Embed Size (px)
Citation preview

Inserting Chromosome 18 into Pancreatic Cancer Cells SwitchesThem to a Dormant Metastatic Phenotype
Liviu P. Lefter, Makoto Sunamura,Toru Furukawa, Kazunori Takeda,Noriko Kotobuki, Mitsuo Oshimura,Seiki Matsuno, and Akira Horii1
Departments of Molecular Pathology [L. P. L., T. F., A. H.] andGastroenterological Surgery [L. P. L., M. S., K. T., S. M.], TohokuUniversity School of Medicine, Sendai 980-8575, Japan, andDepartment of Cell Technology, Tottori University School ofMedicine, Yonago 683-8503, Japan [N. K., M. O.]
ABSTRACTWe demonstrated previously that restoration of chro-
mosome 18 suppressed growth of pancreatic cancer cells invitro, as well as that of tumors inoculated into nude mice. Wealso demonstrated that loss of 18q was associated with poorprognosis. Hence there is the possibility that the 18q armharbors a gene(s) implicated in tumor progression and/ormetastasis. In this study, we evaluated the effect of restoringchromosome 18 on metastasis in a few human pancreaticcancer cell lines with and without inactivation of SMAD4.After microcell-mediated chromosome 18 transfer, hybridcells showed more than a 10-fold weaker metastatic abilitythan corresponding parental cells; mice injected with 1.25 �106/250 �l hybrid clones via tail vein had less than one-tenthof the number of macroscopic metastases in the lung whencompared with the control cells. Microscopic examinationconfirmed the decrease in the number of metastatic lesions.After inoculation of hybrid cells, more than 80% of thehigh-power fields showed no micrometastases, contrastingwith their abundance after using the parental cells. Hybridcells restored maspin expression irrespective of SMAD4 sta-tus in corresponding parental cells. On the other hand,significantly lower vascular endothelial growth factor andmatrix metalloproteinase 2 secretion was observed by mea-suring levels in the conditioned media (CM); the averageswere 22% and 20%, respectively. Angiogenesis assays using
in vivo Matrigel plugs demonstrated that less neovascular-ization was observed in nude mice with hybrid cells thanwith corresponding parental cells. When cells were treatedwith CM from hybrids, the migration of human umbilicalvascular endothelial cells was decreased, but it was partiallyrestored with anti-vascular endothelial growth factor neu-tralizing antibody, as compared with CM from parentalcells. These data represent the first functional evidence sug-gesting that chromosome 18q encodes a gene that stronglysuppresses metastatic activity, possibly through dormancy.
INTRODUCTIONPancreatic cancer is one of the worst diseases worldwide in
terms of prognosis; its mean 5-year survival rate is �5% (1).The mortality rate virtually equals its incidence rate; the reasonsfor this biological aggressiveness include the latency of earlysymptoms and a lack of efficient detection methods at thecurable stages. Thus, acquisition of efficient approaches andmarkers able to accurately detect the earliest stages of pancreaticcancer should be prioritized.
The molecular pathology of pancreatic carcinogenesis is char-acterized by a broad spectrum of mutations in various genes andchromosomal alterations; some of these seem to be specific. Theoncogene KRAS2 and tumor suppressor genes TP53, MTS1, andSMAD4 are mutated in many pancreatic cancers (2–4). Variouschromosomal gains (5p, 7p, 8q, 17q, and 20q) and losses (1p, 6q,9p, 12q, 17p, and 18q) have also been observed in cytogenetic andallelotype studies (5–9). Among these foci for genetic imbalances,loss of 18q is an early event in pancreatic carcinogenesis (10), andthe restoration of chromosome 18 can suppress the growth ofpancreatic cancer cells in vitro (11). Furthermore, LOH2 of 18q isassociated with a poor prognosis (12). LOH of 18q is a commonevent in over 90% of pancreatic carcinomas; only 50% of them arecharacterized by a biallelic inactivation of the SMAD4 gene (4).Although 18q LOH is frequent in intraductal papillary mucinoustumors, one of the premalignant lesions of the pancreas, theSMAD4 protein is expressed in these tumors (13). Moreover, lossof SMAD4 expression occurs biologically later in the neoplasticprogression that leads to the development of infiltrating pancreaticcancer, at the stage of histologically and clinically recognizablecarcinoma (14).
Metastasis is one of the most important factors in poorprognosis. The regulatory genes involved can be broadly cate-
Received 10/28/02; revised 5/30/03; accepted 6/24/03.The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.This work was supported by Japanese Ministries of Education, Culture,Sports, Science and Technology, and Health, Labor and Welfare, Ve-hicle Racing Commemorative Foundation, and Foundation for Promo-tion of Cancer Research in Japan. Part of this article reflects datapresented as a poster at the AACR Annual Meeting in San Francisco,California, on April 6–10, 2002. L. P. L. is a recipient of an AACR-ITOEN, Ltd. Scholar-in-Training Award (http://www.aacr.org/1607a.asp).1 To whom requests for reprints should be addressed, at Department ofMolecular Pathology, Tohoku University School of Medicine, 2-1Seiryo-machi, Aoba-ku, Sendai 980-8575, Japan. Phone: 81-22-717-8042; Fax: 81-22-717-8047; E-mail: [email protected].
2 The abbreviations used are: LOH, loss of heterozygosity; CM, condi-tioned media; MMCT, microcell-mediated chromosome transfer;VEGF, vascular endothelial growth factor; MMP, matrix metallopro-teinase; HUVEC, human umbilical vascular endothelial cell; HPF, high-power field; PECAM-1, platelet/endothelial cell adhesion molecule 1;RT-PCR, reverse transcription-PCR; ATCC, American Type CultureCollection; bFGF, basic fibroblast growth factor; AEc, aminoethyl car-bazole; uPA, urokinase plasminogen activator.
5044 Vol. 9, 5044–5052, October 15, 2003 Clinical Cancer Research
Cancer Research. on January 28, 2020. © 2003 American Association forclincancerres.aacrjournals.org Downloaded from

gorized as either metastasis promoting or metastasis suppress-ing. Because chromosome 18 has been proven to harbor acluster of candidate tumor suppressor and metastasis suppressorgenes, including SMAD2, SMAD4, DCC, SERPINB5 (maspin),and PAI-2, and because poor prognosis is associated with 18qLOH (12), it is of great interest to examine whether genes onchromosome 18 play roles in metastatic processes. Therefore,we herein compared the metastatic ability of pancreatic cancercells after restoration of chromosome 18 with that of theircorresponding parental cells.
MATERIALS AND METHODSPancreatic Cancer Cell Lines. The pancreatic cancer
cell lines used in the present study were PCI-35, BxPC3, Panc-1,and MIAPaCa2. PCI-35 was a generous gift from Dr. HiroshiIshikura (Hokkaido University School of Medicine), and theother three were purchased from ATCC (Manassas, VA).MRC-5 normal human fibroblast cells, HUVECs (ATCC), andpancreatic cancer cells were cultured according to the protocolsof the suppliers and were already well characterized mutation-ally (15). For each cell line, five stable hybrids containing anormal copy of chromosome 18 were established [PCI-35H(18)-1through PCI-35H(18)-5, BxPC3H(18)-1 through BxPC3H(18)-5,Panc-1H(18)-1 through Panc-1H(18)-5, and MIAPaCa2H(18)-1through MIAPaCa2H(18)-5, respectively]. These cells weregenerated by the MMCT technique described in our previousstudy (11) and grown in medium containing 400 �g/ml G418.The A9H(18) mouse fibroblast cell line carrying a single copyof human chromosome 18 tagged with an integrated neomycin-resistant gene was maintained in the same selective medium.
Analysis of Metastatic Ability. To estimate the meta-static ability of the hybrids, we used a lung colonization model(16). Briefly, parental and hybrid cells were prepared as single-cell suspensions in sterile PBS at a concentration of 5 � 106
cells/ml, and a volume of 250 �l (1.25 � 106 cells; viability of95% as determined by trypan blue exclusion) was injected i.v.via the tail vein into 8-week-old male athymic nude mice(BALB/c-�/�; purchased from Japan Clea Inc., Tokyo, Japan).Animals were sacrificed on a day between days 30 and 35, whenthe majority of the control mice became moribund. The leftlungs were snap frozen and used for molecular biological anal-ysis, and the right lungs were fixed in formalin overnight at 4°C.The white surface metastatic tumors were counted, and micro-scopic studies were also performed. This study was approved bythe Ethical Committee of the Tohoku University School ofMedicine. All of the animal experiments were performed ac-cording to Tohoku University Institutional and NIH guidelines.
Immunohistochemical Analysis. Right lungs were ex-cised, fixed in formalin overnight, sectioned discontinuouslyinto five portions, and immunostained with mouse antihumancytokeratin monoclonal antibody (Cosmo Bio, Tokyo, Japan)developed with the AEC system (Zymed Immunomouse Kit). Atleast 100 random HPFs were microscopically examined. Nudemice tumors generated in our previous study (11) as well as 10primary human pancreatic tumors were immunostained usinggoat antihuman maspin monoclonal antibody (Santa Cruz Bio-technology, Santa Cruz, CA) as described previously (17). Im-munohistochemical staining of nude mice for frozen tumors
with anti-PECAM-1 antibody (clone MEC13.3; PharMingen,San Diego, CA) was performed as described previously (18).
Microsatellite Analysis. To test whether or not the in-troduced chromosome 18 was retained in each cell line, weperformed microsatellite analyses by methods described previ-ously (19). Genomic DNAs from the A9H(18) and each parentalcell line and its injected hybrid and the corresponding nude micelung metastatic tumors were analyzed with a panel containingseven highly polymorphic microsatellite markers: D18S1104;D18S463; D18S72; D18S35; D18S1144; D18S483; andD18S58. For each marker, PCR amplification was carried out atleast twice. Nucleotide sequences and conditions for PCR havebeen described previously (11).
RT-PCR. RT-PCR was performed according to meth-ods described previously (20). Primers and conditions used forRT-PCR have been described elsewhere (11, 20, 21). Ten pri-mary human pancreatic cancers as well as their correspondingnormal pancreatic tissues were also used. These tissues wereobtained at Tohoku University Hospital with informed consent.Tissue sections from all patients were reviewed by a board-certificated pathologist (T. F.), and the diagnosis was recon-firmed histologically in all specimens.
Southern Blot Analysis. The SERPINB5 RT-PCR prod-ucts were run on a 3% agarose gel, transferred to a nylonmembrane, and hybridized with a 32P-labeled SERPINB5-specific probe (22). Hybridization procedures were carried outas described previously (23). Each blot was visualized andquantified by the BAS 1500 and Image Gauge 3.3 software(FUJI Photo Film Co., Ltd., Minamiashigara, Japan).
Western Blot Analysis. Western blot analysis was per-formed as described previously (24). A total lysate of humannormal fibroblast MRC-5 was used as the control. CM wereobtained, stored, and analyzed as described previously (25).Antibodies used were goat antihuman MMP2 and MMP9 (SantaCruz Biotechnology) and rabbit antihuman VEGF (Sigma, St.Louis, MO). The relative intensities of signals were analyzedusing the Luminescent Image Analyzer LAS-1000 Plus andImage Gauge 3.3 software (FUJI Photo Film Co., Ltd.).
Matrigel Plug Assays. Each Matrigel plug assay wascarried out by abdominal midline s.c. injection of a 500-�gMatrigel plug alone (negative control) or containing 25% CM.Plugs were removed after 12 days, fixed in formalin, embeddedin paraffin, and stained with H&E as described previously (26).The Matrigel plugs and nude mice tumor vascularization werequantitated by NIH 1.62 software. VEGF concentrations in CMwere quantitated using a VEGF ELISA kit (R&D Systems,Minneapolis, MN) according to the manufacturer’s instructions.
Invasion and Migration Assay. The effects of maspinon the invasion through Matrigel and motility of pancreaticcancer cell lines were analyzed using either a simple modifiedBoyden chamber or one precoated with Matrigel growth factor(Becton Dickinson Labware, Franklin Lakes, NJ) reduced asdescribed previously (25). Five thousand cells, serum-starvedfor 3 h, were seeded in the serum-free medium into the uppercompartment. In the lower compartment of the chamber, mediasupplemented with 10% fetal bovine serum with and withoutfibronectin (5 �g/ml) were added. Antibodies were diluted incell culture medium and added to the lower compartment inconcentrations ranging from 10 to 30 �g/ml. Endothelial cell
5045Clinical Cancer Research
Cancer Research. on January 28, 2020. © 2003 American Association forclincancerres.aacrjournals.org Downloaded from

migration was assayed in 24-well Transwell plates (8.0-�mpores; Costar, Cambridge, MA) using HUVECs purchased fromATCC as described previously (26). CM from the parental cellsor hybrids or CM neutralized with anti-VEGF polyclonal anti-bodies were placed in the lower chamber of the wells. Serum-free medium was used as a negative control. Cell numbers werecounted, averaged, and expressed as the number of migratedcells/HPF. All quantifications were performed in triplicate.
Statistical Analysis. All experiments were performed induplicates or triplicates as indicated. A two-tailed Student’s ttest computed by GraphPad Prism 3.0 software (GraphPad Soft-ware Inc., San Diego, CA) was used to determine the statisticalsignificance of measured differences. The level of significancewas established at P � 0.05.
RESULTSFirst, a panel consisting of seven selected highly polymor-
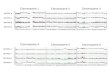
phic microsatellite markers was used to monitor whether or notthe introduced chromosome 18 remained in each hybrid cellline. Genomic DNAs from the parental cell line and its derivedhybrid, A9H(18), were used for PCR. Typical examples areshown in Fig. 1. A complete copy or at least a great majority ofhuman chromosome 18 was transferred and maintained in eachhybrid cell; the band originating from A9H(18) was seen in theDNA of the hybrid cell line in each case.
Once the presence of the introduced chromosome 18 in thehybrids was verified, we estimated the metastatic ability of thehybrids using a lung colonization model. As shown in Table 1,we recorded a significantly decreased number of surface metas-tases in mice lungs injected with hybrids when compared withthose injected with parental cells. These results demonstrate thatthe introduced chromosome 18 provided an important factor thatreduces metastatic activity. Moreover, hybrid cell-injected ani-mals developed a significantly lower number of micrometasta-ses (Table 2). Furthermore, in lungs showing no metastases onsurface examination, we recorded few detectable micrometasta-ses; some of them appeared to be dormant and were not easy todetect. To confirm that these “metastatic lesions” had originatedfrom injected cells, we checked these samples by microsatelliteanalysis and immunostaining; signals for human-specific DNAsequences (see Fig. 1, Lanes M) and human-specific cytokeratin(Fig. 2) were present.
In our previous work, we demonstrated growth suppressioninduced by introduction of chromosome 18 into these pancreaticcancer cell lines (11). There were at least two possibilities for thereduction of metastasis in hybrid cells: one was simply suppressionof cell growth in mice lungs, and the other was involvement offactors regulating the metastatic ability; angiogenesis is one suchfactor. One of the possible targets located on 18q and involved inangiogenesis is SERPINB5 (maspin), so we examined its levelsfirst to consider their functional importance in metastasis. Expres-sion of SERPINB5 mRNA was observed initially only in parentalcells having inactivated SMAD4, PCI-35 and BXPC3, respectively(Fig. 3, Lanes 5 and 7). In sharp contrast, maspin mRNA wasundetectable in SMAD4wt cells but was expressed in the hybridscontaining an extra copy of chromosome 18, namely, Panc-1H(18)and MIAPaCa2H(18) (Fig. 3, A and B). Furthermore, low expres-sion of maspin was observed in 8 of 10 normal human pancreatictissues by RT-PCR and confirmed by Southern hybridization (seeFig. 4, A and B). In contrast, overexpression of maspin was ob-served in all 10 of the human pancreatic primary tumor specimensstudied (see Fig. 4C). Additionally, a subclavicular skin metastasisdetected in one of the patients showed abundant maspin expression(Fig. 4, D and E).
Functional assessment of maspin is depicted in Fig. 5. Basalmigration with serum-free medium in the lower chamber was thesame for both parental and hybrid cells. On the other hand, stim-ulated migration of hybrid clones toward bFGF was significantlyenhanced upon maspin antibody treatment but was seen to a lesser
Table 1 Lung surface metastases generated by parental cells andtheir corresponding hybrid clonesa
Cell lines injectedMean no. of
loci/lung � SD
Mice withoutmetastases/total number
of mice
PCI-35 56.0 � 2.8 0/4PCI-35H(18) 4.6 � 1.3 4/15Panc-1 42.0 � 1.4 0/4Panc-1H(18) 3.6 � 1.8 5/15BxPC3 54.0 � 4.2 0/15BxPC3H(18) 7.6 � 0.9 2/15a P � 0.001 calculated by t test. Macroscopic metastases were
considered to be �1 mm in diameter.
Fig. 1 Representative results of microsatellite analysis are shown. Autoradiograms showed restoration of heterozygosity for each hybrid and the resultantlung metastatic tumor. The primer set for amplification of each microsatellite marker was designed based on nucleotide sequence provided from the GenomeDatabase (http://gdbwww.gdb.org). Lane P, parental pancreatic cancer cell line Panc-1; Lane H, their hybrids, Panc-1(18); Lane M, nude mice lung metastatictumors after injection of Panc-1(18); A9H(18), the donor cell for chromosome 18 in MMCT experiment. Arrows indicate bands corresponding to thechromosome 18 originating from A9H(18). The cytogenetic position of each marker is indicated in parentheses below each marker.
5046 Chromosome 18 Encodes Metastatic Suppressor Activity
Cancer Research. on January 28, 2020. © 2003 American Association forclincancerres.aacrjournals.org Downloaded from

extent in parental cells. This property was only seen in the hybridsgenerated by parental cells originally lacking maspin expression(Fig. 5A), and no effect was seen, regardless of the titer of antibodyused, in hybrids whose parental cells that were expressing maspin(Fig. 5B). In contrast, the invasiveness of hybrids was significantlyreduced and then restored upon the same treatment, regardless ofthe initial status of maspin expression (Fig. 5D). However, neithermotility nor invasion of hybrids was restored to the levels ofuntreated parental cells used as controls. This result suggests thatmaspin expression can explain only part of the reduction of hybridmotility and invasiveness.
As some of most important steps in the establishment ofmetastasis cascade, angiogenesis and invasion were studied byfocusing on activities of some soluble factors such as VEGF orMMPs. When cells were treated with CM from hybrids, themigration of HUVECs was decreased, but it was partially re-stored with anti-VEGF neutralizing antibody, as compared withCM from parental cells (Fig. 5C). Significant decreases in
secretion of VEGF (average, 20.5%; see Fig. 6, A and B) andMMP2 (average, 24.7%; Fig. 6C) in the culture media fromhybrids were observed when the levels were compared withthose of the corresponding parental cells. A relatively similarrate of reduction in migrated endothelial cells was observed inresponse to either antihuman VEGF treatment or Matrigel plugs.These results were confirmed by microvessel assessment intumors that developed in nude mice (Fig. 7); a significantreduction in the hybrid cells was seen when compared withparental cells [by 22 � 2.75% for PCI-35H(18)].
DISCUSSIONOur recent studies have shown that restoration of chromo-
some 18 status suppressed the in vitro growth of human pan-creatic cancer cells and that in vivo growth was also suppressedin nude mice (11). In the present study, we observed a signifi-cant suppression of metastatic ability in pancreatic cancer cells
Table 2 Comparison of metastatic abilities between parental and hybrid cell linesa
Cell lines No metastases Microscopic metastases Macroscopic metastasesb No. of animals
PCI-35 4.9 � 1.9 24.6 � 3.1 77.9 � 9.1 4PCI-35H(18) 82.4 � 7.4 3.9 � 1.1 6.9 � 2.1 15Panc-1 6.1 � 1.9 32.8 � 6.7 72.6 � 5.6 4Panc-1H(18) 78.4 � 9.7 2.8 � 0.91 5.5 � 1.9 15BxPC3 5.3 � 1.1 33.5 � 3.1 69.8 � 6.2 4BxPC3H(18) 73.5 � 7.6 4.1 � 1.2 5.9 � 1.6 15
a Results represent the mean of 100 HPF � SD. Experiments were performed in triplicate, and five discontinuous lung sections were analyzed(�100 magnification) in each experiment.
b The threshold for macroscopic metastases was considered to be �300 cells/positive stained focus; P � 0.001.
Fig. 2 Histological examina-tion of nude mice lungs. Five dis-continuous 4-�m sections wereimmunostained with mouse anti-human cytokeratin monoclonalantibody and developed with theAEC system. Representative as-pects of lungs after injection ofPCI-35 (A and B) and PCI-35H(18) (C and D) are shown atthe indicated magnifications. Ar-rows indicate examples of mac-roscopic metastases (�300 cells/positive stained focus).
5047Clinical Cancer Research
Cancer Research. on January 28, 2020. © 2003 American Association forclincancerres.aacrjournals.org Downloaded from

in which we restored a normal copy of chromosome 18. We alsorecorded a significant suppression in the number of surfacemetastases developing in hybrid cell-injected mice as comparedwith parental cell-injected mice; at least one of every hybridclone had no countable macroscopic metastases. Furthermore,
microscopic examination showed that animals injected withhybrid cells developed a significantly lower number of micro-metastases and only very rare macrometastases when comparedwith those injected with parental cells. Moreover, in lungsshowing no metastases on surface examination, we recorded
Fig. 3 Expression of SERPINB5 (maspin)in pancreatic cancer cell lines was analyzedby RT-PCR (A) and Southern blotting (B).Expression of SMAD4 transcripts by RT-PCR is shown in C. Lane 1, Panc-1; Lane 2,Panc-1H(18); Lane 3, MIAPaCa2; Lane 4,MIAPaCa2H(18); Lane 5, PCI-35; Lane 6,PCI-35H(18); Lane 7, BxPC3; Lane 8,BxPC3H(18); Lane 9, normal colon (A),PCR-amplified maspin cDNA (B), or fibro-blast cell line MRC-5 (C).
Fig. 4 Expression of maspin in 10 normal and 10 cancerous tissues by RT-PCR (A and C). Results in B represent Southern hybridization in 10pancreatic normal tissues. Lanes 1–10 are normal (A) or primary tumor (C) specimens. Lane 11 in C is a positive control, MRC-5. Expression ofmaspin is shown by immunostaining with maspin monoclonal antibody in a skin metastatic lesion in one of the pancreatic cancer patients (D, �100;E, �200).
5048 Chromosome 18 Encodes Metastatic Suppressor Activity
Cancer Research. on January 28, 2020. © 2003 American Association forclincancerres.aacrjournals.org Downloaded from

only a few, hardly detectable micrometastases that appeared tobe dormant. However, the dramatic suppression of macrometas-tases and the presence of apparently dormant micrometastases inhybrid clone-injected animals may simply suggest importantsuppression in cell proliferation.
In an attempt to gain more insight into this effect, wepreviously analyzed the expression of known genes residing onchromosome 18 and related to the metastasis process. MMCTcould be approximated, at least theoretically, with transfectionof a panel encompassing a single allele of each gene residing onthe chromosome 18. Among these genes, only SERPINB5(maspin) has been shown to functionally suppress metastasis ofbreast cancer cells in an in vivo model (27). Interestingly, in theparental cells, maspin expression showed a tendency towardinverse association with a simultaneous inactivation of SMAD4;specifically, only SMAD4-null cells expressed maspin at levelssimilar to those found in human tumors; no expression wasdetected in SMAD4wt cells. Maspin and SMAD4 are closelylocated on chromosome 18 (18q21.1 and 18q21.3, respectively)and share a critical region for pancreatic cancer. Despite theabundance of data concerning SMAD4, there are no detailedstudies describing the frequency of maspin inactivation or itsimplications for pancreatic cancer. To date, LOH of 18q is acommon event in over 90% of pancreatic carcinomas, but only
50% of them are characterized by a biallelic inactivation of theSMAD4 gene (4). Inoue et al. (13) reported that SMAD4 proteinis expressed in virtually all intraductal papillary mucinous tu-mors, suggesting inactivation of SMAD4 as a rather late event inpancreatic tumorigenesis. Based on our findings, we could hy-pothesize that SMAD4 and maspin are inactivated or rathermutated at different stages of pancreatic tumorigenesis. How-ever, to elucidate this attractive aspect, additional studies will benecessary. A low to moderate level of maspin expression wasobserved in the majority of normal human pancreatic tissues byRT-PCR. Maass et al. (17) found by Northern blotting that themRNA level of SERPINB5 in normal pancreas tissue is low.This could be due to a low level of SERPINB5 or to a highersensitivity of the RT-PCR method. Additionally, rather highlevels of SERPINB5 expression have been observed in primarypancreatic tumors. In fact, the latter finding makes SERPINB5an unlikely target of chromosome 18 loss. To date, it has beensuggested that maspin may be useful in separating ductal ade-nocarcinoma from acinar cell carcinoma, pancreatic endocrinetumor, solid pseudopapillary tumor, and chronic pancreatitis(28). Overall, there is a completely different pattern of maspinexpression in pancreatic tumorigenesis than in breast and pros-tate cancers. This paradoxical expression of maspin has beenreported recently by Sood et al. (29) in ovarian carcinoma.
Fig. 5 A and B, motility assays. The indicated pancreatic cancer cells and their hybrids were plated in the upper chambers and allowed to migratefor 8 h toward either fetal bovine serum-free medium or medium containing 10 �g/ml bFGF (basal and stimulated migration, respectively).Antimaspin antibodies were diluted in cell culture medium and added to the lower wells to final concentrations ranging from 10 to 20 �g/ml, asindicated. C, endothelial cell migration assay. HUVECs were allowed to migrate toward CM from either parental cells, hybrids, or CM neutralizedwith anti-VEGF polyclonal antibodies placed in the lower chambers of the wells. Normal human serum medium (IG) was used as a negative control.D, invasion assays. The indicated cells were allowed to invade through a Matrigel reconstituted membrane for 18 h. Antibodies were diluted in cellculture medium and added to the lower wells at 20 �g/ml. Normal human serum medium was used as a negative control. Migrated or invaded cellswere counted, averaged, and expressed either as migrated cells/HPF (�100 magnification) or as relative invasion compared with control cells (D).Data represent the mean � SD of at least three independent experiments. �, P � 0.05; §, P � 0.05.
5049Clinical Cancer Research
Cancer Research. on January 28, 2020. © 2003 American Association forclincancerres.aacrjournals.org Downloaded from

Maspin overexpression was significantly associated with a hightumor grade and a shorter duration of overall survival. In ourstudy, however, functional assessments of maspin expressioncannot fully explain the observed effect on suppression ofmetastatic abilities. It is possible that the maspin expressed inthese tumors may be inactive (29). Degradation and penetrationof the extracellular matrix is a hallmark of tumor invasion andmetastasis (30). In our study, stimulated migration was found tobe significantly enhanced by maspin antibody treatment, but toa lesser extent than that seen with parental cells. This lattereffect can be explained only by the hypothesis that the hybridsgenerated by the parental cells originally lacked maspin expres-sion, and the phenomenon suggests a dose-independent mech-anism. Notably, the invasiveness of hybrids was significantlyreduced and then restored upon the same treatment, regardlessof the initial status of maspin expression in the parental cells.However, neither the motility nor the invasion of hybrids couldbe restored to the levels of untreated parental cells used ascontrols. These results suggest that maspin expression can ex-plain only a part of the reduction of hybrid motility and inva-siveness.
Angiogenesis and invasion, both required steps in the met-astatic process, are under the control of growth factors such asVEGF and proteinases, especially those from MMP members(30, 31). The aggressive phenotype of pancreatic carcinomamay arise from overexpression of MMP2 (32). VEGF maypromote the distribution of metastases, leading to early cancerrecurrence and the poor outcome of pancreatic cancer (33).Therefore, we examined the expression of these factors andshowed a significant reduction in VEGF and MMP2 secretion in
the culture media from hybrids as compared with the levelsproduced by parental cells. The differences among these cells intheir expression of VEGF were more pronounced when theexpression was measured by ELISA than when measured byWestern blotting. The rate of protein secretion is probably morerelevant to the rate of tumor neovascularization than the intra-cellular concentration of protein because angiogenesis is in-duced by the binding of protein to endothelial cell receptors(34). However, no significant differences were found among thecells for secretion of MMP1, MMP9, bFGF, or u-PA (data notshown). Furthermore, relatively similar reduction rates wereobserved in the endothelial cells that migrated through theantihuman VEGF-treated CM and into Matrigel plugs; thesedata suggest that VEGF status is responsible for the majority ofchanges in the angiogenic phenotype throughout the hybridcells. These results were confirmed by PECAM-1 immunoreac-tivity in the nude mice tumors, which showed a significantreduction in the hybrid cells. This fact is concordant with aprevious study showing that SMAD4 restoration in pancreaticcancer cells reduces angiogenesis rates through the down-regu-lation of VEGF expression (35). Taken together, these resultssuggested that the dramatic reduction in the metastatic abilitiesof hybrid cells containing a normal copy of chromosome 18 isonly partially attributable to the changes demonstrated in VEGFor MMP2 secretion rates.
To date, MMCT has been proven to be useful for providingfunctional evidence of the chromosome location of tumor ormetastasis suppressor genes in a variety of cancers, includingpancreatic cancer (11), Nijmegen breaking syndrome (36), andprostate cancer (37). As stated earlier, a whole chromosome
Fig. 6 Estimation of VEGF (A and B) andMMP2 (C) secretion in CM. VEGF was estimatedin CM by ELISA, and the results are presented(A). A total of 106 cells were used. Results ofWestern blotting analyses in either total cell lysate(B) or that of CM (C) are shown. In each lane, 20�g of total cell lysate were loaded, and �-actinwas used as the control. Lane 1, PCI-35; Lane 2,PCI-35H(18); Lane 3, Panc-1; Lane 4, Panc-1H(18); Lane 5, BxPC3; Lane 6, BxPC3H(18);Lane 7, MRC-5 normal fibroblasts (B) or serum-free medium (C). The relative intensity of signalswas analyzed using the Luminescent Image Ana-lyzer LAS-1000 Plus and imaging software Gauge3.3 (FUJI Photo Film Co., Ltd.) and is indicated asnumbers corresponding to each band.
5050 Chromosome 18 Encodes Metastatic Suppressor Activity
Cancer Research. on January 28, 2020. © 2003 American Association forclincancerres.aacrjournals.org Downloaded from

transfer raises at least two presumable effects: (a) direct effectsof the known or unknown genes located on chromosome 18; and(b) indirect effects through possible interactions among thetransferred chromosome and the other genes. Although, in thissetting, these effects could not be clearly delineated, it is con-ceivable that the introduction of an extra copy of chromosome18 confers a less aggressive metastatic phenotype to the pan-creatic cancer cells. In other words, the metastatic inhibitionencoded by 18q can explain the dormant status of the hybridsand, in turn, the presence of micrometastases unable to formmacrometastases.
In summary, metastasis was suppressed when an intactchromosome 18 was transferred into human pancreatic cancercells. This study provides the first functional evidence that oneor more metastasis suppressor gene(s) is located on chromo-some 18. Although this study clearly implicates the importantrole of gene(s) on chromosome 18, the precise subchromosomallocalization of the metastasis suppressor gene(s) is still an openquestion. In the absence of spontaneous revertant hybrids, theprecise localization and identification of a putative metastasissuppressor gene will require further and additional efforts.
ACKNOWLEDGMENTSWe are grateful to Dr. Barbara Lee Smith Pierce at the University
of Maryland University College for editorial work in the preparation ofthe manuscript. We are also grateful to Shinichi Fukushige, EmikoKondo, Hiroko Fujimura, and Emiko Shibuya at Tohoku UniversitySchool of Medicine for their excellent technical assistance and advice,and to Drs. Christian Partensky and Jean-Yves Scoazec at the EdouardHerriot Hospital, Lyon, France, for many helpful discussions.
REFERENCES1. Parker, S. L., Tong, T., Bolden, S., and Wingo, P. A. Cancerstatistics, 1997. CA Cancer J. Clin., 47: 5–27, 1997.
2. Caldas, C., Hahn, S. A., da Costa, L. T., Redston, M. S., Schutte, M.,Seymour, A. B., Weinstein, C. L., Hruban, R. H., Yeo, C. J., and Kern,S. E. Frequent somatic mutations and homozygous deletions of the p16(MTS1) gene in pancreatic adenocarcinoma. Nat. Genet., 8: 27–32,1994.3. Barton, C. M., Staddon, S. L., Hughes, C. M., Hall, P. A., O’Sullivan,C., Kloeppel, G., Theis, B., Russell, R. C. G., Neoptolemos, J., Wil-liamson, R. C. N., Lane, D. P., and Lemoine, N. R. Abnormalities of thep53 tumour suppressor gene in human pancreatic cancer. Br. J. Cancer,64: 1076–1082, 1991.
Fig. 7 Estimation of angiogenesis by Matrigel plug assay (A) was carried out by s.c. injection of a 500-�g Matrigel plug alone (negative control)or containing 25% of the indicated CM. Plugs were removed after 12 days, fixed in formalin, embedded in paraffin, and stained with H&E.Immunohistochemical staining (B) of the indicated frozen tumor sections with anti-PECAM-1 antibody (clone MEC13.3; PharMingen) was performedas described previously. Matrigel plug and nude mice tumor vascularization was quantitated by NIH 1.62 software, and data are presented as the meansof relative microvessel structure areas (in pixels) � SD.
5051Clinical Cancer Research
Cancer Research. on January 28, 2020. © 2003 American Association forclincancerres.aacrjournals.org Downloaded from

4. Hahn, S. A., Schutte, M., Hoque, A. T., Moskaluk, C. A., da Costa,L. T., Rozenblum, E., Weinstein, C. L., Fischer, A., Yeo, C. J., Hruban,R. H., and Kern, S. E. DPC4, a candidate tumor suppressor gene athuman chromosome 18q21.1. Science (Wash. DC), 271: 350–353,1996.
5. Seymour, A. B., Hruban, R. H., Redston, M., Caldas, C., Powell,S. M., Kinzler, K. W., Yeo, C. J., and Kern, S. E. Allelotype ofpancreatic adenocarcinoma. Cancer Res., 54: 2761–2764, 1994.
6. Griffin, C. A., Hruban, R. H., Morsberger, L. A., Ellingham, T.,Long, P. P., Jaffee, E. M., Hauda, K. M., Bohlander, S. K., and Yeo,C. J. Consistent chromosome abnormalities in adenocarcinoma of thepancreas. Cancer Res., 55: 2394–2399, 1995.
7. Hahn, S. A., Seymour, A. B., Hoque, A. T. M. S., Schutte, M., daCosta, L. T., Redston, M. S., Caldas, C., Weinstein, C. L., Fischer, A.,Yeo, C. J., Hruban, R. H., and Kern, S. E. Allelotype of pancreaticadenocarcinoma using xenograft enrichment. Cancer Res., 55: 4670–4675, 1995.8. Kimura, M., Abe, T., Sunamura, M., Matsuno, S., and Horii, A.Detailed deletion mapping on chromosome arm 12q in human pancre-atic adenocarcinoma: identification of a 1-cM region of common allelicloss. Genes Chromosomes Cancer, 17: 88–93, 1996.9. Fukushige, S., Waldman, F. M., Kimura, M., Abe, T., Furukawa, T.,Sunamura, M., Kobari, M., and Horii, A. Frequent gain of copy numberon the long arm of chromosome 20 in human pancreatic adenocarci-noma. Genes Chromosomes Cancer, 19: 161–169, 1997.10. Fukushige, S., Furukawa, T., Satoh, K., Sunamura, M., Kobari, M.,Koizumi, M., and Horii, A. Loss of chromosome 18q is an early eventin pancreatic ductal tumorigenesis. Cancer Res., 58: 4222–4226, 1998.11. Lefter, L. P., Furukawa, T., Sunamura, M., Duda, D. G., Takeda, K.,Kotobuki, N., Oshimura, M., Matsuno, S., and Horii, A. Suppression ofthe tumorigenic phenotype by chromosome 18 transfer into pancreaticcancer cell lines. Genes Chromosomes Cancer, 34: 234–242, 2002.12. Yatsuoka, T., Sunamura, M., Furukawa, T., Fukushige, S.,Yokoyama, T., Inoue, H., Shibuya, K., Takeda, K., Matsuno, S., andHorii, A. Association of poor prognosis with loss of 12q, 17p, and 18q,and concordant loss of 6q/17p and 12q/18q in human pancreatic ductaladenocarcinoma. Am. J. Gastroenterol., 95: 2080–2085, 2000.13. Inoue, H., Furukawa, T., Sunamura, M., Takeda, K., Matsuno, S.,and Horii, A. Exclusion of SMAD4 mutation as an early genetic changein human pancreatic ductal tumorigenesis. Genes Chromosomes Cancer,31: 295–299, 2001.14. Wilentz, R. E., Iacobuzio-Donahue, C. A., Argani, P., McCarthy,D. M., Parsons, J. L., Yeo, C. J., Kern, S. E., and Hruban, R. H. Loss ofexpression of Dpc4 in pancreatic intraepithelial neoplasia: evidence thatDPC4 inactivation occurs late in neoplastic progression. Cancer Res.,60: 2002–2006, 2000.15. Sun, C., Yamato, T., Furukawa, T., Ohnishi, Y., Kijima, H., andHorii, A. Characterization of the mutations of the K-ras, p53, p16, andSMAD4 genes in 15 human pancreatic cancer cell lines. Oncol. Rep., 8:89–92, 2001.16. Luu, H. H., Zagaja, G. P., Dubauskas, Z., Chen, S. L., Smith, R. C.,Watabe, K., Ichikawa, Y., Ichikawa, T., Davis, E. M., Le Beau, M. M.,and Rinker-Schaeffer, C. W. Identification of a novel metastasis-suppressor region on human chromosome 12. Cancer Res., 58: 3561–3565, 1998.17. Maass, N., Hojo, T., Ueding, M., Luttges, J., Kloppel, G., Jonat, W.,and Nagasaki, K. Expression of the tumor suppressor gene Maspin inhuman pancreatic cancers. Clin. Cancer Res., 7: 812–817, 2001.18. Liao, F., Li, Y., O’Connor, W., Zanetta, L., Bassi, R., Santiago, A.,Overholser, J., Hooper, A., Mignatti, P., Dejana, E., Hicklin, D. J., andBohlen, P. Monoclonal antibody to vascular endothelial-cadherin is apotent inhibitor of angiogenesis, tumor growth, and metastasis. CancerRes., 60: 6805–6810, 2000.19. Kimura, M., Furukawa, T., Abe, T., Yatsuoka, T., Youssef, E. M.,Yokoyama, T., Ouyang, H., Ohnishi, Y., Sunamura, M., Kobari, M.,Matsuno, S., and Horii, A. Identification of two common regions ofallelic loss in chromosome arm 12q in human pancreatic cancer. CancerRes., 58: 2456–2460, 1998.
20. Mori, Y., Shiwaku, H., Fukushige, S., Wakatsuki, S., Sato, M.,Nukiwa, T., and Horii, A. Alternative splicing of hMSH2 in normalhuman tissues. Hum. Genet., 99: 590–595, 1997.21. Li, J. J., Colburn, N. H., and Oberley, L. W. Maspin gene expres-sion in tumor suppression induced by overexpressing manganese-containing superoxide dismutase cDNA in human breast cancer cells.Carcinogenesis (Lond.), 19: 833–839, 1988.22. Merrie, A. E., Yun, K., Gunn, J., Phillips, L. V., and McCall, J. L.Analysis of potential markers for detection of submicroscopic lymphnode metastases in breast cancer. Br. J. Cancer, 80: 2019–2024, 1999.23. Hiyama, E., Yokoyama, T., Ichikawa, T., Ishii, T., and Hiyama, K.N-myc gene amplification and other prognostic associated factors inneuroblastoma. J. Pediatr. Surg., 25: 1095–1099, 1990.24. Kondo, E., Horii, A., and Fukushige, S. The human PMS2L pro-teins do not interact with hMLH1, a major DNA mismatch repairprotein. J. Biochem. (Tokyo), 125: 818–825, 1999.25. Attiga, F. A., Fernandez, P. M., Weeraratna, A. T., Manyak, M. J.,and Patierno, S. R. Inhibitors of prostaglandin synthesis inhibit humanprostate tumor cell invasiveness and reduce the release of matrix met-alloproteinases. Cancer Res., 60: 4629–4637, 2000.26. Zhang, L., Yu, D., Hu, M., Xiong, S., Lang, A., Ellis, L. M., andPollock, R. E. Wild-type p53 suppresses angiogenesis in human leiomy-osarcoma and synovial sarcoma by transcriptional suppression of vas-cular endothelial growth factor expression. Cancer Res., 60: 3655–3661,2000.27. Shi, H. Y., Zhang, W., Liang, R., Abraham, S., Kittrell, F. S.,Medina, D., and Zhang, M. Blocking tumor growth, invasion, andmetastasis by maspin in a syngeneic breast cancer model. Cancer Res.,61: 6945–6951, 2001.28. Oh, Y. L., Song, S. Y., and Ahn, G. Expression of maspin inpancreatic neoplasms: application of maspin immunohistochemistry tothe differential diagnosis. Appl. Immunohistochem. Mol. Morphol., 10:62–66, 2002.29. Sood, A. K., Fletcher, M. S., Gruman, L. M., Coffin, J. E., Jabbari,S., Khalkhali-Ellis, Z., Arbour, N., Seftor, E. A., and Hendrix, M. J. Theparadoxical expression of maspin in ovarian carcinoma. Clin. CancerRes., 8: 2924–2932, 2002.30. Price, J. P., Bonovich, M. T., and Kohn, E. The biochemistry ofcancer dissemination. Crit. Rev. Biochem. Mol. Biol., 32: 175–253,1997.31. Zetter, B. R. Angiogenesis and tumor metastasis. Annu. Rev. Med.,49: 407–424, 1998.32. Bramhall, S. R., Neoptolemos, J. P., Stamp, G. W., and Lemoine,N. R. Imbalance of expression of matrix metalloproteinases (MMPs)and tissue inhibitors of the matrix metalloproteinases (TIMPs) in humanpancreatic carcinoma. J. Pathol., 182: 347–355, 1997.33. Niedergethmann, M., Hildenbrand, R., Wostbrock, B., Hartel, M.,Sturm, J. W., Richter, A., and Post, S. High expression of vascularendothelial growth factor predicts early recurrence and poor prognosisafter curative resection for ductal adenocarcinoma of the pancreas.Pancreas, 25: 122–129, 2002.34. Rofstad, E. K., and Halsør, E. F. Vascular endothelial growth factor,interleukin 8, platelet-derived endothelial cell growth factor, and basicfibroblast growth factor promote angiogenesis and metastasis in humanmelanoma xenografts. Cancer Res., 60: 4932–4938, 2000.35. Schwarte-Waldhoff, I., Volpert, O. V., Bouck, N. P., Sipos, B.,Hahn, S. A., Klein-Scory, S., Luttges, J., Kloppel, G., Graeven, U.,Eilert-Micus, C., Hintelmann, A., and Schmiegel, W. SMAD4/DPC4-mediated tumor suppression through suppression of angiogenesis. Proc.Natl. Acad. Sci. USA, 97: 9624–9629, 2000.36. Matsuura, S., Tauchi, H., Nakamura, A., Kondo, N., Sakamoto, S.,Endo, S., Smeets, D., Solder, B., Belohradsky, B. H., der Kaloustian,V. M., Oshimura, M., Nakamura, Y., and Komatsu, K. Positionalcloning of the gene for Nijmegen breakage syndrome. Nat. Genet., 19:179–181, 1998.37. Padalecki, S. S., Johnson-Pais, T. L., Killary, A. M., and Leach,R. J. Chromosome 18 suppresses the tumorigenicity of prostate cancercells. Genes Chromosomes Cancer, 30: 221–229, 2001.
5052 Chromosome 18 Encodes Metastatic Suppressor Activity
Cancer Research. on January 28, 2020. © 2003 American Association forclincancerres.aacrjournals.org Downloaded from

2003;9:5044-5052. Clin Cancer Res Liviu P. Lefter, Makoto Sunamura, Toru Furukawa, et al. Switches Them to a Dormant Metastatic PhenotypeInserting Chromosome 18 into Pancreatic Cancer Cells
Updated version
http://clincancerres.aacrjournals.org/content/9/13/5044
Access the most recent version of this article at:
Cited articles
http://clincancerres.aacrjournals.org/content/9/13/5044.full#ref-list-1
This article cites 36 articles, 16 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/9/13/5044.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://clincancerres.aacrjournals.org/content/9/13/5044To request permission to re-use all or part of this article, use this link
Cancer Research. on January 28, 2020. © 2003 American Association forclincancerres.aacrjournals.org Downloaded from