Embed Size (px)
Citation preview

1
LỜI CẢM ƠNTrong quá trình thực hiện đồ án tốt nghiệp, tôi đã nhận được nhiều sự giúp
đỡ, hướng dẫn, hỗ trợ từ phía gia đình thầy cô và bạn bè. Nhờ đó mà tôi đã hoàn
thành đồ án được như mong muốn, nay cho phép tôi được gửi lời cảm ơn sâu sắc và
chân thành đến:
Trước hết là ThS. Lê Phương Chung, người trực tiếp hướng dẫn đề tài.
Trong suốt thời gian thực hiện, thầy đã tận tình hướng dẫn, truyền đạt nhiều kinh
nghiệm, giúp tôi giải quyết các vấn đề nảy sinh trong quá trình làm đồ án và hoàn
thành đồ án đúng định hướng ban đầu.
Sau đó, tôi xin gửi lời cảm ơn chân thành đến TS. Nguyễn Văn Duy, Viện
phó, Viện Công nghệ sinh học và Môi trường, Trường Đại học Nha Trang đã tận
tình giúp đỡ tạo điều kiện thuận lợi cho tôi thực hiện tốt đồ án này. Đồng thời bày
tỏ lòng biết ơn sâu sắc đến Viện Công nghệ sinh học và Môi trường, Trường Đại
học Nha Trang và các thầy cô trong Bộ môn Công nghệ sinh học đã tạo môi trường
thuận lợi, cung cấp đầy đủ thiết bị, dụng cụ cho tôi thực hiện đề tài một cách xuyên
suốt.
Sau cùng, tôi xin cảm ơn sự ủng hộ, giúp đỡ của gia đình, bạn bè, chị
Nguyễn Minh Nhật, và các anh chị cao học.
Một lần nữa, tôi xin chân thành cảm ơn sự giúp đỡ quý báu đó. Chúc tất cả
mọi người sức khỏe thành đạt.
SVTH: Nguyễn Thị Thảo Trang

2
MỤC LỤC
LỜI CẢM ƠN........................................................................................................i
MỤC LỤC............................................................................................................ii
DANH MỤC BẢNG............................................................................................iv
DANH MỤC HÌNH..............................................................................................v
DANH MỤC TỪ, KÍ TỰ VIẾT TẮT................................................................vi
LỜI MỞ ĐẦU.......................................................................................................2
Chương I. TỔNG QUAN.....................................................................................4
1.1. Bacteriocin................................................................................................4
1.1.1 Khái niệm............................................................................................4
1.1.2 Các chủng vi sinh vật sinh bacteriocin................................................4
1.1.3 Phân loại..............................................................................................6
1.1.4 Tính chất của bacteriocin..................................................................13
1.1.5 Cơ chế hoạt động của bacteriocin.....................................................15
1.1.6 Sinh tổng hợp bacteriocin.................................................................16
1.2. Tôm hùm................................................................................................19
1.2.1 Phân bố..............................................................................................20
1.2.2 Tập tính.............................................................................................20
1.2.3 Đặc điểm sinh trưởng........................................................................21
1.2.4 Đặc điểm sinh sản.............................................................................22
1.2.5 Giá trị của tôm hùm...........................................................................22
1.3 Tình hình nghiên cứu và ứng dụng của bacteriocin...........................23
Chương II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU......................28
2.1 Nguyên liệu.............................................................................................28
2.1.2 Mẫu tôm hùm....................................................................................28
2.1.3 Chủng chỉ thị.....................................................................................28
2.1.4 Môi trường hóa chất và thuốc thử.....................................................28
SVTH: Nguyễn Thị Thảo Trang

3
2.1.5 Thiết bị chuyên dụng.........................................................................29
2.2 Phương pháp nghiên cứu......................................................................30
2.2.1 Phương pháp phân lập vi sinh vật.....................................................32
2.2.2 Tuyển chọn chủng sinh bacteriocin...................................................34
2.2.3 Xác định đặc điểm của chủng vi khuẩn sinh bacteriocin..................38
Chương III. KẾT QUẢ VÀ THẢO LUẬN......................................................44
3.1 Phân lập chủng vi sinh vật từ mẫu tôm Hùm.....................................44
3.2 Hoạt tính kháng khuẩn của các chủng phân lập................................45
3.3 Xác định hoạt tính bacteriocin.............................................................47
3.4 Quan sát hình thái tế bào vi khuẩn......................................................48
3.5 Đặc điểm sinh hóa của chủng vi khuẩn...............................................50
3.5.1 Khả năng chịu muối..........................................................................50
3.5.2 Thử catalase.......................................................................................51
3.5.3 Tính di động......................................................................................52
3.5.4 Khả năng sinh hơi.............................................................................53
3.6 Định danh bằng phương pháp sinh học phân tử................................53
Chương IV : KẾT LUẬN VÀ ĐỀ XUẤT Ý KIẾN.........................................54
4.1 Kết luận..................................................................................................54
4.2 Kiến nghị................................................................................................54
TÀI LIỆU THAM KHẢO.................................................................................55
PHỤ LỤC............................................................................................................58
SVTH: Nguyễn Thị Thảo Trang

4
DANH MỤC BẢNG
Bảng 1.1: Một số chủng vi khuẩn sinh bacteriocin loại I................................................5
Bảng 1.2 : Một số chủng vi khuẩn sinh bacteriocin loại II.............................................6
Bảng 1.3: Phân loại bacteriocin theo Jack [16]...............................................................9
Bảng 1.4: Bacteriocin lớp IIc theo phân loại của Klaenhammer [20]...........................12
Bảng 1.5: Độ bền nhiệt, pH và enzyme của một số bacteriocin được sinh tổng hợp
bởi vi khuẩn....................................................................................................................14
Bảng 3.1: Chủng vi khuẩn phân lập được.....................................................................44
Bảng 3.2: Hoạt tính kháng khuẩn của các chủng vi khuẩn...........................................46
Bảng 3.4: Khả năng chịu muối của N14.......................................................................50
SVTH: Nguyễn Thị Thảo Trang

5
DANH MỤC HÌNH
Hình 1.1: Cấu trúc của nisin............................................................................................7
Hình 1.2 : Cấu trúc sakacin P........................................................................................10
Hình 1.3: Cơ chế hoạt động của bacteriocin.................................................................16
Hình 1.4: Cơ chế tổng hợp bacteriocin nhóm I.............................................................18
Hình 1.5: Cơ chế tổng hợp bacteriocin nhóm II...........................................................19
Hình 1.6: Tôm hùm Bông.............................................................................................22
Hình 2.1: Quy trình thực hiện.......................................................................................31
Hình 2.2: Trình tự pha loãng.........................................................................................32
Hình 2.3: Quy trình thử tính kháng khuẩn của dịch bacteriocin thô với chủng chỉ thị
........................................................................................................................................35
Hình 2.4: Vùng kháng khuẩn của dịch bacteriocin trên đĩa petri..................................36
Hình 2.5 : Sơ đồ bố trí thí nghiệm xác định hoạt tính bacteriocin................................37
Hình 2.6: Sơ đồ bố trí thí nghiệm xác định đặc điểm vi khuẩn....................................38
Hình 3.1: Các chủng vi khuẩn ở nồng độ 10-3...............................................................44
Hình 3.2: Kết quả của 12 chủng nổi bật đối kháng với chủng Bacillus B1.1...............45
Hình 3.3 : Thử nghiệm với enzyme proteinase K và α-chymotripsin...........................48
Hình 3.4 : Hình thái khuẩn lạc và tế bào vi khuẩn N14................................................49
Hình 3.5 : Khả năng chịu muối của chủng N14............................................................51
Hình 3.6 : Khả năng sinh catalase của N14..................................................................52
Hình 3.7 : Khả năng di động của chủng N14................................................................52
Hình 3.8 : Khả năng sinh hơi của chủng N14...............................................................53
SVTH: Nguyễn Thị Thảo Trang

6
DANH MỤC TỪ, KÍ TỰ VIẾT TẮT
STT Kí hiệu viết tắt Nghĩa1 TSB Tryptone Soya Broth
2 TSA Tryptone Soya Agar
3 BC Bacterial Cellulose
4 LAB Lactic Acid Bacteria
5 ATP Adenosine Triphosphate
6 DNA Acid Deoxyribo Nucleic
7 HPK Histidine protein kinase
8 RR Response regulator
SVTH: Nguyễn Thị Thảo Trang

7
LỜI MỞ ĐẦU
Vào thời kỳ khai sinh ra vi sinh học hiện đại ngày nay, nhiều nhà khoa học
đã nghiên cứu và phát hiện ra chất kháng sinh. Từ thế kỷ 17, một thầy thuốc Hoàng
gia Anh đã biết cách chữa bệnh bằng cách dùng rêu áp lên vết thương, hay dùng
mẫu bánh mì để chữa trị thương tích. Đến những năm của thế kỷ 18, thuật ngữ “chất
kháng sinh” lần đầu tiên được Pasteur và Joubert sử dụng để mô tả hiện tượng kìm
hãm khả năng gây bệnh của vi khuẩn Bacillus anthracis trên động vật nhiễm bệnh,
và một số nhà khoa học khác đã phát hiện ra hoạt tính kháng khuẩn của một vài
chủng vi sinh vật. Vào năm 1929, khi Alexander Fleming tìm ra thuốc kháng sinh
penicillin từ nấm Penicillium notanum, có khả năng ức chế vi khuẩn sự sinh trưởng
của các loại vi khuẩn, được ứng dụng để chữa trị các bệnh nhiễm trùng trên phạm vi
rộng.
Việc tìm ra thuốc kháng sinh của Alexander Fleming đã mở ra bước ngoặc
mới cho y học loài người. Chỉ với một số lượng rất nhỏ nhưng hiệu quả tiêu diệt vi
sinh vật của penicillin rất cao. Thuốc kháng sinh không chỉ được ứng dụng trong
điều trị bệnh cho người, mà còn dùng điều trị bệnh cho vật nuôi, tăng trọng cho vật
nuôi và cả trong bảo quản thực phẩm (như chlotetracycllin dùng để bảo quản thịt;
tetracycline, oromycine… để bảo quản sản phẩm nhằm ngăn chặn quá trình thối rữa
của thịt cá, tôm). Tuy nhiên do lạm dụng quá mức kháng sinh đã dẫn đến khả năng
kháng thuốc ở vi sinh vật nên việc nghiên cứu chất kháng khuẩn để ứng dụng trong
bảo quản thực phẩm cần phải đi theo một hướng mới, hiệu quả hơn.
Không những thế, trong những năm gần đây, khi người tiêu dùng đã nhận
thức rõ hơn về tác động của phụ gia thực phẩm đối với sức khỏe và chuộng các loại
thực phẩm chế biến không bổ sung chất bảo quản hóa học tổng hợp vì tính an toàn
cho thực phẩm. Và từ yêu cầu cao về chất lượng, tính an toàn của thực phẩm từ phía
người tiêu dùng cũng như những yêu cầu nghiêm ngặt của chính phủ về an toàn
thực phẩm trong sản xuất, chế biến, bảo quản đã thúc đẩy nghiên cứu rộng rãi các
chất có khả năng bảo quản thực phẩm có nguồn góc tự nhiên.
SVTH: Nguyễn Thị Thảo Trang

8
Xuất phát từ những yêu cầu đó, việc tiếp tục tìm ra những chất kháng khuẩn
mới trở nên quan trọng trong lĩnh vực y dược và chăn nuôi, và trong công nghệ thực
phẩm. Một sự lựa chọn hợp lý là ứng dụng bacteriocin, một nhóm các hợp chất có
bản chất là peptide, nguồn gốc từ vi sinh vật và có khả năng kháng khuẩn mà không
gây ra tác động kháng thuốc ở vi sinh vật. Ngoài ra, bacteriocin cũng không gây dị
ứng cho con người, an toàn với sức khỏe. Đó là một sự lựa chọn được xem là có
nhiều ưu điểm và hiệu quả nhất. Vì thế, tôi chọn tiến hành nghiên cứu đề tài “
Nghiên cứu đặc điểm và phân loại vi sinh vật có khả năng sinh bacteriocin
phân lập được từ tôm hùm ở Khánh Hòa ”, giúp làm phong phú thêm bộ sưu tập
chủng vi sinh vật sinh chất kháng khuẩn, tạo ra nhiều sự lựa chọn cho việc tuyển
chọn chủng, và hiểu được đặc tính của các chủng này để tăng khả năng sinh
bacteriocin.
Đề tài nhằm các mục đích chính sau đây:
- Phân lập các chủng vi sinh vật có khả năng kháng khuẩn từ tôm hùm.
- Sàng lọc các chủng vi sinh vật phân lập từ tôm hùm có khả năng sinh
bacteriocin.
- Xác định các đặc điểm của vi sinh vật sàng lọc được.
- Định danh vi sinh vật có khả năng sinh bacteriocin phân lập được bằng
phương pháp sinh học phân tử.
SVTH: Nguyễn Thị Thảo Trang

9
Chương I. TỔNG QUAN1.1. Bacteriocin
Nghiên cứu đầu tiên và lâu đời nhất về bacteriocin là công trình nghiên cứu
của Gratia và cộng tác viên vào năm 1925 về khả năng kháng khuẩn của
Escherichia coli (colicin V) và thuật ngữ bacteriocin không xuất hiện cho đến
những năm 1950. Định nghĩa về bacteriocin đầu tiên đã dựa trên đặc tính của
colicin, đó là một chất sinh tổng hợp gây tử vong, phổ hoạt động hẹp bị giới hạn ở
những loài tương tự như vi khuẩn sản xuất. Ba chủng vi khuẩn Gram (+) được
nghiên cứu cho việc sản sinh bacteriocin lúc bấy giờ là: Bacillus sp.;Listeria sp. và
Staphylococcus sp. Các nghiên cứu trong những năm 1980 đã cho thấy có sự gia
tăng đáng kể về số lượng các công bố trên bacteriocin. Từ thời điểm này bắt đầu
bùng nổ những nghiên cứu bacteriocin, định hướng như một chất kháng khuẩn an
toàn trong lĩnh vực công nghệ thực phẩm[12],[23].
1.1.1 Khái niệm
Bacteriocin là chất kháng khuẩn có bản chất là peptide hay protein được tổng
hợp trên ribosome ở cả vi khuẩn Gram âm và Gram dương để chống lại vi khuẩn
khác có quan hệ gần gũi với chúng. Như vậy, loại vi khuẩn tạo ra bacteriocin nào
thì có khả năng kháng lại chính bacteriocin đó. Ngoài ra không gây ra phản ứng dị
ứng trong con người và vấn đề về sức khỏe, bị phân hủy nhanh bởi enzym
proteinase, lipase.
1.1.2 Các chủng vi sinh vật sinh bacteriocin
Bacteriocin được sinh tổng hợp bởi vi khuẩn Gram âm hoặc vi khuẩn Gram
dương với những đặc điểm :
- Bacteriocin của vi khuẩn gram âm: gồm nhiều loại khác nhau về kích thước,
nguồn gốc chủng vi sinh vật sản xuất, kiểu tác động và cơ chế miễn dịch.
Bacteriocin của vi khuẩn Gram âm yếu hơn bacteriocin của vi khuẩn Gram dương.
- Bacteirocin của vi khuẩn Gram dương: các bacteriocin này cũng nhiều như ở
vi khuẩn gram âm. Tuy nhiên chúng khác vi khuẩn gram âm ở chỗ là: việc tạo
SVTH: Nguyễn Thị Thảo Trang

10
bacteriocin không cần thiết phải gây chết vi sinh vật chủ và sự sinh tổng hợp
bacteriocin của vi khuẩn Gram dương cần nhiều gen hơn ở vi khuẩn Gram âm .
Những nghiên cứu trước đây về bacteriocin chủ yếu tập trung vào vi khuẩn
Gram âm, điển hình là E. coli sinh tổng hợp colicin. Tuy nhiên những nghiên cứu
về bacteriocin được sinh tổng hợp bởi vi khuẩn Gram dương cũng đã được tiến
hành và đã có nhiều báo cáo cung cấp những thông tin về vấn đề này. Ba chủng vi
khuẩn Gram dương được nghiên cứu cho việc sản sinh bacteriocin lúc bấy giờ là
Bacillus sp.;Listeria sp. và Staphylococcus sp.
Ngày nay thì các chủng vi khuẩn lactic sinh bacteriocin được nghiên cứu
nhiều hơn, do tính ưu việt của bacteriocin này dùng trong bảo quản thực phẩm là an
toàn với người sử dụng, không độc với tế bào nhân chuẩn, bị phân hủy bởi
proteinase trong hệ tiêu hóa nên ít ảnh hưởng đến quần xã sinh vật trong ruột người.
Thêm vào đó chúng hoạt động trong khoảng pH rộng, chống lại nhiều vi khuẩn gây
hư hỏng thực phẩm, cơ chế hoạt động dựa trên màng tế bào vi khuẩn nên không gây
ra tính đề kháng chéo với các kháng sinh và được mã hóa bởi các gen nằm trên
plasmid nên thuận tiện cho thao tác di truyền.
Vi khuẩn lactic từ lâu đã được sử dụng trong thực phẩm vì chúng có hoạt
tính kháng các loại vi sinh vật khác. Đặc tính này là do vi khuẩn lactic sinh acid hữu
cơ và sinh bacteriocin kháng khuẩn kháng nấm.
Bảng 1.1: Một số chủng vi khuẩn sinh bacteriocin loại I
Bacteiocin Vi khuẩnNisin A Nisin Z Mutacin 1140Carnocin U149CytolysinLacticin 481Lactocin SSalivaricin AMersacidi
Lactococcus lactis (nhiều chủng)Lactococcus lactis (nhiều chủng)Streptococcus mutans JH1000Carnobacterium piscicola 149 Enterococcus faecalis DS16Lactococcus Lactis CNRZ481Lactobacillus sake L45Streptococcus Salivarius 20P3Bacillus spp
SVTH: Nguyễn Thị Thảo Trang

11
Bảng 1.2 : Một số chủng vi khuẩn sinh bacteriocin loại II
Bacteiocin Vi khuẩn
Bavaricin A Bavaricin MN Carnobacteriocin B2 Carnobacteriocin BM1 Curvacin A Divercin V41Enterocin A Enterocin P Leucocin A/B –Talla
Mesentericin Y105 Acidocin J1132 Lactacin F Lacticin 3147 Lactobin A Lactococcin G Divergicin A Lactococcin A –Plantaricin 1.25β
Lactobacillus sake MI401Lactobacillus sake MN Carnobacterium pisciocola LV17B Carnobacterium pisciocola LV17B L. curvatus LTH1174Carnobacterium pisciocola divergens V41 Enterococcus faecium CTC492/T136Enterococcus faecium P13 Leuconostoc gelidum UAL187; Leuconostoc carosum Talla Leuconostoc mesenteroides Y105 L. acidophilus JCM 1132L. johnsonii 11088 Lc. lactis DPC3147L. amylovorus LMG P-13139 Lc. latis LMG2081Cb. divergens LV13 Lc. lactis subsp. cremoris LMG 2130L. plantarum TMW
1.1.3 Phân loại
Cho đến nay có khoảng 200 loại bacteriocin được xác định, tuy nhiên việc
phân loại các bacteriocin vẫn chưa được xác định rõ ràng và nó vẫn đang là vấn đề
tranh cãi. Các bacteriocin thường được phân loại dựa trên các tiêu chí khác nhau.
Những tiêu chí chính là họ vi khuẩn sản xuất, trọng lượng phân tử của chúng và
cuối cùng là trình tự chuỗi amino acid [20]. Trong đó, phương pháp phân loại được
chấp nhận và sử dụng phổ biến là bacteriocin được chia thành 4 lớp: lớp I, lớp II lớp
III và lớp IV. Hai nhóm bacteriocin lớp I và lớp II là đối tượng được tìm hiểu và
nghiên cứu nhiều hơn hơn hai nhóm còn lại.
SVTH: Nguyễn Thị Thảo Trang

12
1.1.3.1Lớp I
Bacteriocin lớp I hay còn gọi là Lanbiotic là những peptide nhỏ (<5 kDa),
bền nhiệt và tác động lên cấu trúc màng. Một bacteriocin của nhóm này là nisin.
Các lantibiotic chia thành 2 phân lớp là Ia và Ib dựa trên sự tương đồng cấu trúc.
Phân lớp Ia
Phân lớp Ia gồm các peptide dạng thuôn dài, linh hoạt và tích điện dương,
chúng hoạt động bằng cách kết hợp với các lipid mang điện tích âm hình thành các
lỗ trong màng tế bào chất của các loài vi khuẩn nhạy cảm [24]. Nisin cũng thuộc
nhóm này. Đây là một peptide được hình thành bởi 34 aminoacid, các loại acidamin
phổ biến có trong nisin là lanthionine (Lan), methyllanthionine (MeLan),
didehydroalanine (DHA) và acid didehydroaminobutyric (Dhb). Nisin có 2 biến thể
của nisin được ghi nhận là nisin A và nisin Z. .
Hình 1.1: Cấu trúc của nisin ( Nguồn: Ruiz-Larrea và cộng sự, 2005)
Loại bacteriocin này được sản xuất bởi một vài chủng Lactococcus lactic,
được sử dụng như một chất phụ gia trong thực phẩm. Nó có phổ kháng khuẩn Gram
(+) rộng, E. coli và các vi khuẩn Gram (-) khác chỉ bị ảnh hưởng bởi nisin khi màng
ngoài của chúng bị phá hỏng. Nisin được cho là có hoạt tính kháng khuẩn hiệu quả
đối với Staphylococcus aureus, Listeria monocytogenes, các tế bào sinh dưỡng của
Bacillus spp. và Clostridium spp. [24]. Chúng được sử dụng chủ yếu trong các thực
phẩm đóng hộp và các sản phẩm từ sữa, đặc biệt trong sản xuất phô mai, nhằm
chống lại các vi sinh vật chịu nhiệt như Bacillus và Clostridium .Nisin cũng được
ghi nhận có hiệu quả chống lại các bệnh viêm vú do vi khuẩn Gram (+) gây ra [10].
SVTH: Nguyễn Thị Thảo Trang

13
Phân lớp Ib
Phân lớp Ib gồm các peptide có dạng hình cầu, cấu trúc không linh động, tích
điện âm hoặc không tích điện. Chúng thể hiện hoạt động bằng cách gây ức chế sinh
tổng hợp peptidoglycan [11], gây nhiễu với các phân tử enzyme thiết yếu của vi
khuẩn nhạy cảm. Chúng thường hoạt động trong các phản ứng lên men quan trọng
của vi khuẩn. Nhóm này bao gồm các chất kháng khuẩn như mersacidine ,
actagardin, cinnamycin, mutacin A [20]
SVTH: Nguyễn Thị Thảo Trang

14
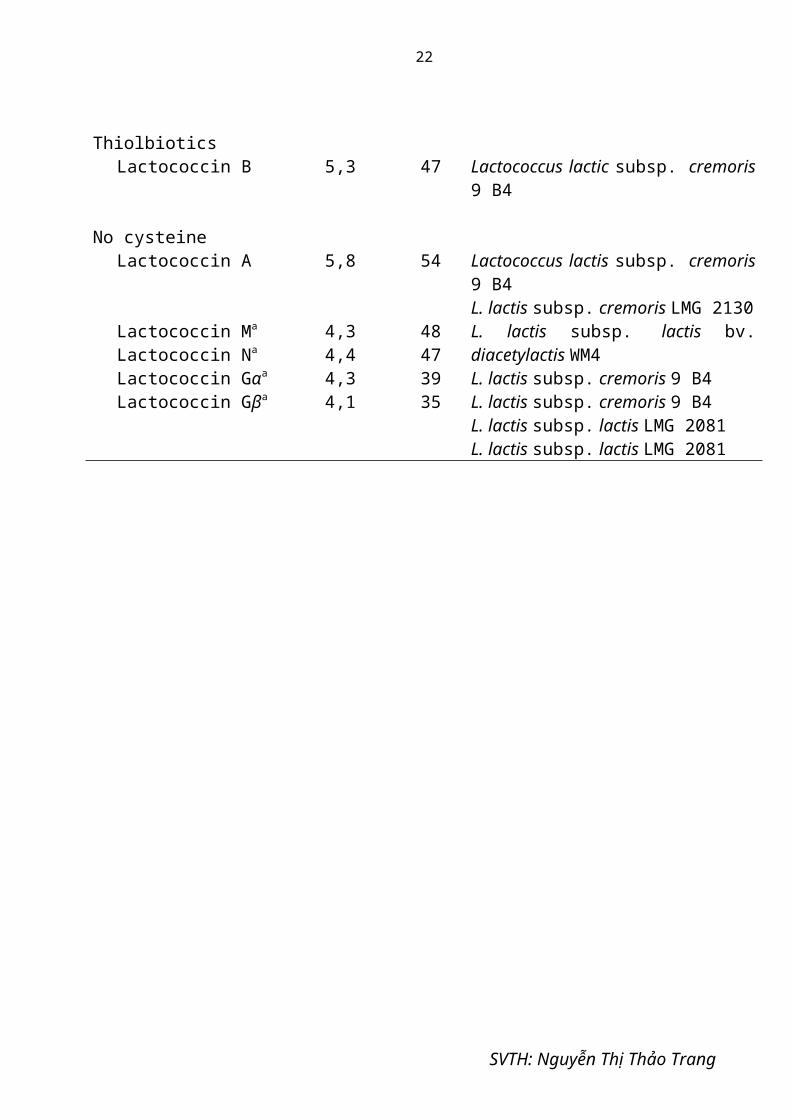
Bảng 1.3: Phân loại bacteriocin theo Jack [16]
Bacteriocin Khối lượng phân tử (kDa)
Amino acids Vi sinh vật sản xuất
Lantibiotic Actagardine AncoveninCinnamycinDuramycinEpiderminGalliderminLanthiopeptinMersacidinNisinPep5Subtilin
1,92,02,02,02,22,22,01,83,43,53,3
1919191922221919343432
Actinoplanes spp.Streptomyces spp.Streptomyces cinnamoneusStreptomyces cinnamoneusStaphylococcus epidermidisStaphylococcus gallinarumStreptoverticullum cinnamoneumBacillus sp.Lactococcus lactisStaphylococcus epidermidisBacillus subtilis
CystibioticsPediocin AcH/PA1Leucocin A/UAL 187Mesentericin Y 105Sakacin ASakacin PLactacin FCarnobacteriocin ACarnobacteriocin BM1Carnobacteriocin B2Cerein 7/8
4,63,93,84,34,45,65,14,54,94,9
44373741435753434856
Pediociccus acidilactici H/PAC 1.0Leuconostoc gelidium UAL 187Leuconostocmesenteroides Y 105Lactobacillus sake LB 706Lactobacillus sake LTH 674Lactobacillus acidophilus 11088Carnobacterium piscicola LV 17 ACarnobacterium piscicola LV 17 BCarnobacterium piscicola LV 17 BBacillus cereus Bc7
ThiolbioticsLactococcin B 5,3 47 Lactococcus lactic subsp. cremoris 9 B4
No cysteineLactococcin A
Lactococcin Ma
Lactococcin Na
Lactococcin Gαa
Lactococcin Gβa
5,8
4,34,44,34,1
54
48473935
Lactococcus lactis subsp. cremoris 9 B4L. lactis subsp. cremoris LMG 2130L. lactis subsp. lactis bv. diacetylactis WM4L. lactis subsp. cremoris 9 B4L. lactis subsp. cremoris 9 B4L. lactis subsp. lactis LMG 2081L. lactis subsp. lactis LMG 2081
SVTH: Nguyễn Thị Thảo Trang

15
1.1.3.2 Lớp II
Còn được gọi là lớp Non-Lanbiotic, bao gồm các bacteriocin có trọng lượng
phân tử nhỏ hơn 10 kDa, bền nhiệt và không chứa lanthionine. Các bacteriocin
nhóm II có thể chia thành 3 phân lớp, bao gồm IIa, IIb và IIc.
Lớp IIa
Lớp IIa là lớp lớn nhất, gồm các peptide hoạt động chống Listeria, đại diện
đặc trưng cho nhóm này là Leucocin A, pediocin PA-1. Các sakacin A, sakacin P
cũng là đại diện đặc trưng cho nhóm này. Các bacteriocin nhóm này hoạt động bằng
cách phá vỡ tính toàn năng của màng tế bào, làm mất cân bằng và thất thoát ion
phosphate hữu cơ do đó mà tiêu diệt vi khuẩn [20].
Sakacin A là bacteriocin được sản xuất bởi Lactobacillus sakei LB 706, được
cấu tạo bởi 41 acid amin (tiền thân là 90 acid amin). Bacteriocin này là một
polypeptide ổn định nhiệt, mang đặc tính di truyền[9].
Sakacin P là một polypeptide nhỏ, ổn định nhiệt, được tổng hợp bởi
ribosome, mang đặc tính di truyền [15], được sản xuất bởi Lactobacillus sakei LB
673.
Hình 1.2 : Cấu trúc sakacin P
Các bacteriocin này hứa hẹn cho nhiều ứng dụng cộng nghiệp nhờ vào hoạt
động kháng Listeria mạnh của chúng. Thậm chí chúng còn là các tác nhân kháng
SVTH: Nguyễn Thị Thảo Trang

16
Listeria được chú ý nhiều hơn là bacteriocin lớp I (nisin). Sakacin được ứng dụng
trong sản xuất xúc xích, xử lý ở các sản phẩm thịt để giữ được lâu (như sản phẩm
jăm bông [18]và thịt đông lạnh [21]), pho mát, các sản phẩm lên men acid lactic
khác. Ngoài ra, còn được sử dụng để ức chế sự phát triển của vi khuẩn không mong
muốn, có thể là nguyên nhân gây nhớt và gây mùi ở các sản phẩm.
Lớp IIb
Lớp IIb hình thành bởi một phức hợp của 2 peptide riêng biệt, những peptide
này ít hoặc không có hoạt động nào và nó không có sự giống nhau giữa các peptide
bổ sung. Các bacteriocin nhóm này có thể hoạt động riêng lẻ, cũng có thể liên hiệp
khi hoạt động cùng nhau (enterocin L50A, L50B) tạo lỗ trên màng tế bào, hoặc
chúng có thể cùng cần thiết cho hoạt động kháng khuẩn (lactococcins Gα/Gβ,
Lactococcins M/N và plantaricins EF, JK). Các bacteriocin đặc trưng cho nhóm này
là lactococcin G, plantaricin EF và plantaricin JK.
Lactococcin G là một bacteriocin có hoạt động phụ thuộc vào hoạt động bổ
sung của 2 peptide, là lactococcins Gα và Gβ. Lactococcin Gα gồm có 39 acid
amin, còn Lactococcin Gβ gồm có 35 acid amin. Bactericoin này được sử dụng như
một chất bảo quản thực phẩm, có khả năng kháng tế bào vi khuẩn Lactococcus
lactis. Hoạt động diệt khuẩn chỉ xảy ra khi có sự hiện diện của cả 2 peptid, mặc dù
2 peptide này có khả năng diệt khuẩn độc lập.
Lớp IIc
Lớp IIc là những peptide nhỏ, bền nhiệt, gồm những bacteriocin không đồng
nhất nên phương thức hoạt động của chúng cũng khác nhau. Nhóm IIc bao gồm tất
cả các bacteriocin lớp II không rơi vào nhóm IIa, IIb. Trong phân lớp này tìm thấy
có 2 dạng [20]:
- Chất kháng khuẩn có 1 hoặc 2 gốc cystein (như thiolbiotics có một và
cystibiotics có 2 gốc cystein).
- Kháng khuẩn không có cystein (lactoccin A và acidocin B)
SVTH: Nguyễn Thị Thảo Trang

17
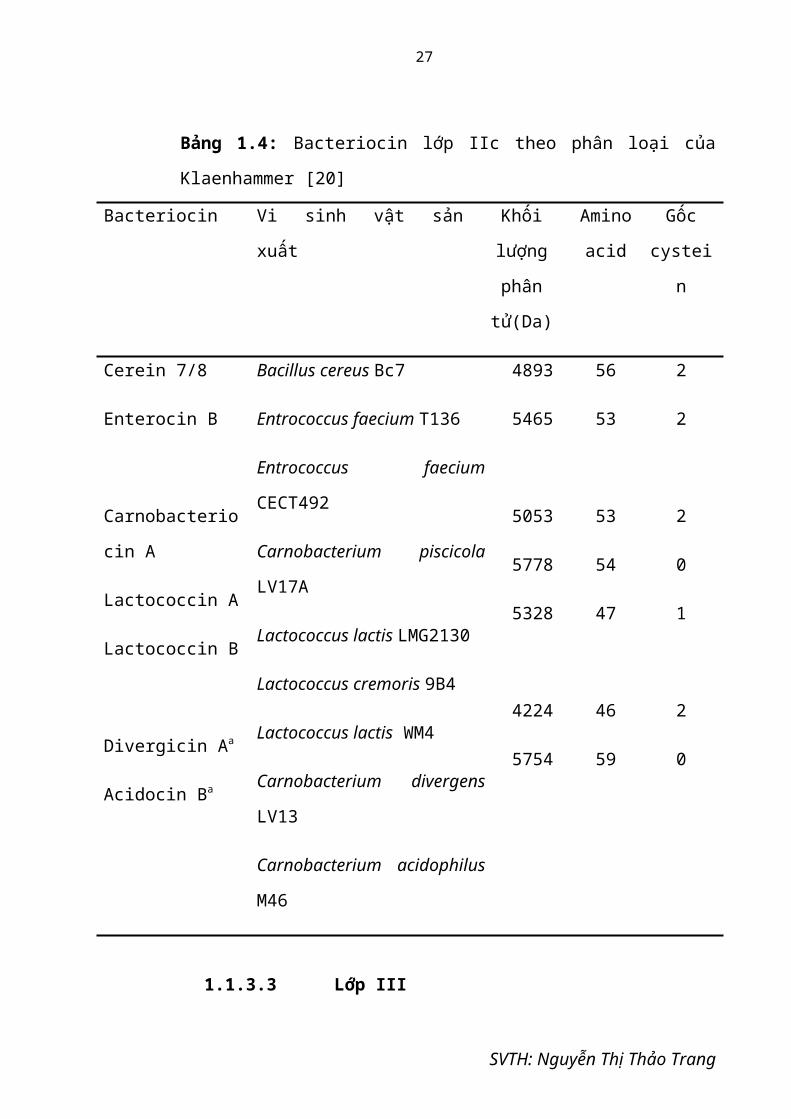
Bảng 1.4: Bacteriocin lớp IIc theo phân loại của Klaenhammer [20]
Bacteriocin Vi sinh vật sản xuất Khối lượng
phân tử(Da)
Amino
acid
Gốc
cystein
Cerein 7/8
Enterocin B
Carnobacteriocin A
Lactococcin A
Lactococcin B
Divergicin Aa
Acidocin Ba
Bacillus cereus Bc7
Entrococcus faecium T136
Entrococcus faecium CECT492
Carnobacterium piscicola
LV17A
Lactococcus lactis LMG2130
Lactococcus cremoris 9B4
Lactococcus lactis WM4
Carnobacterium divergens
LV13
Carnobacterium acidophilus
M46
4893
5465
5053
5778
5328
4224
5754
56
53
53
54
47
46
59
2
2
2
0
1
2
0
1.1.3.3 Lớp III
Lớp này bao gồm những peptide lớn, có trọng lượng phân tử lớn hơn 30
kDa, không bền nhiệt. Nhóm này có thể bao gồm các enzyme ngoại bào kháng lại
các vi khuẩn có thể bắt chước các hoạt động sinh lý của bacteriocin. Các bacteriocin
lớp III cho đến nay chỉ được phân lập từ các thành viên của giống Lactobacillus
[24]. Đại diện cho nhóm này là halveticin J [17] được sản xuất bởi vi khuẩn L.
halveticus 481 và helveticin V, acidifilicin A và lactacin A, lactacin B được sản
xuất bởi L. acidophilus.
SVTH: Nguyễn Thị Thảo Trang

18
1.1.3.4Lớp IV
Hiện nay, có rất ít tài liệu nghiên cứu về nhóm này. Một cách tổng quát các
bacteriocin nhóm IV được định nghĩa là bacteriocin phức tạp có chứa lipid hoặc
carbohydrate moities. Bao gồm các glycoprotein (lactocin 27), hoặc lipoprotein
(lacstrepcins) được quy định bởi non-protein. [20]
1.1.4 Tính chất của bacteriocin
1.1.4.1Thành phần hóa học
Bacteriocin có bản chất là protein, nhưng cũng có thể là hỗn hợp của protein
với các chất khác. Để kiểm tra bản chất protein, người ta sử dụng các enzyme đặc
trưng như proteinase. Dựa vào bảng phân tích hóa học cho thấy một vài bacteriocin
là protein đơn giản. Tuy nhiên, trong khi làm sạch bacteriocin của Staphylococcus,
Clostridium và Lactobacillus cho thấy là những phân tử phức tạp với thành phần
lipid, hydratcarbon và cộng thêm với protein. Một ví dụ khác về phân tử bacteriocin
có nhiều thành phần, streptocin STH1 là một trong những phân tử phức tạp bao
gồm protein, lipid và nhóm gốc phosphate.
1.1.4.2 Độ bền
Độ bền nhiệt
Hiện nay, một số nghiên cứu cho thấy rằng các loại bacteriocin của những
loài khác nhau thì khả năng chịu nhiệt cũng khác nhau. Mỗi loài có khả năng chịu
nhiệt ở một khoảng nhất định nhưng chủ yếu chúng thuộc nhóm I, II. Chủng
ST28MS và ST26MS được sản xuất bởi Lactobacillus plantarum không giảm khả
năng hoạt động chống vi khuẩn sau 90 phút tại 1000C hay 20 phút tại 1210C. Hay
đặc điểm tính chịu nhiệt có thể liên quan đến cấu trúc phân tử của bacteriocin.
Độ bền pH
Cũng như độ bền nhiệt, một số bacteriocin có thể hoạt động tốt dưới những
khoảng pH nhất định. Chủng ST28MS và ST26MS ngoài có độ bền nhiệt cao,
chúng lại có thể hoạt động ổn định trong 2 giờ tại những giá trị pH từ 2-12. Những
đặc tính này đều dựa vào bản chất thành phần cấu trúc của bacteriocin đó. Độ bền
nhiệt độ và pH rất quan trọng trong quá trình bảo quản nguyên liệu thực phẩm. Tùy
SVTH: Nguyễn Thị Thảo Trang

19
vào trạng thái, nhiệt độ thực phẩm mà chúng ta có thể lựa chọn loại bacteriocin bổ
sung và sao cho phù hợp với mục đích sản xuất.
Độ bền enzyme
Mỗi loại bacteriocin có thành phần cấu trúc các aminoacid khác nhau nên sẽ
chịu sự phân cắt đặc hiệu của các enzyme khác nhau, thích hợp với nó. Khi bị phân
cắt bởi các enzyme này sẽ khiến bacteriocin mất đi hoạt tính kháng khuẩn. Đồng
thời đây cũng là dấu hiệu giúp ta nhận biết được đó có phải là bacteriocin không.
Độ bền nhiệt, pH và enzyme của một số bacteriocin được sinh tổng hợp bởi
vi khuẩn Gram dương được trình bày ở bảng sau.
Bảng 1.5: Độ bền nhiệt, pH và enzyme của một số bacteriocin được sinh tổng hợp bởi vi khuẩn
Bacteriocin Độ bền nhiệt Độ bền pH Enzyme thủy phân
Megacin A-216 600C, 30 phút 2 – 7 Chymotrypsin, Pepsin, Trypsin
Clostosin A 1000C, 30 phút 4 – 9 Trypsin, Chymotrypsin, Dnase,
Pronase P, Rnase
Clostosin B 800C, 10 phút 4 – 9 Trypsin, Chymotrypsin, Dnase,
Pronase P, Rnase
Clostosin C 800C, 10 phút 4 – 9 Trypsin, Chymotrypsin, Dnase,
Pronase P, Rnase
Clostosin D 1000C, 30 phút 4 – 9 Trypsin, Chymotrypsin, Dnase,
Pronase P, Rnase
Boticin E-S5 1000C, 10 phút 1,1 – 9,5 Trypsin, Chymotrypsin, Dnase,
Pepsin
Bioticin P 600C, 30 phút 6,5 – 7,5 Trypsin, Rnase, Alkaline
phosphotase, Phospholipases C, D
Butyricin 7423 1000C, 10 phút 2 – 12 Trypsin
Perfringocin 11105 1000C, 30 phút 2 – 12 Trypsin
1.1.4.3 Tính miễn dịch
SVTH: Nguyễn Thị Thảo Trang

20
Bacteriocin có khối lượng phân tử lớn, thành phần cấu tạo là protein và nhiều
bacteriocin có tính kháng nguyên cao. Tuy nhiên, hiện nay chỉ có duy nhất một vài
bài báo cáo nghiên cứu tính kháng nguyên của bacteriocin sinh bởi vi khuẩn Gram
dương. Megacin A-216 là một kháng nguyên và tế bào cũng sinh ra một kháng thể
có khả năng chống lại ảnh hưởng giết chính nó. Việc nghiên cứu những kháng thể
đến bacteriocin trong huyết thanh của con người có lẽ là một lĩnh vực thú vị cho
những nghiên cứu sau này.
Một thuộc tính cần thiết để bảo đảm sự sống sót của vi sinh vật sinh
bacteriocin là có sự miễn dịch đặc biệt. Theo Koponen (2004) tế bào vi khuẩn có
một số cơ chế miễn dịch, đây là khả năng của tế bào trong việc bảo vệ chính nó để
chống lại bacteriocin sinh ra bởi sự trao đổi của chính mình.
Cả cấu tạo thành tế bào và thành phần cấu tạo màng lipid của vi khuẩn cho
thấy sự liên quan đến hoạt động của bacteriocin cũng như sự kháng cự bacteriocin.
Sự biến đổi nhạy cảm của vi khuẩn Gram dương về phía những bacteriocin đáng kể.
Hoạt động kìm hãm của bacteriocin có thể thay đổi giữa những gen khác nhau,
những loài cùng bộ gene và thậm chí đến môi trường nuôi cấy từ những điều kiện
ngoại cảnh khác .
Có hai kiểu miễn dịch được thấy ở Lantibiotic. Một kiểu miễn dịch dựa vào
protein miễn dịch đặc trưng, LanI. Trong khi kiểu miễn dịch khác dựa vào vật
chuyên chở nhiều thành phần riêng biệt (Lan EFG). Bacteriocin sẽ nhận biết được
những vi sinh vật sinh bacteriocin nhờ sự không có mặt của những thụ thể đặc biệt
và kết quả là chúng không bám được vào tế bào vi sinh vật đó[14].
1.1.5 Cơ chế hoạt động của bacteriocin
1.1.5.1Phạm vi hoạt động
Phạm vi hoạt động của bacteriocin được xem xét dựa trên phạm vi kháng
khuẩn, giống vi sinh vật sinh ra, khối lượng phân tử, tính ổn định, tính chất hóa sinh
và kiểu hoạt động của bacteriocin. Hầu hết các bacteriocin được tổng hợp bởi vi
khuẩn Gram âm có hoạt tính ức chế các loài cùng họ hàng. Đối với bacteriocin được
SVTH: Nguyễn Thị Thảo Trang

21
sinh tổng hợp bởi vi khuẩn Gram dương có hoạt tính ức chế các loài vi khuẩn Gram
dương đôi khi nó cũng ức chế một số loài vi khuẩn Gram âm.
1.1.5.2Cơ chế hoạt động
Bacteriocin có khả năng tiêu diệt các vi khuẩn khác do sự tạo thành các kênh làm thay đổi tính thấm của màng tế bào, nhiều loại bacteriocin còn có khả năng phân giải DNA, RNA và tấn công vào peptidoglycan để làm suy yếu thành tế bào.
Một loại bacteriocin là Lacticin 3147 có tác dụng diệt khuẩn trên những tế bào nhạy cảm bởi sự tuong tác đầu tiên với thành tế bào. Đó là nguyên nhân mà trên màng tế bào tạo ra những kênh cho K+ và phosphate vô cơ đi ra khỏi tế bào. Trong sự nổ lực để tái tích lũy lại những ion này, các ATP sẽ bị thủy phân. Khi ATP được yêu cầu cho sự duy trì của những chức năng quan trọng trong tế bào, như gradient pH tại màng tế bào , thì đã không còn nữa, những chức năng sẽ bị phá vỡ và tế bào dần mất năng lượng và chết [13],[1].
Hình 1.3: Cơ chế hoạt động của bacteriocin(Nguồn Fidel Toldrá, 2009)
1.1.6 Sinh tổng hợp bacteriocin
Vì bacteriocin có bản chất là polypeptide nên sinh tổng hợp theo cơ chế của
sinh tổng hợp protein nhờ ribosome.
SVTH: Nguyễn Thị Thảo Trang
22
Trong tế bào vi khuẩn có mang các gene mã hóa cho các protein tham gia
vào quá trình sinh tổng hợp bacteriocin. Qua quá trình phiên mã, dịch mã sẽ tạo ra
các protein đi vào giai đoạn tạo ra bacteriocin hoàn chỉnh, giải phóng nó ra khỏi tế
bào vi khuẩn. Tuy nhiên vì hoạt tính ức chế/tiêu diệt vi khuẩn của bacteriocin nên
nó cũng có thể tác động ngược trở lại lên vi khuẩn sản xuất ra nó, do đó vi khuẩn
sản xuất đòi hỏi phải có protein miễn dịch, giúp nó miễn dịch khỏi tác động của
bacteriocin do chính nó tạo ra.
Các loại gen khác nhau được quy định cho việc tổng hợp bacteriocin:
- Gene tạo ra chất tiền thân của bacteriocin: pre – bacteriocin.
- Gene tạo miễn dịch cho chủng sản xuất (immunity gene)
- Gene mã hóa kênh vận chuyển cho pre – bacteriocin ra khỏi tế bào kích hoạt cơ
chế vận chuyển đặc biệt (kênh vận chuyển ABC – ABC transporter)
- Gene kích hoạt cơ chế tạo ra protein tiếp nhận thông tin (accessing protein), giữ
vai trò quan trọng vận chuyển và kích hoạt hoạt động kháng khuẩn của
bacteriocin khi bacterocin được xuất ra khỏi tế bào (từ pre – bacteriocin thành
bacteriocin). Ngoài ra còn có những gene khác trong sinh tổng hợp bacteriocin
được tìm thấy ở nhóm II.
Khi gặp vi khuẩn đối nghịch, để tổng hợp ra bacteriocin tiêu diệt/ức chế vi
khuẩn đối nghịch cần phải có cơ chế điều hòa tổng hợp bacteriocin, đó chính là
operon.
Gene điều hòa (Promoter) – trình tự nucleotide để 1 gene được phiên mã.
Promoter được nhận diện bởi RNA polymerase, bắt đầu quá trình phiên mã mRNA.
Trong tổng hợp RNA, promoter chỉ thị gene nào cần được sử dụng để tạo mRNA,
do đó kiểm soát tế bào tạo ra protein nào, protein ức chế để gắn vào gene chỉ huy
hay protein cần cho tế bào hoạt động.
Gene chỉ huy (Operator) – một đoạn DNA để protein kìm hãm gắn vào, do
đó ngăn cản RNA polymerase phiên mã gene.
Gene cấu trúc (Structural gene) – các gene được điều hòa tổng hợp bởi
operon. Quay trở lại cơ chế sinh tổng hợp bacteriocin, trên gene cấu trúc có mang
SVTH: Nguyễn Thị Thảo Trang

23
thông tin để phiên mã tạo mRNA chứa thông tin tạo ra pre-bacteriocin (tiền
bacteriocin) cũng như các gene khác như gene miễn dịch (giúp bảo vệ vi khuẩn khỏi
tác động tiêu diệt/ức chế của bacteriocin do chính nó tạo ra)... mRNA sau đó cùng
với ribosome, tRNA đi vào quá trình dịch mã tạo ra pre-bacteriocin, protein miễn
dịch...
Sau khi quá trình phiên mã, dịch mã tạo ra pre-bacteriocin, dưới tác động
của các enzyme trong tế bào chất, pre-bacteriocin khi được giải phóng ra khỏi màng
tế bào được cắt bỏ leader peptide để tạo thành bacteriocin hoàn chỉnh có khả năng
kháng khuẩn và được vận chuyển ra khỏi màng thông qua hệ thống vận chuyển
ABC. Vai trò của leader peptide trong pre-bacteriocin là để bảo vệ vi khuẩn chính
nó khỏi tác động tiêu diệt của bacteriocin trong khi tạo ra nó.
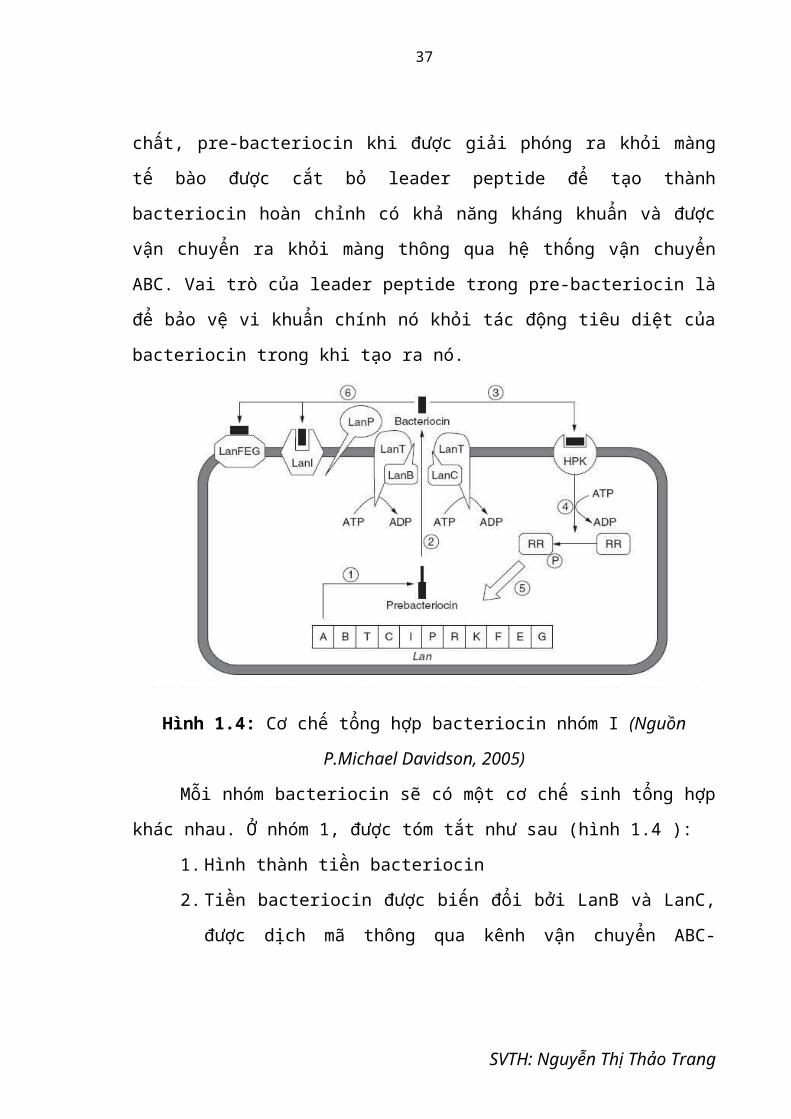
Hình 1.4: Cơ chế tổng hợp bacteriocin nhóm I (Nguồn P.Michael Davidson, 2005)
Mỗi nhóm bacteriocin sẽ có một cơ chế sinh tổng hợp khác nhau. Ở nhóm 1,
được tóm tắt như sau (hình 1.4 ):
1. Hình thành tiền bacteriocin
2. Tiền bacteriocin được biến đổi bởi LanB và LanC, được dịch mã thông
qua kênh vận chuyển ABC-transporter LanT, và được xử lý bởi LanP, kết
quả tạo ra được bacteriocin trưởng thành.
SVTH: Nguyễn Thị Thảo Trang

24
3. HPK nhận diện sự hiện diện của bacteriocin và tự động phosphoryl hóa.
4. Nhóm phosphoryl được truyền đến nhân tố đáp ứng điều hòa RR
5. RR phiên mã gen điều hòa.
6. Sản phẩm chất kháng khuẩn.
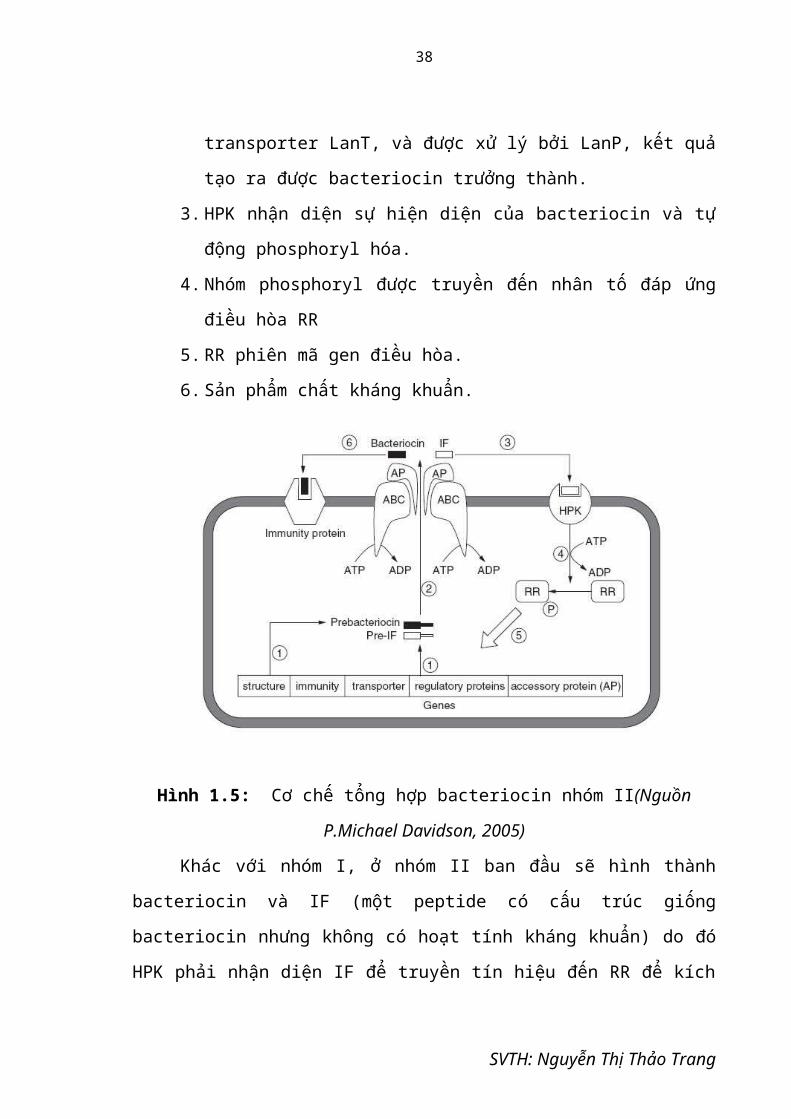
Hình 1.5: Cơ chế tổng hợp bacteriocin nhóm II(Nguồn P.Michael Davidson, 2005)
Khác với nhóm I, ở nhóm II ban đầu sẽ hình thành bacteriocin và IF (một
peptide có cấu trúc giống bacteriocin nhưng không có hoạt tính kháng khuẩn) do đó
HPK phải nhận diện IF để truyền tín hiệu đến RR để kích hoạt tiếp quá trình phiên
mã, tạo được bacteriocin hoàn chỉnh. (hình 1.5 )
1.2. Tôm hùm
Tôm hùm là tên gọi chung của nhóm giáp xác mười chân thuộc 4 họ:
Palinuridae, Scyllaridae, Nephropidae và Synaxidae, giữa chúng có những điểm
đặc trưng về tập tính và môi trường sống. Với sự phong phú về thành phần giống
loài, chúng tạo nên mắt xích quan trọng trong chuỗi thức ăn và có vai trò thiết yếu
trong hệ sinh thái của biển và đại dương. Ở Việt Nam, cho đến nay đã xác định
được 9 loài thuộc họ tôm hùm Gai (Palinuridae), 9 loài thuộc họ tôm Mũ Ni
(Scyllaridae) và 4 loài thuộc họ Nephropidae. Trong đó, một số loài thuộc họ tôm
SVTH: Nguyễn Thị Thảo Trang

25
hùm Gai được nuôi phổ biến hiện nay như tôm hùm Bông (tôm hùm Sao, tôm hùm
Hèo), tôm hùm Đá (xanh chân ngắn), tôm hùm Đỏ (hùm lửa), tôm hùm Sỏi (xanh
chân dài, hùm ghì), tôm hùm Tre (Tề Thiên).
Hệ thống phân loại tôm hùm
Ngành Arthropoda
Lớp Malacostraca
Bộ Decapoda
Phân bộ Pleocyemata
Nhóm Palinura
Họ Paliniridae
Giống Panulirus
1.2.1 Phân bố
Trên thế giới tôm hùm phân bố chủ yếu ở các vùng biển Nhiệt đới đến Á
nhiệt đới như: Úc, Ðài Loan, Trung Quốc, Nhật Bản, In-đô-nê-si-a,…
Ở Việt Nam, tôm hùm phân bố chủ yếu ở các tỉnh miền Trung từ Quảng
Bình đến Bình Thuận, đặc biệt phân bố nhiều ở các tỉnh Khánh Hoà, Ninh Thuận và
Bình Thuận.
Phân bố theo độ sâu: Tùy vào giai đoạn phát triển và tùy vào từng loài mà
chúng phân bố theo độ sâu khác nhau. Ở giai đoạn trưởng thành tôm hùm bông
thường phân bố ở độ sâu từ 20m trở lên, ở giai đoạn ấu trùng và con non chúng
thường phân bố ở các bãi rạn, đá san hô ở độ sâu từ 2 - 10m nước.
Tôm hùm thường sống ở các bãi rạng đá, rạng san hô nơi có nhiều hang hốc,
khe rãnh ven biển. Chúng có tập tính sống quần tụ chủ yếu là ở tầng đáy với chất
đáy sạch, không bùn. Ban ngày trú ẩn trong các hang đá ít hoạt động, ban đêm hoạt
động tích cực tìm mồi.
Tôm hùm sống thích hợp ở các vùng biển có độ mặn từ 30 -36 ‰, nhiệt độ
từ 25 – 32oC.
1.2.2 Tập tính
SVTH: Nguyễn Thị Thảo Trang

26
Tôm hùm là loại động vật ăn tạp, trong tự nhiên chúng ăn chủ yếu là các loại
động vật như: Cá, tôm, cua ghẹ, cầu gai, giáp xác nhỏ, nhuyễn thể,...ngoài ra còn ăn
các loại thực vật như rong rêu., chúng thường có tập tính bắt mồi tích cực vào ban
đêm và tờ mờ sáng. Tuỳ vào giai đoạn phát triển mà nhu cầu dinh dưỡng khác nhau,
tôm càng nhỏ thì nhu cầu dinh dưỡng càng cao, ở giai đoạn trước lột xác 2-4 ngày
tôm ăn rất mạnh, giai đoạn lột xác tôm sẽ ăn chậm lại.
Tôm hùm cũng như nhiều giáp xác khác sinh trưởng thông qua quá trình lột
xác. Ở giai đoạn tôm còn nhỏ chu kỳ lột xác ngắn, tôm càng lớn chu kì lột xác càng
dài, sau mỗi lần lột xác thì kích thước và khối lượng của tôm tăng lên rất nhiều.
Nhìn chung thì tôm hùm có chu kỳ lột xác dài hơn so với các loài giáp xác khác, do
vậy tốc độ tăng tưởng của chúng tương đối chậm.
1.2.3 Đặc điểm sinh trưởng
Sự sinh trưởng ở tôm hùm được xác định bằng sự tăng lên về chiều dài giáp
đầu ngực (CL) hoặc khối lượng cơ thể (W) và mức tăng sinh trưởng được thể hiện
rõ qua chu ký lột xác, điểm đặc trưng ở lớp giáp xác. Có nhiều yếu tố ảnh hưởng
đến chu kỳ lột xác của tôm bao gồm các yếu tố ngoại cảnh lẫn nội tại như: nhiệt độ
nước, ánh sáng, các hormone lột xác và ức chế lột xác, thức ăn, tập tính sống, sự tái
tạo phần phụ,…
Do ảnh hưởng của nhiệt độ nước làm tăng giảm quá trình trao đổi chất trong
cơ thể tôm từ đó hoạt động tích lũy năng lượng cần thiết cho quá trình sinh trưởng
của tôm cũng bị thay đổi theo. Khi nhiệt độ nước thay đổi đột ngột 3 – 50C, tôm con
có thể bị chết do không kịp thích nghi. Khi nhiệt độ tăng cao và ổn định (29 – 300C)
thời gian lột xác là 4 – 6 ngày; khi nhiệt độ hạ thấp xuống 20 – 240C, thời gian giữa
hai lần lột xác kéo dài gấp 1 – 1,2 lần.
Tôm có kích cỡ càng nhỏ càng nhạy cảm với ánh sáng. Những con nuôi
trong điều kiện không có ánh sáng không những bị kéo dài chu kỳ lột xác mà hoạt
động bắt mồi cũng bị ảnh hưởng, đặc biệt là mất dần các sắc tố và biến đổi màu tự
nhiên của vỏ, làm cho tôm phát triển không bình thường[7].
SVTH: Nguyễn Thị Thảo Trang
27
Nhìn chung, tôm hùm lớn lên nhờ quá trình lột xác. Tôm càng nhỏ, quá trình
lột xác càng ngắn và tôm lớn càng nhanh. Tôm hùm có chu kỳ lột xác dài hơn so
với các loài giáp xác khác, do đó, tốc độ tăng trưởng của chúng cũng chậm hơn.
Qua mỗi lần lột xác kế tiếp, tỷ lệ phần trăm chiều dài giáp đầu ngực của tôm giảm
xuống nhưng độ dài vỏ tôm vẫn tăng nhiều [8].
1.2.4 Đặc điểm sinh sản
Tôm hùm sinh sản rải rác quanh năm nhưng mùa vụ sinh sản của giống
Palinurus chủ yếu từ tháng 4-5 và tháng 9 hàng năm. Sức sinh sản của tôm hùm
tương đối lớn và chúng có thể đẻ nhiều lần trong một năm (thường là 2 lần/năm).
Tôm hùm có kích thước vỏ đầu ngực từ 90 – 99 mm đã bắt đầu tham gia sinh sản.
Đến mùa sinh sản tôm thành thục kết đàn di cư ra các vùng biển sâu 10-35m
và có độ mặn 30/340/00 để đẻ. Tôm thụ tinh ngoài, con đực gửi khối túi tinh trên
mảnh ức của con cái. Túi tinh được làm rách nhiều giờ trước khi con cái đẻ để thụ
tinh với trứng ở phần bụng và chân bơi. Tôm giữ trứng ở các đôi chân bụng cho đến
khi trứng nở. Ấu trùng Phyllosoma qua 12 lần lột xác và biến thái thành ấu trùng
Puerulus. Ấu trùng Puerulus qua 4 lần lột xác thành tôm hùm con. Từ lúc trứng nở
đến giai đoạn “tôm trắng” kéo dài khoảng 10 – 12 tháng.
1.2.5 Giá trị của tôm hùm
Tôm hùm được tôn vinh là vua của các loại hải sản giá trị dinh dưỡng cao. Thịt tôm hùm chắc, dai, ngọt ăn một lần không thể nào quên. Tập trung sự tinh túy nhất ở con tôm hùm là gạch san đúng nơi đầu tôm và chạy thành dải vàng dọc sống lưng. Gạch tôm hùm béo, thơm, nhiều đạm.
Hình 1.6: Tôm hùm Bông
SVTH: Nguyễn Thị Thảo Trang

28
Về chất lượng sản phẩm, theo các tài liệu phân tích, phần ăn được của tôm
hùm chiếm 20 – 30% thể trọng. Trong thịt của tôm Hùm tính theo trọng lượng tươi,
hàm lượng protein chiếm 17,62%, lipid chiếm 0,29%, các axit amin chiếm 77,2%
protein cơ thể. Do thịt thơm ngon giàu chất dinh dưỡng, nên tôm Hùm được coi là
món ăn sang trọng quý hiếm và được ưa thích ở nhiều nước. 70% còn lại là vỏ, đầu
thân được sử dụng để sản xuất chitosan ứng dụng trong lĩnh vực y dược, thực phẩm,
bao gói bảo quản thực phẩm và hóa mỹ phẩm phục vụ nhu cầu của nền kinh tế.
1.3 Tình hình nghiên cứu và ứng dụng của bacteriocin
1.3.1 Ứng dụng
Từ rất lâu các bacteriocin này đã được ứng dụng rộng rãi trong bảo quản
thực phẩm, điển hình là Nisin. Năm 1953, thương phẩm Nisaplin xuất hiện trên thị
trường như chất bảo quản thực phẩm và đến năm 1969, tổ chức WHO công nhận
Nisin là chất bảo quản an toàn có nguồn gốc sinh học. Từ đó, các bacteriocin càng
được ứng dụng nhiều trong công nghệ thực phẩm, nhất là trong trong bảo quản thực
phẩm như phomat, xúc xích, sữa chua, một số loại sản phẩm thịt, rau quả đóng
hộp… để thay thế cho các chất kháng sinh để hạn chế tác dụng phụ của nó, nhờ tính
ưu việt của chúng:
- Bacteriocin có tính an toàn cao trong thực phẩm dành cho người. Chúng có ít
hạn chế hơn so với những chất bảo quản hóa học vì là các phân tử được sản
sinh tự nhiên bởi vi sinh vật lên men trong thực phẩm lên men truyền thống .
- Không gây tác động đến môi trường vì chúng bị thoái biến nhanh chóng.
- Các bacteriocin được sử dụng như nguồn thức ăn chủ yếu đối với các tác
nhân gây bệnh mà không làm ảnh hưởng đến vi khuẩn có lợi.
- Bacteriocin không làm thay đổi các tính chất cảm quan của thực phẩm.
- Chúng có phổ hoạt động rõ ràng.
- Có tác dụng bổ sung cho các tác nhân kháng khuẩn.
SVTH: Nguyễn Thị Thảo Trang

29
Không chỉ vậy, bacteriocin còn được ứng dụng trong chăn nuôi làm thuốc
tăng trọng. Trong y dược, thú y, bacteriocin còn được ứng dụng như một chế phẩm
sinh học để điều trị một số bệnh như viêm đường tiết niệu, hay nhiễm trùng đường
tiêu hoá…
1.3.2 Tình hình nghiên cứu
Bacteriocin mang lại nhiều lợi ích, và ứng dụng trong cuộc sống nhờ các tính
chất đặc trưng của chúng. Song còn nhiều vấn đề liên quan đến bacteriocin chưa
được sáng tỏ. Vì vậy, thúc đẩy các nhà khoa học bắt tay vào nghiên cứu lĩnh vực
này. Và việc nghiên cứu và phát triển các loài vi sinh vật có khả năng sinh
bacteriocin có phổ kháng khuẩn lớn được ứng dụng rộng rãi trong nhiều lĩnh vực là
vấn đề mà nhiều nhà khoa học trong nước và trên thế giới quan tâm. Có rất nhiều
công trình nghiên cứu từ trước đến nay đã góp phần nâng cao chất lượng cuộc sống
nhân loại.
Vào tháng 1/1999, Budu-Amoako E cùng cộng sự thuộc trung tâm Công
nghệ thực phẩm PEI, Canada đã nghiên cứu được hiệu quả kết hợp của Nisin và
nhiệt độ trung bình để tăng khả năng tiêu diệt Listeria monocytogenes trong các
thùng chứa bảo quản tôm hùm. Theo đó, khi thêm Nisin ở mức 25mg/kg vào thùng
chứa tôm hùm có muối bao xung quanh, kết hợp với quá trình xử lý nhiệt 60oC
trong 5 phút và 65oC trong 2 phút sẽ giúp giảm đáng kể L.mono.
Năm 2002, Soore cùng cộng sự đã nghiên cứu việc tạo ra một chất kháng
khuẩn đặc biệt của vi khuẩn lactic gọi là bacteriocin. Bacteriocin được sử dụng như
một chất bảo quản thực phẩm tự nhiên có khả năng thay thế các loại hoá chất bảo
quản khác. Bacteriocin có bản chất là protein nên rất an toàn đối với sức khoẻ con
người.
Năm 2004, nhóm tác giả Todorov, Van và Dicks thuộc khoa vi sinh, đại học
Stellenbosch- Nam Phi đã nghiên cứu khả năng sinh tổng hợp bacteriocin của chủng
Lactobacillus Plantarum ST13BR , một chủng được phân lập từ bia Barley.
Bacteriocin được sinh tổng hợp bởi vi khuẩn Lactobacillus plantarum được phân
lập từ bia barley (kích thước phân tử 10kDa) ức chế sự phát triển của vi khuẩn
SVTH: Nguyễn Thị Thảo Trang

30
Lactobacillus casein, Pseudomonas aeruginosa, Enterococcus faecalis, Klebsiella
pneumoniae và Escherichia coli. Nghiên cứu đã đưa ra được các loại môi trường có
ảnh hưởng đến khả năng sinh tổng hợp bacteriocin và hỗn hợp các loại môi trường
mà việc sinh tổng hợp bacteriocin là lớn nhất.
Năm 2006, 50 chủng vi khuẩn lactic được tìm thấy từ sản phẩm lên men
truyền thống suan-tsai của Đài Loan. Với các chủng sau khi phân lập tiếp tục
nghiên cứu các đặc điểm hình thái, sinh lý, sinh hoá và khả năng sinh tổng hợp
bacteriocin.
Năm 2006, Marcinowski, giám đốc điều hành nghiên cứu tại BASF cho biết
kẹo cao su có chứa các chủng vi khuẩn có lợi Lactobacillus do công ty hoá chất
BASF của Đức phát triển. Đây là sản phẩm giúp người dùng loại trừ các bệnh về
răng miệng. Chủng Lactobacillus mới có tên là L anti-caries có khả năng sinh
bacteriocin làm cho vi khuẩn gây sâu răng kết thành khối không thẻ dính trên bề
mặt răng và bị loại bỏ dễ dàng khi súc miệng.
Tháng 3/2007 nhóm tác giả Todorov và Dicks KM thuộc khoa vi sinh, đại
học Stellenbosch- Nam Phi đã nghiên cứu khả năng sinh tổng hợp bacteriocin của
chủng Lactobacillus pentosus ST712BZ được phân lập từ boza. Bacteriocin
ST712BZ (kích thước 14kDa) ức chế sự phát triển của Lactobacillus casei, E.coli,
Pseudomonas aeruginosa, Enterococus faecalis, Klebsiella pneumoniae và
Lactobacillus curvatus. Sự phát triển của chủng ST712BZ trên môi trường BHI,
M17, sữa đậu nành và mật đường tương tự như trên môi trường MRS với việc sinh
tổng hợp bacteriocin cực đại (12800AU/ml) được ghi nhận trên môi trường MRS
sau 24h. Nghiên cứu đã đưa ra được các loại môi trường, nhiệt độ và pH có ảnh
hưởng đến khả năng sinh tổng hợp bacteriocin và thành phần hỗn hợp các loại môi
trường mà việc sinh tổng hợp bacteriocin là lớn nhất.
Các nhà nghiên cứu Csiro để tìm ra một vài bacteriocin và sử dụng chúng để
tiêu diệt những con vi khuẩn gây bệnh giống trong gà và lợn. Bộ gen cho phân tử
bacteriocin sẽ được chèn vào gen vi khuẩn không gây bệnh hoặc vi rút mà có thể
SVTH: Nguyễn Thị Thảo Trang

31
sinh sản và tiết ra bacteriocin. Lợi ích chính của việc sử dụng bacteriocin là nó sẽ
giảm mầm bệnh nguy hiểm của con người chống cự lại thuốc kháng sinh.
Các nhà nghiên cứu thuộc Đại học Tổng hợp Klemson vừa phát hiện ra một
loại thuốc chống vi khuẩn mới không gây tính lờn thuốc từ chính những vi khuẩn
tạo ra loại pho mát Thụy Sĩ. Những vi khuẩn này tiết ra một loại protein có tên
bacteriocin, tạo nên tính chất của pho mát. Các thử nghiệm sơ bộ cho thấy chúng có
tác dụng chống lại hơn 150 loại vi khuẩn thường gặp.
Năm 2009, pinto và cộng sự đã nghiên cứu mô tả đặc tính chống vi khuẩn
Listeria của bacteriocin phân lập từ động vật có vỏ, kháng sinh tiềm năng kiểm soát
chất lượng thủy sản không lên men.
Tháng 11/2009, Satish Kumar R, Arul V thuộc khoa Công nghệ sinh học
trường đại học Pondicherry, Ấn Độ đã tách chiết một loại bacteriocin từ chủng
Streptococus phocae PI80 phân lập bởi tôm thẻ chân trắng Ấn Độ được đặt tên
phocaecin PI80. Chất kháng khuẩn này nhạy cảm với các enzyme: trypsin,
proteinase, pepsin, chymotrypsin. Có hoạt động ức chế một số tác nhân gây bệnh
quan trọng như: Listeria monocytogenes, Vibrio parahaemolyticus, V.fischeri.
Tháng 2/2011, Ozdemir cùng cộng sự thuộc khoa Công nghệ sinh học,
trường đại học Adnan Menderes, Thổ Nhĩ Kỳ đã xây dựng một bộ sưu tập gồm 57
chủng Enterococcal phân lập từ các nguồn gốc khác nhau (bao gồm nước sông, xử
lý cây trồng, nước thải, đất, động vật, và các loại rau) được sử dụng để sản xuất
bacteriocins. Enterococcin được xác định ở các cấp độ loài như Enterococcus
faecium, E. hirae, E. casseliflavus, E. durans, E. faecalis, E. mundtii và E. avium.
40 chủng trong 57 chủng phân lập được có khả năng ức chế sự tăng trưởng của ít
nhất một loại vi khuẩn chỉ thị.
Gần đây vào tháng 6/2012, Samar L cùng cộng sự thuộc phòng thí nghiệm
ERT 62, trường đại học Biopharma Ambrilia S.A, Pháp đã tìm ra một loại
bacteriocin mới lacticin LC14 sản sinh bởi Lactococcus lactis BMG6.14 phân lập từ
môi trường nước thịt kết tủa bởi ammonium sulfate. Lacticin LC14 cho thấy hoạt
động diệt khuẩn, loại kháng sinh chống lại một số vi khuẩn axit lactic và các chủng
SVTH: Nguyễn Thị Thảo Trang

32
gây bệnh bao gồm cả vi khuẩn Listeria monocytogenes. Nó bị bất hoạt bởi
proteinase K và E pronase, nhưng kháng papain, lysozyme, lipase và catalase.
Lacticin LC14 có khả năng chịu nhiệt, ổn định trong một phạm vi rộng của độ pH
(2-10) rất có khả năng ứng dụng làm probiotic và điều trị bệnh.
Cũng như các nước trên thế giới, các nhà khoa học Việt Nam cũng rất quan
tâm đến bacteriocin cũng có nhiều công trình khoa học nghiên cứu trong nhiều lĩnh
vực, được quan tâm nhiều nhất vẫn là ứng dụng trong công nghiệp thực phẩm. Song
các công trình nghiên cứu này vẫn còn hạn chế về nhiều mặt.
Năm 2006, Phạm Thùy Linh nghiên cứu khả năng tạo chất diệt khuẩn
enterocin P tái tổ hợp nhằm ứng dụng trong bảo quản thực phẩm. Đây là nghiên cứu
đầu tiên về tạo bacteriocin tái tổ hợp một cách có hệ thống tại Việt Nam. Sản phẩm
của nghiên cứu này là protein HisentP tái tổ hợp có hoạt tính kháng khuẩn và
các đặc tính sinh hóa tương đồng với enterocin P tự nhiên, bước đầu đã có khả năng
kéo dài thời gian bảo quản thực phẩm và có thể tiếp tục nghiên cứu phát triển làm
phụ gia sinh học dùng cho bảo quản.
Năm 2008, Nguyễn Thúy Hương đã nghiên cứu việc cố định tế bào vi khuẩn
Lactococcus lactic trên chất mang cellulose vi khuẩn (Bacterial Cellulose - BC) để
ứng dụng lên men thu nhận bacteriocin. Kết quả thu được như sau: Hiệu quả
sử dụng chế phẩm tế bào vi khuẩn cố định trên BC để lên men thu nhận
bacteriocin khá cao, có thể tái sử dụng 9 - 10 lần mà vẫn đảm bảo về mặt thời gian
lên men, số lượng và chất lượng bacteriocin so với đối chứng. Kết quả thu được
cũng góp phần thăm dò 2 ứng dụng mới của cellulose vi khuẩn (BC): sử dụng BC
làm chất mangtrong kỹ thuật cố định tế bào vi sinh vật và sử dụng màng mỏng BC
làm màng baothực phẩm.
Các nghiên cứu này đóng góp rất lớn trong việc mở rộng lĩnh vực tìm hiểu
về bacteriocin. Trong tương lại, sẽ có nhiều công trình nghiên cứu hơn về
bacteriocin để vạch ra một hướng đi mới cho chúng.
SVTH: Nguyễn Thị Thảo Trang

33
Chương II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU2.1 Nguyên liệu
2.1.2 Mẫu tôm hùm
Mẫu tôm hùm được thu mua ở trại tôm Bình Ba, Khánh Hòa. Tôm hùm sử
dụng cho thí nghiệm có kích thước khoảng 27cm, nặng 600g. Mẫu tôm được bảo
quản sống đưa về phòng thí nghiệm và tiến hành phân lập vi sinh vật từ máu, nội
tạng, và cơ thịt.
2.1.3 Chủng chỉ thị
Các chủng chỉ thị là chủng Bacillus B1.1 đại diện cho nhóm vi khuẩn Gram
dương và Vibrio V1.1 đại diện cho nhóm vi khuẩn Gram âm. Các chủng này được
lấy từ bộ sưu tập vi sinh vật của phòng thí nghiệm Công nghệ sinh học, trường Đại
học Nha Trang.
Các chủng Bacillus B1.1 và Vibrio V1.1 đều được nuôi cấy trên môi trường
TSB, pH 7,0 ± 0,2, lắc 180 vòng/phút trong 7 - 8 giờ.
Sau khi nuôi dịch đủ thời gian, hút 100µl dịch vi khuẩn để giữ trong dung
dịch glycerol 30%, nhằm mục đích bảo quản để sử dụng trong thời gian dài ở -800C.
Trước khi thử đối kháng, chuẩn bị chủng bằng cách lấy ống giống được bảo
quản, hút 100µl dịch vi khuẩn đem nuôi cấy ở những điều kiện thích hợp trên môi
trường nuôi trong vòng 18-24 giờ.
2.1.4 Môi trường hóa chất và thuốc thử
2.1.4.1 Môi trường phân lập, nuôi cấy vi khuẩn
Môi trường TSB (pH 7,3 ± 0,2; 1,5% NaCl)
Hòa tan 30g môi trường TSB tổng hợp, bổ sung thêm 10g NaCl (để đạt nồng
độ cuối là 1,5%) trong 1000ml nước cất, chuẩn pH = 7, sau đó đem rót môi trường
vào các bình tam giác (ống nghiệm), làm nút bông và giấy bạc, hấp khử trùng 1210C
trong thời gian 15 phút.
SVTH: Nguyễn Thị Thảo Trang

34
2.1.4.2 Môi trường thử hoạt tính TSA
Môi trường TSA được pha từ môi trường TSB có bổ sung thêm 15g agar và
10g NaCl trong 1000ml. Sau đó đun nóng rồi phân phối vào bình tam giác. Hấp khử
trùng 1210C trong thời gian 15 phút. pH cuối 7,3 ± 0,2.
2.1.4.3 Dung dịch nước muối sinh lý
Hòa tan 8,5g NaCl trong 1 lít nước cất. Hấp khử trùng ở nhiệt độ 1210C
trong 15 phút rồi để nguội.
2.1.4.4 Thuốc nhuộm tím Violet
Hòa tan 1g tím violet vào trong 10ml cồn (dung dịch 1)
Hòa tan 2g phenol tinh thể vào trong 10ml nước cất (dung dịch 2)
Trộn chung dung dịch 1 và dung dịch 2 lại với nhau ta có dung dịch thuốc
nhuộm tím violet.
2.1.4.5 Thuốc nhuộm Liugol
Hòa tan iod vào nước cất, sau đó cho KI vào, bảo quản trong lọ tối màu.
2.1.4.6 Thuốc nhuộm Fuschin
Hòa tan Fuschin vào trong 10ml cồn 95% (dung dịch 1).
Hòa tan phenol tinh thể vào 100ml nước cất (dung dịch 2).
Trộn đều dung dịch 1 và dung dịch 2 đem lọc, ta có thuốc nhuộm fuschin.
2.1.4.7 Dung dịch xanh methylen
Dung dịch này bao gồm 25g xanh methylen và 300ml rượu ehtylic.
Được pha chế như sau:
Hòa tan xanh methylen vào rượu rồi lọc (dung dịch 1).
Dịch lọc 1 thêm 1ml KOH 1% và 1000ml nước cất.
2.1.4.8 Thuốc thử Catalase
Dung dịch 30% H2O2 (hydrogen peroxide)
2.1.5 Thiết bị chuyên dụng
- Cân phân tích BL-3200H (L) (Shimadzu, Nhật Bản)
- Máy dập mẫu Smasher (Việt Nam)
- Máy vortex (máy trộn mẫu BE34)
SVTH: Nguyễn Thị Thảo Trang

35
- Thiết bị ly tâm (Eppendorf Centrifuge 5417R, Mỹ)
- Tủ sấy (Binder, Đức)
- Tủ cấy (Telstar AV 100, OSI Co., Ltd, Đài Loan)
- Lò vi sóng (LG, Hàn Quốc)
- Máy lắc (GFL 3005, Đức)
- Tủ ấm Binder BF115 (Binder, Đức)
- Nồi hấp khử trùng autoclave (Sturdy industrial Co., Ltd, Đài loan)
- Kính hiển vi 3 mắt ngắm có camera và máy tính (Motic BA 300 – Mỹ)
- Tủ lạnh (NANO silver, Việt Nam)
2.2 Phương pháp nghiên cứu
Đề tài sử dụng cách tiếp cận kế thừa và chọn lọc các phương pháp phân lập
tuyển chọn các chủng có khả năng sinh bacteriocin, sơ đồ quy trình nghiên cứu dự
kiến như sau:
SVTH: Nguyễn Thị Thảo Trang

Chọn khuẩn lạc đặc trưng
Cấy phân lập
Mẫu tôm
Pha loãng
Nuôi trong môi trường lỏng
Đồng nhất mẫu
Dịch huyền phù
Xác định hoạt tính bacteriocin
Xác định đặc điểm, phân loại chủng vi khuẩn sinh bacteriocin
Ly tâm thu dịch
Xác định hoạt tính kháng khuẩn của chủng vi khuẩn
36
Hình 2.1: Quy trình thực hiện
SVTH: Nguyễn Thị Thảo Trang
9 ml 9 ml 9 ml
0,1 ml0,1 ml0,1 ml0,1 ml
9 ml
0,1 ml0,1 ml0,1 ml0,1 ml
10-5
10-4
10-310-2
10-1
37
Các mẫu máu, nội tạng, và cơ thịt của tôm hùm được tiến hành phân lập độc
lập. Tiến hành đồng nhất mẫu lấy dịch huyền phù để phục vụ cho việc cấy phân lập
tạo khuẩn lạc đặc trưng. Sau quá trình tuyển chọn được khuẩn lạc riêng rẽ, vi khuẩn
được nuôi cấy trên môi trường lỏng, rồi đem ly tâm thu dịch. Lấy dịch ly tâm đi
thực hiện quá trình thử hoạt tính kháng khuẩn của các chủng vi khuẩn phân lập
được với chủng chỉ thị. Khi đã tuyển chọn được chủng vi khuẩn sinh kháng khuẩn,
tiếp tục xác định hoạt tính bacteriocin bằng cách sử dụng enzyme proteinase K và
trypsin. Sau đó, sử dụng chủng sinh bacteriocin để xác định đặc điểm và phân loại
chủng vi khuẩn sinh bacteriocin.
2.2.1 Phương pháp phân lập vi sinh vật
Mục đích của thí nghiệm tạo bộ sưu tập các chủng vi sinh vật có trong mẫu
tôm. Thí nghiệm tiến hành như sau:
2.2.1.1 Xử lý mẫu
Mẫu tôm hùm đem cân khối lượng, sau đó thêm vào dung dịch muối sinh lý
đã được khử trùng với tỷ lệ 1:9. Chuẩn bị mẫu xong cho vào túi PE, có chứa mẫu và
dung dịch nước muối sinh lý, đem đi đồng nhất bằng máy dập mẫu. Thời gian đồng
nhất từ 90 – 120 giây. Sau khi đồng nhất, thu được dung dịch huyền phù.
2.2.1.2 Phân lập
Pha loãng mẫu và cấy đĩa
Tiến hành pha loãng dịch huyền phù trên đến nồng độ 10-5 bằng dung dịch
muối sinh lý đã được hấp khử trùng. Trình tự thao tác được trình bày qua hình sau:
Hình 2.2: Trình tự pha loãng
SVTH: Nguyễn Thị Thảo Trang
38
Trước khi tiến hành, chuẩn bị các ống nghiệm vô trùng, mỗi ống chứa 9ml
dung dịch nước muối sinh lý đã hấp khử .
Lấy dịch mẫu từ túi PE, dịch mẫu có nồng độ 10-1, hút 1ml dịch cho vào 9ml
muối sinh lý đã hấp khử trùng ở ống nghiệm khác, thu được dịch pha loãng có nồng
độ thấp hơn nồng độ ban đầu 10-2, đưa lên máy votex để trộn đều. Tiếp tục làm
tương tự, ta có các ống nghiệm có độ pha loãng mẫu là 10-3, 10-4, 10-5.
Sau đó chuẩn bị môi trường TSA, hấp khử trùng ở 1210C, 15 phút. Chờ môi
trường nguội, rồi phân phối vào các đĩa petri (đã sấy ở 1600C trong 90 phút), mỗi
đĩa 15-20ml. Dùng pipet man hút 0,1ml dịch đã pha loãng ở các nồng độ cho vào
các đĩa thạch đã ghi tên mẫu, nồng độ và ngày phân lập. Dùng que cấy trang đã khử
trùng trang đều lên bề mặt thạch cho đến khi thấy mặt thạch khô. Sau đó, lật ngược
đĩa lại cất và nuôi cấy mẫu phân lập ở 370C. Sau 48 giờ vi khuẩn đã phát triển trên
đĩa thạch tiến hành quan sát, tách và tạo khuẩn lạc thuần.
Tách và tạo khuẩn lạc thuần
Khi khuẩn lạc xuất hiện, chọn những khuẩn lạc riêng rẽ trên đĩa peptri rồi
cấy vào ống nghiệm, mỗi khuẩn lạc cấy vào một ống (kí hiệu đầy đủ), đặt vào tủ ấm
370C. Sau 24 giờ, dùng que cấy ria thu sinh khối khẩn lạc đã chọn, cấy ria trên đĩa
thạch cho đến khi tạo các khuẩn lạc thuần, tiến hành cấy ria nhiều lần. Sau khi cấy,
nuôi trong tủ ấm 370C trong vòng 24 giờ. Khi tạo được khuẩn lạc thuần nhất và
tách rời trên đĩa thạch thì lấy khuẩn lạc này cấy vào ống nghiệm để giữ giống.
Giữ giống và cấy chuyền
Chọn khuẩn lạc riêng rẽ cấy vào ống thạch nghiêng, tiến hành cấy ziczac trên
bề mặt thạch nghiêng để được giống thuần. Các chủng vi khuẩn đã được lựa chọn
cấy trên môi trường thạch nghiêng TSA, được đặt vào tủ ấm 370C. Sau 10-14 ngày
khi vi sinh vật đã phát triển tốt, các ống giống được bảo quản trong tủ lạnh ở 4 –
60C. Sau 2-3 tháng cấy chuyền lại một lần. Để bảo quản được lâu hơn ta có thể giữ
giống trong paraffin lỏng vô trùng hay giữ trong glycerol hoặc đông khô.
Nuôi cấy khuẩn lạc thuần trong các bình tam giác chứa môi trường TSB
trong 7 – 10 giờ. Chia nhỏ huyền phù tế bào vào từng ống eppendorf 1,5ml, sau đó
SVTH: Nguyễn Thị Thảo Trang

39
thêm vào glycerol theo tỷ lệ 7:3. Votex nhẹ cho đều hỗn hợp, để vào tủ lạnh ở ngăn
mát trong 3 giờ rồi đem bảo quản ở - 800C.
2.2.2 Tuyển chọn chủng sinh bacteriocin
Mục tiêu của thí nghiệm là tuyển chọn được các chủng sinh bacteriocin từ bộ
sưu tập chủng phân lập được.
2.2.2.1 Xác định hoạt tính kháng khuẩn của các chủng
Để thử tính đối kháng của các chủng vi khuẩn phân lập được đối với 2 chủng
chỉ thị là V1.1 và B1.1, tiến hành xác định hoạt tính kháng khuẩn theo phương pháp
khếch tán thạch.
Dịch vi khuẩn sau khi nuôi trong 18 – 24 giờ, tiến hành ly tâm thu dịch
bacteriocin thô và điều chỉnh pH về pH 7 – 8 để loại bỏ tác dụng của acid do vi
khuẩn lên men sinh ra có trong dịch, sau đó tiến hành thử hoạt tính với các chủng
chỉ thị.
Hoạt tính kháng khuẩn được xác định bằng phương pháp khếch tán trên
thạch. Đối với phương pháp này chủng vi khuẩn thử nghiệm và chủng chỉ thị cùng
được nuôi trên một môi trường, và tính đối kháng thể hiện ở việc chất ức chế của
chủng thử nghiệm khếch tán vào môi trường và gây ức chế chủng chỉ thị. Sử dụng
đĩa thạch đã nuôi cấy chủng chỉ thị, sau đó tạo các giếng trên thạch bằng cách đục
lỗ, tra dịch chủng thử nghiệm đã được ly tâm và ủ ở nhiệt độ thích hợp. Phương
pháp này thích hợp với kiểm tra những khuẩn lạc riêng rẽ và áp dụng rộng rãi trong
nghiên cứu về mặt di truyền.
Quy trình được tóm tắt theo sơ đồ sau
SVTH: Nguyễn Thị Thảo Trang

40
Hình 2.3: Quy trình thử tính kháng khuẩn của dịch bacteriocin thô với chủng chỉ thị
Chuẩn bị môi trường TSB lỏng, rót 20ml môi trường vào các bình tam giác
có dung tích 100ml, đem hấp khử trùng 1210C trong 15 phút.
Lấy 1 ống vi khuẩn chủng đích hút 1ml dịch vi khuẩn hút vào các bình tam
giác chứa môi trường đã chuẩn bị, nuôi cấy trên máy lắc 180 vòng/phút trong 18 –
20 giờ. Đồng thời, chuẩn bị dịch vi khuẩn cho 2 chủng chỉ thị. Tương tự với vi
khuẩn thử nghiệm, nuôi cấy trên máy lắc180 vòng/phút trong 16 – 20 giờ. Sau đó
tiến hành thử hoạt tính.
Dùng dịch vi khuẩn thử nghiệm đã nuôi cấy qua đêm chia nhỏ dịch huyền
phù vào từng ống eppendorf vô trùng 1,5ml để ly tâm 5000 vòng/phút trong thời
gian 30 phút, ở 40C để loại bỏ sinh khối tế bào và thu dịch nổi. Hút dịch nổi và đo
pH ban đầu, rồi tiến hành chuẩn độ pH bằng NaOH 0,5N để đưa về pH 7. Bước này
nhằm mục đích loại bỏ sự ảnh hưởng của acid do vi khuẩn tiết ra. Nếu pH kiềm thì
dùng HCl 0,5N để chuẩn độ về 7.
SVTH: Nguyễn Thị Thảo Trang
Dịch vi khuẩn nuôi cấy sau 16 – 20 giờ
Hút 1,5ml dịch vi khuẩn vào eppendolf vô trùng
Ly tâm 5000 vòng/phút, 30 phút, 40C
Chuẩn độ dịch đưa về pH = 7
Thử tính kháng khuẩn
Sau 16 – 24 giờ đọc kết quả
Dịch ly tâm

Đĩa petri
Khuẩn lạc mọc trên môi trường thạch
Lỗ chứa dịch bacteriocin thô
Vùng kháng khuẩn ( không có khuẩn lạc)
41
Tiếp theo, tiến hành thử hoạt tính bằng phương pháp khếch tán trên đĩa
thạch. Phương pháp này dựa trên khả năng đối kháng giữa vi sinh vật thử nghiệm và
vi sinh vật chỉ thị. Chuẩn bị các đĩa thạch môi trường TSA, hút 100µl dịch vi khuẩn
chỉ thị được pha loãng theo tỉ lệ 1:9 với nước muối sinh lý trang đều lên bề mặt
thạch cho đến khi bề mặt thạch khô. Sau đó, đục 4 – 6 lỗ, với đường kính 5mm trên
bề mặt đĩa thạch. Các lỗ đục cách thành đĩa petri khoảng 1cm để có thể quan sát
được vòng kháng khuẩn sau này. Dùng micropipet hút 150µl dịch bacteriocin thô
cho vào mỗi lỗ thạch, hút 150µl nước cất đã hấp khử trùng ở 1210C cho vào lỗ làm
mẫu đối chứng. Tiếp theo, bảo quản các đĩa ở 2-40C trong 2 giờ với mục đích ngăn
cản sự phát triển của chủng chỉ thị và tạo điều kiện cho bacteriocin có thời gian
thẩm thấu vào bên trong thạch. Sau đó, các đĩa được ủ ở 370C.
Sau 16 -24 giờ thì đọc kết quả. Khả năng kháng khuẩn của vi khuẩn được
xác định bằng sự hiện diện của vòng kháng khuẩn xung quanh lỗ. Nếu đường kính
vùng kháng khuẩn lớn hơn hoặc bằng 6mm thì được xem là có khả năng ức chế tích
cực.
Đường kính vùng kháng khuẩn được tính như sau:
H = D – d
Trong đó H: đường kính vùng kháng khuẩn xung quanh lỗ
D: đường kính vùng ức chế (bao gồm đường kính lỗ)
d: đường kính lỗ
Hình 2.4: Vùng kháng khuẩn của dịch bacteriocin trên đĩa petri
SVTH: Nguyễn Thị Thảo Trang

Chuẩn độ dịch về pH = 7
Bổ sung Catalase, ủ 370C, 30 phút
Dịch ly tâm
Thử tính kháng khuẩn
Bổ sung proteinase K, ủ 370C, 2 giờ Bổ sung trypsin, ủ 370C, 2 giờ
Sau 16 – 24 giờ đọc kết quả
42
2.2.2.2 Xác định hoạt tính bacteriocin
Để kiểm tra bản chất của chất kháng khuẩn trong dịch bacteriocin thô của vi
khuẩn thử nghiệm có phải là protein không, tiến hành xác định hoạt tính bacteriocin
với enzyme proteinase K và enzyme trypsin.
Bacteriocin có bản chất là protein nên dưới tác dụng của enzyme proteinase
K, enzyme trypsin và một số enzyme khác như lipase, papain… sẽ làm bacteriocin
bị thủy phân, mất hoạt tính kháng khuẩn với các chủng chỉ thị.
Quy trình được tiến hành như sau
Hình 2.5 : Sơ đồ bố trí thí nghiệm xác định hoạt tính bacteriocin
Sử dụng dịch vi khuẩn thử nghiệm đã ly tâm và được chuẩn độ về pH = 7
bằng NaOH 0,5N hoặc HCl 0,5N.
Pha sẵn enzyme proteinase K, enzyme trypsin và catalase với nồng độ
20mg/ml, đem bảo quản lạnh ở -210C, khi cần dùng thì mới lấy ra hút nhanh cho
vào dịch bacteriocin thô.
SVTH: Nguyễn Thị Thảo Trang

Nuôi cấy chủng vi sinh vật được tuyển chọn
Quan sát hình thái tế bào vi khuẩnTest sinh hóa
Phân loại bằng kỹ thuật phân tử
43
Bổ sung enzyme catalase vào dịch bacteriocin thô cho đạt nồng độ cuối cùng
là 0,5ml, ủ ở 370C trong 30 phút, để loại trừ khả năng ức chế của hydrogen
peroxide. Sau đó, bổ sung các enzyme proteinase K, enzyme trypsin vào dịch
bacteriocin thô tới nồng độ enzyme cuối cùng là 1mg/ml. Mẫu đã xử lý enzyme
được ủ ở 370C trong 2 giờ.
Song song với việc chuẩn bị mẫu đối chứng, là dịch không đem xử lý
enzyme, để ở nhiệt độ phòng để dễ dàng giúp ta nhận biết được khả năng kháng
khuẩn của nó qua kích thước vòng kháng khuẩn.
Sử dụng dịch đã xử lý để tiến hành thử enzyme cũng bằng phương pháp
khếch tán trên thạch, với môi trường thạch TSA và 2 chủng chỉ thị V1.1 và B1.1.
Phương pháp tiến hành như mục 2.2.2.1. Ủ đĩa ở 370C trong 16-24 giờ thì kiểm tra
sự tạo thành vòng kháng khuẩn và đo kích thước vòng kháng.
2.2.3 Xác định đặc điểm của chủng vi khuẩn sinh bacteriocin
Xác định đặc điểm của các chủng vi sinh vật kiểm định cũng chính là
phương pháp định danh vi sinh vật theo phương pháp truyền thống dựa trên các đặc
điểm về hình thái, đặc điểm sinh lý, sinh hóa. Dựa vào đó, ta có thể phân loại
chúng.
Sơ đồ bố trí thí nghiệm
Hình 2.6: Sơ đồ bố trí thí nghiệm xác định đặc điểm vi khuẩn
2.2.3.1 Quan sát hình thái tế bào vi khuẩn, nhuộm Gram
SVTH: Nguyễn Thị Thảo Trang

44
Để xác nhận vi khuẩn kiểm định là vi khuẩn Gram dương hay vi khuẩn Gram
âm, tiến hành quan sát tế bào vi khuẩn.
Dựa trên khả năng bắt màu của tế bào chất và màng tế bào với thuốc nhuộm.
Vi khuẩn Gram (-) bắt màu hồng của thuốc nhuộm fuchsin, vi khuẩn Gram (+) bắt
màu tím của violet.
Tiến hành nhuộm như sau:
Chuẩn bị lam kính, dùng bút viết kính đánh dấu giới hạn vùng phết vi khuẩn
mặt sau lam kính. Sau đó, nhỏ 1 giọt nước cất vào vòng tròn đã vẽ.
Dùng que cấy vô trùng lấy một ít khuẩn lạc đại diện cho vi khuẩn kiểm định
từ thạch, rồi đưa vào lam kính, dàn đều vi khuẩn, để khô tự nhiên hoặc hơ nhẹ trên
ngọn đèn cồn, tránh để lâu quá, nóng sẽ vỡ tế bào vi khuẩn, gây sai lệch kết quả.
Tiến hành nhuộm, đầu tiên nhỏ vài giọt tím tinh thể (tím violet) lên tiêu bản,
để trong 30 giây đến 1 phút. Nghiêng đổ rồi rửa lại bằng nước cất. Tiếp đó, nhuộm
màu tiêu bản bằng thuốc nhuộm Liugol trong thời gian 30 giây đến 1 phút, rồi rửa
nhẹ với nước bằng bình tia. Rồi rửa tiêu bản vừa nhuộm với cồn 900 cho đến khu
phiến kính bạc màu trong khoảng 10 – 15 giây, rồi rửa lại với nước.
Nhuộm tiếp tiêu bản với Fuchsin trong thời gian 30 giây đến 1 phút, rồi rửa
lại với nước. Để khô tự nhiên hoặc dùng giấy thấm lau tiêu bản cho sạch xung
quanh vùng vi khuẩn, sau đó đưa lên vật kính dầu quan sát X – 100.
2.2.3.2 Test sinh hóa
Phương pháp xác đinh khả năng chịu muối
Tiến hành xác định khả năng chịu muối nhằm mục đích là biết được khoảng
giới hạn nồng độ muối mà vi khuẩn vẫn sống và phát triển.
Thí nghiệm được tiến hành như sau:
Chuẩn bị các đĩa môi trường TSA bổ sung muối với các nồng độ từ 1 – 12%,
đã được hấp khử trùng ở 1210C, trong 15 phút. Dùng que cấy vòng vô trùng thu
sinh khối dịch vi khuẩn, cấy ria lên các đĩa môi trường theo từng nồng độ. Sau đó,
lật úp đĩa lại giữ trong tủ ấm.
SVTH: Nguyễn Thị Thảo Trang

45
Sau 24 giờ, quan sát kết quả. Nếu vi khuẩn vẫn phát triển chứng tỏ chúng có
khả năng chịu muối.
SVTH: Nguyễn Thị Thảo Trang

46
Thử catalase
Thí nghiệm được tiến hành nhằm xác định sự có mặt của enzyme catalase
trong dịch nuôi cấy. Thường được dùng để phân biệt vi khuẩn hiếu khí với vi khuẩn
kỵ khí.
Các vi sinh vật hiếu khí và kỵ khí tùy tiện chứa chuỗi truyền điện tử có
cytochrome đều có enzyme catalase, có khả năng biến dưỡng năng lượng theo
phương thức hô hấp với oxy là chất nhận điện tử cuối cùng trong chuỗi truyền điện
tử tạo ra H2O2. Catalase sẽ thủy phân hydrogen peroside (H2O2), thành H2O và O2,
ngăn cản sự tích tụ phân tử có độc tính cao này trong tế bào.
Sự thủy phân H2O2 sẽ giải phóng O2 được ghi nhận qua hiện tượng sủi bọt
khí.
Tiến hành thử catalase như sau:
Môi trường TSB lỏng được pha chế cho ống nghiệm mỗi ống chứa 10ml môi
trường và hấp khử trùng theo phương pháp thông thường. Sau khi làm nguội môi
trường, cấy vi khuẩn vào, nuôi ở 370C. Sau một ngày, nhỏ 2 giọt H2O2 30% vào
dịch nuôi cấy.
Thử nghiệm là (+) khi có hiện tượng sủi bọt khí do O2 được tạo ra, ngược lại
là (-) khi không có sủi bọt khí.
Thử nghiệm tính di động
Vi sinh vật di động chủ yếu nhờ các tiêm mao (flagella) có nhiều ở vi khuẩn
hình que, một số vi khuẩn hình cầu. Khả năng di động là một là một trong những
căn cứ có thể dùng để phân biệt vi sinh vật. Khả năng này có thể được quan sát dựa
vào sự tăng trưởng và di động của vi sinh vật vào bên trong môi trường thạch mềm.
Tiến hành thí nghiệm:
Môi trường thử là môi trường thạch TSA, rót môi trường vào ống nghiệm
(10ml/ống), hấp khử trùng như thông thường. Làm nguội môi trường ở trạng thái
đứng. Sau đó dùng que cấy đâm xuyên vô trùng thu sinh khối từ khuẩn lạc, cấy
thẳng khuẩn lạc vào tâm môi trường trong ống nghiệm, độ sâu khoảng 2cm. Ủ các
ống nghiệm đã cấy trong tủ ấm ở 370C. Quan sát kết quả sau 24 giờ.
SVTH: Nguyễn Thị Thảo Trang

47
Thử nghiệm là (+) khi vi sinh vật mọc lan ra khỏi đường cấy và làm đục môi
trường xung quanh; là (-) khi vi sinh vật chỉ mọc dọc theo đường cấy trong khi môi
trường xung quanh vẫn trong. Ống đối chứng không có vi sinh vật tăng trưởng, môi
trường trong.
Khả năng sinh hơi
Khi sử dụng nguồn carbon để lên men, tùy phương thức lên men sẽ tạo ra
được các sản phẩm khác nhau bao gồm rượu, các acid hữu cơ, CO2... Trường hợp,
CO2 được tạo thành sẽ được bẫy lại thành bọt khí trong ống chuông Durham làm nổi
ống chuông này (trường hợp môi trường lỏng), hoặc làm vỡ thạch khi cấy trong ống
thạch sâu (trường hợp sử dụng môi trường rắn).
Để biết được sản phẩm cuối cùng của quá trình lên men có CO2 không, tiến
hành thí nghiệm xác định khả năng sinh hơi.
Chuẩn bị môi trường TSB, phân phối vào các ông nghiệm, đem hấp khử
trùng ở 1210C trong 15 phút. Chờ môi trường nguội, dùng que cấy thu sinh khối từ
khuẩn lạc cấy vào môi trường, sau đó cho các ống Durham vô trùng vào ống
nghiệm, đặt nhẹ nhàng tránh tạo khí trong ống Durham. Các ống nghiệm này sẽ
được đem ủ trong tủ ấm 370C. Sau 18 – 24 giờ, xem kết quả.
Thử nghiệm là (+) nếu ống Durham nổi lên trên bề mặt môi trường.
2.2.3.3 Định danh bằng phương pháp sinh học phân tử
Tách chiết DNA hệ gen
Tiến hành tách chiết DNA để thu được dịch DNA hệ gen tinh sạch có nồng
độ đáp ứng cho yêu cầu chạy PCR
Quá trình tách chiết được tiến hành như sau :
Các chủng vi khuẩn được nuôi trên môi trường TSB ở nhiệt độ 37OC trong
24 giờ. Dịch nuôi cấy được ly tâm ở 13.000 vòng/phút trong 2 phút ở 4OC để thu tế
bào (Luan et al., 2007). Sau đó, DNA tổng số được tách chiết bằng bộ kit Wizard®
SV Genomic DNA System (Promega) theo hướng dẫn của nhà sản xuất và bảo quản
ở -20OC.
SVTH: Nguyễn Thị Thảo Trang

48
Phản ứng PCR
Thực hiện phản ứng PCR nhắm mục đích nhân lên đoạn gen mong muốn với
số lượng lớn.
Phương pháp PCR được Kary Mullis phát minh vào năm 1985. PCR là
phương pháp dùng enzyme khuếch đại chọn lọc một đoạn DNA theo luật số mũ.
Phản ứng nhân gen được thực hiện với enzyme Taq polymerase hoạt động tổng hợp
mạch DNA mới từ khuôn và mồi đặc hiệu, số lượng DNA đích sẽ tăng theo mỗi chu
kì của phản ứng.
Mỗi chu kì phản ứng gồm 3 bước :
- Bước 1 : phân tử DNA biến tính ở nhiệt độ cao 91 – 970C , làm mạch kép
tách thành 2 mạch đơn
- Bước 2 : nhiệt độ hạ thấp đến 40 – 650C, cho phép các đoạn mồi
oligonucleotide ngắn gắn với các chuỗi DNA đơn.
- Bước 3 : kéo dài mạch ở nhiệt độ 68–730C, nhờ enzyme Taq polymerase
(thermostable DNA polymerase) để kéo dài các dNTP lại đầu 3’ của đoạn mồi đang
bắt cặp trên đầu 5’ của sợi DNA đích để bắt đầu cho sự tổng hợp nên mạch bổ sung.
Sau mỗi chu kì, số lượng các bản sao DNA tăng lên gấp đôi. Theo lý thuyết,
kết quả sau n chu kì phản ứng sẽ có 2n bản sao phân tử DNA mạch kép nằm giữa 2
đoạn mồi.
Tiến hành phản ứng PCR :
Cặp mồi sử dụng cho phản ứng PCR được tổng hợp bởi IDT có trình tự như
sau:
- 16S – 27F : 5’ – AGA GTT TGA TCC TGG CTC AG – 3’
- 16S – 1492R : 5’ – ACG GCT ACC TTG TTA CGA CT – 3’
Phản ứng PCR được tiến hành với tổng thể tích 50µl với các thành phần ở
nồng độ cuối như nhau : DNA (2-20 ng), mỗi mồi (0,1 µM), dNTP (0,1 mM mỗi
loại), MgCl2 (2 mM), Taq polymerase (1,25 U) và đệm Tag (1X).Chu trình nhiệt
của phản ứng bao gồm :
SVTH: Nguyễn Thị Thảo Trang

49
Bước 1 940C 3phút
940C 1phút
Bước 2 500C 1phút 35 chu kỳ
720C 1phút
Bước 3 720C 7phút
Bước 4 40C ∞
Sản phẩm PCR được điện di trên gel agarose 1-1,5% trong đệm TBE 0,5X,
sau đó được nhuộm với ethidium bromide và đọc kết quả trên máy chụp ảnh gel
(Biorad).
Giải và phân tích trình tự
Sản phẩm PCR được tinh sạch bằng bộ kit PCR clean up system (Promega)
theo hướng dẫn của nhà sản xuất và sử dụng làm khuôn trực tiếp cho phản ứng tiền
giải trình tự theo nguyên tắc dye-labelled dideoxy terminator (Big Dye Terminator
v. 3.1, Applied Biosystems) với các cặp mồi 16S-27F/16S-1492R theo chương trình
nhiệt như sau: 96°C trong 20 giây, 50°C trong 20 giây và 60°C trong 4 phút. Sản
phẩm phản ứng được phân tích trên máy phân tích trình tự tự động ABI Prism 3700
DNA Analyser (Applied Biosystems).
Các trình tự được phân tích, chỉnh sửa bằng phần mềm BioEdit 7.1.3.0 và
Genious Pro 5.5.7, và được so sánh với các trình tự nucleotide tương đồng trên
Genbank bằng chương trình BLAST (www. ncbi . nlm . nih . gov / blast / ).
SVTH: Nguyễn Thị Thảo Trang

50
Chương III. KẾT QUẢ VÀ THẢO LUẬN3.1 Phân lập chủng vi sinh vật từ mẫu tôm Hùm
Tiến hành phân lập trên môi trường TSB : từ 3 mẫu tôm giống phân lập được
46 chủng vi khuẩn .
Bảng 3.1: Chủng vi khuẩn phân lập được
STT Mẫu Ngày phân lập Số chủng phân lập
1 Tôm giống 1 22/02/2012 21 (N1 – N21)
2 Tôm giống 2 09/04/2012 12 (N22 – N33)
3 Tôm giống 3 16/04/2012 13 (N34 – N46)
Hình 3.1: Các chủng vi khuẩn ở nồng độ 10-3
Sau khi nuôi cấy vi khuẩn ở các nồng độ pha loãng khác nhau trên môi
trường TSA, kết quả cho thấy tổng mật độ vi khuẩn hiếu khí là 6,7 x 105 CFU/g,
quan sát kết quả thấy xuất hiện 3 dạng khuẩn lạc chính :
SVTH: Nguyễn Thị Thảo Trang
10-3

ĐC
N9N4
N1N200
N10
N16
N12
N13
N3
N18
N14
51
- Màu vàng, trơn bóng.
- Màu trắng sữa, trơn bóng.
- Màu trắng sữa, bề mặt nhăn.
Lựa chọn khuẩn lạc riêng rẽ, tiến hành quá trình tuyển chọn chủng vi khuẩn
sinh bacteriocin.
3.2 Hoạt tính kháng khuẩn của các chủng phân lập
Để xác định được hoạt tính kháng khuẩn, việc thử nghiệm được tiến hành
trên môi tường rắn có chủng chỉ thị Vibrio V1.1 và Bacillus B1.1. Tính đối kháng
thể hiện ở việc chủng vi khuẩn khếch tán vào môi trường và gây ức chế sự phát
triển của chủng chỉ thị. Vi khuẩn sinh bacteriocin có hoạt tính đối kháng với các
chủng chỉ thị càng nhiều thì phổ kháng khuẩn càng rộng.
Hình 3.2: Kết quả của 11 chủng nổi bật đối kháng với chủng Bacillus B1.1(ĐC : đối chứng là mẫu nước cất)
Kết quả kiểm tra tính kháng khuẩn bằng phương pháp khếch tán đĩa thạch
cho thấy sau 24 giờ, vòng kháng xuất hiện xung quanh bề mặt giếng có chủng vi
khuẩn thử nghiệm trên đĩa thạch, kích thước của chúng được thể hiện ở bảng 3.2.
SVTH: Nguyễn Thị Thảo Trang

52
Chú thích bảng 3.2 : (-) : Không có hoạt tính kháng khuẩn với chủng chỉ thị
Bảng 3.2 : Hoạt tính kháng khuẩn của các chủng vi khuẩn
SVTH: Nguyễn Thị Thảo Trang
STT Chủng vi khuẩn phân lập đượcĐường kính vùng kháng khuẩn (D, mm)
Vibrio V1.1 Bacillus B1.1
1 N1 - 18
2 N3 - 22
3 N4 19 -
4 N5 13 -
5 N6 9 12
6 N7 13 -
7 N8 10 10
8 N9 - 14
9 N10 19 18
10 N11 - 9
11 N12 20 18
12 N13 - 18
13 N14 - 20
14 N15 - 12
15 N16 12 12
16 N17 8 -
17 N18 14 14
18 N19 - 8
19 N20 - 22
20 N21 15 -

53
Từ 46 chủng vi khuẩn đã được kiểm tra tính kháng khuẩn, có 20 chủng thể
hiện được hoạt tính kháng trong đó có 5 chủng đối kháng cả 2 chủng chỉ thị, có phổ
kháng khuẩn rộng (kháng được cả vi khuẩn Gram dương và Gram âm) ; 6 chủng có
vùng kháng khuẩn ≥18mm với chủng chỉ thị Bacillus B1.1 ; 3 chủng có vùng
kháng khuẩn lớn ≥18 mm với chủng chỉ thị Vibrio V1.1.
Qua đó cho thấy rằng các chủng vi khuẩn đối kháng với các chủng chỉ thị
nhờ vào các thành phần kháng khuẩn tồn tại trong dịch kháng khuẩn thô. Những
thành phần kháng khuẩn mà khi chúng sinh trưởng, phát triển có thể sẽ tạo ra như
acid, bacteriocin, CO2, H2O2. Do đó, đến đây chưa khẳng định được các chủng này
sinh bacteriocin. Chọn ra 11 chủng nổi bật, ổn định (các chủng trong ô màu ở bảng
3.2) qua các lần thử để kiểm tra tiếp.
3.3 Xác định hoạt tính bacteriocin
Sau khi tuyển chọn được các chủng có hoạt tính kháng khuẩn, để kiểm tra
dịch kháng khuẩn có phải là bacteriocin không, ta tiến hành xử lý proteinase K và
tripsin. Bacteriocin vốn có bản chất là protein, cũng có thể là hỗn hợp của protein
với các chất khác. Vì vậy để kiểm tra bản chất protein, ta sử dụng các enzyme đặc
trưng như proteinase K. Ngoài ra thì độ bền enzyme cũng là một dấu hiệu để nhận
biết bacteriocin. Mỗi loại bacteriocin có thành phần cấu trúc các aminoacid khác
nhau nên sẽ chịu sự phân cắt đặc hiệu của các enzyme khác nhau, thích hợp với nó.
Khi bị phân cắt bởi các enzyme này sẽ khiến bacteriocin mất đi hoạt tính kháng
khuẩn.
Trước khi thực hiện quá trình thử nghiệm, tiến hành thử với enzyme catalase
để loại bỏ khả năng sinh H2O2. Kết quả sau khi thử nghiệm, tuyển chọn được chủng
N14 bị bất hoạt sau khi xử lý với các enzyme proteinase K và tripsin. Do các
peptide kháng khuẩn trong dịch đã bị phân hủy bởi enzyme nên không thể kháng lại
chủng chỉ thị được nữa, hay không thể tạo vòng kháng với B1.1.
SVTH: Nguyễn Thị Thảo Trang

B1.1N14
Proteinase K
Trypsin
ĐC
54
Hình 3.3 : Thử nghiệm với enzyme proteinase K và tripsin
Các chủng khác cho kết quả thử nghiệm là không có vòng kháng sau khi xử
lý proteinase K nhưng lại có vòng kháng sau khi xử lý enzyme tripsin. Do đó, chưa
thể xác định được dịch kháng khuẩn có phải là dịch bacteriocin thô hay không.
3.4 Quan sát hình thái tế bào vi khuẩn
Nuôi cấy chủng được tuyển chọn trên môi trường thạch TSA quan sát hình
thái khuẩn lạc của chủng tuyển chọn N14 để phục vụ cho việc định danh sau này.
Kết quả thể hiện như sau :
- Hình thái khuẩn lạc : Khuẩn lạc tròn, màu trắng sữa, trơn, hơi lồi lên trên bề
mặt thạch, bờ đều.
SVTH: Nguyễn Thị Thảo Trang

55
- Hình thái tế bào : Vi khuẩn Gram (+), trực khuẩn ngắn.
Hình 3.4 : Hình thái khuẩn lạc và tế bào vi khuẩn N14
A : khuẩn lạc trên đĩa cấy ria ; B : hình thái khuẩn lạc phóng to
C : hình thái tế bào khuẩn lạc ; D : cấy điểm
Kết luận chủng N14 là vi khuẩn gram dương là vi tế bào của nó bắt màu tím
của thuốc nhuộm tím violet. Điều này được giải thích như sau, do vi khuẩn Gram
dương có thành tế bào dày, dạng lưới, cấu tạo bởi peptidoglycan, có khả năng giữ
phức hợp tím kết tinh – liugol khi nhuộm. Khi tẩy cồn, có lỗ petidoglycan co lại,
khiến thành tế bào bắt giữ phức hợp này bên trong tế bào. Do đó tế bào không bắt
màu nhuộm fuchsin bổ sung, kết quả là tế bào vi khuẩn vẫn có màu tím.
SVTH: Nguyễn Thị Thảo Trang
A BCD

56
3.5 Đặc điểm sinh hóa của chủng vi khuẩn
3.5.1 Khả năng chịu muối
Chủng chọn lọc được nuôi trong môi trường TSB ở các nồng độ muối khác
nhau từ 1% - 12% ở 370C, để kiểm tra khả năng chịu muối của các chủng. Kết quả
sau 24 giờ nuôi được thể hiện ở bảng sau :
Bảng 3.4: Khả năng chịu muối của N14
Nồng độNaCl(%)Chủng 1 2 3 4 5 6 7 8 9 10 11 12
N14 + + + + + + + - - - - -
Từ bảng 3.4, cho thấy chủng N14 là chủng chịu mặn, có khả năng sinh
trưởng và phát triển tốt ở môi trường có nồng độ muối thấp. Do đó chủng có khả
năng sinh sống ở các vùng gần bờ, ở các vùng vịnh và cửa sông, nhu cầu về Na+ là
cần thiết đối với chủng này.
Sau 24 giờ, qua hình 3.5 ta thấy khuẩn lạc mọc dày đều trên các đường ria ở
các nồng độ từ 1-3% ở cả 2 chủng. Chủng phát triển mạnh nhất ở nồng độ 2%, có
thể kết luận rằng chủng N14 sống tốt ở vùng biển có độ mặn không cao. Ở nồng độ
muối 6%, khuẩn lạc bắt đầu thưa dần, đến nồng độ 7% vẫn có khả năng phát triển
nhưng rất yếu. Vậy nồng độ 7% chính là giới hạn trên của khả năng chịu muối . Từ
nồng độ 8% trở đi vi khuẩn không thể phát triển được nữa do nồng độ quá cao.
Qua đây có kết luận rằng, chủng N14 có khoảng giới hạn khả năng chịu
muối là khá cao từ 1% - 7%, môi trường nuôi cấy thích hợp nhất là bổ sung muối có
nồng độ 1 - 2%.
SVTH: Nguyễn Thị Thảo Trang

10%
1% 2% 4%3%
7%6%
11% 12%9%
8%5%
57
Hình 3.5 : Khả năng chịu muối của chủng N14
3.5.2 Thử catalase
Nuôi cấy dịch vi khuẩn sau 24 giờ, tiến hành nhỏ một giọt H2O2 30% vào
dịch vi khuẩn, kết quả cho thấy dương tính đối với khả năng sinh enzyme catalase.
Enzyme này là enzyme chỉ hiện diện ở vi khuẩn hiếu khí và kỵ khí tùy tiện có khả
năng biến dưỡng năng lượng theo phương thức hô hấp hiếu khí. Sự thủy phân
hydrogen peroxide sẽ giải phóng O2 gây ra hiện tượng sủi bọt.
2H2O2 2H2O + O2
SVTH: Nguyễn Thị Thảo Trang
Catalase

58
Hình 3.6 : Khả năng sinh catalase của N143.5.3 Tính di động
Để kiểm tra khả năng này, ta cấy thẳng chủng vi khuẩn vào môi trường thạch
TSA. Nuôi cấy sau 24 giờ, kết quả cho thấy cả chủng vi khuẩn chỉ mọc theo đường
cấy, chứng tỏ rằng chúng không có khả năng di động. Qua hình 3.7, ta còn thấy
rằng chủng N14có khả năng sinh hơi làm nứt thạch, và đẩy thạch lên trên.
Hình 3.7 : Khả năng di động của chủng N14
SVTH: Nguyễn Thị Thảo Trang
N14
N14ĐC

59
3.5.4 Khả năng sinh hơi
Khi vi khuẩn sử dụng nguồn carbon để lên men, tùy phương thức lên men sẽ
tạo ra được các sản phẩm khác nhau bao gồm rượu, các acid hữu cơ, CO2... Qua quá
trình thử nghiệm khả năng này, cho kết quả dương tính, song khả năng sinh hơi là
rất yếu, chỉ xuất hiện bóng khí trong ống Durham nhưng không thể đẩy ống nổi lên
trên môi trường. Vậy có thể kết luận rằng sản phẩm cuối cùng của quá trình lên men
ở chủng N14 có CO2.
Hình 3.8 : Khả năng sinh hơi của chủng N14
3.6 Định danh bằng phương pháp sinh học phân tử
Kết quả giải trình tự DNA của chủng N14 thu được đoạn gen có kích thước
là 835bp. Trình tự sau khi được phân tích, chỉnh sửa bằng phần mềm BioEdit
7.1.3.0 và Genious Pro 5.5.7, và được so sánh với các trình tự nucleotide tương
đồng trên Genbank bằng chương trình BLAST (www. ncbi . nlm . nih . gov / blast / ). Kết
quả cho thấy chủng N14 có độ tương đồng cao nhất với chủng Proteus sp. là 99%.
Như vậy, có thể kết luận sơ bộ chủng vi khuẩn N14 phân lập được, có khả năng
sinh bacteriocin thuộc chủng Proteus sp.
SVTH: Nguyễn Thị Thảo Trang
N14 ĐC

60
Chương IV : KẾT LUẬN VÀ ĐỀ XUẤT Ý KIẾN4.1 Kết luận
Từ những kết quả nghiên cứu ở trên chúng em đã đưa ra một số kết luận như
sau :
1) Đã phân lập được 46 chủng trong đó có 20 chủng có hoạt tính kháng
khuẩn.
2) Từ các chủng đã phân lập trên tuyển chọn được chủng N14 sinh
bacteriocin.
3) Xác định được sơ bộ các đặc điểm của chủng N14 :
- Có khuẩn lạc tròn màu trắng sữa, trơn,bờ đều
- Khả năng chịu muối ở nồng độ 1 - 7%, có khả năng sống ở những nơi có độ
mặn không quá cao.
- Sinh hơi, hay sản phẩm cuối cùng của quá trình lên men nguồn carbon có
chứa CO2
- Không có khả năng di động
- Sinh enzyme catalase.
4) Chủng được xác định là chủng Proteus sp.N14 có kích thước gen là
835bp.
4.2 Kiến nghị
1) Nghiên cứu sâu hơn về đặc điểm sinh hóa của chủng N14.
2) Xác định điều kiện nuối cấy tối ưu để chủng N14 phát triển và sinh
trưởng tối ưu nhất.
3) Kiểm tra lại các chủng còn lại trong số các chủng đem thử nghiệm
enzyme để đưa ra kết luận chính xác hơn, làm phong phú thêm bộ sưu tập
chủng vi khuẩn sinh bacteriocin.
4) Tiến hành phân loại bacteriocin.
5) Nghiên cứu khả năng ứng dụng của bacteriocin trong các lĩnh vực khác
nhau như y học, công nghiệp thực phẩm…
SVTH: Nguyễn Thị Thảo Trang

61
TÀI LIỆU THAM KHẢO Tài liệu tiếng Việt
1. Lê Thị Hồng Tuyết (2004). “Nghiên cứu bcateriocin sản xuất bởi Lactobacillus acidophilus NrrlB – 2092”. Luận văn thạc sĩ Sinh học, ĐHQG TpHCM.
2. Ngô Thị Phương Dung, Huỳnh Thị Yến Ly, Huỳnh Xuân Phong, Đại
học Cần Thơ. “Phân lập và tuyển chọn vi khuẩn lactic có khả năng sinh
chất kháng khuẩn”. Tạp chí Khoa học 2011 : số 19a 176-184.
3. Nguyễn Đức Hùng, Lê Đình Hùng, Huỳnh Lê Tâm (2004). “Sổ tay
kiểm nghiệm vi sinh thực phẩm thủy sản”. Nhà xuất bản nông nghiệp, Hà
Nội.
4. Nguyễn Lân Dũng, Nguyễn Đình Quyến, Phạm Văn Ty(2005). “Vi
sinh vật học”. Nhà xuất bản giáo dục.
5. Nguyễn Thị Đà, Hoa Thị Minh Tú, Lê Thanh Bình (2008). “Một số
tính chất của bcteriocin được tổng hợp bởi vi khuẩn lactic phân lập từ sữa
bò tươi”. Tạp chí Khoa học và Công nghệ, tập 46, số 6, 2008, Tr. 33-34.
6. Nguyễn Thị Bích Thủy (2004). “Một số dẫn liệu về ảnh hưởng của các
điều kiện môi trường lên sự sinh trưởng của tôm con (juvenile) tôm hùm
bông (Panulirus ornatus) ở vùng biển miền trung Việt Nam”. Tuyển tập
các công trình nghiên cứu khao học công nghệ (1984 – 2004), trung tâm
nghiên cứu thủy sản III – nha trang. Tr 73 - 81
7. Nguyễn Thị Bích Thủy(2006). “Kỹ thuật nuôi tôm hùm. GT kỹ thuật
nuôi giáp xác”. Nhà xuất bản Nông Nghiệp, TP Hồ Chí Minh.
8. Trần Linh Thước (2006). “Phương pháp phân tích vi sinh vật trong
nước, thực phẩm và mĩ phẩm”. Nhà xuất bản giáo dục, Hà Nội.
SVTH: Nguyễn Thị Thảo Trang

62
Tài liệu tiếng Anh
9. Axelsson L, Holck A (1995). “The genes involved in production of and
immunity to sakacin A, a bacteriocin from Lactobacillus sake Lb706”.
J.Bacteriol 177 (8): 2125-37.
10. Broadbent JR, Chou YC, Gillies K, Kondo JK (1989). “Ninsin inhitbits several gram – positive, mastitis–causing pathogens”. J.Dairy Sci, 1989, Dec;72(12) :3342-5.
11. Brötz H and Sahl H-G (2000). “New insights into the mechanism of
action of lantibiotics—diverse biological effects by binding to the same
molecular target”. Journal of Antimicrobial Chemotherapy (2000) 46, 1-6.
46 (1): 1–6. DOI:10.1093/jac/46.1.1. PMID 10882681
12. Desriac F, Defer D, Bourgougnon N, Brillet B, Le Chevalier P, Fleury
Y (2010). “Bacteriocin as Weapons in the Marine Animal-Associated
Bacteria Warfare: Inventory and Potential Applications as an Aquaculture
Probiotic”. Marine Drugs 2010 , 8 , 1153-1177, DOI
10.3390/md8041153.
13. Gert N.Moll, Wil N.Konings, Arnold J.M.Driessen (1999). “Bacteriocin: mechanism of membrane insertion and pore formation”. Antonie van Leeuwenhoek, 76, 185 – 198.
14. H.chen, D.G.Hoover (2003). “Bacteriocin and their food applications”. Comprehensive review in food sience and food safety. 2, 82 – 85.
15. Hühne K, Axelsson L, Holck A, Kröckel L (1996). “Analysis of the
sakacin P gene cluster from Lactobacillus sake Lb674 and its expression
in sakacin-negative Lb. sake strains". Microbiology (Reading, Engl.)
142(6): 1437–48. PMID 8704983 .
16. Jack RW, Tagg JR, Ray B (1995). “bacteriocin of gram-positive
bacteria”. Microbiol Rev 59:171-200
17. Joerger MC, Klaenhammer TR (1986). “Characterization and purification of helveticin J and evidence for a chromosomally determined bacteriocin produced by Lactobacillus helveticus 481”. J.Bacteriol, 1986 Aug167(2):439-46.
SVTH: Nguyễn Thị Thảo Trang

63
18. Jofré A, Garriga M, Aymerich T (2007). “Inhibition of Listeria
monocytogenes in cooked ham through active packaging with natural
antimicrobials and high-pressure processing”. J. Food Prot, 70 (11): 2498-
502, PMID 18044426.
19. Jose´ L Balcázar, Sara Loureiro, Yolanda J Da Silva, José Pintado
and Miquel Planas (2010). “Identification and characterization of
bacteria with antibacterial activities isolated from seahorses
(Hippocampus guttulatus)”. The Journal of Antibiotics (2010) 63, 271–
274, DOI:10.1038/ja.2010.27, published online 26 March 2010.
20. Juan C. Oscáriz, Antonio G. Pisabarro (2001). “Classification and
mode of action of membrane-active bacteriocin produced by gram positive
bacteria”. Int. Microbiol (2001) 4: 13-19, DOI 10.1007/s101230100003,
published online 25 August 2001.
21. Katla T, Møretrø T, Sveen I, Aasen IM, Axelsson L, Rørvik LM,
Naterstad K (2002). “Inhibition of Listeria monocytogenes in chicken
cold cuts by addition of sakacin P and sakacin P-producing Lactobacillus
sakei”. J.Appl. Microbiol 93 (2): 191-196, DOI : 10.1046/j.1365-
2672.2002.01675.x PMID 12147066
22. M.A. Riley, M.A. Chavan (Eds.). “Bacteriocins ecology and evolution”
23. Poeta P, Costa D, Sáenz Y, Klibi N, Ruiz-Larrea F, Rodrigues J, Torres C (2005). “Characterization of antibiotic resistance genes and virulence factors in faecal enterococci of wild animals in Portugal”. J.Vet Med B Infect Dis Vet Public Health, 2005, Nov;52(9):396-402.
24. Vesterlund S, Paltta J, Lauková A, Karp M, Ouwehand AC (2004). “Rapid screening method for the detection of antimicrobial substances”. J.Microbiol Methods, 2004 , Apr;57(1):23-31.
SVTH: Nguyễn Thị Thảo Trang

64
PHỤ LỤCPhụ lục 1 : Thành phần các hỗn hợp
a. Môi trường TSB
Trypticase peptone 17g
Phytone peptone 3g
NaCl 5g
K2HPO4 2,5g
Glucose 2,5g
Nước cất 1000ml
b. Thuốc nhuộm tím violet
Tím violet 1g
Rượu 1g
Phenol tinh thể 2g
Nước cất 100ml
c. Thuốc nhuộm liugol
Iod tinh thể 1g
KI 2g
Nước cất 200ml
d. Thuốc nhuộm Fuschin
Fuschin kiềm 1g
Rượu ethylic 95% 10ml
Phenol tinh thể 5g
Nước cất 100ml
SVTH: Nguyễn Thị Thảo Trang

65
Phụ lục 2 : Giải trình tự
Hình trình tự đoạn gen 16S rDNA của chủng Proteus sp. N14 (835
bp)
1 ACGGGAGGCA GCAGTGGGGA ATATTGCACA ATGGGCGCAA GCCTGATGCA 50
51 GCCATGCCGC GTGTATGAAG AAGGCCTTAG GGTTGTAAAG
TACTTTCAGC 100
101 GGGGAGGAAG GTGTTAAGAT TAATACTCTT ATCAATTGAC
GTTACCCGCA 150
151 GAAGAAGCAC CGGCTAACTC CGTGCCAGCA GCCGCGGTAA
TACGGAGGGT 200
201 GCAAGCGTTA ATCGGAATTA CTGGGCGTAA AGCGCACGCA
GGCGGTCAAT 250
251 TAAGTCAGAT GTGAAAGCCC CGAGCTTAAC TTGGGAATTG
CATCTGAAAC 300
301 TGGTTGGCTA GAGTCTTGTA GAGGGGGGTA GAATTCCACG
TGTAGCGGTG 350
351 AAATGCGTAG AGATGTGGAG GAATACCGGT GGCGAAGGCG
GCCCCCTGGA 400
401 CAAAGACTGA CGCTCAGGTG CGAAAGCGTG GGGAGCAAAC
SVTH: Nguyễn Thị Thảo Trang

66
AGGATTAGAT 450
451 ACCCTGGTAG TCCACGCTGT AAACGATGTC GATTTAGAGG
TTGTGGTCTT 500
501 GAACCGTGGC TTCTGGAGCT AACGCGTTAA ATCGACCGCC
TGGGGAGTAC 550
551 GGCCGCAAGG TTAAAACTCA AATGAATTGA CGGGGGCCCG
CACAAGCGGT 600
601 GGAGCATGTG GTTTAATTCG ATGCAACGCG AAGAACCTTA
CCTACTCTTG 650
651 ACATCCAGCG AATCCTTTAG AGATAGAGGA GTGCCTTCGG
GAACGCTGAG 700
701 ACAGGTGCTG CATGGCTGTC GTCAGCTCGT GTTGTGAAAT
GTTGGGTTAA 750
751 GTCCCGCAAC GAGCGCAACC CTTATCCTTT GTTGCCAGCG
CGTGATGGCG 800
801 GGAACTCAAA GGAGACTGCC GGTGATAAAC CGGAG 835
SVTH: Nguyễn Thị Thảo Trang

67
Bảng so sánh trình tự đoạn gen 16S rDNA của chủng Proteus sp.N14 với các
trình tự tương đồng trên Genbank bằng công cụ BLAST
Accession Description Max
score
Total
score
Query
coverage
E
value
Max
ident
Links
NR_043998.
1
Proteus penneri strain
NCTC 12737 16S
ribosomal RNA, partial
sequence
1498 1498 100% 0.0 99%
NR_043997.
1
Proteus mirabilis strain
NCTC 11938 16S
ribosomal RNA, partial
sequence
1487 1487 100% 0.0 99%
NR_025336.
1
Proteus vulgaris strain
DSM 30118 16S ribosomal
RNA, partial sequence
1476 1476 100% 0.0 99%
NR_043999.
1
Proteus myxofaciens strain
NCIMB 13273 16S
ribosomal RNA, partial
sequence
1423 1423 100% 0.0 97%
SVTH: Nguyễn Thị Thảo Trang





























