresës
së dyfishtë lipidike (shih Figurën 11-17).
PROTEINAT E MEMBRANËS
Edhe pse shtresa e dyfishtë lipidike ofron strukturën bazë për gjithë membranat qelizore dhe shërben
si një barierë e lëshueshmërisë (permeabilitetit) për molekulat, shumica e funksioneve të membranës
kryhet nga proteinat e membranës. Te shtazët, proteinat përbëjnë rreth 50% të masës së shumicës së
membranave plazmatike, ndërsa pjesa tjetër janë lipide, pak karbohidrate që gjenden në glikolipide,
dhe proteina të glikoziluara. Për arsye se molekulat lipidike janë shumë më të vogla se proteinat, një
membranë qelizore normalisht përmban rreth 50 herë më shumë molekula lipidike sesa molekula
proteinike (shih Figurën 11-4).
Proteinat e membranës jo vetëm që transportojnë materie ushqyese të caktuar, metabolite dhe jonë
përgjatë shtresës së dyfishtë lipidike; ato gjithashtu kanë shume funksione të tjera. Disa e “ankorojnë”
(ngjesin) membranën për makromolekulat në të dy anët. Të tjerat funksionojnë si receptorë që
dallojnë sinjalet kimike në ambientin e qelizës dhe i përçojnë ato në brendësi të qelizës, dhe tjerat
funksionojnë si enzima për të katalizuar reaksione të caktuara (Figura 11-20 dhe Tabela 11-1). Secili
lloj membranë qelizore përmban një grup të ndryshëm proteinash, dhe kryen funksionet e veçanta të

membranës specifike. Në këtë kapitull, do të diskutojmë strukturën e proteinave të membranës dhe
ilustrojmë mënyrat e ndryshme si ato mund të lidhen me shtresën e dyfishtë lipidike.
Figura 11-20 Proteinat e membranës palzmatike kanë shumë funksione.
Proteinat e membranës lidhen me shtresën e dyfishtë lipidike në mënyra të ndryshme
Proteinat mund të lidhen me shtresën e dyfishtë lipidike të një membrane qelizore në disa mënyra
(Figura 11-21).
1. Shumë proteina të membranës përhapen përgjatë shtresës së dyfishtë, ndërsa një pjesë e
masës së tyre mbetet në të dyja anët (Figura 11-21 A). Njësoj si lipidet, këto proteina
transmembranore kanë gjithashtu zona hidrofobe dhe hidrofile. Zonat e tyre hidrofobe
ndodhen në brendësi të shtresës së dyfishtë, afër bishtave hidrofob të molekulave lipidike.
Zonat hidrofile janë të ekspozuara ndaj ambienteve ujore nga të dyja anët e membranës.
2. Proteinat e tjera të membranës ndodhen tërësisht në citosol, të lidhura me faqen e
brendshme të shtresës së dyfishtë lipidike me anë të një a heliksi amfipatik i ekspozuar në
sipërfaqe të proteinës (Figura 11-21B).
3. Disa proteina qëndrojnë krejtësisht jashtë shtresës së dyfishtë, në njërën anë ose
tjetrën, të lidhura me membranën vetëm me një ose më shumë grupe lipidike me anë të
lidhjeve kovalente (Figura 11-21C).
4. Proteina të tjera janë të lidhura indirekt me njërën nga anët e membranës, dhe rrinë
fiksuar avetëm nga ndërveprimi i tyre me proteinat e tjera të membranës (Figura 11-21D).
Proteinat që janë të lidhura drejtpërdrejt me shtresën e dyfishtë lipidike—qofshin ato
transmembranore, të lidhura me shtresën e njëfishtë, apo të lidhura me lipide—mund të zhvendosen
vetëm duke shkatërruar shtresën e dyfishtë me detergjent. Këto proteina njihen si proteina integrale të
membranës. Grupi tjetër i proteinave të membranës njihet si proteinat periferike të membranës; ato
mund të shkëputen nga membrana më lehtësisht, me procedura që ndërhyjnë në reagimet proteinë-
proteinë por e ruajnë shtresën e dyfishtë lipidike të paprekur.

Figura 11-21 Proteinat e membranës mund të lidhen me shtresën e dyfishtë lipidike në disa mënyra të
ndryshme. (A) Proteinat transmembranore mund të shtrihen përgjatë shtresës së dyfishtë si një α heliks i vetëm,
si disa α helikse, ose të mbështjella në një fletë β (që fuçi β). (B) Disa proteina të membranës lidhen me
sipërfaqen citosolike me anë të një α heliksi amfipatik. (C) Të tjera lidhen me njërën nga anët e shtresës së
dyfishtë vetëm me anë të një lidhje kovalente me një molekulë lipidike (vijat zig zage të kuqe). (D) Shumë
proteina janë të lidhura me membranën vetëm më anë të ndërveprimeve relativish të dobëta jokovalente me
proteinat e tjera të membranës.
Një zinxhir polipeptid zakonisht e përshkon shtresën e dyfishtë si një α Heliks
Të gjitha proteinat e membranës kanë një orientim unik në shtresën e dyfishtë lipidike, çka është
esenciale për funksionin e tyre. Për shembull, për një proteinë receptore transmembranore, pjesa që
shërben për marrjen e sinjalit nga ambienti duhet të jetë nga ana e jashtme e qelizës, ndërsa pjesa që
përçon sinjalin duhet të jetë në citosol (shih Figurën 11-20). Ky orientim është rrjedhojë e mënyrës si
proteinat e membranës sintetizohen (siç trajtohet në Kapitullin 15). Pjesëzat e proteinës
transmembranore të vendosura në secilën anë të shtresës së dyfishtë lipidike lidhen nga segmente të
veçanta të zinxhirit polipeptid të cilat shtrihen përgjatë membranës (shih Figurën 11-21 A). Këto
segmente, të cilat shtrihen përgjatë gjithë ambientit hidrofobik të brendësisë së shtresës së dyfishtë
lipidike, përbëhen gjerësisht nga amino acide me zinxhirë anësorë hidrofob. Për arsye se këta zinxhirë
anësorë nuk mund të ndërveprime të favorshme me molekulat e ujit, ato preferojnë ambientin lipidik,
ku uji mungon.
Për dallim nga zinxhirët anësorë hidrofob, lidhjet peptide që bashkojnë aminno acidet në një proteinë
janë zakonisht të polarizuara, duke e kthyer shtyllën polipeptide në hidrofile (Figura 11-22). Për arsye
se uji mungon në shtresën e dyfishtë, atomet që formojnë shtyllën bazë kanë tendencën të krijojnë
lidhje hidrogjenore me njëri-tjetrin. Lidhjet hidrogjenore përforcohen nëse zinxhiri polipeptid formon

Figura 11-22 Lidhjet peptide (të hijëzuara me ngjyrë hiri) që bashkojnë amino acidet së bashku në një
zinxhir polipeptid janë të polarizuara, pra janë hidrofile. Ngarkesat e pjesshme (δˉ tregon një ngarkesë të
pjesshme negative ndërsa δ+ një ngarkesë të pjesshme pozitive) u lejojnë këtyre atomeve të lidhen me lidhje
hidrogjenore me njëri-tjetrin kur polipeptidi paloset duke formuar një α heliks që zgjeron membranën (shih
Figurën 11-23).
një α heliks të rregullt, dhe kështu shumica e segmenteve të zinxhirëve polipeptidik përshkojnë
shtresën e dyfishtë si α helikse (shih Figurën 4-10). Në këto α helikse që shtrihen në tërë membranën,
zinxhirët anësorë hidrofob janë të ekspozuar nga ana e jashtme e heliksit, ku ata mund të kontaktojnë
bishtat hidrofob lipidik, ndërsa atomet në shtyllën polipeptide formojnë lidhje hidrogjenore me njëri-
tjetrën në brendësi të heliksit (Figura 11-23).
Figura 11-23 Një segment i një α heliksi përshkon
shtresën e dyfishtë lipidike. Zinxhirët anësor
hidrofob të amino acideve formojnë kontaktin mes α
heliksit dhe bishtave hidrokarbonik hidrofibik të
molekulave fosfolipidike, ndërsa pjesët hidrofile të
shtyllës së polipeptidit formojnë lidhje hidrogjenore
me njëra-tjetrën në brendësi të heliksit. Një α heliks
duhet të përmbaj rreth 20 amino acide që ta përshkojë
tërë membranën.
Në shumë proteina transmembranore, zinxhiri polipeptid përshkon membranën vetëm një herë (shih
Figurën 11-21 A). Shumë nga këto proteina janë receptorë për sinjalet jashtëqelizore. Proteinat e tjera

transmemranore formojnë pore ujore që lejojnë molekula të tretshme në ujë të kalojnë membranën.
Këto pore nuk mund të formohen nga proteina me një α heliks transmembranor të vetëm dhe
uniformisht hidrofob. Proteinat që formojnë poret janë më të ndërlikuara, zakonisht kanë një seri α
heliksesh që përshkojnë shtresën e dyfishtë disa herë (shih Figurën 11 -21A). Në shumë nga këto
proteina, një ose më shumë rajone transmembranore formohen nga α helikset që përmbajnë zinxhirë
anësore të aminoacideve hidrofobe ashtu edhe të atyre hidrofile. Këto aminoacide kanë tendencën të
organizohen në mënyrë që zinxhirët anësor hidrofob të bien në njërën anë të heliksit, ndërsa zinxhirët
hidrofil të përqëndrohen në anën tjetër. Në ambientin hidrofob të shtresës së dyfishtë lipidike, α-
helikset e këtij lloji rrinë pran njëri tjetrit në rreth, me zinxhirët anësor hidrofob të ekspozuar ndaj
lipideve të membranës, dhe zinxhirët anësor hidrofil që formojnë sipërfaqen e brendshme të një pori
hidrofil përgjatë shtresës së dyfishtë lipidike (Figura 11-24). Mënyra se si këto pore funksionojnë në
transportin përzgjedhës të molekulave të vogla të tretshme në ujë, do të shqyrtohet në Kapitullin 12.
Edhe pse α helksi është dukshëm forma më e zakonshme në të cilën një zinxhir polipeptid përshkon
shtresën e dyfishtë lipidike, zinxhiri polipeptid i disa proteinave transmembranore përshkon shtresën e
dyfishtë lipidike si një fletë β që harkohet në formë cilindri, duke formuar një strukturë në formë
fuçie, të quajtur fuçi β. Siç pritet, zinxhirët anësorë të aminoacideve që shtrihen nga brendësia e
fuçisë, dhe japin shtresën e brendshme të kanalit ujor, janë në të shumtën e rasteve hidrofile, ndërsa
ato në pjesën e jashtme të fuçisë, të cilët kontaktojnë pjesën qendrore hidrofobe të shtresës së dyfishtë
lipidike, janë vëtëm hidrofob. Shembulli më i qartë i strukturës së një fuçie β gjendet në proteinat
porine, të cilat krijojnë pore të mëdha të mbushura me ujë, në membranat mitokondriale dhe bakterore
(Figure 11-25). Mitokondriet dhe disa baktere janë të rrethuara nga një membranë e dyfishtë, ndërsa
porinet lejojnë kalimin e materieve ushqyese dhe joneve të vogla përmes membranave të tyre të
jashtme, duke penguar hyrjen e molekulave më të mëdha si antibiotikët dhe toksinat. Për dallim nga α
helikset, fuçitë β mund të formojnë vetëm kanale të gjera, sepse kanë një kufi për këndin e harkimit të
fletës β për formimin e fuçisë. Në këtë aspekt, një β fuçi është më pak e lakueshme se një koleksion α
heliksesh.
Figura 11-24 Një pore hidrofobe transmembranore
formohet nga disa α helikse. Në këtë shembull, pesë α helikse transmembranor formojnë një kanal të mbushur
me ujë përgjatë shtresës së dyfishtë lipidike. Zinxhirët anësor hidrofob të amino acideve (e gjelbër) në njërën
anë të helikseve kontaktojnë bishtat hidrofob hidrokarbonik, ndërsa zinxhirët anësor hidrofil (e kuqe) në krahun
tjetër të helikseve formojnë një pore të mbushur me ujë.

Figura 11-25 Proteinat porine formojnë kanale të
mbushura me ujë në membranën e jashtme të një bakteri (Rhodobacter capsulatus) . Proteina
konsiston në një β shtresë me 16 fije të harkuar rreth vetes për të formuar një kanal transmembranor të mbushur
me ujë, siç tregohet në këtë strukturë tre-dimensionale të determinuar me anë të kristalografisë me rreze X. Edhe
pse nuk është paraqitur në vizatim, tre proteina porine lidhen së bashku për të formuar një trimer, i cili ka tre
kanale të veçanta. (Nga S.W. Kowan, Curr. Opin. Struc.Biol.3,501-507, 1993. Me leje nga Elsevier).
Proteinat e membranës mund të treten në detergjentë dhe të purifikohen
Për të kuptuar plotësisht një proteinë, duhet të njohim strukturën e saj në detaje, dhe në rastin e
proteinave të membranës, kjo paraqet disa probleme të veçanta. Shumica e procedurave biokimike
janë krijuar për studimin e molekulave që treten në ujë apo në një tretës të thjeshtë; nga ana tjetër,
proteinat e membranës janë ndërtuar të veprojnë në ambient pjesërisht ujor dhe pjesërisht të yndyror,
prandaj ti nxjerrësh ato jashtë këtij ambienti dhe ti purifikosh duke i ruajtur në të njëjtën kohë dhe
strukturën e tyre themelore, nuk është një punë e lehtë.
Përpara se të studiohet në detaje një proteinë, ajo duhet të ndahet nga të gjitha proteinat tjera të
qelizës. Për shumë proteina të membranës, hapi i parë në këtë proces të ndarjes konsiston në tretjen e
membranës me agjentë që shkatërrojnë shtresën e dyfishtë lipidike duke prishur lidhjet hidrofobe.
Agjentët më të përhapur shkatërrues janë detergjentët (Filmimi 11.3). Këto janë molekula të vogla
lipidike amfipatike që kanë si pjesë hidrofobe ashtu dhe hidrofile (Figura 11-26). Detergjentët
dallojnë nga fosfolipidet e membranës sepse ata kanë vetëm një bisht hidrofob, dhe si rrjedhojë, sillen
krejt ndryshe. Për arsye se ata kanë vetëm një bisht, molekulat e detergjentëve kanë formë konike; në
ujë ato kanë tendencën të mblidhen dhe formojnë copëza të quajtura micele, në vend që të formojnë
një shtresën të dyfishtë siç bëjnë fosfolipidet, të cilat kanë formë më cilindrike.
Kur përzihen në masë të madhe me membranat, skajet hidrofobe të molekulave të detergjentëve
lidhen me zonën hidrofobe të proteinave transmembranore si dhe me bishtat hidrofob të molekulave
fosfolipidike, duke prishur shtresën e dyfishtë lipidike dhe duke ndarë proteinat nga shumica e

Figura 11-26 SDS dhe Triton X-100 janë dy detergjentë që
përdoren gjerësisht. Sodium dodecil sulfati (SDS) është një
detergjent i fortë jonik (ka një grup jonik në anën e tij
hidrofile), dhe Triton X-100 është një detergjent i bute jo-
jonik (ka një strukturë jo-jonike por të polarizuar në anën e tij
hidrofile). Ana hidrofobe e secilit detergjent është paraqitur
me të kaltër, ndërsa ajo hidrofile me të kuqe. Pjesa në kllapa e
Triton X-100 përsëritet rreth 8 herë. Detergjentët e fortë jonik
si SDS jo vetëm që zhvendosin molekulat lipidike nga
proteinat por ata edhe shpalosin proteinat (shih Panelin 4-5, p.
166).
fosfolipideve. Për arsye se ana tjetër e molekulës së detergjentit është hidrofile, ky asocim i sjellë
proteinat e membranës në tretësirë si komplekse proteinë-detergjent (Figura 11-27). Në të njëjtën
kohë, detergjenti tret fosfolipidet. Komplekset proteina-detergjent më pas mund të ndahen njëri-tjetri
dhe nga komplekset lipid-detergjent, me anë të një teknike si poliakrilamid-xhel elektroforezë (e
diskutuar në Kapitullin 4).
Figura 11-27
Proteinat e
membranës
mund të treten
nga një
detergjent i
butë të tillë si
Triton X-100. Molekulat e
detergjentit
(ngjyrë ari)
identifikohen si
monomere dhe
micele, forma në
të cilën
molekulat e
detergjentit kanë
tendencën të
përhapen në ujë.
Koka hidrofile e

detergjentit është skaji me një rreth. Detergjenti shkatërron shtresën e dyfishtë lipidike dhe i sjell proteinat në
tretësirë si kompleks proteinë-detergjent. Fosfolipidet në membranë gjithashtu treten nga detergjentët.
Struktura e plotë njihet relativisht për pak proteina të membranës
Për shumë vite, shumëçka ne dimim rreth strukturës së proteinave të membranës ishte mësuar me
mjete të tërthorta. Metoda standarde e drejtpërdrejtë për përcaktimin e strukturës është kristalografia
me rreze X (shih Figurën 4-48), por kjo kërkon një rreshtim të rregullt të kristaleve në molekulë, dhe
proteinat e membranës janë më të vështira për tu kristalizuar sesa proteinat e tretura që gjenden në
citosolin e qelizës apo lëngjet jashtëqelizore. Gjithsesi, me përparimet e fundit në kristalografinë me
rreze X, strukturat e shumë proteinave të membranës tashmë janë të dukshme në rezolucion të lartë,
përfshirë bakteriorodopsinën dhe qendrën e reaksionit fotosintetik – proteina të membranës të cilat
kanë role të rëndësishme në kapjen dhe përdorimin e energjisë nga dielli, aftësi kjo që do të studiohet
në Kapitullin 14. Struktura e bakteriorodopsinës tregoi për herë të parë se si α helikset e përshkojnë
shtresën e dyfishtë lipidike, dhe struktura e qendrës së reaksionit fotosintetik paraqiti në detaje se si
një grup i molekulave të ndryshme proteinike mund të mblidhen dhe të formojnë një kompleks
funksional në një membranë.
Bakteriorodopsina është një proteinë e vogël (rreth 250 amino acide) e gjendur gjerësisht në
membranën plazmatike të një arkeani të quajtur Halobacterium halobium, që jeton në kneta të
njelmëta. Bakteriorodopsina vepron si një proteinë membranore transportuese e cila pompon H+
(protone) jashtë bakterit. Pompimi kërkon energji, dhe bakteriorhodopsina e merr atë drejtpërdrejt nga
drita e diellit. Secila molekulë e bakteriorodopsinës përmban një molekulë të vetme jo-proteinike që
absorbon dritën – e quajtur retinal - e cila i jep proteinës (dhe bakterit) një ngjyrë të theksuar vjollcë.
Kjo molekulë e vogël hidrofobe është e lidhur në mënyrë kovalente me një nga shtatë α helikset
transmembranor të bakteriorhodopsinës dhe shtrihet në rrafshin e shtresës së dyfishtë lipidike, e
rrethuar tërësisht nga shtatë α helikset (Figura 11-28). Kur retinali absorbon një foton drite, ky i’a
ndryshon formën dhe kështu e detyron proteinën në shtresën e dyfishtë lipidike të pësojë disa
ndryshime. Këto ndryshime konsistojnë në transferimin e një H+ nga retinali te pjesa e jashtme e
bakterit: H+ lëvizë përgjatë shtresës së dyfishtë në një rrugë me zinxhirë anësor të aminoacideve të
polarizuara (shih Figurën 11-28) të vendosura në mënyrë strategjike. Retinali më tej rigjenerohet duke
marrë një H+ nga citosoli, dhe e kthen proteinën në formacionin e saj paraprak nga ku ajo do mund ta
përsërisë ciklin. Rezultati përfundimtar është lëvizja e një H+ nga ana e brendshme në anën e jashtme
të qelizës. Në prezencë të dritës, mijëra molekula të bakteriorodopsinës pompojnë H+ jashtë qelizës,
duke gjeneruar një përqëndrim të gradientit të H+ rreth membranës plazmatike. Qeliza e përdor këtë
gradient protonik për të grumbulluar energji dhe më pas ta shndërrojë atë në ATP, siç do ta
diskutojmë në detaj në Kapitullin 14. Bakteriorodopsina është një lloj proteine transportuese, që i
përket klasës së proteinave transmembranore që i lëvizin molekulat dhe jonet brenda dhe jashtë
qelizës (shih Figurën 11-20). Me transportues të tjerë do të njihemi në Kapitullin 12.

Figura 11-28 Bakteriorodopsina vepron si një pompë protonesh. Zinxhiri polipeptid përshkon shtresën e
dyfishtë lipidike si shtatë α helikse. Këtu janë paraqitur vendndodhja e retinalit (vjollce) dhe rruga e mundshme
e ndërmarrë nga protonet gjatë ciklit të pompimit të aktivizuar nga drita: zinxhirët anësor të aminoacideve të
polarizuara të përfshira në procesin e transferimit të H+ paraqiten me të kuqe, të verdhë dhe kaltër. Vini re që
rruga e ndërmarrë nga protonet (shigjetat e kuqe) i mundëson atyre të shmangin kontaktin me shtresën e dyfishtë
lipidike.Gjurmët e transferimit të protonit tegohen në Filmimin 11.4. Retinali gjithashtu përdoret për të dalluar
dritën në sytë tanë, ku është i bashkangjitur me një proteinë me strukturë shumë të ngjashme me
bakteriorodopsinën. (E adaptuar nga H. Luecke et al., Science 286:255-260. Me leje të AAAS).
Struktura e një qendre bakteriale të reaksionit fotosintetik është paraqitur në Figurën 11-29. Është një
kompleks i madh i përbërë nga katër nën-njesi proteinore. Tre janë proteina transmembranore; dy nga
këto (M dhe L) kanë disa α helikse që përshkojnë shtresën e dyfishtë lipidike, ndërsa tjetra (H) ka
vetëm një. Proteina e katërt (citokromi) është në kontakt me sipërfaqen e jashtme të membranës, e
lidhur me proteinat transmembranore. I gjithë kompleksi proteinor shërben si një makineri
molekulare, që merr energjinë e dritës të thithur nga klorofili dhe prodhon elektrone me energji të
lartë që nevojiten për reaksioni fotosintetike (që do të diskutohet në Kapitullin 14). Shumë proteina
membranore organizohen në komplekse të mëdha, dhe struktura e qendrës së reaksionit fotosintetik
zbulon shumë parime që vlejnë për mijëra proteina të tjera membranorë, strukturat e të cilave akoma
nuk njihen.

Figura 11-29 Qendra e reaksionit fotosintetik e bakterit Rhodopseudomonas v ir idis merr energji nga
drita e diellit. Struktura tre-dimensionale u identifikua përmes analizës së kristaleve të këtij kompleksi
proteinash transmembranore me anë të difraksionit me rreze X. Kompleksi konsiston në katër nën-njesi – L, Μ,
H, dhe një citokrom. L dhe Μ formojnë bërthamën e qendrës së reaksionit, dhe secila përmban pesë α helikse që
zgjaten përgjatë shtresës së dyfishtë lipidike. Të gjitha α helikset paraqiten si cilindra. Vendndodhja e grupeve të
ndryshme që bartin elektrone, të cilët janë të lidhur në mënyrë kovalente me nën-njesitë e proteinave, është e
paraqitur me ngjyrë të zezë,përveç çiftit të veçantë të molekulave të klorofilit të cilat reagojnë ndaj dritës, dhe
këtu janë paraqitur me ngjyrë gjelbër të errët. Kini parasysh që citokromi është i lidhur me sipërfaqen e
membranës vetëm nëpërmjet lidhjes së tij me nën-njesitë transmembranore (shih Figurën 11-21D).
Membrana plazmatike përforcohet nga korteksi qelizor
Një membranë qelizore është tepër e hollë dhe shume delikate. Do të duheshin rreth 10000 membrana
qelizore mbi njëra tjetrën që të arrihej trashësia e kësaj letre. Shumica e membranave qelizore
forcohen dhe mbështeten nga një strukturë proteinash, të lidhura me membranën me anë të proteinave
transmembranore. Në veçanti, forma e një qelize vetitë mekanike të membranës plazmatike të saj
përcaktohen nga rrjeti i proteinave fibroze, të quajtur korteksi qelizor, i cili është i ngjitur me
sipërfaqen citosolike të membranës.
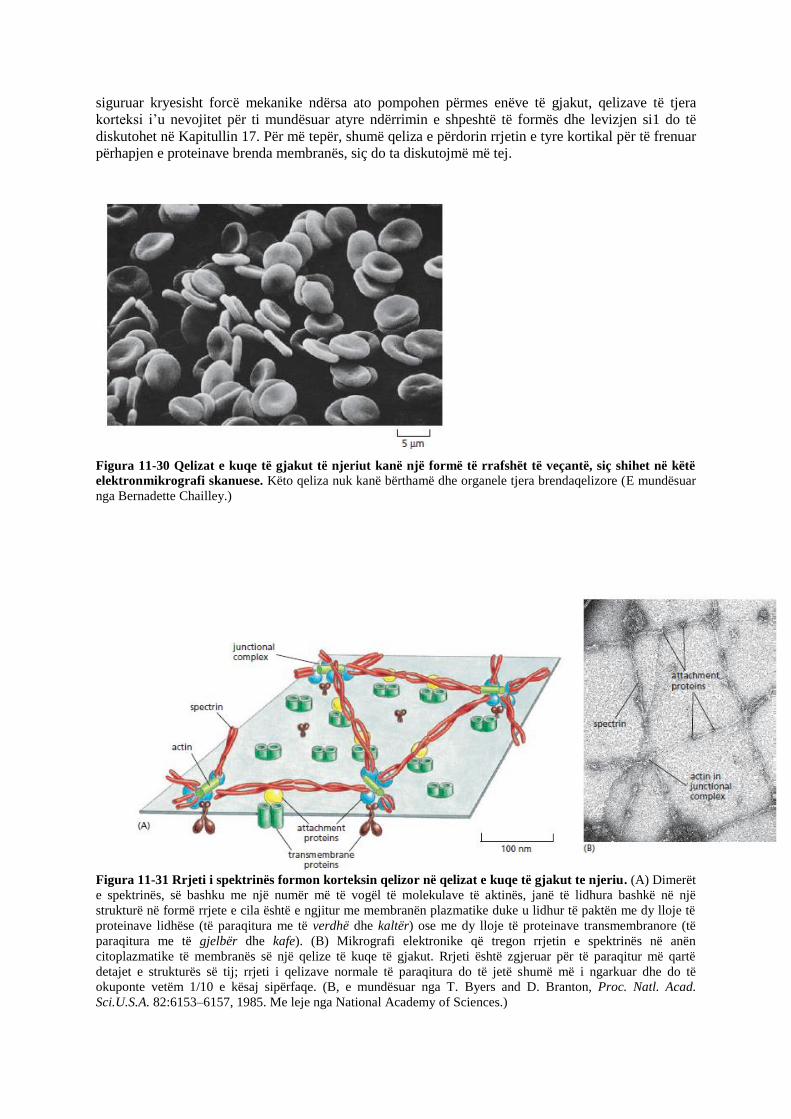
Korteksi i qelizave të kuqe të gjakut tek njeriu është relativisht i thjeshtë dhe me strukturë të rregullt,
si dhe është korteksi i egzaminuar më me hollësi deri më tani. Qelizat e kuqe të gjakut janë të vogla
dhe kanë një formë të veçantë të rrafshët (Figura 11-30). Përbërësi kryesor i korteksit të tyre është
proteina spektrinë, një shufër elastike e hollë dhe e gjatë rreth 100 nm. Ajo krijon një rrjet që i ofron
mbështetje membranës plazmatike dhe ruan formën e qelizës. Rrjeti i spektrinës është i lidhur me
membranën nëpërmjet proteinave lidhëse ndërqelizore që lidhin spektrinën me proteina specifike
transmembranore (Figura 11-31). Rëndësia e këtij rrjeti vërëhet te minjtë dhe te njerëzit që kanë
anomali gjenetike në strukturën e spektrinës. Këta individë janë anemikë: ata kanë më pak qeliza të
kuqe dhe i kanë në formë sferike, jo të rrafshëta.
Proteinat e ngjashme më spektrinën dhe me proteinat e bashkangjitura me të mund të gjenden në
korteksin e qelizës së shumicës së qelizave shtazore, por korteksi i këtyre qelizave është shumë më i
ndërlikuar se ai i qelizave të kuqe të gjakut. Derisa qelizat e kuqe kanë nevojë për korteksin për ti
siguruar kryesisht forcë mekanike ndërsa ato pompohen përmes enëve të gjakut, qelizave të tjera
korteksi i’u nevojitet për ti mundësuar atyre ndërrimin e shpeshtë të formës dhe levizjen si1 do të
diskutohet në Kapitullin 17. Për më tepër, shumë qeliza e përdorin rrjetin e tyre kortikal për të frenuar
përhapjen e proteinave brenda membranës, siç do ta diskutojmë më tej.
Figura 11-30 Qelizat e kuqe të gjakut të njeriut kanë një formë të rrafshët të veçantë, siç shihet në këtë
elektronmikrografi skanuese. Këto qeliza nuk kanë bërthamë dhe organele tjera brendaqelizore (E mundësuar
nga Bernadette Chailley.)
Figura 11-31 Rrjeti i spektrinës formon korteksin qelizor në qelizat e kuqe të gjakut te njeriu. (A) Dimerët
e spektrinës, së bashku me një numër më të vogël të molekulave të aktinës, janë të lidhura bashkë në një
strukturë në formë rrjete e cila është e ngjitur me membranën plazmatike duke u lidhur të paktën me dy lloje të
proteinave lidhëse (të paraqitura me të verdhë dhe kaltër) ose me dy lloje të proteinave transmembranore (të
paraqitura me të gjelbër dhe kafe). (B) Mikrografi elektronike që tregon rrjetin e spektrinës në anën
citoplazmatike të membranës së një qelize të kuqe të gjakut. Rrjeti është zgjeruar për të paraqitur më qartë
detajet e strukturës së tij; rrjeti i qelizave normale të paraqitura do të jetë shumë më i ngarkuar dhe do të
okuponte vetëm 1/10 e kësaj sipërfaqe. (B, e mundësuar nga T. Byers and D. Branton, Proc. Natl. Acad.
Sci.U.S.A. 82:6153–6157, 1985. Me leje nga National Academy of Sciences.)

Qelizat mund të kufizojnë lëvizjen e proteinave membranore
Për shkak se një membrana është një lëng dy-dimensional, shumë prej proteinave të saj, si lipidet e
saj, mund të lëvizin lirisht përmes shtresës së dyfishtë lipidike. Kjo lëvizje mund të demonstrohet
qartë nga bashkimi e një qelizë të miut me një qelizë humane për të formuar një qelizë hibride me
madhësi të dyfishtë dhe pastaj duke monitoruar shpërndarjen e proteinave të membranës plazmës të
miut dhe të njeriut. Në fillim proteinat e miut dhe ato të njeriut qëndrojnë të mbyllura në gjysmat e
tyre të qelizës, por pas rreth gjysëm ore, shohim që dy grupet e proteinave përzihen në mënyrë
uniforme mbi gjithë sipërfaqen e qelizave (Figura 11-32).
Pamja e një membrane si një det lipidik në të cilin gjithë proteinat notojnë lirisht është tepër e
thjeshtë. Qelizat kanë disa mënyra si ti kufizojnë disa proteina të membranës plazmatike në zona të
zaktuara brenda shtresës së dyfishtë, duke krijuar kështu regjione me funksione të specializuara ose
domene të membranës në sipërfaqen e qelizës apo organelit. Disa teknika për studimin e lëvizjeve të
proteinave membranore janë përshkruar në Si Duhet Studiuar, pp.382-383.
Siç ilustrohet në Figurën 11-33, proteinat e membranës plazmatike mund të lidhen me struktura
statike jashtë qelizës – për shembull, me molekulat në matriksin jashtëqelizore (e diskutuar ne
Kapitullin 20) – ose me strukturat relativisht të palëvizshme brenda qelizës, veçanërisht me korteksin
Figura 11-32 Formimi i qelizave hibride mi-njeri tregon që proteinat e membranës plazmatike
mund të lëvizin anash në shtresën e dyfishtë lipidike. Proteinat e miut dhe të njeriut fillimisht rrinë në
gjysmat e tyre të membranës plazmatike të qelizës hibride të sapo formuar, por ato më pas bashkohen brenda një
kohe të shkurtër. Për ti zbuluar këto proteina, dy antitrupa që lidhen me proteinat e njeriut the ato të miut,
etiketohet me ngjyra të ndryshme fluoreshente (rodaminë dhe fluoresceinë) dhe i shtohen qelizave. Dy antitrupat
fluoreshent mund të dallohen me mikroskop fluoreshent sepse fluoresceina është e gjelbër ndërsa rodamina
është e kuqe. (Bazuar në observimet e L.D. Frye and M. edidin, J. Cell Sci. 7:319–335, 1970. Me leje nga The
Company of Biologists Ltd.)

Figura 11-33 Lëvizja anësore e proteinave të membranës plazmatike mund të kufizohet në disa mënyra. Proteinat mund të lidhen me korteksin brenda qelizës (A), me molekula të matriksit jashtëqelizore jashtë
qelizës (B), apo me proteinat në sipërfaqen e një qelize tjetër (C). Barierat e shpërndarjes (të paraqitura si sbreza
të zi) kufizojnë proteinat brenda një domeni të caktuar të membranës (D).
e qelizës (shih Figurën 11-31). Së fundi, qelizat mund të krijojnë bariera që kufizojnë përbërësit e
caktuar membranor në një domen të membranës. Në qelizat epiteliale që veshin zorrën, për shembull,
është e rëndësishme që proteinat transportuese që marrin materiet ushqyese nga zorra, të qëndrojnë në
sipërfaqen apikale të qelizave (sipërfaqja në kontakt me përmbajtjen e zorrës) dhe që proteina të tjera
që bartin materiet e tretura jashtë qelizës epiteliale për te indet dhe sistemin e qarkullimit të gjakut, të
kufizohen në sipërfaqet bazale dhe anësore (Figura 11-34). Kjo shpërndarje asimetrike e proteinave
membranore ruhet nga një barierë e formuar përgjatë vijës ku qeliza është e ngjitur me qelizat e afërta
epiteliale me anë të një lidhje që quhet lidhje e ngushtë. Në këtë zonë, proteina të specializuara lidhëse
formojnë një rrip që vazhdon përreth qelizës ku ajo kontakton qelizat fqinje të saj, duke krijuar një
ngjitje në ndërmjet membranave të qelizave të afërta (shih Figurën 20-23). Proteinat membranore nuk
mund të përhapen përtej kësaj lidhje.
Figura 11-34 Një proteinë membranore kufizohet në një domen të caktuar të membranës plazmatike të
një qelize epiteliale në zorrë. Proteina A (në membranën apikale) dhe proteina Β (në membranat bazale dhe
anësore) mund të shpërndahen lateralisht në domenet e tyre të membranës, por parandalohen që të hyjnë në një
domen tjetër nga një lidhje qelizore e specializuar, e quajtur lidhje e ngushtë.

Sipërfaqja e qelizës është e mbuluar me karbohidrate
Më parë mësuam që shumë nga lipidet në shtresën e jashtme të membranës plazmatike kanë sheqerna
të bashkangjitur me lidhje kovalente. E njëjta vlen për shumicën e proteinave në membranën
plazmatike. Shumica e këtyre proteinave kanë zinxhirë të shkurtër sheqernash, të quajtura
oligosaharide, të bashkangjitura me to; ato quhen glikoproteina. Proteina të tjera membranore kanë
një ose më shumë zinxhirë të gjatë polisaharid të bashkangjitur; ata quhen proteoglikane. I tërë
karbohidrati në glikoproteina, proteoglikane dhe glikolipide gjendet në njërën anë të membranës, atë
jocitosolike, ku krijon një mbulesë sheqeri të quajtur shtresa karbohidrate (Figura 11-35).
Figura 11-35 Qelizat eukariote janë të mbuluara me sheqerna. Shtresa karbohidrate përbëhet nga zinxhirët
anësor oligosaharid të ngjitur në glikolipidet dhe glikoproteinat e membranës, dhe nga zinxhirët polisaharid në
proteoglikanet e membranës. Glikoproteinat dhe proteoglikanet që janë sekretuar nga qeliza dhe më pas janë
absorbuar sërish në sipërfaqen e saj mund të kontribuojnë në shtresën karbohidrate. Keni parasysh që tërë
karbohidrati ndodhet në sipërfaqen e jashtme (jo-citosolike) të membranës plazmatike.
Duke formuar një shtresë materiali që mbulon shtresën e dyfishtë lipidike, shrtesa karbohidrate
ndihmon në mbrojtjen nga dëmtimet mekanike dhe kimike. Pasi që oligosaharidet dhe polisaharidet
në shtresën karbohidrate thithin ujin, ato krijojnë në qelizë një sipërfaqe të rrëshkitshme. Kjo mbulojë
i ndihmon qelizat e lëvizshme si qelizat e bardha të gjakut të depërtojnë nëpër hapësira të ngushta, dhe
parandalon ngjitjen e qelizave te kuqe të gjakut me njëra-tjetrën apo në muret e enëve të gjakut.
Karbohidratet e sipërfaqes së qelizës bejnë më tepër sesa thjesht të mbrojnë dhe lubrifikojnë qelizën.
Ato luajnë një rol të rëndësishëm në njohjen qelizë-qelizë dhe bashkimin mes tyre. Pikërisht si shumë
proteina njohin dhe ngjiten në një vend të caktuar apo me një proteinë tjetër, disa proteina (të quajtura
lektine) janë të specializuara në njohjen e zinxhirëve specifik anësor oligosaharid dhe bashkohen me
ta. Zinxhirët anësore oligosaharid të glikoproteinave dhe glikolipideve, edhe pse të shkurtër
(zakonisht më pak se 15 njësi sheqeri), janë jashtëzakonisht të ndryshëm. Për dallim nga zinxhirët
polipeptid (proteina), në të cilët aminoacidet janë të gjitha të bashkuara në mënyrë lineare me anë të
lidhjeve identike peptide (shih Figurën 11-22), sheqernat mund të bashkohen në shumë mënyra,
shpesh duke formuar zinxhirë të degëzuar (shih Panelin 2-3, pp. 68-69). Edhe vetëm tre grupe

sheqernash mund të bashkohen bashkë në aq shumë kombinime lidhjesh kovalente sa që ata mund të
formojnë qindra trisaharide.
Në një organizëm shumëqelizor, shtresa karbohidrate shërben si një mbulojë dalluese, si një uniformë
e zyrtarit policor, që është karakteristike për qelizat e specializuara në një funksion specifik që njihet
nga qelizat e tjera me të cilat secila duhet të ndërveprojë. Oligosaharidet specifike në shtresën
karbohidrate janë të përfshira për shembull në njohjen e qelizës veze nga një spermatozoidi (e
diskutuar në Kapitullin 19). Ato gjithashtu përfshihen në përgjigjet tona ndaj infeksionit. Ne fazat e
para të infeksionit bakterial, për shembull, karbohidrati në sipërfaqen e qelizave të bardha të gjakut, të
quajtura neutrofile, njihet nga një lektinë që gjendet në qelizat që veshin enët e gjakut në zonën e
infektuar. Kjo njohje bënë që neutrofilet të ngjiten për enët e gjakut dhe pastaj migrojnë nga rrjedha e
gjakut në drejtim të indeve të infektuara, ku ato ndihmojnë në largimin e baktereve që kanë invaduar
(Figura 11-39).
Glikoproteinat janë anëtarë të rëndësishëm në familjen e proteinave membranore. Në kapitullin tjetër
do të ekzaminohen funksionet e ndërlikuara të proteinave transmembranore që transportojnë
molekulat nga jashtë-brenda qelizës dhe anasjelltas.
Figura 11-39 Njohja e karbohidratit të sipërfaqes së qelizës tek neutrofilet është faza e parë e migrimit të
tyra nga gjaku në drejtim të vendit të infeksionit. Proteinat transmembranore të specializuara (të quajtura
lektine) krijohen nga qelizat që veshin enët e gjakut (qelizat endoteliale) si përgjigje ndaj sinjaleve kimike që
vijnë nga vendi i infeksionit. Këto proteina njohin grupe të caktuara të sheqernave të bartura nga glikolipidet
dhe glikoproteinat në sipërfaqe të neutrofileve që qarkullojnë në gjak. Si rrjedhojë, neutrofilet ngjiten për murin
e enës së gjakut. Kjo ngjitje nuk është shumë e fortë, por çon te lidhja tjetër proteinë-proteinë, e cila është shumë
më e fortë (që nuk është paraqitur këtu) që ndihmon neutrofilet të migrojnë jashtë rrjedhës së gjakut, ndërmjet
qelizave endoteliale, në indin te vendi i infeksionit (Filmimi 11.6).





























