Embed Size (px)
Citation preview

1
Anthranilate deteriorates biofilm structure of Pseudomonas 1
aeruginosa and antagonizes the biofilm-enhancing indole effect 2
Soo-Kyoung Kim, Ha-Young Park, and Joon-Hee Lee* 3
Department of Pharmacy, College of Pharmacy, Pusan National University, Busan, 4
609-735, South Korea 5
*Send correspondence to: Joon-Hee Lee 6
Laboratory of Microbiology, Department of Pharmacy, 7
College of Pharmacy, Pusan National University. 8
Research building 537, San 30, Jangjun-Dong, Geumjung-9
Gu, Busan, 609-735, South Korea 10
Phone: 82-051-510-2821 11
FAX: 82-051-513-6754 12
Email: [email protected] 13
Running Title; Anthranilate effect on biofilm formation 14
Key words; Pseudomonas aeruginosa, anthranilate, quorum sensing, biofilm 15
16
AEM Accepted Manuscript Posted Online 23 January 2015Appl. Environ. Microbiol. doi:10.1128/AEM.03551-14Copyright © 2015, American Society for Microbiology. All Rights Reserved.
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

2
Abstract 17
Anthranilate and indole are alternative degradation products of tryptophan depending on 18
bacterial species. While indole enhances the biofilm formation of Pseudomonas 19
aeruginosa, we found that anthranilate, the tryptophan degradation product of P. 20
aeruginosa has an opposite effect on P. aeruginosa biofilm formation to indole, in 21
which anthranilate deteriorated the mushroom structure of biofilm. The anthranilate 22
effect on biofilm formation was differentially exerted depending on the developmental 23
stage and the presence of shear force. Anthranilate slightly accelerated the initial 24
attachment of P. aeruginosa at the early stage of biofilm development and appeared to 25
build more biofilm without shear force. But, anthranilate weakened the biofilm structure 26
in the late stage, deteriorating the mushroom structure of biofilms with shear force to 27
make a flat biofilm. To investigate the interplay of anthranilate with indole in biofilm 28
formation, biofilms were co-treated with anthranilate and indole, and the result showed 29
that anthranilate antagonized the biofilm-enhancing effect of indole. Anthranilate was 30
able to deteriorate the pre-formed biofilm. The effect of anthranilate and indole on 31
biofilm formation was quorum sensing-independent. AntR, a regulator of anthranilate-32
degrading metabolism was synergistically activated by co-treatment with anthranilate 33
and indole, suggesting that indole might enhance the biofilm formation by facilitating 34
the degradation of anthranilate. Anthranilate slightly but significantly affected the c-di-35
GMP level and transcription of major extracellular polysaccharide (Psl, Pel, and 36
alginate) operons. These results suggest that anthranilate may be a promising anti-37
biofilm agent and antagonize the indole effect on P. aeruginosa biofilm formation.38
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

3
Introduction 39
Biofilms are a representative example of bacterial group behavior that provides cells 40
with many biological advantages like high infectivity, antibiotic resistance, and strong 41
survivability (1, 2). Currently, most persistent bacterial infections are believed to be 42
associated with antibiotic-resistant biofilms of pathogenic bacteria (3, 4). Pseudomonas 43
aeruginosa, a Gram-negative bacterium, is a ubiquitous and major opportunistic human 44
pathogen. The colonization and biofilm formation of P. aeruginosa cause great loss in 45
many industrial facilities and serious infections such as cystic fibrosis, microbial 46
keratitis, and burn wound infections in humans (4-8). Therefore, control of P. 47
aeruginosa biofilms is a very important issue in medicine, public health, and industry. 48
Anthranilate and indole are both aromatic compounds produced from tryptophan 49
metabolism. In bacteria, tryptophan is metabolized differently depending on the 50
bacterial species, and the key enzyme of this differentiation is tryptophanase, encoded 51
by the tnaA gene (9). Many bacteria, such as Escherichia coli, Haemophilus influenzae, 52
and Vibrio vulnificus produce indole from tryptophan, since they have tnaA that 53
converts tryptophan into indole, pyruvate, and ammonia (9, 10). However, some other 54
bacteria including P. aeruginosa degrade tryptophan to anthranilate through kynurenine 55
pathway using kynBAU genes (9). Therefore, anthranilate and indole are alternative 56
degradation products of tryptophan in the microbial community and if P. aeruginosa 57
exists in tryptophan-rich environment with other indole-producing bacteria, it will 58
encounter indole from other bacteria as it produces anthranilate. 59
The effects of tryptophan and indole on the biofilm formation of P. aeruginosa have 60
been recently reported. Tryptophan had an inhibitory effect on the biofilm formation by 61
E. coli and P. aeruginosa (11, 12). Indole also inhibited the biofilm formation of E. coli 62
that metabolizes tryptophan to indole (13). Interestingly, indole enhanced the biofilm 63
formation of P. aeruginosa that degrades tryptophan to anthranilate, but not to indole 64
(13, 14). While P. aeruginosa does not produce indole, P. aeruginosa may encounter 65
indole produced by indole-producing bacteria like E. coli in mixed bacterial community 66
in nature and its physiology can be influenced by indole (10, 14). However, the effect of 67
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

4
anthranilate on the biofilm formation of P. aeruginosa has not been addressed yet, 68
although it is a real product of the tryptophan metabolism of P. aeruginosa. A recent 69
study showed that P. aeruginosa biofilm cells have enhanced anthranilate-degrading 70
activity (15), implying the possible involvement of anthranilate in biofilm physiology. 71
Biofilm formation by P. aeruginosa can be controlled in a cell density-dependent 72
manner by a quorum-sensing (QS) system that allows bacteria to communicate with 73
each other via signaling molecules, acyl-homoserine lactones (AHLs) and Pseudomonas 74
quinolone signal (PQS; 2-heptyl-3-hydroxy-4-quinolone) (16, 17). Major AHLs of P. 75
aeruginosa are N-3-oxododecanoyl-L-homoserine lactone (3OC12-HSL) and N-76
butyryl-L-homoserine lactone (C4-HSL), which are synthesized by LasI and RhlI, 77
respectively, and bind to their cognate receptors, LasR (for 3OC12-HSL), QscR (for 78
3OC12-HSL), and RhlR (for C4-HSL) to regulate target genes (18-20). PQS, another 79
important QS signal, also plays a significant role in the regulation of virulence genes 80
and biofilm formation (21). In P. aeruginosa, indole inhibits PQS production (13) and 81
anthranilate is a precursor of PQS biosynthesis (22). Moreover, the metabolism of 82
anthranilate is growth phase-differentially regulated by QS system in P. aeruginosa (23). 83
Anthranilate and indole therefore intersect with QS regulation and biofilm formation. 84
Since anthranilate is also a precursor of tryptophan biosynthesis and an intermediate 85
that is metabolized through the TCA cycle, it is a key metabolite of P. aeruginosa at the 86
metabolic branch point (22, 23). The complex relationship among QS, biofilm, 87
anthranilate and indole in P. aeruginosa is schematically described in Fig. S1. 88
In this study, we investigated the effects of anthranilate and indole on the biofilm 89
formation of P. aeruginosa and found that anthranilate deteriorated the biofilm, making 90
a flat biofilm. We suggest that anthranilate may be a promising anti-biofilm agent and 91
can antagonize the indole effect on P. aeruginosa biofilm formation. 92
Materials and Methods 93
Bacterial strains, culture conditions, and plasmids The bacterial strains and plasmids 94
used in this study are listed in Table 1. P. aeruginosa strains were grown at 37°C in 95
Luria-Bertani (LB; yeast 5 g/L, bacto-tryptone 10 g/L, NaCl 5 g/L) medium with 96
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

5
vigorous shaking at 170 rpm. Growth was measured by optical density at 600 nm 97
(OD600). Antibiotics were used at the following concentrations: carbenicillin, 100 μg/ml; 98
ampicillin, 50 μg/ml; gentamicin, 10 μg/ml (for E. coli) and 50 μg/ml (for P. 99
aeruginosa). Indole and anthranilate were dissolved at 1 M in DMSO for stock solution 100
and diluted into the media at final concentrations of 0.4 mM and 0.1 mM, respectively. 101
Measurement of QS-regulators and AntR in P. aeruginosa To make P. aeruginosa 102
reporter strains for measuring the activity of the QS-regulators, the specific promoter-103
lacZ fusion plasmids, pSC11 (lasIp-lacZ fusion for measuring the LasR activity), 104
pJL101 (PA1897p-lacZ fusion for measuring the QscR activity), and pJL201 (antAp-lacZ 105
fusion for measuring the AntR activity) were transformed into PAO1 (Table 1). The 106
overnight cultures of each reporter strain were inoculated at 1% into fresh LB broth 107
containing carbenicillin and cultivated with/without 0.1 mM anthranilate or 0.4 mM 108
indole, or both. During growth, aliquots were taken every hour and β-galactosidase 109
activities were measured as described below. 110
Measurement of AntR activity in E. coli (E. coli reconstitution analysis) To 111
overexpress AntR of P. aeruginosa in E. coli, pJN105A plasmid was used (Table 1). 112
Two compatible plasmids, pJL201 and pJN105A, were transformed into E. coli DH5α 113
and the transformants were grown overnight in LB medium. The cells were then 114
inoculated into fresh LB medium at initial OD600 = 0.04 and grown to OD600 = 0.3 115
without arabinose. After 0.4% arabinose was added for the induction of AntR, 0.1 mM 116
anthranilate or 0.4 mM indole or both were added for 2 h. The β-galactosidase activity 117
was then measured as described below. As a control, pJN105, the parental plasmid of 118
pJN105A, was co-transformed with pJL201, the transformed cells were grown in the 119
same way, and the β-galactosidase activity was measured. 120
β-Galactosidase activity assay β-galactosidase activity was assayed with Tropix-121
plusTM kit (Applied Biosystems, USA) with slight modification of the manufacturer’s 122
instruction, as described elsewhere (24). First, OD600 of cultures was measured. One 123
hundred microliters of the cultures were taken and 10 μl of chloroform was added. After 124
vigorous vortex and 15-min incubation at room temperature, 10 μl from the top was 125
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

6
transferred to a new tube and 100 μl of 1:100-diluted substrate solution was added. 126
After 45-min incubation at room temperature in the dark, 150 μl of light emission 127
solution (Accelerator II) was added and the luminescence was promptly measured with 128
multi-well plate reader (Tristar LB941, Berthold). The luminescence was normalized 129
with the OD600 of the cultures and the activity was presented in luminescence/OD600. 130
Biofilm formation and assay The static biofilm assay was carried out as described 131
elsewhere (25, 26). P. aeruginosa PAO1 cells were grown to OD600 = 3 in LB broth with 132
vigorous shaking. These cells were diluted to OD600 = 0.06 in fresh M63 medium [M63 133
salt (KH2PO4 12 g/L, K2HPO4 28 g/L, NH4SO4 8 g/L), 1 mM MgSO4, 0.5% casamino 134
acid, 0.2% citrate] on 96-well polystyrene plates and incubated at 37°C for 24 h without 135
shaking. After cell growth was measured by OD600, planktonic cells were poured out 136
and the plate was washed with water and dried for 10 min. Then, 180 μl of crystal violet 137
(0.1%, wt/vol) was added to each well and incubated for 7 min to stain biofilms 138
attached to the well surface. After a brief wash, the biofilm-staining crystal violet was 139
dissolved in 200 μl of absolute ethanol. The absorbance was then measured at 550 nm 140
(A550) to determine the amount of crystal violet, which was normalized with cell growth 141
(OD600). 142
The drip-flow biofilm formation was carried out as previously described (27) with 143
slight modification. P. aeruginosa cells harboring the GFP-expressing plasmid, pAB1 144
(Table 1), were cultured overnight and diluted to OD600 = 1 in fresh LB broth, 1 ml of 145
which was dropped on to slide glass. This inoculation was repeated 3 times with 10-min 146
intervals. After 40-min incubation, continuous dropping of 1% bacto tryptic soy broth 147
(TSB; tryptone 5.6 g/L, soytone 1.6 g/L, NaCl 1.6 g/L, dextrose 0.8 g/L, K2HPO4 0.8 148
g/L) containing 100 μg/ml carbenicillin and 1 mM IPTG started at a flow rate = 0.5 149
ml/min. 150
For flow-cell biofilm formation, the same GFP-expressing P. aeruginosa cells were 151
grown overnight and diluted to OD600 = 0.5 in LB, 200 μl of which was injected into 152
flow-cell chamber (2 mm by 2 mm by 50 mm). Cells were incubated for 1 h without 153
flow for attachment, and then the TSB media containing 100 μg/ml carbenicillin and 1 154
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

7
mM IPTG flowed at 200 μl /min. Biofilms in the flow-cell were grown for 5 d at room 155
temperature and visualized by GFP or staining with 0.1% calcofluor white for 15 min. 156
For the dispersion of pre-formed biofilm, biofilm was formed in flow-cell for 3 d and 157
treated with 0.1 mM anthranilate for 1 d. Biofilms were observed on fluorescence 158
microscopes as described below. 159
Biofilm imaging and quantification Biofilm images were obtained using Confocal 160
Laser Scanning Microscopy (CLSM; Olympus, FV10i) or fluorescence microscopy 161
(Zeiss, Axioskop FL). The excitation wavelength for GFP was 488 nm and the emission 162
wavelength was 500 nm. For calcofluor white, the excitation and emission wavelengths 163
were 355 nm and 440 nm, respectively. The 3-dimensional images of biofilms were 164
reconstructed from plane images by using Bitplane Imaris 6.3.1 image analysis software. 165
Quantification of biofilms was performed using integrated morphometry analysis in 166
Metamorph 7.7 version (Molecular Devices, USA) and statistical analysis in AutoQuant 167
X (Mediacy, USA). 168
Measurement of cell surface hydrophobicity Bacteria were cultured overnight in LB 169
broth at 37°C with vigorous shaking. An aliquot (1.2 ml) of cell suspension was mixed 170
with 200 μl of hexane with vigorous shaking for 2 min. After separating the aqueous 171
and organic phases, the aqueous phase was carefully taken and OD600 was measured. 172
The hydrophobicity of cells was calculated based on the cell adhesion to organic phase 173
as described elsewhere (26, 28) from the following equation: percentage of adhesion = 174
100 × [(OD600 of the initial bacterial suspension – OD600 of the aqueous phase)/OD600 of 175
the initial bacterial suspension]. 176
Quantitative analysis of anthranilate by HPLC P. aeruginosa cells were grown 177
overnight in LB medium with or without 0.4 mM indole. Cells were then removed by 178
centrifugation at 5000 rpm for 20 min at 4°C. The cell-free supernatant was extracted 179
with an equal volume of ethylacetate and the ethylacetate fraction was carefully taken. 180
Ethylacetate was then evaporated to dryness and the pellet was dissolved in a small 181
volume of ethylacetate. This was analyzed for anthranilate by reversed-phase HPLC 182
(Gilson) using C18 reverse-phase column and UV detector (wavelength; 220 nm) at 0.5 183
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

8
ml/min flow rate in linear gradient elution (solvent A: H2O + 0.1% TFA, solvent B: 184
100% methanol). Commercial anthranilate (DAEJUNG, Korea) was analyzed under the 185
same conditions as a standard. For the quantification of peaks, we used the integration 186
mode of the HPLC-operating software (Unipoint, Gilson, USA). 187
Reporter-based measurement of cyclic diguaniylate (c-di-GMP) For the 188
construction of reporter plasmid (pSKcdrA in Table 1) to measure the c-di-GMP level, 189
the DNA fragment containing cdrA promoter region was prepared by PCR amplification 190
and cloned into BamHI and HindIII site of pQF50, a promoter probing plasmid (29). 191
The cdrA promoter cloned in pSKcdrA includes -492 ~ +1 region relative to start codon 192
of cdrA gene. The PCR primer sequences were 193
GATCGGATCCTTGTTGCTGATCGCGGACCCG (forward) and 194
GACGAAGCTTGAAAATCTCCCTATCTGCGT (reverse). For the measurement of c-195
di-GMP level, P. aeruginosa cells harboring pSKcdrA were grown with or without 0.1 196
mM anthranilate for 20 h. For comparison, the reporter cells were grown with various 197
concentration of sodium nitroprusside (SNP, Sigma) in the same condition. Then, β-198
galactosidase activity that reflects the c-di-GMP level was measured. The level of non-199
treated cells was set to 100% and the relative levels of treated cells were calculated from 200
the β-galactosidase activity. 201
RNA isolation and quantification of transcripts The transcription levels of three 202
major extracellular polysaccharide (EPS) gene clusters of P. aeruginosa were measured 203
by quantitative real-time PCR analysis. The specific primer sets for pslA (PA2231, for 204
Psl operon), pelA (PA3064, for Pel operon), and alg44 (PA3542, for alginate operon) 205
were designed as follows; Psl-forward, CGCTCACGGTGATTATGTTC; Psl-reverse, 206
TACATGAACAACAGCAGGCA; Pel-forward, ACAGCCAGGTAATGGACCTC; Pel-207
reverse, AAGCTGTCCAGGGTATCGAG; alginate-forward, 208
CTACCTTCTCGGCCAACCT; alginate-reverse, GTCAGGGTCCCTTTCATCTG. P. 209
aeruginosa cells were cultivated with or without 0.1 mM anthranilate. The cultures 210
were directly mixed with RNA Protect Bacteria reagent (Qiagen) to stabilize RNA, and 211
lysed by lysozyme treatment and brief sonication. RNA was purified by RNeasy mini 212
columns (Qiagen) as instructed by the manufacturer's manual. Contaminated DNA was 213
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

9
removed by on-column DNase I (Qiagen) digestion and additional RNeasy column 214
purification. The cDNA synthesis was accomplished by HelixCripTM Thermo Reverse 215
Transcriptase Kit (NanoHelix, Korea) as the supplied protocol, in which 10 μg of RNA, 216
100 pmoles of semi-random primer with average G+C content of 75%, and dNTP were 217
used. For the quantitative real-time PCR, 10 ng of cDNA and specific primers were 218
mixed with 20 μl of SYBR Premix Ex Taq (TaKaRa) and analyzed in Thermal CyclerTM 219
Real-time PCR system TP8000 (TaKaRa, Japan). Genomic DNA was used as a standard 220
for quantification and nadB (PA0761) was used as an internal control, as described 221
elsewhere (20). 222
Statistical analysis In order to ensure the significance of the results, the data were 223
statistically analyzed using t-test (two-sample assuming equal variances) in MS office 224
Excel (Microsoft, USA). Any P-value lower than 0.05 was considered significant. 225
Results 226
Biofilm-enhancing effect of indole on P. aeruginosa is QS-independent. 227
Indole was reported to both inhibit the PQS production and enhance the biofilm 228
formation in P. aeruginosa (10, 13). Since both PQS production and biofilm formation 229
were positively regulated by QS systems (Fig. S1), we investigated whether the indole 230
effect on the biofilm formation is exerted through QS regulation. When we treated a QS 231
mutant strain, MW1 (lasI-, rhlI-), which is deficient in the production of QS signals, 232
with 0.4 mM indole, the biofilm formation of MW1 was still enhanced by the indole 233
treatment (Fig. 1). When we investigated the influence of indole on the activities of QS 234
regulators, LasR and QscR, by using reporter fusions, none was significantly affected 235
by 0.4 mM indole treatment (Fig. S2AB). Indole did not affect cell growth at this 236
concentration (data not shown). These results demonstrate that the indole effect on 237
biofilm formation is exerted independently of the acyl-HSL-based QS systems. 238
Anthranilate effect differs depending on the developmental stage and the presence 239
of shear force. 240
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

10
We investigated the anthranilate effect on the biofilm formation of P. aeruginosa. 241
When we measured the static biofilm formation with anthranilate treatment in shear 242
force-free conditions, anthranilate appeared to enhance the biofilm formation like indole 243
(Fig. 2). When the biofilm was co-treated with anthranilate and indole together, static 244
biofilm assay showed the additive enhancement of biofilm formation (Fig. 2). 245
To better investigate this collaborative effect of indole and anthranilate, the biofilm 246
formation was measured in the presence of shear force in the flow-cell system. We first 247
investigated the biofilm formation in the early stage of biofilm development and found 248
that anthranilate and indole were each able to facilitate the attachment of cells. But 249
interestingly, co-treatment of both failed to additively enhance the cell attachment (Fig. 250
3A). Instead, anthranilate appeared to antagonize the biofilm-enhancing effect of indole 251
(Fig. 3A). When we further grew the biofilm and investigated the anthranilate effect at 252
the late stage of biofilm development, anthranilate hindered the maturation of the 253
biofilm structure, finally making a flat biofilm (Fig. 3B). As previously reported, indole 254
treatment enhanced and advanced the maturation of biofilm and facilitated the 255
dispersion of biofilm in the late stage, leaving the “central void form” at the center of a 256
mushroom structure (Fig. 3B). Anthranilate was also able to dramatically antagonize 257
this indole effect in the late stage of biofilm development (Fig. 3B). Anthranilate did not 258
affect cell growth at the concentration used in this experiment (data not shown). 259
Taken together, our results demonstrate that anthranilate has a complex effect on 260
biofilm formation dependent on the presence of shear force and the developmental stage 261
of the biofilm. Without shear force, anthranilate was able to enhance the biofilm 262
formation and this effect was additive with indole. But with shear force, although 263
anthranilate facilitated the initial colonization in the early stage, it ultimately hindered 264
the maturation of the biofilm and antagonized the indole effect. 265
Anthranilate deteriorates the biofilm structure. 266
Since the biofilm inhibition by anthranilate occurred in the presence of shear force, 267
we hypothesized that anthranilate weakens the biofilm and destabilizes the structure. To 268
better see the influence of anthranilate and indole on the biofilm, we used calcofluor 269
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

11
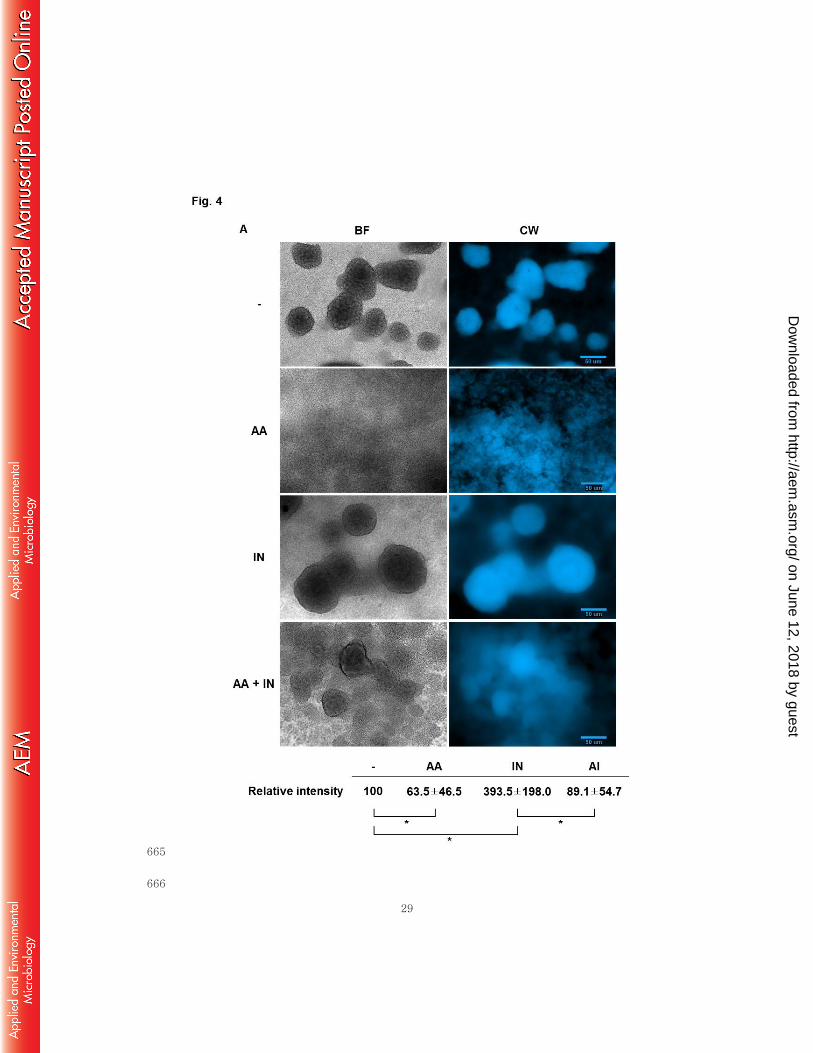
white-staining of extracellular polysaccharides (EPS) in the biofilms and simple bright 270
field microscopy. Calcofluor white is a fluorescent dye that strongly binds to 271
extracellular structural polysaccharides, specifically to β-1,4 linkage of glycosidic bonds 272
(30). Both bright field and EPS staining microscopy showed that the anthranilate-treated 273
biofilm contained more deteriorated structure, implying that anthranilate likely weakens 274
the biofilm and crumbles the structure in the presence of shear force (Fig. 4A). In 275
contrast, indole enhanced the biofilm structure, forming bigger mushroom-bodies. The 276
anthranilate treatment seemed to cause biofilm cells to fall away to the interstitial space 277
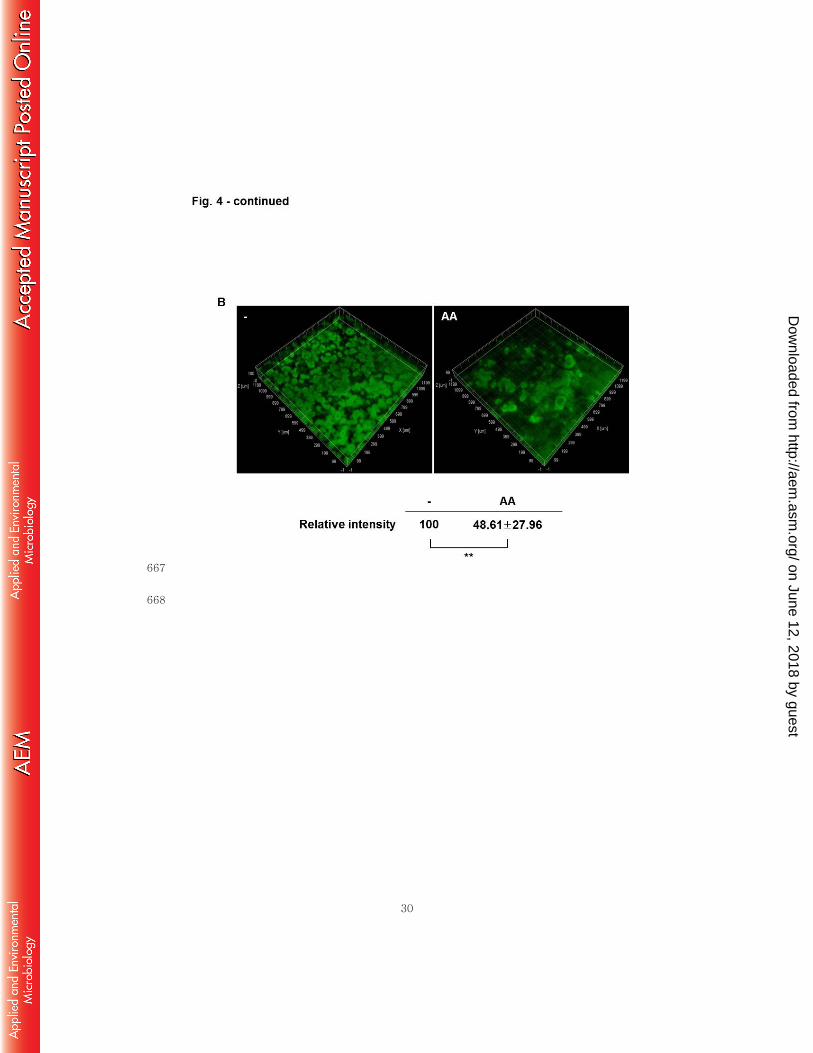
and even crumbled the indole-enhanced biofilm (Fig. 4A). In order to confirm this 278
biofilm-deteriorating effect of anthranilate, we treated pre-formed biofilm with 279
anthranilate. As shown in Fig. 4B, the pre-formed biofilm was destroyed by the 280
anthranilate treatment. These results also demonstrate that anthranilate deteriorates the 281
biofilm structure. 282
The anthranilate effect is also QS-independent. 283
We investigated whether the anthranilate effects occurred in a QS-independent 284
manner like indole. We treated the QS mutant strain, MW1, with anthranilate and 285
observed the biofilm formation using fluorescence microscopy. The facilitation of the 286
initial attachment by the anthranilate treatment was still observed in the QS mutant 287
strain as in the wild type (Fig. 5A). Anthranilate also antagonized the biofilm-enhancing 288
effect of indole in the QS mutant (Fig. 5B). These results demonstrate that the 289
anthranilate effects are exerted in a QS-independent manner like the indole effect. We 290
note that the attachment-facilitating effect of anthranilate in the early stage was more 291
dramatic in the QS mutant than in wild type. 292
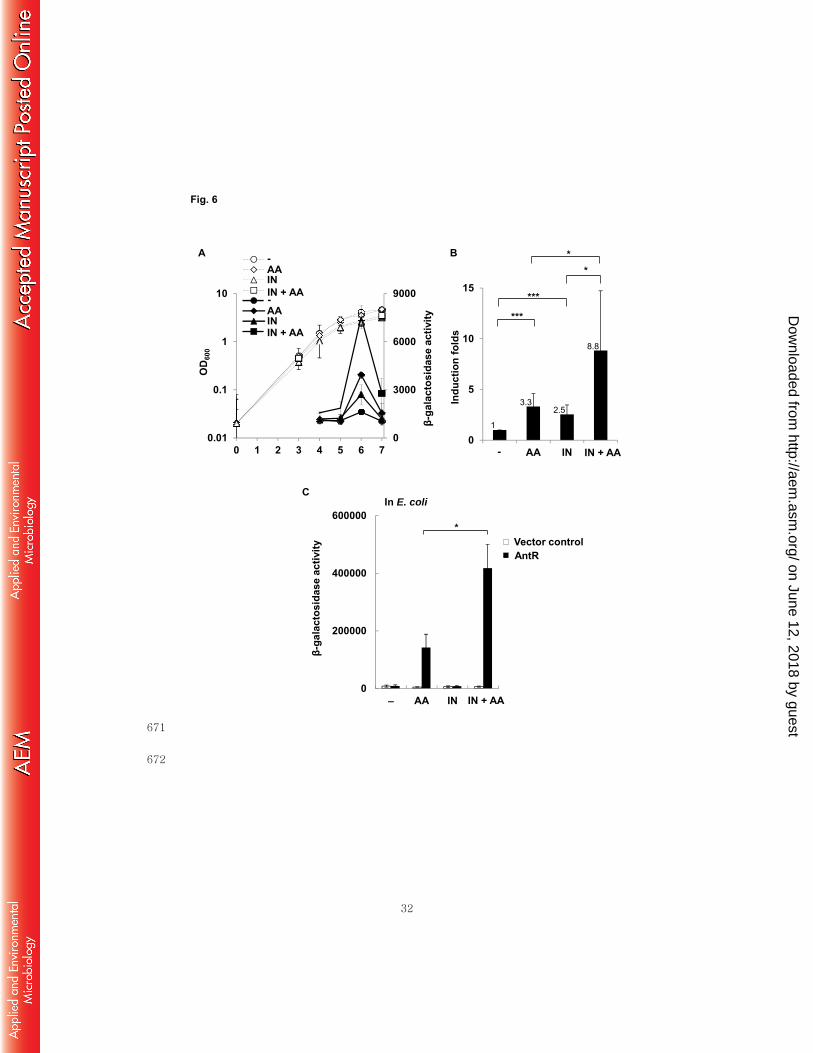
Indole boosts the anthranilate-degrading activity. 293
Anthranilate is an activating cofactor of AntR, a transcriptional regulator of the 294
antABC operon encoding the anthranilate dioxygenase complex that functions to 295
degrade anthranilate through the TCA cycle (22, 31). So, anthranilate treatment induces 296
the expression of antABC through direct activation of AntR (22, 31). A recent 297
microarray study showed that indole treatment also increased the transcription of the 298
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

12
antABC operon (10). To examine the effects of anthranilate and indole on the expression 299
of the antABC operon, P. aeruginosa cells harboring antAp-lacZ fusion were treated 300
with either anthranilate or indole, or both. The measurement of β-galactosidase activity 301
showed that antA expression was collaboratively augmented by co-treatment of 302
anthranilate and indole in P. aeruginosa (Fig. 6A). This collaborative induction 303
appeared significant and synergistic (Fig. 6B). While it is well documented that 304
anthranilate is a cofactor of AntR to induce antA transcription (22, 31), indole was never 305
reported to activate AntR. To investigate whether indole can activate AntR, we carried 306
out the E. coli reconstitution analysis using two compatible plasmids, the AntR-307
expressing plasmid and antAp-lacZ fusion plasmid, as previously described (23, 31). 308
Interestingly, although indole alone did not induce antA expression, it boosted the 309
induction of antA by anthranilate (Fig. 6C). This is consistent with the result of the P. 310
aeruginosa experiment in Fig. 6AB, because indole also synergistically induced antA 311
expression when co-treated with anthranilate. The slight activation of antA by single 312
treatment with indole in P. aeruginosa is likely due to endogenously produced 313
anthranilate. 314
We hypothesized that the extra-enhanced expression of antABC by indole might 315
accelerate the degradation of anthranilate and reduce the local concentration of 316
anthranilate within the biofilm, which might cause the biofilm enhancement by indole 317
treatment. To prove this hypothesis, we measured the anthranilate level in P. aeruginosa 318
culture supernatant with or without the indole treatment. The result showed that the 319
anthranilate level reduced to half by the indole treatment (Fig. 7), supporting our 320
hypothesis. 321
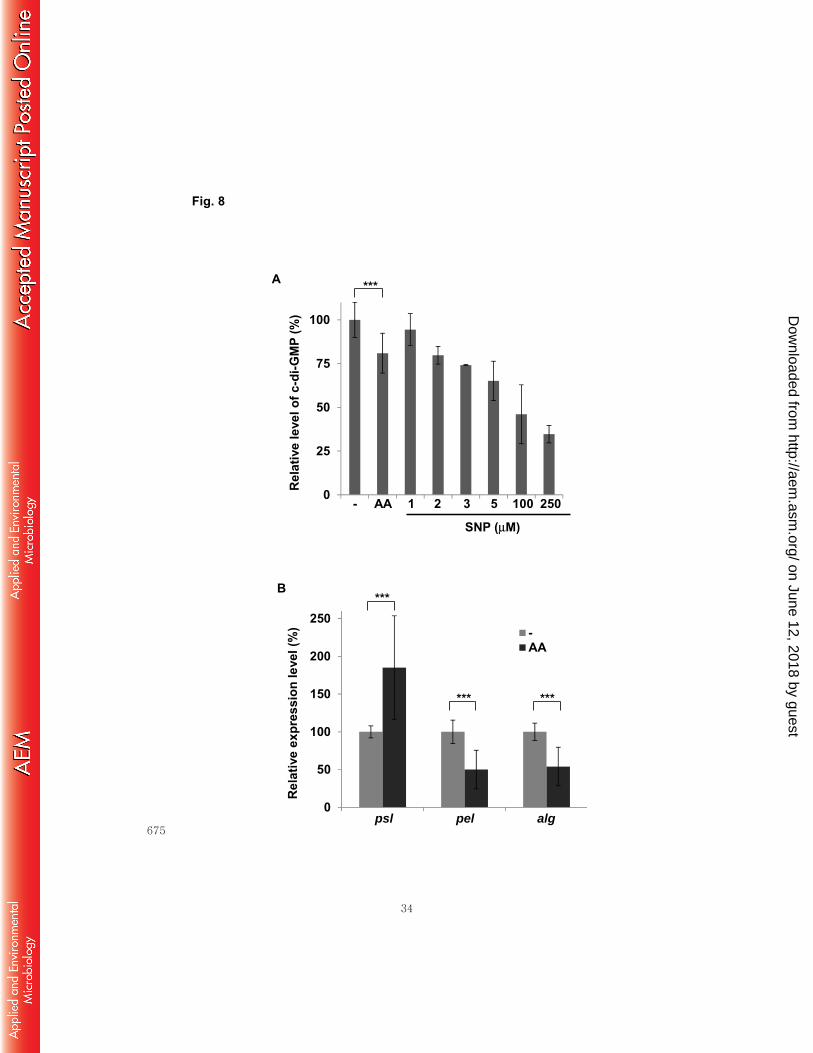
Anthranilate affects the c-di-GMP level and transcription of EPS operons. 322
We tried to address how anthranilate deteriorates the biofilm structure. We first tested 323
whether the treatment of anthranilate could change the hydrophobicity of cell surface, 324
but there was no significant change, indicating that it is not the cause (Fig. S3). Next, 325
we investigated the level of an intracellular signaling molecule, c-di-GMP that plays an 326
important role in controlling biofilm formation in many Gram-negative bacteria (32). 327
We constructed cdrA-lacZ fusion for gauging c-di-GMP level, since cdrA gene 328
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

13
(PA4625) encoding a large adhesin has been used for c-di-GMP-responsive reporter 329
(33). cdrA is highly upregulated in response to increased level of c-di-GMP and 330
downregulated by decreased level of c-di-GMP (34). When we measured the β-331
galactosidase activity that reflects the intracellular c-di-GMP level, the anthranilate-332
treated cells showed decreased level by 20% (Fig. 8A). Sodium nitroprusside (SNP), a 333
nitrogen monoxide (NO)-releasing agent has been reported to decrease the intracellular 334
c-di-GMP level (35). Previous study using LC-MS-MS analysis reported about 44% 335
reduction of intracellular c-di-GMP by 5 μM SNP treatment (35) and our result showed 336
35% reduction by same concentration of SNP (Fig. 8A). When the reporter cells were 337
treated with various concentration of SNP for comparison, it demonstrated that the 0.1 338
mM anthranilate treatment had a comparable effect to 2 μM SNP treatment (Fig. 8A). 339
The c-di-GMP level regulates the production of EPSs in P. aeruginosa. In order to 340
find a clue about what happens to the biofilm matrix with anthranilate treatment, we 341
measured the transcription levels of the EPS biosynthetic operons. P. aeruginosa has 342
three major EPSs in the biofilm matrix; Psl, Pel, and alginate (36). Our result showed 343
that the anthranilate treatment increased the transcription of Psl operon by 85%, but 344
decreased the transcriptions of Pel and alginate operons by 50% and 54%, respectively 345
(Fig. 8B). 346
Discussion 347
In this study, we investigated the anthranilate effects on P. aeruginosa biofilm 348
formation. Our results demonstrated followings: 1) anthranilate causes the 349
destabilization of biofilm, deteriorating the mushroom structure; 2) anthranilate affects 350
biofilm formation differently depending on the developmental stage and the presence of 351
shear force; 3) the effects of anthranilate and indole on biofilm formation do not involve 352
QS-regulation; 4) indole boosts the anthranilate degrading activity. 353
Anthranilate facilitated the attachment of cells to the surface in the early stage of 354
biofilm development, but it seemed to deteriorate the mushroom structure in the late 355
stage. Shear force played an important role in this effect: Without shear force, 356
anthranilate treatment increased the total amount of biofilm even with long cultivation 357
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

14
(Fig. 2), but with shear force, the anthranilate treatment crumbled the biofilm (Fig. 3B; 358
Fig. 4AB). Apparently, the shear force holds the key to determine the final form of the 359
biofilm structure between the two anthranilate effects: increasing the cell attachment to 360
surface or weakening the mushroom structure. Strong shear force will crumble and 361
wash away the weakened biofilm. Still, the cells falling from the weakened biofilm may 362
be retained in the interstitial space around the biofilm stalks if the shear force is not 363
strong enough to wash the cells off, because anthranilate facilitates the surface 364
attachment while weakening the biofilm structure. 365
Then, how anthranilate deteriorates and weakens the biofilm structure? Multiple 366
studies have suggested that Psl is important in initial attachment to abiotic and biotic 367
surfaces (32, 37), Pel primarily plays a role after surface attachment (38), and alginate is 368
associated with chronic stages of biofilm-mediated infection (39). Therefore, our result 369
about the expression of EPS operons implies that the anthranilate treatment may 370
facilitate initial attachment by increasing the Psl production, but weaken the biofilm 371
structure at late stage by decreasing the Pel and alginate production. This is consistent 372
with our observation about the anthranilate effect on biofilm formation. Recently, it was 373
reported that the mucoid strains recovered from chronic pulmonary infections in CF 374
patients express elevated level of alginate and reduced levels of Psl, and the 375
transcription factor responsible for this inverse regulation of alginate and Psl operons is 376
AmrZ (40). Since AmrZ positively regulates the expression of alginate operon, but 377
represses the transcription of Psl operon, it was suggested that AmrZ may mediate 378
transition of P. aeruginosa biofilm infection from colonizing to chronic stages. 379
Interestingly, the anthranilate effect on the transcription of Psl and alginate operons is 380
opposite to the AmrZ function. 381
Among three major EPSs of P. aeruginosa, the production of Pel is positively 382
regulated by c-di-GMP at the transcriptional and allosteric level in P. aeruginosa. FleQ, 383
a transcriptional repressor derepresses the transcription of pel operon when intracellular 384
c-di-GMP levels are high (41). c-di-GMP also binds to PelD, an inner membrane protein 385
and enhances Pel production (36). The production of alginate is positively regulated c-386
di-GMP at allosteric level. Like PelD, the c-di-GMP binding to Alg44, an inner 387
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

15
membrane protein is required for alginate production (36). Since the anthranilate 388
treatment decreased both c-di-GMP and the transcription of Pel and alginate operons 389
(Fig. 8AB), final production of Pel and alginate should decrease by anthranilate 390
treatment at both transcriptional and allosteric levels. 391
Anthranilate is naturally produced by P. aeruginosa. It is produced at very low level 392
in P. aeruginosa culture medium during exponential growth, but rapidly accumulates at 393
late stationary phase up to 0.05 mM (23). The accumulation of anthranilate activates 394
AntR that activates the expression of antABC operon to degrade anthranilate (23, 31). 395
Our results imply that anthranilate should be degraded for P. aeruginosa to form mature 396
biofilms. Actually, a recent study showed that P. aeruginosa cells in biofilm enhanced 397
anthranilate-degrading activity (15). Another study showed that tryptophan had an 398
inhibitory effect on the biofilm formation by P. aeruginosa (11). This also supports our 399
results in that P. aeruginosa degrades tryptophan to anthranilate. The anthranilate 400
concentration will goes up with the supplement of tryptophan to culture medium, which 401
can deteriorates biofilm. A study actually showed that the anthranilate concentration 402
increased with the tryptophan supplement (42). 403
Indole has recently received much attention due to its diverse biological roles in 404
bacterial physiology and its potential to modulate biofilm formation (14). In P. 405
aeruginosa, indole enhances biofilm formation, but it has the opposite effect in E. coli, 406
repressing the biofilm formation (13). Since there are generally many indole-producing 407
bacteria in environmental habitat where P. aeruginosa lives (e.g., in gut where E. coli is 408
abundant), occasional input of tryptophan-rich nutrients may increase both indole and 409
anthranilate around P. aeruginosa. Our study suggests that anthranilate produced by P. 410
aeruginosa can antagonize the indole effect on biofilm. So, the indole effect on P. 411
aeruginosa may be finely tuned in the balance between endogenously produced 412
anthranilate and exogenous indole. 413
In addition, our results suggest that the biofilm-enhancing effect of indole might be 414
exerted through the reduction of anthranilate by indole treatment, as shown in Fig. 7. 415
The extra-induction of antABC by indole may accelerate the degradation of anthranilate, 416
which can remove the biofilm-deteriorating effect of anthranilate by reducing the local 417
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

16
concentration of anthranilate within the biofilm. This may result in the enhancement of 418
biofilm formation. This postulation is also supported by the study showing that P. 419
aeruginosa cells in biofilm have enhanced anthranilate-degrading activity (15). 420
Acknowledgement 421
This research was supported by Basic Science Research Program through the 422
National Research Foundation of Korea (NRF) funded by the Ministry of Education 423
(grant number: 2013R1A1A2012220). This research was also supported by Basic 424
Science Research Program through the National Research Foundation of Korea (NRF) 425
funded by the Ministry of Education, Science and Technology (grant number: 2010-426
0006622) 427
428
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

17
Reference 429
1. Bassler BL, Losick R. 2006. Bacterially speaking. Cell 125:237-246. 430
2. Parsek MR, Greenberg EP. 2005. Sociomicrobiology: the connections between 431
quorum sensing and biofilms. Trends Microbiol 13:27-33. 432
3. Ymele-Leki P, Ross JM. 2007. Erosion from Staphylococcus aureus biofilms 433
grown under physiologically relevant fluid shear forces yields bacterial cells 434
with reduced avidity to collagen. Appl Environ Microbiol 73:1834-1841. 435
4. Costerton JW, Stewart PS, Greenberg EP. 1999. Bacterial biofilms: a common 436
cause of persistent infections. Science 284:1318-1322. 437
5. Hancock RE, Speert DP. 2000. Antibiotic resistance in Pseudomonas aeruginosa: 438
mechanisms and impact on treatment. Drug resistance updates : reviews and 439
commentaries in antimicrobial and anticancer chemotherapy 3:247-255. 440
6. Willcox MD, Zhu H, Conibear TC, Hume EB, Givskov M, Kjelleberg S, Rice SA. 441
2008. Role of quorum sensing by Pseudomonas aeruginosa in microbial keratitis 442
and cystic fibrosis. Microbiology 154:2184-2194. 443
7. Huq A, Whitehouse CA, Grim CJ, Alam M, Colwell RR. 2008. Biofilms in water, its 444
role and impact in human disease transmission. Curr Opin Biotechnol 19:244-445
247. 446
8. Kumar CG, Anand SK. 1998. Significance of microbial biofilms in food industry: a 447
review. International journal of food microbiology 42:9-27. 448
9. Kurnasov O, Jablonski L, Polanuyer B, Dorrestein P, Begley T, Osterman A. 2003. 449
Aerobic tryptophan degradation pathway in bacteria: novel kynurenine 450
formamidase. FEMS Microbiol Lett 227:219-227. 451
10. Lee J, Attila C, Cirillo SL, Cirillo JD, Wood TK. 2009. Indole and 7-hydroxyindole 452
diminish Pseudomonas aeruginosa virulence. Microb Biotechnol 2:75-90. 453
11. Brandenburg KS, Rodriguez KJ, McAnulty JF, Murphy CJ, Abbott NL, Schurr MJ, 454
Czuprynski CJ. 2013. Tryptophan inhibits biofilm formation by Pseudomonas 455
aeruginosa. Antimicrob Agents Chemother 57:1921-1925. 456
12. Shimazaki J, Furukawa S, Ogihara H, Morinaga Y. 2012. L-Tryptophan prevents 457
Escherichia coli biofilm formation and triggers biofilm degradation. Biochem 458
Biophys Res Commun 419:715-718. 459
13. Lee J, Jayaraman A, Wood TK. 2007. Indole is an inter-species biofilm signal 460
mediated by SdiA. BMC microbiology 7:42. 461
14. Lee JH, Lee J. 2010. Indole as an intercellular signal in microbial communities. 462
FEMS microbiology reviews 34:426-444. 463
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

18
15. Costaglioli P, Barthe C, Claverol S, Brozel VS, Perrot M, Crouzet M, Bonneu M, 464
Garbay B, Vilain S. 2012. Evidence for the involvement of the anthranilate 465
degradation pathway in Pseudomonas aeruginosa biofilm formation. 466
Microbiologyopen 1:326-339. 467
16. Davies DG, Parsek MR, Pearson JP, Iglewski BH, Costerton JW, Greenberg EP. 468
1998. The involvement of cell-to-cell signals in the development of a bacterial 469
biofilm. Science 280:295-298. 470
17. Bassler BL. 2002. Small talk. Cell-to-cell communication in bacteria. Cell 471
109:421-424. 472
18. Fuqua C, Greenberg EP. 2002. Listening in on bacteria: acyl-homoserine lactone 473
signalling. Nat Rev Mol Cell Biol 3:685-695. 474
19. Park SJ, Liu HB, Park S, Lee JH. 2013. Modulation of QscR, a quorum sensing 475
receptor of Pseudomonas aeruginosa, by truncation of a signal binding domain. 476
Research in microbiology 164:375-381. 477
20. Ha C, Park SJ, Im SJ, Lee JH. 2012. Interspecies signaling through QscR, a 478
quorum receptor of Pseudomonas aeruginosa. Mol Cells 33:53-59. 479
21. Diggle SP, Matthijs S, Wright VJ, Fletcher MP, Chhabra SR, Lamont IL, Kong X, 480
Hider RC, Cornelis P, Camara M, Williams P. 2007. The Pseudomonas aeruginosa 481
4-quinolone signal molecules HHQ and PQS play multifunctional roles in quorum 482
sensing and iron entrapment. Chem Biol 14:87-96. 483
22. Oglesby AG, Farrow JM, 3rd, Lee JH, Tomaras AP, Greenberg EP, Pesci EC, Vasil 484
ML. 2008. The influence of iron on Pseudomonas aeruginosa physiology: a 485
regulatory link between iron and quorum sensing. J Biol Chem 283:15558-15567. 486
23. Choi Y, Park HY, Park SJ, Kim SK, Ha C, Im SJ, Lee JH. 2011. Growth phase-487
differential quorum sensing regulation of anthranilate metabolism in 488
Pseudomonas aeruginosa. Mol Cells 32:57-65. 489
24. Lee JH, Lequette Y, Greenberg EP. 2006. Activity of purified QscR, a 490
Pseudomonas aeruginosa orphan quorum-sensing transcription factor. Mol 491
Microbiol 59:602-609. 492
25. O'Toole GA, Kolter R. 1998. Initiation of biofilm formation in Pseudomonas 493
fluorescens WCS365 proceeds via multiple, convergent signalling pathways: a 494
genetic analysis. Mol Microbiol 28:449-461. 495
26. Lee KJ, Kim JA, Hwang W, Park SJ, Lee KH. 2013. Role of capsular 496
polysaccharide (CPS) in biofilm formation and regulation of CPS production by 497
quorum-sensing in Vibrio vulnificus. Mol Microbiol 90:841-857. 498
27. Xu KD, Stewart PS, Xia F, Huang CT, McFeters GA. 1998. Spatial physiological 499
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

19
heterogeneity in Pseudomonas aeruginosa biofilm is determined by oxygen 500
availability. Appl Environ Microbiol 64:4035-4039. 501
28. Balebona MC, Morinigo MA, Borrego JJ. 2001. Hydrophobicity and adhesion to 502
fish cells and mucus of Vibrio strains isolated from infected fish. International 503
microbiology : the official journal of the Spanish Society for Microbiology 4:21-504
26. 505
29. Farinha MA, Kropinski AM. 1990. Construction of broad-host-range plasmid 506
vectors for easy visible selection and analysis of promoters. J Bacteriol 507
172:3496-3499. 508
30. Herth W, Schnepf E. 1980. The fluorochrome, calcofluor white, binds oriented to 509
structural polysaccharide fibrils. Protoplasma 105:129-133. 510
31. Kim SK, Im SJ, Yeom DH, Lee JH. 2012. AntR-mediated bidirectional activation 511
of antA and antR, anthranilate degradative genes in Pseudomonas aeruginosa. 512
Gene 505:146-152. 513
32. Wei Q, Ma LZ. 2013. Biofilm matrix and its regulation in Pseudomonas 514
aeruginosa. Int J Mol Sci 14:20983-21005. 515
33. Rybtke MT, Borlee BR, Murakami K, Irie Y, Hentzer M, Nielsen TE, Givskov M, 516
Parsek MR, Tolker-Nielsen T. 2012. Fluorescence-based reporter for gauging 517
cyclic di-GMP levels in Pseudomonas aeruginosa. Appl Environ Microbiol 518
78:5060-5069. 519
34. Borlee BR, Goldman AD, Murakami K, Samudrala R, Wozniak DJ, Parsek MR. 2010. 520
Pseudomonas aeruginosa uses a cyclic-di-GMP-regulated adhesin to reinforce 521
the biofilm extracellular matrix. Mol Microbiol 75:827-842. 522
35. Barraud N, Schleheck D, Klebensberger J, Webb JS, Hassett DJ, Rice SA, 523
Kjelleberg S. 2009. Nitric oxide signaling in Pseudomonas aeruginosa biofilms 524
mediates phosphodiesterase activity, decreased cyclic di-GMP levels, and 525
enhanced dispersal. J Bacteriol 191:7333-7342. 526
36. Franklin MJ, Nivens DE, Weadge JT, Howell PL. 2011. Biosynthesis of the 527
Pseudomonas aeruginosa Extracellular Polysaccharides, Alginate, Pel, and Psl. 528
Frontiers in microbiology 2:167. 529
37. Ma L, Jackson KD, Landry RM, Parsek MR, Wozniak DJ. 2006. Analysis of 530
Pseudomonas aeruginosa conditional psl variants reveals roles for the psl 531
polysaccharide in adhesion and maintaining biofilm structure postattachment. J 532
Bacteriol 188:8213-8221. 533
38. Vasseur P, Vallet-Gely I, Soscia C, Genin S, Filloux A. 2005. The pel genes of 534
the Pseudomonas aeruginosa PAK strain are involved at early and late stages of 535
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

20
biofilm formation. Microbiology 151:985-997. 536
39. Schurr MJ. 2013. Which bacterial biofilm exopolysaccharide is preferred, Psl or 537
alginate? J Bacteriol 195:1623-1626. 538
40. Jones CJ, Ryder CR, Mann EE, Wozniak DJ. 2013. AmrZ modulates Pseudomonas 539
aeruginosa biofilm architecture by directly repressing transcription of the psl 540
operon. J Bacteriol 195:1637-1644. 541
41. Hickman JW, Harwood CS. 2008. Identification of FleQ from Pseudomonas 542
aeruginosa as a c-di-GMP-responsive transcription factor. Mol Microbiol 543
69:376-389. 544
42. Farrow JM, 3rd, Pesci EC. 2007. Two distinct pathways supply anthranilate as a 545
precursor of the Pseudomonas quinolone signal. J Bacteriol 189:3425-3433. 546
43. Pearson JP, Pesci EC, Iglewski BH. 1997. Roles of Pseudomonas aeruginosa las 547
and rhl quorum-sensing systems in control of elastase and rhamnolipid 548
biosynthesis genes. J Bacteriol 179:5756-5767. 549
44. Whiteley M, Lee KM, Greenberg EP. 1999. Identification of genes controlled by 550
quorum sensing in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 551
96:13904-13909. 552
45. Sambrook J, E. F. Fritsch, and T. Maniatis 1989. Molecular cloning: a laboratory 553
manual, 2nd ed ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 554
NY. 555
46. Walters MC, 3rd, Roe F, Bugnicourt A, Franklin MJ, Stewart PS. 2003. 556
Contributions of antibiotic penetration, oxygen limitation, and low metabolic 557
activity to tolerance of Pseudomonas aeruginosa biofilms to ciprofloxacin and 558
tobramycin. Antimicrobial agents and chemotherapy 47:317-323. 559
47. Newman JR, Fuqua C. 1999. Broad-host-range expression vectors that carry the 560
L-arabinose-inducible Escherichia coli araBAD promoter and the araC regulator. 561
Gene 227:197-203. 562
48. Chugani SA, Whiteley M, Lee KM, D'Argenio D, Manoil C, Greenberg EP. 2001. 563
QscR, a modulator of quorum-sensing signal synthesis and virulence in 564
Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 98:2752-2757. 565
566
567
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

21
Legends 568
Fig. 1. Biofilm-enhancing effect of indole is QS-independent. PAO1 (wild type, WT) 569
and MW1 (QS mutant; lasI- rhlI-) cells harboring pAB1 were grown in the drip-flow 570
chamber to form biofilms with indole treatment. WT and MW1 cells in the left column 571
were treated with dimethyl sulfoxide (DMSO) as a buffer control, because indole was 572
dissolved in DMSO before use. Biofilms were imaged by green fluorescence using 573
CLSM at 40 h after inoculation. IN, 0.4 mM indole. 574
Fig. 2. Anthranilate enhances static biofilm formation. Static biofilm assay was 575
carried out as described in Materials and Methods. Wild type PAO1 cells were grown in 576
96-well plates and the measurement was taken at 24 h after inoculation. -, DMSO as a 577
buffer control; IN, 0.4 mM indole; AA, 0.1 mM anthranilate; AA + IN, 0.1 mM 578
anthranilate plus 0.4 mM indole. P-value, *< 0.05, ***< 0.005 579
Fig. 3. Shear force differentiates the anthranilate effect according to developmental 580
stage. A, initial attachment of PAO1 cells was observed in the early stage of biofilm 581
development (15 h after inoculation) with indole or anthranilate treatment. The 582
fluorescence of images was quantified and presented below as relative intensity. B, P. 583
aeruginosa biofilm was observed at late stage of biofilm development (4 d after 584
inoculation). GFP-expressing PAO1 cells that harbor pAB1 were used in these 585
experiments. -, DMSO as a buffer control; IN, 0.4 mM indole; AA, 0.1 mM 586
anthranilate; AA + IN, 0.1 mM anthranilate plus 0.4 mM indole. P-value, *< 0.05, **< 587
0.01 588
Fig. 4. Anthranilate deteriorates the biofilm structure. A, biofilm of PAO1 cells was 589
formed in flow-cell system with treatment of anthranilate or indole. At 5 d after 590
inoculation, biofilms were directly observed on bright field microscope (BF) or stained 591
with calcofluor white and imaged by fluorescence microscopy (CW). -, DMSO as a 592
buffer control; IN, 0.4 mM indole; AA, 0.1 mM anthranilate; AA + IN, 0.1 mM 593
anthranilate plus 0.4 mM indole. The fluorescence intensity was quantified and 594
presented below as relative intensity. B, biofilms of GFP-expressing PAO1 cells were 595
formed without anthranilate for 3 d in the flow-cell system and this pre-formed biofilm 596
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

22
was then treated with 0.1 mM anthranilate for 24 h (AA). As a control, the pre-formed 597
biofilm was further incubated without anthranilate under the same conditions (-). The 598
fluorescence was quantified and presented below. P-value, *< 0.05, **< 0.01 599
Fig. 5. The anthranilate effect is QS-independent. A, MW1 harboring pAB1 was 600
used for the biofilm formation. The biofilms of GFP-expressing MW1 cells were grown 601
with 0.1 mM anthranilate (AA) or 0.4 mM indole (IN) in flow-cell system for 24 h. 602
Biofilms were observed on fluorescence microscope by green fluorescence. The 603
intensity of fluorescence was quantified and presented below. B, the biofilms of GFP-604
expressing MW1 cells were grown in a flow-cell system for 3 d with 0.4 mM indole 605
(IN) or 0.4 mM indole plus 0.1 mM anthranilate (AA + IN). Fluorescence was 606
quantified and presented below. P-value, ***< 0.005 607
Fig. 6. Indole activates antABC expression synergistically with anthranilate. A, 608
AntR activity in P. aeruginosa was measured through growth using the reporter strain 609
(pJL201) with treatment of indole or anthranilate, or both. Solid lines indicate β-610
galactosidase activity and dotted lines, cell growth (OD600). B, induction of AntR 611
activity at stationary phase was presented as fold-induction with statistical significance. 612
C, AntR-expressing plasmid (pJN105A) and pJL201 were co-transformed into E. coli. 613
The transformed E. coli cells were grown to OD600 = 0.3 and treated with indole or 614
anthranilate for 2 h; β-galactosidase activity was then measured. -, buffer control; IN, 615
0.4 mM indole; AA, 0.1 mM anthranilate; AA + IN, 0.1 mM anthranilate plus 0.4 mM 616
indole. P-value, *< 0.05, ***< 0.005 617
Fig. 7. Anthranilate level in culture supernatant of P. aeruginosa was reduced by 618
indole treatment. P. aeruginosa cells were cultivated to OD600 = 3 with or without 0.4 619
mM indole, and the anthranilate level in the culture supernatant was measured by HPLC 620
analysis. P-value; ***< 0.005. 621
Fig. 8. Anthranilate effects on the levels of c-di-GMP and expression of EPS 622
biosynthetic genes. A, P. aeruginosa cells harboring pSKcdrA (cdrA-lacZ fusion) were 623
grown with or without 0.1 mM anthranilate for 20 h and β-galactosidase activity that 624
reflects the c-di-GMP level was measured. For comparison, the reporter cells were 625
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

23
grown with various concentration of SNP in the same condition. The c-di-GMP level of 626
non-treated cells was set to 100% and the relative c-di-GMP levels of treated cells were 627
presented from β-galactosidase activity. B, P. aeruginosa cells were cultivated with or 628
without 0.1 mM anthranilate and the transcripts of Psl, Pel, and alginate biosynthetic 629
operons were measured by quantitative real-time PCR analysis. The transcription levels 630
were presented relatively to the transcription level of non-treated cells. P-value; ***< 631
0.005. 632
Supporting information 633
Additional Supporting Information may be found in the online version of this article at 634
the publisher’s website: 635
Fig. S1. Physiological relationship among anthranilate, indole, QS, and biofilm 636
formation in P. aeruginosa. Anthranilate is a precursor of PQS and tryptophan 637
biosynthesis, as well as a degradation product of tryptophan in P. aeruginosa. Indole 638
that is not produced by P. aeruginosa inhibits PQS synthesis, enhances biofilm 639
formation, and reduces virulence in P. aeruginosa. QS regulation positively regulates 640
both PQS production and biofilm formation. Anthranilate is an activating cofactor of 641
AntR, a transcriptional regulator of antABC operon, which encodes the anthranilate 642
dioxygenase complex that functions to degrade anthranilate through the TCA cycle. 643
AntR therefore activates anthranilate degradation by inducing the antABC operon. 644
Fig. S2. Indole has no effect on the activity of LasR and QscR, the QS regulators. 645
The activities of LasR (A) and QscR (B) were measured with 0.4 mM indole treatment 646
using the reporters, lasIp-lacZ (pSC11) and PA1897p-lacZ (pJL101). Solid lines 647
indicate β-galactosidase activity and dotted lines, cell growth (OD600). 648
Fig. S3. Hydrophobicity of cells was not influenced by anthranilate or indole 649
treatment P. aeruginosa cells were cultured overnight in LB broth with 0.1 mM 650
anthranilate or 0.4 mM indole or both, and the cell surface hydrophobicity was 651
measured by cell adhesion to organic solvent. The hydrophobicity was presented by 652
percentage of adhesion to organic solvent as described in Materials and Methods.653
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

24
Table 1. Bacterial strains and plasmids used in this study. 654
Name Genotype References
P. aeruginosa
PAO1 Wild type P. aeruginosa (43)
MW1 lasI-,rhlI- double mutant of PAO1 (44)
E. coli
DH5α supE44 ΔlacU169 (ф80 lacZΔM15) hsdR17 recA1
endA1 gyrA96 thi‐1 relA1
(45)
Plasmids
pAB1 gfp-mut2 gene in pMF54, ApR (46)
pQF50 Broad-host-range lacZ fusion plasmid, ApR (29)
pJN105 araC-pBAD cassette cloned in pBBR1MCS-5,
GmR
(47)
pJN105A Plasmid for AntR overexpression. antR orf in
pJN105, GmR
(23)
pSC11 lasIp-lacZ reporter in pQF50, ApR (48)
pJL101 PA1897p-lacZ reporter in pQF50, ApR (24)
pJL201 antAp-lacZ fusion in pQF50, ApR (23)
pSKcdrA cdrA-lacZ fusion in pQF50, ApR This study
GmR, gentamicin-resistance; ApR, ampicillin and carbenicillin resistance 655
656
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

25
WT WT, IN
MW1 MW1, IN
Fig. 1
657
658
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

26
0
2
4
6
Fig. 2
A 600
/OD
600
- IN AA + INAA
***
***
****
659
660
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
32
0
3000
6000
9000
0.01
0.1
1
10
0 1 2 3 4 5 6 7
xAAINAA INxAAINAA IN
0
5
10
15
x AA IN AA IN
OD 6
00
β-ga
lact
osid
ase
activ
ity
Indu
ctio
n fo
lds
***
*
***
Fig. 6
A B*
IN + AA
IN + AA
-
-
-
0
200000
400000
600000
– AA IN AI
β-ga
lact
osid
ase
activ
ity
In E. coli
IN + AA
IN + AA
*
C
Vector controlAntR
3.32.5
8.8
1
671
672
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

33
AA le
vel
***
Fig. 7
0
0.04
0.08
0.12
Not treated IN-treated 673
674 on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
34
0
50
100
150
200
250-AA
***
***
*** ***
0
25
50
75
100
AA 1
SNP (μM)
3 52 250100-
Rel
ativ
e le
vel o
f c-d
i-GM
P (%
)Re
lativ
e ex
pres
sion
leve
l (%
)
psl algpel
A
B
Fig. 8
675
on June 12, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from