Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Nov. 1994, p. 6965-69730021-9193/94/$04.00+0Copyright X 1994, American Society for Microbiology
Vol. 176, No. 22
Isolation and Characterization of Point Mutations in theEscherichia coli grpE Heat Shock Gene
BIN WU,1* DEBBIE ANG,2 MARK SNAVELY,3 AND COSTA GEORGOPOULOS1'2Department of Cellular, Viral and Molecular Biology, University of Utah School of Medicine, Salt Lake City,
Utah 841321; Departement de Biochimie Medicale, Centre Medical Universitaire, 1211 Gene've 4,Switzerland2; and Amgen, Inc., Thousand Oaks, California 91320-17893
Received 17 May 1994/Accepted 28 August 1994
The Escherichia coli grpE gene (along with dnaK, dnaJ, groEL, and groES) was originally identified as one ofthe host factors required for phage X growth. The classical grpE280 mutation was the only grpE mutation thatresulted from the initial screen and shown to specifically block the initiation of A DNA replication. Here wereport the isolation of several new grpE missense mutations, again using phage X resistance as a selection. Allmutants fall into two groups based on their temperature-dependent phenotype for A growth. Members of thefirst group (I), including grpE17 and gjpE280, which was obtained again, are resistant to A growth at both 30and 42°C. Members of the second group (II), includinggrpE25,grpE66, grpE103, grpE13a,grpE57b, and grpE61,are sensitive to A growth at 30°C but resistant at 42°C. All mutations are recessive, since an E. coli grpE nullmutant strain carrying these mutant alleles on low-copy-number plasmids are sensitive to infection by theAgrpE' transducing phage. Both group I and group II mutants are temperature sensitive for E. coli growthabove 42°C. The nucleotide changes were identified by sequencing analyses and shown to be dispersedthroughout the latter 75% of the grpE coding region. Most of the amino acid changes occur at conservedresidues, as judged by sequence comparisons between E. coli and other bacterial and yeast GrpE homologs. Theisolation of these new mutations is the first step toward a structure-function analysis of the GrpE protein.
The Escherichia coli DnaK chaperone machine is composedof three member proteins: DnaK (the homolog of eukaryoticHsp7O), DnaJ (the homolog of eukaryotic Hsp4O), and GrpE.These proteins were initially identified about 20 years ago ashost factors required for phage X growth (13, 14). Subse-quently, these proteins were shown to be essential for E. coligrowth as well. DnaK, DnaJ, and GrpE belong to the heatshock protein family, whose expression is transiently inducedunder various stress conditions, such as heat, ethanol treat-ment, viral infection, and exposures to heavy metals (33).Earlier observations, combined with ample genetic and bio-chemical studies from different fields, have led to the appreci-ation of a more generalized chaperone function for the DnaK,DnaJ, and GrpE proteins (10, 16). The three proteins worktogether as a team to carry out a variety of cellular functions,including replication of phage X DNA, the E. coli chromosome,and the P1 and F plasmids (14), autoregulation of the heatshock response (11, 25, 45, 48), protection of other proteinsfrom misfolding or aggregation (23), reactivation of damagedproteins under stress conditions (41, 44, 54), facilitating pro-tein export by maintaining polypeptides in a translocation-competent conformation (38), and proteolysis (43).DnaK, DnaJ, and GrpE are highly conserved in nature.
DnaK is 50% identical at the amino acid sequence level to theeukaryotic Hsp7O proteins. It has two functional domains, theN-terminal 44-kDa ATPase domain and the C-terminal 24-kDa substrate-binding domain. A highly conserved DnaJ pro-tein family has also emerged during the last few years. DnaJhas four distinct domains: the highly conserved N-terminal Jregion, followed by a short Gly/Phe-rich region, a Cys-richregion of four CXXCXGXG motifs, and the less conservedC-terminal domain (3). In E. coli, dnaK and dnaJ are not
* Corresponding author. Phone: (801) 581-3848. Fax: (801) 581-3607.Electronic mail address: [email protected].
single-gene families, since at least one homolog of each hasbeen discovered recently (21, 42, 49). For grpE, the long-suspected homolog in eukaryotic cells has been identifiedrecently in mitochondria of the fungi Saccharomyces cerevisiaeand Neurospora crassa (19, 50) and shown to be an essentialgene whose product interacts with mitochondrial Hsp7O (2).The individual activities of DnaK, DnaJ, and GrpE, as well
as their combined action as a chaperone team, have been bestcharacterized in the well-defined X DNA replication systemreconstituted by using highly purified phage X- and E. coli-encoded replication proteins (30, 56). The XO protein binds tothe defined oriA sequences and attracts XP complexed with theDnaB helicase. DnaJ stabilizes this preprimosomal complexand tags it for recognition by DnaK. In the presence of GrpE,10-fold less DnaK protein is required for efficient in vitro XDNA replication (56).DnaK and DnaJ are bona fide chaperones which interact
with various target polypeptide substrates. GrpE acts only asan accessory factor for DnaK. DnaK has been shown to possessa very weak ATPase activity (approximately 1 ATP hydrolyzedper 10 min per monomer) (26, 57). The presence of both theDnaJ and GrpE cohorts synergistically stimulates DnaK'sATPase up to 50-fold, with DnaJ specifically accelerating thehydrolysis of bound ATP and GrpE accelerating the release ofbound nucleotides (26). The hydrolysis of ATP causes confor-mational changes in DnaK which may help release DnaK fromits bound substrates, thus allowing DnaK to recycle moreefficiently (27). Recently, Palleros et al. (36) suggested that inthe absence of the DnaJ and GrpE proteins, exchange ofADPto ATP bound to DnaK triggers the release of substrate fromDnaK.There exists both in vivo and in vitro evidence which
supports an intimate physical interaction between DnaK andGrpE: (i) extragenic suppressors of the grpE280 temperature-sensitive mutation which map in the dnaK gene can be isolated(20), (ii) the two proteins can be cross-linked by glutaraldehyde
6965
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

6966 WU ET AL.
TABLE 1. Bacterial strains, phages, and plasmids used
Strain, phage, or Genotype and description Reference or sourceplasmid
StrainB178 W3110 galE sup' 12C600 supE44 hsdR thi-1 thr-1 leuB6 lacYl tonA21 Our collectionDH5a recAl Bethesda Research
LaboratoriesDA259 C600 AgrpE::fl-Camr thr::TnlO 1BW452 DA259 (pBW401) This workBW496 DA259 (pWSK29/grpE25) This workBW506 DA259 (pWSK29/grpE280) This workBW509 DA259 (pWSK29/grpE103) This work
PhageXcI- XcI60; clear-plaque former Our collection434cl- Clear-plaque former Our collectionXgrpE+ X imm int xis aft cI857 grpE+ 39
PlasmidspDK124 pBluescript SK(+) carrying 600 bp of the grpE' open reading frame obtained by PCR This workpEMBL/grpE280 1.7-kbp grpE280 with flanking sequences cloned in pEMBL8(+) vector Our collectionpBW375 pEMBL/grpE+ This workpBW401 pWSK29 with 1.3-kbp grpE+ from pBW375 cloned into the Sall and EcoRI sites This workpGpl-2 Kanr 46pEMBL8(+) Ampr Tetr 6pWSK29 Ampr, low-copy-number vector 51
and can be coprecipitated in the presence of antibody againstone or the other (20, 35), and (iii) the GrpE protein can bepurified on the basis of its retention on a DnaK affinity column.The DnaK-GrpE complex is stable in the presence of 2 M KCI,suggesting that the interaction is very hydrophobic in nature.The complex, however, is efficiently destroyed in the presenceof ATP (55). It has been shown in vitro that GrpE probablybinds to a conserved, exposed loop (around codons 28 to 33)near the nucleotide-binding site of DnaK, because deletion ofthis loop inhibits formation of the DnaK-GrpE complex (5).This finding is consistent with the in vivo result, namely, thatthe mutant DnaK756 protein, which has a Gly-to-Asp changeat codon 32, has a lower affinity for GrpE (5, 20).GrpE is essential for bacterial growth at all temperatures;
however, it is dispensable in certain dnaK mutant backgrounds(1). Further analysis showed that it is not the lack of DnaKfunction per se that allows E. coli to tolerate the deletion of thegrpE+ gene. Rather, it is the presence of various extragenicsuppressors of the dnaK mutation that somehow compensatefor the deficiency in both DnaK and GrpE functions (1).To gain a better understanding of how GrpE carries out its
function(s), we isolated several new grpE missense mutations,using phage X resistance as a selection. The analyses of thesegrpE mutations and the biochemical characterization of thedefects of the mutant proteins should furnish insights into themechanism of action of the DnaK chaperone machine.
MATERIALS AND METHODS
Strains and media. The bacterial strains, phages, and plas-mids used in this study are listed in Table 1. L broth and L agarsupplemented with the appropriate antibiotics (when neces-sary) were used for the growth of the E. coli strains asdescribed by Miller (31).
Construction of pBW401. Two cloning steps were involvedin obtaining a low-copy-number plasmid carrying the wild-typegrpE gene. First, the 900-bp BamHI-EcoRI fragment frompEMBL8(+)/grpE280 containing part of the grpE open readingframe and its 3' flanking region was replaced by a 480-bp
BamHI-EcoRI fragment from plasmid pDK124 to obtainpBW375, resulting in a wild-type grpE gene cloned in thepEMBL8(+) vector. Second, the 1.3-kbp SalI-EcoRI fragmentfrom pBW375, containing the entire grpE+ open reading frameand its 5' flanking region, was moved into the low-copy-number plasmid pWSK29 (51). The resulting plasmid,pBW401, which has the grpE+ gene under the transcriptionalregulation of a T7 promoter and its own natural promoter, wasused in this study for mutagenesis purposes. The grpE+ openreading frame of pBW401 was confirmed to be wild type bysequencing and comparison with that of wild-type grpE' fromthe E. coli chromosome (data not shown).Plasmid mutagenesis with NH20H. Ten micrograms of
pBW401 plasmid DNA were treated with 40 mM hydroxy-lamine (NH2OH) in 50 mM sodium phosphate buffer (pH 6.0)at 72°C for 60 to 90 min, and the reaction was stopped withTNE buffer (100 mM NaCl, 1 mM EDTA, 10 mM Tris HCl[pH 8.0]). The mutagenized plasmid DNA was ethanol precip-itated and transformed into E. coli DH5at at 37°C on L-ampicillin (100 ,ug/ml) plates. The DNA (designatedpBW401*) extracted from the pooled transformants was usedfor the isolation of grpE mutations.Random PCR mutagenesis. One nanogram of XmnI-linear-
ized pBW401 DNA was used in each PCR as described byLeung et al. (24). The reverse primer, 5'-AACAGCTATGACCATG-3', and the -40 primer, 5'-G'TlTCCCAGTCACGAC-3', were used to amplify the 1.4-kbp fragment containingthe grpE gene and flanking DNA regions. After experimentingwith different conditions, we used a dGTP/dATP ratio of 5 andan otherwise normal PCR protocol to perform the mutagenesiswith a total of 35 cycles. The 1.4-kbp PCR DNA products weredigested with Sall and EcoRI to yield a -1.3-kbp fragment,which was cloned back into pBW401 treated with the samerestriction enzymes to replace the corresponding wild-typegrpE+ DNA fragment. The resulting plasmid pool was subse-quently screened for the presence of grpE mutations.DNA sequencing. The nucleotide changes caused by the
various grpE mutations were identified by using a double-stranded DNA cycle sequencing protocol from GIBCO BRL
J. BACTERIOL.
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
MUTATIONAL ANALYSIS OF THE grpE GENE 6967
as instructed by the manufacturer, with appropriate 32P-end-labeled primers complementary to both strands of the grpEDNA sequence.Phage burst size analysis. Bacteria were grown at 30°C in L
broth supplemented with ampicillin (100 ,ug/ml) and 0.2%maltose to an optical density at 595 nm of approximately 0.25.Phage XcI was added to the culture at a multiplicity ofinfection of 0.05, i.e., one phage per 20 bacteria. After incu-bation at room temperature for 30 min, the cells were pelleted,washed with 10 mM MgSO4 twice, and resuspended in thesame volume of L broth. The cell cultures were divided intotwo aliquots. One was kept at 30°C, and the other was kept at42°C. Fifteen-microliter aliquots were removed every 15 min,vortexed with 1 drop of chloroform, diluted, and spotted ontoan E. coli C600 lawn to determine the number of plaque-forming units (PFU) at each time point.
Induction and identification of the mutant grpE gene prod-ucts. The grpE null mutant bacterial strain (DA259), carryingvarious mutant grpE plasmids, was transformed with a secondplasmid, pGpl-2, that carries the T7 RNA polymerase gene 1cloned under the inducible control of the PL promoter (re-pressed by the thermolabile cI857 repressor) (46). Cells weregrown in L broth supplemented with ampicillin (100 ,ug/ml)and kanamycin (50 ,ug/ml) at 30°C to an optical density at 595nm of 0.4 and then shifted to 42°C for 30 to 45 min to inducesynthesis of the T7 RNA polymerase. Following induction ofthe T7 polymerase, the cell cultures were shifted back to 30°C.At various time points, aliquots were taken, pelleted, andresuspended in 100 RI of H20 per ml of cell culture, and 15-pulaliquots were loaded per well of a sodium dodecyl sulfate(SDS)-15% polyacrylamide gel (22).
RESULTS
Isolation of new grpE point mutations. Initially we at-tempted to directly isolate single-copy grpE mutations whichconfer temperature sensitivity for bacterial growth at 42°C orresistance mutations in the E. coli chromosome by P1-localizedmutagenesis (18), using a closely linked drug resistance marker(Camr), but were unsuccessful. Perhaps the open readingframe of grpE (0.6 kbp) is too small for efficient in vivomutagenesis and this is why originally only one grpE280mutation was isolated (40). Consistent with this possibility, amore recent attempt to isolate mutations by using a screen forthe high constitutive expression of heat shock proteins resultedin the isolation of many dnaK and dnaJ mutations but failed toyield any in grpE, which was also an expected target (53).Therefore, we set out to construct a wild-type gipE-carryingplasmid and perform plasmid mutagenesis in order to increasethe probability of obtaining mutations in the grpE gene.
Mutagenized plasmids, obtained by either NH2OH treat-ment or random PCR mutagenesis as described in Materialsand Methods, were transformed into the grpE null strainDA259 and incubated at 30°C overnight on L-ampicillin plates.Ampr transformants were pooled, and approximately 10' cellswere spread onto L-ampicillin plates preseeded with a mixtureof 4 x 107 XcI- and 4 x 107 434cI- phages. The plates wereincubated at either 37 or 42°C for 24 h. Colonies were streakedon L-ampicillin plates to obtain single colonies free of phage.They were then checked for phage resistance by cross-streak-ing against both XcI- and 434cI-. Plasmid DNA was extractedfrom those candidates that were resistant to both phages andretransformed into DA259. Lawns of the transformants were
spot tested with a dilution series of Xcl- and 434cI- to confirmtheir mutant phenotype.The rationale behind using both XcI- and 434cI- phages
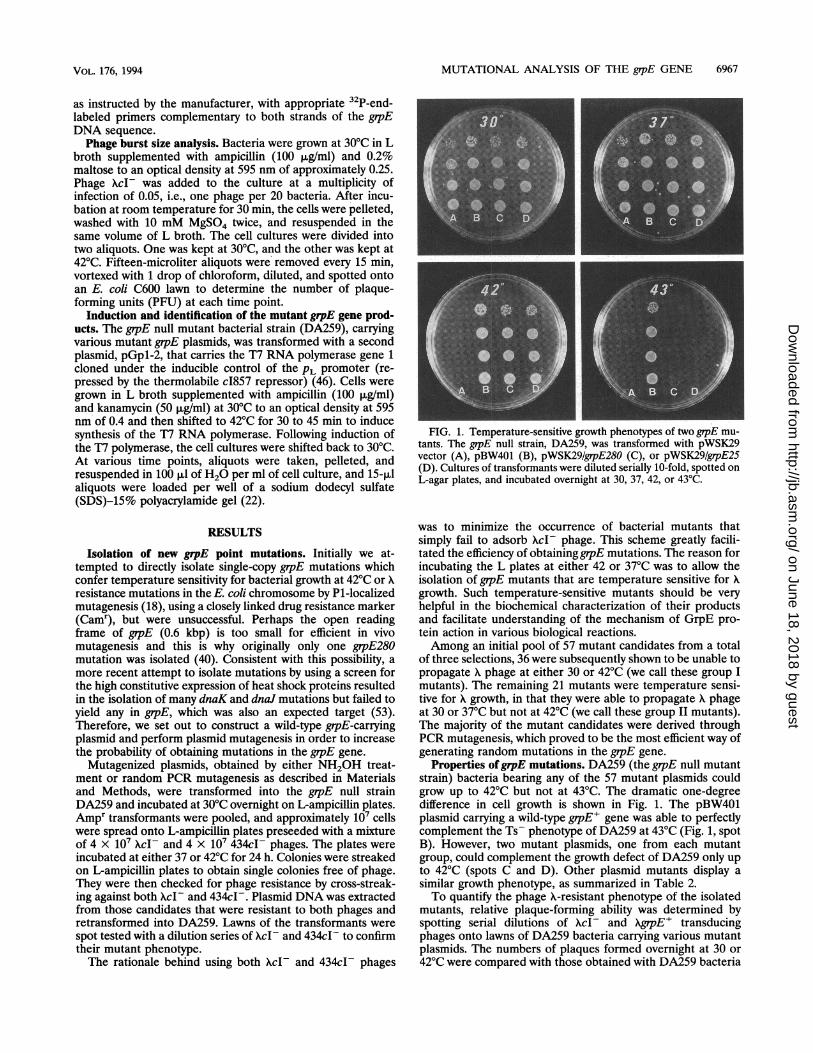
FIG. 1. Temperature-sensitive growth phenotypes of two grpE mu-tants. The grpE null strain, DA259, was transformed with pWSK29vector (A), pBW401 (B), pWSK29/grpE280 (C), or pWSK29/grpE25(D). Cultures of transformants were diluted serially 10-fold, spotted onL-agar plates, and incubated overnight at 30, 37, 42, or 43°C.
was to minimize the occurrence of bacterial mutants thatsimply fail to adsorb XcI- phage. This scheme greatly facili-tated the efficiency of obtaininggrpE mutations. The reason forincubating the L plates at either 42 or 37°C was to allow theisolation of grpE mutants that are temperature sensitive for Xgrowth. Such temperature-sensitive mutants should be veryhelpful in the biochemical characterization of their productsand facilitate understanding of the mechanism of GrpE pro-tein action in various biological reactions.Among an initial pool of 57 mutant candidates from a total
of three selections, 36 were subsequently shown to be unable topropagate X phage at either 30 or 42°C (we call these group Imutants). The remaining 21 mutants were temperature sensi-tive for X growth, in that they were able to propagate X phageat 30 or 37°C but not at 42°C (we call these group II mutants).The majority of the mutant candidates were derived throughPCR mutagenesis, which proved to be the most efficient way ofgenerating random mutations in the grpE gene.
Properties ofgrpE mutations. DA259 (the grpE null mutantstrain) bacteria bearing any of the 57 mutant plasmids couldgrow up to 42°C but not -at 43°C. The dramatic one-degreedifference in cell growth is shown in Fig. 1. The pBW401plasmid carrying a wild-type grpE' gene was able to perfectlycomplement the Ts- phenotype of DA259 at 43°C (Fig. 1, spotB). However, two mutant plasmids, one from each mutantgroup, could complement the growth defect of DA259 only upto 42°C (spots C and D). Other plasmid mutants display asimilar growth phenotype, as summarized in Table 2.To quantify the phage X-resistant phenotype of the isolated
mutants, relative plaque-forming ability was determined byspotting serial dilutions of Xcl- and XgrpE' transducingphages onto lawns of DA259 bacteria carrying various mutantplasmids. The numbers of plaques formed overnight at 30 or42°C were compared with those obtained with DA259 bacteria
VOL. 176, 1994
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
6968 WU ET AL. J. BACrERIOL.
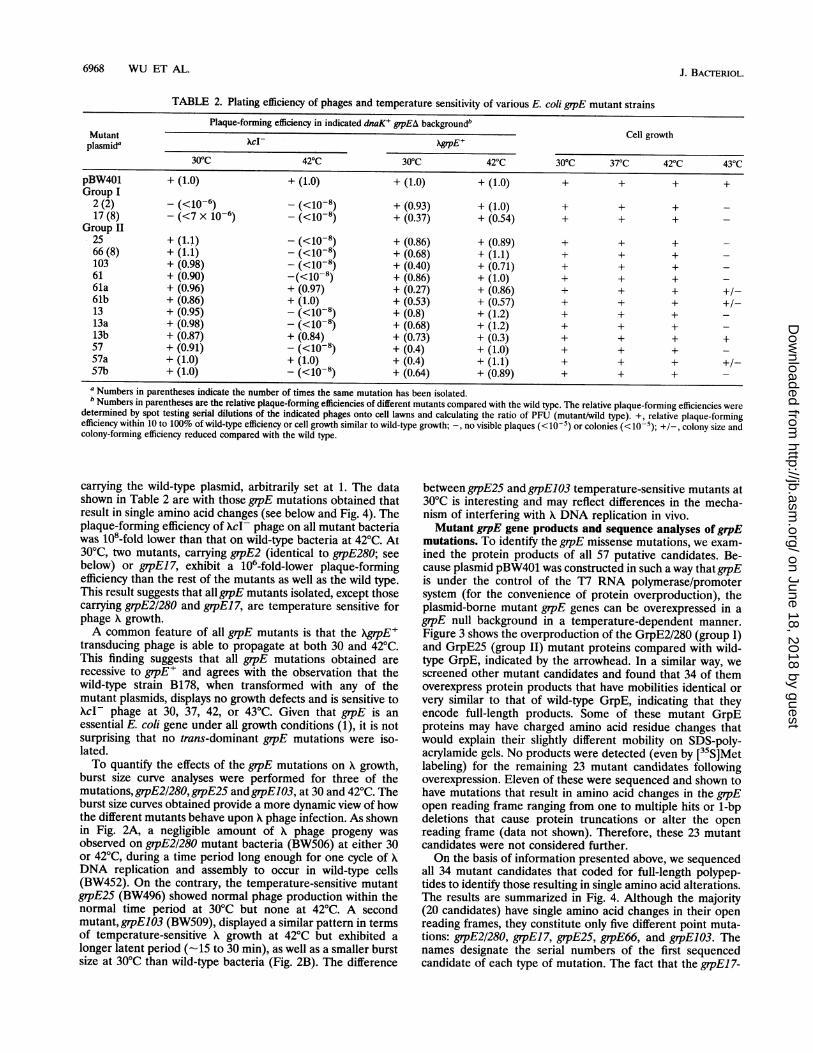
TABLE 2. Plating efficiency of phages and temperature sensitivity of various E. coli grpE mutant strainsPlaque-forming efficiency in indicated dnaK+ gipEA backgroundb
Mutant _ Cell growthplasmida ___ A.gpE+
300C 42°C 30°C 42°C 300C 37°C 42°C 43°CpBW401 + (1.0) + (1.0) + (1.0) + (1.0) + + + +Group I
2 (2) - (<10-6) - (<10-8) + (0.93) + (1.0) + + + -17 (8) - (<7 X 10-6) - (<10-8) + (0.37) + (0.54) + + + -
Group II25 + (1.1) - (<10-8) + (0.86) + (0.89) + + + -66 (8) + (1.1) - (<10-8) + (0.68) + (1.1) + + + -103 + (0.98) - (<10-8) + (0.40) + (0.71) + + + -61 + (0.90) -(<10-8) + (0.86) + (1.0) + + + -61a + (0.96) + (0.97) + (0.27) + (0.86) + + + +/-61b + (0.86) + (1.0) + (0.53) + (0.57) + + + +/-13 + (0.95) - (<10-8) + (0.8) + (1.2) + + + -13a + (0.98) - (<10-8) + (0.68) + (1.2) + + +13b + (0.87) + (0.84) + (0.73) + (0.3) + + + +57 + (0.91) _ (<10-8) + (0.4) + (1.0) + + + -57a + (1.0) + (1.0) + (0.4) + (1.1) + + + +/-57b + (1.0) - (<10-8) + (0.64) + (0.89) + + + -a Numbers in parentheses indicate the number of times the same mutation has been isolated.b Numbers in parentheses are the relative plaque-forming efficiencies of different mutants compared with the wild type. The relative plaque-forming efficiencies were
determined by spot testing serial dilutions of the indicated phages onto cell lawns and calculating the ratio of PFU (mutant/wild type). +, relative plaque-formingefficiency within 10 to 100% of wild-type efficiency or cell growth similar to wild-type growth; -, no visible plaques (<10-5) or colonies (<10-5); +/-, colony size andcolony-forming efficiency reduced compared with the wild type.
carrying the wild-type plasmid, arbitrarily set at 1. The datashown in Table 2 are with those grpE mutations obtained thatresult in single amino acid changes (see below and Fig. 4). Theplaque-forming efficiency of XcI- phage on all mutant bacteriawas 108-fold lower than that on wild-type bacteria at 42°C. At30°C, two mutants, carrying grpE2 (identical to grpE280; seebelow) or grpEJ7, exhibit a 106-fold-lower plaque-formingefficiency than the rest of the mutants as well as the wild type.This result suggests that all grpE mutants isolated, except thosecarrying grpE2/280 and grpE17, are temperature sensitive forphage X growth.A common feature of all grpE mutants is that the XgrpE+
transducing phage is able to propagate at both 30 and 42°C.This finding suggests that all grpE mutations obtained arerecessive to grpE' and agrees with the observation that thewild-type strain B178, when transformed with any of themutant plasmids, displays no growth defects and is sensitive toXcl- phage at 30, 37, 42, or 43°C. Given that grpE is anessential E. coli gene under all growth conditions (1), it is notsurprising that no trans-dominant grpE mutations were iso-lated.To quantify the effects of the grpE mutations on A growth,
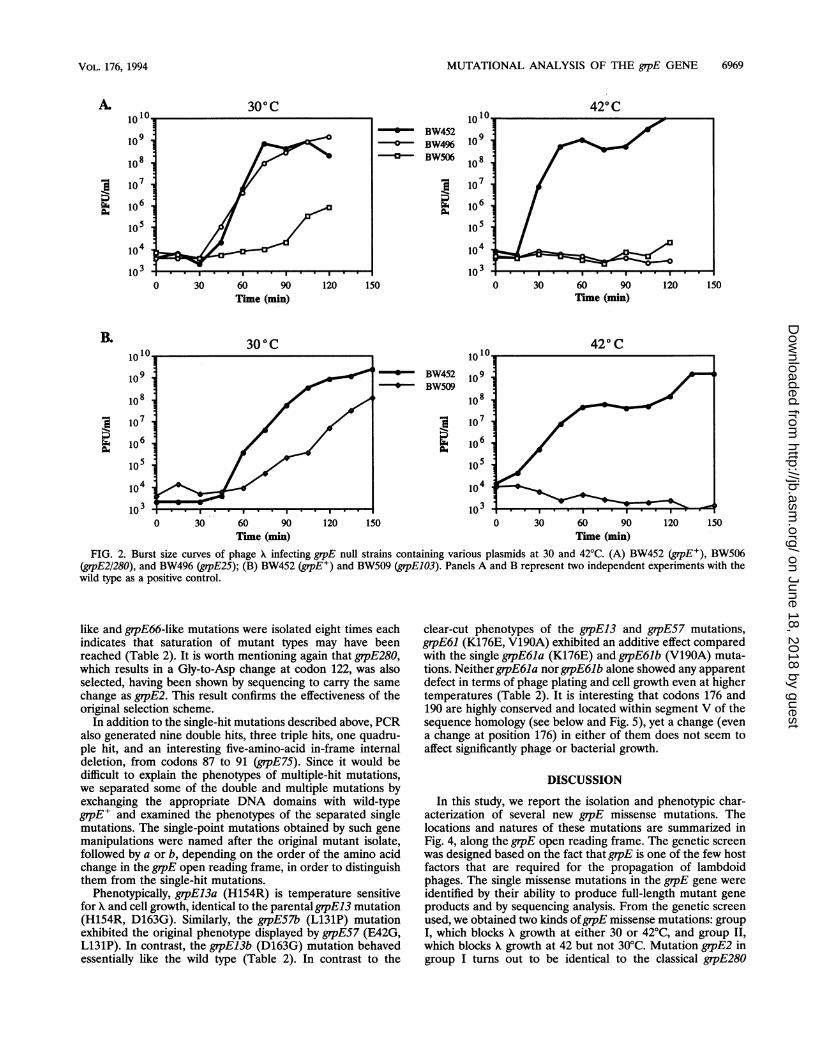
burst size curve analyses were performed for three of themutations, grpE2/280, grpE25 and grpEl03, at 30 and 42°C. Theburst size curves obtained provide a more dynamic view of howthe different mutants behave upon A phage infection. As shownin Fig. 2A, a negligible amount of X phage progeny wasobserved on grpE2/280 mutant bacteria (BW506) at either 30or 42°C, during a time period long enough for one cycle of XDNA replication and assembly to occur in wild-type cells(BW452). On the contrary, the temperature-sensitive mutantgrpE25 (BW496) showed normal phage production within thenormal time period at 30°C but none at 42°C. A secondmutant, grpE103 (BW509), displayed a similar pattern in termsof temperature-sensitive A growth at 42°C but exhibited alonger latent period (-15 to 30 min), as well as a smaller burstsize at 30°C than wild-type bacteria (Fig. 2B). The difference
between grpE25 and grpE103 temperature-sensitive mutants at30°C is interesting and may reflect differences in the mecha-nism of interfering with A DNA replication in vivo.Mutant grpE gene products and sequence analyses of grpE
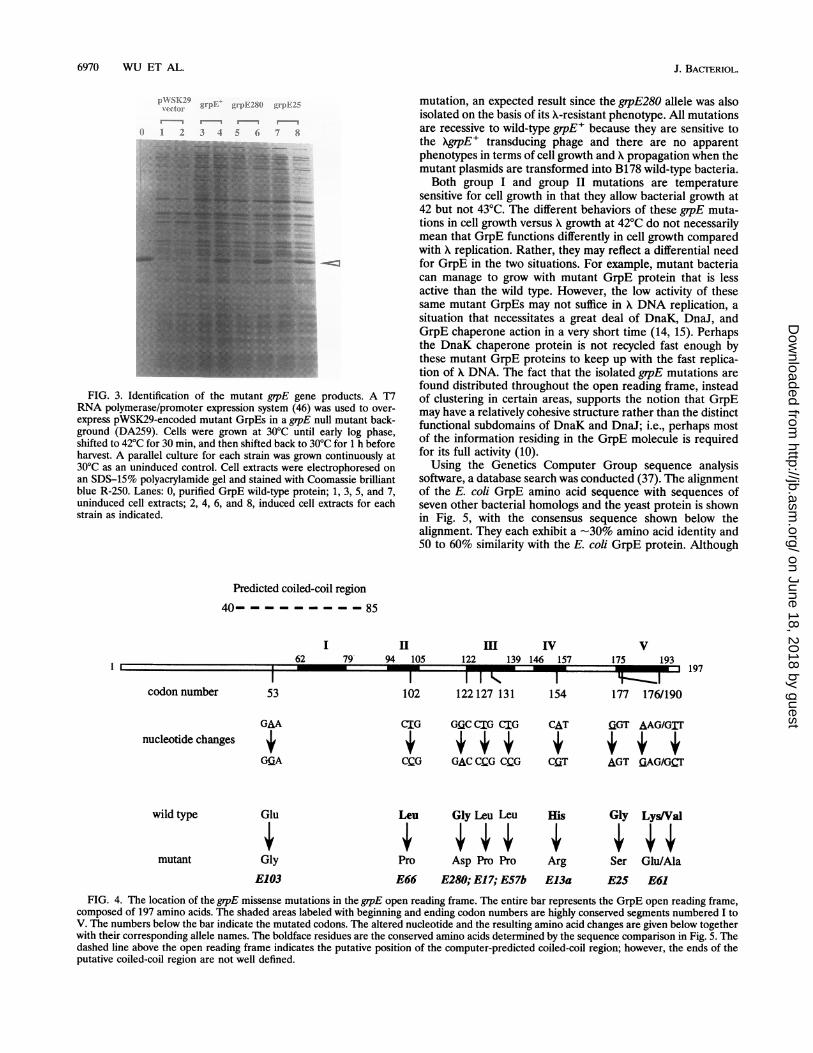
mutations. To identify the grpE missense mutations, we exam-ined the protein products of all 57 putative candidates. Be-cause plasmid pBW401 was constructed in such a way that grpEis under the control of the T7 RNA polymerase/promotersystem (for the convenience of protein overproduction), theplasmid-borne mutant grpE genes can be overexpressed in agrpE null background in a temperature-dependent manner.Figure 3 shows the overproduction of the GrpE2/280 (group I)and GrpE25 (group II) mutant proteins compared with wild-type GrpE, indicated by the arrowhead. In a similar way, wescreened other mutant candidates and found that 34 of themoverexpress protein products that have mobilities identical orvery similar to that of wild-type GrpE, indicating that theyencode full-length products. Some of these mutant GrpEproteins may have charged amino acid residue changes thatwould explain their slightly different mobility on SDS-poly-acrylamide gels. No products were detected (even by [35S]Metlabeling) for the remaining 23 mutant candidates followingoverexpression. Eleven of these were sequenced and shown tohave mutations that result in amino acid changes in the grpEopen reading frame ranging from one to multiple hits or 1-bpdeletions that cause protein truncations or alter the openreading frame (data not shown). Therefore, these 23 mutantcandidates were not considered further.On the basis of information presented above, we sequenced
all 34 mutant candidates that coded for full-length polypep-tides to identify those resulting in single amino acid alterations.The results are summarized in Fig. 4. Although the majority(20 candidates) have single amino acid changes in their openreading frames, they constitute only five different point muta-tions: grpE2/280, grpE17, grpE25, grpE66, and grpE103. Thenames designate the serial numbers of the first sequencedcandidate of each type of mutation. The fact that the grpE17-
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
MUTATIONAL ANALYSIS OF THE grpE GENE 6969
300C 420C
Time (min)
300 C
-0-a-
0 30 60 90 120 150Time (min)
BW452BW496BW506
BW452BW509
0 30 60 90Time (min)
FIG. 2. Burst size curves of phage X infecting grpE null strains containing various plasmids at 30 and 42°C. (A) BW452 (grpE+), BW506(grpE2/280), and BW496 (grpE25); (B) BW452 (gipE+) and BW509 (grpE103). Panels A and B represent two independent experiments with thewild type as a positive control.
like and grpE66-like mutations were isolated eight times eachindicates that saturation of mutant types may have beenreached (Table 2). It is worth mentioning again that grpE280,which results in a Gly-to-Asp change at codon 122, was alsoselected, having been shown by sequencing to carry the samechange as grpE2. This result confirms the effectiveness of theoriginal selection scheme.
In addition to the single-hit mutations described above, PCRalso generated nine double hits, three triple hits, one quadru-ple hit, and an interesting five-amino-acid in-frame internaldeletion, from codons 87 to 91 (grpE75). Since it would bedifficult to explain the phenotypes of multiple-hit mutations,we separated some of the double and multiple mutations byexchanging the appropriate DNA domains with wild-typegrpE' and examined the phenotypes of the separated singlemutations. The single-point mutations obtained by such genemanipulations were named after the original mutant isolate,followed by a or b, depending on the order of the amino acidchange in the grpE open reading frame, in order to distinguishthem from the single-hit mutations.
Phenotypically, grpEl3a (H154R) is temperature sensitivefor and cell growth, identical to the parental grpE13 mutation(H154R, D163G). Similarly, the grpE57b (L131P) mutationexhibited the original phenotype displayed by grpE57 (E42G,L131P). In contrast, the grpEl3b (D163G) mutation behavedessentially like the wild type (Table 2). In contrast to the
clear-cut phenotypes of the grpE13 and grpE57 mutations,grpE61 (K176E, V190A) exhibited an additive effect comparedwith the single grpE61a (K176E) and grpE61b (V19OA) muta-tions. NeithergrpE61a nor grpE61b alone showed any apparentdefect in terms of phage plating and cell growth even at highertemperatures (Table 2). It is interesting that codons 176 and190 are highly conserved and located within segment V of thesequence homology (see below and Fig. 5), yet a change (evena change at position 176) in either of them does not seem toaffect significantly phage or bacterial growth.
DISCUSSION
In this study, we report the isolation and phenotypic char-acterization of several new grpE missense mutations. Thelocations and natures of these mutations are summarized inFig. 4, along the grpE open reading frame. The genetic screenwas designed based on the fact that grpE is one of the few hostfactors that are required for the propagation of lambdoidphages. The single missense mutations in the grpE gene wereidentified by their ability to produce full-length mutant geneproducts and by sequencing analysis. From the genetic screen
used, we obtained two kinds ofgrpE missense mutations: groupI, which blocks A growth at either 30 or 42°C, and group II,which blocks X growth at 42 but not 30°C. Mutation grpE2 ingroup I turns out to be identical to the classical grpE280
B.
0 30 60 90 120 150Time (min)
120 150
VOL. 176, 1994
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
6970 WU ET AL.
vector grpE+ grpE280 grpE25
0 1 2 3 4 5 6 7 8
FIG. 3. Identification of the mutant grpE gene products. A T7RNA polymerase/promoter expression system (46) was used to over-express pWSK29-encoded mutant GrpEs in a grpE null mutant back-ground (DA259). Cells were grown at 30°C until early log phase,shifted to 42°C for 30 min, and then shifted back to 30°C for 1 h beforeharvest. A parallel culture for each strain was grown continuously at30°C as an uninduced control. Cell extracts were electrophoresed onan SDS-15% polyacrylamide gel and stained with Coomassie brilliantblue R-250. Lanes: 0, purified GrpE wild-type protein; 1, 3, 5, and 7,uninduced cell extracts; 2, 4, 6, and 8, induced cell extracts for eachstrain as indicated.
mutation, an expected result since the grpE280 allele was alsoisolated on the basis of its X-resistant phenotype. All mutationsare recessive to wild-type grpE+ because they are sensitive tothe XgrpE+ transducing phage and there are no apparentphenotypes in terms of cell growth and A propagation when themutant plasmids are transformed into B178 wild-type bacteria.Both group I and group II mutations are temperature
sensitive for cell growth in that they allow bacterial growth at42 but not 43°C. The different behaviors of these grpE muta-tions in cell growth versus X growth at 42°C do not necessarilymean that GrpE functions differently in cell growth comparedwith X replication. Rather, they may reflect a differential needfor GrpE in the two situations. For example, mutant bacteriacan manage to grow with mutant GrpE protein that is lessactive than the wild type. However, the low activity of thesesame mutant GrpEs may not suffice in X DNA replication, asituation that necessitates a great deal of DnaK, DnaJ, andGrpE chaperone action in a very short time (14, 15). Perhapsthe DnaK chaperone protein is not recycled fast enough bythese mutant GrpE proteins to keep up with the fast replica-tion of X DNA. The fact that the isolated grpE mutations arefound distributed throughout the open reading frame, insteadof clustering in certain areas, supports the notion that GrpEmay have a relatively cohesive structure rather than the distinctfunctional subdomains of DnaK and DnaJ; i.e., perhaps mostof the information residing in the GrpE molecule is requiredfor its full activity (10).
Using the Genetics Computer Group sequence analysissoftware, a database search was conducted (37). The alignmentof the E. coli GrpE amino acid sequence with sequences ofseven other bacterial homologs and the yeast protein is shownin Fig. 5, with the consensus sequence shown below thealignment. They each exhibit a -30% amino acid identity and50 to 60% similarity with the E. coli GrpE protein. Although
Predicted coiled-coil region
40- - - - - - - - - 85
I II62 79 94 105
HII IV122 139 146 157
I I I k 1102 122127 131 154 177 176/19053
nucleotide changes
wild type
mutant
GMA
G!IA
Glu
Gly
E103
CIG G(IC CIG CrG CAT
CCG GAC CCG CCG CQT
IZGT AAG/GIT
AGT £iAG/GCT
Leu Gly Leu Leu Eis Gl LysVal
Pro Asp Pro Pro Arg Ser Glu/AlaE66 E280; E17; E57b E13a E25 E61
FIG. 4. The location of the grpE missense mutations in the grpE open reading frame. The entire bar represents the GrpE open reading frame,composed of 197 amino acids. The shaded areas labeled with beginning and ending codon numbers are highly conserved segments numbered I toV. The numbers below the bar indicate the mutated codons. The altered nucleotide and the resulting amino acid changes are given below togetherwith their corresponding allele names. The boldface residues are the conserved amino acids determined by the sequence comparison in Fig. 5. Thedashed line above the open reading frame indicates the putative position of the computer-predicted coiled-coil region; however, the ends of theputative coiled-coil region are not well defined.
codon number
V175 193
197I I sI I I I I I I.P. I
J. BAcrERIOL.
I
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

MUTATIONAL ANALYSIS OF THE grpE GENE 6971
1
* - - ... .....---.. --....
*-.....---- ...X......
.......... ...mqekdskdmtnkdesvek ntestveetn
.... mrafsa atvrattrks
.msskeqktp egqapeeiim dqheeieave
.mseekqtve qneteeqeii eeqaaadeqq
.... . . . . . . . . . . . . . . . . etkrikptnk.......... .mseetkeeiknekvdeevte.................... mtetpntsse
vtmedeetia sqeeievegn seesskeeesikqniddsve qaeeskghlq deaieetsde......... mteetpgfeek pdvpsgatpdfipmaprtpf vtpsftknvg smrrrnrfysd
70peasaeqvdp rdekvanle.eetnes..el lqnqinel..iikikiakke nlnlvnsdkkelteealedi veeeineldeeiqtsepssd n.elqtlqqennseisdenl seenlklkdenvieei..dp kdqkinel..daepqaasee gaapagdaseeakseesken nedlteeqse
Consensus ---------- ---------- ---------- ---------- ---------- ---------- ----------
71 _ _I____ I 14CEco a. ......eaqtrerd gi 4teli ekah alek fi 1pvi s 1ev..Bsu .q.4 ...n fe s ri easq rsqn iv 1 plv..Burg iae.... .. snlkdl fe ke4 dnfv danet imi vf n...Lacla aq a ....n qrgn lqlv;rsqd lakki sa av..Ctra n .... .. kneknd e Irq temgk4raen all lgf..Cace ne. .... e...d ek f eiktdacr vi ta n t..Staph .a...... de... e e einktdqacr vi ti si..Strep na aqldq vrtalnerta q vavkevavan 1s1 v dvYeast ik sqlsa kt)4aselkd fr k qkakclalqk falcits 4 lnafk
Consensus ---L------ ---t------ -LLR--AX-- MY--R---3 - -----Y----
EcoBsuBurgLacla
CtraCaceStaphStrepYeast
Consensus
EcoBsuBurgLaclaCtraCaceStaphStrepYeast
---D-L---D -LaRA-----* (grPE66 P)
141 1 III IV I 210..adkan4 samv lks f eviaet .vesdd vapgnVlgii 175
..eadne.qt ksll hr e.k veaieav .mqaeden ygsniVve4 167
..sskks f dnll eneils lkkfgen ie.ekeg lknpelvevy 157
..egltd. vk... qe e evsy. .es tvpadde hpadsIvq 159
..asqts4v knwa f lq d aeyssk ie.ettd ipegtIle 156
..egsae. i k... vk s .8 eeipse . .mhiedeg ygeneVve..egdde.tf kslq he n .k evikte .vqddnpd fesgeltq 188
....... .....gelv ae ttva. qqfgke .mhsyapd vtettcva 167
eedlqks s rdr ntl.r ekldpl .felpqpd kepgtVfhvq 204
- ---- --Mv ---L---L-- 9 --F--- V---------L*(grpE280 D) *(gz.pEl3a R)
__V I
akaka ....... .........
lw v p8 q ....................... ..........
1a qsk..................
.................. ..........
ak fptkgnndsn eeke.
r s an ..........
1ps n. ..........
iigt pa ise pqpgaqtvkp aedaaeaqds1tn ipa kgeene..
268.......... .........197
.......... .........187
.......... .........179
.......... .........179
.......... .........190
.......... .........200
.......... .........208
sgaeddagtk esggpdeg 225.......... .........228
Consensus QKGY---DR- -R---V-V-- ---------- ---------- ---------- --------
* (grpE25 S) * (grpE61b A)* (grpE61a E)
FIG. 5. Comparison of the amino acid sequences of GrpE proteins (or homologs) from E. coli (Eco) (28), B. subtilis (Bsu) (52), B. burgdorferi(Burg) (47), Lactococcus lactis (Lacla) (7), C. trachomatis (Ctra) (8), Clostridium acetobutylicum (Cace) (32), Staphylococcus aureus (Staph) (34),Streptomyces coelicolor A3(2) (Strep) (4), and the yeast S. cerevisiae (Yeast) (19). The number at the end of each line indicates the position of thelast amino acid shown. Highlighted residues are those that are conserved in at least seven of nine sequences. The asterisks under the consensussequence indicate the positions of the most conserved missense mutations discussed in this work (in parentheses). The analysis was performed withthe Genetics Computer Group sequence analysis program (9, 17).
sequence homology exists almost throughout the entire lengthof the protein, there are five distinct, highly conserved seg-ments, which we have numbered I to V (Fig. 4 and 5). SegmentI spans codons 62 to 79, segment II spans codons 94 to 105,segment III spans codons 122 to 139, segment IV spans codons
146 to 157, and segment V spans codons 175 to 193, the codonnumbers referring to those of the E. coli GrpE protein. Whenthe amino acid changes caused by the grpE missense mutationswere compared with the consensus sequence, five of them,grpE66 (L102P), grpE2I280 (G122D), grpEl3a (H154R),
EcoBsuBurgLaclaCtraCaceStaphStrepYeast
484530402958664166
108102909890
119123106136
VOL. 176, 1994
0
--------D-
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

6972 WU ET AL.
grpE25 (G177S), and grpE61 composed of 61a (K176E) and61b (V19OA), were changes occurring at very conserved resi-dues (asterisks in Fig. 5) located in the highly conservedsegments II, III, IV, and V, respectively. Although the otherthree mutations, grpE103 (E53G), grpEl 7 (L127P), andgrpE57b (L131P), do not result in changes of conserved aminoacid residues, nevertheless they are all located next to highlyconserved ones, Glu at codon 54 for grpEl03, Val at codon 126for grpE17, and Leu at codon 130 for grpE57b. The overallgood correlation between the locations of the grpE missensemutations obtained in this study and the conserved residuesrevealed through sequence homology comparisons stronglysuggests that these conserved amino acids are important forGrpE function.
Because there is no available information on GrpE's sec-ondary structure, we entered the GrpE amino acid sequenceinto a computer program that predicts the probability ofcoiled-coil secondary structure based on the context of theamino acid sequence (29). All four bacterial protein sequencesentered, namely, those from E. coli, Bacillus subtilis, Borreliaburgdorferi, and Chlamydia trachomatis, were predicted topossess a possible coiled-coil region between E. coli GrpEcodons 40 and 85, which span the conserved segment I (Fig. 4).One of the missense mutations, grpE103, falls in the middle ofthis predicted coiled-coil region. When the Glu residue ofcodon 53 was substituted with that of Gly found in grpEl03, theresulting sequence showed a 30% reduced probability offorming the coiled-coil structure compared with wild-typeGrpE. Coiled-coil regions have been postulated to facilitateprotein-protein interactions. Previously, GrpE had been shownto form a salt-resistant but ATP-sensitive complex with DnaKand to be a dimer by glutaraldehyde cross-linking (35). It willbe interesting to determine whether the mutant GrpE103protein is deficient in DnaK-GrpE interaction and/or dimer-ization.Another interesting feature of the distribution of the grpE
mutations is that the only two mutations (grpE2/280 andgrpE17) that block X growth at all temperatures are locatedonly within the conserved segment III, while all other muta-tions are temperature sensitive for X growth and are spreadthroughout the open reading frame, either before or aftergrpE280. This result may imply that codons around amino acid122 in E. coli are very crucial for GrpE's function. However,the significance of such mutant distributions must await the invitro biochemical characterization of the defects of eachmutant protein.
ACKNOWLEDGMENTS
This work was supported by grants from the NIH (AI21029), theFNS (31-31129.91), and the Canton of Geneva.We thank Dorothy Lowry for assisting us with the coiled-coil
secondary structure analysis.
REFERENCES1. Ang, D., and C. Georgopoulos. 1989. The heat-shock-regulated
grpE gene of Escherichia coli is required for bacterial growth at alltemperatures but is dispensable in certain mutant backgrounds. J.Bacteriol. 171:2748-2755.
2. Bolliger, L., 0. Deloche, B. S. Glick, C. Georgopoulos, P. Jeno, N.Kronidou, M. Horst, N. Morishima, and G. Schatz. 1994. Amitochondrial homolog of bacterial GrpE interacts with mitochon-drial hsp7O and is essential for viability. EMBO J. 13:1998-2006.
3. Bork, P., C. Sander, A. Valencia, and B. Bukau. 1992. A module ofthe DnaJ heat shock proteins found in malaria parasites. TrendsBiochem. Sci. 17:129.
4. Bucca, G., C. P. Smith, M. Alberti, G. Seidita, R. Passantino, andA. M. Puglia. 1993. Cloning and sequencing of the dnaK region of
Streptomyces coelicolor A3(2). Gene 130:141-144.5. Buchberger, A., H. Schroder, M. Buttner, A. Valencia, and B.
Bukau. 1994. A conserved loop in the ATPase domain of theDnaK chaperone is essential for stable binding of GrpE. Struct.Biol. 1:95-101.
6. Dente, L., G. Cesareni, and R. Cortese. 1983. pEMBL: a newfamily of single-stranded plasmids. Nucleic Acids Res. 11:1645-1655.
7. Eaton, T. J., C. A. Shearman, and M. J. Gasson. 1993. Cloning andsequence analysis of the dnaK gene region of Lactococcus lactissubsp. lactis. J. Gen. Microbiol. 139:3253-3263.
8. Engel, J. N., J. Pollack, E. Perara, and D. Ganem. 1990. Heatshock response of murine Chlamydia trachomatis. J. Bacteriol.172:6959-6972.
9. Feng, D. F., and R. F. Doolittle. 1987. Progressive sequencealignment as a prerequisite to correct phylogenetic trees. J. Mol.Evol. 35:351-360.
10. Frydman, J., and F.-U. Harti. 1994. Molecular chaperone func-tions of hsp70 and hsp60 in protein folding, p. 251-283. In R. I.Morimoto, A. Tissieres, and C. Georgopoulos (ed.), The biologyof heat shock proteins and molecular chaperones. Cold SpringHarbor Laboratory Press, Cold Spring Harbor, N.Y.
11. Gamer, J., H. Bujard, and B. Bukau. 1992. Physical interactionbetween heat shock proteins DnaK, DnaJ, and GrpE and thebacterial heat shock transcription factor &r32. Cell 69:833-842.
12. Georgopoulos, C. 1971. Bacterial mutants in which the gene N ofphage lambda is blocked have an altered RNA polymerase. Proc.Natl. Acad. Sci. USA 68:2977-2981.
13. Georgopoulos, C. 1992. The emergence of the chaperone machines.Trends Biochem. Sci. 17:295-299.
14. Georgopoulos, C., D. Ang, K. Liberek, and M. Zylicz. 1990.Properties of the Escherichia coli heat shock proteins and theirrole in phage A growth, p. 191-221. In R. I. Morimoto, A.Tissieres, and C. Georgopoulos (ed.), Stress Proteins in biologyand medicine. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, N.Y.
15. Georgopoulos, C., K. Liberek, M. Zylicz, and D. Ang. 1994.Properties of the heat shock proteins of Escherichia coli and theautoregulation of the heat shock response, p. 209-249. In R. I.Morimoto, A. Tissieres, and C. Georgopoulos (ed.), The biologyof heat shock proteins and molecular chaperones. Cold SpringHarbor Laboratory Press, Cold Spring Harbor, N.Y.
16. Georgopoulos, C., and W. J. Welch. 1993. Role of the major heatshock proteins as molecular chaperones. Annu. Rev. Cell Biol.9:601-634.
17. Higgins, D. G., and P. M. Sharp. 1989. Fast and sensitive multiplesequence alignment on a microcomputer. CABIOS 5:151-153.
18. Hong, J., and B. Ames. 1971. Localized mutagenesis of any specificsmall region of the bacterial chromosome. Proc. Natl. Acad. Sci.USA 68:3158-3162.
19. Ikeda, E., S. Yoshida, H. Mitsuzawa, I. Uno, and A. Toh-e. 1994.YGE1 is a yeast homologue of Escherichia coli grpE and is requiredfor maintenance of mitochondrial functions. FEBS Lett. 339:265-268.
20. Johnson, C., G. N. Chandrasekhar, and C. Georgopoulos. 1989.Escherichia coli DnaK and GrpE heat shock proteins interact bothin vivo and in vitro. J. Bacteriol. 171:1590-1596.
21. Kawula, T. H., and M. J. Lelivelt. 1994. Mutations in a geneencoding a new hsp7o suppress rapid DNA inversion and bglactivation, but notproU derepression, in hns-I mutant Escherichiacoli. J. Bacteriol. 176:610-619.
22. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
23. Langer, T., C. Lu, H. Echols, J. Flanagan, M. K. Hayer, and F.-U.Hartl. 1992. Successive action of DnaK, DnaJ and GroEL alongthe pathway of chaperone-mediated protein folding. Nature (Lon-don) 356:683-689.
24. Leung, D. W., E. Chen, and D. V. Goeddel. 1989. A method forrandom mutagenesis of a defined DNA segment using a modifiedpolymerase chain reaction. Technique 1:11-15.
25. Liberek, K., and C. Georgopoulos. 1993. Autoregulation of theEscherichia coli heat shock response by the DnaK and DnaJ heat
J. BACFERIOL.
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

MUTATIONAL ANALYSIS OF THE grpE GENE 6973
shock proteins. Proc. Natl. Acad. Sci. USA 90:11019-11023.26. Liberek, K., J. Marszalek, D. Ang, C. Georgopoulos, and M.
Zylicz. 1991. Eschenichia coli dnaJ and grpE heat shock proteinsjointly stimulate ATPase activity of DnaK Proc. Natl. Acad. Sci.USA 88:2874-2878.
27. Liberek, K., D. Skowyra, M. Zylicz, C. Johnson, and C. Georgo-poulos. 1991. The Escherichia coli DnaK chaperone protein, theHsp7O eukaryotic equivalent, changes its conformation upon ATPhydrolysis, thus triggering its dissociation from a bound targetprotein. J. Biol. Chem. 266:14491-14496.
28. Lipinska, B., J. King, D. Ang, and C. Georgopoulos. 1988. Se-quence analysis and transcriptional regulation of the Escherichiacoli grpE gene encoding a heat shock protein. Nucleic Acids Res.16:7545-7562.
29. Lupas, A., M. Van Dyke, and J. Stock. 1991. Predicting coiled-coilsfrom protein sequences. Science 252:1162-1164.
30. Mensa-Wilmot, K., R. Seaby, R, C. Alfano, M. S. Wold, B. Gomes,and R. MacMacken. 1989. Reconstitution of a nine-protein systemthat initiates phage X DNA replication. J. Biol. Chem. 264:2853-2861.
31. Miller, J. H. 1972. Experiments in molecular genetics. Cold SpringHarbor Laboratory Press, Cold Spring Harbor, N.Y.
32. Narberhaus, F., K. Giebeler, and H. Bahl. 1992. Molecularcharacterization of the dnaK gene region of Clostridium acetobu-tylicum, including grpE, dnaJ, and a new heat shock gene. J.Bacteriol. 174:3290-3299.
33. Neidhardt, F. C., and R A. VanBogelen. 1987. Heat shockresponse, p. 1334-1345. In F. C. Neidhardt, J. L. Ingraham, K. B.Low, B. Magasanik, M. Schaechter, and H. E. Umbarger (ed.),Escherichia coli and Salmonella typhimurium: cellular and molec-ular biology. American Society for Microbiology, Washington,D.C.
34. Ohta, T., K. Saito, M. Kuroda, K. Honda, H. Hirata, and H.Hayashi. 1993. Molecular characterization of the gene operon,S70, comprising grpE, dnaK, dnaJ homologues, and two new heatshock genes of Staphylococcus aureus. GenBank accession numberD14715.
35. Osipiuk, J., C. Georgopoulos, and M. Zylicz. 1993. Initiation of XDNA replication: the Escherichia coli small heat shock proteins,DnaJ and GrpE, increase DnaK's affinity for the XP protein. J.Biol. Chem. 268:4821-4827.
36. Palleros, D. R, K. L. Reid, L, Shi, W. J. Welch, and A. L. Fink.1993. ATP-induced protein-Hsp7O complex dissociation requiresK' but not ATP hydrolysis. Nature (London) 365:664-666.
37. Pearson, W. R, and D. J. Lipman. 1988. Improved tools forbiological sequence comparison. Proc. Natl. Acad. Sci. USA85:2444 2448.
38. Phillips, G. J., and T. J. Silhavy. 1990. Heat-shock proteins DnaKand GroEL facilitate export of LacZ hybrid proteins in E. coli.Nature (London) 344:882-884.
39. Saito, H., Y. Nakamura, and H. Uchida. 1978. A transducing Xphage carrying g7pE, a bacterial gene necessary for X DNAreplication, and two ribosomal protein genes rpsP (S16) and rpsS(L19) of Escherichia coli K12. Mol. Gen. Genet. 165:247-256.
40. Saito, H., and H. Uchida. 1977. Initiation of the DNA replicationof bacteriophage lambda in Escherichia coli K12. J. Mol. Biol.113:1-25.
41. Schroder, H., T. Langer, F.-U. Hartl, and B. Bukau. 1993. DnaK,DnaJ and GrpE form a cellular chaperone machinery capable of
repairing heat-induced protein damage. EMBO J. 12:4137-4144.42. Seaton, B., and L. E. Vickery. 1994. A gene encoding a DnaK/
hsp7o homolog in Escherichia coli. Proc. Natl. Acad. Sci. USA91:2066-2070.
43. Sherman, M. Y., and A. L. Goldberg. 1992. Involvement of thechaperonin dnaK in the rapid degradation of a mutant protein inEscherichia coli. EMBO J. 11:71-77.
44. Skowyra, D., C. Georgopoulos, and M. Zylicz. 1990. The E. colidnaK gene product, the hsp7o homolog, can reactivate heat-inactivated RNA polymerase in an ATP hydrolysis-dependentmanner. Cell 62:939-944.
45. Straus, D., W. Walter, and C. A. Gross. 1990. DnaK, DnaJ, andGrpE heat shock proteins negatively regulate heat shock geneexpression by controlling the synthesis and stability of a2. GenesDev. 4:2202-2209.
46. Tabor, S., and C. C. Richardson. 1985. A phage T7 RNApolymerase/promoter system for controlled exclusive expression ofspecific genes. Proc. Natl. Acad. Sci. USA 82:1074-1078.
47. Tilly, K., R. Hauser, J. Campbell, and G. J. Ostheimer. 1993.Isolation of dnaJ, dnaK, and grpE homologues from Borreliaburgdorferi and complementation of Escherichia coli mutants. Mol.Microbiol. 7:359-369.
48. Tilly, K., and N. McKittrick, M. Zylicz, and C. Georgopoulos.1983. The dnaK protein modulates the heat-shock response ofEscherichia coli. Cell 34:641-646.
49. Ueguchi, C., M. Kakeda, H. Yamada, and T. Mizuno. 1994. Ananalogue of the DnaJ molecular chaperone in Escherichia coli.Proc. Natl. Acad. Sci. USA 91:1054-1058.
50. Voos, W., B. D. Gambill, S. Layorala, D. Ang, E. A. Craig, and N.Pfanner. 1994. Mitochondrial GrpE is present in a complex withhsp70 and preproteins in transit across membranes. Mol. Cell.Biol. 14:6627-6634.
51. Wang, R F., and S. R Kushner. 1991. Construction of versatilelow-copy-number vectors for cloning, sequencing and gene expres-sion in Escherichia coli. Gene 100:195-199.
52. Wetzstein, M., and W. Schumann. 1990. Nucleotide sequence of aBacillus subtilis gene homologous to the grpE gene of E. colilocated immediately upstream of the dnaK gene. Nucleic AcidsRes. 18:1289.
53. Wild, J., A. Kamath-loeb, E. Ziegelhoffer, M. Lonetto, Y. Ka-wasaki, and C. A. Gross. 1992. Partial loss of function mutations inDnaK, the Escherichia coli homologue of the 70-kDa heat shockproteins, affect highly conserved amino acids implicated in ATPbinding and hydrolysis. Proc. Natl. Acad. Sci. USA 89:7139-7143.
54. Ziemienowicz, A., D. Skowyra, J. Zeilstra-Ryalls, 0. Fayet, C.Georgopoulos, and M. Zylicz. 1993. Both the Escherichia colichaperone systems, GroEL/GroES and DnaK/DnaJ/GrpE, canreactivate heat-treated RNA polymerase. J. Biol. Chem. 268:25425-25431.
55. Zylicz, M., D. Ang, and C. Georgopoulos. 1987. The grpE proteinof Escherichia coli: purification and properties. J. Biol. Chem.262:17437-17442.
56. Zylicz, M., D. Ang, K. Liberek, and C. Georgopoulos. 1989.Initiation of A DNA replication with purified host- and phage-encoded proteins: the role of the dnaK, dnaJ and grpE heat shockproteins. EMBO J. 8:1601-1608.
57. Zylicz, M., and C. Georgopoulos. 1984. Purification and propertiesof the Escherichia coli dnaK replication protein. J. Biol. Chem.259:8820-8825.
VOL. 176, 1994
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from












![Heterogeneous binding of the SH3 client protein to the ... · representationofthe SBD of DnaK[PDBIDcode1DKX (18)].Hydrophobic sidechains with a fractionalburied area>0.74(43)are shown](https://img.dokumen.tips/doc/110x75/5fc0d07e0b8f763bd91ad51b/heterogeneous-binding-of-the-sh3-client-protein-to-the-representationofthe-sbd.jpg)
















