Embed Size (px)
Citation preview

J. CeU Set. 58, 225-240 (1982) 225Printed in Great Britain © Company of Biologists Limited 1982
ISOLATION AND PROPERTIES OF
VITELLOGENIC FERRITIN FROM SNAILS
W. BOTTKEZoologiiches Institut der Unwersitdt, Badestr. 9,4400 MUnster, F.R.G.
SUMMARYThe iron storage protein ferritin is the principal yolk protein in oocytes of the snails Planor-
barius corneus L. and Lymnaea ttagnalis L. This report gives an account of the isolation pro-cedure and of some properties of snail ferritins. The isolation procedure includes a heat-denaturation step, gel filtration on Sepharose 6B and two ultracentrifugation steps followed byelectrophoresis. Ferritins from both snails are highly reminiscent of vertebrate and plantferritins in terms of heat stability, absorption spectrum and ultrastructure, and both sharecommon antigen determinants with horse spleen ferritin. In different electrophoresis systemssnail ferritins display considerable heterogeneity and microheterogeneity. Electrophoresis in thepresence of sodium dodecyl sulphate (SDS) yields two major polypeptides with molecularweights of 19000 and 24000, which are interpreted to be authentic subunits of the ferritinmolecule.
Different organs and tissues of the snails differ in subunit composition. Midgut gland ferritinconsists predominantly of the 19000 M, polypeptide, while in embryos only the 24000 M, bandwas found. No carbohydrates or lipids could be detected by staining acrylamide gels. Resultsfrom SDS/acrylamide electrophoresis, electrophoresis under non-denaturing conditions ongradient gels and from isoelectric focusing indicate that the ferritins of both snails are composedof at least two different types of ferritin that are tissue-specific. One ferritin is typical of somatictissue (midgut gland) and is most probably a homopolymer of the 19000 MT subunit. The otherferritin is typical of oocytes, but since it is an exogenous protein it is also encountered in themidgut gland (the presumed site of yolk synthesis) and the haemolymph. Vitellogenic ferritinis either a homopolymer of the 24000 MT subunit or is predominantly composed of it. So far,there is no evidence for a precursor-product relationship between the two subunits.
INTRODUCTION
Iron has a central role in all living organisms as a constituent of many enzymes,electron-transfer complexes and oxygen carriers. Since free iron is highly toxic tocells, organisms have to protect against its harmful effects and simultaneously toconserve supplies of the metal for the synthesis of functional iron compounds. Bothfunctions are combined in the ferritin molecule (see reviews by Crichton, 1973;Munro & Linder, 1978; Clegg, Fitton, Harrison & Treffry, 1980; Aisen & Listowsky,1980).
Ferritin is a water-soluble spherical molecule consisting of a multisubunit proteinshell in the form of a hollow sphere that surrounds an inorganic iron core of approx-imate composition (FeOOH)8(FeOOPO3H2) in the form of microcrystalline particles.It is assumed that the iron is made accessible to the cell by reduction or by proteolyticdegradation of whole ferritin molecules inside lysosomes. Ferritin is distributedubiquitously in eukaryotes and prokaryotes (Crichton, 1973; Crichton et al. 1978;

226 W. Bottke
Yariv et al. 1981). Ferritins from diverse sources - bacteria, plants and mammals,including man - have been investigated. However, reports dealing with the occurrenceand properties of the protein in invertebrates are scarce: almost nothing is known ofthe function of ferritin in these animal groups. In molluscs, ferritin has been foundin a chiton (Towe, Lowenstam & Nesson, 1963), a freshwater bivalve (Baba, 1969),a gastropod snail (Heneine, Gazzinelli & Tafuri, 1969) and Octopus (Nardi, Muzii &Puca, 1971). The most detailed investigation of a molluscan ferritin has been carriedout on a ferritin from the coelomic fluid of the mussel Corbicula (Baba, May & Fish,1977). Interestingly, this ferritin has been shown to deviate considerably in immuno-logical and physical properties from other better-known ferritins, suggesting that thestructure of ferritin in general is not as consistent as was anticipated previously (cf.Clegg et al. 1980). The method as devised by Baba et al. for the isolation of molluscanferritin is time-consuming.
Recently, we presented evidence that ferritin is the principal yolk protein of certainfreshwater snails (Bottke & Sinha, 1979; Bottke, Sinha & Keil, 1982). In these snailsferritin is an exogenously synthesized yolk protein, which is sequestered by the oocytesby adsorptive endocytosis, thus suggesting a highly specific uptake of the molecules(Bottke, Sinha & Keil, 1982). This ferritin is easily accessible in sufficient amountsfor biochemical studies on structure and properties. In our opinion this ferritin deservesspecial attention, not only because investigations on this invertebrate protein bridge thegap between bacteria and plant ferritins on one side and mammalian ferritin on theother, but also because this snail ferritin can be expected to fulfil special tasks in embryo-genesis. Moreover, this ferritin most probably carries a signal that allows its specificrecognition and uptake by the oocyte. In this report I describe the isolation procedureand some properties of this ferritin. As a result evidence is obtained for considerableheterogeneity of the protein, suggesting that there are at least two different types offerritin, of which only one is vitellogenic.
MATERIALS AND METHODS
Materials
Adult laboratory-bred specimens of the pulmonate snails Planorbarius corneas L. andLymnaea ttagnalis L. were used. The animals were reared as described previously (Bottke et al.1982). In Planorbarius, gonads and midgut glands were dissected separately, in Lymnaea wholevisceral masses were removed. Embryos of both snails were isolated manually from egg capsuleswith needles and micropipettes. All tissues were frozen in liquid nitrogen and stored at — 50 °Cuntil used. Haemolymph from Planorbarius and Lymnaea was obtained by puncturing the foot,and the solution was clarified by centrifugation at 23000 g for 20 min.
Chemicals
Sepharose 6B-C1, Sephacryl S-300 and precast polyacrylamide gradient gels (PAA 4/30)were bought from Pharmacia (Uppsala, Sweden); anti-horse-spleen-ferritin antiserum, carrierampholytes and precast polyacrylamide gels for isoelectric focusing were obtained from LKB-Produkter (Bromma, Sweden) and Serva (Heidelberg, FRG). Acrylamide was bought fromServa. All chemicals were analytical grade.

Vitellogenic ferritin isolated from mails 227
Ferritin preparation
The procedures used for the purification of snail ferritins are outlined in Fig. 1. They are acombination of several methods suggested for the purification of ferritins from different sources(cf. Crichton, 1973). Our isolation procedure starts with the preparation of an acetone powderin order to eliminate the large amounts of lipids present in the gonads and midgut glands ofsnails. The powder was prepared essentially as described by Morton (1955) and Nardi et al.(1971). About 30 g of dry acetone powder are obtained from 180 g tissue. Extraction of theacetone powder, column chromatography and dialysis of the samples were carried out withhomogenization buffer (o-i M-Tris-HCl, pH8, o-i M-NaCl, 0-02%NaN,, 1 mM-phenyl-methylsulfonylfluoride as a protease inhibitor).
Homogenize frozen tissue in a blender and prepare acetone powderI
Extract proteins from powder and subject to heat denaturation (8 min at 75 °C)I
Centrifuge at 10000 g for 20 min and make supernatant up to 60 % saturation with ammoniumsulphate
ICollect proteins by centrifugation at 4000 g and dissolve in buffer. Dialyse against buffer for
at least 24 hICentrifuge suspension for 45 min at 23 000 g and pass supernatant over a column of Sepharose-6B
IConcentrate ferritin by ultrafiltration (filter PMio, Amicon)
ICentrifuge ferritin solution at 200000 g for 2-5 h and redissolve precipitate in buffer. Removeinsoluble material by centrifugation at 23 000 g
ICentrifuge ferritin solution on a cushion of CsCl (£) = 1 -4) dissolved in buffer. Dissolve ferritin
pellet in buffer and dialyse against the same bufferI
Repeat the last step. Dialyse ferritin against buffer as desired for further useI
Fractionate ferritin into monomers and oligomers by passage over a column ofSephacryl S-300
IPrepare apoferritin by treatment with thioglycollic acid
Fig. 1. Outlines of ferritin preparation from gonads and midgut glands of pulmonatesnails (P. corneus L. and L. stagnalis L.). D, density.
Apoferritin was prepared by dialysing samples of ferritin against 1 % thioglycollic aciddissolved in 01 M-acetate buffer (pH 5) until the solution was colourless. Thioglycollate wasremoved by extensive dialysis against water. About 50 mg apoferritin are obtained from 1 kgsnail tissue. For separation of monomers and higher oligomers, ferritin samples were passedover a column of Sephacryl S-300, equilibrated with homogenization buffer but made up to0-5 M-sodium chloride (column: 140 cm x 1-4 cm). Ferritins were concentrated by ultrafiltra-tion (filter PMio, Amicon). Protein concentrations were determined routinely by the methodof Lowry, Rosebrough, Farr & Randall (1951) using bovine serum albumin as a standard.
Preparation of ferritin from haemolymph and embryos
Haemolymph proteins were extensively dialysed against homogenization buffer and pre-cipitated by 60% saturation with ammonium sulphate. The proteins were passed over aSepharose 4B column. Fractions at the elution position of ferritin were inspected for presence

228 W. Bottke
of the protein by electron microscopy and concentrated by ultrafiltration. Finally, the proteinwas obtained by ultracentrifugation at 200000 g for 2-5 h. The small amounts of ferritin thusobtained precluded any further purification.
Embryo ferritin was obtained by homogenizing 1-day-old embryos and subjecting thehomogenate to heat denaturation. The suspension was clarified by centrifugation for 20 min at23 ooo.g and the supernatant was lysed directly in electrophoresis buffer.
ImTnunodiffusion
Radial immunodiffusion of snail ferritins was performed according to the Ouchterlonymethod in 1 % agarose gels prepared in o-1 M-phosphate buffer (pH 7-2), o-i M-sodium chlorideand o-oa% sodium azide. A commercial antiserum against horse spleen ferritin (rabbit) wasused. Hor3e spleen ferritin was diluted with buffer and the appropriate conditions tested byserial dilution.
Electron microscopy
Preparation of samples and measurements on micrographs were done as described previously(Bottke et al. 1982).
Absorbancy
Snail ferritins were dialysed against 0-05 M-Tris • HC1 buffer (pH 7) and the absorptionspectra in the ultraviolet and visible regions were recorded in a Zeiss PMQ2 photometer.
Polyacrylamide electrophoresis
Disk electrophoresis essentially followed the method of Ornstein-Davis (Maurer, 1971).Samples of up to 50 fig were run on 5 % gels with and without spacer gels. The electrophoresiswas carried out at a constant current of 3-5 mA per gel for 45-90 min. Gels were stained withCoomassie Blue or Prussian Blue for ferric iron. For detection of carbohydrates and lipids, gelswere stained by the periodic acid-Schiff (PAS) reaction according to Zacharius et al. (1969) orwith Sudan Black B according to Ressler (1961) and Prat (1969) (all three references quotedby Maurer, 1971).
For better resolution, samples were run on commercial PAA 4/30 gradient gels (Pharmacia;gradient: 4% to 30% acrylamide) or on 4/20 gradient gels (gradient: 4% to 20% acrylamide)prepared in Tris-EDTA-borate buffer (0-09 M-Tris, 0-08 M-borate, 0-003 M-Na,EDTA,pH 8-4). Electrophoresis was carried out at 20 °C for 15 h at a potential difference of 120 V.Ferritin bands were cut out of the gels and the proteins eluted by diffusion in homogenizationbuffer, O'5 M-sodium chloride.
Add/urea electrophoresis
For electrophoresis at low pH, ferritin was dissociated into subunits by treatment with 67 %acetic acid at o °C for 8 h. Processing of the samples and electrophoresis was carried out asdescribed by Panyim & Chalkley (1969) and Arosio, Adelman & Drysdale (1978).
SDS/acrylamide gel electrophoresis
Solutions of ferritin (1-3 mg/ml) were dissociated by heating at 100 °C for 8 min in o-oi M-phosphate or imidazole buffer (pH 7), 2-5% (v/v) in /?-mercaptoethanol, 1-5% (w/v) insodium dodecyl sulphate (SDS). Samples of 10—40 fig protein were electrophoresed on 10 %acrylamide slab gels (260 mm x 125 mm x 2 mm) on an LKB Multiphor electrophoresisapparatus at a potential difference of 120 V and a temperature of 8 °C. Alternatively, ferritinsamples were run on cylindrical gels (80 mm x 5 mm). Estimation of the molecular weights offerritin subunits was carried out essentially as described by Weber & Osborn (1969). Subunitbands were cut out of the gels and homogenized in imidazole buffer, 1% in SDS and /?-mercaptoethanol. Recovered proteins were dialysed extensively against water and concentratedby lyophilization.

Vitellogenk ferritin isolated from snails 229
Isoelectric focusing
Ferritin samples of 10-50 fig were fractionated by isoelectric focusing in the pH range from3-5 to 10 on precast slab gels (LKB PAG-plates, Bromma, Sweden) or on 5% acrylamide gelsprepared with ampholytes from LKB or Serva (Heidelberg, FRG).
Isoelectric focusing was carried out on a Multiphor apparatus (LKB) at a temperature of2 °C at an average power of 5 W/gel. pH value readings were made with a flat-membranemicroelectrode (Ingold LOT 403-M8).
RESULTS
Isolation
We had to isolate ferritins from whole organs since the oocytes are rather small andnot obtainable in sufficient quantities. Consequently a large amount of soluble non-ferritin proteins was extracted together with the ferritins (Fig. 3 A).
A number of different isolation procedures are recommended for ferritins fromvarious sources (cf. Crichton, 1973). However, the isolation of ferritin from snailspresents special problems. These are due to the presence in snails of mucus substances,large amounts of lipids in the gonad, high molecular weight haemoglobin (Svedberg,1933) and proteolytic enzymes from the midgut gland and somatic cells of the gonad.We have been unsuccessful in isolating ferritins by ultracentrifugation as describedby Penders, De Rooij-Dijk & Leijnse (1968), since haemoglobin is isolated togetherwith the ferritin, and the yields are low, probably due to the low iron content of snailferritin. Likewise, ferritin could not be obtained in a pure state by ion-exchangechromatography on DEAE-Sephadex. A large amount of the material is eluted overa broad range of ionic strengths or is immobilized on the gel, resulting in low yieldsalso. Extraction of the tissues with butanol/water, as recommended for fungal ferritin(David & Easterbrook, 1971), resulted in poor yields that were only sufficient forobservation in the electron microscope.
Since difficulties arise from the large amounts of lipids in the yolk and somaticauxiliary cells of the gonad our procedure starts with an acetone treatment. Weassume that the effectiveness of this step is due not only to the removal of lipids butalso to the breakdown of the yolk platelets. Acetone has been used in the purificationof ferritins from other sources (cf. Crichton, 1973). However, electron microscopicalobservations suggest that acetone causes a considerable disruption of the protein shellsof ferritin (Massover, 1977). Comparing electron micrographs of different prepara-tions with and without the acetone step we could not detect any difference in thefrequency of disrupted particles in our material. Since the heat-denaturation stepmight also be harmful for snail ferritin we compared samples of ferritin, isolated withor without this step, by isoelectric focusing. As a result, no qualitative difference wasrecorded between the focusing profiles of the samples. As shown by the presence offerritin in the supernatants following ultracentrifugation, only ferritin molecules witha high iron content are isolated.
We could not detect any difference between the ferritin in the pellet and that in the

230 W. Bottke
supernatant, using isoelectric focusing. However, quantitative differences in theproportions of individual bands in isoelectric focusing have not been excluded.
Properties offerritin
Snail ferritins dissolve readily in dilute buffers at pH values above 6. Followingextensive dialysis against water adjusted to pH 4-5-5-5 the bulk of the ferritin isprecipitated. Preparations of Planorbarius ferritin are slightly turbid, probably becausethey contain higher amounts of oligomeric ferritin. The apoferritins of both snails,obtained by chemical treatment of the respective ferritins, do not completely dissolvein dilute buffers. In particular, the material from Planorbarius is poorly soluble. Wewere unsuccessful in crystallizing the ferritins of both snails with 5 % cadmiumsulphate, producing only an amorphous yellow precipitate. Concentrated samples offerritin exhibit the typical brownish yellow colour. The absorption spectra of snailferritins correspond to that of horse spleen ferritin. The absorption spectra of apoferri-tins likewise correspond to that of vertebrate apoferritin. No characteristic absorptionwas recorded in the region of longer wavelength, indicating that the ferritin prepara-tions are practically free from other intrinsic or contaminating chromophores, e.g.haem groups from haemoglobin. Staining of ferritin samples (run on 5 % acrylamidegels or on SDS/acrylamide gels for detection of carbohydrates) using the PAS reactionyielded negative results. Likewise, no lipids could be detected by staining gels withSudan Black B.
infrastructure of ferritins
Electron micrographs of negatively stained snail ferritins demonstrate the strikingarrangement of the multi-subunit protein shells (Fig. 2 c). In preparations that havenot been subjected to ultracentrifugation, apoferritin molecules or ferritin moleculeswith a rather low iron content are also seen. Measurements on micrographs yieldeddiameters of 11-13 nm for individual molecules. In addition to complete shells, in-complete ones, which probably arise by loss of subunits, are seen infrequently. Thus,snail ferritins do not differ in size and shape from the better-known ferritins of otherspecies (cf. Massover & Cowley, 1975; Massover, 1977). Aggregates offerritin mole-cules are often seen in addition to free particles, and typically form monolayers inwell-spread preparations. It is not clear whether these aggregates still consist offerritin particles or represent haemosiderin.
Immunology
Purified snail ferritins were inspected for the presence of antigen determinantssimilar to those of horse spleen ferritin using an antiserum from rabbit. Interestingly,both ferritins react with the antiserum (Fig. 2 A, B). Since a distinct spur was detectedwhen the homologous antigen was included in the test, the reaction was judged to beone of partial identity. The precipitin bands could be stained for protein and iron.

Vitellogertic ferritin isolated from snails
B
Fig. 2A, B. Double diffusion plates showing reactions of partial identity for horsespleen ferritin (h) and snail ferritins (s), when tested with rabbit antiserum againsthorse spleen ferritin (central well). L. stagnalis (A), P. corneus (B). Coomassie Bluestaining, c, D. Purified preparations of Planorbarhu ferritins obtained from differentbands of an acrylamide gradient gel on which whole gonadal ferritin was fractionated(see Fig. 6 A). Protein coats outlined by means of 2 % neutral potassium phosphotung-state. c. Ferritin extracted from band I of the gradient gel; D, ferritin extracted fromband II of the gradient gel. c, D, X 330000.
Polyacrylaim.de gel electrophoresis
The purity of preparations was routinely assessed by electrophoresis on 5 %acrylamide gels. The patterns obtained with ferritins from both snails are shown inFig. 3. They yield several bands, which all stain for protein and iron (Fig. 3B, F). Weinterpret the different bands as the monomers and oligomers of the ferritin molecule,by analogy with observations on other ferritins (Lee & Richter, 1976).
All bands observed in 5 % gels yield the same subunit pattern when run under

232 W. Bottke
1 0 -
8 0-5-
0-25-
Planorbariusgonad
Seph. S-300
K26/100
Oligo- Di- Monomeric-Ferritin fractions
90
A .... B
e o
I II II
: 1 1# • •

Vitellogenic ferritin isolated from snails 233
denaturing conditions (Fig. 3E, F, G). The amount of higher oligomers that do notenter the gels differs from isolation to isolation and increases with the storage time ofthe protein following its isolation.
The ferritin monomers form a conspicuous broad band (Fig. 3B, F). In Planorbariusthis band splits up into at least two sub-bands if a spacer gel is used. If much proteinis applied to the gels a distinct smearing of the protein is observed between themonomer and dimer band (Fig. 3B). The ferritins from both snails have the samemobility but are well-separated from horse spleen ferritin, which runs considerablyfaster. An even greater heterogeneity of the proteins is revealed when they are frac-tionated on polyacrylamide gradient gels (4% to 30% and 4% to 20% acrylamide).In both ferritins the monomer band splits up into four sub-bands that all stain foriron. Two dimer bands are seen (Figs. 3 c, D and 6A). Proteins were eluted from thetwo most prominent bands in the monomer region and inspected by electron micro-scopy. From both bands typical ferritin molecules were obtained (Fig. 2C, D). Thus,the different sub-bands most probably represent populations of ferritins that differ insize, but differences in charges cannot be excluded.
SDSI'acrylamide gel electrophoresis
Molecular weights of the SDS-subunit complexes were estimated by runningsuitable standard proteins together with the ferritin. The standard curve was basedon the mean values of eight runs obtained from six different isolations. The resultsin Fig. 4 show that snail ferritins display multiple bands. In both ferritins there aretwo principal polypeptides with molecular weights of 19000 and 24000 (Fig. 4c, D,E). In addition, some weaker bands in the high and low molecular weight region areseen. For the two most prominent small polypeptides molecular weights of 11000 and13000, respectively, were calculated. A 33000 MT polypeptide is regularly encoun-tered in the high molecular weight region. The molecular weights of the larger poly-peptides could not be estimated with accuracy since they are beyond the linear rangeof the standard curve.
Fig. 3. Polyacrylamide gel electrophoresis of ferritin preparations. A. Extract from thegonad of Planorbarius; acrylamide gradient gel (4% to 30% acrylamide); PrussianBlue positive bands ( • ) . B. Planorbarius ferritin, run under non-dissociating condi-tions on a s % gel. The strong band is due to ferritin monomers while the weak bandpresumably represents dimer3. Prussian Blue staining, c. Fractionation of gonadferritin from Planorbarius on a non-denaturing acrylamide gradient gel (4 % to 30 %).Prussian Blue staining. Four monomer bands and two dimer bands can be recognized.D. Fractionation of midgut gland ferritin from Planorbarius on an acrylamide gradientgel (4 % to 30 %). Prussian Blue staining. Only two monomer bands are seen. E. Wholegonad ferritin from Planorbarius resolved into three fractions by gel chromatographyon a Sephacryl S-300 column (I, II and III). F. The three different fractions from Eyield different electropheretograms, which presumably show higher oligomers, dimersand monomers of the ferritin molecule. Electrophoresis in non-dissociating conditions.Prussian Blue staining, 5 % acrylamide gel. G. The three different fractions shown inE essentially yield the same SDS-subunit complexes under denaturing conditions.Equal amounts of protein applied to the gels (10% acrylamide/SDS gels).
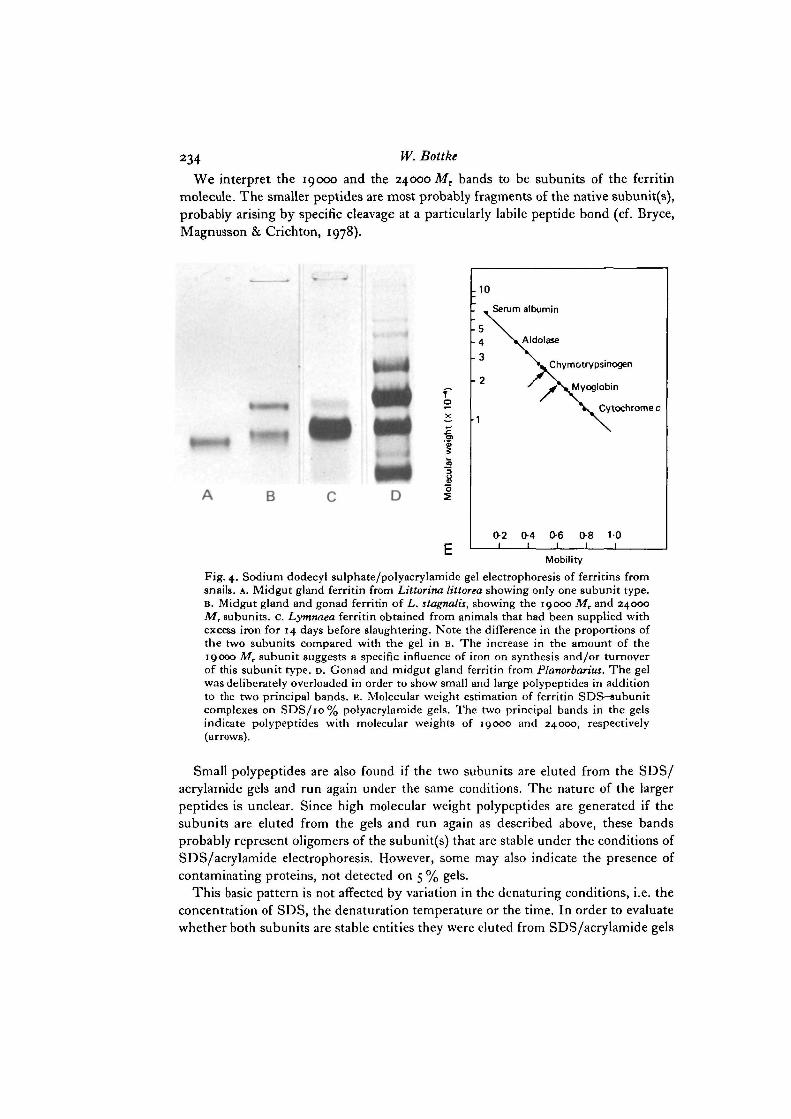
234 w- Bottke
We interpret the 19000 and the 24000 Mr bands to be subunits of the ferritinmolecule. The smaller peptides are most probably fragments of the native subunit(s),probably arising by specific cleavage at a particularly labile peptide bond (cf. Bryce,Magnusson & Crichton, 1978).
. 1 0
fa* To
•§>
B
Serum albumin
lAldolase
L Chymotrypsinogen
^Myoglobin
Cytochrome c
0-21
0-4I
O61
0-8 101
Mobility
Fig. 4. Sodium dodecyl sulphate/polyacrylamide gel electrophoresis of ferritins fromsnails. A. Midgut gland ferritin from Littorina littorea showing only one subunit type.B. Midgut gland and gonad ferritin of L. stagnalis, showing the 19000 Mr and 24000Mr subunits. C. Lymnaea ferritin obtained from animals that had been supplied withexcess iron for 14 days before slaughtering. Note the difference in the proportions ofthe two subunits compared with the gel in B. The increase in the amount of the19000 Mr subunit suggests a specific influence of iron on synthesis and/or turnoverof this subunit type. D. Gonad and midgut gland ferritin from Planorbarius. The gelwas deliberately overloaded in order to show small and large polypeptides in additionto the two principal bands, E. Molecular weight estimation of ferritin SDS-subunitcomplexes on SDS/10% polyacrylamide gels. The two principal bands in the gelsindicate polypeptides with molecular weights of 19000 and 24000, respectively(arrows).
Small polypeptides are also found if the two subunits are eluted from the SDS/acrylamide gels and run again under the same conditions. The nature of the largerpeptides is unclear. Since high molecular weight polypeptides are generated if thesubunits are eluted from the gels and run again as described above, these bandsprobably represent oligomers of the subunit(s) that are stable under the conditions ofSDS/acrylamide electrophoresis. However, some may also indicate the presence ofcontaminating proteins, not detected on 5 % gels.
This basic pattern is not affected by variation in the denaturing conditions, i.e. theconcentration of SDS, the denaturation temperature or the time. In order to evaluatewhether both subunits are stable entities they were eluted from SDS/acrylamide gels

Vitellogenic ferritin isolated from snails 235
and run again under the same conditions (Fig. 5 G, H). Apart from the generation ofseveral peptides from both subunits no alteration of the banding pattern was seen. No19000 Mr band was produced from the 24000 MT band and vice versa.
In Planorbarius it is possible to dissect the midgut gland, the presumed site ofsynthesis of vitellogenic ferritin, from the gonad and to inspect the ferritins from thetwo organs separately (Fig. 3 c, D). AS a result striking differences in the proportionsof the subunits are seen (Fig. 5 c, D). Whereas in the midgut gland the 19000 MT bandprevails, in the gonad the 24000 MT band is the predominant one. In embryos lysed
1
rm
cc
B C D E F G H
Fig. 5. Sodium dodecyl sulphate/polyacrylamide gel electrophoresis of ferritins fromdifferent organs and tissues of Lymnaea and Planorbarius. A. Molecular weight markers(serum albumin, s; aldolase, a; chymotrypsinogen A, c; myoglobin, m; cytochromec, cc). B. Ferritin from haemolymph of Planorbarius. c. Midgut gland ferritin; the19000 Mr band is the prevailing one; Planorbarius. D. Gonad ferritin from Planor-barius; the 24000 Mr polypeptide prevails. E. Whole embryos lysed in SDS buffer.Whereas the 24000 Mr subunit is prominent the presence of a 19000 M, subunit isdoubtful; Planorbarius. F. Ferritin from the visceral mass of Lymnaea shows bothsubunit-SDS complexes in nearly equal amounts. However, also compare with Fig.4B and c. G. The 19000 Mr polypeptide of Lymnaea ferritin isolated by SDS/poly-acrylamide gel electrophoresis and run again under the same conditions. H. The24000 M, polypeptide of Lymnaea ferritin isolated by SDS/polyacrylamide gelelectrophoresis and run again under the same conditions, j . Whole embryos ofLymnaea lysed in SDS buffer. Note the distinct 24000 M, subunit.
directly in SDS buffer only the 24000 MT band is encountered (Fig. SE, J). Thus, the24000 Mr subunit seems to be typical for embryo ferritin, the 19000 Mr subunit formidgut gland ferritin. In addition to earlier observations in which only the 19000 Mr
band was detected in the haemolymph (Bottke & Sinha, 1979), we can now confirmthe presence of the 24000 MT band for the haemolymph also (Fig. 5B). In Lymnaeano discrimination between the two ferritin-bearing organs is possible. However, inLymnaea also the 24000 MT band is typical for the embryo (Fig. 5J), whereas the19000 MT band is the prevailing band for the whole visceral mass (Fig. 4c). Sincethe gonad in Lymnaea comprises only a rather small part of the visceral mass it isreasonable to ascribe the bulk of the 19000 Mr material to the midgut gland ferritin.
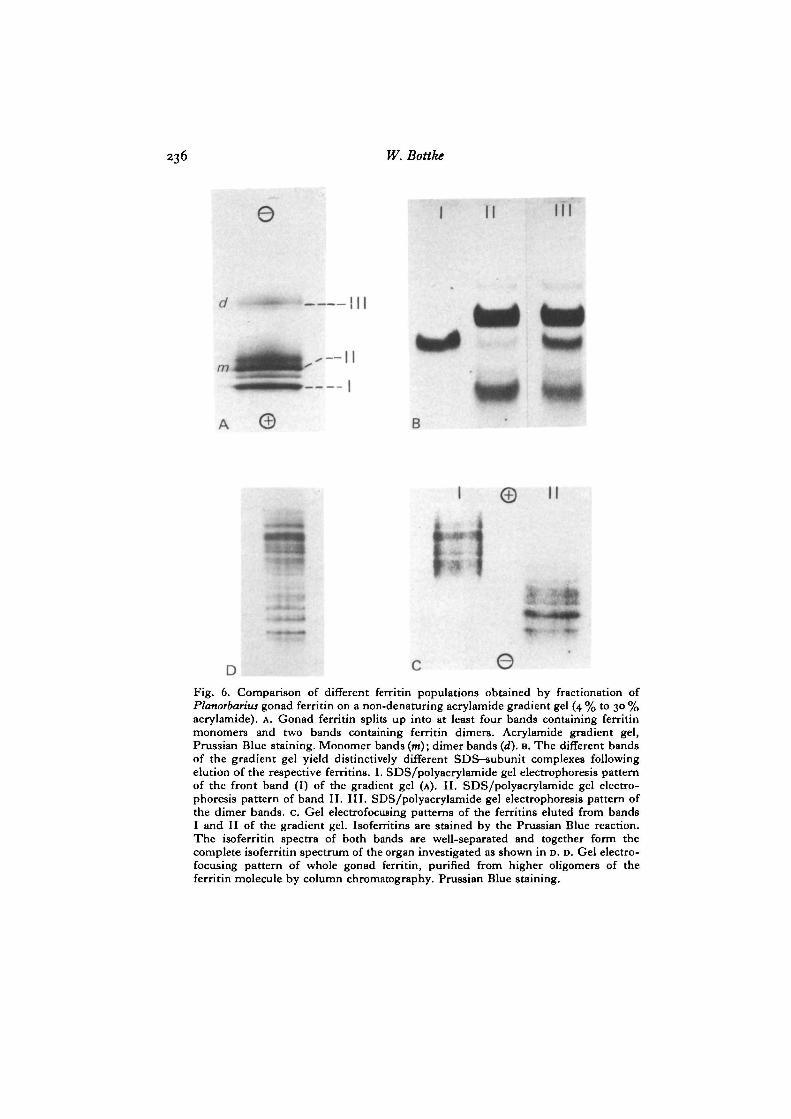
236 W. Bottke
Ill
---II
- - - I
A © B
1:1D u w
Fig. 6. Comparison of different ferritin populations obtained by fractionation ofPlanorbarius gonad ferritin on a non-denaturing acrylamide gradient gel (4 % to 30 %acrylamide). A. Gonad ferritin splits up into at least four bands containing ferritinmonomers and two bands containing ferritin dinners. Acrylamide gradient gel,Prussian Blue staining. Monomer bands (m); dimer bands (d). B. The different bandsof the gradient gel yield distinctively different SDS-subunit complexes followingelution of the respective ferritins. I. SDS/polyacrylamide gel electrophoresis patternof the front band (I) of the gradient gel (A). II. SDS/polyacrylamide gel electro-phoresis pattern of band II. III. SDS/polyacrylamide gel electrophoresis pattern ofthe dimer bands, c. Gel electrofocusing patterns of the ferritins eluted from bandsI and II of the gradient gel. Isoferritins are stained by the Prussian Blue reaction.The isoferritin spectra of both bands are well-separated and together form thecomplete isoferritin spectrum of the organ investigated as shown in D. D. Gel electro-focusing pattern of whole gonad ferritin, purified from higher oligomers of theferritin molecule by column chromatography. Prussian Blue staining.

ViteUogenic ferritin isolated from snails 237
Acid/urea electrophoresis
On acid/urea gels a remarkable parallel is obtained to the pattern recorded inSDS/acrylamide electrophoresis in terms of the number of bands and their relativeintensity of staining. This is especially striking in the patterns seen with midgut glandand gonad ferritin of Planorbarius, which largely correspond to the patterns of bothorgans in SDS/acrylamide electrophoresis.
Isoelectric focusing
Ferritin samples from both snails enriched in monomers by gel filtration onSephacryl S-300 were run in the pH range from 3-5 to 10. Both ferritins show amarked tendency towards smearing. This could be partly overcome by prefocusingand addition of 0-5 % Triton X-100 to the samples. The smearing is especially strongin Planorbarius ferritin and is evidently due to the presence of the 24000 Mr subunit.Like other ferritins, snail ferritins are acidic proteins with a pronounced negativecharge at physiological pH values. Both snail ferritins split up into a lot of bands,isoelectric between pH 49 and 6-5, that we term isoferritins, analogous to otherferritins for which the molecular basis of the heterogeneity is not established (Fig. 6D).
Lymnaea and Planorbarius ferritins have many isoferritins in common. All bandsare well-separated from horse spleen isoferritins when mixed with snail ferritins orrun on an adjacent track of the gel. In order to relate the observed charge hetero-geneity of the protein shells to the subunit composition, ferritin samples were firstfractionated on gradient gels (Fig. 6A). We then cut out the most prominent bandsand investigated the eluted proteins by SDS/acrylamide electrophoresis and iso-electric focusing. The results of this experiment (see Fig. 6) show that ferritin mono-mers with the highest mobility consist predominantly of the 19000 MT subunit andfocus in the most acidic part of the spectrum (Fig. 6B, C). Ferritin monomers withslower mobility in the gradient gel are predominantly composed of the 24000 MT
subunit (Fig. 6 B) and focus in the more basic region of the spectrum (Fig. 6 c). Theisoferritin profiles of both fractions from the gradient gel are partly overlapping andtogether form the complete spectrum of the organ investigated (Fig. 6D).
DISCUSSION
Ferritin as a yolk protein deserves special interest on account of its restrictedbiological function and on account of its regulation by iron, which is in strikingcontrast to the regulation of hormonally induced typical yolk proteins. Previousreports dealing with iron metabolism in snails suggest that ferritin synthesis in thisgroup is induced by iron, as it is in vertebrates or plants. Snails accumulate con-siderable amounts of iron if kept in an environment with an excess of the metal(Deschiens, Bertrand & Molinari, 1957). From the different heavy metals accumulatedby snails (Fe, Cu, Zn, Cd), only iron is obviously transferred to the gonad andappears in the young in appreciable quantities (Johnson, Angel & Erickson, 1962;Yager & Harry, 1966). The isolation of ferritin from different organs of the snailsreported here indicates that the protein is not only a detoxifying agent but also acts

238 W. Bottke
as the transport vehicle for iron. On account of this dual function, snail ferritin canbe expected to exhibit unusual properties, i.e. different from those of other tissueferritins.
In terms of heat stability, absorption spectrum and gross molecular structure asrevealed by electron microscopy, snail ferritin is fairly similar to other better-knownferritins. In the Ouchterlony test both snail proteins yield a partial cross-reaction withhorse spleen ferritin, thus confirming the notion that ferritin is a rather conservativeprotein on the evolutionary scale. Our observations extend previous reports thatestablished immunological relatedness of ferritins from horse, dog, man and frog(Granick, 1943; Richter, 1967; Crichton, Millar, Cumming & Bryce, 1973). Weexpect to find some common sequences in horse spleen and snail ferritin in the lightof recent sequencing results, which show extensive regions of sequence homology inferritins from different sources, both animal and plant (cf. Clegg et al. 1980). Interest-ingly, apoferritin from the freshwater mussel Corbicula has been reported to beimmunologically unreactive with rabbit antihuman ferritin antiserum (Baba et al.1977). In snail ferritin the heterogeneity and microheterogeneity displayed by differentelectrophoresis systems are most striking.
It is unclear how this microheterogeneity originates at a molecular level. However,in the light of results obtained with vertebrate ferritins some explanations can beconsidered. Clearly, some heterogeneity can be expected from methodological artifactsin isoelectric focusing, as suggested by Shinjo & Harrison (1979). Likewise, someheterogeneity probably originates from disruption of ferritin molecules during theisolation procedure or from proteolytic degradation of yolk in vivo, as shown byincomplete ferritin shells in the electron microscope. Since we have no indication ofthe presence of carbohydrates in our ferritins, either by staining of gels, by treatingferritins with neuraminidase or by isoelectric focusing, we assume that carbohydratesare not responsible for the heterogeneity. However, since the staining methods arenot sensitive enough to detect small amounts of carbohydrates and since human serumferritin is a glycoprotein (Cragg, Wagstaff & Worwood, 1981), this point deservesfurther consideration.
In our opinion, the heterogeneity is due to the presence of two types of subunit ofthe ferritin molecule, which differ in size and charge. The molecular weight of thesmaller subunit (19000), as inferred from SDS/acrylamide electrophoresis, cor-responds fairly well to those obtained for other ferritins. Apoferritin from Lymnaea,which consists predominantly of the 19000 Mr subunit, yielded a value of 16-2 S byboundary sedimentation ultracentrifugation, similar to sedimentation coefficients ofother ferritins (Crichton, 1973). Therefore, we expect ferritin, which consists mainly(or exclusively) of the smaller (19000 MT) subunit, to show the typical ferritin archi-tecture of 24 subunits. It is an open question whether this holds also for 'heavyferritin', i.e. the one that consists of the 24000 Mr subunit. We have not been able toobtain the sedimentation coefficient of this apoferritin since it is poorly soluble indilute buffers. The molecular weight of the heavy subunit as estimated by SDS/acrylamide electrophoresis is comparatively high compared with the subunit size ofother ferritins (cf. Clegg et al. 1980).

Vitellogenic ferritin isolated from snails 239
Interestingly, a similar value has been reported for another molluscan ferritin(23000), using two different analytical approaches (Baba et al. 1977). Since we havenot succeeded in dissociating the 24000 MT polypeptide from the ferritin withoutdisrupting the structure of the ferritin molecule as a whole, and since we cannot infera simple precursor-product relationship between both subunits from our gels, weconsider the 24000 Mr polypeptide to be an authentic subunit of the ferritin molecule.Of course this assumption needs more stringent evidence, such as amino acid analysisor fingerprinting (Bottke, unpublished data).
Our results are most easily explained on the basis of the heteropolymer hypothesisof ferritin structure (Drysdale, Hazard & Righetti, 1975; Arosio et al. 1978; Otsuka,Maruyama & Listowsky, 1981). According to this model, microheterogeneity offerritin is generated at a molecular level from the presence of at least two differenttypes of subunits, termed H- and L-subunit: both assemble in different proportionsin the multimeric ferritin molecule. Both subunits have been demonstrated to beauthentic polypeptides that are similar in amino acid composition, but which alsoshow marked dissimilarities in conformation and properties (Otsuka et al. 1981). Inour material we find two polypeptides that differ in size and charge. Both occur indifferent organs of the snails in different proportions. Since gonad and embryoferritins are highly enriched in the heavy polypeptide, vitellogenic ferritin most prob-ably consists of the 24000 Mt subunit. In contrast, ferritin from somatic cells (midgutgland) preferentially (or exclusively) consists of the 19000 MT subunit. It will be veryinteresting to compare the structure of the two subunits, since it may provide a clueto understanding the basic mechanisms of specific recognition and uptake of vitello-genic ferritin at the oolemma.
I acknowledge the advice given to me by Mrs I. Sinha, Professor DrW. K6hnlein (analyticalultracentrifugation) and Dr U. Mays. This work was supported by the Deutsche Forschungs-gemeinschaft and the Stiftung Volkswagenwerk.
REFERENCES
AISEN, PH. & LISTOWSKY, I. (1980). Iron transport and storage proteins. A. Rev. Biochem. 49,357-393-
AROSIO, P., ADELMAN, TH. G. & DRYSDALE, J. W. (1978). On ferritin heterogeneity. J. biol.Chem. 253, 4451-4458.
BABA, A. (1959). Crystallization of ferritin from coelomic fluid of Corbicula sandai with am-monium sulfate. J. Biochem. (Tokyo) 65, 915-923.
BABA, A., MAY, M. E. & FISH, W. W. (1977). The properties of Corbicula sandai apoferritin.Biochim. biophys. Acta 491, 491-496.
BOTTKE, W. & SINHA, I. (1979). Ferritin as an exogenous yolk protein in snails. Wilhelm RouxArch. EntviMech. Org. 186, 71-75.
BOTTKE, W., SINHA, I. & KEIL, I. (1982). Coated vesicle-mediated transport and deposition ofvitellogenic ferritin in the rapid growth phase of snail oocytes. J. Cell Set. 53, 173-191.
BRYCE, CH. F. A., MAGNUSSON, C. M. & CRICHTON, R. R. (1978). A reappraisal of the electro-phoretic patterns obtained from ferritin and apoferritin in the presence of denaturants.FEBS Lett. 96, 257-262.
CLEGC, G. A., FITTON, J. E., HARRISON, P.M. & TREFFRY, A. (1980). Ferritin: molecularstructure and iron-storage mechanisms. Prog. Biophys. molec. Biol. 36, 56-86.
CRAGG, S. J., WAGSTAFF, M. & WORWOOD, M. (1981). Detection of a glycosylated subunit inhuman serum ferritin. Biochem. J. 199, 565-571.

240 W. Bottke
CRICHTON, R. R. (1973). Ferritin. Structure and Bonding 17, 67-134.CRICHTON, R. R., MILLAR, J. A., CUMMINC, R. L. C. & BRYCE, C. F. A. (1973). The organ-
specificity of ferritin in human and horse liver and spleen. Biochem. J. 131, 51-59.CRICHTON, R. R., PONCE-ORTIZ, Y., KOCH, M. H., PARFAIT, R. & STUHRMANN, H. B. (1978).
Isolation and characterization of phytoferritin from pea {Pisum sativum) and lentil {Lensesculenta). Biochem. J. 171, 349-356.
DAVID, C H . N. & EASTERBROOK, K. (1971). Ferritin in the fungus Phycomyces. J. Cell Biol. 48,15-28.
DESCHIENS, R., BERTRAND, D. & MOLINARI, V. (1957). Capacity d'accumulation de certainsmdtaux par les mollusques de la famille des Planorbides. C. r. Sianc. Soc. Biol. 151, 1356-I3S8.
DRYSDALE, J. W., HAZARD, J. T. & RIGHETTI, P. G. (1975). Microheterogeneity in tissue ferri-tins: evidence for hybrid molecules. In Progress in Isoelectric Focusing and Isotachophoresis(ed. P. G. Righetti), pp. 193-203. Amsterdam: North-Holland.
GRANICK, S. (1943). Ferritin. IV. Occurrence and immunological properties of ferritin. J. biol.Chem. 149, 157-167.
HENEINE, I. F., GAZZINELLI, G. & TAFURI, W. L. (1969). Iron metabolism in the snail Biom-phalariaglabrata: uptake, storage and transfer. Comp. Biochem. Physiol. 28, 391-399.
JOHNSON, C H . R., ANGEL, C H . R. & ERICKSON, D. G. (1962). The uptake, distribution andexcretion of four radionuclides in Australorbis glabratus (Planorbidae). Am.jf. trop. Med. Hyg.11, 855-860.
LEE, S. C. & RICHTER, G. W. (1976). The monomers and oligomers of ferritin and apoferritin:association and dissociation. Biochemistry 15, 65-70.
LOWRY, O. H., ROSEBROUGH, N. J., FARR, A. L. & RANDALL, R. J. (1951). Protein measure-ment with the Folin phenol reagent. .7. biol. Chem. 193, 265-275.
MASSOVER, W. H. (1977). Ultrastructural studies of disrupted ferritin. In Proteins of IronMetabolism (ed. E. B. Brown, P. Ai3en, J. Fielding & R. R. Crichton), pp. 23-30. New York:Grune and Stratton.
MASSOVER, W. H. & COWLEY, J. M. (1975). Ultrastructure of ferritin and apoferritin. InProteins of Iron Storage and Transport in Biochemistry and Medicine (ed. R. R. Crichton),pp. 237-244. Amsterdam: North-Holland.
MAURER, H. R. (1971). Disc Electrophoresis. New York: de Gruyter.MORTON, R. K. (1955). Methods of extraction of enzymes from animal tissues. In Methods in
Enzymology (ed. S. P. Colowick & N.O.Kaplan), vol. 1, pp. 25-51. New York: Academic Press.MUNRO, H. N. & LINDER, M. C. (1978). Ferritin: structure, biosynthesis and role in iron
metabolism. Physiol. Rev. 58, 317-396.NARDI, G., MUZII , E. O. & PUCA, M. (1971). Ferritin in the hepatopancreas of Octopus vulgaris
Lam. Comp. Biochem. Physiol. 40B, 199-205.OTSUKA, SH. , MARUYAMA, H. & LISTOWSKY, I. (1981). Structure, assembly, conformation, and
immunological properties of the two subunit classes of ferritin. Biochemistry 20, 5226-5232.PANYIM, S. & CHALKLEY, R. (1969). High resolution acrylamide gel electrophoresis of histones.
Archs Biochem. Biophys. 130, 337-346.PENDERS, T. J., D E ROOIJ-DIJK, H. H. & LEIJNSE, B. (1968). Rapid isolation of ferritin by means
of ultracentrifugation. Biochim. biophys. Acta 168, 588-590.RICHTER, G. W. (1967). Serological cross-reactions of human, rat and horse ferritins. Expl
molec. Pathol. 6, 96-105.SHINJO, S. & HARRISON, P. M (1979). Artifacts in ferritin isoelectric focusing profiles. FEBS
Lett. 105, 353-356.SVEDBERG, T . (1933). Sedimentation constants, molecular weights and isoelectric points of the
respiratory proteins. .7. biol. Chem. 103, 311-325.TOWE, K. M., LOWENSTAM, H. A. & NESSON, M. H. (1963). Invertebrate ferritin: occurrence
in mollusca. Science, N. Y. 142, 63-64.WEBER, K. & OSBORN, M. (1969). The reliability of molecular weight determinations by
dodecyl sulfate-polyacrylamide gel electrophoresis. J. biol. Chem. 244, 4406—4412.YAGER, C H . M. & HARRY, H. W. (1966). Uptake of heavy metal ions by Taphius glabratus, a
snail host of Schistosoma mansoni. Expl Parasit. 19, 174-182.YARIV, J., KALB, A. J., SPERLING, R., BAUMINGER, E. R., COHEN, S. G. & OFER, S. (1981). The
composition and structure of bacterioferritin of Escherichia coli. Biochem. J. 197, 171-175.
(Received 5 March 1982)





























