Embed Size (px)
Citation preview

1
Modulating ER-Golgi cargo receptors for improving secretion of carrier-fused 1
heterologous protein in the filamentous fungus Aspergillus oryzae 2
3
Running title 4
ER-Golgi cargo receptor and heterologous proteins 5
6
Authors 7
Huy-Dung Hoang, Jun-ichi Maruyama and Katsuhiko Kitamoto* 8
9
Department of Biotechnology, The University of Tokyo, Tokyo, Japan. 10
11
*Corresponding author 12
Mailing address: Department of Biotechnology, The University of Tokyo, 1-1-1 Yayoi, Bunkyo-13
ku, Tokyo, Japan. 14
Tel: +81-3-5841-5161 15
Fax: +81-3-5841-8033 16
Email: [email protected] 17
AEM Accepts, published online ahead of print on 31 October 2014Appl. Environ. Microbiol. doi:10.1128/AEM.02133-14Copyright © 2014, American Society for Microbiology. All Rights Reserved.
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

2
Abstract 18
19
Filamentous fungi are excellent hosts for industrial protein production due to their superior 20
secretory capacity; however, the yield of heterologous eukaryotic proteins is generally lower 21
compared to that of fungal or endogenous proteins. Although activating protein folding 22
machinery in the ER improves the yield, the importance of intracellular transport machinery for 23
heterologous protein secretion is poorly understood. Here, using Aspergillus oryzae as a model 24
filamentous fungus, we studied the involvement of two putative lectin-like cargo receptors, 25
AoVip36 and AoEmp47, in secretion of heterologous protein expressed in fusion with the 26
endogenous enzyme α-amylase as carrier. Fluorescence microscopy revealed that mDsRed-27
tagged AoVip36 localized in the Golgi, whereas AoEmp47 showed localization in both the ER 28
and Golgi. Deletion of AoVip36 and AoEmp47 improved heterologous protein secretion, but 29
only AoVip36 deletion had a negative effect on secretion of α-amylase. Analysis of ER-enriched 30
cell fractions revealed that AoVip36 and AoEmp47 were involved in the retention of 31
heterologous proteins in the ER. However, the overexpression of each cargo receptor had 32
different effects on heterologous protein secretion: AoVip36 enhanced the secretion, whereas 33
AoEmp47 promoted the intracellular retention. Taken together, our data suggest that AoVip36 34
and AoEmp47 hinder the secretion of heterologous proteins by promoting their retention in the 35
ER, but that AoVip36 also promotes the secretion of heterologous proteins. Moreover, we found 36
that genetic deletion of these putative ER-Golgi cargo receptors significantly improves 37
heterologous protein production. The present study is the first to propose that ER-Golgi transport 38
is a bottleneck for heterologous protein production in filamentous fungi. 39
40
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

3
Introduction 41
42
Filamentous fungi possess a prominent secretory capacity and thus are excellent hosts for 43
recombinant protein production. Numerous approaches and attempts have been made to produce 44
industrially valuable proteins in filamentous fungi, such as Aspergillus and Trichoderma (1). 45
However, higher eukaryotic proteins are generally inefficiently produced and secreted in these 46
fungi compared to fungal or endogenous proteins. Several bottlenecks in the heterologous 47
protein production process have been identified to date and a few limiting factors have been 48
overcome by genetically modifying the expression host. In particular, reducing protease activity 49
is necessary to limit the degradation of heterologous proteins, as was demonstrated by the three-50
fold increase in the level of heterologous proteins in the culture supernatant of an Aspergillus 51
oryzae strain with ten successively deleted protease genes (2). Heterologous protein production 52
by A. oryzae was also effectively improved by the repression of vacuolar protein sorting and 53
autophagy (3, 4). The genetic fusion of a target protein with an endogenous protein carrier is a 54
commonly used strategy to increase heterologous protein yields in filamentous fungi. Effective 55
carrier proteins are abundantly secreted enzymes, such as glucoamylase, α-amylase, and 56
cellobiohydrolase (5–7), and are thought to allow the heterologous fusion protein to overcome 57
bottlenecks in transcription and post-transcriptional processes (8). Despite the demonstrated 58
efficacy of these approaches, heterologous protein secretion in recombinant strains remains far 59
below that of endogenous proteins, indicating that other bottlenecks for heterologous protein 60
production and secretion must be overcome before filamentous fungi can reach maximal 61
production capacity. 62
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

4
One approach for increasing the expression of heterologous proteins is the use of a strong 63
promoter; however, increased target protein synthesis may lead to overloading of the folding 64
capacity of the ER. In Aspergilli, expression of human protein tissue plasminogen activator or a 65
fused form of α-amylase-prochymosin triggered the unfolded protein response (UPR) due to ER 66
stress (9, 10). Overexpression of molecular chaperones such as BiP and protein disulfide 67
isomerase to assist protein folding improves the yield of heterologous proteins (11, 12), a 68
response that may be attributable to alleviation of ER stress resulting from increased folding 69
activity. When the UPR is triggered, the expression of a set of UPR-associated proteins, 70
including molecular chaperones, vesicular traffic components, and ER-associated degradation 71
(ERAD) proteins, is up-regulated in an attempt to resolve the ER stress by increasing the folding, 72
transport, and degradation of proteins (13). Constitutive up-regulation of UPR by expression of 73
the activated form of the transcription factor HacA was shown to improve the production of 74
bovine chymosin in Aspergillus niger (14) and the plant taste-modifying protein neoculin in A. 75
oryzae (7). Although these observations suggest that ER stress is a major hindrance for 76
heterologous protein production in filamentous fungi, the underlying cause of the ER stress 77
associated with heterologous protein expression remains unclear. 78
One possible cause of ER stress is the inefficient transport of heterologous proteins between 79
the ER and Golgi. Cargo proteins are transported between the ER and Golgi by vesicular 80
trafficking. Coat protein complex II (COPII)-coated vesicles transport secretory proteins in the 81
anterograde direction, whereas COPI-coated vesicles carry proteins in the retrograde direction, 82
from the Golgi back to the ER (15). During the budding of COPII vesicles, a class of membrane 83
proteins called ER-Golgi cargo receptors is responsible for recruiting cargo proteins into the 84
vesicles, thereby promoting their transport (16). The binding of cargo receptors to target proteins 85
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

5
is highly specific, as demonstrated by the defective secretion of a specific set of proteins upon 86
deletion of a single cargo receptor. For example, the mammalian lectin-type cargo receptor 87
ERGIC-53 selectively exports a subset of glycosylated proteins (17). Previous studies have also 88
found that another lectin-type cargo receptor, VIP36, promotes secretion of the glycoproteins rat 89
parotid gland α-amylase and clusterin (18). VIP36 has also been shown to recycle human α1-90
antitrypsin from the Golgi back to the ER, and upon silencing of VIP36, α1-antitrypsin secretion 91
is increased (19), demonstrating that this cargo receptor has a protein retention function. As 92
VIP36 forms a stable complex with the molecular chaperone BiP (20), it appears to be involved 93
in the quality control of secretory proteins. Because glycoproteins such as α-amylase are 94
commonly used as carrier proteins, lectin-type cargo receptors might affect the secretion of 95
carrier-fused heterologous proteins and are therefore potentially good models for studying the 96
interaction between cargo receptors and heterologous proteins in A. oryzae. This study aimed to 97
investigate the involvement of lectin-type cargo receptors in the intracellular trafficking of 98
carrier-fused heterologous proteins in the filamentous fungus A. oryzae. 99
100
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

6
MATERIALS AND METHODS 101
102
Strains, media and growth conditions. A. oryzae strain RIB40 (21) was used as a DNA 103
donor. Escherichia coli DH5α was used as a host for DNA manipulation. A. oryzae strains 104
NSlD1 (niaD- sC
- ΔligD) and NSPlD1 (niaD
- sC
- ΔpyrG ΔligD) (22) were used as hosts for gene 105
expression and deletion, respectively. A. oryzae strain NS-CaG1 (niaD- sC
- adeA
- AoclxA-106
egfp::adeA) expressing the ER marker calnexin (23) fused to EGFP was used as a host for 107
transformation in the microscopic observation experiments. All strains were maintained on PD 108
agar medium (Nissui Pharmaceutical, Tokyo, Japan). M medium (2% glucose, 0.2% NH4Cl, 109
0.1% (NH4)2SO4, 0.05% KCl, 0.05% NaCl, 0.1% KH2PO4, 0.05% MgSO4·7H2O, and 0.002% 110
FeSO4·7H2O [pH 5.5]) supplemented with 0.15% methionine (M+Met) was used as selective 111
medium for pyrG+ niaD
- sC
- strains used for gene deletion. CD medium (2% glucose, 0.3% 112
NaNO3, 0.2% KCl, 0.1% KH2PO4, 0.05% MgSO4·7H2O, and 0.002% FeSO4·7H2O [pH 5.5]) 113
supplemented with 0.0015% methionine (CD+Met) was used as selective medium for niaD+ sC
- 114
strains and as a growth medium for microscopic observation experiments. 5×DPY medium (10% 115
dextrin, 5% polypeptone, 2.5% yeast extract, 0.5% KH2PO4, and 0.05% MgSO4·7H2O [pH 5.5]) 116
was used as general growth medium unless otherwise stated. 117
118
Generation of a putative cargo receptor-deletion mutant. For generation of a plasmid for 119
the deletion of Aovip36, the 1.5-kb upstream and downstream regions of Aovip36 120
(AO090026000428) were amplified using the primer pairs X-uVIP36-F/X-uVIP36-R and X-121
dVIP36-F/X-dVIP36-R, respectively. The amplified fragments were cloned into the entry vector 122
pDONR-P4-P1R and PDONR-p2R-p3 using the BP reaction (MultiSite Gateway Cloning Kit; 123
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

7
Life Technologies, Carlsbad, CA) to generate the plasmids p5’uVp and p3’dVp, respectively. 124
Using a similar method, the plasmids p5’uEp and p3’dEp, containing the 1.5-kb upstream and 125
downstream regions of Aoemp47 (AO090102000145), respectively, were constructed using the 126
primer sets X-uEp-F/X-uEp-R and X-dEp-F/X-dEp-R, respectively. The two fragments were 127
then connected with upstream and downstream ends of the selective marker pyrG from the 128
plasmid pgEpG using the LR reaction of MultiSite Gateway cloning kit. Linear deletion 129
fragments were then amplified by PCR with the primer sets X-uVIP36-F/X-dVIP36-R for 130
Aovip36 and X-uEp-F/X-dEp-R for Aoemp47. The amplified products were then purified and 131
introduced into A. oryzae strain NSPlD1 using a previously described transformation method 132
(24), and deletion strains were selected on M+Met agar medium. Deletion of cargo receptor 133
genes by the pyrG marker was confirmed by Southern analysis using the upstream region of each 134
gene as probes. The sequences of all primers used in this study are listed in Table S1 in the 135
supplemental material. 136
137
Construction of expression plasmids. For the generation of an expression plasmid for N-138
terminally mDsRed-tagged AoVip36, the signal sequence of AoVip36, mDsRed without a stop 139
codon, and the remaining coding sequence of Aovip36 were amplified using the primer sets 140
attB1-Vp-F/ssVp-mDsRed-R, ssVp-mDsRed-F/mDsRed-Vp-R, and mDsRed-Vp-F/AttB2-Vp-R, 141
respectively. Fusion PCR with the primer pair attB1-Vp-F and attB2-Vp-R was used to insert 142
mDsRed between the signal sequence and the remaining Aovip36 coding sequence. Similarly, the 143
signal sequence of AoEmp47, mDsRed without a stop codon, and the remaining coding sequence 144
of Aoemp47 were amplified using the primer sets attB1-Ep-F/ssEp-mDsRed-R, ssEp-mDsRed-145
F/mDsRed-Ep-R, and mDsRed-Ep-F/AttB2-Ep-R, respectively. Fusion PCR with the primer pair 146
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

8
attB1-Ep-F and attB2-Ep-R was used to insert the mDsRed sequence between the Aoemp47 147
signal sequence and the remaining coding sequence. The fusion products for AoVip36 and 148
AoEmp47 were then cloned into center entry vector pDONR-P221 using the BP clonase reaction 149
to generate pCRVp and pCREp, respectively. The LR clonase reaction with the 5’ entry clone 150
pg5’PaB, containing the amyB promoter, and the 3’ entry clone pg3’TaN, containing the amyB 151
terminator and niaD marker, was then performed to generate a plasmid expressing mDsRed 152
fused versions of AoVip36 (pEpARVpN) and AoEmp47 (pEpAREpN) under control the of 153
amyB promoter. 154
To express heterologous proteins, plasmids for expressing AmyB-fused prochymosin 155
(proCHY) (pgAKCN) (25) or EGFP (pNamyBEGFP) (23) were used for transformation. 156
Integration of a single copy of the plasmids into the niaD locus was confirmed by Southern 157
analysis. Two to three transformants were selected and expression of heterologous protein was 158
confirmed by enzyme activity (proCHY) or immunoblot (AmyB-EGFP). There were no 159
differences in the secretion levels, one transformants was selected for further analysis. 160
For overexpressing mDsRed-AoVip36 and mDsRed-AoEmp47, LR clonase reactions of 161
pCRVp or pCREp with pg5’PaB and pg3’TaSO, containing the amyB terminator and A. oryzae 162
sC marker, were performed to generate an overexpression plasmid. The plasmids were 163
transformed into the strain SlD-aG2 expressing AmyB-EGFP to ensure similar levels of AmyB-164
EGFP expression in the resulted transformants. Expression of mDsRed-tagged cargo receptors in 165
the transformants were confirmed by fluorescence microscopy. There were no differences in the 166
fluorescence pattern of both mDsRed-tagged cargo receptor and AmyB-EGFP between two 167
independent transformants for each construct, and therefore one transformant was selected for 168
further analysis. 169
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

9
170
Construction of a strain endogenously expressing AoGrh1-EGFP. For inserting EGFP at 171
the C-terminus of AoGrh1 (AO090001000523), a 1.5-kb region containing the 0.5-kb 5’ 172
upstream region and entire open reading frame (ORF) of Aogrh1 without a stop codon was PCR 173
amplified with the primers aB4-uGr1-F and aB1r-uGr1-R, and was then cloned into a 5’ entry 174
vector to generate the plasmid p5’uGr1. A 1.5-kb downstream region of Aogrh1 was amplified 175
by PCR with primers aB2r-dGr1-F and aB3-dGr1-R and then cloned into a 3’ entry vector to 176
generate the plasmid p3’dGr1. The two entry clones were subjected to the LR clonase reaction 177
with the center entry clone pgEGFPadeA, which contained the egfp gene and amyB terminator, 178
and destination vector pDEST R4-R3, generating the plasmid pgGr1GA. The insertion cassette 179
for expressing the AoGrh1-EGFP fusion protein from the Aogrh1 gene locus was amplified by 180
PCR using primers aB4-uGr1-F and aB3-dGr1-R from the plasmid pgGr1GA, and was then 181
introduced into strain NSR-ΔlD-2. Insertion of the egfp gene at the 3’ end of the Aogrh1 ORF 182
was confirmed by colony PCR with the primer pair aB4-uGr1-F and aB3-dGr1-R (data not 183
shown). 184
185
Milk-clotting assay for quantification of secreted chymosin. Milk-clotting assay for 186
chymosin activity was performed according to a previously reported method (3). Briefly, 100 µl 187
of sample was mixed with 900 μl of a skim milk solution (12% skim milk in 10 mM CaCl2) in a 188
2-ml eppendorf tube, which was then incubated at 30°C with constant shaking at 50 rpm. The 189
clotting point was defined at the time the solution stopped visibly moving due to formation of a 190
milk clot. The clotting times were recorded and compared with a standard solution prepared with 191
rennin (Sigma-Aldrich, St. Louis, MO) to determine the chymosin concentration in the test 192
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

10
samples. The total amount of proteins in the culture supernatant was measured with a Protein 193
Assay kit (Bio-Rad, Hercules, CA) according to the manufacturer’s instructions. 194
195
Fluorescence microscopy observation. For fluorescence microscopy, 105 conidia were 196
inoculated into 100 μl M+Met liquid medium in 35-mm glass-based dishes (Asahi Techno Glass, 197
Chiba, Japan) and incubated for 18 h at 30°C. Hyphae were then observed with an IX71 inverted 198
microscope (Olympus, Tokyo, Japan) equipped with a 100×neofluor objective lens (1.4 199
numerical aperture). GFP and mDsRed excitations were performed with 488 nm (Furukawa 200
Electric, Tokyo, Japan) and 561 nm (Melles Griot, Carlsbad, CA) semiconductor lasers, 201
respectively, and emitted light was filtered with GFP, DsRed, and DualView filters (Nippon 202
Roper, Chiba, Japan). The CSU22 confocal scanning system (Yokogawa Electronic, Tokyo, 203
Japan) equipped with an Andor iXon cooled digital charge-coupled device camera (Andor 204
Technology PLC, Belfast, UK) and Andor iQ Software (Andor Technology) was used for 205
acquiring and analyzing images. 206
For quantifying colocalization, mean fraction of mDsRed-tagged cargo receptors overlapping 207
the ER or Golgi in the three independent images was calculated using Mander’s coefficient (MC) 208
(26) and the ImageJ (National Institute of Health, Bethesda, MD) plug-in JaCoP. The MC value 209
of 1 means that the two fluorescence signals completely overlap with each other while a MC 210
value of 0 means that there is no overlap between the two fluorescence signals. 211
Localization analysis of AmyB-EGFP in the strains overexpressing mDsRed-tagged AoVip36 212
and AoEmp47 was performed by observing at least 20 independent hyphae. All the hyphae 213
showed similar localization patterns of AmyB-EGFP and mDsRed-tagged cargo receptors, and 214
the representative images were captured. 215
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

11
216
SDS-PAGE and immunoblotting. For SDS-PAGE, protein samples were mixed with 1/5 217
volume 5×Laemli Sample buffer and were then boiled in a water bath for 5 min and immediately 218
placed on ice for 5 min. Samples and molecular markers were separated on a 10% 219
polyacrylamide gel. For immunoblotting, proteins in the gel were transferred onto Immobilon-P 220
PVDF membranes (Millipore, Bedford, MA) using a semi-dry blotting system (Nihon Eido, 221
Tokyo, Japan). For detecting chymosin, a polyclonal rabbit anti-chymosin antibody (Nordic 222
Immunological Laboratories, Tilburg, Netherlands) was used. For detecting EGFP, Living 223
Colors® A.V. Monoclonal Antibody (Clontech, Palo Alto, CA) was used. Chemiluminescence 224
was detected using an ECL Advance Western blotting Detection Kit (GE Healthcare, 225
Buckinghamshire, UK) and images were obtained with an LAS-4000 Mini System (GE 226
Healthcare). Quantification of immunoblotting analysis was performed using ImageJ software 227
(National Institute of Health, Bethesda, MD). The band intensities in the same gel were 228
compared between the control and deletion/overexpression strains, and the relative values were 229
calculated by normalizing that of the control strain as 100%. 230
231
Cell disruption, ER enrichment, and immunoblotting. Conidia were inoculated into 20 ml 232
5×DPY liquid medium and cultivated at 30°C with shaking at 150 rpm. Mycelia were collected 233
and disrupted using metal beads in a Multi-Bead Shocker (Yasui Kikai, Osaka, Japan). Disrupted 234
mycelia were homogenized in extraction buffer (250 mM sucrose, 50 mM Tris, and 1 mM 235
PMSF) supplemented with 1% Protease Inhibitor Cocktail (Sigma) and briefly centrifuged at 236
500×g for 5 min to remove unbroken cells and cell debris. For enrichment of ER contents, the 237
obtained supernatant was further centrifuged at 20,000×g for 30 min, and both the resulting 238
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

12
pellet and supernatant were collected for subsequent analysis. Protein concentration of samples 239
was determined using a Protein Assay kit (Bio-Rad). Cell lysate samples containing 2 µg protein 240
were subjected to SDS-PAGE and immunoblotting, as described above. 241
242
α-Amylase activity assay. α-Amylase in the culture supernatant was quantified using an α-243
Amylase Quantification Kit (Kikkoman, Tokyo, Japan), as instructed by the manufacturer. All 244
culture supernatant samples were diluted 1/50 before quantification. 245
246
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

13
RESULTS 247
248
Subcellular localization of putative lectin-type cargo receptors in A. oryzae. A search for 249
homologs of VIP36 in two A. oryzae genome databases, the Comprehensive A. oryzae Genome 250
Database (CAoGD; http://nribf21.nrib.go.jp/CFGD/) and Database of the Genome Analyzed at 251
NITE (DOGAN; http://www.bio.nite.go.jp/dogan/project/view/AO), resulted in the identification 252
of two entries with high sequence similarity to VIP36: AO090026000428 and AO090102000145. 253
The predicted product of the AO090026000428 gene shared high similarity to human VIP36 and 254
was therefore named AoVip36. AoVip36 had a domain organization similar to that of human 255
VIP36, consisting of a signal sequence, lectin-like domain, and a predicted transmembrane 256
region near the C-terminus (Fig. 1A). AoVip36 showed 27% overall identity with human VIP36, 257
with the lectin-like domain showing 34% identity with the corresponding domain of human 258
VIP36. 259
The predicted AO090102000145 gene product had higher amino acid sequence similarity to 260
the Saccharomyces cerevisiae cargo receptor Emp47p than human VIP36, and was therefore 261
named AoEmp47. The domain organization of AoEmp47 was also similar to that of S. cerevisiae 262
Emp47p and consisted of a signal sequence, a lectin-like domain followed by a coiled-coil 263
domain, and a predicted transmembrane region located near the C-terminus (Fig. 1B). The 264
coiled-coil domain of S. cerevisiae Emp47p was reported to promote self-oligomerization, which 265
is essential for the exit from the ER (27). AoEmp47 showed 16% overall identity with S. 266
cerevisiae Emp47p, and the lectin-like domain shared 18% identity to that of S. cerevisiae 267
Emp47p. Moreover, a tyrosine containing motif (YxxΦ) and a dilysine motif were also found at 268
the C-terminus of AoEmp47 (Fig. 1C). Previous studies in S. cerevisiae Emp47p found that the 269
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

14
tyrosine-containing motif is important for Golgi localization and interaction with COPI and 270
COPII components, while the dilysine motif is required for ER recycling (28, 29). 271
To determine the subcellular localization of AoVip36 and AoEmp47, mDsRed-tagged 272
versions of the two putative cargo receptors were expressed in A. oryzae under control of the 273
amyB promoter. In the fusion construct, mDsRed was inserted after the N-terminal signal 274
sequence of the receptor (Fig. S1A in the supplemental material). The ER membrane protein 275
AoClxA fused to EGFP was used as an ER marker (23), and an A. oryzae homolog 276
(AO090001000523) of Aspergillus nidulans GrhA, called AoGrh1, was used as a Golgi marker. 277
GrhA is a homolog of the human Golgi-associated protein GRASP65 and localizes to cis-Golgi 278
in A. nidulans (30). EGFP was inserted at the C-terminal region of the Aogrh1 gene locus, 279
allowing for the resulting AoGrh1-EGFP fusion protein to be expressed under control of the 280
native Aogrh1 promoter. 281
Fluorescent microscopy analysis revealed that mDsRed-AoVip36 localized to punctuate 282
structures labeled by AoGrh1-EGFP, but did not clearly colocalize with the ER marker AoClxA-283
EGFP (Fig. 2A). Quantification analysis of colocalization indicated that a higher fraction of 284
mDsRed-AoVip36 overlapped with the Golgi marker than the ER (Fig. 2C). These results 285
supported that AoVip36 predominantly localizes to the Golgi. This finding is consistent with the 286
previously reported localization of mammalian VIP36 in MDCK (31) and HeLa cells (19). In 287
contrast, mDsRed-AoEmp47 localized to tubular structures labeled by AoClxA-EGFP and to 288
punctuate structures labeled by AoGrh1-EGFP (Fig. 2B). Quantification analysis of 289
colocalization suggested that a high fraction of mDsRed-AoEmp47 overlapped with both the ER 290
and Golgi markers (Fig. 2C). It is noteworthy to mention that when co-expressed with AoClxA-291
EGFP, the localization shifted predominantly to the ER, which might be due to an unintentional 292
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

15
interaction between AoClxA-EGFP and mDsRed-AoEmp47. These data suggested that 293
AoEmp47 might cycle between the ER and Golgi. This pattern slightly differed from that of S. 294
cerevisiae Emp47p, which was shown to localize to Golgi localization (29), which is discussed 295
later (See Discussion). 296
297
Effects of AoVip36 and AoEmp47 deletion on endogenous α-amylase secretion. To 298
investigate the function of AoVip36 and AoEmp47, the genes encoding these putative receptors 299
were deleted (Fig. S1B and C in the supplemental material) and α-amylase activities in the 300
culture supernatant of the deletion strains were measured. The analysis revealed that the deletion 301
of Aovip36 reduced the α-amylase activity by approximately 30% compared with the control 302
strain, whereas the deletion of AoEmp47 did not affect the α-amylase activity (Fig. 3A). Deletion 303
of each putative cargo receptor did not affect the growth and the total amount of proteins in the 304
culture supernatant (Fig. 3B and C). SDS-PAGE/Coomassie staining analysis showed that most 305
of protein band patterns from the culture supernatant did not significantly differ among the 306
control and deletion strains (Fig. 3D). These data suggested that the decrease in α-amylase 307
activity of the ΔAovip36 strain was not caused by the defects in growth, expression of secreted 308
proteins and extracellular protein stability. The α-amylase activity is specifically correlated with 309
a protein band of approximately 50 kDa in SDS-PAGE/Coomassie staining analysis as evidenced 310
by the fact that deletion of all three A. oryzae α-amylase genes (amyA, amyB and amyC) 311
encoding the identical amino acid sequences except for two amino acids led to a complete loss of 312
both the activity and the protein band (32). The intensity of the protein band corresponding to α-313
amylase was reduced in the ΔAovip36 strain (Fig. 3D). Taken together, these results suggested 314
that AoVip36 was involved in the secretion of α-amylase in A. oryzae. 315
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

16
316
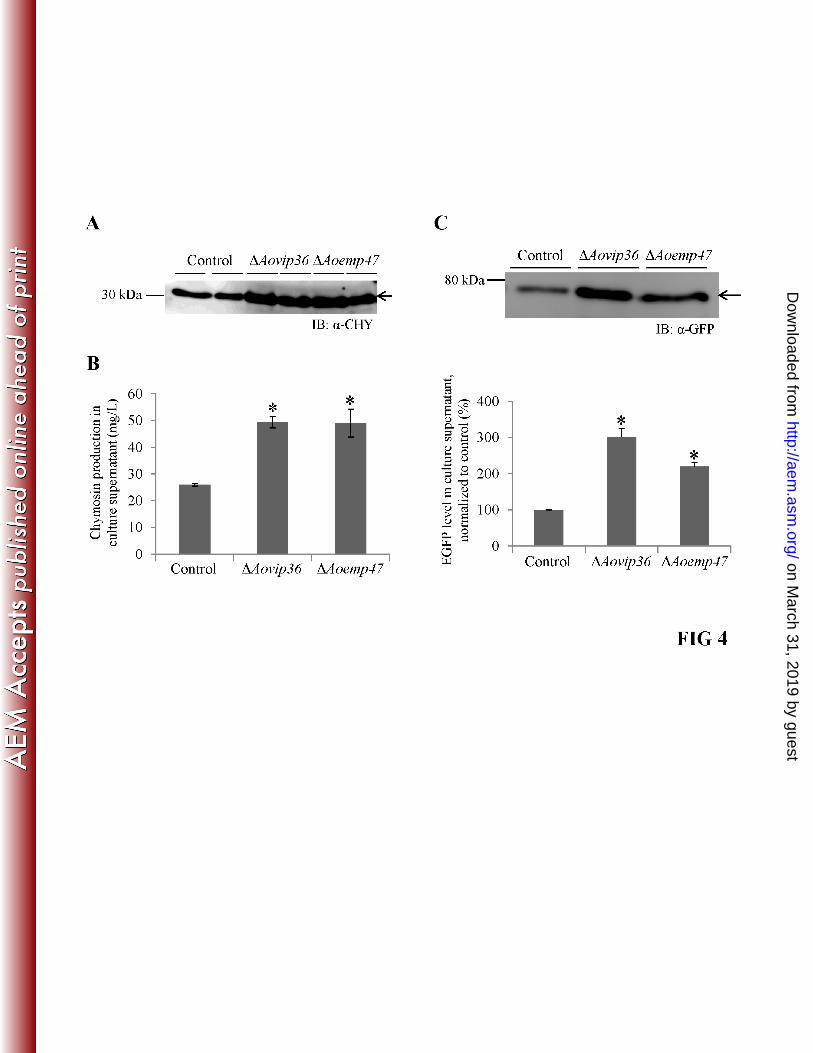
Effects of AoVip36 and AoEmp47 deletion on carrier-fused heterologous protein secretion. 317
To determine if deletion of the putative cargo receptors affected carrier-fused heterologous 318
protein secretion in A. oryzae, the α-amylase fused form of bovine prochymosin (AmyB-319
proCHY) was used as a carrier-fused heterologous protein model to investigate secretory protein 320
trafficking in the ΔAovip36 and ΔAoemp47 strains. The AmyB-proCHY fusion protein is thought 321
to be cleaved at a Kex2-like protease cleavage site in the Golgi, and the released prochymosin 322
autocatalytically converts into mature CHY after further cleavage of the prosequence (33). We 323
first evaluated the secretion of chymosin on the second day of cultivation to minimize the effects 324
of protease degradation. Immunoblotting analysis of the culture supernatant of the ΔAovip36 and 325
ΔAoemp47 strains showed that the band corresponding to prochymosin was increased compared 326
to the control (Fig. 4A). Notably, the size of the detected band corresponded to that of 327
prochymosin after cleavage from AmyB, suggesting that the secreted protein had passed through 328
the complete secretory pathway and did not enter the supernatant as a result of an unconventional 329
secretion process or leakage due to cell lysis. The enzymatic activity of chymosin was quantified 330
on the fourth day of cultivation, because the majority of prochymosin is converted into active 331
chymosin by this day (3, 10). An assay for milk clotting activity in the culture supernatant 332
revealed that deletion of AoVip36 or AoEmp47 increased the amount of mature CHY in the 333
culture medium by approximately two folds compared to the control cells (Fig. 4B). 334
To confirm that deletion of the putative cargo receptors improved carrier-fused heterologous 335
protein secretion, we examined the effect of their deletion on α-amylase-fused EGFP (AmyB-336
EGFP). Immunoblotting analysis of the culture supernatant after two days of cultivation 337
demonstrated that a band corresponding to the size of AmyB-EGFP fusion protein 338
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

17
(approximately 73 kDa) was present. Quantitative analysis of the band intensity indicated that a 339
larger amount of AmyB-EGFP was secreted into the culture medium of the ΔAovip36 and 340
ΔAoemp47 strains compared to the control strain, similar to the case of AmyB-proCHY (Fig. 4C). 341
These results indicated that the putative cargo receptors negatively affected carrier-fused 342
heterologous protein secretion. 343
344
Effects of AoVip36 and AoEmp47 deletion on ER retention of carrier-fused heterologous 345
proteins. As AoVip36 and AoEmp47 were found to localize to the ER and Golgi, we reasoned 346
that they might interfere with the normal intracellular trafficking of carrier-fused heterologous 347
proteins. Thus, we examined and compared the retention of fusion proteins in the ER of the wild-348
type, ΔAovip36, and ΔAoemp47 strains. ER-enriched fractions were obtained by centrifuging 349
mycelial extracts at 20,000×g and collecting the pellet. To test the efficiency of this method, the 350
ER-enriched pellet and supernatant isolated from mycelia expressing the ER marker AoClxA-351
EGFP were immunoblotted for GFP. The analysis showed that AoClxA-EGFP was concentrated 352
in the pellet and was only barely detectable in the supernatant (Fig. S2 in the supplemental 353
material). Thus, the 20,000×g pellet was used an ER-enriched fraction in subsequent analyses. 354
ER-enriched fractions were prepared from the wild-type, ΔAovip36, and ΔAoemp47 strains 355
expressing AmyB-proCHY and then analyzed by immunoblotting for CHY. A band 356
corresponding to the predicted size of AmyB-proCHY (approximately 87 kDa) was detected in 357
the ER-enriched fractions of all strains (Fig. 5A, upper blot), indicating that the fusion protein 358
had not been processed by the Kex2-like protease. As the S. cerevisiae homolog of Kex2p 359
resides in the Golgi (34), the detection of intact AmyB-proCHY supported the validity of the ER 360
enrichment process. Quantitative analysis of the immunoblots revealed that the amount of 361
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

18
AmyB-proCHY in the ER-enriched fraction was reduced in the ΔAovip36 and ΔAoemp47 strains 362
compared to that in the wild-type strain (Fig. 5A, lower). 363
Immunoblotting analysis of the ER-enriched fraction of mycelia expressing AmyB-EGFP 364
revealed that a band consistent with the predicted size of the fusion protein (approximately 73 365
kDa) was present in all strains (Fig. 5B, upper blot). However, the amount of AmyB-EGFP in the 366
ER-enriched fraction was decreased in the ΔAovip36 and ΔAoemp47 strains as compared with 367
the wild-type strain (Fig. 5B, lower). These results suggest that AoVip36 and AoEmp47 interfere 368
with secretion of carrier-fused heterologous proteins by increasing the ER retention rate, a 369
conclusion that is consistent with the increased secretion of AmyB-proCHY and AmyB-EGFP in 370
the ΔAovip36 and ΔAoemp47 strains (Fig. 4). 371
372
Secretion of carrier-fused heterologous protein in strains overexpressing AoVip36 and 373
AoEmp47. Fluorescent microscopic observation of the intracellular trafficking of AmyB-EGFP 374
in strains ΔAovip36 and ΔAoemp47 did not detect any differences compared with that in the 375
wild-type strain (data not shown). We speculated that the putative cargo receptors may be 376
expressed at low levels in the wild-type strain, thereby making it difficult to detect differences in 377
the ER retention rate. For this reason, AoVip36 and AoEmp47 were overexpressed in mDsRed-378
tagged forms under control of the amyB promoter in the presence of the inducing substrate 379
dextrin (35), which was added to cultures as a carbon source. The mDsRed-tagged AoVip36 and 380
AoEmp47 expressed under their native promoters complemented the ΔAovip36 and ΔAoemp47 381
strains, respectively, by suppressing the increased CHY production (Fig. S3 in the supplemental 382
material), which indicated that the mDsRed-tagged forms of cargo receptors are functional. 383
AmyB-EGFP was co-expressed in these strains to detect any effects from the overexpression of 384
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

19
the cargo receptors on the intracellular trafficking of heterologous proteins. In the wild-type 385
strain, an intense signal corresponding to AmyB-EGFP was detected at the hyphal tip (Fig. 6A), 386
similar to the localization of the Spitzenkörper, the location where exocytotic secretory 387
components and secretory proteins concentrate in filamentous fungi (23, 36–38). Under 388
overexpression conditions, mDsRed-AoVip36 was detected as punctate structures, whereas the 389
intracellular AmyB-EGFP signal seemed to be decreased and the localization was more 390
prominent at the cell periphery (Fig. 6B). This observation implied that the polarity of secretion 391
may have shifted from the hyphal tip to the cell periphery, likely as a result of increased 392
intracellular trafficking. In contrast to mDsRed-AoVip36, overexpression of mDsRed-AoEmp47 393
significantly increased the intracellular retention of AmyB-EGFP in tubular structures and 394
aggregates, where mDsRed-AoEmp47 was also predominantly localized (Fig. 6C). These 395
contrasting observations between the two putative cargo receptors suggested that overexpression 396
of AoVip36 enhanced, AmyB-EGFP trafficking, whereas overexpression of AoEmp47 promoted 397
the ER retention of AmyB-EGFP. 398
To confirm this speculation, the effect of overexpressing the two putative cargo receptors 399
on AmyB-EGFP secretion was examined. Immunoblotting of the culture supernatant revealed 400
that overexpression of mDsRed-AoVip36 significantly enhanced AmyB-EGFP secretion, 401
whereas overexpression of mDsRed-AoEmp47 impaired the secretion (Fig. 6D). Overexpression 402
of each cargo receptor did not affect the growth (Fig. S4A in the supplemental material), 403
indicating that these secretion effects were not due to the altered growth rate. Based on these 404
findings, we concluded that the overexpression of AoEmp47 promoted the intracellular retention 405
of AmyB-EGFP, whereas overexpressing AoVip36 alleviated the retention and even stimulated 406
the trafficking of AmyB-EGFP. 407
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

20
DISCUSSION 408
409
Although ER-Golgi cargo receptors are important for ER-Golgi trafficking, no reports prior 410
to the present study have examined the relationship between heterologous protein secretion and 411
cargo receptors. This study has provided evidence that two putative lectin-type cargo receptors 412
affect the intracellular trafficking of heterologous proteins in filamentous fungi. By genetically 413
deleting the putative receptors AoVip36 and AoEmp47, we succeeded in increasing the secretion 414
of carrier-fused heterologous proteins in A. oryzae. 415
The mDsRed-tagged form of AoVip36 predominantly localized to the Golgi (Fig. 2A), 416
similar to mammalian VIP36 (19, 31), suggesting that AoVip36 functions as a cargo receptor by 417
cycling between the Golgi and ER. We found that deletion of AoVip36 impaired the secretion of 418
α-amylase (Fig. 3), whereas AoVip36 overexpression increased the secretion of AmyB-EGFP 419
(Fig. 6D). Together, these results indicate that AoVip36 promotes the anterograde transport of 420
both endogenous and the EGFP-fused form of α-amylase, a property that is in agreement with 421
the fact that overexpression of mammalian VIP36 promotes secretion of the secretory 422
glycoprotein clusterin (18). It was also previously shown that mutation of the carbohydrate-423
binding domain of mammalian VIP36 leads to the loss of lectin activity, but does not result in 424
increased clusterin secretion (18). These results, taken together with our present findings, suggest 425
that the promotion of anterograde transport of VIP36 depends on its lectin activity. As the 426
affinity of mammalian VIP36 to high-mannose glycan is reportedly high in the pH range of 6.5 - 427
7.0, which corresponds to the pH of the ER, but is reduced sharply in the acidic conditions of 428
late-Golgi (39), VIP36 likely binds to glycoproteins in the ER and then transports and releases 429
them at the late-Golgi. Because A. oryzae α-amylase is a glycoprotein (40) and is also N-430
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

21
glycosylated when fused to heterologous proteins, it is assumed that the secretion-promoting 431
effect of AoVip36 might depend on its lectin activity. 432
In contrast to the anterograde transport-promoting function of AoVip36, deletion of AoVip36 433
increased heterologous protein secretion in A. oryzae (Fig. 3), providing evidence that this 434
putative cargo receptor has a protein retention function. In mammals, the glycoprotein α1-435
antitrypsin, which was identified as a retention target of VIP36, exhibits increased intracellular 436
transport upon silencing of VIP36 (19). The interacting complex between VIP36 and α1-437
antitrypsin was shown to repeatedly cycle from the Golgi to the ER, and was also found to co-438
immunoprecipitate with the ER molecular chaperone BiP (19). VIP36 also forms a complex with 439
BiP independently of its N-glycan binding activity (20), and overproduction of a lectin-deficient 440
form of VIP36 decreases clusterin secretion (18). These observations suggest that VIP36 also 441
negatively interferes with protein transport in an N-glycan-independent manner. Consistent with 442
this speculation, deletion of AoVip36 and AoEmp47 alleviated the ER retention of the 443
heterologous proteins chymosin and EGFP in A. oryzae (Fig. 5). The fusion proteins AmyB-444
proCHY and AmyB-EGFP induce UPR (10; data not shown), suggesting that they may be 445
abnormally folded. We speculate that misfolded AmyB-heterologous protein fusions might bind 446
to the chaperone BiP and subsequently form a protein complex with AoVip36, which then 447
recycles back to the ER, as demonstrated with the ER retention (Fig. 5). This hypothesis conflicts 448
with the fact that overexpression of AoVip36 leads to increased secretion of AmyB-EGFP (Fig. 449
6D). To explain these apparently conflicting findings, it is possible that the retention effect might 450
be saturated at a certain concentration of overexpressed AoVip36 when BiP molecules are fully 451
engaged. After exceeding this threshold concentration, AoVip36 overexpression would still 452
promote anterograde transport, leading to induction of the intracellular transport of glycoproteins. 453
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

22
The α-amylase activity in the culture supernatant was not significantly changed by 454
overexpression of AoVip36 (Fig. S4B in the supplemental material), which suggests that the 455
overexpression effect is mainly on the secretion of AmyB-EGFP but not endogenous α-amylase. 456
The observed localization of AoEmp47 in the ER and Golgi (Fig. 2B) is consistent with that 457
of S. cerevisiae Emp47p, which cycles between these two organelles (28, 29). When co-458
expressed with AoClxA-EGFP, mDsRed-AoEmp47 shifted the localization to the ER (Fig. 2B). 459
This observation is similar to the shift of AoEmp47 localization to a tubular structure when 460
overexpressed with AmyB-EGFP (Fig. 6C), suggesting that the overexpression and/or co-461
expression of AmyB-EGFP inhibited the ER exit of AoEmp47. Although AoClxA-EGFP 462
expressed from the native gene locus was shown to be functional (23), we hypothesize that 463
expression of the EGFP fusion of AoClxA might give some negative effect such as the ER stress 464
and inhibition of ER exit, leading to the increase in the retention of AoEmp47 in the ER. 465
Deletion of AoEmp47 increased the secretion of heterologous proteins and reduced their ER 466
retention (Figs. 4 and 5), whereas AoEmp47 overexpression led to the intracellular retention and 467
reduced secretion of AmyB-EGFP (Fig. 6C and D), suggesting that AoEmp47 functions to retain 468
heterologous proteins in the ER. These results are in clear contrast to the observations that 469
deletion of yeast Emp47p impairs the secretion of certain glycoproteins (28), and that the 470
mammalian homolog of AoEmp47, ERGIC-53, mediates the ER export of a subset of 471
glycoproteins, including α1-antitrypsin (17). The present study of cargo receptors in A. oryzae is 472
the first to reveal that an Emp47 homolog has a retention effect for the intracellular transport of 473
secretory proteins. Overexpression of AoEmp47 reduced the α-amylase activity in the culture 474
supernatant (Fig. S4B in the supplemental material). This raises the possibility that the secretion 475
of endogenous α-amylase could be inhibited by AoEmp47 overexpression. However, it remains 476
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

23
unknown whether the interaction of AoEmp47 with target proteins involves N-glycan binding via 477
lectin-like domains. This key question regarding the molecular basis of AoEmp47 interaction 478
with target cargo proteins, and how this interaction results in protein retention in the ER, 479
warrants further investigation. 480
A. oryzae possesses an inherently high potential for protein secretion, which is an attractive 481
property for recombinant protein production. However, before A. oryzae can be used as a cost-482
effective protein secretion host, its capacity for heterologous protein production needs to be 483
markedly improved. The present study has provided the first evidence that the transport of 484
proteins from the ER to Golgi represents a bottleneck in the secretory pathway of A. oryzae and 485
selectively hinders heterologous protein transport. Moreover, our findings suggest that modifying 486
the cargo receptor involved in the trafficking of intracellular proteins can improve heterologous 487
protein secretion. Further investigation on the interaction mechanism between cargo receptors 488
and their target proteins might provide useful information for not only the optimization of A. 489
oryzae, but also for other eukaryotic hosts, for protein production and contribute to the general 490
knowledge of intracellular vesicular trafficking. 491
492
493
ACKNOWLEDGEMENTS 494
H-D.H. was financially supported by an Ajinomoto ASEAN Scholarship. This study was also 495
supported by a Grant-in-Aid for Scientific Research (B) to K. Kitamoto from the Ministry of 496
Education, Culture, Sports, Science and Technology, Japan. 497
498
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

24
REFERENCES 499
500
1. Punt PJ, van Biezen N, Conesa A, Albers A, Mangnus J, van den Hondel C. 2002. 501
Filamentous fungi as cell factories for heterologous protein production. Trends Biotechnol. 502
20:200–206. 503
2. Yoon J, Maruyama J, Kitamoto K. 2011. Disruption of ten protease genes in the 504
filamentous fungus Aspergillus oryzae highly improves production of heterologous 505
proteins. Appl. Microbiol. Biotechnol. 89:747–759. 506
3. Yoon J, Aishan T, Maruyama J, Kitamoto K. 2010. Enhanced production and secretion 507
of heterologous proteins by the filamentous fungus Aspergillus oryzae via disruption of 508
vacuolar protein sorting receptor gene Aovps10. Appl. Environ. Microbiol. 76:5718–5727. 509
4. Yoon J, Kikuma T, Maruyama J, Kitamoto K. 2013. Enhanced production of bovine 510
chymosin by autophagy deficiency in the filamentous fungus Aspergillus oryzae. PLoS 511
One 8:e62512. 512
5. Tsuchiya K, Nagashima T, Yamamoto Y, Gomi K, Kitamoto K, Kumagai C, Tamura 513
G. 1994. High level secretion of calf chymosin using a Glucoamylase-prochymosin fusion 514
gene in Aspergillus oryzae. Biosci. Biotechnol. Biochem. 58:895-899. 515
6. Nyyssönen E, Keränen S. 1995. Multiple roles of the cellulase CBHI in enhancing 516
production of fusion antibodies by the filamentous fungus Trichoderma reesei. Curr. 517
Genet. 28:71–79. 518
7. Nakajima K, Asakura T, Maruyama J, Morita Y, Oike H, Shimizu-ibuka A, 519
Sorimachi H, Arai S, Abe K. 2006. Extracellular production of neoculin, a sweet-tasting 520
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

25
heterodimeric protein with taste-modifying activity, by Aspergillus oryzae. Appl. Environ. 521
Microbiol. 72:3716–3723. 522
8. Gouka RJ, Punt PJ. 1997. Glucoamylase gene fusions alleviate limitations for protein 523
production in Aspergillus awamori at the transcriptional and (post) translational level. 524
Appl. Environ. Microbiol. 63:488–497. 525
9. Guillemette T, van Peij NNME, Goosen T, Lanthaler K, Robson GD, van den Hondel 526
C a MJJ, Stam H, Archer DB. 2007. Genomic analysis of the secretion stress response 527
in the enzyme-producing cell factory Aspergillus niger. BMC Genomics 8:158. 528
10. Ohno A, Maruyama J, Nemoto T, Arioka M, Kitamoto K. 2011. A carrier fusion 529
significantly induces unfolded protein response in heterologous protein production by 530
Aspergillus oryzae. Appl. Microbiol. Biotechnol. 92:1197–1206. 531
11. Lombraña M, Moralejo FJ, Pinto R, Martín JF, Lombran M, Martı JF. 2004. 532
Modulation of Aspergillus awamori thaumatin secretion by modification of bipA gene 533
expression. Appl. Environ. Microbiol. 70:5145–5152. 534
12. Moralejo F, Watson AJ, Jeenes DJ, Archer DB, Martin JF. 2001. A defined level of 535
protein disulfide isomerase expression is required for optimal secretion of thaumatin by 536
Aspergillus awamori. Mol. Genet. Genomics 266:246–253. 537
13. Vembar SS, Brodsky JL. 2008. One step at a time: endoplasmic reticulum-associated 538
degradation. Nat. Rev. Mol. Cell Biol. 9:944–957. 539
14. Valkonen M, Ward M, Wang H, Penttila M. 2003. Improvement of foreign protein 540
production in Aspergillus niger var. awamori by constitutive induction of the unfolded-541
protein response. Appl. Environ. Microbiol. 69:6979–6986. 542
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

26
15. Spang A. 2009. On vesicle formation and tethering in the ER-Golgi shuttle. Curr. Opin. 543
Cell Biol. 21:531–536. 544
16. Dancourt J, Barlowe C. 2010. Protein sorting receptors in the early secretory pathway. 545
Annu. Rev. Biochem. 79:777–802. 546
17. Hauri HP, Kappeler F, Andersson H, Appenzeller C. 2000. ERGIC-53 and traffic in 547
the secretory pathway. J. Cell Sci. 113:587–596. 548
18. Hara-Kuge S, Ohkura T, Ideo H, Shimada O, Atsumi S, Yamashita K. 2002. 549
Involvement of VIP36 in intracellular transport and secretion of glycoproteins in polarized 550
Madin-Darby canine kidney (MDCK) cells. J. Biol. Chem. 277:16332–16339. 551
19. Reiterer V, Nyfeler B, Hauri H-P. 2010. Role of the lectin VIP36 in post-ER quality 552
control of human alpha1-antitrypsin. Traffic 11:1044–1055. 553
20. Nawa D, Shimada O, Kawasaki N, Matsumoto N, Yamamoto K. 2007. Stable 554
interaction of the cargo receptor VIP36 with molecular chaperone BiP. Glycobiology 555
17:913–921. 556
21. Machida M, Asai K, Sano M, Tanaka T, Kumagai T, Terai G, Kusumoto K-I, Arima 557
T, Akita O, Kashiwagi Y, Abe K, Gomi K, Horiuchi H, Kitamoto K, Kobayashi T, 558
Takeuchi M, Denning DW, Galagan JE, Nierman WC, Yu J, Archer DB, Bennett 559
JW, Bhatnagar D, Cleveland TE, Fedorova ND, Gotoh O, Horikawa H, Hosoyama A, 560
Ichinomiya M, Igarashi R, Iwashita K, Juvvadi PR, Kato M, Kato Y, Kin T, 561
Kokubun A, Maeda H, Maeyama N, Maruyama J, Nagasaki H, Nakajima T, Oda K, 562
Okada K, Paulsen I, Sakamoto K, Sawano T, Takahashi M, Takase K, Terabayashi 563
Y, Wortman JR, Yamada O, Yamagata Y, Anazawa H, Hata Y, Koide Y, Komori T, 564
Koyama Y, Minetoki T, Suharnan S, Tanaka A, Isono K, Kuhara S, Ogasawara N, 565
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

27
Kikuchi H. 2005. Genome sequencing and analysis of Aspergillus oryzae. Nature 566
438:1157–1161. 567
22. Maruyama J, Kitamoto K. 2008. Multiple gene disruptions by marker recycling with 568
highly efficient gene-targeting background (DeltaligD) in Aspergillus oryzae. Biotechnol. 569
Lett. 30:1811–1817. 570
23. Kimura S, Maruyama J, Watanabe T, Ito Y, Arioka M, Kitamoto K. 2010. In vivo 571
imaging of endoplasmic reticulum and distribution of mutant α-amylase in Aspergillus 572
oryzae. Fungal Genet. Biol. 47:1044–1054. 573
24. Kitamoto K. 2002. Molecular biology of the Koji molds. Adv. Appl. Microbiol. 51:129–574
153. 575
25. Nemoto T, Watanabe T, Mizogami Y, Maruyama J, Kitamoto K. 2009. Isolation of 576
Aspergillus oryzae mutants for heterologous protein production from a double proteinase 577
gene disruptant. Appl. Microbiol. Biotechnol. 82:1105–1114. 578
26. Bolte S, Cordelières FP. 2006. A guided tour into subcellular colocalization analysis in 579
light microscopy. J. Microsc. 224:213–232. 580
27. Sato K, Nakano A. 2003. Oligomerization of a cargo receptor directs protein sorting into 581
COPII-coated transport vesicles. Mol. Biol. Cell 14:3055–3063. 582
28. Sato K, Nakano A. 2002. Emp47p and its close homolog Emp46p have a tyrosine-583
containing endoplasmic reticulum exit signal and function in glycoprotein secretion in 584
Saccharomyces cerevisiae. Mol. Biol. Cell 13:2518–2532. 585
29. Schröder S, Schimmöller F, Singer-Krüger B, Riezman H. 1995. The Golgi-586
localization of yeast Emp47p depends on its di-lysine motif but is not affected by the ret1-587
1 mutation in alpha-COP. J. Cell Biol. 131:895–912. 588
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

28
30. Pantazopoulou A, Peñalva MA. 2009. Organization and dynamics of the Aspergillus 589
nidulans Golgi during apical extension and mitosis. Mol. Biol. Cell 20:4335–4347. 590
31. Füllekrug J, Scheiffele P, Simons K. 1999. VIP36 localisation to the early secretory 591
pathway. J. Cell Sci. 112:2813–2821. 592
32. Nemoto T, Maruyama J, Kitamoto K. 2012. Contribution ratios of amyA, amyB, amyC 593
genes to high-level α-amylase expression in Aspergillus oryzae. Biosci. Biotechnol. 594
Biochem. 76:1477–1483. 595
33. Nemoto T, Maruyama J, Kitamoto K. 2009. Improvement of heterologous protein 596
production in Aspergillus oryzae by RNA interference with α-Amylase genes. Biosci. 597
Biotechnol. Biochem. 73:2370–2373. 598
34. Redding K, Holcomb C, Fuller RS. 1991. Immunolocalization of Kex2 protease 599
identifies a putative late Golgi compartment in the yeast Saccharomyces cerevisiae. J. Cell 600
Biol. 113:527–538. 601
35. Tsuchiya K, Tada S, Gomi K, Kitamoto K, Kumagai C, Jigami Y, Tamura G. 1992. 602
High level expression of the synthetic human lysozyme gene in Aspergillus oryzae. Appl. 603
Microbiol. Biotechnol. 38:109–114. 604
36. Khalaj V, Brookman JL, Robson GD. 2001. A study of the protein secretory pathway of 605
Aspergillus niger using a glucoamylase-GFP fusion protein. Fungal Genet. Biol. 32:55–65. 606
37. Taheri-talesh N, Horio T, Araujo-baza L, Dou X, Espeso EA, Pen MA, Osmani SA, 607
Oakley BR. 2008. The tip growth apparatus of Aspergillus nidulans 19:1439–1449. 608
38. Hayakawa Y, Ishikawa E, Shoji J-Y, Nakano H, Kitamoto K. 2011. Septum-directed 609
secretion in the filamentous fungus Aspergillus oryzae. Mol. Microbiol. 81:40–55. 610
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

29
39. Kamiya Y, Yamaguchi Y, Takahashi N, Arata Y, Kasai K-I, Ihara Y, Matsuo I, Ito Y, 611
Yamamoto K, Kato K. 2005. Sugar-binding properties of VIP36, an intracellular animal 612
lectin operating as a cargo receptor. J. Biol. Chem. 280:37178–37182. 613
40. Minobe S, Nakajima H, Ito N, Funakoshi I, Yamashina I. 1979. Structure of a major 614
oligosaccharide of Taka-amylase A. J. Biochem. 86:1851–1854. 615
616
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

30
TABLE 1 Strain list 617
Strain name Host Genotype References 618
NSlD1 NSPlD-1 niaD- sC- adeA- argB::adeA- ΔligD::argB ΔpyrG::adeA pgEpG[pyrG] Yoon et al., 2011 619
NSR-ΔlD-2 NSAR1 niaD- sC- adeA- ΔargB::adeA- ΔligD::argB Maruyama et al., 2008 620
NS-CaG-1 NSR-ΔlD-2 niaD- sC- adeA- ΔargB::adeA- ΔligD::argB AoclxA-egfp::adeA Kimura et al., 2010 621
NS-Grh1G-1 NSR-ΔlD-2 niaD- sC- adeA- ΔargB::adeA- ΔligD::argB Aogrh1-egfp::adeA This study 622
S-CaGRVp NS-CaG-1 niaD- sC- adeA- ΔargB::adeA- ΔligD::argB AoclxA-egfp::adeA pEpARVpN[PamyB::Aovip36-mdsred::TamyB::niaD] This study 623
S-CaGREp NS-CaG-1 niaD- sC- adeA- ΔargB::adeA- ΔligD::argB AoclxA-egfp::adeA pEpAREpN[PamyB::Aoemp47-mdsred::TamyB::niaD] This study 624
S-G1GRVp NS-Grh1G-1 niaD- sC- adeA- ΔargB::adeA- ΔligD::argB Aogrh1-egfp::adeA pEpARVpN[PamyB::Aovip36-mdsred::TamyB::niaD] This study 625
S-G1GREp NS-Grh1G-1 niaD- sC- adeA- ΔargB::adeA- ΔligD::argB Aogrh1-egfp::adeA pEpAREpN[PamyB::Aoemp47-mdsred::TamyB::niaD] This study 626
NSlDVp-1 NSPlD-1 niaD- sC- adeA- ΔargB::adeA- ΔligD::argB ΔpyrG::adeA ΔAovip36::pyrG This study 627
NSlDEp-1 NSPlD-1 niaD- sC- adeA- ΔargB::adeA- ΔligD::argB ΔpyrG::adeA ΔAoemp47::pyrG This study 628
SlD-aG-2 NSlD-1 niaD-::pNamyBEGFP[PamyB::amyB-egfp::TagdA::niaD] sC- adeA- ΔargB::adeA- ΔligD::argB ΔpyrG::adeA pgEpG[pyrG] This study 629
SlDVp-aG-2 NSlDVp-1 niaD-::pNamyBEGFP[PamyB::amyB-egfp::TagdA::niaD] sC- adeA- ΔargB::adeA- ΔligD::argB ΔpyrG::adeA ΔAovip36::pyrG This study 630
SlDEp-aG-2 NSlDEp-1 niaD-::pNamyBEGFP[PamyB::amyB-egfp::TagdA::niaD] sC- adeA- ΔargB::adeA- ΔligD::argB ΔpyrG::adeA ΔAoemp47::pyrG This study 631
SlD-AKC NSlD-1 niaD-::pgAKCN[PamyB::amyB-(kex2)-proCHY::TamyB::niaD] sC- adeA- ΔargB::adeA- ΔligD::argB ΔpyrG::adeA pgEpG[pyrG] This study 632
SlDVp-AKC NSlDVp-1 niaD-::pgAKCN[PamyB::amyB-(kex2)-proCHY::TamyB::niaD] sC- adeA- ΔargB::adeA- ΔligD::argB ΔpyrG::adeA pgEpG[pyrG] This study 633
ΔAovip36::pyrG 634
SlDEp-AKC NSlDEp-1 niaD-::pgAKCN[PamyB::amyB-(kex2)-proCHY::TamyB::niaD] sC- adeA- ΔargB::adeA- ΔligD::argB ΔpyrG::adeA pgEpG[pyrG] This study 635
ΔAoemp47::pyrG 636
lD-aGRVp SlD-aG-2 niaD-::pNamyBegfp[PamyB::amyB-egfp::TagdA::niaD] sC- adeA- ΔargB::adeA- ΔligD::argB ΔpyrG::adeA This study 637
pgEpG[pyrG] pNRVp[PamyB::Aovip36-mdsred::TamyB::sC] 638
lD-aGREp SlD-aG-2 niaD-::pNamyBegfp[PamyB::amyB-egfp::TagdA::niaD] sC-adeA- ΔargB::adeA- ΔligD::argB ΔpyrG::adeA This study 639
pgEpG[pyrG] pNRVp[PamyB::Aoemp47-mdsred::TamyB::sC] 640
641
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

31
Figure legends 642
643
FIG 1 Comparison of domain organization between putative cargo receptors in A. oryzae and 644
their known homologs. (A) AoVip36 and Homo sapiens VIP36 (Accession number 645
NP_006807.1). (B) AoEmp47 and S. cerevisiae Emp47p (Accession number CAA60953.1). 646
Signal sequence (SS) was predicted by SignalP 4.0 (http://www.cbs.dtu.dk/services/SignalP/). 647
Coiled-coil domain (CCD) was predicted by COILS (http://embnet.vital-648
it.ch/software/COILS_form.html). Transmembrane region (TM) was predicted by TMHMM 649
server 2.0 (http://www.cbs.dtu.dk/services/TMHMM/). Lectin-like domain was predicted by 650
Search for Conserved Domains in The National Center for Biotechnology Information 651
(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi). (C) Tyrosine-containing motifs and 652
dilysine motifs of Emp47 and AoEmp47 are boxed and indicated in italic and bold characters, 653
respectively. 654
655
FIG 2 Subcellular localizations of two putative cargo receptors. mDsRed-tagged AoVip36 or 656
AoEmp47 was co-expressed with the EGFP-tagged version of the ER marker AoClxA-EGFP or 657
the Golgi marker AoGrh1-EGFP. Approximately 105 conidia were inoculated in 100 μl CD+Met 658
and incubated for 18 h at 30°C. (A) AoVip36 localized exclusively to the Golgi. Representative 659
fluorescence microscopy images of the apical area of hyphae expressing AoClxA-EGFP or 660
AoGrh1-EGFP with mDsRed-AoVip36. Arrowheads indicate the punctuate structures labeled by 661
both AoGrh1-EGFP and mDsRed-AoVip36 (Bars = 5 μm). (B) AoEmp47 localized to both the 662
ER and Golgi. Representative fluorescence microscopy images of the apical regions of hyphae 663
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

32
expressing AoClxA-EGFP or AoGrh1-EGFP with mDsRed-AoEmp47. Arrowheads indicate 664
punctuate structures labeled by both AoGrh1-EGFP and mDsRed-AoEmp47 (Bars = 5 μm). (C) 665
Graph representing mean Mander’s coefficient quantifying fraction of cargo receptors 666
overlapping with the ER and Golgi (n = 3 biological replicates, error bars = standard errors,). 667
668
FIG 3 Effect of cargo receptor gene deletion on α-amylase secretion. Approximately 105 conidia 669
of the wild-type, ΔAovip36, or ΔAoemp47 strains were cultivated in 20 ml 5×DPY (pH 5.5) at 670
30°C. (A) α-Amylase activities in the culture supernatant on the second day of cultivation were 671
quantified (n = 3 biological replicates, error bars = standard errors, *P<0.05 unpaired t-test). (B) 672
Wet mycelia weight of the control and deletion strains on the second day of cultivation. (C) Total 673
amount of proteins in the culture supernatants of the control and deletion strains on the second 674
day of cultivation. (D) SDS-PAGE/Coomassie staining of 10 µl of culture supernatant on the 675
second day of cultivation. The arrow indicates the band predicted to correspond to α-amylase. 676
677
FIG 4 Deletion of AoVip36 or AoEmp47 increased the secretion of carrier-fused heterologous 678
proteins in A. oryzae. Conidia of the wild-type, ΔAovip36, or ΔAoemp47 strains expressing 679
AmyB-proCHY or AmyB-EGFP were cultivated in 20 ml 5×DPY (pH 5.5) at 30°C. (A) 680
Immunoblotting of 10 μl culture supernatants of 107
conidia expressing AmyB-proCHY on the 681
second day of cultivation using anti-chymosin antibody. The arrow indicates the estimated 682
position of mature chymosin. (B) Milk clotting activity in the culture supernatant of 105 conidia 683
expressing AmyB-proCHY on the fourth day of cultivation (n = 3 biological replicates, error 684
bars = standard errors, *P<0.05 unpaired t-test). (C) Representative immunoblot image of 10 μl 685
culture supernatants of 107
conidia expressing AmyB-EGFP on the second day of cultivation 686
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

33
using anti-GFP antibody. The arrow indicates the estimated position of the AmyB-EGFP fusion 687
protein. The bar graph shows the quantitative analysis of the immunoblot results (n = 3 688
biological replicates, error bars = standard errors, *P<0.05 unpaired t-test). 689
690
FIG 5 Cargo receptor protein deletion alleviated ER retention of carrier-fused heterologous 691
proteins. Approximately 107 conidia of the wild-type, ΔAovip36, or ΔAoemp47 strains 692
expressing AmyB-proCHY or AmyB-EGFP were cultivated in 20 ml 5×DPY (pH 5.5) for 2 days 693
at 30°C. The ER-enriched fractions with 2 μg proteins were immunoblotted with the indicated 694
antibodies. (A) Representative immunoblot image of the ER-enriched fraction of the strain 695
expressing AmyB-proCHY for CHY. The arrow indicates the estimated position of AmyB-696
proCHY. The bar graph shows the quantitative analysis of the immunoblotting results (n = 3 697
biological replicates, error bars = standard errors, *P<0.05 unpaired t-test). (B) Representative 698
immunoblot image of the ER-enriched fraction of the strain expressing AmyB-EGFP for GFP. 699
The arrow indicates the estimated position of fusion AmyB-EGFP. The bar graph shows the 700
quantitative analysis of the immunoblotting results (n = 3 biological replicates, error bars = 701
standard errors, *P<0.05 unpaired t-test). 702
703
FIG 6 Effect of cargo receptor overexpression on the intracellular trafficking of AmyB-EGFP. 704
An N-terminally mDsRed-tagged version of AoVip36 or AoEmp47 was overexpressed under 705
control of the amyB promoter (P-amyB) with AmyB-EGFP. Approximately 105 conidia were 706
inoculated in 100 μl CD+Met and incubated for 24 h at 30°C. (A) Representative fluorescence 707
microscopy image of the apical region of hyphae expressing AmyB-EGFP (bars = 5 μm). (B) 708
Representative fluorescence microscopy images of the apical region of a hyphae overexpressing 709
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from

34
mDsRed-AoVip36 with AmyB-EGFP (bars = 5 μm). AoVip36 overexpression shifted the 710
intracellular AmyB-EGFP signal to the cell periphery. (C) Representative fluorescence 711
microscopy images of the apical region of hyphae overexpressing mDsRed-AoEmp47 with 712
AmyB-EGFP. AoEmp47 overexpression promoted intracellular retention of AmyB-EGFP in ER-713
like tubular structures. The arrowhead indicates an abnormal aggregate containing both AmyB-714
EGFP and mDsRed-AoEmp47. (D) Representative immunoblot image of the culture supernatant 715
of the wild-type, mDsRed-AoVip36, or mDsRed-AoEmp47 overexpressing strains for GFP. 716
Overexpression of mDsRed-AoVip36 improved AmyB-EGFP secretion, whereas overexpression 717
of mDsRed-AoEmp47 impaired AmyB-EGFP secretion. The arrow indicates the estimated 718
position of AmyB-EGFP. 719
720
on March 31, 2019 by guest
http://aem.asm
.org/D
ownloaded from