Embed Size (px)
Citation preview


GGGEEENNNEEETTTYYYKKKAAA III EEEWWWOOOLLLUUUCCCJJJAAA
ZZZaaadddaaannniiiaaa iii PPPrrrooobbbllleeemmmyyy
KKKooorrrnnneeellliiiaaa PPPooollloookkk KKKaaattteeedddrrraaa GGGeeennneeetttyyykkkiii
UUUnnniiiwwweeerrrsssyyyttteeettt WWWaaarrrmmmiiińńńssskkkooo---MMMaaazzzuuurrrssskkkiii www OOOlllssszzztttyyynnniiieee
Olsztyn 2010

OOOkkk łłłaaadddkkkaaa Projekt: K. Polok, Fotografie: K. Polok Przód: Skan oryginalnego tekstu Grzegorza Mendla opisującego wyniki doświadczeń nad krzyżowaniem grochu oraz segregacja barwy kwiatu w pokoleniu F2 grochu. Fotografia wykonana na poletku doświadczalnym Katedry Genetyki UWM w Tomaszkowie, 2008. Tył: Przykłady mutantów/mutacji u grochu (Pisum sativum), owsa szorstkiego (Avena strigosa) oraz jęczmienia (Hordeum vulgare). Fotografie wykonano na poletku doświadczalnym Katedry Genetyki UWM w Tomaszkowie, 2008, 2009. Mutant grochu clavicula pochodzi z kolekcji Prof. W. Gottschalka i otrzymano go dzięki uprzejmości Prof. Hansa-Jorga Jacobsena z Leibniz Universitat Hannover. Forma MRS jęczmienia, u której pokazano mutację wst, jest wielogenową homozygotą recesywną (Multiple Recessive Stock), otrzymaną w wyniku wielokrotnego krzyżowania (Franckowiak 1986). Mutanty fasciata grochu, ereriferum owsa szorstkiego, tigrina jęczmienia oraz formę jęczmienia z mutacją ości otrzymano w Katedrze Genetyki UWM w wyniku traktowania nasion N-nitrozo-N-metylomocznikiem i azydkiem sodu. Stanowią one zaawansowane pokolenia M6-M7. GGGeeennneeetttyyykkkaaa iii EEEwwwooollluuucccjjjaaa... ZZZaaagggaaadddnnniiieeennniiiaaa iii PPPrrrooobbbllleeemmmyyy Website: http://www.uwm.edu.pl/katgenbiol, e-mail: [email protected], [email protected]
Recenzent: Prof. dr hab. Roman Zielinski
Korekta: Prof. dr hab. Roman Zielinski
Zebranie i pomoc techniczna: Mgr Katarzyna Wołosz

SSSPPPIIISSS TTTRRREEEŚŚŚCCCIII
GGGEEENNNEEETTTYYYKKKAAA
1 Terminologia genetyczna . . . . . . . . . . . . . . . . . . . . . 9
2 Pierwsze Prawo Mendla . . . . . . . . . . . . . . . . . . . . . . 11
3 Drugie Prawo Mendla . . . . . . . . . . . . . . . . . . . . . . . 14
4 Kodominacja . . . . . . . . . . . . . . . . . . . . . . . . . . 21
5 Allele letalne . . . . . . . . . . . . . . . . . . . . . . . . . . 24
6 Allele wielokrotne . . . . . . . . . . . . . . . . . . . . . . . . 27
7 Współdziałanie genów . . . . . . . . . . . . . . . . . . . . . . . 29
8 Geny sprzężone z płcią . . . . . . . . . . . . . . . . . . . . . . 33
9 Geny sprzężone . . . . . . . . . . . . . . . . . . . . . . . . . 37
10 Mapowanie genów . . . . . . . . . . . . . . . . . . . . . . . . 48
EEEWWWOOOLLLUUUCCCJJJAAA
11 O ewolucji… . . . . . . . . . . . . . . . . . . . . . . . . . . 53
12 Genetyka populacyjna . . . . . . . . . . . . . . . . . . . . . . . 58
13 Genetyka ewolucyjna. . . . . . . . . . . . . . . . . . . . . . . . 63

ZZZAAAMMMIIIAAASSSTTT WWWSSSTTTĘĘĘPPPUUU………
“Każda z siedmiu cech mieszańców Pisum albo jest tak bardzo podobna do cechy jednego z dwóch rodziców, że cecha przeciwstawna jest niewidoczna lub też jest tak podobna do cechy jednego z rodziców, że nie można cechy u rodzica i mieszańca rozróżnić. Ten wniosek ma ogromne znaczenie dla zdefiniowania i klasyfikacji form, które występują wśród mieszańców. W dalszych rozważaniach, cechy, które przekazywane są mieszańcom niezmienione lub prawie niezmienione są określone jako dominujące, te zaś, które stają się niewidoczne określane są jako recesywne. Termin recesywny został wybrany ponieważ cechy te są osłabione lub znikają całkowicie u mieszańca, ale ukazują się ponownie jako niezmienione w potomstwie”
Johan Gregor MendelExperiments on Plant Hybrids. 1865
(Tłumaczenie z języka angielskiego: Kornelia Polok)
Geniusz Mendla polegał na umiejętnym zaplanowaniu i starannym przeprowadzeniu doświadczenia, a
następnie analizie wyników przy pomocy metod matematyczno-statystycznych. Dzięki temu stworzył On podstawy
genetyki, ale nade wszystko podał zasady dziedziczenia cech. Zasady te nie uległy zmianie pomimo nowych
odkryć, jakich dostarczyła genetyka molekularna. Co więcej, w dobie zachłyśnięcia się nowoczesnymi
technologiami, okazuje się, że bez podstaw jakie stworzył Mendel nie można przeprowadzić najbardziej
zaawansowanych analiz z zakresu genomiki. Dlatego także i dzisiaj studenci powinni uczyć się od Mendla jak
prawidłowo prowadzić oraz analizować doświadczenia. Bez wykazania dziedziczenia mendlowskiego cechy
wprowadzonej do rośliny metodą inżynierii genetycznej nie można zarejestrować odmiany transgenicznej. Hodowla
roślin czy analiza rodowodów w medycynie od lat czerpią garściami z dwóch prostych twierdzeń o wykluczaniu się
alleli tego samego genu w gametach oraz niezależnego dziedziczenia się genów zlokalizowanych na różnych
chromosomach. Prawa Mendla mają w genetyce taki status jak prawa Newtona w fizyce. Dzięki nim genetyka stała
się pierwszą nauką biologiczną w rozumieniu nauk fizycznych, nauką, w której teoria może być zweryfikowana
eksperymentalnie, tak jak w fizyce (Theodosius Dobzhansky 1962). Dalszy postęp polegał na rozwinięciu reguł
Mendla.
Odkrycie Morgana, że geny zlokalizowane są na chromosomach ma istotne konsekwencje. Niektóre
cechy nie dziedziczą się niezależnie, tylko mają tendencję do wspólnego występowania u potomstwa. Jest to efekt
położenia genów blisko siebie na jednym chromosomie – sprzężenia. Odkrycie to pozwoliło Morganowi i jego
współpracownikom po raz pierwszy „ułożyć” geny muszki owocowej (Drosophila melanogaster) na chromosomach i
podać odległość pomiędzy nimi tak jakby to były punkty na mapie drogowej. W ten sposób Morgan stworzył
pierwszą mapę genetyczną i wytyczył ramy wszelkich doświadczeń genetycznych. Dzisiaj mapy genetyczne są

niezbędnym narzędziem hodowli, diagnostyki, stanowią istotny element izolowania genów i sekwencjonowania
genomów. Mapy genetyczne oparte o markery molekularne są podstawą genomiki.
Rozszerzenie analiz osobniczych, które są domeną genetyki mendlowskiej, na poziom populacji
stworzyło podstawy genetyki ewolucyjnej. Jak prawa Mendla realizowane są w populacji? Jak zmieniają się
częstości alleli w czasie? Kiedy zmiana to tylko różnicowanie populacji, a kiedy mamy do czynienia ze zmianą
gatunkową? Czy analiza jednego genu jest wystarczająca do odtworzenia ewolucji? Procesy ewolucyjne można
określić słowami: zmienność – dziedziczność – czas (ang. „Variation – Inheritance – Time” = VST). Zmienność i
dziedziczenie to procesy genetyczne. Ewolucja to zmiany genetyczne w populacji na przestrzeni czasu. Każdy
wykształcony człowiek powinien mieć przynajmniej pojęcie o tych procesach, gdyż od tego w dużej mierze zależy
przetrwanie człowieka i jego środowiska.
Wraz z natłokiem informacji o najnowszych osiągnięciach genetyki studenci często są oburzeni, że „każe
im się liczyć groszek i rozwiązywać zadania zamiast pokazać im jak sekwencjonuje się DNA, bada ekspresję,
wprowadza obce geny metodą inżynierii genetycznej”. Nie rozumieją, że analiza dziedziczenia cech i mapowanie
są niezbędnym warsztatem współczesnego genetyka. Jest to tak oczywiste, że naukowcy przedstawiając
publicznie swoje osiągnięcia często pomijają te szczegóły metodyczne skupiając się na nowatorskich wynikach.
Stąd powstaje mylne wrażenie jakoby metody genetyki klasycznej były nieistotne, a nawet przestarzałe. Z
przerażeniem ongiś obserwowałam zdumienie jednego z polskich biologów molekularnych na wieść, iż w genetyce
roślin jeszcze się krzyżuje na potrzeby mapowania. Dziw bierze, ze można w Polsce znaleźć naukowców
kwestionujących znaczenie mapowania i badania dziedziczenia cech. A przecież te metody dostarczają
podstawowej wiedzy o genomie i jego ewolucji. Wystarczy przejrzeć publikacje z zakresu genomiki porównawczej.
Genetyka i nauka o ewolucji są nierozłącznym duetem, to „dwoje drzwi prowadzących do tego samego pokoju”.
Tymczasem są w Polsce ewolucjoniści, którzy uważają, że badania nad zmiennością nie mają nic wspólnego z
ewolucją. Takie poglądy niewątpliwie były inspiracją do powstania obecnej książki.
Podręczniki obecne na polskim rynku również nie ułatwiają zrozumienia znaczenia podstaw genetyki.
Naszpikowane danymi molekularnymi, pełne szczegółowych opisów doświadczeń niczym w fachowych
publikacjach, nie uczą studenta myślenia genetycznego i umiejętności rozwiązywania problemów. Bądź też są to
podręczniki genetyki medycznej, siłą rzeczy skupione na człowieku. Brak tak podstawowych pozycji jak choćby
amerykański podręcznik „Principles of Genetics” (Snustad i Simmons 2006). Jakże daleko polskim pozycjom do
prostoty wywodu i logiki prezentowanej w tejże książce. Jakże w niewielkim stopniu uczą one kreatywności i
niezależnego myślenia tak akcentowanych w podręczniku amerykańskim.
Dlatego obecna książka będąca zbiorem zadań i zagadnień z genetyki i genetyki ewolucyjnej ma nade
wszystko wyrobić w studentach umiejętność samodzielnego myślenia, analizy danych i ustalania dziedziczenia
cech. Część dotycząca ewolucji ma za zadanie uświadomić studentom znaczenie wiedzy o ewolucji w ich
codziennym życiu, w wyborach, gdy powstrzymają się od zniszczenia rzadkiego okazu rośliny, zaśmiecenia
cennego terenu czy uświadomienia sobie, w jakim stopniu nasze dziedzictwo ewolucyjne wpływa na nasze
zachowania społeczne.
Podręcznik niniejszy jest uzupełnieniem problemów zawartych w Plant Genetics and Genomics in
Practice (Polok 2010). Dlatego też zrezygnowano w nim z treści teoretycznych. Zawarte w tym podręczniku
zadania i problemy są efektem wieloletnich rozmów ze studentami oraz obserwowania trudności ze zrozumieniem
genetyki podczas ćwiczeń. Aby ułatwić studentom pracę, pozycja „Genetyka i Ewolucja” została przygotowana w
języku polskim. Wraz z papierową wersją dostępna będzie wersja elektroniczna.
Kornelia Polok, Olsztyn 2010

PPPooodddzzziiieeekkkooowwwaaannniiiaaa Publikacja wykonana w ramach European Community Grant: Marie Curie Host Fellowship for the Transfer of Knowledge, GenCrop, Contract N° MTKD-CT-2004-509834

9
TTTEEERRRMMMIIINNNOOOLLLOOOGGGIIIAAA GGGEEENNNEEETTTYYYCCCZZZNNNAAA
Pokolenia
P: pokolenie rodzicielskie, ang. parental generation, łac. parentes. F1: pierwsze pokolenie mieszańców, ang. first filial generation, łac. Filus; powstaje w wyniku krzyżowania
linii czystych. F1 jest fenotypowo jednorodne i składa się z heterozygot. F2: drugie pokolenie mieszańców, ang. secondo filial generation; powstaje w wyniku samozapłodnienia
lub zapłodnienia krzyżowego osobników F1. Fn: F3, F4…dalsze pokolenia powstałe w wyniku krzyżowania F2, F3… BC1: potomstwo krzyżówki wstecznej, ang. backcross generation; jest efektem krzyżowania osobników F1 z
którymkolwiek z rodziców; dalsze pokolenia powstałe z krzyżowania osobników BC1 to BC2, BC3… TC1: potomstwo krzyżówki testowej, ang. testcross generation, jest efektem krzyżowania heterozygoty F1 z
homozygotą recesywną; dalsze pokolenia powstałe z krzyżowania osobników TC1 to TC2, TC3… Krzyżówka testowa jest wykorzystywana do określenia typów i częstości gamet wytwarzanych przez F1. Umożliwia ona ocenę odległości genetycznej i mapowanie genów. O krzyżowaniu testowym mówi się także, gdy krzyżuje się osobnika o nieznanym genotypie z homozygotą recesywną. Wówczas krzyżówka testowa pozwala określić czy krzyżowany osobnik był homo- czy heterozygotą.
Pojęcia
Fenotyp: zestaw ujawnionych cech danego osobnika. Genotyp: zestaw genów danego osobnika. Allel: alternatywna forma genów; u osobników diploidalnych zawsze występują dwa allele w locus;
w populacji alleli może być wiele. Cecha dzika: normalna, pojęcie używane w stosunku do allela genu, który najczęściej występuje w
przyrodzie; pojęcie najczęściej stosuje się u organizmów modelowych (Arabidopsis thaliana, Drosophila melanogaster). Cechę dziką oznacza się jako „+”. Cecha dzika nie jest równoznaczna z cechą dominującą.
Homozygota: osobnik mający dwa takie same allele w locus. Heterozygota: osobnik mający dwa różne allele w locus. Dominacja: w heterozygocie ujawnia się fenotypowo tylko jeden z alleli; allel ujawniający się w
heterozygocie to allel dominujący, allel nieprzejawiający się fenotypowo w heterozygocie to allel recesywny, jego efekt można obserwować tylko u homozygot.
Kodominacja: heterozygota posiada jednocześnie cechy obu homozygot rodzicielskich. Linia czysta: populacja składająca się z homozygot względem danej cechy, ang. pure line, true breeds. U
roślin linie homozygotyczne względem wielu cech otrzymuje się między innymi w wyniku hodowli wsobnej, krzyżowania oddalonego oraz androgenezy.
Rekombinant: osobnik posiadający kombinację alleli rodzicielskich. Krzyżówka dwupunktowa: rodzaj krzyżówki testowej, w której podwójna heterozygota jest krzyżowana z
podwójną homozygotą recesywną. Krzyżówka trójpunktowa: rodzaj krzyżówki testowej, w której potrójna heterozygota jest krzyżowana z
potrójną homozygotą recesywną.

1. TERMINOLOGIA GENETYCZNA
10
Sprzężenie z płcią: geny zlokalizowane na chromosomach płci; ich segregacja jest różna u poszczególnych płci, wyniki krzyżówek odwrotnych są różne, sprzężenie może być widoczne już u osobników F1 jako różne fenotypy osobników męskich i żeńskich.
Płeć homogametyczna: płeć, u której oba chromosomy płci są identyczne. Płeć heterogametyczna: płeć, u której chromosomy płci są różne, najczęściej X i Y. Sprzężenie genów: geny zlokalizowane na tym samym chromosomie i dziedziczące się wspólnie. Sprzężenie całkowite: na obszarze pomiędzy genami nie występuje crossing-over i w związku z tym nie
obserwuje się gamet i osobników o cechach zrekombinowanych. Interferencja: ograniczenie możliwości zajścia drugiego crossing-over w pobliżu pierwszego. Faza coupling: konfiguracja cis, faza skupiona, faza wzajemnego przyciągania; heterozygota pod względem
dwóch genów; posiada sprzężone allele dominujące na jednym chromosomie homologicznym i allele recesywne na drugim, np. AB i ab.
Faza repulsion: konfiguracja trans, faza rozproszona, faza wzajemnego odpychania; heterozygota pod względem dwóch genów; posiada sprzężony allel recesywny i allel dominujący na obu chromosomach homologicznych, np. Ab i Ab.
CentyMorgan: jednostka mapowa, miara odległości genetycznej obliczana jako częstość crossing-over; 1 cM (1 jm) oznacza, że w danym odcinku chromosomu crossing-over wystąpił tylko raz na 100 chromatyd tego chromosomu odzyskanych w mejozie; najczęściej odległość oblicza się na podstawie częstości fenotypów w krzyżówce testowej. 1 cM = 1 jednostka mapowa = 1% rekombinantów.
Mapa genetyczna: liniowy układ genów na chromosomach wraz ze wzajemnymi odległościami w jednostkach mapowych.
Grupa sprzężeń: grupa genów na mapie genetycznej, które dziedziczą się wspólnie, są sprzężone ze sobą. Dla danego gatunku liczba grup sprzężeń powinna odpowiadać haploidalnej liczbie chromosomów. Grupy ustala się na podstawie wzajemnej odległości i może się zdarzyć, że geny leżące w jednej grupie, ale w znacznej odległości (np. na przeciwległych końcach) dziedziczą się niezależnie.
Zapis genetyczny
A, a: allele jednego genu oznacza się tą samą literą alfabetu, najczęściej stosuje się Italic. A: allel dominujący oznacza się dużą literą. a: allel recesywny oznacza się małą literą. A1, A2…: allele kodominujące oznacza się tą samą literą z indeksem dolnym. A, B, lk, vg…: allele różnych genów oznacza się różnymi literami lub kombinacjami liter. w+, vg+ allele „dzikie” u Drosophila melanogaster i Arabidopsis thaliana często oznacza się znakiem
plus z indeksem górnym wskazującym na warunkujący cechę gen. Małe litery w indeksie górnym oznaczają, że cecha zmieniona (zmutowana) jest recesywna, duże litery symbolizują cechę dominującą.
AA, Aa, aa: zapis genotypu, gdy rozpatrywany jest jeden gen. AA BB: zapis genotypu dla dwóch genów niesprzężonych, czyli zlokalizowanych na dwóch
chromosomach niehomologicznych. A―B―: zapis genotypu, gdy jego ustalenie nie jest jednoznaczne. Kreski oznaczają, że może być A
lub a i analogiczne B lub b. zapis genotypu dla genów sprzężonych, kreska ułamkowa oddziela allele zlokalizowane na
dwóch chromosomach homologicznych, z których każdy pochodzi od innego rodzica. Inaczej, kreska oddziela allele odziedziczone od jednego rodzica od alleli drugiego rodzica. Zamiast ułamka można zastosować ukośnik AB/AB.
zapis genotypu dla dwóch genów sprzężonych oraz genu niesprzężonego z nimi. zapis genotypu dla dwóch grup genów sprzężonych zlokalizowanych na różnych
chromosomach. zapis genotypu dla genów sprzężonych z płcią; symbol w mianowniku oznacza chromosom Y.
ABAB
CC ABAB
CDCD
ABAB
ABf

11
PPPIIIEEERRRWWWSSSZZZEEE PPPRRRAAAWWWOOO MMMEEENNNDDDLLLAAA PPPRRRAAAWWWOOO CCCZZZYYYSSSTTTOOOŚŚŚCCCIII GGGAAAMMMEEETTT
AAAlllllleeellleee ttteeegggooo sssaaammmeeegggooo gggeeennnuuu wwwyyykkkllluuuccczzzaaajjjąąą sssiiięęę wwwzzzaaajjjeeemmmnnniiieee www gggaaammmeeetttaaaccchhh...
Przykład 1
Podaj rozszczepienie, jakie wystąpi w pokoleniach F1 i F2 uzyskanych ze skrzyżowania linii czystych jęczmienia o kłosie ościstym z jęczmieniem o kłosie kapturkowym, jeżeli ościstość kłosa jest uwarunkowana allelem dominującym,K, a kapturek allelem recesywnym, k.
ROZWIĄZANIE
Linia czysta o kłosie kapturkowym (cecha recesywna) musi być homozygotą recesywną. Jej genotyp to kk.
Osobnik o cesze dominującej, ościsty może być heterozygotą o genotypie Kk lub homozygotą dominującą o genotypie KK. Ponieważ jest to linia czysta, a więc homozygotyczna, to jej genotyp jest KK.
W pokoleniu F1 nie obserwowano rozszczepienia. U wszystkich roślin wystąpiły kłosy ościste i były one heterozygotami. W pokoleniu F2 wystąpiło rozszczepienie na rośliny o kłosach ościstych (cecha dominująca) i kłosach przekształconych w kapturek w stosunku 3:1. Wśród roślin z kłosami ościstymi wystąpiły homozygoty dominujące i heterozygoty w stosunku 2:1. Rośliny z kłosem przekształconym w kapturek były homozygotami recesywnymi.
K
K
k
k
kK
F1:
F2:
K k
P:X
K kapturkiemłos z kk
Kłos ościstyKK
Kłos ościstyKk
Kk
Kk
KK
kk
Fenotypy w F ch cz :2 i i ęstości•Kłosy ościste:•Kłosy z kapturkiem:
Genotypy w F :2 i ich częstości•Kłosy ościste - ąca: -heterozygota:
homozygota dominuj
•Kłosy z kapturkiem -homozygota recesywna:
31
12
1

2. PIERWSZE PRAWO MENDLA
12
Przykład 2
Życice (rajgrasy) są cennymi gatunkami pastwiskowymi i trawnikowymi. Dwa gatunki taksonomiczne, życica trwała (Lolium perenne ) oraz życica wielokwiatowa (L. multiflorum) nie są gatunkami biologicznymi i proponuje się je klasyfikować jako podgatunki w obrębie L. perenne. Świadczą o tym między innymi wyniki krzyżowań. Po skrzyżowaniu tych podgatunków życic o nieznanych genotypach i fenotypach otrzymano 157 roślin wykazujących fluorescencję korzonków zarodkowych w świetle UV i 44 rośliny, których korzonki nie fluoryzowały. Jakie były genotypy i fenotypy krzyżowanych roślin?
ROZWIĄZANIE
Skrzyżowano 2 rośliny i otrzymano potomstwo różniące się jedną cechą – zdolnością do fluorescencji korzonków zarodkowych. Prawdopodobnie cecha ta jest kodowana przez jeden gen.
Otrzymano 157 roślin z fluoryzującymi korzonkami i 44 rośliny, które nie wykazywały tej cechy. Stosunek obu typów wynosił 3,56 do 1, co w przybliżeniu odpowiada stosunkowi 3:1. Można to sprawdzić za pomocą testu chi2. W omawianym przypadku wartość chi2 wynosiła 1,04 i świadczyła, że odchylenie od zakładanego stosunku 3:1 jest nieistotne statystycznie(Polok 2007). Zgodnie z pierwszym prawem Mendla stosunek 3:1 występuje, gdy krzyżujemy 2 heterozygoty pod względem 1 genu, przy czym 3 (75%) to osobniki, u których przejawia się fenotypowo cecha dominująca, a 1 (25%) to osobnik z cechą recesywną.
Możemy, więc przyjąć:
157 roślin to osobniki o cesze dominującej, a więc cechą dominującą jest fluorescencja korzonków zarodkowych. Warunkujący ją allel dominujący oznaczymy jako SRF.
44 rośliny to osobniki o cesze recesywnej, a więc brak fluorescencji jest cechą recesywną. Allel ją warunkujący oznaczymy jako srf.
Krzyżowane rośliny były heterozygotami, o genotypie SRFsrf. Rośliny te wykazywały fluorescencję korzonków zarodkowych.
Przykład 3
Skrzyżowano 2 rośliny pomidora. W potomstwie tej krzyżówki otrzymano 106 roślin o owocach czerwonych i 110 roślin o owocach żółtych. Podaj wszystkie możliwe genotypy i fenotypy roślin wziętych do krzyżowania.
ROZWIĄZANIE
Skrzyżowano 2 rośliny i otrzymano rośliny różniące się barwą – czerwone i żółte. Prawdopodobnie segregowała cecha kodowana przez jeden gen.
Rośliny o różnych barwach wystąpiły stosunku zbliżonym do 1:1 (106:110 = 1,04:1).
Stosunek 1:1 występuje, gdy krzyżujemy heterozygotę z homozygotą recesywną pod względem jednego genu. Oznaczmy gen warunkujący barwę owocu jako A dla allela dominującego i a dla allela recesywnego.
Nie jesteśmy w stanie określić, która cecha jest dominująca, a która recesywna, gdyż nie znamy fenotypu heterozygoty. W zadaniu nie podano jak wyglądały krzyżowane rośliny.
Możemy, więc przyjąć,
Genotypy krzyżowanych roślin pomidora to Aa x aa. Fenotypy krzyżowanych roślin to rośliny o owocach czerwonych i żółtych, ale nie jesteśmy w stanie jednoznacznie przypisać ich do genotypów.
Genotypy krzyżowanych roślin mogły być Aa (czerwone) x aa (żółte) lub Aa (żółte) x aa (czerwone).

2. PIERWSZE PRAWO MENDLA
13
Zadania
1. U wyżlinu różowa barwa kwiatów jest wynikiem niepełnej dominacji allela warunkującego czerwoną barwę kwiatów nad allelem decydującym o białej barwie kwiatów. Allel warunkujący kwiaty grzbieciste dominuje nad allelem decydującym o kwiatach promienistych. Podaj wszystkie możliwe genotypy roślin o kwiatach różowych, grzbiecistych, które skrzyżowane ze sobą dały rozszczepienie na rośliny czerwone o kwiatach grzbiecistych, różowe o kwiatach grzbiecistych oraz białe o kwiatach grzbiecistych w proporcjach 1:2:1.
2. W potomstwie uzyskanym ze skrzyżowania 2 roślin pszenicy otrzymano 267 roślin o czarno zabarwionym ziarniaku oraz 810 roślin o normalnym zabarwieniu ziarniaków. Podaj wszystkie możliwe genotypy i fenotypy krzyżowanych roślin.
3. W pokoleniu F2 75% stanowią rośliny o kwiatach barwnych i 25% rośliny o kwiatach białych. Jeżeli spośród roślin o kwiatach barwnych wybrać losowo jedną i samozapylić, to jakie jest prawdopodobieństwo, że w jej potomstwie będzie się obserwowało segregację.
4. Odmiany fasoli różnią się wrażliwością na zakażenia wirusowe. Niektóre reagują na infekcję wytwarzaniem niewielkich plam nekrotycznych, które nie mają większego wpływu na roślinę, inne wykazują zakażenie systemowe znacznie obniżające żywotność roślin. Po skrzyżowaniu odmian fasoli o różnym pochodzeniu i różniących się reakcją na infekcję wirusem (jedna wykazywała jedynie nekrotyczne plamy, druga zakażenie systemowe) w pokoleniu F1 otrzymano rośliny z nekrotycznymi plamami, natomiast w pokoleniu F2 otrzymano 785 roślin z nekrotycznymi plamami oraz 269 roślin z zakażeniem systemowym. W jaki sposób dziedziczy się cecha wrażliwości na zakażenia wirusem u fasoli. Podaj genotypy i fenotypy potomstwa krzyżówki testowej.
5. U bydła bezrożność dominuje nad występowaniem rogów. Bezrożnego buhaja skojarzono z 3 krowami. Cielę uzyskane z skojarzenia z krową nr 1, rogatą było rogate, cielę uzyskane z krowy nr 2, bezrożnej było rogate, natomiast cielę uzyskane z krowy nr 3, bezrożnej było bezrożne. Podaj genotyp buhaja i 3 krów użytych do krzyżowań. Jakich stosunków rozszczepień należy spodziewać się w każdej krzyżówce?
6. U cocker spanieli jednolite zabarwienie sierści dominuje nad łaciatym. Jakie jest prawdopodobieństwo, że pierwsze szczenię pokolenia F2 krzyżówki pomiędzy samicą linii czystej o jednolitym zabarwieniu a samcem linii czystej łaciatej będzie łaciate? Jakie jest prawdopodobieństwo, że w miocie składającym się z czterech szczeniąt wszystkie będą miały jednolite zabarwienie?
7. Opierzone skoki u kur są cechą dominującą nad brakiem opierzenia (allele O, o). Koguta o opierzonych skokach skojarzono z dwoma kurami o skokach opierzonych. Po kurze nr 1 otrzymano kurczęta mające tylko opierzone skoki. Kura nr 2 dała kurczęta zarówno o skokach opierzonych, jak i gładkich. Podaj genotypy koguta i obu kur.
8. Dzikie rude lisy mają czasami szczenięta czarne. Czarne szczenięta odławia się i po uzyskaniu dojrzałości są one rozmnażane w niewoli. W ten sposób otrzymuje się wyłącznie lisy czarne. Wytłumacz, jaka jest dziedziczna podstawa tej cechy, dlaczego tak łatwo wytworzyć czysty typ czarny, który nie będzie dawał rudego potomstwa? Jakie zwierzęta powstałyby w wyniku wstecznego kojarzenia czarnych lisów z ich własnymi rudymi rodzicami? Podaj ich genotypy potencjalnych rodziców, fenotypy oraz częstość występowania czarnych szczeniąt w ich potomstwie.
9. Barwa oczu u ludzi zależy od jednego genu. Dominujący allel B jest odpowiedzialny za barwę ciemną, a recesywny allel b za barwę jasną. Jakie jest prawdopodobieństwo urodzenia jasnookiego dziecka w małżeństwie kobiety o oczach ciemnych z mężczyzną o oczach jasnych, jeżeli matka kobiety miała oczy jasne?
10. Czy jest prawdziwe twierdzenie: jeżeli w małżeństwie heterozygotycznych ciemnookich rodziców urodziło się troje dzieci ciemnookich, to czwarte dziecko będzie miało oczy jasne? Uzasadnij odpowiedź.

©Kornelia Polok 14
DDDRRRUUUGGGIIIEEE PPPRRRAAAWWWOOO MMMEEENNNDDDLLLAAA DDDZZZIIIEEEDDDZZZIIICCCZZZEEENNNIIIEEE NNNIIIEEEZZZAAALLLEEEŻŻŻNNNEEE
AAAlllllleeellleee rrróóóżżżnnnyyyccchhh gggeeennnóóówww ssseeegggrrreeeggguuujjjąąą nnniiieeezzzaaallleeeżżżnnniiieee oooddd sssiiieeebbbiiieee pppooodddccczzzaaasss tttwwwooorrrzzzeeennniiiaaa gggaaammmeeettt... JJJeeesssttt tttooo zzzaaasssaaadddaaa nnniiieeezzzaaallleeeżżżnnneeejjj ssseeegggrrreeegggaaacccjjjiii ccceeeccchhh... RRRóóóżżżnnneee gggeeennnyyy dddzzziiieeedddzzziiiccczzząąą sssiiięęę nnniiieeezzzaaallleeeżżżnnniiieee...
Przykład 1
U bielunia kwiaty purpurowe (P) dominują nad kwiatami białymi (p), a torebki kolczaste (S) nad gładkimi (s). Skrzyżowano homozygotyczną roślinę o kwiatach białych i torebkach kolczastych z homozygotyczną rośliną o kwiatach purpurowych i torebkach gładkich.
a) Podaj fenotyp i genotyp pokolenia F1 oraz rozszczepienie w pokoleniu F2. b) Jakie będą fenotypy i genotypy otrzymame w potomstwie krzyżówki wstecznej z rodzicem o kwiatach
białych i torebkach kolczastych? c) Jakie będą rozszczepienia fenotypów w potomstwie krzyżówki testowej?
ROZWIĄZANIE – PUNKT A)
Homozygotyczna roślina o kwiatach białych i torebkach kolczastych ma genotyp ppSS. Roślina ta wytwarza gamety zawierające po jednym allelu z każdego genu, czyli pS.
Homozygotyczna roślina o kwiatach purpurowych i torebkach gładkich ma genotyp PPss. Roślina ta wytwarza gamety zawierające po jednym allelu z każdego genu, czyli Ps.
Pokolenie F1 powstaje z połączenia gamet rodzicielskich, pS i Ps. Jest ono heterozygotą pod względem obu genów, PpSs. W heterozygocie przejawiają się cechy dominujące. W związku z tym F1 musi mieć purpurowe kwiaty i kolczaste torebki.
Pokolenie F1 wytworzą 4 typy gamet, z których każda zawiera po jednym allelu z obu genów. Allele P, p oraz S, s przekazywane są do gamet niezależnie od siebie, co prowadzi do powstania wszystkich możliwych kombinacji.
F1:
P:Białe,Kolczaste
Purpurowe,Gładkie
Purpurowe,Kolczaste
X
ppSS PPss
PpSs
pS Ps
PS pSPs ps

3. DRUGIE PRAWO MENDLA
©Kornelia Polok 15
Pokolenie F2 powstaje poprzez losowe łączenie się 4 typów gamet wytworzonych przez krzyżowane osobniki F1.
Uwzględniając każdą cechę oddzielnie, rozszczepienie na fenotypy o cesze dominującej i recesywnej jest zgodne z Pierwszym Prawem Mendla i wynosi 3:1. Jeżeli obie cechy uwzględnimy łącznie to otrzymujemy rozszczepienie na:
9 osobników o obu cechach dominujących; można je zapisać jako P―S―; 3 + 3 osobniki o jednej cesze dominującej (purpurowe kwiaty lub kolczasty owoc); można je zapisać jako P―ss lub ppS―;
1 osobnik o obu cechach recesywnych, można go zapisać jako ppss. Rozszczepienie fenotypów w stosunku 9:3:3:1 jest charakterystyczne dla segregacji dwóch cech, które dziedziczą się zgodnie z Drugim Prawem Mendla, a więc niezależnie.
ROZWIĄZANIE – PUNKT B
Krzyżówka wsteczna jest w tym przypadku krzyżówką pomiędzy pokoleniem F1 otrzymanym w punkcie a) oraz rodzicem o białych kwiatach I kolczastych torebkach. Pokolenie F1 jest heterozygotą pod względem obu cech o genotypie PpSs, natomiast rodzic ma genotyp ppSS.
Pokolenie F1 wytwarza 4 typy gamet, PS, Ps, pS, ps Rodzic wytwarza tylko jeden typ gamet, pS (patrz: punkt a).
BC1PpSS PpSs ppSS ppSs
pSPs psPS
pSFenotypy w :BC•
1
Kwiaty purpurowe, torebki kolczaste:•Kwiaty białe, torebki kolczaste:
22
F2:
pSPs psPS
pS
Ps
ps
PS
PPSS PPSs PpSS PpSs
PPSs PPss PpSs Ppss
PpSS PpSs ppSS ppSs
PpSs Ppss ppSs ppss Pod względem barwy kwiatu:purpurowe : białe =12 : 4 = 3 : 1
Pod względem kształtu torebki:kolczasty : gładki =12 : 4 = 3 : 1
Fenotypy w F:
2
pod względem obu cech•Kwiaty purpurowe, torebki kolczaste:•••
Kwiaty purpurowe, torebki gładkie:Kwiaty białe, torebki kolczaste:Kwiaty białe,torebki gładkie:
9331

3. DRUGIE PRAWO MENDLA
©Kornelia Polok 16
W potomstwie obserwujemy rozszczepienie tylko pod względem barwy kwiatu. Wystąpią 2 osobniki o kwiatach purpurowych i 2 o kwiatach białych, co odpowiada stosunkowi 1:1. Osobniki o kwiatach purpurowych są heterozygotami, Pp, natomiast osobniki o kwiatach białych są homozygotami recesywnymi pp.
Nie obserwujemy rozszczepienia pod względem kształtu torebki. Wszystkie rośliny mają torebki kolczaste. Jednakże genotypowo różnią się między sobą. Połowa osobników to homozygoty dominujące, SS, a połowa to heterozygoty, Ss.
ROZWIĄZANIE – PUNKT C
Krzyżówka testowa polega na skrzyżowaniu homozygoty recesywnej z heterozygotą F1. Krzyżowano więc homozygotę recesywną pod względem obu cech, czyli osobnika o kwiatach białych i gładkich torebkach, o genotypie ppss z osobnikiem F1 o kwiatach purpurowych i kolczastej torebce, o genotypie PpSs
Heterozygota F1 wytwarza 4 typy gamet (punkt a), natomiast homozygota recesywna tylko jeden typ, ps.
W krzyżówce testowej powstają 4 klasy fenotypowe. Każda z nich stanowi ¼ (25%) potomstwa. Stosunek rozszczepień jest 1:1:1:1 na osobniki o kwiatach purpurowych i kolczastych torebkach, o kwiatach purpurowych i gładkich torebkach, o kwiatach białych i kolczastych torebkach oraz o kwiatach białych i gładkich torebkach. Otrzymane klasy fenotypowe odpowiadają typom gamet wytwarzanych przez heterozygotę F1. W uproszczeniu, homozygota recesywna wytwarza gamety, które nie przeszkadzają w fenotypowym ujawnieniu się cech wnoszonych przez gamety wytwarzane przez heterozygotę.
Rozszczepienie 1:1:1:1 jest charakterystyczne dla krzyżowania testowego heterozygoty pod względem dwóch genów. Rodzaje klas genotypowych i ich częstość odpowiadają rodzajom i częstościom gamet wytworzonych przez heterozygotę. Gdy segreguje więcej genów zwiększa się odpowiednio liczba klas genotypowych (np. 8 dla 3 genów), ale częstości poszczególnych klas są identyczne. Występowanie w potomstwie krzyżówki testowej wszystkich przewidywanych klas fenotypowych z równą częstością jest charakterystyczne dla genów, które dziedziczą się niezależnie – nie są sprzężone. Jakiekolwiek odstępstwo od równych częstości klas fenotypowych w potomstwie krzyżówki testowej świadczy o sprzężeniu genów.
TC1
pSPs psPS
ps
Fenotypy w :TC•
1
Kwiaty purpurowe, torebki kolczaste:•
•
Kwiaty purpurowe, torebki gładkie:
Kwiaty białe, torebki gładkie:•Kwiaty białe, torebki kolczaste:
1111
Białe,Gładkie
Purpurowe,Kolczaste
Xppss PpSs
Homozygotarecesywna
Heterozygota
PpSs Ppss ppSs ppss

3. DRUGIE PRAWO MENDLA
©Kornelia Polok 17
Przykład 2
Po skrzyżowaniu stokrotki o kwiatach białych, niskiej ze stokrotką o kwiatach czerwonych, wysoką otrzymano w F1 rośliny różowe i wysokie, a w F2 nastąpiło rozszczepienie na następujące klasy osobników:
180 roślin czerwonych, wysokich; 360 roślin różowych, wysokich; 180 roślin białych, wysokich; 60 roślin czerwonych, niskich; 120 roślin różowych, niskich; 60 roślin białych, niskich.
Ustal genotypy rodziców i wyjaśnij jak dziedziczy się każda z tych cech?
ROZWIĄZANIE
W krzyżówce segregują 2 cechy – barwa kwiatu i wysokość rośliny. W wyniku skrzyżowania dwóch roślin o przeciwstawnych cechach (biała – czerwona, niska – wysoka) otrzymano jednolite pokolenie F1, co świadczy, że krzyżowane rośliny były liniami czystymi – homozygotami pod względem obu cech.
Pokolenie F1 musi być heterozygotą pod względem obu cech – barwy kwiatu i wysokości rośliny. Możemy zapisać jego genotyp jako AaBb, gdzie A, a oznacza barwę kwiatu, a B, b – wysokość.
Na podstawie pokolenia F1 ustalamy, które cechy są dominujące, a które recesywne.
Różowa, czyli pośrednia barwa kwiatów w F1 wskazuje na niepełną dominację barwy czerwonej nad białą. Stokrotka biała musi mieć genotyp aa, natomiast stokrotka czerwona AA.
Fenotyp F1 wskazuje, że wysoki wzrost był cechą dominującą na wzrostem niskim. W takim razie możemy zapisać bb dla stokrotki niskiej i BB dla stokrotki wysokiej.
Sumując powyższe twierdzenia wnioskujemy, że krzyżowano stokrotkę o kwiatach białych, niską o genotypie aabb ze stokrotką o kwiatach czerwonych, wysoką o genotypie AABB.
Wnioski z punktu 3 znajdują potwierdzenie w rozszczepieniu obserwowanym w pokoleniu F2, które otrzymano w wyniku krzyżowania osobników F1. Dla uproszczenia rozważmy każdą cechę oddzielnie.
Pod względem barwy kwiatu występuje 240 roślin o kwiatach czerwonych, 480 o kwiatach różowych i 240 roślin o kwiatach białych. Odpowiada to stosunkowi 1:2:1, typowemu dla krzyżowania 2 heterozygot względem genu barwy kwiatu oraz w przypadku, gdy barwa czerwona nie dominuje w pełni nad barwą białą. Wówczas stosunek fenotypów w F2 odpowiada stosunkowi genotypów zgodnie z Pierwszym Prawem Mendla.
Pod względem wysokości występuje 720 roślin wysokich i 240 niskich, co odpowiada mendlowskiemu stosunkowi 3:1 wynikającego ze skrzyżowania 2 heterozygot.
Podsumowując, osobniki F1 były heterozygotami pod względem obu genów, AaBb i otrzymano je w wyniku krzyżowania homozygot rodzicielskich.
Przykład 3
Geny a, b i c dziedziczą się niezależnie i są recesywne w stosunku do alleli A, B, C. Jakie jest prawdopodobieństwo otrzymania w potomstwie powstałym ze skrzyżowania dwóch heterozygot pod względem wszystkich genów osobników o wszystkich cechach dominujących? Jakie jest prawdopodobieństwo otrzymania osobników homozygotycznych pod względem wszystkich genów?
ROZWIĄZANIE
Genotyp heterozygoty pod względem 3 genów można zapisać AaBbCc.
Heterozygota ta wytwarza 8 typów gamet. Liczbę tę można obliczyć ze wzoru 2n, gdzie n to liczba genów w stanie heterozygotycznym. Ponieważ 23 = 8, to heterozygota pod względem 3 genów wytwarza 8 typów gamet.

3. DRUGIE PRAWO MENDLA
©Kornelia Polok 18
Pokolenie F2 powstanie przez połączenie się 8 typów gamet osobnika matecznego z 8 typami gamet osobnika ojcowskiego. Powstanie 8 x 8 = 64 kombinacji genotypów oraz 8 klas fenotypowych.
Dla każdej z cech oddzielnie stosunek osobników o cesze dominującej do tych o cesze recesywnej będzie 3:1. Klasy fenotypowe to wszystkie możliwe kombinacje cech recesywnych i dominujących. Jest ich 8. Najwięcej będzie osobników o 3 cechach dominujących – 27, A―B―C―, następnie o dwóch cechach dominujących – 9 A―B―cc, 9 A―bbC― i 9 aaB―C―, o 1 cesze dominującej – 3 A―bbcc, 3 aaB―cc, 3 aabbC― oraz 1 homozygota recesywna aabbcc.
Z powyższego wynika, że osobniki o wszystkich cechach dominujących wystąpią z prawdopodobieństwem 27/64.
Osobniki homozygotyczne pod względem wszystkich genów to homozygoty dominujące oraz recesywne. Pierwsze powstają z połączenia gamety matecznej ABC z ojcowską ABC. Jest tylko jeden taki przypadek. Analogicznie, homozygoty recesywne powstają z połączenia gamet abc i abc. Też jest tylko jedna taka możliwość. Podsumowując osobniki homozygotyczne stanowią 2/64 wszystkich możliwości, czyli 1/32.
Przykład 4
Po skrzyżowaniu pomidora o wzroście karłowym, liściach całobrzegich, łodydze antocjanowej z pomidorem o wzroście wysokim, liściach całobrzegich, łodydze o barwie antocjanowej otrzymano:
180 roślin wysokich, o liściach całobrzegich, łodydze antocjanowej; 20 roślin wysokich, o liściach ząbkowanych, łodydze zielonej; 60 roślin wysokich, o liściach całobrzegich, łodydze zielonej; 60 roślin wysokich, o liściach ząbkowanych, łodydze antocjanowej; 180 roślin karłowych, o liściach całobrzegich, łodydze antocjanowej; 60 roślin karłowych, o liściach całobrzegich, łodydze zielonej; 60 roślin karłowych, o liściach ząbkowanych, łodydze antocjanowej; 20 roślin karłowych, o liściach ząbkowanych, łodydze zielonej.
Podaj wszystkie możliwe genotypy rodziców.
ROZWIĄZANIE
W potomstwie wystąpiło rozszczepienie pod względem wszystkich cech, co świadczy, iż krzyżowane rośliny nie były liniami czystymi i wykazywały różny stopień heterozygotyczności.
Genotypy krzyżowanych roślin oraz cechy dominujące można ustalić na podstawie stosunków rozszczepień w potomstwie. Dla uproszczenia przeanalizujmy każdą cechę osobno.
Wzrost – gen kodujący wzrost oznaczmy literą A, a. Otrzymano 320 roślin wysokich i 320 roślin niskich. Stosunek roślin wysokich do niskich wynosił 1:1, a więc był charakterystyczny dla krzyżowania heterozygoty z homozygotą recesywną (krzyżówka testowa). Genotypy krzyżowanych roślin to Aa x aa. Nie można ustalić, która z cech jest dominująca.
Liście – oznaczmy literą B, b. Otrzymano 480 roślin o liściach całobrzegich i 160 o liściach ząbkowanych. Stosunek fenotypów wynosi 3:1. Jest zgodny ze stosunkiem rozszczepień wynikającym z krzyżowania 2 heterozygot. Cechą dominującą są liście całobrzegie. Genotypy krzyżowanych roślin to Bb x Bb. Łodyga – oznaczmy literą C, c. Otrzymano 480 roślin o łodydze antocjanowej i 160 o łodydze zielonej. Stosunek fenotypów wynosi 3:1, czyli krzyżowano 2 heterozygoty. Cechą dominującą są jest łodyga antocyjanowa. Genotypy krzyżowanych roślin to Cc x Cc.
Krzyżowane rośliny miały następujące genotypy:
pomidor karłowy o liściach całobrzegich i łodydze antocjanowej - aaBbCc x pomidor wysoki o liściach całobrzegich, łodydze antocjanowej – AaBbCc lub
pomidor karłowy o liściach całobrzegich i łodydze antocjanowej - AaBbCc x pomidor wysoki o liściach całobrzegich, łodydze antocjanowej – aaBbCc.

3. DRUGIE PRAWO MENDLA
©Kornelia Polok 19
Zadania
1. Po skrzyżowaniu ogórka o kwiatach żółtych i łodygach owłosionych z ogórkiem o takim samym kolorze kwiatów i łodydze nagiej otrzymano:
94 rośliny o kwiatach żółtych i łodydze owłosionej; 81 roślin o kwiatach żółtych i łodydze nagiej; 29 roślin o kwiatach białych i łodydze nagiej; 30 roślin o kwiatach białych i łodydze owłosionej.
Podaj wszystkie możliwe genotypy rodziców.
2. Podaj genotypy rodziców oraz roślin pokolenia F1 wiedząc, że w wyniku krzyżówki testowej pszenicy o kłosach luźnych i wysokim wzroście otrzymano:
10 roślin o kłosie luźnym, wysokich; 10 roślin o kłosie zbitym, wysokich; 10 roślin o kłosie luźnym, karłowych; 10 roślin o kłosie zbitym, karłowych.
3. Podaj wszystkie możliwe genotypy rodziców, a z jednym z nich wykonaj krzyżówkę testową, wiedząc, że po skrzyżowaniu melona o kwiatach żółtych, łodydze spłaszczonej, owłosionej z melonem o takim samym fenotypie otrzymano:
180 roślin o kwiatach żółtych, łodydze spłaszczonej, owłosionej; 60 roślin o kwiatach żółtych, łodydze okrągłej, owłosionej; 60 roślin o kwiatach białych, łodydze spłaszczonej, owłosionej; 20 roślin o kwiatach białych, łodydze okrągłej, owłosionej.
4. U dyni barwa owocu jest uwarunkowana parą alleli W, w a kształt owocu parą alleli D, d. Białe owoce są dominujące w stosunku do owoców żółtych, owoce płaskie są dominujące w stosunku do owoców kulistych.
a) Podaj genotypy dyni o owocach białych, płaskich oraz dyni o owocach żółtych, kulistych, które skrzyżowane ze sobą dały 169 roślin o owocach białych, płaskich oraz 165 o owocach białych, kulistych.
b) Podaj genotypy dwóch dyni o owocach białych, kulistych, które skrzyżowane ze sobą dały 77 roślin o owocach białych, kulistych oraz 25 o owocach żółtych, kulistych.
c) Podaj genotypy dyni o owocach żółtych, płaskich oraz dyni o owocach białych, kulistych, które skrzyżowane ze sobą dały jedynie rośliny o owocach białych, płaskich.
d) Podaj genotypy dyni o owocach białych, płaskich oraz dyni o owocach żółtych, kulistych, które skrzyżowane ze sobą dały 52 rośliny o owocach białych, płaskich, 56 roślin o owocach białych, kulistych, 54 rośliny o owocach żółtych, płaskich oraz 50 o owocach żółtych, kulistych.
e) Podaj genotypy dyni o owocach białych, płaskich oraz dyni o owocach białych, kulistych, które skrzyżowane ze sobą dały 3/8 roślin o owocach białych, płaskich, 3/8 roślin o owocach białych, kulistych, 1/8 roślin o owocach żółtych, płaskich oraz 1/8 roślin o owocach żółtych, kulistych.
5. Podaj genotyp Drosophila melanogaster o oczach eozynowych, barwie ciała czarnej (black), która skrzyżowana testowo dała 106 osobników o oczach eozynowych i ciele czarnym oraz 111 osobników o oczach białych i ciele czarnym.
6. U królików himalajski typ futerka jest ustępujący w stosunku do dzikich, jednolicie zabarwionych osobników, a włos długi jest recesywny w stosunku do krótkiego. Skrzyżowano dziką, krótkowłosą samicę z samcem o nieznanym fenotypie. W potomstwie otrzymano 6 królików dzikich (jednolicie szarych) i krótkowłosych, 5 królików szarych, o włosie długim oraz 4 o futerku himalajskim, w tym 2 posiadały włos krótki, a 2 włos długi. Podaj genotypy rodziców.
7. U królików umaszczenie łaciate dominuje nad jednolitym (S, s), a czarna barwa sierści nad brązową (B, b). Jednolicie czarnego samca skojarzono z jednolicie brązową samicą i otrzymano dwie tak samo liczne dwie grupy potomstwa o fenotypach takich jak rodzice. Jaki był genotyp samca?

3. DRUGIE PRAWO MENDLA
©Kornelia Polok 20
8. Podejrzewamy, że czarny bezrożny buhaj jest heterozygotyczny pod względem obu cech. W jaki sposób można najłatwiej sprawdzić jego genotyp? Udowodnij to na schemacie.
9. Bezrożność jest u bydła cechą dominującą nad rogatością i zależy od pary alleli P, p. Rasę shorthorn charakteryzują trzy różne barwy umaszczenia uwarunkowane jednym genem. Osobniki czerwone mają genotyp RR, dereszowate Rr, a białe rr. W wyniku krzyżowania bezrogiego, czerwonego bydła o genotypie PPRR z rogatym, dereszowatym uzyskano zwierzęta bezrogie, przy czym 1/2 była czerwona i 1/2 dereszowata. Jeżeli skojarzymy ze sobą zwierzęta bezrogie, dereszowate z tego potomstwa, to jaki będzie udział zwierząt o takim samym jak ich genotypie w kolejnym pokoleniu?
10. Barwa upierzenia u kur andaluzyjskich wyznaczona jest przez parę alleli A, a. Homozygoty AA są czarne, aa - białe, natomiast heterozygoty mają barwę niebieskopopielatą. Gen O warunkuje opierzone skoki, a jego allel recesywny o - skoki nieopierzone. Niebieskiego koguta o opierzonych skokach skojarzono z trzema kurami I, II, III. Kura I była biała o nieopierzonych skokach, kura II - niebieska o nieopierzonych skokach, kura III - czarna o opierzonych skokach. Otrzymano 3 pisklęta, po jednym z każdej kury. Pisklę po kurze I było białe o nieopierzonych skokach, po kurze II - niebieskie o opierzonych skokach, po III - niebieskie o skokach nieopierzonych. Jakie są genotypy koguta i skojarzonych z nim kur? Jakiego potomstwa można oczekiwać po tych samych ptakach i w jakich proporcjach, jeżeli będą się wylęgać pisklęta zewszystkich zniesione jaj?
11. U świni krótka sierść jest dominująca (L) w stosunku do długiej sierści (l), natomiast barwa kremowa sierści jest wynikiem niepełnej dominacji allela warunkującego barwę żółtą sierści (W) nad allelem warunkującym barwę białą (w). W wyniku skojarzenia kremowych świń o krótkiej sierści z świniami białymi o długiej sierści urodził się kremowy prosiak o długiej sierści. Jakich prosiąt i z jaką częstotliwością należy się spodziewać po skojarzeniu tego prosiaka z rodzicem kremowym o krótkiej sierści?
12. Po skrzyżowaniu D. melanogaster o oczach czerwonych, skrzydłach vestigal, barwie ciała szarej i włoskach szczecinek zredukowanych z osobnikiem o oczach czerwonych, skrzydłach normalnych, barwie ciała szarej, włoskach szczecinek zredukowanych otrzymano:
90 much o oczach czerwonych, skrzydłach vestigal, ciele szarym, szczecinkach zredukowanych; 90 much o oczach czerwonych, skrzydłach normalnych, ciele szarym, szczecinkach zredukowanych; 30 much o oczach czerwonych, skrzydłach normalnych, ciele yellow (żółtym), szczecinkach
zredukowanych; 30 much o oczach czerwonych, skrzydłach vestigal, ciele yellow (żółtym), szczecinkach zredukowanych; 30 much o oczach białych, skrzydłach vestigal, ciele szarym, szczecinkach zredukowanych; 30 much o oczach białych, skrzydłach normalnych, ciele szarym, szczecinkach zredukowanych; 10 much o oczach białych, skrzydłach vestigal, ciele yellow (żółtym), szczecinkach zredukowanych; 10 much o oczach białych, skrzydłach normalnych, ciele yellow (żółtym), szczecinkach zredukowanych.
Podaj wszystkie możliwe genotypy rodziców.
13. U człowieka oczy ciemne dominują nad niebieskimi, a włosy proste nad kręconymi. Jakie jest prawdopodobieństwo urodzenia dziecka o oczach niebieskich i kręconych włosach w małżeństwie kobiety o oczach niebieskich i włosach prostych z mężczyzną o oczach ciemnych i włosach prostych? Rodzice kobiety oraz matka mężczyzny mieli oczy niebieskie. Zarówno matka kobiety jak i matka mężczyzny mieli włosy kręcone. Podaj genotypy wszystkich członków rodziny.
14. U kotów szkockich występują skręcone uszy uwarunkowane allelem dominującym F. Allel recesywny f warunkuje powstanie typowych uszu. Allel recesywny b jest odpowiedzialny za rozjaśnienie czarnej sierści kotów, które w efekcie mają barwę czekoladową. Skojarzono dwa czarne koty, z których jeden miał uszy skręcone, a drugi normalne. W miocie pojawiły się dwa kocięta czarne i dwa czekoladowe, ale wszystkie miały normalne uszy. Podaj genotypy krzyżowanych kotów. Czy jest możliwe i z jakim prawdopodobieństwem pojawienie się w tej parze kotów czekoladowych o uszach skręconych?

21
KKKOOODDDOOOMMMIIINNNAAACCCJJJAAA
WWW hhheeettteeerrrooozzzyyygggoooccciiieee uuujjjaaawwwnnniiiaaajjjąąą sssiiięęę fffeeennnoootttyyypppyyy dddeeettteeerrrmmmiiinnnooowwwaaannneee ppprrrzzzeeezzz ooobbbaaa aaalllllleeellleee dddaaannneeegggooo gggeeennnuuu... KKKaaażżżdddyyy zzz aaalllllleeellliii uuullleeegggaaa nnniiieeezzzaaallleeeżżżnnniiieee eeekkksssppprrreeesssjjjiii...
Przykład 1
U człowieka zdolność do wytwarzania antygenów krwinkowych M i N jest warunkowana przez gen L z dwoma allelami. Jeden z alleli genu L warunkuje produkcję antygenu typu M, a drugi jest odpowiedzialny za wytworzenie antygenu typu N. W małżeństwie kobiety z grupą krwi M oraz mężczyzny z grupą krwi N urodziło się troje dzieci. W krwi wszystkich dzieci stwierdzono występowanie zarówno antygenu M jak i N, a ich grupę krwi oznaczono jako MN. Wyjaśnij genetyczne uwarunkowania produkcji antygenów M i N oraz podaj genotypy rodziców i dzieci.
ROZWIĄZANIE
Dzieci otrzymały po jednym allelu od każdego z rodziców. Matka przekazała allel LM odpowiedzialny za produkcję antygenu M. Ojciec przekazał allel LN odpowiedzialny za produkcję antygenu N.
Wszystkie dzieci są heterozygotami o genotypie LMLN. U dzieci produkowany jest zarówno antygen M jak i N, co prowadzi do powstania grupy MN. Oba allele dają efekt fenotypowy „niezależnie od siebie”. Oba allele przejawiają się w heterozygocie. Żaden z nich nie jest dominujący.
Ten typ działania alleli jednego genu nosi nazwę kodominacji. Od niepełnej dominacji różni się tym, iż w przypadku kodominacji widoczne są efekty działania obu alleli, natomiast niepełna dominacja charakteryzuje się fenotypem pośrednim, który powstaje na skutek osłabienia efektu genotypowego allela dominującego. W przypadku kodominacji, żaden z alleli nie jest dominujący, ani nawet częściowo dominujący. Dlatego nie zapisuje się ich dużymi i małymi literami. Zapisuje się je jako indeksy górne przy literze symbolizującej gen. Kodominacja najczęściej występuje na poziomie molekularnym. Obserwuje się ją między innymi w odniesieniu do grup krwi AB, izoenzymów, niektórych markerów DNA (SSR, RFLP).
.
P:
F:
X
LM
Grupa M Grupa N
Grupa MN
L LM M L LN N
L LM N
LN

4. KODOMINACJA
22
Przykład 2
Izoenzymy są to formy danego enzymu pełniące taką samą funkcję, ale różniące się niektórymi właściwościami fizycznymi, w tym ruchliwością w polu elektrycznym. Izoenzymy o różnej ruchliwości często uwarunkowane są różnymi allelami jednego genu. Różnice w ruchliwości można wykryć przy pomocy elektroforezy enzymów. Metoda ta polega na nałożeniu ekstraktu białkowego na stały nośnik (skrobia, agaroza, poliakrylamid). Następnie żel podłącza się do prądu o określonym napięciu. W powstałym polu elektrycznym enzymy migrują, przy czym kierunek i szybkość migracji zależą od znaku i wielkości ładunku elektrycznego. Po odłączeniu napięcia, enzymy zostają unieruchomione w określonym miejscu nośnika i można je ujawnić za pomocą specyficznej reakcji histochemicznej. W efekcie otrzymuje się obraz barwnych prążków w określonej odległości od miejsca naniesienia ekstraktu. Gen Per1 u Lolium perenne jest odpowiedzialny za wytwarzanie peroksydazy. W locus tym występuje allel Per1F
kodujący enzym (allozyme) o ruchliwości 45 mm oraz allel Per1S kodujący enzym migrujący wolniej, o ruchliwości 35 mm. Po skrzyżowaniu rośliny homozygotycznej z szybko migrującą peroksydazą (45 mm) z rośliną homozygotyczną, u której wystąpił wariant wolnomigrujący (35 mm) w F2 pojawiły się dwa rodzaje fenotypów jednoprążkowych oraz fenotypy dwuprążkowe (patrz rysunek: obraz rozdziału elektroforetycznego w pokoleniu F2).
a) Wyjaśnij skąd wzięły się fenotypy dwuprążkowe? b) Jakie były genotypy roślin obserwowanych w F2 oraz jaki był genotyp i fenotyp pokolenia F1? c) W jakich proporcjach wystąpiły poszczególne fenotypy w pokoleniu F2?
ROZWIĄZANIE
P: XRuchliwość - 45 mm
(forma szybka)Ruchliwo 3
wolnaść - 5 mm
(forma )
Ruchliwo i 35 mm
ść45 mm
Per1 Per1F F Per1 Per1S S
Per1 Per1F S
F
F
S
S
Per1
Per1
Per1
Per1
Per1 F
Per1 F
Per1 S
Per1 S
F1:
F2:
Obraz rozdzia
żelu otrzymany w wynikułu elektroforetycznego
peroksydaz w pokoleniu F .2

4. KODOMINACJA
23
Allel Per1F jest odpowiedzialny za wytworzenie cząsteczki enzymu o ruchliwości 45 mm. Homozygoty pod względem allela Per1F wytwarzają jedynie formy peroksydazy o ruchliwości 45 mm, co jest widoczne na żelu jako jeden, szybko migrujący prążek, tzn. widoczny jako prążek położony dalej od początku żelu (ścieżka na żelu nr 1).
Podobnie, homozygoty względem allela Per1S wytwarzają jedynie cząsteczki peroksydazy o ruchliwości 35 mm, co jest widoczne na żelu jako wolno migrujący prążek, tzn. widoczny jako prążek położony bliżej od początku żelu (ścieżki nr 5 i 6 na żelu).
W wyniku skrzyżowania dwóch homozygot rodzicielskich o genotypach Per1FPer1F i Per1SPer1S otrzymujemy heterozygotyczne pokolenie F1 o genotypie Per1FPer1S. U heterozygot allel Per1F jest odpowiedzialny za wytwarzanie szybko migrujących cząsteczek peroksydazy (45 mm), zaś allel Per1S warunkuje wytwarzanie cząsteczek wolno migrujących (35 mm). Heterozygoty będą wytwarzały dwa typy cząsteczek peroksydazy, które będą widoczne na żelu jako dwa prążki (ścieżki nr 2-4 na żelu). Fenotypy dwuprążkowe w F2 pochodzą od heterozygot F2 i podobnie jak dwuprążkowe fenotypy F1 są efektem zjawiska kodominacji.
Obserwowane w pokoleniu F2 fenotypy jednoprążkowe odpowiadają genotypom Per1FPer1F oraz Per1SPer1S, a fenotypy dwuprążkowe, genotypowi Per1FPer1S.
Stosunki rozszczepień w F2 będą zgodne z częstościami genotypów zakładanych przez Pierwsze Prawo Mendla i będą wynosiły 1:2:1, z podziałem na rośliny Per1FPer1F (jeden prążek o ruchliwości 45 mm), Per1FPer1S (dwa prążki o ruchliwości 45 mm i 35 mm) i Per1SPer1S (jeden prążek o ruchliwości 35 mm).
Zadania
1. Na rysunku przedstawiono obraz rozdziału elektroforetycznego dysmutazy ponadtlenkowej (SOD) w pokoleniu F2 Lolium perenne. Ujawniono 2 geny, Sod1 i Sod2 kodujące formy tego enzymu. W pokoleniu F2 zaobserwowano występowanie fenotypów jedno- i dwuprążkowych pod względem genu Sod1. Wyjaśnij skąd wzięły się fenotypy dwuprążkowe w F2, podaj genotypy dla wzorów roślin F2 widocznych na rysunku. Ustal genotypy rodziców i pokolenia F1.
2. W pokoleniu F2 otrzymanym ze skrzyżowania Lolium perenne x L. multiflorum przeprowadzono analizę sekwencji mikrosatelitarnych (SSR) wykorzystując w reakcji PCR specyficzne startery. Za pomocą startera C11 ujawniono wzory jedno i dwuprążkowe przedstawione na rysunku. Oznaczając locus mikrosatelitarne jako C11, a jego allele jako C11a i C11b, wyjaśnij dziedziczenie się ujawnionych prążków. Podaj genotypy rodziców i F1.
3. Kamelia to krzew o lśniących, ciemnozielonych liściach i okazałych kwiatach, niegdyś będących wyznacznikiem dobrego stylu. Roślina pochodzi z wschodniej i południowo-wschodniej Azji, a w Europie znana między innymi za sprawą powieści A. Dumas „Dama kameliowa”. Czerwono-biały kolor kwiatów (część płatków jest czerwona, a część biała) tej rośliny jest wynikiem kodominacji. Pewien ogrodnik skrzyżował rośliny o czerwono-białych kwiatach w celu wyhodowania stabilnych, czyli, nierozszczepiających się w potomstwie, form białych. Czy postąpił słusznie? Czy w wyniku tego krzyżowania można również uzyskać formy stabilne czerwone?
4. Pewna kobieta o grupie krwi AB urodziła dziecko o grupie krwi A. Który z wymienionych mężczyzn był ojcem dziecka – a) o grupie krwi A, b) o grupie B?
5. Jeżeli każdy z rodziców ma grupę krwi AB, to jakich genotypów i z jakim prawdopodobieństwem możemy spodziewać się wśród ich dzieci?

24
F:
P:X
ŻółtaBb
ŻółtaBb
B bbB
Bb
Bb bb
B
B
b
bFenotypy w potomstwie:•Żółte:•Szarobrązowe:
21
Osobniki o genotypie powinny mieBB ćżółtą sierść, jednakże gen w stanie homozygotycznym jest letalny i powodujeśmierć w stanie embrionalnym.
B
Genotypy •
w potomstwie:Homozygota dominująca, :BB
•• Heterozygoty, Homozygota recesywna,
Bbbb
121
AAALLLLLLEEELLLEEE LLLEEETTTAAALLLNNNEEE
NNNiiieeekkktttóóórrreee aaalllllleeellleee pppooowwwoooddduuujjjąąą śśśmmmiiieeerrrććć ooorrrgggaaannniiizzzmmmuuu... SSSąąą tttooo aaalllllleeellleee llleeetttaaalllnnneee... NNNiiieeekkktttóóórrreee aaalllllleeellleee ooobbbnnniiiżżżaaajjjąąą żżżyyywwwoootttnnnooośśśććć ooorrrgggaaannniiizzzmmmuuu... SSSąąą tttooo aaalllllleeellleee sssuuubbbllleeetttaaalllnnneee...
Przykład 1
Skrzyżowano 5 par myszy o żółtej sierści. W potomstwie otrzymano 10 myszy szarobrązowych i 20 myszy żółtych. a) Jakie były genotypy krzyżowanych osobników? b) Jak dziedziczy się cecha barwy sierści u myszy? c) Jakie potomstwo i w jakich proporcjach wystąpi po skrzyżowaniu myszy szarobrązowej z myszą żółtą?
ROZWIĄZANIE
W potomstwie wystąpiło rozszczepienie na formy szarobrązowe i żółte, co świadczy o krzyżowaniu osobników heterozygotycznych względem genu warunkującego barwę sierści u myszy. Żółta barwa u heterozygot wskazuje, że allel ją warunkujący był dominujący. Możemy więc oznaczyć literą B allel warunkujący barwę żółtą, a literą b allel kodujący barwę szarobrązową. Genotypy krzyżowanych osobników to Bb x Bb.

5. ALLELE LETALNE
25
S r ąza obr zowabb
b
F:
P:X
B b
ŻółtaBb
B
b
b
Bb bb Fenotypy w potomstwie:•Żółte:•Szarobrązowe:
11
Genotypy w potomstwie:•• Heterozygoty, Homozygota recesywna,
Bbbb
11
Zgodnie z Pierwszym Prawem Mendla, w wyniku krzyżowania dwóch heterozygot Bb powinniśmy otrzymać w potomstwie osobniki żółte i szarobrązowe w stosunku 3:1. W odniesieniu do genotypów powinniśmy otrzymać homozygoty dominujące, heterozygoty i homozygoty recesywne w stosunku 1:2:1. Tymczasem stosunek osobników żółtych do szarobrązowych wynosił 2:1. Takie proporcje mogą świadczyć, że brakuje któregoś z genotypów o przewidywanej częstości ¼, czyli brakuje jednego typu homozygot. Ponieważ szarobrązowe myszy są homozygotami recesywnymi, to musi brakować homozygoty dominującej. W tym przypadku formy te obumierają we wczesnych stadiach embrionalnych. Allel dominujący w stanie homozygotycznym jest letalny ponieważ powoduje śmierć organizmu. Większość genów może mutować w kierunku alleli letalnych. Oznacza to, że większość genów jest absolutnie niezbędna do zachowania funkcji życiowych organizmu. Recesywne allele letalne mogą być utrzymywane w populacji. Dominujące allele letalne mogą być utracone w ciągu jednego pokolenia, jeżeli powodują śmierć osobnika przed fazą rozrodczą. Jeżeli działają w okresie późniejszym, po wydaniu potomstwa, dominujące allele letalne utrzymują się w populacji. Krzyżowanie myszy szarobrązowej z myszą żółtą jest krzyżowaniem homozygoty recesywnej z heterozygotą pod względem jednego genu. Jest to przykład krzyżówki testowej. W potomstwie wystąpią osobniki żółte i szarobrązowe w stosunku 1:1.
Przykład 2
U petunii występują rośliny z ciemnozielonymi, lśniącymi liśćmi. Ze względu na atrakcyjny wygląd rośliny te są poszukiwane przez hodowców. Niestety próby otrzymania linii czystych o ciemnozielonych liściach na ogół kończą się niepowodzeniem. Po skrzyżowaniu roślin o ciemnozielonych liściach zawsze otrzymuje się 50% roślin o liściach ciemnozielonych i 25% roślin o jasnych liściach. Wyjaśnij dziedziczenie ciemnozielonych liści u petunii.
ROZWIĄZANIE
Rośliny o ciemnozielonych liściach są heterozygotami (zapiszmy je jako Aa) pod względem genu kodującego barwę liści, ponieważ po skrzyżowaniu zawsze uzyskuje się rozszczepienie w potomstwie.
W wyniku skrzyżowania 2 heterozygot o ciemnozielonych liściach uzyskuje się rozszczepienie 2:1. Brakuje jednego z genotypów, a mianowicie homozygot dominujących AA o ciemnozielonych liściach. Allel A w stanie homozygotycznym jest letalny. Rośliny o jasnozielonych liściach to homozygoty recesywne, aa.

5. ALLELE LETALNE
26
W rzeczywistości efekt letalny allela A u petunii jest efektem bliskiego położenia (sprzężenia) genu warunkującego barwę liści i innego genu L, warunkującego istotne funkcje fizjologiczne. U roślin o ciemnozielonych liściach, w genie L występuje recesywna mutacja (allel l) o efekcie letalnym. Homozygoty o ciemnozielonych liściach mają genotyp AAll, i właśnie obecność allela ll w stanie homozygotycznym jest odpowiedzialna za obumieranie homozygot AA we wczesnym stadium rozwojowym. W potomstwie heterozygot o ciemnozielonych liściach, bardzo rzadko udaje się uzyskać osobniki zrekombinowane (w wyniku crossing‐over) o genotypie AALL. Osobniki te mają ciemnozielone liście i są homozygotami dominującymi, dzięki czemu nie rozszczepiają w potomstwie. Te osobniki wykorzystywane są do tworzenia odmian o ciemnozielonych liściach. Zadania
1. Rośliny rzodkiewnika, Arabidopsis thaliana posiadają szybkomigrujący wariant dehydrogenazy izocytrynianowej kodowany przez allel IdhF. Z liści A. thaliana założono kulturę in vitro. W pierwszym etapie powstała tkanka kalusowa, z której następnie zregenerowano kilkadziesiąt roślin. U jednej z nich stwierdzono dwuprążkowy fenotyp dehydrogenazy izocytrynianowej, przy czym nowy prążek był wolnomigrujący. Założono, że w trakcie kultury w locus Idh zaszła mutacja punktowa, odpowiedzialna za pojawienie się nowego, wolnomigrującego prążka. Zmutowany allel oznaczono jako IdhS. Roślinę o fenotypie dwuprążkowym, o przypuszczalnym genotypie IdhFIdhS samozapylano w celu otrzymania homozygot IdhSIdhS. W potomstwie otrzymano tylko formy posiadające jeden szybkomigrujący prążek oraz formy o fenotypie dwuprążkowym. Ponownie samozapylono wszystkie otrzymane formy o fenotypie dwuprążkowym. Niestety znowu nie udało się otrzymać osobników IdhS IdhS. Wyjaśnij przyczynę niepowodzenia. W jakich proporcjach występowały fenotypy jednoprążkowe, szybkomigrujące oraz dwuprążkowe w potomstwie samozapylanych heterozygot.
2. U brzoskwini owłosiona skórka jest dominująca (F) w stosunku do gładkiej (nektaryny) skórki (f). Brzoskwinie z owalnymi gruczołami u podstawy liści są zawsze heterozygotyczne względem genu O, natomiast u homozygot recesywnych (oo) gruczoły nie występują. Podaj fenotypy i genotypy pokolenia F1 i F2 uzyskanego po skrzyżowaniu odmiany brzoskwini o owłosionej skórce, pozbawionej gruczołów u postawy liści z odmianą o gładkiej skórce (nektaryną) i owalnych gruczołach u podstawy liści.
3. Jeden ze szczepów Drosophila melanogaster ma oczy o kolorze śliwkowym. Po skrzyżowaniu dwóch much z tego szczepu o śliwkowych oczach otrzymuje się 2/3 much o oczach śliwkowych i 1/3 o oczach czerwonych (dzikich). Inny ze szczepów D. melanogaster charakteryzuje się krótkimi szczecinkami zamiast normalnych, długich szczecinek. Heterozygoty mają szczecinki krótkie, przy czym allel warunkujący krótkie szczecinki jest letalny w stanie homozygotycznym. Podaj genotypy i fenotypy potomstwa otrzymanego po skrzyżowaniu muchy o oczach śliwkowych z muchą o szczecinkach krótkich?
4. Po skrzyżowaniu dwóch kotów o krótkich ogonach otrzymano 6 kotów o ogonach krótkich i 3 koty o ogonach normalnych. Ustal genotypy kojarzonych kotów oraz wyjaśnij sposób dziedziczenia się długości ogonów u kotów.
5. U owiec karakułowatych gen S warunkuje siwą barwę i dominuje nad genem barwy czarnej s, ale w stanie homozygotycznym daje efekt letalny. Jakich genotypów i fenotypów należy oczekiwać kojarząc owce czarne?
6. Gen p dający u lisów srebrzystą barwę włosa zmutował w dwóch kierunkach: allela P - dającego barwę platynową oraz allela Pb dającego lisy białopyskie. Zmutowane geny w stanie homozygotycznym dają efekt letalny. Heterozygoty PPb również nie są zdolne do życia i giną przed urodzeniem. Jakiego potomstwa i w jakich ilościach można oczekiwać po skojarzeniu a) dwóch lisów platynowych, b) dwóch lisów białopyskich, c) dwóch lisów srebrzystych, d) platynowego i białopyskiego, e) platynowego i srebrzystego, f) białopyskiego i srebrzystego?

27
AAALLLLLLEEELLLEEE WWWIIIEEELLLOOOKKKRRROOOTTTNNNEEE
AAAlllllleeellleee wwwiiieeellloookkkrrroootttnnneee --- www pppooopppuuulllaaacccjjjiii wwwyyyssstttęęępppuuujjjeee 333 llluuubbb wwwiiięęęccceeejjj aaalllllleeellliii dddaaannneeegggooo gggeeennnuuu...
PPPooollliiimmmooorrrfffiiizzzmmm --- www pppooopppuuulllaaacccjjjiii wwwyyyssstttęęępppuuujjjąąą cccooo nnnaaajjjmmmnnniiieeejjj 222 aaalllllleeellleee dddaaannneeegggooo gggeeennnuuu...
Przykład 1
Czy możliwe jest aby w rodzinie złożonej z rodziców i dwojga dzieci każda osoba miała inną grupę krwi? Uzasadnij odpowiedź na schemacie.
ROZWIĄZANIE
Grupy krwi u człowieka, A, B, AB i 0 determinowane są allelami wielokrotnymi. Allel IA jest odpowiedzialny za wytworzenie antygenu krwinkowego A, allel IB za wytworzenie antygenu B, natomiast trzeci allel i jest odpowiedzialny za brak jakiegokolwiek antygenu krwinkowego.
Allele IA i IB są kodominujące, czyli każdy z nich daje efekt fenotypowy w heterozygocie. Oba allele są dominujące względem allela i. Grupa krwi A występuje u homozygot dominujących IAIA oraz heterozygot IAi. Grupa krwi B u homozygot dominujących IBIB oraz heterozygot IBi. Osobniki o genotypie ii mają grupę krwi 0.
Jeżeli rodzice mają grupę krwi A i B, to dzieci mogą mieć grupę AB i 0 (schemat poniżej). Podobnie, jeżeli rodzice mają grupę krwi AB i 0, to dzieci mogą mieć grupę krwi A i B.
Allele IA, IB i i występują w populacji ludzkiej, a więc gen I jest polimorficzny. Rozkład częstości ma charakter klinalny i nieciągły, co odzwierciedla złożoną ewolucję człowieka. Częstość występowania allela IB jest najniższa (pB = 0,16). Allel ten jest obecny w Centralnej Azji, a najrzadziej występuje w Ameryce i Australii. Częstość allela IA wynosi 0,21. Najczęściej spotyka się go w małych, izolowanych populacjach (Indianie z Montany, Aborygeni, Północna Skandynawia). Allel i występuje często (pi = 0,63), natomiast grupa krwi 0 jest charakterystyczna dla Centralnej i Południowej Ameryki (blisko 100% populacji), Aborygenów i Europy Zachodniej, zwłaszcza potomków Celtów. Rzadko spotyka się ją w Europie Wschodniej, gdzie przeważa grupa B.
Grupa A
Grupa AB Grupa 0
F:
P:
IA i IB i
X

6. ALLELE WIELOKROTNE
28
Zadania
1. Na podstawie uzyskanych wyników krzyżówek ustal kolejność dominowania alleli warunkujących kolor kwiatu u wyżlinu:
czerwony x brązowy = czerwony żółty x kremowy = żółty brązowy x pomarańczowy = brązowy zielony x biały = biały pomarańczowy x żółty = pomarańczowy kremowy x biały = kremowy.
2. U Anthirrinum majus o zabarwieniu kwiatów decyduje seria alleli wielokrotnych w następującej kolejności dominowania:
allel dziki - kwiaty czerwone - A; allel niva - kwiaty paskowane - an; allel palida - kwiaty pstrokate - ap; allel alba - kwiaty białe - aa.
Podaj wszystkie możliwe genotypy rodziców oraz proporcje, w jakich pojawią się w ich potomstwie różne fenotypy, jeśli po skrzyżowaniu:
a) formy palida z formą niva uzyskano rozszczepienie na palida, niva i alba; b) formy dzikiej z formą niva uzyskano rozszczepienie na czerwone, niva i palida; c) formy niva z formą niva uzyskano rozszczepienie na niva i palida; d) formy czerwonej z formą czerwoną uzyskano rozszczepienie na czerwone i niva.
3. Umaszczenie u królików uwarunkowane jest szeregiem alleli wielokrotnych, które pod względem dominowania dadzą się uszeregować w następującej kolejności: C - umaszczenie jednolicie ciemne, Cch - umaszczenie szynszylowe, Cm - umaszczenie kuny, Ch- umaszczenie himalajskie, c - albinosy. Skojarzono ciemnego jednolitego samca z himalajskimi samicami i otrzymano królika ciemnego, królika himalajskiego i królika albinotycznego. Podaj genotypy rodziców oraz prawdopodobne proporcje podanych fenotypów, gdybyśmy mieli wiele miotów od tych samych zwierząt.
4. Barwa świnek morskich zależy od szeregu alleli wielokrotnych podanych w kolejności dominowania: C - kolor intensywnie rudy, Ck - ciemna sepia, Cd. - sepia, Cr - sepia jasna, c - albinos. W potomstwie osobników intensywnie rudych i sepia otrzymano potomstwo intensywnie rude, sepia i jasna sepia. Jakie były genotypy rodziców i potomstwa?
5. U kotów barwa sierści zależy od szeregu alleli wielokrotnych składającego się z genów: C - futerko ciemne, cr - srebrzyste, cs - syjamskie. Srebrzysta kotka dała miot złożony z kociąt srebrzystych, ciemnych i syjamskich. Jaki był genotyp i fenotyp ojca tych kociąt?
6. Czy dziecko z grupą krwi 0 może pochodzić z małżeństwa kobiety i mężczyzny z grupą krwi A? Uzasadnij odpowiedź.
7. W małżeństwie mężczyzny z grupą krwi 0, M z kobietą o grupie A, M urodziło się dziecko z grupą krwi A, MN. Mężczyzna twierdzi, że dziecko nie jest jego i oddał sprawę do sądu, który zlecił badanie grup krwi. Jaki będzie wyrok sądu? Uzasadnij odpowiedź.
8. Jeżeli matka i dziecko mają grupę krwi 0, to jakiej grupy krwi na pewno nie ma ojciec?
9. Czy dziecko z grupą krwi AB może pochodzić z małżeństwa kobiety z grupą krwi A i mężczyzny z grupą krwi 0? Uzasadnij odpowiedź.

29
WWWSSSPPPÓÓÓŁŁŁDDDZZZIIIAAAŁŁŁAAANNNIIIEEE GGGEEENNNÓÓÓWWW
WWWiiieeellleee gggeeennnóóówww mmmooożżżeee wwwpppłłłyyywwwaaaććć nnnaaa jjjeeedddnnnąąą ccceeeccchhhąąą...
Przykład 1
Skrzyżowano kurę o grzebieniu różyczkowym z kurą o grzebieniu groszkowym. W F1 otrzymano kury o grzebieniu orzeszkowym, a w F2 otrzymano 271 kur o grzebieniu orzeszkowym, 89 kur o grzebieniu różyczkowym, 92 kury o grzebieniu groszkowym i 30 kur grzebieniu pojedynczym. Jak dziedziczy się kształt grzebienia u kur? Podaj genotypy rodziców.
ROZWIĄZANIE
Krzyżowane kury różniły się jedną cechą – kształtem grzebienia. Należałoby więc przypuszczać, że cecha ta zależy od jednego genu i dziedziczy się zgodnie z Pierwszym Prawem Mendla. Przy takim założeniu w F1 powinniśmy uzyskać formy o kształcie grzebienia któregoś z rodziców lub pośrednim. Tymczasem uzyskujemy nową cechę – grzebień orzeszkowy. Pojawienie się nowej cechy w pokoleniach F1 lub/i F2 otrzymanych ze skrzyżowania homozygot (linii czystych) różniących się jedną cechą może świadczyć, iż analizowana cecha jest uwarunkowana więcej niż jednym genem. Jeżeli kształt grzebienia dziedziczyłby się zgodnie z Pierwszym Prawem Mendla to w pokoleniu F2 powinniśmy otrzymać dwie (lub 3) klasy fenotypowe w stosunku 3:1 (lub 1:2:1). Tymczasem otrzymane wartości odpowiadają w przybliżeniu stosunkowi 9:3:3:1, charakterystycznemu dla dwóch genów, które dziedziczą się niezależnie.
Załóżmy, że kształt grzebienia jest uwarunkowany dwoma genami R, r oraz P, p. Jednolite pokolenie F1
otrzymuje się tylko w wyniku krzyżowania dwóch homozygot pod względem obu genów. F1 jest heterozygotą o genotypie RrPp. Wiadomo też, że ma ono grzebień orzeszkowy.

7. WSPÓŁDZIAŁANIE GENÓW
30
Na podstawie pokolenia F1 można stwierdzić, że grzebień orzeszkowy powstaje, gdy obecny jest przynajmniej jeden allel dominujący w każdym locus. Żaden z rodziców nie miał grzebienia orzeszkowego, a więc żaden nie mógł posiadać jednocześnie allela dominującego R i P. Jeden z rodziców, o grzebieniu różyczkowym miał genotyp RRpp, a drugi o grzebieniu groszkowym rrPP.
W pokoleniu F2 otrzymujemy osobniki o fenotypie F1 – z grzebieniami orzeszkowymi, o fenotypach rodzicielskich – z grzebieniami różyczkowymi i groszkowymi oraz pojawiają się osobniki o nowym fenotypie – z grzebieniami pojedynczymi. Proporcje poszczególnych klas genotypowych wynoszą 9:3:3:1.
Jeżeli w wyniku krzyżowania homozygot (linii czystych) różniących się jedną cechą otrzymujemy w pokoleniu F2 stosunek rozszczepień 9:3:3:1, charakterystyczny dla niezależnego dziedziczenia dwóch genów, to świadczy, że cecha uwarunkowana jest współdziałaniem dwóch genów. Przykład 2
Po skrzyżowaniu dwóch karłowych odmian ogórka otrzymano w F1 rośliny wysokie. Po samozapyleniu, w F2
otrzymano 97 roślin wysokich oraz 79 roślin o wzroście karłowym. Podaj genotypy i fenotypy rodziców oraz F1. Jak dziedziczy się wzrost u ogórka?
ROZWIĄZANIE Pokolenie F1 jest jednolite, co świadczy, że krzyżowano linie czyste (homozygotyczne).
Pokolenie rodzicielskie miało wzrost karłowy, natomiast w pokoleniu F1 pojawił się całkowicie nowy fenotyp – wzrost wysoki. Pojawienie się nowego fenotypu świadczy, że na wzrost u ogórka wpływają co najmniej dwa współdziałające geny.
Aby ustalić, w jaki sposób geny współdziałają ze sobą w warunkowaniu wzrostu należy ustalić stosunek rozszczepień w pokoleniu F2. W omawianym przypadku rośliny wysokie i karłowe wystąpiły w proporcjach 9:7.
Otrzymane rozszczepienie fenotypów 9:7 możemy porównać z proporcjami fenotypów w pokoleniu F2 wynoszącymi 9:3:3:1 i które wystąpiłyby w przypadku niezależnego dziedziczenia dwóch genów zgodnie z Drugim Prawem Mendla. Załóżmy, że wzrost jest uwarunkowany genami A i B. Możemy więc przyjąć, że:

7. WSPÓŁDZIAŁANIE GENÓW
31
osobniki o wzroście wysokim, stanowiące 9/16 pokolenia F2 mają genotyp A B , oznacza to, że wzrost wysoki występuje tylko w obecności alleli dominujących w każdym z dwóch loci.
osobniki karłowe, stanowiące 7/16 potomstwa mają genotypy A bb, aaB oraz aabb; w uproszczeniu można przyjąć, że powstają one z połączenia 3 klas fenotypowych obserwowanych przy niezależnym dziedziczeniu i stanowiących odpowiedni 3/16, 3/16 i 1/16 pokolenia F2.
Karłowi rodzice musieli mieć genotypy AAbb i aaBB, natomiast wysokie F1 AaBb. Współdziałanie obserwowane u ogórka to przykład komplementacji. Komplementacja to takie współdziałanie genów, w którym do powstania danego fenotypu niezbędne są allele dominujące we wszystkich współdziałających loci. Brak allela dominującego w którymkolwiek locus powoduje ujawnienie się zmienionego‐ zmutowanego fenotypu. Test komplementacji pozwala sprawdzić czy mutacje są alleliczne, to znaczy czy zaszły w tym samym genie (locus), czy też są niealleliczne i zaszły w różnych genach (loci). Jeżeli w wyniku krzyżowania dwóch mutantów otrzymujemy w F1 formę zmutowaną to mutacje są alleliczne. Jeżeli w wyniku krzyżowania dwóch mutantów w F1 otrzymujemy formę niezmutowaną to mutacje są niealleliczne.
Przykład 3
Skrzyżowano dwie odmiany dyni o owocach kulistych. W F1 otrzymano tylko dynie o owocach płaskich, natomiast w F2 dynie o owocach płaskich, kulistych i wydłużonych w stosunku 9:6:1. Ustal jak dziedziczy się kształt owoców u dyni. Podaj genotypy rodziców i F1.
ROZWIĄZANIE
Pokolenie F1 jest jednolite, a więc krzyżowano dwie linie czyste.
W przykładzie segreguje jedna cecha, jednakże zarówno w F1 jak i w F2 pojawiają się nowe fenotypy – płaskie dynie w F1 i dynie wydłużone w F2. Świadczy to o uwarunkowaniu kształtu owocu u dyni przez co najmniej dwa współdziałające geny.
Załóżmy, że kształt owocu dyni jest uwarunkowany genami A, a i B, b. Pokolenie F1 powstało w wyniku krzyżowania linii czystych, miało więc genotyp AaBb. Ponieważ osobniki F1 miały owoce płaskie, można przyjąć, że ten kształt owocu powstaje, gdy w obu współdziałających loci obecne są allele dominujące.
Stosunki rozszczepień w F2 można przyrównać do stosunku 9:3:3:1, przy czym osobniki płaskie stanowiące 9/16 to osobniki o genotypie A B , osobniki kuliste stanowiące 6/16 powstają „z połączenia dwóch klas” – klasy o częstości 3/16 i genotypie aaB oraz klasy o częstości 3/16 i genotypie A bb. Owoc kulisty powstaje gdy jest obecny allel dominujący tylko w jednym z dwóch loci. Brak allela dominującego powoduje powstanie owoców wydłużonych. Mają one genotyp aabb i stanowią 1/16 F2. Stosunek rozszczepień fenotypów w F2 9:3:3:1 jest charakterystyczny dla niezależnego dziedziczenia dwóch genów i obserwujemy go w F2, gdy krzyżowane osobniki różniły się dwoma cechami. Jeżeli rozszczepienie fenotypowe 9:3:3:1 lub jego modyfikacje występują w pokoleniu F2 uzyskanym w wyniku krzyżowania form różniących się jedną cechą, to świadczy o współdziałaniu genów. W przypadku współdziałania genów modyfikacje stosunków rozszczepień fenotypowych w F2 mogą być następujące – 9:7 (komplementacja), 9:6:1 (addytywne współdziałanie genów dominujących), 9:3:4 (recesywna epistaza), 12:3:1 oraz 13:3 (epistaza genów dominujących). Współdziałanie genów może także polegać na sumowaniu lub mnożeniu się efektów alleli dominujących we wszystkich loci. Mówimy wówczas o współdziałaniu addytywnym (geny kumulatywne) lub multiplikatywnym, które są charakterystyczne dla genów warunkujących cechy ilościowe. Współdziałania te mogą być dalej komplikowane poprzez kombinacje działania addytywnego, multiplikatywnego i dominacji.

7. WSPÓŁDZIAŁANIE GENÓW
32
Zadania
1. U groszku pachnącego gen A warunkuje czerwoną barwę kwiatu i jest komplementarny do genu B. Recesywne allele tych genów powodują powstanie białych kwiatów. Po skrzyżowaniu groszku o kwiatach czerwonych z groszkiem o kwiatach białych otrzymano 94 rośliny o kwiatach czerwonych i 277 roślin o kwiatach białych. Podaj genotypy roślin użytych do krzyżowań.
2. Podaj wszystkie możliwe genotypy dyni o owocach płaskich, która po skrzyżowaniu testowym dała rozszczepienie 1:1 na dynie o owocach płaskich i kulistych, wiedząc, że kształt owocu u dyni jest wynikiem współdziałania genów.
3. Skrzyżowano dwie rośliny pszenicy o wzroście karłowym. W pokoleniu F1 uzyskano tylko rośliny wysokie. Po samozapyleniu F1, w F2 otrzymano 143 rośliny karłowe i 98 roślin wysokich. Podaj genotypy rodziców. Wyjaśnij sposób dziedziczenia się karłowatości u pszenicy oraz sprawdź, stosując test chi2 czy postawiona hipoteza jest słuszna.
4. Skrzyżowano ogórek o pomarańczowym zabarwieniu owocu z rośliną o barwie owocu żółtej. F1 uzyskane z tej krzyżówki miało owoce zielone, a w F2 wystąpiły:
264 rośliny o owocach zielonych, 87 roślin o owocach pomarańczowych, 92 roślin o owocach żółtych, 26 roślin o owocach kremowych. Ustal w jaki sposób dziedziczy się cecha barwy owocu u ogórka. Podaj genotypy rodziców i pokolenia F1.
5. Podaj genotypy i fenotypy dwóch roślin jęczmienia, które skrzyżowane ze sobą dały w potomstwie:
186 roślin o plewach zielonych, 62 rośliny o plewach pomarańczowych, 84 rośliny o plewach białych. Jak dziedziczy się barwa plew u jęczmienia? Podaj genotypy krzyżowanych roślin.
6. U dyni para alleli Y, y warunkuje żółtą względnie zieloną barwę owocu, a para alleli W, w barwę białą względnie zieloną, przy czym gen W jest epistatyczny w stosunku do Y. Para alleli M, m odpowiada za prążkowaną względnie jednolicie zabarwioną powierzchnię owocu. Jaki fenotyp i genotyp będzie mieć pokolenie F1 uzyskane ze skrzyżowania dyni o owocu zielonym, prążkowanym z rośliną o owocu białym, zabarwionym jednolicie? Jaki będzie wynik krzyżówki testowej?
7. Czy dwa mutanty petunii o kwiatach białych są alleliczne czy niealleliczne jeżeli po ich skrzyżowaniu w F1
uzyskano rośliny o kwiatach białych?
8. Jaki będzie fenotyp F1 oraz stosunek rozszczepień fenotypowych w F2 uzyskany po skrzyżowaniu dwóch erektoidalnych mutantów jęczmienia (oba mutanty były recesywne w stosunku do normalnego, niesztywnoowłosionego fenotypu rośliny), w przypadku gdy:
a) geny odpowiedzialne za erektoidalność każdego z mutantów są niealleliczne? b) geny odpowiedzialne za erektoidalność każdego z mutantów są alleliczne?

33
GGGEEENNNYYY SSSPPPRRRZZZĘĘĘŻŻŻOOONNNEEE ZZZ PPPŁŁŁCCCIIIĄĄĄ
GGGeeennnyyy zzzllloookkkaaallliiizzzooowwwaaannneee nnnaaa ccchhhrrrooommmooosssooommmaaaccchhh pppłłłccciii tttooo gggeeennnyyy sssppprrrzzzęęężżżooonnneee zzz pppłłłccciiiąąą... SSSaaammmiiiccceee iii sssaaammmccceee mmmooogggąąą rrróóóżżżnnniiiććć sssiiięęę fffeeennnoootttyyypppooowwwooo pppoooddd wwwzzzggglllęęędddeeemmm kkkooodddooowwwaaannnyyyccchhh ppprrrzzzeeezzz nnniiieee ccceeeccchhh...
Przykład 1
Skrzyżowano samicę Drosophila melanogaster o ciele żółtym z samcem o ciele szarym (typ dziki). W pokoleniu F1
samice miały szarą barwę ciała (typ dziki), a samce ciało żółte. W pokoleniu F2, zarówno wśród samic jak i samców wystąpiły osobniki o ciele szarym i żółtym w równych proporcjach.
a) Jak dziedziczy się cecha barwy ciała u D. melanogaster. Podaj genotypy rodziców oraz pokolenia F1. b) Jakie będzie pokolenie F1 i F2 w krzyżówce odwrotnej?
ROZWIĄZANIE – PUNKT A
Skrzyżowano osobniki różniące się jedną cechą, barwą ciała. W pokoleniu F1 otrzymano osobniki o barwie ciała żółtej i szarej. Porównywalna liczebność form żółtych i szarych (prawdopodobieństwo wystąpienia samicy jak i samca wynosi 0,5) mogłaby wskazywać, zgodnie z Pierwszym Prawem Mendla, że krzyżowano heterozygotę z homozygotą recesywną. Jednakże, w takim przypadku osobniki o ciele żółtym i szarym wystąpiłyby zarówno wśród samców jak i samic. Tymczasem, otrzymano tylko szare samice i tylko żółte samce. Różny fenotyp samic i samców w F1 świadczy, że gen warunkujący barwę ciała zlokalizowany jest na chromosomie płci (rys., str. 34) U D. melanogaster płeć jest determinowana przez chromosomy płci X, Y, które różnią się morfologią. Samice mają zawsze dwa identyczne chromosomy, XX. Jest to płeć homogametyczna. Samce mają dwa różne chromosomy, XY. Jest to płeć heterogametyczna. Geny sprzężone z płcią znajdują się na chromosomie X. Chromosom Y nie zawiera genów determinujących cechy – jest „pusty”. Dlatego w odniesieniu do genów zlokalizowanych na chromosomie X, samce zawsze mają tylko jedną kopię – są hemizygotami (półzygotami). Z tego względu u samców zawsze ujawniają się allele recesywne genów sprzężonych z płcią.
Krzyżowane formy były homozygotami. Samica pokolenia rodzicielskiego posiada allel żółtej barwy ciała na obu chromosomach X. Jej genotyp możemy zapisać jako yy lub XyXy. Samiec posiada allel szarej barwy ciała na chromosomie X oraz chromosom Y. Jego genotyp można zapisać jako y+Y lub Xy+Y. Samica wytwarza jeden typ gamet – z chromosomem X i zlokalizowanym na nim allelem żółtej barwy (y), natomiast samiec wytwarza dwa typy gamet – z chromosomem X niosącym allel szarej barwy ciała (y+) oraz z chromosomem Y.
W wyniku połączenia gamet samicy i gamety samca z chromosomem X w F1 powstają samice, które są heterozygotami pod względem genu barwy ciała (y+y). Od matki otrzymały allel żółtej barwy ciała a od ojca allel barwy szarej. Szara barwa ciała heterozygotycznych samic F1, świadczy o dominacji allela warunkującego barwę szarą nad allelem barwy żółtej.
W wyniku połączenia gamet samicy z gametą samca zawierającą chromosom Y, w F1 powstają samce, u których fenotypowo ujawnia się allel zlokalizowany na chromosomie X otrzymanym od matki. W tym przypadku jest to allel żółtej barwy ciała.
Samice pokolenia F1 wytwarzają dwa typy gamet – z allelem szarej barwy ciała na chromosomie X (y+) oraz z allelem żółtej barwy ciała na chromosomie X (y). Samiec wytwarza gamety z allelem żółtej barwy ciała na chromosomie X (y) oraz gamety z chromosomem Y. W wyniku połączenia tych typów gamet w pokoleniu F2
powstają zarówno samice jak i samce o żółtej i szarej barwie ciała. Proporcje osobników o szarym i żółtym ciele wynoszą 1:1, zamiast spodziewanego stosunku mendlowskiego 3:1.
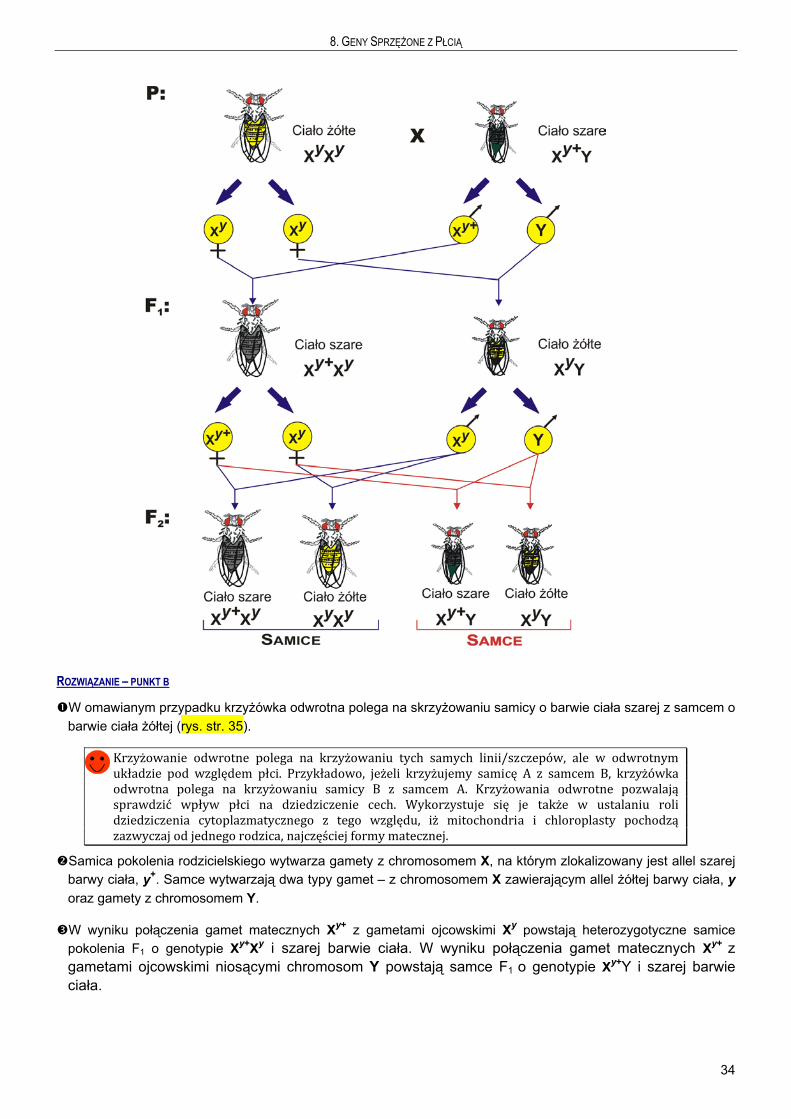
8. GENY SPRZĘŻONE Z PŁCIĄ
34
ROZWIĄZANIE – PUNKT B
W omawianym przypadku krzyżówka odwrotna polega na skrzyżowaniu samicy o barwie ciała szarej z samcem o barwie ciała żółtej (rys. str. 35). Krzyżowanie odwrotne polega na krzyżowaniu tych samych linii/szczepów, ale w odwrotnym układzie pod względem płci. Przykładowo, jeżeli krzyżujemy samicę A z samcem B, krzyżówka odwrotna polega na krzyżowaniu samicy B z samcem A. Krzyżowania odwrotne pozwalają sprawdzić wpływ płci na dziedziczenie cech. Wykorzystuje się je także w ustalaniu roli dziedziczenia cytoplazmatycznego z tego względu, iż mitochondria i chloroplasty pochodzą zazwyczaj od jednego rodzica, najczęściej formy matecznej. Samica pokolenia rodzicielskiego wytwarza gamety z chromosomem X, na którym zlokalizowany jest allel szarej barwy ciała, y+. Samce wytwarzają dwa typy gamet – z chromosomem X zawierającym allel żółtej barwy ciała, y oraz gamety z chromosomem Y.
W wyniku połączenia gamet matecznych Xy+ z gametami ojcowskimi Xy powstają heterozygotyczne samice pokolenia F1 o genotypie Xy+Xy i szarej barwie ciała. W wyniku połączenia gamet matecznych Xy+ z gametami ojcowskimi niosącymi chromosom Y powstają samce F1 o genotypie Xy+Y i szarej barwie ciała.

8. GENY SPRZĘŻONE Z PŁCIĄ
35
Samice pokolenia F1 wytwarzają dwa typy gamet – z chromosomem X i allelem szarej barwy ciała, y+ oraz z chromosomem X i allelem żółtej barwy ciała, y. Samce pokolenia F1 również wytwarzają dwa typy gamet – z chromosomem X i allelem szarej barwy ciała, y+ oraz z chromosomem Y.
W wyniku połączenia gamet F1 powstają samice F2, które mają szarą barwę ciała, a pod względem genotypu 50% samic to homozygoty dominujące, a 50% to heterozygoty. Samce pokolenia F2 są szare i żółte w stosunku 1:1. Jeżeli nie uwzględnimy płci, to stosunek osobników szarych i żółtych wynosi 3:1, jak w przypadku Pierwszego Prawa Mendla. Jednakże w przypadku dziedziczenia mendlowskiego, stosunek fenotypowy 3:1 byłby obserwowany zarówno wśród samic jak i samców. Tak więc, pomimo, iż na pierwszy rzut oka wydaje się, że mamy do czynienia z mendlowskim dziedziczeniem, dokładna analiza pokazuje, że pomiędzy płciami występują różnice pod względem barwy ciała (samice są szare, a samce szare i żółte) i te różnice jednoznacznie wskazują na odstępstwo od dziedziczenia mendlowskiego. Sprzężenie z płcią możemy poznać po różnicach w fenotypie samic i samców pod względem danej cechy. Dodatkowo na sprzężenie genu warunkującego daną cechę z płcią wskazują także różne wyniki krzyżowań odwrotnych. W przypadku genów zlokalizowanych na autosomach wyniki krzyżowań odwrotnych są identyczne. U D. melanogaster na chromosomie X zlokalizowane są także inne geny, w tym geny barwy oczu (w – białe oczy, v – oczy vermilion), gen odpowiedzialny za miniaturowe skrzydła (m) a także geny kodujące kształt szczecinek (f, sn). U człowieka płeć jest zdeterminowana obecnością chromosomów XX u kobiet i XY u mężczyzn. Na ludzkim chromosomie X zlokalizowany jest między innymi gen wywołujący daltonizm oraz gen odpowiedzialny za hemofilię. U kur płeć determinowana jest odwrotnie niż u człowieka, samice są XY, a samce XX. Na chromosomie X zlokalizowany jest gen kodujący żółtą bądź zieloną barwę nóg.

8. GENY SPRZĘŻONE Z PŁCIĄ
36
Zadania
1. W krzyżówce dwóch much Drosophila melanogaster o fenotypie dzikim otrzymano:
60 samic dzikich; 22 samice o skrzydłach dzikich i oczach glass; 31 samców dzikich; 30 samców o skrzydłach hair i oczach dzikich; 12 samców o skrzydłach dzikich i oczach glass; 10 samców o skrzydłach hair i oczach glass;
Podaj genotypy krzyżowanych much i wyjaśnij jak dziedziczą się obie cechy.
2. Samicę D. melanogaster o oczach white (w) skrzyżowano z samcami o skrzydłach vestigal (vg). W pokoleniu F1 uzyskano wszystkie samice dzikie oraz samce o oczach white. W F2 otrzymano:
301 much o oczach white; 298 much dzikich; 102 muchy będące podwójnym mutantem; 99 much o skrzydłach vestigal.
Podaj genotypy rodziców, F1 oraz ustal sposób dziedziczenia się tych cech.
3. W krzyżówce kury pasiastej o dziobie czerwonym (cecha dominująca) z kogutem pasiastym o dziobie żółtym otrzymano:
20 kogutów pasiastych o dziobie czerwonym; 21 kogutów pasiastych o dziobie żółtym; 10 kur pasiastych o dziobie czerwonym; 10 kur pasiastych o dziobie żółtym; 10 kur czarnych o dziobie czerwonym; 12 kur czarnych o dziobie żółtym.
Ustal genotypy krzyżowanych ze sobą osobników i wyjaśnij sposób dziedziczenia obu cech.
4. Po skrzyżowaniu pewnej kury z kogutem otrzymano 20 piskląt. Wszystkie koguty miały nogi żółte, natomiast połowa kur miała nogi zielone, a połowa żółte. Podaj genotypy i fenotypy krzyżowanego koguta i kury oraz wyjaśnij jak dziedziczy się barwa nóg u kur.
5. U kur barwa nóg jest cechą uwarunkowaną genem sprzężonym z płcią, przy czym nogi żółte (lub białe) dominują nad zielonymi. Koguta mieszańca po ojcu leghorn (nogi żółte) i matce zielononóżce skojarzono z kurą o nieznanym fenotypie. W wyniku krzyżowań wylęgło się 5 kurek leghorn, 6 kurek zielononóżek, 7 kogutków zielononogich i 6 kogutków leghorn. Jaki był fenotyp i genotyp krzyżowanych ptaków? Jakie rozszczepienie uzyskamy po skrzyżowaniu koguta mieszańca z kurą o przeciwnym fenotypie?
6. Czarne lub żółte zabarwienie sierści u kotów zależy od jednej pary alleli B, b, mającej locus na chromosomie X. Osobniki heterozygotyczne pod względem tego genu są łaciate. Jaki jest genotyp i fenotyp kota, który skojarzony z żółtą kotką dał 5 kociąt: 2 żółte i 3 łaciate? Określ płeć kociąt.
7. W pewnym małżeństwie, gdzie matka była daltonistką urodziła się zdrowa dziewczynka. Jakie jest prawdopodobieństwo urodzenia się w tym małżeństwie zdrowego chłopca, jeżeli wiadomo, że gen warunkujący daltonizm jest zlokalizowany w chromosomie X i jest on recesywny w stosunku do allela odpowiedzialnego za normalne rozróżnianie barw?
8. Pewien jasnooki mężczyzna poślubił kobietę o oczach ciemnych, której matka miała oczy jasne, a ojciec był chory na hemofilię. Podaj prawdopodobieństwo urodzenia się w tym małżeństwie zdrowego chłopca o oczach ciemnych.

©Kornelia Polok 37
GGGEEENNNYYY SSSPPPRRRZZZĘĘĘŻŻŻOOONNNEEE
GGGeeennnyyy,,, kkktttóóórrreee llleeeżżżąąą bbbllliiissskkkooo sssiiieeebbbiiieee iii dddzzziiieeedddzzziiiccczzząąą sssiiięęę wwwssspppóóólllnnniiieee sssąąą gggeeennnaaammmiii sssppprrrzzzęęężżżooonnnyyymmmiii... GGGeeennnyyy sssppprrrzzzęęężżżooonnneee llleeeżżżąąą nnnaaa jjjeeedddnnnyyymmm ccchhhrrrooommmooosssooommmiiieee...
Przykład 1. Sprzężenie całkowite
Po skrzyżowaniu dwóch roślin pomidora otrzymano rośliny o owocach gładkich, łodygach antocjanowych i liściach zielonych. W krzyżówce testowej roślin pokolenia F1 otrzymano:
45 roślin o owocach gładkich, łodygach antocjanowych, liściach mozaikowych (żółto-zielonych); 52 rośliny o owocach gładkich, łodygach zielonych, liściach mozaikowych (żółto-zielonych); 49 roślin o owocach owłosionych, łodygach zielonych, liściach zielonych; 44 rośliny o owocach owłosionych, łodygach antocjanowych, liściach zielonych.
Ustal sposób dziedziczenia się tych cech. Podaj wszystkie możliwe fenotypy i genotypy rodziców tej krzyżówki.
ROZWIĄZANIE
W wyniku skrzyżowania, dwóch roślin pomidora o przeciwstawnych cechach otrzymano jednolite potomstwo (wszystkie rośliny były identyczne), co oznacza, że krzyżowane rośliny były homozygotami, a potomstwo uzyskane z ich skrzyżowania – heterozygotami. Było to pokolenie F1.
W krzyżówce segregują trzy cechy – typ owocu, zabarwienie łodygi i zabarwienie liścia. Załóżmy, że cechy te uwarunkowane są odpowiednio genami A, B i D. Możemy zapisać „roboczy” genotyp pokolenia F1 jako AaBbDd. Na podstawie wyglądu pokolenia F1 ustalimy jakie cechy warunkowane są przez allele dominujące, a jakie przez recesywne.
F1 miało owoce gładkie, a więc allel dominujący A, jest odpowiedzialny za wystąpienie gładkich owoców, a allel recesywny, a za owoce owłosione.
Allel dominujący B odpowiada za antocjanowe zabarwienie łodygi (wystąpiło w F1), a allel recesywny b za zabarwienie zielone łodygi.
Liście zielone są uwarunkowane allelem dominującym, D, a liście mozaikowe – allelem recesywnym, d.
Pokolenie F1 skrzyżowano testowo, a więc było to krzyżowanie między heterozygotą AaBbDd i homozygotą recesywną aabbdd. Zgodnie z Drugim Prawem Mendla, w potomstwie krzyżówki testowej powinny wystąpić równe proporcje wszystkich przewidywanych fenotypów. W pierwszej kolejności należy określić, ile klas fenotypowych powinno powstać w krzyżówce AaBbDd x aabbdd, a następnie porównać oczekiwaną liczbę klas z liczbą otrzymanych klas fenotypowych.
Liczba klas fenotypowych w krzyżówce testowej odpowiada liczbie gamet wytworzonych przez heterozygotę. Obliczymy ją ze wzoru 2n, gdzie n oznacza liczbę genów w stanie heterozygotycznym. W naszym przykładzie mamy 3 geny w stanie heterozygotycznym. Liczba gamet wytwarzanych przez heterozygotę wynosi 23 = 8 i tyle samo wynosi oczekiwana liczba klas w krzyżówce testowej.
W potomstwie naszej krzyżówki testowej wystąpiły tylko 4 klasy fenotypowe. Brakuje 4 klas fenotypowych. Oznacza to, że geny kodujące analizowane cechy pomidora nie dziedziczą się niezależnie, ale są sprzężone.
.

9. GENY SPRZĘŻONE
©Kornelia Polok 38
W drugim etapie należy sprawdzić ile genów jest sprzężonych. Mogą być sprzężone wszystkie 3 geny lub tylko 2. W krzyżówce testowej otrzymano 4 klasy fenotypowe, co oznacza, że heterozygota wytwarzała 4 typy gamet. Heterozygota wytwarzała tyle gamet, ile powstałoby w przypadku 2 genów dziedziczących się niezależnie. Wynika z tego, że dwa geny kodujące cechy pomidora były sprzężone i tworzyły jedną grupę, natomiast trzeci dziedziczył się od nich niezależnie. Liczbę sprzężonych genów można łatwo określić porównując liczbę klas fenotypowych z oczekiwaną liczbą klas w potomstwie krzyżówki testowej. Jeżeli 3 geny lub grupy genów sprzężonych segregują niezależnie to powinno powstać 23 czyli 8 klas fenotypowych. Przykładowo, jeżeli krzyżujemy heterozygoty względem 8 genów, a w potomstwie krzyżówki testowej otrzymujemy 8 klas to geny są sprzężone w 3 grupach. Analogicznie ‐ 4 klasy fenotypowe oznaczają, że segregują 2 geny lub 2 grupy genów sprzężonych, 2 klasy fenotypowe – segreguje 1 gen lub 1 grupa wielu sprzężonych genów. Jeżeli krzyżujemy testowo heterozygotę pod względem 10 genów, a w potomstwie otrzymujemy tylko dwie klasy fenotypowe to znaczy, że wszystkie geny były ze sobą sprzężone – tworzyły jeden blok.
W ostatnim etapie należy ustalić, które geny były sprzężone. Dla uproszczenia dalszych rozważań zapiszmy klasy fenotypowe w potomstwie krzyżówki testowej za pomocą gamet, z których one powstały, pamiętając, że jedną z gamet zawsze była gameta abd pochodząca od homozygoty recesywnej. Gametę pochodzącą od heterozygoty F1 łatwo ustalimy, zapisując symbolami alleli ujawnione cechy fenotypowe. Przykładowo roślina o owocach gładkich, łodygach antocjanowych i liściach mozaikowych powstała z połączenia gamety F1 niosącej allel A warunkujący gładkie owoce, allel B warunkujący antocjanowe łodygi i allel d warunkujący mozaikowe liście (możemy tę gametę zapisać ABd) z gametą homozygoty recesywnej abd. Genotypy otrzymanych klas zapisane za pomocą gamet są następujące:
45 roślin o owocach gładkich, łodygach antocjanowych, liściach mozaikowych ABd x abd 52 rośliny o owocach gładkich, łodygach zielonych, liściach mozaikowych Abd x abd; 49 roślin o owocach owłosionych, łodygach zielonych, liściach zielonych abD x abd; 44 rośliny o owocach owłosionych, łodygach antocjanowych, liściach zielonych aBD x abd;
Sprawdźmy kolejno, które geny są sprzężone. Najłatwiej to zrobić analizując dwa geny jednocześnie i przyrównując typy i częstości gamet wytworzonych przez pokolenie F1 z częstościami oczekiwanymi w przypadku niezależnego dziedziczenia dwóch genów.
Sprzężenie między A i B – jeżeli geny A i B dziedziczą się niezależnie to heterozygota AaBb wytwarza 4 typy gamet, AB, Ab, aB, ab w proporcji 1:1:1:1. Korzystając z naszego zapisu genotypów za pomocą gamet widzimy gamety wytwarzane przez heterozygotę F1, a ich proporcja jest identyczna jak proporcja fenotypów w potomstwie krzyżówki testowej. Dla genu A i B otrzymano wszystkie 4 gamety w proporcjach 45:52:49:44, a więc zbliżonych do oczekiwanej proporcji 1:1:1:1. Oznacza to, że gen A warunkujący typ owocu oraz gen B odpowiedzialny za zabarwienie łodygi dziedziczą się niezależnie.
Sprzężenie między A i D – jeżeli geny A i D dziedziczą się niezależnie heterozygota AaDd wytwarza 4 typy gamet, AD, Ad, aD, ad w proporcji 1:1:1:1. Korzystając z zapisu fenotypów za pomocą gamet, widzimy, że F1 pomidora wytworzyło 0 gamet AD (0 roślin AD x ad w potomstwie krzyżówki testowej), 45+52 Ad, 49+44 aD oraz 0 gamet ad, a więc proporcje wynosiły 0:1:1:0. Wystąpiło odstępstwo od przewidywanej częstości gamet, co uwidoczniło się jako odstępstwo od przewidywanych rozszczepień fenotypowych 1:1:1:1 w potomstwie krzyżówki testowej. Geny A i D nie dziedziczą się niezależnie, lecz są ze sobą sprzężone całkowicie.
Sprzężenie między B i D – wykonując podobne obliczenia możemy ustalić sprzężenie pomiędzy genami B i D. Jednakże w omawianym przypadku z poprzednich ustaleń wynika również niezależne dziedziczenie B i D. Skoro gen B dziedziczył się niezależnie od A, a gen D jest z A sprzężony to również gen B musi dziedziczyć się niezależnie od genu D. Dla 3 genów nie ma innej możliwości. Jednakże dla większej liczby genów zawsze lepiej posłużyć się obliczeniami, aby uniknąć przypadkowych błędów.

9. GENY SPRZĘŻONE
©Kornelia Polok 39
Brak części oczekiwanych klas fenotypowych w potomstwie krzyżówki testowej świadczy o sprzężeniu całkowitym genów. Geny takie zawsze dziedziczą się wspólnie i nie występują formy zrekombinowane w stosunku do form rodzicielskich. Sprzężenie całkowite wykorzystywane jest w praktyce w selekcji wspomaganej markerami. Poszukuje się markerów DNA, które faktycznie są fragmentem genu kodującego jakąś cechę, np. odporność na choroby, a następnie w trakcie hodowli wykorzystuje się taki marker do identyfikacji roślin odpornych. Procedura ta ułatwia selekcję, gdyż można dokonać jej na etapie siewek i jest bardziej jednoznaczna. Symptomy choroby jest trudno określić i często zależą od warunków środowiskowych. Sprzężenie całkowite w przypadku genów kodujących cechy morfologiczne na ogół jest efektem położenia genów na chromosomie na tyle blisko, że w przeciętnej populacji segregującej, liczącej 100‐200 osobników nie obserwuje się rekombinantów. W ostatnim etapie należy ustalić genotypy rodziców z uwzględnieniem położenia na chromosomach, a także fenotypy rodziców.
Genotyp rodziców dla genów sprzężonych można łatwo ustalić wiedząc, jakie gamety wytwarzało pokolenie F1. W naszym przypadku sprzężone były geny A i D, a F1 wytworzyło tylko gamety Ad i aD. Oznacza to, że w takim układzie musiały występować oba geny u rodziców, czyli jeden z rodziców wytwarzał gamety Ad, a drugi aD. Gamety Ad i aD to gamety rodzicielskie. Gamety AD i ad, które nie powstały w F1, byłyby gametami zrekombinowanymi. Genotypy rodziców pod względem genów A i D były Ad/Ad oraz aD/aD, gdzie kreska oddziela allele zlokalizowane na dwóch chromosomach homologicznych. Genotyp F1 odpowiednio należy zapisać Ad/aD – kreska oddziela allele dziedziczone od jednego rodzica od alleli odziedziczonych od drugiego rodzica. W przypadku całkowitego sprzężenia genów heterozygota wytwarza tylko gamety rodzicielsie – tzn. takie, które wytwarzali jej homozygotyczni rodzice. Jeżeli rodzice wytwarzali gamety AB i ab, to tylko takie wytworzy heterozygota. Nie wystąpią gamety zrekombinowane Ab i aB. Jeżeli rodzice wytwarzali gamety Ab i aB to takie wytworzy heterozygota, a gamety zrekombinowane AB i ab nie pojawią sie. Dzięki temu można odtworzyć w jakim układzie geny były sprzężone a także genotypy i fenotypy rodziców, jeżeli nie są znane.
W odniesieniu do genu B, który dziedziczył się niezależnie od A i D, F1 było heterozygotyczne, Bb i tym samym rodzice musieli mieć genotypy BB i bb.
Podsumowując rozważania w punkcie 7 możemy zapisać łączne genotypy rodziców i ich fenotypy.
krzyżowano rośliny o owocach gładkich, łodygach antocjanowych i liściach mozaikowych i z roślinami o owocach owłosionych, łodygach i liściach zielonych (ten przypadek pokazano na rysunku, str. XX),
lub
krzyżowano rośliny o owocach gładkich, łodygach zielonych i liściach mozaikowych z roślinami o owocach owłosionych, łodygach antocjanowych i liściach zielonych.
Nie znając fenotypów rodziców nie jesteśmy w stanie określić czy allel dominujący B występował z allelami Ad czy aD, dlatego nie możemy jednoznacznie określić fenotypów i genotypów, a jedynie podać wszystkie możliwości.
Genotyp F1 z uwzględnieniem położenia na chromosomach:
bbaDaD
X BBAdAd
BBaDaD
X bbAdAd
BbaDAd
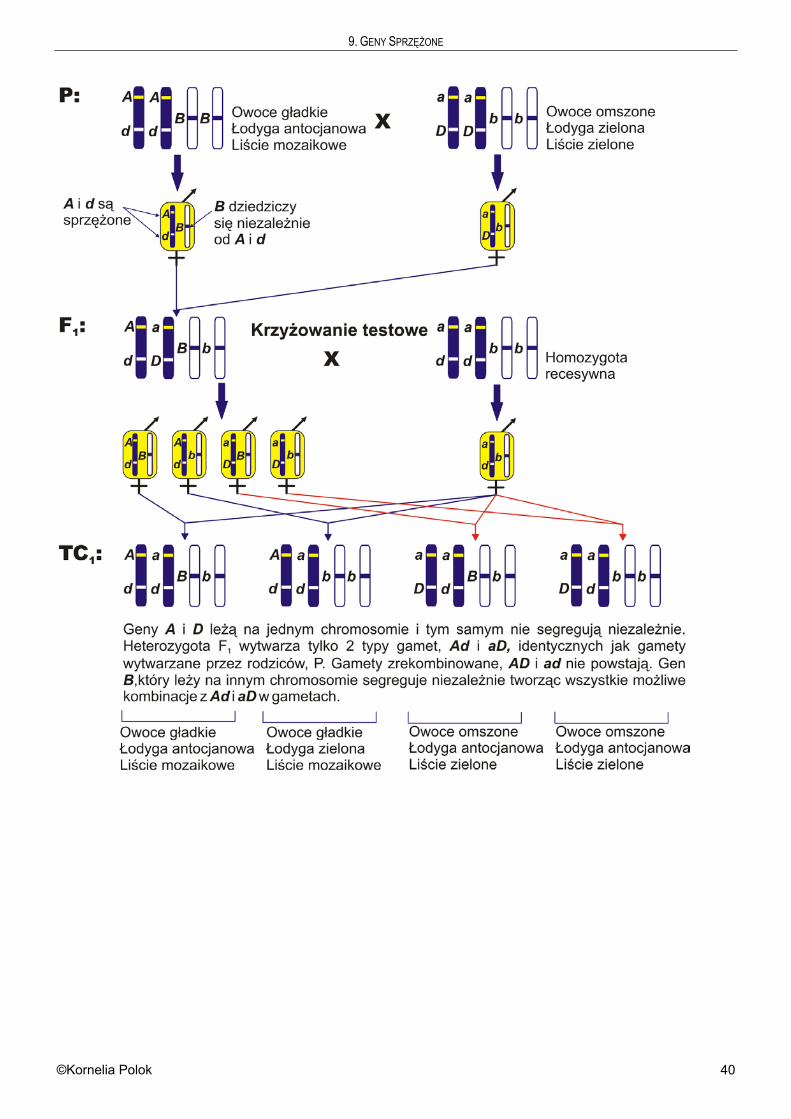
9. GENY SPRZĘŻONE
©Kornelia Polok 40

9. GENY SPRZĘŻONE
©Kornelia Polok 41
Przykład 2. Sprzężenie częściowe, krzyżówka dwupunktowa
Po skrzyżowaniu dwóch roślin jęczmienia otrzymano rośliny o kłosie luźnym i normalnie wykształconej nasadce kłosa. W krzyżówce testowej pokolenia F1 otrzymano:
221 roślin o kłosie luźnym i osadce kłosowej z kryzą, 229 roślin o kłosie zbitym i normalnej osadce kłosowej; 24 rośliny o kłosie luźnym i osadce kłosowej normalnej, 26 roślin o kłosie zbitym i osadce kłosowej z kryzą.
a) Ustal sposób dziedziczenia się obu cech i podaj genotypy rodziców oraz pokolenia F1. b) Podaj prawdopodobieństwo wystąpienia w pokoleniu F2 roślin o kłosie luźnym i osadce kłosowej z kryzą.
ROZWIĄZANIE – PUNKT A
W wyniku skrzyżowania, dwóch roślin jęczmienia otrzymano jednolite potomstwo (wszystkie rośliny były identyczne), co oznacza, że krzyżowane rośliny były homozygotami, a potomstwo uzyskane z ich skrzyżowania – heterozygotami. Było to pokolenie F1.
W krzyżówce segregują dwie cechy – kształt kłosa i kształt osadki kłosowej, Załóżmy, że cechy te uwarunkowane są odpowiednio genami X, i B. Możemy zapisać „roboczy” genotyp pokolenia F1 jako XxBb. Na podstawie wyglądu pokolenia F1, możemy stwierdzić, że allel dominujący X warunkuje wystąpienie luźnego kłosa, a jego allel recesywny, x – kłosa zbitego. Allel B jest odpowiedzialny za normalnie wykształconą osadkę kłosową, a jego allel recesywny, b za obecność kryzy. Nie ma jednolitej zasady nadawania nazw genom i symbolom alleli. W przeszłości nazwy genów i ich symbole na ogół nadawano na podstawie fenotypów dominujących. U człowieka nazwy genów często pochodzą od wywoływanych chorób. Wraz z rozwojem technik mapowania i indukowania mutacji, symbole genów zaczęto wywodzić od fenotypów zmutowanych, np. gen żółtej barwy ciała u D. melanogaster oznacza się symbolem y ( od „yellow”). Projekty sekwencjonowania genomów przyczyniły się do gwałtownego wzrostu liczby poznanych genów, które często oznaczane są kombinacją liter i cyfr wymaganą przez banki genów.
Pokolenie F1 skrzyżowano testowo, a więc było to krzyżowanie między heterozygotą XxBb, a homozygotą recesywną xxbb. Zgodnie z Drugim Prawem Mendla w potomstwie tej krzyżówki testowej powinny wystąpić 4 klasy fenotypowe w proporcjach 1:1:1:1.
W otrzymanym potomstwie pojawiły się przewidywane 4 klasy fenotypowe. Oznacza to, że geny X i B nie są całkowicie sprzężone.
Czy obecność 4 klas fenotypowych świadczy o niezależnym dziedziczeniu genów X i B? Aby to sprawdzić należy porównać proporcje fenotypów w potomstwie krzyżówki testowej z oczekiwaną proporcją 1:1:1:1, która występuje w przypadku dziedziczenia niezależnego. Okazuje się, że proporcja oczekiwana nie wystąpiła w naszej krzyżówce testowej. Klasy fenotypowe wystąpiły w przybliżeniu w proporcjach 10:10:1:1. Odstępstwo od proporcji fenotypów 1:1:1:1 jest dowodem, że geny X i B nie dziedziczą się niezależnie. Geny te są częściowo sprzężone. Występowanie w potomstwie krzyżówki testowej wszystkich oczekiwanych klas fenotypowych, ale z różną częstością oznacza częściowe sprzężenie genów. Przykładowo, dla dwóch genów dziedziczących się niezależnie klasy fenotypowe powinny wystąpić w proporcjach 1:1:1:1, a więc po 25% dla każdej z klas – jeżeli proporcje są inne (np. 4:4:1:1) to geny są sprzężone. Jeżeli dwa geny są sprzężone, a trzeci dziedziczy się niezależnie, to w potomstwie krzyżówki testowej pojawi się 8 klas, przy czym 4 będą znacznie bardziej liczne (np. 4:4:4:4:1:1:1:1).

9. GENY SPRZĘŻONE
©Kornelia Polok 42
Wiedząc, że geny X i B są sprzężone, następnym krokiem jest ustalenie, w jakim układzie są one sprzężone – czy w fazie skupionej (występowanie u heterozygoty alleli dominujących na jednym chromosomie, a alleli recesywnych na drugim), czy w fazie rozproszonej (sprzężenie między allelem dominującym i allelem recesywnym). Podobnie jak w przypadku sprzężeń całkowitych, zapiszmy klasy fenotypowe w potomstwie krzyżówki testowej za pomocą gamet, z których one powstały, pamiętając, że jedna z gamet to xb wytwarzana przez homozygotę recesywną.
221 roślin o kłosie luźnym i osadce kłosowej z kryzą Xb x xb; 229 roślin o kłosie zbitym i normalnej osadce kłosowej xB x xb; 24 rośliny o kłosie luźnym i osadce kłosowej normalnej XB x xb; 26 roślin o kłosie zbitym i osadce kłosowej z kryzą xb x xb.
Jeżeli geny dziedziczą się niezależnie to heterozygota F1 wytwarza 4 typy gamet, każdą z taką samą częstością, co znajduje odzwierciedlenie w równej częstości poszczególnych klas fenotypowych w potomstwie krzyżówki testowej. W naszej krzyżówce testowej 4 klasy fenotypowe wystąpiły z różną częstością, dwie klasy były znacznie liczniejsze niż pozostałe. Oznacza to, że heterozygota wytwarzała 4 typy gamet, ale w przeciwieństwie do niezależnego dziedziczenia genów, 2 typy gamet powstawały znacznie częściej. W naszym przypadku najliczniejsze były klasy o liczebnościach 221 i 229. Z fenotypów odczytujemy, że klasy te powstały z gamet Xb oraz xB, które musiały być najczęściej wytwarzane przez heterozygotę. Gamety te to tzw. gamety rodzicielskie. Widzimy, że geny X i B były sprzężone w układzie rozproszonym. Możemy ustalić genotypy rodziców i F1 oraz fenotypy rodziców.
P1 kłos luźny, osadka kłosowa z kryzą
P2 kłos zbity, osadka kłosowa normalna (bez kryzy)
F1 kłos luźny, osadka kłosowa normalna (fenotyp znamy z treści zadania)
W przypadku genów sprzężonych częściowo, heterozygota wytwarza najliczniej gamety, które odpowiadają gametom jej homozygotycznych rodziców – są to tzw. gamety rodzicielskie. Jeżeli genotypy rodziców nie są znane, to genotyp gamet rodzicielskich można ustalić na podstawie potomstwa krzyżówki testowej – klasy fenotypowe występujące najczęściej powstają z gamet rodzicielskich. Znając gamety rodzicielskie można ustalić genotypy rodziców, heterozygoty F1 oraz można określić układ genów na chromosomach heterozygoty (faza skupiona czy rozproszona).
Wiemy już, że geny X i B są sprzężone, czyli leżą na jednym chromosomie. Znamy także genotypy rodziców oraz F1 z uwzględnieniem położenia tych genów na chromosomie. Kolejnym etapem jest odpowiedź na pytanie jak silnie geny są sprzężone. Intensywność sprzężenia można określić na podstawie częstości rekombinacji pomiędzy sprzężonymi genami. Im silniej geny są sprzężone tym mniejsza częstość rekombinacji i przy sprzężeniu całkowitym wynosi ona zero. W takim przypadku heterozygota nie wytwarza gamet ze zrekombinowanym układem genów w stosunku do form rodzicielskich, co widzieliśmy w przykładzie 1. Przy sprzężeniu częściowym gamety zrekombinowane są wytwarzane, ale powstają znacznie rzadziej niż gamety zawierające rodzicielski układ genów. Częstość, z jaką heterozygota wytwarza gamety zrekombinowane (z układem genów zmienionym w stosunku do gamet rodzicielskich) jest równa częstości rekombinacji między genami sprzężonymi i stanowi miernik intensywności sprzężenia. Częstość rekombinacji wyrażona w procentach nosi nazwę odległości genetycznej. Wartość 1 jednostki mapowej lub 1 centyMorgana (cM) odpowiada 1% rekombinacji. W 1931 roku H. Creighton i B. McClintock po raz pierwszy udowodnili, że rekombinacja pomiędzy genami sprzężonymi jest związana z wymianą materiału genetycznego pomiędzy chromosomami homologicznymi. Proces ten znany jest jako crossing-over i zachodzi w pachytenie mejozy. Dlatego możemy powiedzieć, że odległość genetyczna pomiędzy dwoma genami jest równa częstości crossing-over.
XbXb
xBxB
xBXb

9. GENY SPRZĘŻONE
©Kornelia Polok 43
Aby określić intensywność sprzężenia pomiędzy genami Xb oraz xB należy obliczyć odległość genetyczną. W tym celu obliczamy częstość występowania gamet zrekombinowanych, która jest równa częstości klas fenotypowych w potomstwie krzyżówki testowej, które z tych gamet powstały. Dzięki zapisowi genotypów za pomocą gamet, widzimy, że w naszym przypadku gamety zrekombinowane to XB i xb, które dały klasy fenotypowe o liczebnościach 24 i 26. Częstość wystąpienia gamet zrekombinowanych obliczamy jako iloraz sumy osobników powstałych z gamet zrekombinowanych i wszystkich otrzymanych osobników. Dla genów Xb i xB wartość ta wynosi 24+26(221+229+24+26), czyli 50/500 czyli 0,1.
Dla jakichkolwiek dwóch genów sprzężonych częstość rekombinacji nie przekracza wartości 0,5 (50%). Z częstością 0,5 powstają gamety zrekombinowane, gdy geny leżą na różnych chromosomach – czyli dziedziczą się niezależnie. Zdarza się także, że geny zlokalizowane na przeciwległych końcach długich chromosomów, a czasami nawet ich ramion rekombinują z częstością ≥50%, czyli spełniają warunek niezależnego dziedziczenia, tak jakby były na różnych chromosomach. Dlatego dla genów zlokalizowanych na jednym chromosomie używa się pojęcia grupa sprzężeń. Geny leżące w dużej odległości na jednym chromosomie, pomimo iż z matematycznego punktu widzenia mogą być niesprzężone, nadal należą do jednej grupy sprzężeń.

9. GENY SPRZĘŻONE
©Kornelia Polok 44
Częstość gamet zrekombinowanych jest równa częstości rekombinacji między genami Xb i xB. Częstość rekombinacji jest więc równa 0,1. Odległość genetyczna to częstość rekombinacji wyrażona w procentach. W takim razie 0,1 x 100% = 10%. Odległość pomiędzy tymi genami wynosi 10 jednostek mapowych, czyli 10 cM. W genetyce wyróżnia się odległość fizyczną i genetyczną. Odległość fizyczna jest mierzona w parach zasad i ma znaczenie w sekwencjonowaniu genów. Odległość fizyczna nie zawsze odpowiada intensywności sprzężenia. Odległość genetyczna określa częstość rekombinacji i ma znaczenie w mapowaniu, a także w hodowli, gdzie istotna jest wiedza, z jaką częstością pojawią się rekombinanty o pożądanych cechach. Ponieważ crossing-over zachodzi z różną częstością wzdłuż chromosomów, określonej odległości genetycznej, np. 1 cM odpowiada różna odległość fizyczna. Przy dużej częstości crossing-over geny oddalone o zaledwie kilkaset par zasad mogą wykazywać znaczną odległość genetyczną. Jeżeli częstość crossing-over jest mała to geny mogą być oddalone o tysiące par zasad, a mimo to sprzężone całkowicie. Ponieważ obie miary odległości nie są tożsame i mają różne zastosowania, są one na równi podstawą zaawansowanych badań z zakresu genomiki.
ROZWIĄZANIE – PUNKT B
Mamy obliczyć prawdopodobieństwo wystąpienia osobników o kłosie luźnym i osadce kłosowej z kryzą w pokoleniu F2. Pokolenie F2 powstaje w wyniku krzyżowania dwóch heterozygot, w naszym przypadku XxBb. W przypadku niezależnego dziedziczenia genów, heterozygota XxBb wytwarzałaby 4 typy gamet z równą częstością i znając proporcje fenotypów w pokoleniu F2 wynikające z Drugiego Prawa Mendla (9:3:3:1), można łatwo obliczyć częstość wystąpienia interesującego nas fenotypu. W naszym przypadku fenotyp o kłosie luźnym (cecha dominująca) oraz osadce kłosowej z kryzą (cecha recesywna) powstałby z częstością 3/16 (częstość osobników o genotypie X bb). Jednakże geny X i B nie dziedziczą się niezależnie, są sprzężone i w związku z tym heterozygoty XxBb wytwarzają 4 typy gamet z różną częstością, co uniemożliwia nam zastosowanie obliczeń wynikających z Drugiego Prawa Mendla.
Ponieważ geny X i B są sprzężone, prawdopodobieństwo wystąpienia w F2 osobników o kłosie luźnym i osadce kłosowej z kryzą musimy obliczyć na podstawie częstości gamet, z których ten fenotyp powstaje. W przypadku sprzężenia genów, heterozygota częściej wytwarza gamety rodzicielskie niż zrekombinowane. Dlatego, w pierwszej kolejności należy ustalić, które gamety są zrekombinowane, a które rodzicielskie, a to ustalamy na podstawie fazy, w jakiej geny były sprzężone. W punkcie a) ustaliliśmy, że geny X i B były sprzężone w fazie rozproszonej i heterozygota miała genotyp Xb/xB. Stąd wiemy, że:
gamety zrekombinowane to XB, xb; gamety rodzicielskie to Xb i xB.
Następnie należy obliczyć częstość występowania gamet rodzicielskich i zrekombinowanych wykorzystując odległość genetyczną, która dla X i B wynosi 10 cM.
Częstość gamet zrekombinowanych XB i xb - odległość pomiędzy Xb i xB wynosząca 10 cM to częstość rekombinacji między X i B wyrażona w procentach – 10%, a więc częstość rekombinacji między X i B wynosi 0,1. Wartość ta odpowiada sumarycznej częstości wystąpienia gamet zrekombinowanych. Ponieważ mamy dwa typy gamet zrekombinowanych, XB i xb, to częstość każdego z nich wynosi 0,1/2 = 0,05.
Częstość gamet rodzicielskich Xb i xB – wszystkie gamety stanowią 100%, gamety zrekombinowane stanowią 10%, a więc gamety rodzicielskie to 100% - 10%, czyli 90%, co odpowiada częstości gamet rodzicielskich 0,9. Częstość gamet rodzicielskich możemy też obliczyć jako 1 (częstość wszystkich gamet) minus częstość gamet zrekombinowanych, czyli 1 – 0,1 = 0,9. Ponieważ mamy 2 typy gamet rodzicielskich, to częstość każdego z nich wynosi 0,45.

9. GENY SPRZĘŻONE
©Kornelia Polok 45
Częstość gamet zrekombinowanych wynikająca z odległości genetycznej jest częstością sumaryczną dla wszystkich gamet zrekombinowanych (np. 0,2). Jeżeli mamy 2 typy gamet zrekombinowanych (np. Ab i aB) to częstość każdego z nich jest równa połowie częstości sumarycznej (np. 0,2/2 = 0,1). Jeżeli są 4 typy gamet zrekombinowanych (np. AbD, Abd, aBD, aBd) to częstość każdego z nich równa się jednej czwartej częstości sumarycznej (np. 0,2/4 = 0,05). Sumaryczna częstość gamet rodzicielskich jest różnicą pomiędzy wartością 1 a sumaryczną częstością gamet zrekombinowanych (np. 1 – 0,2 = 0,8). Podobnie jak w przypadku gamet zrekombinowanych, częstość danego typu gamety rodzicielskiej (np. AB) jest ilorazem częstości sumarycznej i liczby typów gamet rodzicielskich (np. dla dwóch typów – 0,8/2 = 0,4; dla 4 typów 0,8/4 = 0,2).
W ostatnim etapie należy ustalić, z jakich gamet powstaną osobniki o luźnych kłosach (cecha dominująca) i osadce kłosowej z kryzą (cecha recesywna). Zapiszmy interesujący nas fenotyp za pomocą genotypu jako X bb. Z zapisu widzimy, że są 3 możliwości kombinacji gamet, które prowadzą do interesującego nas fenotypu: Xb x Xb; Xb x xb; xb x Xb. Obliczmy prawdopodobieństwo każdej kombinacji gamet – z rachunku prawdopodobieństwa wynika, że prawdopodobieństwo zdarzenia A x B jest iloczynem prawdopodobieństwa A i prawdopodobieństwa B. Częstość wyraża prawdopodobieństwo, czyli częstość pojawienia się kombinacji gamet np. Xb x xb jest iloczynem częstości występowania gamety Xb oraz gamety xb.
Częstość Xb x Xb – częstość gamet Xb wynosi 0,45, czyli 0,45 x 0,45 = 0,2025. Częstość Xb x xb – częstość gamet Xb wynosi 0,45, częstość gamet xb, 0,05, czyli 0,45 x 0,05 = 0,0225. Częstość xb x Xb – 0,05 x 0,45 = 0,0225.

9. GENY SPRZĘŻONE
©Kornelia Polok 46
Podsumowując, istnieją 3 możliwości powstania w pokoleniu F2 fenotypu o kłosie luźnym i osadce kłosowej z kryzą. Prawdopodobieństwo pojawienia się takiego fenotypu jest sumą częstości, z jaką wystąpi każda z 3 możliwości, a więc 0,2025 + 0,0225 + 0,0225 = 0,2025 + 2 x 0,0225 = 0,2025 + 0,05 = 0,2475 Prawdopodobieństwo wystąpienia jakiegokolwiek fenotypu w pokoleniu F2 w przypadku sprzężenia genów oblicza się na podstawie częstości gamet, z których interesujący nas fenotyp powstaje. W tym celu musimy znać fazę sprzężenia (skupiona lub rozproszona) oraz odległość genetyczną pomiędzy genami sprzężonymi (lub wyniki krzyżówki testowej). Na podstawie fazy sprzężenia ustalamy gamety rodzicielskie i zrekombinowane, a na podstawie odległości obliczamy częstość gamet rodzicielskich i zrekombinowanych. Jeżeli fenotyp odpowiada tylko jednemu genotypowi, to obliczamy prawdopodobieństwo wystąpienia danego fenotypu/genotypu jako iloczyn częstości gamet (np. 0,1 x 0,1), z których on powstaje. Gdy genotypów składających się na nasz fenotyp jest więcej (np. 3), prawdopodobieństwo wystąpienia określonego fenotypu jest sumą prawdopodobieństw pojawienia się każdego z genotypów dających interesujący nas fenotyp. (np. 0,01 + 0,04 + 0,04 = 0,09). W bardziej skomplikowanych przypadkach, gdy segreguje wiele genów i na dany fenotyp składa się wiele genotypów najlepiej posłużyć się szachownicą.

9. GENY SPRZĘŻONE
©Kornelia Polok 47
Zadania
1. Pokolenie F1 otrzymane po skrzyżowaniu dwóch roślin jęczmienia miało 2-rzędowy (Vrs) oraz ościsty kłos (Lk), antocjanową łodygę (Ant) i normalnie zabarwione liście (Wst). W krzyżówce testowej rośliny F1 otrzymano:
66 roślin o kłosie 2-rzędowym, bezostnym, zielonej łodydze i normalnie zabarwionych liściach;
64 rośliny o kłosie 2-rzędowym, ościstym, zielonej łodydze i biało paskowanych liściach;
68 rośliny o kłosie 6-rzędowym, bezostnym, antocjanowej łodydze i normalnie zabarwionych liściach;
60 roślin o kłosie 6-rzędowym, ościstym, antocjanowej łodydze i biało paskowanych liściach.
Ustal sposób dziedziczenia omawianych cech. Podaj wszystkie możliwe genotypy i fenotypy krzyżowanych roślin jęczmienia oraz określ prawdopodobieństwo pojawienia się w pokoleniu F2 roślin o kłosie 2-rzędowym, ościstym, zielonej łodydze i normalnie zabarwionych liściach.
2. U jęczmienia w chromosomie 1 zmapowano gen warunkujący oplewienie ziarniaka (N, n) oraz gen warunkujący kształt kłosa (Ert, ert). Odległość między tymi genami wynosi 12 j.m. W chromosomie 2 zlokalizowano gen odpowiedzialny za zabarwienie plewek (Or, or). W pokoleniu F1 uzyskanym ze skrzyżowania rośliny o nagim ziarniaku, kłosie zbitym, i pomarańczowej plewie z rośliną o oplewionym ziarniaku, kłosie normalnym i zielonej plewie otrzymano wyłącznie osobniki o oplewionym ziarniaku, kłosie normalnym i zielonej plewie. Podaj genotypy krzyżowanych form, pokolenia F1 oraz oblicz prawdopodobieństwo pojawienia się w pokoleniu F2 osobników o oplewionym ziarniaku, kłosie zbitym i pomarańczowej plewie.
3. U pomidora w chromosomie 7 zmapowano gen warunkujący barwę owocu (R, r) oraz gen warunkujący kształt owocu (Ov, ov). Odległość między tymi genami wynosi 8 j.m. W chromosomie 2 zlokalizowano gen odpowiedzialny za owłosienie łodygi (H, h). W pokoleniu F1 uzyskanym ze skrzyżowania rośliny o owocach czerwonych, okrągłych i łodygach nagich z rośliną o owocach żółtych, wydłużonych i łodygach owłosionych otrzymano wyłącznie osobniki o owocach czerwonych, okrągłych i łodygach owłosionych. Podaj genotypy krzyżowanych form, pokolenia F1 oraz oblicz prawdopodobieństwo pojawienia się w pokoleniu F2 osobników o owocach żółtych, okrągłych i łodygach owłosionych.
4. Pokolenie F1 otrzymane ze skrzyżowaniu dwóch much D. melanogaster było całkowicie dzikie. W krzyżówce testowej samicy F1 otrzymano:
85 much o oczach vermilion, 89 much o skrzydłach cut, 12 much będących podwójnym mutantem, 14 much całkowicie dzikich.
Podaj genotypy rodziców oraz F1 oraz ustal odległość między genem barwy oczu i kształtu skrzydeł.
5. W pewnej rodzinie, gdzie matka była miała grupę krwi Rh- a ojciec był daltonistą i miał grupę krwi Rh+ urodziła się zdrowa dziewczynka o grupie krwi Rh-. Babka ze strony matki była chora na łuszczycę, ale prawidłowo rozróżniała barwy, natomiast dziadek był daltonistą. Oblicz prawdopodobieństwo urodzenia przez tę dziewczynkę zdrowego chłopca o grupie krwi Rh+ w małżeństwie ze całkowicie zdrowym mężczyzną, którego oboje rodzice oraz dziadkowie mieli grupę Rh+, wiedząc, że gen łuszczycy (p) i daltonizmu są zlokalizowane na chromosomie X w odległości 18 j.m. i są recesywne w stosunku do alleli warunkujących normalny fenotyp; natomiast gen warunkujący czynnik Rh na chromosomie 20. Podaj genotypy matki i ojca.

©Kornelia Polok 48
MMMAAAPPPOOOWWWAAANNNIIIEEE GGGEEENNNÓÓÓWWW
MMMaaapppooowwwaaannniiieee gggeeennnóóówww pppooollleeegggaaa nnnaaa uuussstttaaalllaaannniiiuuu wwwzzzaaajjjeeemmmnnneeegggooo pppooołłłooożżżeeennniiiaaa gggeeennnóóówww nnnaaa ccchhhrrrooommmooosssooommmiiieee ooorrraaazzz ooodddllleeegggłłłooośśśccciii pppooommmiiięęędddzzzyyy gggeeennnaaammmiii...
Przykład 1. Krzyżówka trójpunktowa
U rzodkiewnika, Arabidopsis thaliana gen PY, py odpowiada za zdolność do syntezy tiaminy, gen ER, er za osadzenie łuszczynki, a gen AS, as odpowiada za kształt liści. Wszystkie rośliny F1 otrzymane ze skrzyżowania dwóch linii A. thaliana o przeciwstawnych cechach były zdolne do syntezy tiaminy, miały normalnie zwisającą łuszczynkę i owalne liście. W krzyżówce testowej pokolenia F1 otrzymano:
898 roślin syntetyzujących tiaminę, o erektoidalnych łuszczynkach i owalnych liściach; 36 roślin syntetyzujących tiaminę, o zwisających łuszczynkach i owalnych liściach; 44 rośliny syntetyzujące tiaminę, o erektoidalnych łuszczynkach i asymetrycznych liściach; 2 rośliny syntetyzujące tiaminę, o zwisających łuszczynkach i asymetrycznych liściach; 926 roślin niezdolnych do syntezy tiaminy, o zwisających łuszczynkach i asymetrycznych liściach; 52 roślin niezdolnych do syntezy tiaminy, o zwisających łuszczynkach i owalnych liściach; 40 roślin niezdolnych do syntezy tiaminy, o erektoidalnych łuszczynkach i asymetrycznych liściach; 2 rośliny niezdolne do syntezy tiaminy, o erektoidalnych łuszczynkach i owalnych liściach.
Podaj wszystkie możliwe genotypy i fenotypy rodziców oraz pokolenia F1. Określ, które geny są sprzężone i podaj odległość między nimi.
Z treści zadania wiemy, że krzyżowano linie czyste – homozygotyczne (otrzymano jednolite pokolenie F1). Ponadto wiadomo, że segregują trzy cechy (trzy cechy są wymienione w odniesieniu do F1 oraz trzy cechy segregują w krzyżówce testowej), kodowane przez trzy geny (geny wymieniono w treści zadania). Roboczy genotyp pokolenia F1 możemy zapisać jako PYpyERerASas. Na podstawie fenotypu pokolenia F1 możemy ustalić, które allele są dominujące, a które recesywne.
Allel dominujący, PY odpowiada za zdolność do syntezy tiaminy, a jego allel recesywny, py warunkuje brak tej zdolności.
Allel dominujący, ER warunkuje normalnie zwisającą łuszczynkę, a jego allel recesywny, er łuszczynkę erektoidalną.
Allel dominujący, AS odpowiada za owalne liście a allel recesywny, as za liście asymetryczne.
Pokolenie F1 krzyżowano testowo i otrzymano 8 klas fenotypowych, jednakże liczebności poszczególnych klas były różne. Wystąpiło odstępstwo od oczekiwanego stosunku rozszczepień 1:1:1:1:1:1:1:1, co świadczy o sprzężeniu genów.
W pierwszej kolejności należy ustalić, które geny są sprzężone. Możemy to zrobić sprawdzając każde dwa geny oddzielnie, tak jak to robiliśmy w przypadku sprzężeń całkowitych oraz krzyżówki dwupunktowej. Jednakże można to też stwierdzić analizując liczebności klas fenotypowych. W przypadku częściowego sprzężenia dwóch genów obserwowaliśmy dwie klasy liczebności – liczne klasy fenotypowe, które miały takie same fenotypy jak pokolenie rodzicielskie oraz klasy nieliczne, które miały fenotypy zrekombinowane. W obecnym przykładzie występują 2 klasy bardzo liczne, 2 klasy nieliczne i 4 klasy o pośrednich liczebnościach. Taki układ świadczy, że sprzężone są 3 segregujące geny.
.

10. MAPOWANIE GENÓW
©Kornelia Polok 49
Krzyżowanie testowe heterozygoty pod względem trzech genów nosi nazwę krzyżówki trójpunktowej, ponieważ pozwala ona określić położenie 3 punktów (genów) na chromosomie oraz ustalić odległość pomiędzy nimi. Krzyżówka trójpunktowa jest najprostszym sposobem mapowania genów. Pozwala ona na zmapowanie pojedynczych genów. W praktyce, do tworzenia wysoce wysyconych map genetycznych wykorzystuje się populacje, w których segreguje wiele genów warunkujących cechy morfologiczne, różne izoenzymy, a także segregują tzw. markery DNA – sekwencje DNA wykrywane przy pomocy hybrydyzacji (RFLP) oraz techniki PCR (markery RAPD, AFLP, SSAP, SSR i inne). Obecnie coraz częściej wykorzystuje się sekwencje podlegające ekspresji (EST). Przeciętnie na mapę nanosi się 100‐200 markerów, ale wysoce wysycone mapy mogą zawierać nawet 500 markerów, jak u L. multiflorum x L. perenne (Polok 2007), a u gatunków modelowych 1000 i więcej, jak u jęczmienia. Takie mapy są podstawą genomiki.
Wiedząc, że 3 geny są sprzężone należy ustalić, w jakim układzie były one sprzężone. Pozwoli nam to zapisać genotypy rodziców i F1 z uwzględnieniem układu na chromosomach. Dodatkową komplikacją są 3 sprzężone geny, z których jeden musi być w środku. W pierwszej kolejności należy ustalić gen środkowy.
a) Istnieją 3 możliwości wzajemnego położenia genów, które możemy roboczo zapisać jako:
1. Py-Er-As, 2. Py-As-Er, 3. Er-Py-As. b) Aby ustalić, która z tych możliwości jest prawdziwa należy przeanalizować otrzymane klasy fenotypowe.
Klasy najliczniejsze powstały z gamet rodzicielskich. Możemy zapisać robocze genotypy tych klas za pomocą gamet pamiętając, że jedna z gamet pyeras pochodzi od homozygoty recesywnej:
898 roślin syntetyzujących tiaminę, o erektoidalnych łuszczynkach i owalnych liściach - PYerAS x pyeras; 926 roślin niezdolnych do syntezy tiaminy, o zwisających łuszczynkach i asymetrycznych liściach - pyERas x pyeras;
c) Pozostałe 6 klas fenotypowych powstało z gamet zrekombinowanych, przy czym 4 z nich, o liczebności
pośredniej, musiały pochodzić z gamet po pojedynczym crossing-over, który zaszedł w jednym z dwóch regionów pomiędzy 3 genami, np. pomiędzy genem 1 i 2 lub pomiędzy genem 2 i 3. Dwie klasy najmniej liczne powstały z gamet po podwójnym crossing-over, a więc takim, który zaszedł jednocześnie w regionie pomiędzy genami 1 i 2 oraz pomiędzy genami 2 i 3. Zapiszmy klasy powstałe z gamet po podwójnym crossing-over jako:
2 rośliny syntetyzujące tiaminę, o zwisających łuszczynkach i asymetrycznych liściach - PYERas x pyeras; 2 rośliny niezdolne do syntezy tiaminy, o erektoidalnych łuszczynkach i owalnych liściach - pyerAS x pyeras;
d) Porównajmy gamety rodzicielskie z gametami po podwójnym crossing-over. Gen, który zmienił się w
gametach po podwójnym crossing-over w stosunku do gamet rodzicielskich jest genem środkowym.
Gamety rodzicielskie
Gamety po podwójnym
crossing-over
Allele identyczne w gametach rodzicielskich i gametach po
podwójnym crossing-over
Allele zmienione w gametach po podwójnym crossing-over
PYerAS pyerAS erAS PY
pyERas PYERas ERas py

10. MAPOWANIE GENÓW
©Kornelia Polok 50
Klasy fenotypowe występujące najliczniej w potomstwie krzyżówki trójpunktowej powstały z gamet rodzicielskich. Klasy o najniższych liczebnościach powstały z gamet po podwójnym crossing-over. Porównanie gamet rodzicielskich z gametami po podwójnym crossing-over pozwala ustalić gen środkowy. Jest to ten gen, który zmienia się w gametach po podwójnym crossing-over w stosunku do gamet rodzicieskich.
Znając gen środkowy oraz układ genów w gametach rodzicielskich możemy ustalić genotypy rodziców i F1 z uwzględnieniem położenia na chromosomach.
P1 łuszczynki erektoidalne, zdolność do syntezy tiaminy, liście owalne
P2 łuszczynki zwisające, brak zdolności do syntezy tiaminy, liście asymetryczne
F1 łuszczynki zwisające, zdolność do syntezy tiaminy, liście owalne
Wiemy już, który gen jest środkowy oraz znamy układ genów sprzężonych. W ostatnim etapie obliczamy odległość pomiędzy genami sprzężonymi, czyli odległość pomiędzy genami er i PY (ER–py) oraz pomiędzy genami PY a AS (py–as). W tym celu należy obliczyć częstość rekombinacji (crossing-over) pomiędzy odpowiednimi genami. Częstość rekombinacji odpowiada częstości gamet zrekombinowanych, a ta z kolei odpowiada częstości klas fenotypowych powstałych z tych gamet. Dlatego należy ustalić, które klasy fenotypowe w potomstwie krzyżówki trójpunktowej powstały z gamet po crossing-over w obszarze er-PY (ER-py), a które po crossing-over w obszarze PY-AS (py-as). Są to 4 klasy o częstościach pośrednich oraz 2 klasy po podwójnym crossing-over. Zapiszmy je za pomocą gamet, z których powstały uwzględniając gen PY, py jako środkowy.
ERpyasERpyas
ERpyaserPYAS
erPYASerPYAS

10. MAPOWANIE GENÓW
©Kornelia Polok 51
Po pojedynczym crossing-over 36 roślin syntetyzujących tiaminę, o zwisających łuszczynkach i owalnych liściach - ERPYAS x erpyas; 44 rośliny syntetyzujące tiaminę, o erektoidalnych łuszczynkach i asymetrycznych liściach - erPYas x erpyas; 52 roślin niezdolnych do syntezy tiaminy, o zwisających łuszczynkach i owalnych liściach - ERpyAS x erpyas; 40 roślin niezdolnych do syntezy tiaminy, o erektoidalnych łuszczynkach i asymetrycznych liściach - erpyas x erpyas. Po podwójnym crossing-over 2 rośliny syntetyzujące tiaminę, o zwisających łuszczynkach i asymetrycznych liściach - ERPYas x erpyas; 2 rośliny niezdolne do syntezy tiaminy, o erektoidalnych łuszczynkach i owalnych liściach - erpyAS x erpyas.
Widzimy, że klasy o liczebności 36 i 40 powstały z gamet po pojedynczym crossing-over w obszarze pomiędzy ER-PY, natomiast klasy o liczebności 44 i 52 powstały z gamet po pojedynczym crossing-over w obszarze pomiędzy genami PY-AS. Znając liczebność wszystkich klas możemy obliczyć częstość gamet zrekombinowanych, a tym samym odległość genetyczną.
a) Odległość między ER-PY. Na podstawie gamet rodzicielskich ustaliliśmy, że geny ER i PY są sprzężone w układzie rozproszonym, czyli erPY oraz ERpy. Gamety zrekombinowane to gamety ERPY oraz erpy. Liczebność odpowiadających im klas fenotypowych to 36 i 40. Całkowita liczba otrzymanych roślin wynosi 2000. Częstość gamet zrekombinowanych wynosi 36 +2 + 40 + 2 /2000 = 80/2000 = 0,040. Odległość genetyczna jest wyrażona w procentach czyli wynosi 0,040 x 100% = 4,0 cM.
b) Odległość między PY-AS. Z układu genów w gametach rodzicielskich wiemy, że geny są sprzężone w fazie skupionej czyli PYAS i pyas. Gamety zrekombinowane to PYas oraz pyAS. Odpowiadają one klasom fenotypowym o liczebnościach 44 i 52. Odległość wynosi 44 + 2 + 52 + 2 /2000 = 100/2000 = 0,050. Odległość genetyczna wynosi 5,0 cM.

10. MAPOWANIE GENÓW
©Kornelia Polok 52
Zadania
1. U Lolium na czwartej grupie sprzężeń (LG4) zlokalizowano gen LF, lf, odpowiedzialny za długie, względnie krótkie liście, gen H, h, warunkujący wczesne, względnie późne dojrzewanie oraz gen A, a – kodujący obecność lub brak ości. Krzyżowanie testowe roślin heterozygotycznych pod względem wszystkich cech dało w potomstwie następujące osobniki:
411 roślin dojrzewających późno, bezostnych, o liściach długich; 43 rośliny dojrzewające wcześnie, bezostne, o liściach krótkich; 5 roślin dojrzewających późno, bezostnych, o liściach krótkich; 37 roślin dojrzewających wcześnie, bezostne, o liściach długich; 34 roślin dojrzewających późno, ościste, o liściach krótkich; 418 roślin dojrzewających wcześnie, ościstych, o liściach krótkich; 4 rośliny dojrzewających wcześnie, ościste, o liściach długich; 48 roślin dojrzewających późno, ościstych, o liściach długich.
Podaj genotypy i fenotypy rodziców, pokolenia F1 oraz oblicz odległość pomiędzy genami sprzężonymi.
2. Pokolenie F1 powstałe ze skrzyżowania dwóch much D. melanogaster miało oczy i ciało dzikie oraz szczecinki typu Dichaete (D). W krzyżówce testowej otrzymano następujące klasy osobników:
360 much całkowicie dzikich; 44 muchy o oczach dzikich, szczecinkach Dichaete i ciele stripe (st); 78 much o oczach dzikich, szczecinkach Dichaete i ciele dzikim; 9 much o oczach i szczecinkach dzikich i ciele stripe; 370 much o oczach cardinal (c), szczecinkach Dichaete i ciele stripe (st); 56 much o oczach cardinal, szczecinkach i ciele dzikim; 72 muchy o oczach cardinal,szczecinkach dzikich i ciele stripe; 11 much o oczach cardinal, szczecinkach Dichaete i ciele dzikim.
Podaj genotypy i fenotypy krzyżowanych much, genotyp i fenotyp pokolenia F1 oraz określ, które geny są sprzężone i oblicz odległość miedzy nimi.
3. U królików jednolite zabarwienie sierści dominuje nad ubarwieniem typu himalajskiego, czarna barwa sierści nad brązową, a biały kolor tłuszczu nad żółtym. Potomstwo uzyskane ze skrzyżowania królików należących do dwóch linii było jednolicie czarne i miało biały tłuszcz. W wyniku krzyżówki testowej otrzymano:
405 form himalajskich, czarnych o żółtym tłuszczu, 68 form himalajskich, brązowych o żółtym tłuszczu, 24 formy himalajskie, czarne o białym tłuszczu, 3 formy himalajskie, brązowe o białym tłuszczu, 392 formy jednolicie ubarwione, brązowe o białym tłuszczu, 75 form jednolicie ubarwionych, czarnych o białym tłuszczu, 29 form jednolicie ubarwionych, brązowych o żółtym tłuszczu, 4 formy jednolicie ubarwione, czarne o żółtym tłuszczu,
Podaj genotypy rodziców, F1 z uwzględnieniem połóżenia genów na chromosomach oraz oblicz odległości pomiędzy genami sprężonymi.

JJJaaakkkaaa tttaaammm eeewwwooollluuucccjjjaaa!!! ZZZaaannniiimmm tttrrraaafffiiięęę nnnaaa jjjeeedddnnneeegggooo mmmyyyśśś llląąąccceeegggooo,,, mmmuuussszzzęęę ssstttoooccczzzyyyććć bbbiiitttwwwęęę zzz dddzzziiieeewwwiiięęęccciiiooommmaaa ooorrraaannnggguuutttaaannnaaammmiii...
CCCaaarrrlllooosss RRRuuuiiisss ZZZaaafffooonnn,,, 111999555444
Wszystkie fotografie G.W. Busha pochodzą z portalu Political Humor, http://politicalhumor.about.com (Grudzień 2010)
Seria dowcipów politycznych ukazujących George W. Busha na tle szympansów była szczególnie chętnie rozpowszechniana po wypowiedziach Prezydenta na temat ewolucji w 2005 r. Celem było ośmieszenie, ale patrząc na fotografie wydaje się, że potwierdzają one teorię ewolucji. George W. Bush nigdy nie zaprzeczał teorii ewolucji, ale uważał, że teorie kreacjonistyczne również powinny być przedstawiane w szkołach. Dziennikarze zinterpretowali te poglądy jednoznacznie, że George W. Bush nie uznaje teorii ewolucji. Na dowód chętnie jest przytaczane zdanie Prezydenta wypowiedziane dla Washington Post w 2008 r., że ewolucja nie tłumaczy w pełni cudu życia”. Poglądy i zdania wyrwane z kontekstu doprowadziły do stworzenia wizerunku Prezydenta USA jako człowieka odrzucającego teorię ewolucji. Pomijając aspekt polityczny, cała historia pokazuje, ile emocji nadal wzbudza teoria ewolucji.

©Kornelia Polok 54
OOO EEEWWWOOOLLLUUUCCCJJJIII BBBIIIOOOLLLOOOGGGIIICCCZZZNNNEEEJJJ………
EEEwwwooollluuucccjjjaaa tttooo zzzmmmiiiaaannnaaa nnnaaa ppprrrzzzeeessstttrrrzzzeeennniii ccczzzaaasssuuu... EEEwwwooollluuucccjjjaaa bbbiiiooolllooogggiiiccczzznnnaaa tttooo zzzmmmiiiaaannnaaa ccceeeccchhh dddzzziiieeedddzzziiiccczzznnnyyyccchhh pppooopppuuulllaaacccjjjiii nnnaaa ppprrrzzzeeessstttrrrzzzeeennniii pppoookkkooollleeeńńń... EEEwwwooollluuucccjjjaaa mmmooollleeekkkuuulllaaarrrnnnaaa tttooo eeewwwooollluuucccjjjaaa nnnaaa
pppooozzziiiooommmiiieee kkkwwwaaasssóóówww nnnuuukkkllleeeiiinnnooowwwyyyccchhh iii bbbiiiaaałłłeeekkk...
Wydawałoby się, że teoria ewolucji opracowana przez Karola Darwina wiele lat temu jest ugruntowana. Tymczasem, cytat Carlosa Ruisa Zafona jak i fotografie G.W. Busha unaoczniają jak wiele emocji budzi pojęcie ewolucji oraz jej mechanizmy. I nie chodzi tutaj tylko o niewinne żarty porównujące G.W. Busha do szympansa, czy protesty kreacjonistów w USA przeciwko mówieniu o ewolucji w szkołach. O wiele bardziej niebezpieczna jest postawa wielu naukowców, którzy nie rozumieją i często nawet nie chcą zrozumieć idei ewolucji. Z jednej strony są to niektórzy biolodzy molekularni zafascynowani postępami w analizie DNA, którzy ewolucję odnoszą często jedynie do molekuł. I chociaż mają rację mówiąc o ewolucji genów i białek, i chociaż to im zawdzięczamy wykrycie, że niektóre białka o odmiennych funkcjach mają wspólnego przodka (wazopresyna i oksytocyna), to niestety często zapominają, że ewolucja organizmów odbywa się na poziomie całego fenotypu, a nie pojedynczych cząsteczek. Nie każda zmiana na poziomie DNA przekłada się na zmianę fenotypową. Populacje mogą ewoluować na poziomie molekularnym podczas, gdy ich fenotyp pozostaje niezmienny. Zdarza się tak zwłaszcza, gdy mutacje zachodzą w niekodującym DNA, który stanowi większość genomu. Jednakże to fenotyp reaguje na zmiany środowiska i jako całość podlega Darwinowskiemu doborowi naturalnemu. Obecny postęp technologiczny pozwala na pozyskiwanie ogromnej ilości danych molekularnych. Dzięki temu wiemy coraz więcej o efektach zmian w niekodującym DNA, co więcej zmiany te nie zawsze mają charakter neutralny, a więcmogá wpływa© na fenotyp. Okazuje się, że mutacje wywołane przemieszczeniem transpozonów mogą szczególnie często występować w warunkach stresowych, co zaobserwowano u Hordeum spontaneum w Kanionie Ewolucji w Izraelu. W porównaniu z biologami molekularnymi, znacznie bardziej „niebezpieczni” są Ci naukowcy, którzy utożsamiają ewolucję jedynie z filogenezą (historią rodową), a swoje wnioskowania opierają na pojedynczych genach. Szczególnie dotyczy to taksonomii roślin, w mniejszym stopniu zwierząt. Dzięki działalności Theodosiusa Dobzhansky’ego oraz Ernsta Mayra zoolodzy przyjmują, że ewolucja jest procesem genetycznym i znacznie częściej niż botanicy akceptują biologiczną koncepcję gatunku. Niektórzy taksonomowie, zachłyśnięci metodami sekwencjonowania DNA analizują setki gatunków w celu określenia ich pochodzenia i przedstawienia wzajemnych powiązań w postaci drzewa filogenetycznego. Analizują pojedyncze zmiany zasad, jednakże najczęściej wnioskowanie opierają na pojedynczym przedstawicielu gatunku. Niektóre prace ukazujące się w nawet w renomowanych czasopismach oparte są zaledwie na kilku reakcjach PCR, podczas, gdy kilkadziesiąt innych sekwencji zostało pobranych z banku genów. Wystarczy sięgnąć do baz Springer, Elsevier, HighWire i innych. Nie oznacza to, że takich prac nie należy publikować, bowiem dostarczają one określonych danych. Jednakże do stworzonych drzew i wniosków należy podchodzić z rezerwą pamiętając, że są one oparte na pojedynczych sekwencjach.

11. O EWOLUCJI…
©Kornelia Polok 55
Analiza pojedynczej sekwencji u wielu gatunków, pomimo zastosowania najnowszych technik nie różni się niczym od XIX-wiecznej taksonomii, kiedy to naukowcy niczym muzealnicy spędzali lata na liczeniu włosków lub liści u pojedynczych okazów zielnikowych, a których jedynym celem było nadanie nazwy jakiemuś okazowi. Podejście to doprowadziło do wielu kuriozalnych sytuacji, jak nadawani rangi gatunkowej mutantom jęczmienia otrzymanym w drodze mutagenezy. Dopiero rewizja taksonomiczna w latach 70-tych ubiegłego wieku uporządkowała systematykę tego zboża. Obecnie rolę cech morfologicznych cech zastępują geny, którym próbuje się nadać rangę genu diagnostycznego. Zapomina się przy tym, że jeden gen stanowi niewielki ułamek genomu. Jeden gen o długości 1000 par zasad stanowi zaledwie 0,0086% genomu Arabidopsis thaliana, a jest to roślina o jednym z najmniejszych genomów. Ewolucja może także zachodzić na poziomie sekwencji powtarzalnych, często transpozonowych jak to stwierdzono u Lolium multiflorum i L. perenne (Polok 2007). Do tego analizując niewielkie próby (1-2 osobniki na gatunek) zapomina się o wariantach genu (allelach), których częstość może się zmieniać w populacji na przestrzeni lat, a u gatunków o wielu cyklach rozrodczych w roku, nawet sezonowo, jak to wykazał T. Dobzhansky u muszki owocowej.
Opierając wnioskowanie na
pojedynczych osobnikach, pomija się całkowicie zmienność wewnątrzgatunkową i tym samym nie rozróżnia się procesów zachodzących na poziomie populacji od różnicowania gatunków. Nie oznacza to, że analizy filogenetyczne oparte o pojedyncze sekwencje DNA nie mają zastosowania. Oznacza jedynie tyle, że jest to ewolucja genu/sekwencji u wybranych gatunków, a nie ewolucja gatunków. Drzewa oparte na pojedynczych sekwencjach mogą się różnić w zależności od zastosowanego genu i nie muszą odpowiadać ewolucyjnym
powiązaniom pomiędzy gatunkami. Podobnie jak drzewa oparte na morfologii nie zawsze są zgodne z ewolucją danej grupy taksonomicznej. Przykładem może być kłos u traw. Gdyby przyjąć tę cechę morfologiczną za wyznacznik przebiegu filogenezy to jęczmień i życica powinny być zaklasyfikowane jako blisko spokrewnione, a tak nie jest. Z drugiej strony analizy pojedynczych sekwencji mogą być pomocne zwłaszcza, gdy niewiele wiemy o danej grupie systematycznej, trudno jest pozyskać materiał, czy też poszukujemy markerów różniących dobrze wyodrębnione gatunki. Nie zwalnia to jednak od analizy większej próby niż pojedyncze osobniki. Ewolucja to coś więcej niż tylko pobieżne śledzenie historii gatunków, rysowanie drzew filogenetycznych, a nawet poszukiwanie markerów. Ewolucja to także mechanizmy będące przyczyną różnicowania gatunków. Mutacje w genie nie muszą przełożyć się na zmianę gatunkową. W locus mlo u jęczmienia odpowiedzialnym za odporność na mączniaka stwierdzono kilkanaście mutacji, które różnią jedynie odmiany. Region bz kukurydzy różni się u poszczególnych odmian całym spektrum transpozonów oraz liczbą genów w regionie, która wynosi od 6 do 10. Pomimo tak istotnych zmian nie ma podstaw do mówienia o różnych gatunkach, gdyż nie powstała bariera reprodukcyjna. Co więcej mieszańce między genetycznie różnymi odmianami wykazują zwiększoną żywotność i plenność.
Termin ewolucja pochodzi od łacińskiego słowa „evolvere”, który oznacza rozwijać, unaoczniać.
Współcześnie ewolucja biologiczna oznacza zmiany dziedziczne zachodzące pod wpływem środowiska w określonym czasie. Tak więc podstawą ewolucji jest zmienność – różnorodność form, cech w obrębie gatunku, a także pomiędzy gatunkami. Bez zmienności nie ma mowy o ewolucji i dlatego zmiana jest podstawowym zdarzeniem ewolucyjnym. Znaczenie ewolucyjne mają tylko zmiany dziedziczne. Zmiany niedziedziczne, nawet jeżeli mają podłoże genetyczne (mutacja w komórkach somatycznych) giną wraz z osobnikiem. Zmienność dziedziczna to zmienność rekombinacyjna i mutacyjna. Rekombinacja umożliwia tworzenie nowych układów genów, natomiast mutacje prowadzą do powstania nowych genów. Mutacje całego genomu, takie jak poliploidyzacja, mogą być przyczyną szybkiej specjacji. Tak powstała pszenica – podstawa diety współczesnego

11. O EWOLUCJI…
©Kornelia Polok 56
człowieka. Żeby jednak zmienność miała znaczenie ewolucyjne musi działać dodatkowy czynnik – selekcja. Teoria powstawania gatunków w wyniku selekcji najlepiej przystosowanych form była największym osiągnięciem Karola Darwina. Selekcja działa na przestrzeni czasu. Tak, więc mamy czwarty czynnik będący podstawą ewolucji - czas.
Obecnie nie ulega wątpliwości, że nie można zrozumieć mechanizmów ewolucji bez zrozumienia
zmienności i jej genetycznego podłoża. Jednak nie zawsze było to oczywiste i nadal, dla niektórych nie jest oczywiste. Pierwszym człowiekiem, który powiązał Darwinowską teorię powstawania gatunków ze zmiennością genetyczną był Theodosius Dobzhansky. Jako pierwszy zrozumiał on znaczenie zmienności genetycznej w populacjach naturalnych. Wywodząc się jednocześnie ze szkoły Thomasa Morgana (był doktorantem w jego laboratorium), a także posiadając tradycyjną wiedzę i doświadczenie dotyczące historii naturalnej (wykształcił się w Kijowie), Dobzhansky stworzył ramy, które połączyły materialne podstawy ewolucji (geny) opisywane przez genetykę klasyczną ze zróżnicowaniem gatunków obserwowanym przez ewolucjonistów. Było to możliwe dzięki jego kreatywności i interdyscyplinarnym umiejętnościom. Był jednym z pierwszych naukowców, który zrozumiał, że do poznania mechanizmów ewolucji niezbędna jest wiedza z wielu obszarów.
W książce „Genetics and the Origin of Species (1937, 1951; ISBN 0-231-05474-0) T. Dobzhanski pierwszy
wykazał związek pomiędzy mikroewolucją (procesy obserwowane na poziomie populacji i gatunku), a makroewolucją (duże zmiany ewolucyjne prowadzące do powstawania wyższych taksonów). Przekonująco zademonstrował on, że badania laboratoryjne materiału genetycznego dostarczają informacji o mechanizmach ewolucji. Książka Dobzhansky’ego nie tylko przyczyniła się do lepszego zrozumienia mechanizmów ewolucji, ale także stworzyła ramy dla współpracy pomiędzy różnymi specjalistami. Dlatego ma ona nie tylko znaczenie naukowe, ale należy ją uznać za podstawę współczesnego zrozumienia badań – podejścia interdiscyplinarnego, w którym istotne jest połączenie wysiłków różnych specjalistów. Obserwowane cechy należy widzieć w szerszym kontekście, nawet wówczas, gdy analiza dotyczy takich zagadnień, wydawałoby się odległych od rozważań ewolucyjnych, jak odporność na aluminium u roślin, wyższa plenność czy rozprzestrzenianie się chorób w populacjach ludzkich. Mechanizmy ewolucji powinny być rozumiane przez każdego wykształconego człowieka, zwłaszcza gdy chce zajmować się naukami biologicznymi.

11. O EWOLUCJI…
©Kornelia Polok 57
Dlatego mówiąc o T. Dobzhansky’m nie można zapominać, że jego niezaprzeczalny wkład w biologię
polegał także na zmianie sposobu myślenia. T. Dobzhansky przekonująco wytłumaczył matematyczne podstawy teorii opisującej zachowanie genów w populacjach, zebrał dane z różnych obszarów biologii reprezentowanych przez różne szkoły, przełamał bariery koncepcyjne i stworzył nowy spójny sposób myślenia. Dodatkowo, jasny, łatwy do zrozumienia dla niespecjalisty wywód, umożliwił zainspirowanie i zjednoczenie większości zantagonizowanych szkół biologicznych. Istotne było uproszczenie matematycznych teorii Fishera, Wrighta i Haldana, którzy faktycznie dali podwaliny pod syntetyczną teorię ewolucji. Dzięki prostocie wywodu, więcej niż połowa czytelników pierwszego wydania wysoko oceniła tę monografię jako wyjątkowo zrozumiale napisaną. William B. Brierley, recenzent wydawniczy, który określił się jako typowy botanik/zoolog napisał, że
„Prof. Dobzansky napisał najbardziej stymulującą książkę. Posiada on zdolność prostego mówienia
o rzeczach trudnych bez ich upraszczania, opisuje idee zwięźle, interesującym językiem”. Dobzansky uzyskal ten efekt
poprzez definiowanie używanych terminów, ilustrowanie przykładami rozważań teoretycznych, opis eksperymentów i dokumentację graficzną – atrybuty typowe dla współczesnego, dobrego języka naukowego. Dzięki wpływowi jaki jego książka wywarła na środowisko naukowe T. Dobzhansky faktycznie doprowadził do przekształcenia się biologii dziewiętnastowiecznej, dla której charakterystyczne były wąskie specjalności, w biologię współczesną, której najbardziej zaawansowanym przejawem jest zintegrowana genomika i biologia systemowa. Nie bez przyczyny, książka Dobzhansky’ego została uznana za jedną z najważniejszych pozycji będących podstawą współczesnej syntetycznej teorii ewolucji i w 1990 roku została zaliczona do najważniejszych książek świata zachodniego (Great books of the Western Word) razem z pracami Karola Darwina.
W duchu tejże interdyscyplinarności zostały przedstawione zagadnienia ewolucyjne w niniejszej książce. Nie jest jej celem wykład na temat ewolucji, ale pokazanie, ze teoria ewolucji jest najbardziej ogólną teorią w biologii oraz, że jej zrozumienie ma istotne znaczenie we współczesnym świecie. Przedstawione problemy i pytania mają także unaocznić, że nie można rozpatrywać ewolucji w oderwaniu od ewolucji Ziemi i Kosmosu. To właśnie unikalne właściwości naszej planety umożliwiły rozwój życia biologicznego. Nie można ewolucji rozpatrywać w oderwaniu od zachowań współczesnego człowieka. Rasizm nie ma prawa bytu nie dlatego, że tak jest poprawnie politycznie, ale dlatego, że kolor skóry czy inne cechy różniące rasy to przejaw zmienności wewnątrzgatunkowej powstałej w wyniku przystosowania się populacji do lokalnych warunków.

©Kornelia Polok 58
GGGEEENNNEEETTTYYYKKKAAA PPPOOOPPPUUULLLAAACCCYYYJJJNNNAAA
GGGeeennneeetttyyykkkaaa pppooopppuuulllaaacccyyyjjjnnnaaa zzzaaajjjmmmuuujjjeee sssiiięęę bbbaaadddaaannniiieeemmm gggeeennnóóówww www pppooopppuuulllaaacccjjjaaaccchhh... AAAnnnaaallliiizzzuuujjjeee ooonnnaaa zzzmmmiiieeennnnnnooośśśććć gggeeennneeetttyyyccczzznnnąąą,,, ccczzzęęęssstttooośśśććć aaalllllleeellliii iii zzzmmmiiiaaannnyyy ccczzzęęęssstttooośśśccciii aaalllllleeellliii pppoooddd wwwpppłłłyyywwweeemmm
ppprrrzzzyyypppaaadddkkkooowwwyyyccchhh zzzdddaaarrrzzzeeeńńń iii sssyyysssttteeemmmaaatttyyyccczzznnnyyyccchhh ppprrroooccceeesssóóówww eeewwwooollluuucccyyyjjjnnnyyyccchhh...
Przykład 1. Prawo Hardy-Weinberga
U Dreissena polymorpha z jeziora Czarnogłowy (zachodniopomorskie) występują dwa allele w locus Mdh kodującym dehydrogenazę jabłczanową. Analiza enzymatyczna 200 osobników wykazała, że 111 z nich posiadało jeden prążek charakterystyczny dla homozygot względem allela F, 13 posiadało jeden prążek charakterystyczny dla homozygot względem allela S, natomiast 76 osobników miało fenotyp dwuprążkowy, typowy dla heterozygot FS.
a) Oblicz częstości obu alleli w populacji D. polymorpha. b) Określ czy populacja znajduje się w równowadze Hardy-Weinberga.
Populacja to zbiór osobników danego rodzaju, np. zbiór osobników jednego gatunku (ang. total population) lub grupa organizmów jednego gatunku na określonym obszarze. Populacja na ogół jest zbyt duża, aby badać wszystkie osobniki do niej należące, dlatego wybiera się grupę reprezentatywną (próbę). Wybór osobników musi być losowy, a grupa dostatecznie duża. W genetyce populacyjnej przyjmuje się minimalną liczebność próby wynoszącą 20‐30 osobników w przypadku analiz morfologicznych i enzymatycznych. Jednakże liczba ta może być mniejsza (np. 10), jeżeli wykorzystujemy dużą liczbę markerów DNA (np. 100). ROZWIĄZANIE – PUNKT A
Każdy gen może istnieć w różnych formach – allelach. Jest to tak zwany polimorfizm. W analizowanej populacji wystąpiły dwa allele, przy czym allele te są kodominujące. Dzięki temu możliwe jest rozróżnienie wszystkich genotypów. Znając liczebności poszczególnych genotypów znamy również liczebności poszczególnych alleli.
Ponieważ D. polymorpha jest organizmem diploidalnym, każdy osobnik posiada dwa allele. Analizowano łącznie 200 osobników, czyli analizowano 2 x 200 = 400 alleli.
Liczebności alleli były następujące:
Allel F występował u homozygot FF oraz u heterozygot FS. Stwierdzono 111 homozygot FF, z których każda posiadała 2 allele F, w sumie, 2 x 111 = 222 allele. Dodatkowo zaobserwowano 76 heterozygot FS, każda tylko z jednym allelem F, czyli 76 alleli F. Liczebność allela F wynosiła 222 + 76 = 298.
Allel S występował u homozygot SS oraz heterozygot FS. Stwierdzono 13 homozygot SS, każda miała 2 allele S, czyli liczebność wynosiła 2 x 13 = 26 alleli. Dodatkowo po jednym allelu S posiadała każda z 76 heterozygot. Całkowita liczba alleli S wynosiła 26 + 76 = 102.

11. GENETYKA POPULACYJNA
©Kornelia Polok 59
Oznaczmy częstość allela F jako p, a allela S jako q. Częstości alleli były następujące:
Allel F 298/400 = 0,745 p = 0,745 Allel S 102/400 = 0,255 q = 0,255
Zauważmy, że częstości obu alleli sumują się do jedności, p + q = 1. Oba allele występują także ze stosunkowo wysoką częstością, znacznie przekraczającą 0,01. Mówimy, że locus Mdh jest polimorficzne.
Polimorfizm oznacza występowanie co najmniej dwóch alleli w locus, przy czym częstość każdego allela musi być większa od 0,01. Częstości niższe od 0,01 traktowane są jako jednorazowe mutacje. Częstości alleli oznacza się małymi literami. Dla dwóch alleli w locus jest to najczęściej p i q, przy czym jeżeli jeden z alleli jest dominujący to p odnosi się do allela dominującego, a q recesywnego. Dla większej liczby alleli wprowadza się dodatkowe oznaczenia, np. k, r itd. Częstości genotypów oznacza się dużymi literami. Dla dwóch alleli są to odpowiednio P i Q. Literą H oznacza się heterozygoty. W przypadku większej liczby alleli, częstości homozygot oznacza się dużymi literami odpowiadającymi oznaczeniom częstości alleli (np. K, R), natomiast częstość heterozygot zapisuje się za pomocą symboli dwuliterowych (np. PK, PR, KR).
ROZWIĄZANIE – PUNKT B
Obliczenie częstości alleli na podstawie liczebności genotypów było możliwe dzięki możliwości ich rozróżnienia, wynikającej z kodominacji. Metoda ta nie może być zastosowana, gdy jeden z alleli jest dominujący. Nie jest możliwe wówczas rozróżnienie homozygot dominujących od heterozygot i tym samym nie można policzyć liczby alleli. W takim przypadku częstości alleli można określić na podstawie częstości genotypów korzystając z Prawa Hardy-Weinberga. Prawo to można zastosować, gdy populacja jest dostatecznie duża, osobniki swobodnie się krzyżują i nie działa selekcja. Prześledźmy założenia Prawa Hardy-Weinberga wykorzystując nasz przykład.
Zależności pomiędzy częstościami alleli a częstościami genotypów opisywane przez Prawo Hardy-Weinberga najłatwiej pokazać na szachownicy. Jeżeli osobniki w danej populacji krzyżują się losowo, to częstość danego genotypu zależy od prawdopodobieństwa wystąpienia gamet, z których ten genotyp powstaje.
Homozygota FF powstaje z połączenia gamety z allelem F jednego osobnika, z taką samą gametą drugiego osobnika. Prawdopodobieństwo, że dana gameta zawiera allel F wynosi p (w praktyce jest to częstość występowania allela F). Prawdopodobieństwo powstania homozygoty FF jest iloczynem prawdopodobieństwa wystąpienia gamet z allelem F, czyli p x p = p2 = P. Podobnie obliczany prawdopodobieństwo wystąpienia homozygot SS - q x q = q2 = Q. Heterozygoty FS powstają z połączenia gamety z allelem F oraz gamety z allelem S. Istnieją dwie możliwości takiego połączenia, gameta z allelem F pochodzi od matki, a z allelem S od ojca lub odwrotnie. Ponieważ obie możliwości są równie prawdopodobne, prawdopodobieństwo wystąpienia heterozygoty wynosi 2 x p x q = 2pq = H.
Zauważmy, że częstości genotypów sumują się do jedności, P + Q + H = 1. Jeżeli genotypy zapiszemy za pomocą prawdopodobieństwa ich wystąpienia to otrzymamy równanie Hardy-Weinberga.
P + Q + H = p2 + 2pq + q2 Ponieważ P + Q + H = 1 to p2 + 2pq + q2 = 1 Wyrażenie p2 + 2pq + q2 jest rozszerzeniem wyrażenia (p + q)2

11. GENETYKA POPULACYJNA
©Kornelia Polok 60
Wyrażenie (p + q)2 = p2 + 2pq + q2 nosi nazwę Prawa (Równanie) Hardy‐Weinberga. Opisuje ono zależności między częstościami alleli oraz częstościami genotypów w populacjach. Równanie to jest słuszne w dużych populacjach, gdy osobniki krzyżują się losowo i nie występuje selekcja. Takie populacje noszą nazwę populacji mendlowskich. W przypadku alleli dominujących Równanie Hardy‐Weinberga pozwala obliczyć prawdopodobne częstości alleli na podstawie częstości genotypów. Prawo Hardy‐Weinberga służy do oceny czy obserwowane częstości genotypów są zgodne z częstościami genotypów obliczonymi na podstawie równania. Jeżeli obserwowane i oczekiwane częstości genotypów są zgodne mówimy, że populacja jest w równowadze Hardy‐Weinberga. Brak takiej zgodności może świadczyć o selekcji zachodzącej w populacji.
Prawo Hardy-Weinberga pozwala sprawdzić czy obserwowane częstości genotypów są zgodne z częstościami oczekiwanymi, obliczonymi na podstawie częstości alleli. Częstości genotypów w naszym przykładzie były następujące:
Homozygoty FF 111/200 = 0,555 P = 0,555 Homozygoty SS 13/200 = 0,065 Q = 0,065 Heterozygoty FS 76/200 = 0,380 H = 0,380
Wykorzystajmy zależność pomiędzy częstościami genotypów, a częstościami alleli wynikająca z Prawa Hardy-Weinberga:
Homozygoty FF P = p2 P = p2 = 0,7452 = 0,555 Homozygoty SS Q = q2 Q = q2 = 0,2552 = 0,065 Heterozygoty FS H = 2pq H = 2pq = 2 x 0,745 x 0,255 = 0,379.
Ponieważ obserwowane I oczekiwane częstości genotypów są prawie identyczne możemy powiedzieć, że populacja D. polymorpha z jeziora Czarnogłowy jest w stanie równowagi Hardy-Weinberga.
Jeżeli jeden z alleli jest dominujący, to częstości alleli możemy obliczyć na podstawie częstości genotypów.
Częstość allela recesywnego,
Częstość allela dominującego, p + q = 1, p = 1 – q Jeżeli powyższe zależności zastosujemy do naszego przykładu, gdzie Q = 0,065 to q = 0,255 Wartość p obliczamy pośrednio z równania p = 1 – 0,255 = 0,745.
Ćwiczenia i zadania
Ćwiczenie 1. Wpływ mutacji na populację
Celem ćwiczenia jest symulacja zmian częstości alleli w populacji pod wpływem mutacji. Dla uproszczenia symulacji i zmniejszenia liczby możliwych kombinacji założono, że jest to populacja roślin samopylnych. W takiej populacji krzyżowanie nie jest całkowicie losowe (nie następuje zapłodnienie krzyżowe). Osoby zainteresowane mogą podobną symulację przeprowadzić dla roślin obcopylnych.
PRZEBIEG ĆWICZENIA
Zakładamy, że populacja roślin jednorocznych, samopylnych składa się z 5 osobników o genotypie AA (pokolenie nr 1). Jaka jest częstość allela A w tej populacji?
W populacji u jednego z osobników zachodzi mutacja allela A w kierunku allela recesywnego a. Populacja rozmnaża się swobodnie i daje następne pokolenie (nr 2). Jakie są częstości alleli A i a w tym pokoleniu nr 2?
Pokolenie nr 2 rozmnaża się swobodnie dając pokolenie nr 3. Podaj częstości alleli w pokoleniu nr 3 powstałym z samozapylenia pokolenia nr 2.
Q q q =2=

11. GENETYKA POPULACYJNA
©Kornelia Polok 61
Podaj częstości alleli A i a w trzech następujących po sobie pokoleniach roślin z punktu przy założeniu, że w każdym pokoleniu (nr 1 i 2 i 3) zachodzi mutacja A w kierunku a u jednego osobnika.
Jakie będą częstości alleli jeżeli osobniki AA będą wydawały o połowę mniej potomstwa niż osobniki Aa i aa?.
Ćwiczenie 2. Wpływ selekcji
Podobnie jak w ćwiczeniu 1, w celu uproszczenia symulacji przyjęto, że populacja składa się z roślin samopylnych. Należy jednak pamiętać, że w takiej populacji krzyżowania nie są w pełni losowe.
PRZEBIEG ĆWICZENIA
Sporządź model homozygot dominujących, recesywnych i heterozygot wykorzystując np. różnobarwne kartoniki, kule lub nasiona łubinu. Przykładowo oznacz kolorem czarnym homozygoty dominujące AA, kolorem białym, homozygoty recesywne aa, a kolorem szarym heterozygoty Aa. Każdy genotyp powinien być reprezentowany przez co najmniej 20 osobników.
Umieść wszystkie osobniki w jednym pojemniku, a następnie wybierz spośród nich losowo 10 modeli. Podaj liczebności wybranych genotypów. Załóżmy, że otrzymana w ten sposób populacja, to populacja roślin jednorocznych, samopylnych. Podaj częstości alleli w tej populacji.
Przez 5 pokoleń prowadź selekcję kierunkową skierowaną przeciwko:
a) homozygotom dominującym, b) homozygotom recesywnym, c) heterozygotom.
Podaj częstości alleli w 4 i 5 pokoleniu. Jak zmieniły się częstości alleli?
Ćwiczenie 3. Wykorzystanie izoenzymów
Izoenzymy są powszechnie wykorzystywane w badaniach populacyjnych. Ich zastosowanie w latach 60-70-tych ubiegłego wieku umożliwiło po raz pierwszy poznanie zmienności genetycznej w populacjach naturalnych. To enzymom zawdzięczamy wiedzę o procesach mikroewolucyjnych. Celem ćwiczenia jest wykorzystanie tej metody do oceny częstości alleli u dowolnego gatunku. Jednakże najlepiej wykonać to ćwiczenie dla gatunku, u którego metoda nie stwarza problemów. Do takich gatunków należą groch, jęczmień, owies, życice i wiele innych. Najłatwiej wykonać rozdział w żelu skrobiowym, wybarwiając esterazy lub peroksydazy.
PRZEBIEG ĆWIZENIA Do elektroforezy enzymów wykorzystaj bufor litowo-borowy, pH 8,1 o składzie:
a) Bufor elektrodowy, pH 8,1 0,028 M LiOH x H2O 0,192 M H3BO3
2,4 g LiOH oraz 23,78 g H3BO3 rozpuść w 1,5 l wody dejonizowanej, doprowadź pH do 8,1 i uzupełnij wodą dejonizowaną do 2 l.
b) Bufor żelowy, pH 8,1
0,5 M TRIS 0,8 M kwas cytrynowy
27,8 g TRIS oraz 7,8 g kwasu cytrynowego rozpuść w 450 ml wody dejonizowanej, doprowadź pH do 8,1 i uzupełnij wodą dejonizowaną do 500 ml.
Do wykonania żelu użyj 10-krotnego rozcieńczenia buforu żelowego i elektrodowego. Doprowadź pH roztworu do 8,1 za pomocą 1 N LiOH lub 1 N H3BO3. Do wykonania bloku żelowego o długości 9,7 cm, szerokości 19,5 cm i grubości 0,6 cm wykorzystaj 125 ml roztworu, z czego 100 ml zagotuj, a w pozostałych 25 ml rozpuść skrobię

11. GENETYKA POPULACYJNA
©Kornelia Polok 62
firmy Sigma w ilości 13,75 g (11% żel skrobiowy). Roztwór po zagotowaniu wlej do kolby ssawkowej z rozpuszczoną skrobią, całość intensywnie mieszaj i wylej na płytę szklaną ograniczoną plastikowymi listewkami.
W celu uzyskania ekstraktu z siewki zerwij liść z rośliny bezpośrednio przed wykonaniem analizy. Fragment liścia przepłucz w wodzie dejonizowanej, osusz na bibule filtracyjnej, potnij i umieść w naczyniu ekstrakcyjnym wykonanym z plexi, po czym rozetrzyj w 2-3 kroplach wody dejonizowanej (peroksydazy) lub β-merkaptoetanolu (esterazy i inne enzymy)z dodatkiem oczyszczonego i sterylizowanego piasku kwarcowego. Uzyskanym ekstraktem nasącz paski bibuły Whatman 3 MM o wymiarach 6 mm x 5 mm.
Nasączone ekstraktem bibułki w liczbie 20 sztuk (dla 20 roślin) umieść w szczelinie żelu powstałej w wyniku jego przecięcia w odległości 2,5 cm od początku. Wypełnij komory buforem elektrodowym i umieść naczynie z żelem na komorach łącząc żel z buforem za pomocą gąbek. Przykryj żel folią i połóż szklaną płytkę na górę. Żel umieść w lodówce i podłącz prąd ("-" od strony bibułek). Elektroforezę prowadź w lodówce w temp. 4 C, najpierw przez 0,5 h przy napięciu 250 V i natężeniu 50 mA. Po tym czasie wyjmij bibułki i kontynuuj elektroforezę przez 2 h przy 300 V i 50 mA. Po zakończonej elektroforezie żel pozostaw w lodówce na około 0,5 h celem jego schłodzenia. Następnie żel wyjmij z lodówki i potnij na 2 warstwy, każda o grubości 0,2 cm. Warstwę zewnętrzną odrzuć, a pozostałą przepłucz w wodzie dejonizowanej i wybarw specyficznie na peroksydazę.
Barwienie peroksydazy.
3-amino-9-etylokarbazol, 100 mg dwumetyloformamid, 3,5 ml 3% H2O2, 1 ml 1% MgCl2 x 6 H2O, 2 ml 1 M bufor octanowy, pH 5,0, 5 ml (1 N NaOH doprowadzony CH3COOH do pH 5,0) H2O dejonizowana, 90 ml
Składniki dodaj w kolejności wyżej podanej, po czym tak przygotowanym roztworem zalej żel i inkubuj w temp. pokojowej przez 3-4 h.
Żel utrwal w mieszaninie alkoholu metylowego, kwasu octowego i wody w stosunku 5:1:5. Utrwalone żele zamknij w folię i przechowuj w lodówce.
Odczytaj żel
a) Ile stref aktywności możesz wyróżnić na żelu? b) Ile loci enzymatycznych możesz wyróżnić? c) Ile prążków o różnej ruchliwości można wyróżnić w każdej strefie? d) Ile różnych alleli obserwujesz? e) Ile i jakie zaobserwowałeś homozygoty? f) Ile i jakie zaobserwowałeś heterozygoty? g) Oblicz częstość każdego z obserwowanych przez Ciebie alleli. h) Czy analizowana populacja jest w równowadze Hardy-Weinberga?
Zadania
1. Jakie są częstości grup krwi MM, NN, MN populacji ludzkiej w Afryce, jeżeli częstości alleli wynoszą 0,80 dla allela LM oraz 0,2 dla LN?
2. U Dreissena polymorpha z jeziora Ińsko stwierdzono występowanie czterech alleli w locus transaminazy asparaginianowej, AAT. Częstości alleli wynosiły odpowiednio p(A) = 0,866, q(B) = 0,055, k(D) = 0,068 oraz r(E) = 0,011. Podaj odpowiednie częstości wszystkich genotypów jakie mogą pojawić się w populacji.
3. W populacji ludzkiej osobniki albinotyczne występują z częstością 0,0004. Albinizm jest wywołany recesywnym allelem a. Jaka jest częstość tego allela w populacji ludzkiej? Jaka jest częstość allela dominującego A?

©Kornelia Polok 63
GGGEEENNNEEETTTYYYKKKAAA EEEWWWOOOLLLUUUCCCYYYJJJNNNAAA
EEEwwwooollluuucccjjjaaa jjjeeesssttt ppprrroooccceeessseeemmm gggeeennneeetttyyyccczzznnnyyymmm... WWWyyykkkrrryyyccciiieee PPPrrraaawww MMMeeennndddlllaaa pppoootttwwwiiieeerrrdddzzziiiłłłooo iiidddeeeęęę DDDaaarrrwwwiiinnnaaa... ZZZmmmiiieeennnnnnooośśśććć gggeeennneeetttyyyccczzznnnaaa jjjeeesssttt pppooodddssstttaaawwwąąą eeewwwooollluuucccjjjiii... BBBiiiooolllooogggiiiaaa eeewwwooollluuucccyyyjjjnnnaaa zzzaaajjjmmmuuujjjeee
sssiiięęę aaannnaaallliiizzząąą zzzmmmiiieeennnnnnooośśśccciii nnnaaa wwwssszzzyyyssstttkkkiiiccchhh pppooozzziiiooommmaaaccchhh ooorrrgggaaannniiizzzaaacccjjjiii bbbiiiooolllooogggiiiccczzznnneeejjj...
Przykład 1. Powiązania filogenetyczne w rodzaju Pinus
Markery molekularne są powszechnie wykorzystywane w badaniach ewolucyjnych. Jednym z lepszych tego typu przykładów jest P. sylvestris oraz trzy blisko spokrewnione gatunki z kompleksu P. mugo, P. mugo Turra (kosodrzewina), P. uncinata (sosna hakowata) i P. uliginosa (sosna błotna). Sosna zwyczajna jest monokormicznym drzewem o bardzo szerokim zasięgu sięgającym od południowych części Hiszpanii po Krąg Polarny, kosodrzewina jest rośliną polikormiczną o zasięgu ograniczonym do piętra sub-alpejskiego Europy Centralnej i Południowej, natomiast sosna hakowata jest monokormicznym drzewem typowym dla Pirenejów i Alp. Sosna błotna jest również monokormicznym taksonem znanym głównie z torfowiska „Wielkie Batorowskie” w południowo-zachodniej Polsce. Wszystkie wymienione gatunki mają takie same wzory enzymatyczne, a podobieństwo genetyczne wyznaczone na podstawie elektroforezy enzymów mogłoby je uplasować w randze populacji jednego gatunku. Uważa się także, że P. sylvestris i P. mugo mogą tworzyć mieszańce na stanowiskach gdzie występują razem. Jednym z lepiej poznanych takich stanowisk jest Bór na Czerwonem, gdzie wyodrębniono nothospecies Pinus x rhaetica. Dyskusja toczy się także odnośnie statusu taksonomicznego P. uliginosa, która jest uważana albo za mieszańca pomiędzy P. mugo i P. sylvestris, albo za marginalną populację P. uncinata. Dostępne dane biometryczne i enzymatyczne nie pozwalają na jednoznaczne ustalenie powiązań filogenetycznych pomiędzy blisko spokrewnionymi sosnami. Blisko spokrewnione gatunki często są bardziej podobne morfologicznie niż gatunki odległe. Jednakże nawet gatunki blisko spokrewnione na ogół różnią się wieloma, a nie pojedynczymi cechami uwarunkowanymi genetycznie.
W celu oceny powiązań filogenetycznych pomiędzy blisko spokrewnionymi sosnami wykorzystano kilka kategorii markerów DNA, w tym RAPD, ISJ i wysoce efektywne w badaniach filogenetycznych markery B-SAP, wyprowadzone z bakteryjnego genu KatG, kodującego katalazę-peroksydazę, sekwencję inercyjną IS6110 oraz genów Pol i Rpl. W tabeli podano wynik sumaryczny w postaci łącznej liczby prążków zidentyfikowanych u każdego taksonu (T) oraz liczbę prążków wspólnych dla każdej pary taksonów (S). Dla uproszczenia obliczeń pominięto dane dla poszczególnych osobników.

13. GENETYKA EWOLUCYJNA
©Kornelia Polok 64
P. sylvestris P. mugo P. uliginosa P. uncinata P. sylvestris T = 103 S = 65 S = 50 S = 79 P. mugo T = 137 S = 116 S = 91 P. uliginosa T = 109 S = 74 P. uncinata T = 103
a) Na podstawie uzyskanych wyników określ powiązania ewolucyjne pomiędzy taksonami sosen. b) Czy wszystkie cztery taksony można uznać za gatunki?
Podstawą analiz nad ewolucją jest rozróżnienie procesów zachodzących wewnątrz gatunku od procesów prowadzących do powstawania gatunków (specjacji). W praktyce oznacza to odróżnienie zmienności wewnątrzgatunkowej od zmienności pomiędzy gatunkami. Na poziomie molekularnym (enzymy, DNA) należy odpowiedzieć na pytanie, czy obserwowane warianty u różnych genów, markerów prezentują jeszcze zmienność alleliczną, czy też różnicują gatunki. Dlatego istotna jest analiza wielu osobników z danego gatunku. Analizy pojedynczych osobników nawet na poziomie sekwencji DNA nie pozwalają określić charakteru obserwowanej zmienności. Istotnym aspektem jest także konieczność analizy wielu sekwencji, markerów, cech. Dopóki analizuje się jedną sekwencję dopóty nie można mówić o ewolucji gatunków. Analiza jednej sekwencji pozwala wypowiadać się tylko na temat ewolucji badanej sekwencji.
ROZWIĄZANIE – PUNKT A
Ewolucja jest procesem genetycznym. Gatunki blisko spokrewnione są bardziej podobne genetycznie niż gatunki bardziej odległe. Inaczej mówiąc, gatunki blisko spokrewnione mają bardziej podobny genom i tym samym więcej sekwencji/genów wspólnych niż odległe taksony. Dlatego, aby ocenić zależności ewolucyjne pomiędzy poszczególnymi taksonami należy określić na ile podobne są ich genomy. W tym celu należy ocenić udział sekwencji wspólnych dla każdej pary gatunków, a następnie pogrupować gatunki na podstawie sekwencji wspólnych tworząc drzewo filogenetyczne.
W celu oceny różnic genetycznych pomiędzy taksonami, ale także pomiędzy populacjami tego samego gatunku należy obliczyć współczynnik podobieństwa genetycznego (I) lub jego odwrotność – odległość genetyczną (D). Pozwala on ilościowo ocenić stopień różnic genetycznych pomiędzy taksonami (populacjami). Najpowszechniej stosowaną miarą jest współczynnik podobieństwa genetycznego Nei’a (I), który wprowadzono dla enzymów, ale znajduje on również zastosowanie dla DNA.
I – podobieństwo genetyczne D – odległość genetyczna px – częstość allela i w populacji x py – częstość allela i w populacji y
lnI D ; pp
pp I
yx
yx ==22∑ ∑
∑

13. GENETYKA EWOLUCYJNA
©Kornelia Polok 65
Standardowy współczynnik Nei’a wymaga obliczenia podobieństwa dla każdego locus osobno, a następnie podsumowania danych dla wszystkich loci. Dla dużej liczby markerów podobieństwo genetyczne Nei’a rutynowo oblicza się przy pomocy programów komputerowych. W naszym przykładzie, posłużymy się uproszczonym współczynnikiem podobieństwa genetycznego (Nei i Li 1979), który został opracowany dla analizy fragmentów RFLP i stosowany jest także w przypadku markerów DNA opartych o reakcję PCR.
S – liczba prążków wspólnych dla dwóch taksonów T – liczba wszystkich prążków dla danego taksonu
Pojęcie podobieństwa genetycznego i odległości genetycznej ma fundamentalne znaczenie dla analiz ewolucyjnych. Odległość genetyczna jest odwrotnością podobieństwa genetycznego i określa ona ilościowo na ile różne są dwa taksony, populacje. Najczęściej stosowaną miarą podobieństwa i odległości jest współczynnik Nei’a. Wartość podobieństwa genetycznego Nei’a mieści się w przedziale od 0 do 1. Odległość Nei’a oblicza się jako logarytm naturalny podobieństwa i wynosi ona od 0 do nieskończoności.
Wykorzystując uproszczony wzór Nei i Li (1979), obliczmy współczynnik podobieństwa genetycznego dla analizowanych taksonów. Wyniki umieśćmy w tabeli. Przykładowo, podobieństwo genetyczne pomiędzy P. sylvestris i P. mugo obliczamy następująco:
I = 2 x 65/(103+137) = 130/240 = 0,54
Podobnie obliczamy wartości podobieństwa pomiędzy pozostałymi parami taksonów.
Dane wstawiamy do tabeli. Poniżej przedstawiono tabelę z programu STATISTICA z prawidłowo wprowadzonymi danymi. W tabeli nie może być pustych komórek, dlatego wartości muszą być wstawione dwukrotnie. Kursywą zaznaczono powtórzone dane
Kolejnym etapem jest pogrupowanie taksonów na podstawie uzyskanej wartości podobieństwa. Wykorzystuje się do tego rutynowe metody grupowania (clustering) dostępne w wielu programach statystycznych. W naszym przypadku wykorzystamy metodę UPGMA dostępną w pakiecie STATISTICA w module STATISTICS/MULTIVARIATE../CLUSTER ANALYSIS. Należy wybrać opcję JOINING (TREE CLUSTERING) Do grupowania taksonów wykorzystuje się rutynowe metody statystyczne, które pozwalają uporządkować taksony na podstawie danych (np. podobieństwa) w przestrzeni 2‐ lub 3‐wymiarowej. Najczęściej stosuje się metody pozwalające na konstrukcję dendrogramów (drzew), a obecnie także analizę składowych głównych (PCA) oraz skalowanie wielowymiarowe (MDS). Metod tworzenia dendrogramów jest wiele, każda ma zalety i wady. Często stosuje się grupowanie metodą UPGMA, która pomimo pewnych wad, stosunkowo dobrze oddaje relacje pomiędzy obiektami. Danymi wejściowymi może być podobieństwo genetyczne przekształcone na macierz odległości np. na macierz odległości euklidesowej. Można też wykorzystać odległość genetyczną jako macierz odległości. Obliczenia można wykonać w programie STATISTICA, ale także w programach specjalistycznych jak POPGENE, NTSYS i wiele innych.
21
2TT
S I
+=

13. GENETYKA EWOLUCYJNA
©Kornelia Polok 66
W kolejnym etapie należy wybrać typ danych („surowe” lub macierz odległości). W naszym przypadku należy wybrać dane „surowe” (podobieństwo nie jest macierzą odległości) i przekształcić je na macierz odległości za pomocą odległości euklidesowej (distance measure). Możemy też wprowadzić wartość odległości genetycznej i zaznaczyć „macierz” jako typ danych. Należy jednak pamiętać, że tabela musi mieć wówczas charakter macierzy. Następnie należy wybrać sposób grupowania UPGMA – Unweighted pair-group average.
W wyniku przeprowadzonego grupowania otrzymaliśmy dendrogram, który przedstawia zależności pomiędzy badanymi taksonami. Na dendrogramie widoczne są dwie grupy (klastry), jedna łączy P. sylvestris i P. uncinata, a druga P. mugo i P. uliginosa, przy czym gatunki z grupy drugiej są znacznie bardziej podobne, a więc bliższe ewolucyjnie niż gatunki grupy pierwszej. Dendrogram pokazuje także, że obie grupy są dość odległe genetycznie i ewolucyjnie. Grupowanie pokazuje zależności ewolucyjne pomiędzy taksonami, jednakże na podstawie samego grupowania nie możemy stwierdzić, czy widoczne różnice mają charakter gatunkowy.
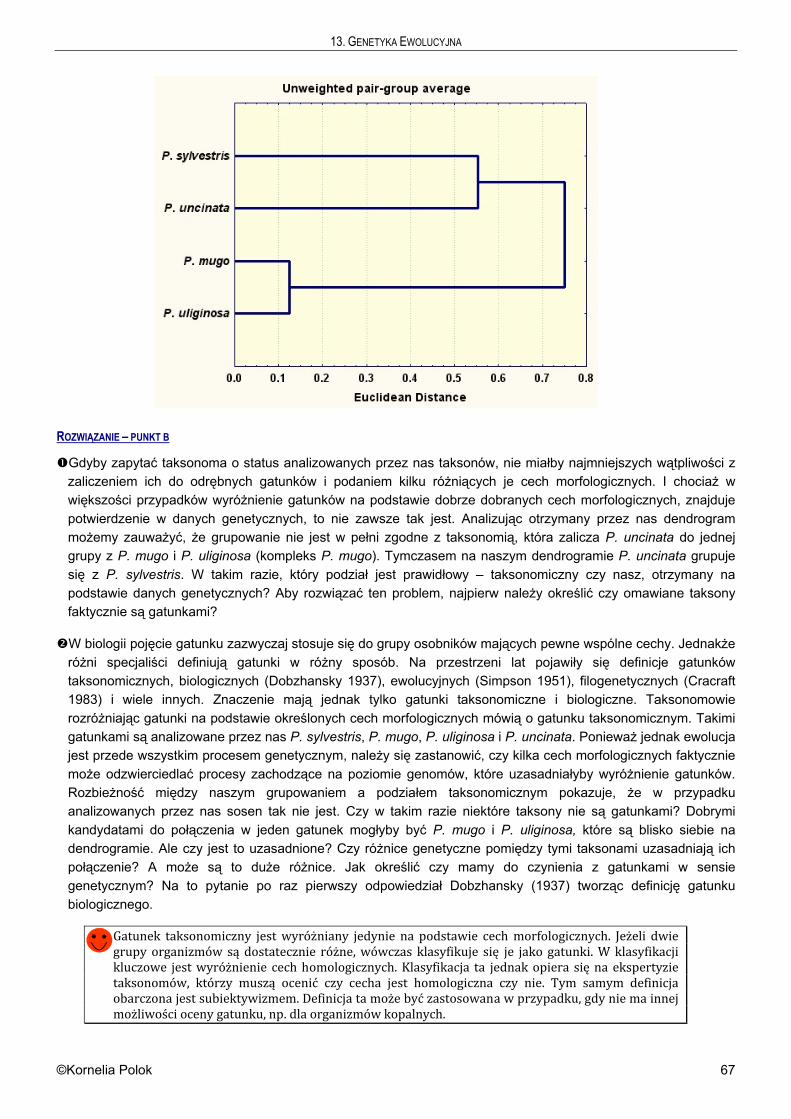
13. GENETYKA EWOLUCYJNA
©Kornelia Polok 67
ROZWIĄZANIE – PUNKT B
Gdyby zapytać taksonoma o status analizowanych przez nas taksonów, nie miałby najmniejszych wątpliwości z zaliczeniem ich do odrębnych gatunków i podaniem kilku różniących je cech morfologicznych. I chociaż w większości przypadków wyróżnienie gatunków na podstawie dobrze dobranych cech morfologicznych, znajduje potwierdzenie w danych genetycznych, to nie zawsze tak jest. Analizując otrzymany przez nas dendrogram możemy zauważyć, że grupowanie nie jest w pełni zgodne z taksonomią, która zalicza P. uncinata do jednej grupy z P. mugo i P. uliginosa (kompleks P. mugo). Tymczasem na naszym dendrogramie P. uncinata grupuje się z P. sylvestris. W takim razie, który podział jest prawidłowy – taksonomiczny czy nasz, otrzymany na podstawie danych genetycznych? Aby rozwiązać ten problem, najpierw należy określić czy omawiane taksony faktycznie są gatunkami?
W biologii pojęcie gatunku zazwyczaj stosuje się do grupy osobników mających pewne wspólne cechy. Jednakże różni specjaliści definiują gatunki w różny sposób. Na przestrzeni lat pojawiły się definicje gatunków taksonomicznych, biologicznych (Dobzhansky 1937), ewolucyjnych (Simpson 1951), filogenetycznych (Cracraft 1983) i wiele innych. Znaczenie mają jednak tylko gatunki taksonomiczne i biologiczne. Taksonomowie rozróżniając gatunki na podstawie określonych cech morfologicznych mówią o gatunku taksonomicznym. Takimi gatunkami są analizowane przez nas P. sylvestris, P. mugo, P. uliginosa i P. uncinata. Ponieważ jednak ewolucja jest przede wszystkim procesem genetycznym, należy się zastanowić, czy kilka cech morfologicznych faktycznie może odzwierciedlać procesy zachodzące na poziomie genomów, które uzasadniałyby wyróżnienie gatunków. Rozbieżność między naszym grupowaniem a podziałem taksonomicznym pokazuje, że w przypadku analizowanych przez nas sosen tak nie jest. Czy w takim razie niektóre taksony nie są gatunkami? Dobrymi kandydatami do połączenia w jeden gatunek mogłyby być P. mugo i P. uliginosa, które są blisko siebie na dendrogramie. Ale czy jest to uzasadnione? Czy różnice genetyczne pomiędzy tymi taksonami uzasadniają ich połączenie? A może są to duże różnice. Jak określić czy mamy do czynienia z gatunkami w sensie genetycznym? Na to pytanie po raz pierwszy odpowiedział Dobzhansky (1937) tworząc definicję gatunku biologicznego. Gatunek taksonomiczny jest wyróżniany jedynie na podstawie cech morfologicznych. Jeżeli dwie grupy organizmów są dostatecznie różne, wówczas klasyfikuje się je jako gatunki. W klasyfikacji kluczowe jest wyróżnienie cech homologicznych. Klasyfikacja ta jednak opiera się na ekspertyzie taksonomów, którzy muszą ocenić czy cecha jest homologiczna czy nie. Tym samym definicja obarczona jest subiektywizmem. Definicja ta może być zastosowana w przypadku, gdy nie ma innej możliwości oceny gatunku, np. dla organizmów kopalnych.

13. GENETYKA EWOLUCYJNA
©Kornelia Polok 69
Znając wartości podobieństwa odpowiadające poszczególnym etapom ewolucji możemy określić na jakim etapie znajdują się 4 analizowane przez nas taksony Pinus.
Pinus sylvestris jest dobrze wyodrębnionym gatunkiem biologicznym od P. mugo i P. uliginosa, o czym świadczą wartości podobieństwa wynoszące odpowiednio 0,54; 0,47. Wartość 0,47 pomiędzy P. sylvestris i P. uliginosa mieści się w przedziale dla gatunków bliźniaczych podobnych morfologicznie, ale o pełnej barierze reprodukcyjnej. Wiemy także, że P. sylvestris i P. uliginosa różnią się cechami morfologicznymi, a więc nie spełniają kryterium gatunków bliźniaczych o podobieństwie morfologicznym. Dlatego należy je uznać za w pełni wyodrębnione gatunki biologiczne. Wartość podobieństwa pomiędzy P. sylvestris i P. mugo jest nieco wyższa od granicy 0,517, jednakże trudności z krzyżowaniem tych gatunków wskazują na pełną barierę reprodukcyjną.
Pinus sylvestris i P. uncinata – wartość podobieństwa 0,62 odpowiadałaby statusowi podgatunków z częściową barierą reprodukcyjną.
Pinus uncinata wykazuje podobieństwo względem P. mugo i P. uliginosa odpowiednio 0,63 i 0,57, a więc również odpowiadające statusowi podgatunków z częściową barierą reprodukcyjną. Wskazywałoby to na pośredni charakter tego taksonu pomiędzy P. sylvestris, a grupą P. mugo-P. uliginosa. Aby ten status potwierdzić należałoby poszerzyć badania o liczbę prób z różnych rejonów geograficznych oraz uwzględniając dodatkowe kategorie markerów, zwłaszcza markerów transpozonowych (SSAP). Transpozony są pierwszymi sekwencjami, które różnicują populacje (Polok 2007).
P. mugo i P. uliginosa – podobieństwo wynosi 0.94, co klasyfikuje te dwa gatunki taksonomiczne jako populacje jednego gatunku. Biorąc pod uwagę fakt, że oba gatunki różnią się jedynie wysokością (karłowy i wysoki) i typem wzrostu (polikormicznym i monokormicznym) oraz, że P. mugo na niżu może tworzyć formy wysokie zaś P. uliginosa może być polikormiczna, zaklasyfikowanie obu taksonów jako jednego gatunku wydaje się uzasadnione. Różnice pomiędzy nimi są porównywalne z tymi, jakie obserwowaliśmy pomiędzy różnymi krzyżowanymi formami roślin grochu (karłowy, wysoki), jęczmienia, życic i wielu innych, w części dotyczącej genetyki. Możemy więc przyjąć, że P. mugo jest mutantem karłowym P. uliginosa.
Specjacja jest to proces powstawania gatunków. Gatunki powstają, gdy osobniki należące do subpopulacji nie mogą się krzyżować na skutek nagromadzenia różnic genetycznych pomiędzy nimi. Powstanie gatunku jest nierozerwalnie związane z powstaniem bariery reprodukcyjnej. Na ogół nie jest to proces skokowy (wyjątkiem jest specjacja przez poliploidyzację) i populacje znajdują się na różnym etapie ewolucji – od populacji jednego gatunku, przez półgatunki, podgatunki, gatunki bliźniacze, i w pełni wyodrębnione gatunki biologiczne. Podobieństwo genetyczne pozwala określić na jakim etapie dywergencji są populacje. Gatunki mogą powstawać z subpopulacji odległych geograficznie (specjacja allopatryczna) lub z subpopulacji na tym samym obszarze (specjacja sympatryczna). Niewielki przepływ genów może następować nawet pomiędzy dobrymi gatunkami biologicznymi i proces ten nosi nazwę introgresji.

13. GENETYKA EWOLUCYJNA
©Kornelia Polok 68
W genetyce ewolucyjnej przyjmuje się definicję gatunku biologicznego, zgodnie z którą o gatunkach możemy mówić gdy istnieje pomiędzy nimi bariera reprodukcyjna. O jej występowaniu można się przekonać krzyżując potencjalne gatunki. Jeżeli potomstwo nie występuje, jest nieżywotne (ginie we wczesnych etapach rozwoju) lub sterylne (mieszaniec powstaje, ale nie wytwarza żywotnych gamet i tym samym potomstwa) to mamy do czynienia z barierą reprodukcyjną. Jednakże krzyżowanie nie zawsze jest łatwe do wykonania, chociażby ze względu na długi czas oczekiwania na wynik, jak w przypadku naszych taksonów. Czy istnieje inna możliwość oceny, czy nasze taksony są gatunkami biologicznymi?
Występowanie bariery reprodukcyjnej świadczy, że gatunki nie wymieniają między sobą genów. Niemożność wymiany genów wynika z różnic genetycznych między gatunkami. Muszą one przynajmniej w części posiadać inne sekwencje DNA. Już zmiana jednej zasady drastycznie obniża prawdopodobieństwo zajścia crossing-over na danym obszarze chromosomu. Skoro gatunki nie wymieniają genów, to znaczy muszą różnić się wieloma sekwencjami DNA, a więc także posiadają różne allele w poszczególnych loci. Różnice można wykryć za pomocą markerów molekularnych takich jak enzymy i markery DNA. Nawet blisko spokrewnione gatunki muszą różnić się tysiącami markerów. Innymi słowy możemy powiedzieć, że gatunki biologiczne różnią się wieloma genami lub mają różne pule genowe.
Zgodnie z definicją Ernsta Mayra gatunki biologiczne to gatunki, które mają różne pule genowe. Gatunki takie nie wymieniają genów, a więc istnieje pomiędzy nimi bariera reprodukcyjna. Definicja gatunku biologicznego jest powszechnie przyjęta w genetyce ewolucyjnej, a także w zoologii. Botanicy często kwestionują tę definicję twierdząc, że różne gatunki roślin mogą tworzyć płodne mieszańce, a mimo to są gatunkami, bo w naturze krzyżują się rzadko i różnią się morfologicznie. Należy zaznaczyć, że bariery o charakterze etologicznym i geograficznym mogą być łatwo zniesione, natomiast krzyżowanie świadczy o braku różnic genetycznych i tym samym wyróżnienie gatunków na podstawie morfologii jest nieuzasadnione. Mutanty często różnią się szeregiem cech morfologicznych od odmian, z których je otrzymano i wiele z tych cech jest stosowanych do wyróżniania taksonów przez botaników. Mikrobiolodzy kwestionują definicję gatunku biologicznego twierdząc, że bakterie rozmnażają się wegetatywnie i nie można stwierdzić istnienia bariery reprodukcyjnej. Ten pogląd jest wyrazem niezrozumienia definicji biologicznej gatunku. Nawet dla organizmów rozmnażających się wegetatywnie można określić czy ich pule genowe są wspólne czy nie.
Aby odpowiedź na pytanie czy nasze taksony sosen mają takie same czy różne pule genowe należy przeanalizować wartości podobieństwa genetycznego. Widzimy, że waha się ono od 0,47 pomiędzy P. sylvestris i P. uliginosa do 0,94 pomiędzy P. mugo i P. uliginosa. Czy wartości w pierwszym przypadku świadczą o różnych pulach genowych, a w drugim o jednej? Gdybyśmy tak uznali byłoby to arbitralne. Skąd bowiem wiemy, że I = 0,47 świadczy o różnych pulach genowych? Bez porównania tych wartości z wartościami uzyskiwanymi dla gatunków nie budzących wątpliwości lub bez porównania podobieństwa Nei’a z krzyżowalnością nie jesteśmy w stanie ocenić znaczenia wartości I. Na szczęście począwszy od lat 60-70-tych ubiegłego wieku, gdy analiza enzymów pozwoliła poznać zróżnicowanie populacji i gatunków, do dnia dzisiejszego dysponujemy tysiącami przykładów dla różnych grup roślin i zwierząt, które powiązały istnienie bariery reprodukcyjnej z wartością podobieństwa genetycznego (Futuyma 1987; Avise 2004; Zielinski i Polok 2005). I tak, gatunki biologiczne z pełną barierą reprodukcyjną mają wartość I poniżej 0,352, natomiast populacje jednego gatunku od 0,873 do 1,00. Pomiędzy tymi wartościami istnieje cały szereg form pośrednich, z częściowo wykształconą barierą reprodukcyjną. Gatunki kryptyczne lub bliźniacze (sibling species), podobne morfologicznie, ale z pełną barierą reprodukcyjną, mają wartość podobieństwa pomiędzy 0,352 – 0,517. Wartości pomiędzy 0,517 a 0,795 są typowe dla podgatunków, które występują na różnym obszarze i posiadają częściową barierę reprodukcyjną (subspesies). Wartości pomiędzy 0,795 a 0,873 są typowe dla gatunków na początkowym etapie ewolucji (półgatunki - semispecies), które nie krzyżują się z przyczyn zewnętrznych, jak różna etologia czy różny obszar występowania. Oczywiście wartości te nie są arbitralne i istnieją liczne wyjątki. Przykładowo poliploidyzacja prowadzi do powstania gatunków bez różnicowania puli genowej. Podobieństwo może wynosić np. 0,95 a pomimo tego istnieje bariera reprodukcyjna spowodowana różną liczbą chromosomów. Jednakże wartości powyższe pokazują ramy podobieństwa genetycznego pomiędzy organizmami na różnych etapach ich ewolucji.

13. GENETYKA EWOLUCYJNA
©Kornelia Polok 70
Ćwiczenia i problemy
Zaprezentowane ćwiczenia mogą być przeprowadzone indywidualnie lub też w grupach 3-4 osób. Wówczas grupy mogą przedstawić efekty swojej pracy w postaci prezentacji, a pozostali studenci mogą dokonać oceny.
Ćwiczenie 1. Markery molekularne
Celem ćwiczenia jest podsumowanie danych o markerach molekularnych, ze szczególnym uwzględnieniem w badaniach ewolucyjnych. Można wykorzystać literaturę dostępną w naukowych bazach danych, a także liczne edukacyjne strony WWW.
PYTANIA I ZAGADNIENIA
Na czym polega metoda RFLP. Przedstaw schemat analizy przy pomocy RFLP (etapy) oraz podaj jej wykorzystanie w badaniach ewolucyjnych na wybranym przykładzie.
Na czym polega metoda RAPD. Czym różni się ona od reakcji PCR sekwencji unikalnej? Przedstaw schemat analizy przy pomocy RAPD. Podaj przykład wykorzystania w badaniach ewolucyjnych.
Na czym polega analiza sekwencji mikrosatelitarnych? Przedstaw schemat analizy oraz wykorzystanie na podstawie wybranego przykładu.
Na czym polega analiza AFLP. Przedstaw schemat analizy oraz wykorzystanie na wybranym przykładzie.
ZADANIA
1. Korzystając z Internetu lub dostępnych protokołów (Polok 2007) przeprowadź analizę kosztów dla metod AFLP, RAPD, SSR oraz analizy enzymatycznej uwzględniając analizę:
a) pojedynczej próbki b) 20 próbek c) 2 000 próbek.
2. Na podstawie charakterystyki poszczególnych typów markerów sporządź tabelę porównującą ich właściwości, w tym identyfikowane sekwencje, sposób detekcji, poziom polimorfizmu, typ dziedziczenia, ekspresję. Na podstawie sporządzonej tabeli podaj najbardziej efektywne możliwości zastosowania każdego typu markera w badaniach ewolucyjno-populacyjnych. Czy każdy marker nadaje się do każdej analizy?
3. Enzymy mogą występować w postaci cząsteczek zbudowanych z jednej podjednostki (monomer) lub większej liczby podjednostek, np. z dwóch (dimer), czterech (tetrametr). Obecność różnych form izoenzymatycznych powoduje, że dimery i tetramery mogą składać się z cząsteczek o identycznych właściwościach (homomery) lub różnych właściwościach (heteromery). Liczba możliwych homo i heteromerów zależy od liczby podjednostek. Z ilu i z jakich podjednostek składa się enzym określa się na podstawie ruchliwości na żelu oraz wzorów u homo i heterozygot. Wynika to z kodominacji. Przedstaw na schemacie wszystkie możliwe wzory, jakie można obserwować dla enzymu u homozygot i heterozygot w przypadku
a) monomeru, b) dimeru, c) tetrameru.
4. Przedstaw na schemacie wzory dla homozygot i heterozygot w przypadku następujących typów markerów DNA: RFLP, RAPD, SSR, AFLP.

13. GENETYKA EWOLUCYJNA
©Kornelia Polok 71
Ćwiczenie 2. Jak badamy procesy ewolucyjne?
Celem ćwiczenia jest odpowiedź na pytanie jaką rolę pełni teoria ewolucji w biologii oraz we współczesnym świecie. Ćwiczenie ma także na celu zapoznanie z metodologią projektowania badań ewolucyjnych, ich prowadzenia oraz wyciągania wniosków.
PYTANIA I ZAGADNIENIA
Jak teoria ewolucji jednoczy wszystkie nauki biologiczne.
Czy teoria ewolucji może mieć znaczenie w polityce?
Czy zmiany morfologiczne mają znaczenie w ewolucji? Uzasadnij odpowiedź.
Czy analizując filogenezę na podstawie dwóch różnych genów można uzyskać różne powiązania filogenetyczne? Uzasadnij odpowiedź.
ZADANIA
Na podstawie danych literaturowych zaproponuj doświadczenie, w którym wykorzystasz markery molekularne w badaniach nad ewolucją wybranej grupy organizmów. Określ cel główny doświadczenia, zadania badawcze z podaniem parametrów ilościowych, przedstaw schemat analizy (wykres) oraz metodykę, a także podaj oczekiwane rezultaty. Zastanów się nad możliwością publikacji oraz komercjalizacji uzyskanych wyników. 1. Wybierz dowolną grupę złożoną z kilku gatunków roślin (grzybów lub zwierząt) najlepiej należących do jednej
rodziny, plus jeden gatunek odrębny. Uzgodnij z prowadzącym.
2. Zapoznaj się z dostępną literaturą dotyczącą filogenezy wybranej grupy.
3. Przedstaw systematykę oraz powiązania filogenetyczne na podstawie dostępnej literatury w postaci tabelarycznej (systematyka) i jednej strony wstępu.
4. Określ nierozwiązane problemy w omawianej grupie i nakreśl cel badań.
5. Przygotuj program badań prowadzący do odpowiedzi na zadane pytania. Zaproponuj sposób poboru prób, techniki, które wykorzystasz oraz niezbędne parametry informujące o zmienności i podobieństwie genetycznym. Zaproponuj metody statystyczne.
6. Zaproponuj kosztorys i czasokres prowadzenia badań.
7. Przedyskutuj program badań z prowadzącym i grupą. W tym celu przygotuj 10 minutowe wystąpienie, które zaprezentujesz grupie.
8. Po zaakceptowaniu programu badań, na podstawie dostępnej literatury przedstaw wyniki, jakie mógłbyś uzyskać i omów je w postaci krótkiej dyskusji. Przedstaw swoją pracę grupie, która ją zrecenzuje podając jej mocne i słabe strony.

13. GENETYKA EWOLUCYJNA
©Kornelia Polok 72
Ćwiczenie 3. Molekularne podstawy ewolucji. Zmienność
PYTANIA I ZAGADNIENIA
1. Kreacjonizm odrzuca teorię ewolucji na podstawie braku ciągłości w śladach kopalnych. Koronnym argumentem jest Archeopteryx ponieważ ma pióra i lata, a więc był ptakiem, a nie formą pośrednią. Czy można uznać tę hipotezę? Uzasadnij odpowiedź.
2. Czy jest możliwe, że jakaś cecha wielokrotnie pojawia się w trakcie ewolucji? Uzasadnij odpowiedź na wybranym przykładzie.
3. Czy jest możliwy Świat według Gwiezdnych Wojen? Uzasadnij odpowiedź opierając się na danych z historii Życia na Ziemi. Jak można wyobrazić sobie taki Świat?
4. Czy należy obawiać się ocieplenia klimatu? Uzasadnij odpowiedź wykorzystując wiedzę dotyczącą
zlodowaceń i ociepleń klimatu w przeszłości.
ZADANIA
1. Każda era w historii Ziemi wytworzyła różnorodne organizmy przystosowane do środowiska. Opisz zróżnicowanie organizmów w wybranej erze.
2. Dla wybranej przez siebie grupy organizmów opisz rozmieszczenie geograficzne oraz zróżnicowanie morfologiczne populacji.
3. Wybierz gatunek zagrożony i przedstaw plan jego ochrony. Jakie czynniki ograniczają rozmieszczenie gatunku?
4. Określ nierozwiązane problemy w omawianej grupie i nakreśl cel badań.
Ćwiczenie 4. Ewolucja wybranych taksonów. Ewolucja a cechy ilościowe
PYTANIA I ZAGADNIENIA
1. Hodowcy wykorzystują selekcję sztuczną do zmiany wielu cech u zwierząt i roślin. Czy jest możliwe uzyskanie np. Drosophila melanogaster wielkości trzmiela? Uzasadnij odpowiedź.
2. Czy możesz wymienić na podstawie danych literaturowych (Internet) mutacje, które miały największe znaczenie w rozwoju ludzkości (ewolucji biologicznej, społecznej itp.). Nie muszą to być koniecznie mutacje u człowieka. Uwzględnij także rośliny i zwierzęta.
3. Czy udomowienie jest przykładem ewolucji? Odpowiedź przedstaw na przykładzie wybranych roślin uprawnych.
4. Niektóre QTL zostały zidentyfikowane i zsekwencjonowane. Czy wiedza ta przyczyniła się do lepszego zrozumienia ewolucji?
ZADANIA
1. W ostatnich latach śledzimy rozprzestrzenianie się różnych chorób, co przybiera rozmiary pandemii. Jak rozwój pandemii można wytłumaczyć w świetle ewolucji? Jak zapobiegać rozwojowi pandemii? Zagadnienie omów na przykładzie wybranej choroby, np. grypy, w tym A/H1N1; AIDS (HIV); SARS; gruźlicy.
a) Przygotuj krótką prezentację na wybrany przez Ciebie temat, omawiający czynniki wywołujące chorobę, rozprzestrzenianie się (epidemiologia), ewolucję wirusa/bakterii, sposoby leczenia i zapobiegania.
b) Wyobraź sobie, że jesteś pracownikiem firmy medycznej mającej opracować lek/szczepionkę przeciwko wybranej chorobie. Napisz plan działań.
c) Wyobraź sobie, że jesteś Ministrem Zdrowia. Przygotuj plan działań na wypadek pandemii wybranej przez Ciebie choroby.

13. GENETYKA EWOLUCYJNA
©Kornelia Polok 73
2. Selekcja sztuczna jest podstawą hodowli. Wybierz gatunek udomowiony przez człowieka i w postaci krótkiej publikacji przeglądowej opisz, na czym polegało jego udomowienie.
3. Jak potraktować modyfikacje przy pomocy inżynierii genetycznej w świetle ewolucji. Opracuj zagadnienie wykorzystując modyfikację wybraną na podstawie danych literaturowych. Podaj istotę modyfikacji, wywołane zmiany i symulację ewentualnych scenariuszy ewolucyjnych.
Ćwiczenie 5. Ewolucja genów i genomów. Gatunek biologiczny
PYTANIA I ZAGADNIENIA
1. Zastanów się nad definicją gatunku na podstawie danych w Internecie. Jak sądzisz, która z definicji najpełniej oddaje sytuację wytworzoną w drodze ewolucji biologicznej? Uzasadnij ilustrując przykładami.
2. W badaniach filogenetycznych mutacja punktowa jest często traktowana jako wyznacznik powstania nowej linii czy gatunku. Kiedy zamiana pojedynczego nukleotydu może prowadzić do powstania nowej cechy?
3. Systematyka wyróżnia gatunki na podstawie cech morfologicznych analizowanych u pojedynczych osobników. O zaklasyfikowaniu osobnika jako nowego gatunku niejednokrotnie decyduje pojedyncza cecha. Jak myślisz ile jest genów „specjacji”? (Ile genów różni gatunki?).
4. Znaczną część genomu stanowią sekwencje niekodujące, np. transpozony lub inne sekwencje powtarzalne. Czy są one jedynie „śmietnikiem ewolucji” czy może odgrywają istotną rolę w procesach ewolucyjnych? Uzasadnij odpowiedź na przykładach.
5. Rośliny transgeniczne mają wprowadzone geny od odległych gatunków, często są to geny bakteryjne, jak gen Bt u kukurydzy, który uodparnia ją na działanie herbicydów? Czy taka roślina mogłaby powstać bez udziału inżynierii genetycznej, a nawet bez udziału człowieka? Jakie są możliwości przekraczania barier gatunkowych przez geny? Podaj przykłady.
ZADANIA
1. Jednym z najważniejszych źródeł informacji są bazy danych ze zdeponowanymi sekwencjami nukleotydowymi.
a) Krótko scharakteryzuj bazę NCBI pod kątem jej zawartości i przydatności w analizach ewolucyjnych. b) Opisz dostępny w bazie software ze szczególnym uwzględnieniem grupy programów BLAST c) Opisz strukturę pliku przedstawiającego sekwencję DNA. Podaj różne formaty. d) Podaj zastosowanie CLUSTALX (można wykorzystać prace magisterskie i stronę http://www.clustal.org)
oraz COBALT (NCBI).
2. Duplikacja jest ważnym procesem prowadzącym do powstawania nowych genów, a tym samym ich ewolucji. Omów wybrany przykład ewolucji genu poprzez duplikację. Zastanów się nad czynnikami, które przyczyniają się do uzyskania nowej funkcji przez zduplikowany gen.
3. Wybierz dowolny gen i w bazie NCBI wyszukaj jego sekwencję nukleotydową. Wykorzystując odpowiedni program z grupy BLAST wyszukaj:
a) Podobne sekwencje nukleotydowe. Ile jest takich sekwencji, które wykazują największe, a które najmniejsze podobieństwo?
b) Z uzyskanej listy wybierz sekwencje dla 10-15 gatunków w miarę blisko spokrewnionych oraz dla jednego gatunku bardziej odległego. Zaznacz interesujące sekwencje na liście sumarycznej i wyświetl w postaci formatu FASTA. Wybierz opcję, zapisz jako tekst, skopiuj wszystkie sekwencje do pliku w notatniku i zapisz na dysku jako plik nukleotydy.txt Sekwencje powinny być mniej więcej jednakowej długości. Najlepiej wybierać pomiędzy 1000-2000 bp. Sekwencje dla wszystkich gatunków mają być w jednym pliku.

13. GENETYKA EWOLUCYJNA
©Kornelia Polok 74
UWAGA! Unikaj bardzo długich sekwencji, które najczęściej są sekwencją całych chromosomów lub znacznych fragmentów genomu w klonach BAC
c) Dla wyjściowej sekwencji nukleotydowej pozyskaj sekwencję białkową. Za pomocą uzyskanej sekwencji białkowej oraz programu BLAST dokonaj przeszukiwania bazy NCBI w celu znalezienia podobnych sekwencji białkowych. Przedstaw domeny konserwatywne i strukturę przestrzenną białka (program Cn3D).
d) Z uzyskanej listy sekwencji białkowych pozyskaj sekwencje dla tych samych gatunków, dla których pozyskano sekwencje nukleotydowe. Uzyskane sekwencje białkowe zapisz w formacie FASTA i przenieść do notatnika podobnie jak w punkcie b. Zapisz plik jako protein.txt
e) Dokonaj analizy zbieżności w programie CLUSTAL i przedstaw drzewo no podstawie sekwencji DNA, a następnie na podstawie sekwencji białkowej. Porównaj uzyskane drzewa ze znaną ewolucją analizowanych przez Ciebie gatunków.
f) Przeprowadź podobną analizę w programie COBALT i porównaj wyniki z programem CLUSTALX.





















![Genetyka Zwierząt - 2004 - Krystyna M. Charon [PL]](https://img.dokumen.tips/doc/110x75/5571f26e49795947648c8e67/genetyka-zwierzat-2004-krystyna-m-charon-pl.jpg)








