Embed Size (px)
Citation preview

Distribution and abundance of infective, soilborneFrankia and host symbionts Shepherdia, Alnus,and Myrica in a sand dune ecosystem
Janet McCray Batzli, Jeff F. Zimpfer, Valérie Huguet, Charles A. Smyth,Maria Fernandez, and Jeffrey O. Dawson
Abstract: We describe presence, abundance, and distribution of three sympatric nitrogen-fixing shrubs and their symbi-otic diazatroph, Frankia, in a sand dune ecosystem differing in successional stage, vegetative cover, edaphic character-istics, and topography. Distribution of actinorhizal Myrica gale L., Alnus incana (L.) Moench subsp. rugosa (Du Roi)Clausen, and Shepherdia canadensis (L.) Nutt. was analyzed among 120 sampling locations representing a gradient ofsuccessional stages in a sand dune system along Lake Michigan. In a greenhouse study, seedlings of these species wereemployed to bioassay the presence and abundance of infective Frankia in soils. Shepherdia-infective Frankia was de-tected in 80% of the plots, while Alnus- and Myrica-infective Frankia were found in 65% and 64% of the plots, re-spectively, with no Frankia found in 18% of the plots. Only 14% of the plots supported actinorhizal host-plant species.Infective Frankia were present in soils of young dunes prior to the establishment of any actinorhizal hosts. Shepherdia-infective Frankia were more abundant in soils from drier, earlier successional sites, while Alnus- and Myrica-infectiveFrankia were more abundant in moister soils of later successional communities. A previous study had revealed thatnodular Frankia strains at this site were host specific for Shepherdia and largely so for Myrica and Alnus, which hadonly a small proportion of shared strains (Huguet et al. 2001). The likelihood of host-plant nodulation by soilborneFrankia was increased by the presence of actinorhizal plants in general, but not by the presence of their respective spe-cific host plants. Submerged soils had no infectious capacity, whereas soils with greater in situ moisture content andsoils subject to intermittent saturation tended to have lower infectious capacities overall. Our results suggest thatsoilborne, infective Frankia genotypes are not only host specific, but are also associated with spatially and chronologi-cally distinct sets of ecological conditions.
709Key words: speckled alder, sweet gale, Canada buffalo berry, actinorhizal, nitrogen fixation, Frankia, root nodules.
Résumé : Les auteurs décrivent la présence, l’abondance et la distribution de trois arbustes sympatriques fixateursd’azote, ainsi que du Frankia, leur symbionte diazotrophe, dans un écosystème de dunes de sable, selon le stade desuccession, la couverture végétative, les caractéristiques édaphiques et la position topographique. Ainsi, ils ont étudié ladistribution de trois espèces actinorhiziennes, les Myrica gale L., Alnus incana (L.) Moench subsp. rugosa (Du Roi)Clausen et Shepherdia (L.) Nutt., en analysant 120 stations échantillons représentant un gradient de stades de succes-sion, dans un système de dunes de sable, le long du lac Michigan. En serre, ils ont utilisé les plantules de ces trois es-pèces pour évaluer, par piégeage, la présence et l’abondance des propagules colonisatrices du Frankia dans les sols. Onretrouve des propagules du Frankia colonisatrices du Shepherdia, de l’Alnus et du Myrica dans 80 %, 65 % et 64 %des parcelles, respectivement; le Frankia s’avère absent dans 18 % des parcelles. Seulement 14 % des parcelles suppor-tent des espèces de plantes actinorhiziennes. Les propagules colonisatrices du Frankia sont présentes dans les sols desjeunes dunes, avant même l’arrivée de toute plante hôte actinorhizienne. Les Frankia colonisateurs du Shepherdia sontplus abondants dans les sols plus secs des sites de début de succession, alors que les propagules colonisatrices del’Alnus et du Myrica sont plus abondantes dans les sols plus humides des communautés de fin de succession. Uneétude antécédente a montré que ce site comporte des souches de Frankia nodulateur spécifiques au Shepherdia, ce quiest également vrai en bonne partie pour le Myrica et l’Alnus, qui n’ont que de rares souches en commun. La probabi-lité de nodulation des plantes hôtes par les Frankia du sol est augmentée par la présence de plantes actinorhiziennes en
Can. J. Bot. 82: 700–709 (2004) doi: 10.1139/B04-044 © 2004 NRC Canada
700
Received 21 March 2003. Published on the NRC Research Press Web site at http://canjbot.nrc.ca on 7 June 2004.
J.M. Batzli,1 J.F. Zimpfer, and J.O. Dawson.2 Department of Natural Resources & Environmental Sciences, University of Illinois,Urbana, IL 61801, USA.C.A. Smyth. Department of Crop Sciences, University of Illinois, Urbana, IL 61801, USA.V. Huguet3 and M. Fernandez. Laboratoire d’écologie microbienne, Unité mixte de recherche (UMR), Centre national de larecherche scientifique (CNRS), 5557 Université Claude Bernard Lyon 1, 69622 Villeurbanne CEDEX, France.
1Present address: Biocore Program, University of Wisconsin, Madison, WI 53706, USA.2Corresponding author (e-mail: [email protected]).3Present address: Microbiologie et géochimie des sols, UMR INRA 1229, 17 Rue Sully, B.P. 86510, 21065 Dijon CEDEX, France.

général, mais non par la présence de leur plante hôte spécifique respective. Les sols submergés n’ont pas de capacitécolonisatrice, alors que les sols avec des teneurs en humidité in situ plus élevées et les sols sujets à la saturation parintermittence ont, en général, tendance à montrer de plus faibles capacités colonisatrices. Les résultats suggèrent queles génotypes des Frankia colonisateurs des sols ne sont pas seulement spécifiques à l’hôte, mais aussi associés avecdes ensembles de conditions écologiques spatialement et chronologiquement distinctes.
Mots clés : aune rugueux, myrique baumier, shepherdie du Canada, actinorhize, fixation de l’azote, Frankia, nodulesracinaires.
[Traduit par la Rédaction] Batzli et al.
Introduction
Actinorhizal plants form root nodules with nitrogen-fixingactinomycetal symbionts in the genus Frankia. Actinorhizalplants are prominent as early-successional plants in a varietyof terrestrial ecosystems (Lawrence et al. 1967; Chapin et al.1994; Dalling 1994; Vitousik and Walker 1989). Nitrogen-fixing legumes and actinorhizal plants tend to colonize soilslacking available nitrogen and can, in some instances, con-tribute significantly to soil nitrogen pools. Nitrogen-fixinglegumes are known to be rare in the sand dune ecosystemsalong the shores of the Great Lakes of North America, asopposed to nonleguminous fixers such as Myrica and Alnus,which can become locally dominant (Walker 1993), as in ourstudy location in Wisconsin on the western shore of LakeMichigan (Trick 1983). Lichter (1998) noted the rare in-stance of nitrogen-fixing plants in a dune–ridge ecosystemon the eastern shore of Lake Michigan, suggesting that pre-cipitation is likely the primary source of soil nitrogen indune successional communities. Other research on a ridge–swale chronosequence along Grand Traverse Bay in northernMichigan found Myrica gale L. (sweet gale) to dominate onyounger ridges and the presence of large amounts of Alnusincana (L.) Moench subsp. rugosa (Du Roi) Clausen (speck-led alder) and M. gale pollen associated with depositionalstrata occurring after fire and logging in the mid- to late 19thcentury (Booth and Jackson 2000). The role of actinorhizalplants in the nitrogen economy of these dune–ridge ecosys-tems has not been established. Whatever their nitrogen con-tribution to these ecosystems might be, their symbioticrelationship probably plays an important role in enablinghost-plant establishment in nitrogen-poor sand dune ecosys-tems.
During primary succession, or wherever soil nitrogen islimited in its availability to plants, establishment and growthof actinorhizal plants likely depends on their ability to formfunctional root nodules. Little is known about the spatial dis-tribution and abundance of infective Frankia in relation totheir host plants and environments in sand dune or otherearly-successional soils. Many factors are thought to affectthe abundance of soilborne Frankia able to infect their hostplants. These factors include soil pH (Smolander 1990),moisture (Dawson et al. 1989), aeration (Dawson et al.1989), soil nutrients (Righetti et al. 1986; Smolander 1990),competition from other soil organisms (Oremus 1980), claycontent (Smolander et al. 1988), salinity (Young et al. 1992),and the nature of the dominant vegetation (Smolander 1990;Elo et al. 2000; Gauthier et al. 2000). Therefore, symbioticinteractions may vary in successional plant communities
with variable physical, biological, and chemical soilconditions. Understanding factors affecting the nodulation ofactinorhizal host plants will help explain their potential forestablishment, nitrogen fixation, and growth.
Several studies have indicated that the presence of Fran-kia in young soils is not dependent on host-plant presence.In soils of barrier islands off the coast of Virginia, coloniza-tion of Frankia able to infect actinorhizal Myrica ceriferaoccurs prior to colonization by host plants (Young et al.1992). On the islands of Java (Becking 1970) and Hawaii(Burleigh and Dawson 1994), Myrica nodulates on volcanicsoils. On the island of Jamaica, 1-year-old reclaimed bauxitemines harbored low levels of M. cerifera-infective Frankia(Zimpfer et al. 1997). In Alaska’s Glacier Bay, Dryas drum-mondii and Alnus sinuata nodulate readily on newly formedglacial soils (Lawrence et al. 1967), indicating the earlypresence in glacial deposits of infectious Frankia.
Most actinorhizal plants are found to be nodulated in theirnative habitats, and Frankia is usually found in abundance insoils beneath nodulated host plants (Arveby and Huss-Danell1988; Oremus 1980; Van Dijk 1979; Smolander 1990; Zimp-fer et al. 1999). However, some actinorhizal genera are oftensparsely nodulated or not nodulated at all, even within theirnative habitats (Dawson et al. 1989) and, therefore, probablycontribute little to host plant success or the overall accumu-lation of soil nitrogen. In addition, there are numerous re-ports of Frankia in soils lacking actinorhizal host plants(Rodriguez-Barrueco 1968; Bermudez de Castro et al. 1976;Huss-Danell and Frej 1986; Dawson and Klemp 1987;Paschke and Dawson 1992b; Smolander and Sundman 1987;Smolander 1990; Young et al. 1992; Zimpfer et al. 1997),providing indirect evidence that Frankia can grow and sur-vive in soils in the absence of symbiotic hosts.
Three species of actinorhizal plants, A. incana subsp.rugosa, Shepherdia canadensis (L.) Nutt. (Canada buffaloberry), and M. gale occur in Lake Michigan sand dune –swale primary successional communities in the Door Penin-sula in east-central Wisconsin. The objective of the presentstudy was to describe the presence, abundance, and distribu-tion of soilborne Frankia as associated with host-plant pres-ence, successional stage, edaphic factors, and topographicfeatures. Frankia nodulation capacities, actinorhizal plantcover, and soil characteristics are described for sample plotsstratified over sand dune communities on the Door CountyPeninsula along Lake Michigan. Nodulation capacities weredetermined from soil samples that were serially diluted andused to inoculate seedlings of the three actinorhizal host spe-cies in a greenhouse bioassay.
© 2004 NRC Canada
Batzli et al. 701

Materials and methods
Site descriptions
The Ridges siteThis study site is located within the Ridges Sanctuary in
Bailey’s Harbor, Wisconsin, USA (45°05′N, 87°06′W),which has long been recognized for its unique, cool lake-shore microclimate and flora (Curtis 1959) (Fig. 1). Oneside of the site is bounded by a 2.5 km wide embayment thatopens southward to Lake Michigan, and both east and westsides are bounded by Silurian dolomite uplands (Sherrill1978). Between the uplands there are 29 sand dune ridgesthat formed between 1050 and 250 years BP (Thompson andBaedke 1997) and run parallel to the Lake Michigan shore-line. These ridges are low in relief (0.5–2.5 m in height), andthe distance from ridge top to ridge top varies from 26 to51 m (Fig. 1). The soils of the ridges are sandy with a dis-tinct, surficial organic horizon that deepens with increaseddistance from the lake. The soils of the swales consist of anorganic muck (Link et al. 1977). The dune ridges remain asremnants of ancient shorelines that developed through wave
action and annual fluctuations in lake water level over thelast 1260 years (Thomson and Baedke 1997). Sand was de-posited during times of rapid rise in lake levels followed byprolonged low levels that created ridges with intervening de-pressions called swales. Water in the swales is stagnant,fluctuating with seasonal high water levels in the spring andlow levels, with some swales drying to soil level, in the fall.The swales support open pond vegetation, peat lands, andconifer swamp. Swales alternate with ridge upland commu-nities that are dominated by Thuja occidentalis L. (white ce-dar), Abies balsamea (L.) Mill. (balsam fir), Picea glauca(Moench) Voss (white spruce), Pinus strobus L. (whitepine), Pinus resinosa Ait. (red pine), and Larix laricina (DuRoi) K. Koch (tamarack).
Plots, vegetation analysis, and sample collectionAt the Ridges site, soil and vegetative samples were taken
from twelve 4 m × 4 m plots randomly selected within unde-veloped portions of each of 10 communities paralleling thedunes and representing the extremes of the range of commu-nity types and topographic positions. The biotic communi-
© 2004 NRC Canada
702 Can. J. Bot. Vol. 82, 2004
Fig. 1. Aerial photograph of Ridges Sanctuary, Bailey’s Harbor, Wisconsin. Sampling plots in the Ridges were randomly selectedwithin 10 communities parallel to ridges 1, 2, and 8, with numbers shown above ridge location. MLB refers to Moonlight Bay studysite in companion study (Huguet et al. 2004). (Photograph courtesy of US Geological Survey; scale 1 : 24 000.)

ties were stratified over the successional gradient at thefollowing locations: (1) the beach along the lakeshore whereno vegetation occurred; (2) a small dune supporting the firstoccurrence of vegetation inland from the beach; (3–5) againmoving inland, the successive foreslope, top, and afterslopeof ridge 1; (6) the swale between ridges 1 and 2; (7) the topof ridge 2; (8) the swale between ridges 2 and 3; (9) the topof ridge 8; and (10) the swale between ridges 8 and 9(Fig. 1). A total of 120 plots were sampled.
Surface soil samples were collected aseptically using a2 cm diameter steel probe to a depth of 15 cm in each 4 m ×4 m plot. In plots with standing water, a modified samplingmethod was used in which soil was collected using a 6 cmdiameter probe to a depth of 15 cm. Soil samples were col-lected in June and July from four random locations withineach plot and bulked. Additional samples were collected forphysical and chemical soil analyses. The samples weresealed in sterile plastic bags and transported on ice to thegreenhouse in Urbana, Illinois, where they were uniformlyair dried (at 25 °C) in a room with a filtered air supply freeof Frankia for 5 d. Sample bags were then sealed, and soilswere stored at 20 °C until they were used in the greenhousebioassay (~2 weeks). Because air-dried samples have beenshown to remain infectious for Frankia for months, the useof dried preparations is standard practice for assaying rela-tive infectious capacities of soil (Tortosa and Cusato 1991).Nevertheless, the change in infectious capacity was deter-mined by comparing a subsample of plants inoculated withfresh soil relative to those that had been air dried for2 weeks. In the present study we found that nodulation ca-pacity was diminished equally in all three species by roughly50% compared with fresh soil inoculum when soil wasdried, therefore estimates of nodulation capacity should beviewed as conservative. The lack of a significant species ×drying treatment interaction suggests that the infectious ca-pacity of the soil for each species was not differentially af-fected by drying.
Vegetation frequency and density was estimated forS. canadensis, M. gale, and A. rugosa using the line inter-cept method (Bonham 1989). The vegetation estimates wereobtained by stretching a 4-m measuring tape taut at groundlevel and measuring the number of intercepts and the lengthof tape covered by the crowns of each actinorhizal species.Vegetation intercept was measured midway between andalong the north and south edges of the 4 m × 4 m plot. Thepercentages of the line transects intersected by the canopiesof each species were used to estimate the percent cover ofeach actinorhizal species per plot.
Soil bioassay for Frankia presence and abundanceMethods used to determine Frankia abundance were those
of Paschke and Dawson (1993) in which nodule quantity onthe roots of “bait plants”, in this case greenhouse-grownseedlings of S. canadensis, A. incana subsp. rugosa, andM. gale, inoculated with serial dilutions of soil samples wereused to estimate the number of infective units (IU) of Fran-kia in the soil. Seeds of S. canadensis, M. gale, andA. rugosa were obtained from commercial distributors (Shef-field’s Seed Co., Inc. Locke, New York), washed in deio-nized water for 24 h, surface sterilized in 30% (v/v)hydrogen peroxide, rinsed in sterile deionized water, and
sown directly in an artificial soil. Bait plants were grown in4 cm × 14 cm cone-shaped containers (tubes) with bottomdrainage filled with a 1:1:1 steam-pasteurized mixture offine vermiculite – mixed sand – fine gravel. Containers weredisinfected prior to use in a 0.4% (v/v) aqueous solution ofsurface sterilant (Greenshield® Whitmire Research Labora-tories, St. Louis, Missouri).
The plants were watered weekly with one-eighth strength(0.179 µmol/L NH4NO3) nutrient solution (Huss-Danell1978). Two weeks prior to inoculation, the nitrogen supplywas eliminated from the applied nutrient solution to facili-tate nodulation, which is reduced or eliminated by high lev-els of nitrogen fertility. After 12 weeks, when the roots ofthe seedlings fully occupied the growing medium withineach tube, the plants were inoculated with serial dilutions ofsoil. At this stage the plants were chlorotic given the lack ofnodulation and substrate nitrogen. Initial mean plant heights(standard deviation) 12 weeks after germination were 3.4 cm(0.6) for Shepherdia, 3.5 cm (0.5) for Myrica, and 5.7 (1.1)for Alnus. The initial limited availability of nitrogen to theexperimental plants favored root growth over shoot growth,a well-known and common response of plants to low levelsof available nitrogen. This explains the full occupation byroots of the artificial soil in the small conical tubes, eventhough shoot growth was modest. However, increased shootgrowth of nodulating seedlings and greening of chlorotic fo-liage became evident several weeks after inoculation, as thenitrogen status of the seedlings apparently improved withnodulation by effective, soilborne Frankia.
The bulk densities of soil samples from the field were de-termined prior to their use in the bioassay. The soil serial di-lutions were prepared by adding 50 g of soil to 175 mL ofdeionized H2O and mechanically stirring them for 30 min,after which time the contents were filtered through cheese-cloth. From these solutions, four 10-fold dilutions were pre-pared and applied to the bait plants. Ten millilitres of theappropriate inoculum was applied to each plant, with threeplants inoculated with solutions at each dilution level. To as-say the presence of Frankia in lake and swale water, 10-mLsamples of water, filtered and unfiltered for removal of de-bris, were used as inoculum with the same dilution levels. Inaddition to inoculated plants, noninoculated controls of eachplant species were distributed within the inoculated treat-ments to assay for contamination.
Plants were grown under greenhouse conditions at a tem-perature of 22 ± 5 °C and a photoperiod of 16 h, extendedby 1000 W Sylvania LU1000 lamps when natural lightintensity at plant level decreased below 400 µmol·m–2·s–1
photosynthetic photon flux density. To eliminate Frankiacontamination and minimize other microbial contaminants,floors and greenhouse benches were disinfected with a 0.4%(v/v) aqueous solution of the surface sterilant Greenshield®
once every 10 d.Twelve weeks after inoculation, the plants were harvested
and the root nodules counted. Shoot mean heights (standarddeviation) of nodulated seedlings at time of harvest were5.4 cm (0.8) for Shepherdia, 5.5 cm (0.9) for Myrica, and10.1 cm (1.8) for Alnus. Controls had not grown after theinitial 12 weeks and had chlorotic foliage with some necro-sis. Soil bulk density values together with dry mass of inocu-lated soil were used to determine the equivalent volume of
© 2004 NRC Canada
Batzli et al. 703

soil applied to each plant. Standard regression with the inter-cept forced through zero was used to determine the numberof nodules formed per cubic centimetre of soil inoculum.One nodule represented an estimate of one infective unit ofFrankia.
Soil analysesSix dried soil samples (2 cm × 10 cm deep) from each
plot were analyzed using the following techniques: pH (Mc-Lean 1980), phosphorus (Knudsen 1980); cation exchangecapacity (Rhoades 1982); potassium (Carson 1980); magne-sium (Whitney 1980); and calcium (Lanyon and Heald1982). Three dried soil samples from each plot were usedfor total nitrogen determination by weighing 50 mg of soiland analyzing it in a N2000 analyzer (Thermo InstrumentSystems, Inc., Waltham, Massachussetts) according to themethods of Kirsten (1983). Bulk density was expressed asgrams of dry mass per cubic centimetre, and percent mois-ture was determined by dividing the soil moisture content bysoil dry mass. Percent soil organic matter was determined bycombustion of soil at 700 °C in a Thermolyne type 10500muffle furnace for 24 h (Barnstead/Thermolyne Corp.,Dubuque, Iowa). Soil moisture was determined by differen-tial mass of fresh soil samples and those that were dried at121 °C in a drying oven for 48 h.
Statistical analysisUnivariate analysis of variance with mean separation pro-
cedures was used to determine differences in Frankia nodu-lation capacity at the Ridges location, the percentage ofvegetative cover of actinorhizal shrubs, and soil propertiesassociated with each community along the successional gra-dient. Estimated values for infectious capacity were derivedfrom regression analysis of serial dilution × nodulation re-sults and were included in statistical analyses only if the pvalue was < 0.05. Because the capacity to nodulate could bedependent on intrinsic characteristics of each species, abso-lute infectious capacity was used only to make comparisonswithin a species across the successional gradient. Correla-tion, χ2 analysis, and multiple regression were employed toexplore the relationships of Frankia nodulation capacity withhost-plant abundance and soil characteristics.
Results
The greenhouse bioassay for estimating nodulation capac-ities of soil samples was controlled effectively. Only inocu-lated plants that also became nodulated had dark greenfoliage when grown without substrate nitrogen, indicatingthat actinorhizal nodules formed on bioassay trap plantswere effective at fixing nitrogen symbiotically.
Infectious Frankia occurred in soils from 82% of the 120plots, while only 14% of the plots contained actinorhizalhost species. Only one of the plots had two host-plant spe-cies present, and no plots supported growth of all three acti-norhizal plant species. Infective Frankia was detected in soilfrom all plots where actinorhizal plants were found growing.Shepherdia-infective Frankia were found in 80% of sampledplots (96 of 120 plants sampled), while Alnus- and Myrica-infective Frankia were found in 65% and 64% of the plots,respectively. A χ2 analysis of Frankia presence or absence
data indicated that the presence of specific Frankia in a plotwas not dependent on the presence of a specific host plant.However, there was a positive relationship between Frankiapresence and the presence of actinorhizal plant species ingeneral (χ2 (1, N = 101) = 3.85, p < 0.05). The distributionof soils capable of nodulating Myrica was similar to that ofsoils capable of infecting Alnus, while the distribution ofShepherdia-infective Frankia was independent of the soildistribution of the other two types of Frankia. No Frankiawas detected in lake water or filtered swale water, and lessthan 1 IU Frankia/cm3 was found in unfiltered swale water.No Frankia was detected in soil of the swales betweenridges if it was submerged to a depth of greater than 21 cmat the time we sampled.
Infective populations of Frankia occur prior to the estab-lishment of plant host species (Fig. 2). Only Shepherdia-in-fective Frankia was detected in soils of the earliestsuccessional communities at the Ridges. It was found in soilsamples of a dune forming between the beach and ridge 1.Shepherdia-infective Frankia was found in this young dunecommunity at a level of 7 Frankia IU/cm3 soil, whereas nei-ther Alnus- nor Myrica-infective Frankia was detected in thesame soils.
The foreslope plots of ridge 1 were located an average of8 m inland from the first new dune. Shepherdia-infectiveFrankia differed significantly among seral communities andwas found to have its greatest abundance (20 FrankiaIU/cm3 soil) on the fore slope of ridge 1 facing the lake(Fig. 2). Shepherdia nodulation capacity declined in abun-dance in later successional communities, with lesser infec-tious capacities in soils from the top of ridge 2 and theswales on the inland sides of ridges 2 and 8.
The earliest successional site in which Alnus-infectiveFrankia occurred was the top of ridge 1 (Fig. 2). The Alnusnodulation capacity of soils differed significantly accordingto seral community, but was similar for the ridge tops ofboth early- and late-successional communities, with lesserinfectious capacities for Alnus in the swales. The earliestsuccessional site in which Myrica-infective Frankia was de-tected in soils was on the foreslope of ridge 1, and thenodulation capacity of soils increased significantly overall,with greatest capacities found inland with increasing age andmore advanced successional stages of the communities.
There was a significant species × seral community inter-action (F = 2.55; p = 0.0002) in overall infectious capacitytrends across the successional gradient, suggesting differ-ences in response to changes across ecological gradients ofthe host-specific Frankia populations. Shepherdia-infectiveFrankia had a different distribution compared with eitherAlnus- or Myrica-infective Frankia, while the distributionsof Myrica- and Alnus-infective Frankia could not be differ-entiated (Fig. 2). Trends along the successional gradienthighlight an abundance of Shepherdia-infective Frankiaearlier in succession, with much less infectious capacity atridge 8. In contrast, Alnus- and Myrica-infective Frankiawere more abundant in soils of later successional communi-ties and stayed at a moderate to low level throughout thegradient, with declines detected only in samples from thewet swale locations.
The vegetative cover of actinorhizal plant species wasspotty and localized at the Ridges location. Although per-
© 2004 NRC Canada
704 Can. J. Bot. Vol. 82, 2004
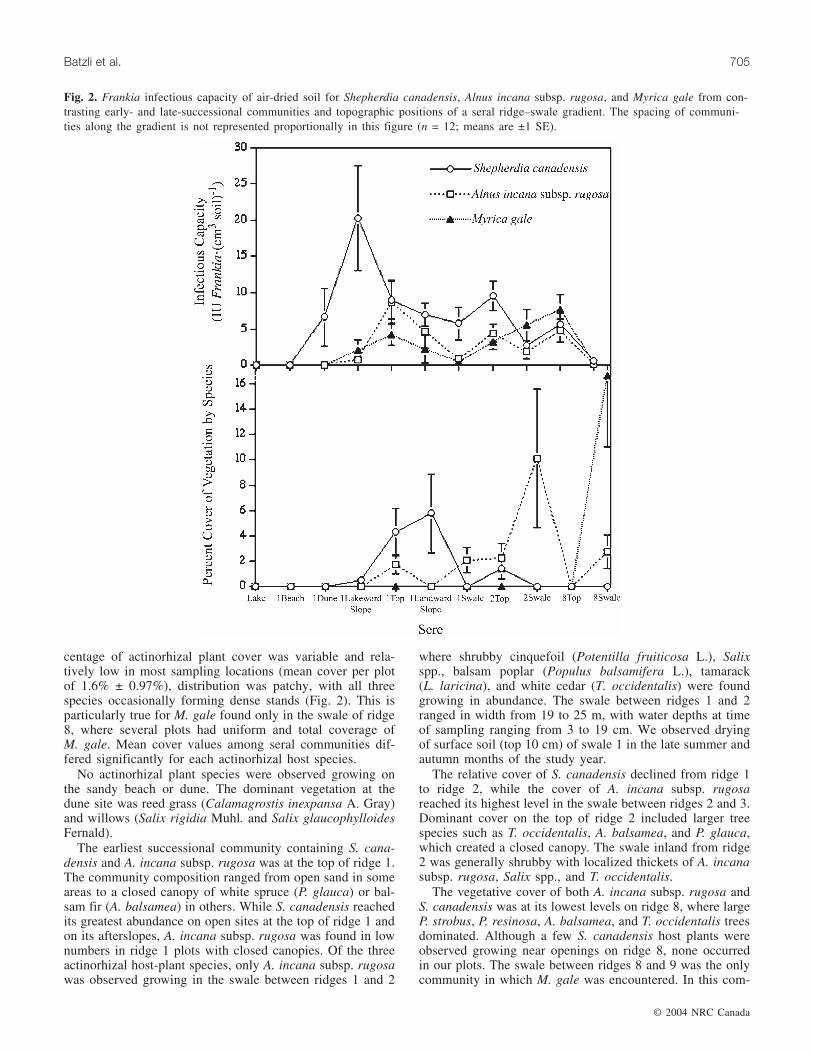
centage of actinorhizal plant cover was variable and rela-tively low in most sampling locations (mean cover per plotof 1.6% ± 0.97%), distribution was patchy, with all threespecies occasionally forming dense stands (Fig. 2). This isparticularly true for M. gale found only in the swale of ridge8, where several plots had uniform and total coverage ofM. gale. Mean cover values among seral communities dif-fered significantly for each actinorhizal host species.
No actinorhizal plant species were observed growing onthe sandy beach or dune. The dominant vegetation at thedune site was reed grass (Calamagrostis inexpansa A. Gray)and willows (Salix rigidia Muhl. and Salix glaucophylloidesFernald).
The earliest successional community containing S. cana-densis and A. incana subsp. rugosa was at the top of ridge 1.The community composition ranged from open sand in someareas to a closed canopy of white spruce (P. glauca) or bal-sam fir (A. balsamea) in others. While S. canadensis reachedits greatest abundance on open sites at the top of ridge 1 andon its afterslopes, A. incana subsp. rugosa was found in lownumbers in ridge 1 plots with closed canopies. Of the threeactinorhizal host-plant species, only A. incana subsp. rugosawas observed growing in the swale between ridges 1 and 2
where shrubby cinquefoil (Potentilla fruiticosa L.), Salixspp., balsam poplar (Populus balsamifera L.), tamarack(L. laricina), and white cedar (T. occidentalis) were foundgrowing in abundance. The swale between ridges 1 and 2ranged in width from 19 to 25 m, with water depths at timeof sampling ranging from 3 to 19 cm. We observed dryingof surface soil (top 10 cm) of swale 1 in the late summer andautumn months of the study year.
The relative cover of S. canadensis declined from ridge 1to ridge 2, while the cover of A. incana subsp. rugosareached its highest level in the swale between ridges 2 and 3.Dominant cover on the top of ridge 2 included larger treespecies such as T. occidentalis, A. balsamea, and P. glauca,which created a closed canopy. The swale inland from ridge2 was generally shrubby with localized thickets of A. incanasubsp. rugosa, Salix spp., and T. occidentalis.
The vegetative cover of both A. incana subsp. rugosa andS. canadensis was at its lowest levels on ridge 8, where largeP. strobus, P. resinosa, A. balsamea, and T. occidentalis treesdominated. Although a few S. canadensis host plants wereobserved growing near openings on ridge 8, none occurredin our plots. The swale between ridges 8 and 9 was the onlycommunity in which M. gale was encountered. In this com-
© 2004 NRC Canada
Batzli et al. 705
Fig. 2. Frankia infectious capacity of air-dried soil for Shepherdia canadensis, Alnus incana subsp. rugosa, and Myrica gale from con-trasting early- and late-successional communities and topographic positions of a seral ridge–swale gradient. The spacing of communi-ties along the gradient is not represented proportionally in this figure (n = 12; means are ±1 SE).

munity, M. gale was locally dominant along with sedges(Carex spp.). At our sampling locations, this swale varied inwidth from 19 to 50 m, with areas of open water that rangedin depth from 0 to 53 cm.
Soil nitrogen levels increased with increasing distancefrom the lakeshore, while phosphorus and potassium weremore variable (Table 1). The higher concentrations of nitro-gen in soils from swales between ridges 8 and 9 were associ-ated with higher levels of organic matter at this samplinglocation. Soil pH was lower in older soils with higher levelsof organic matter. Soil moisture on dunes, slopes, and sandyridge tops was similar and consistently drier than the soil inthe swales.
There was no significant correlation between most soilproperties measured and soil nodulation capacities. Therewas, however, a relationship between soil nodulation capac-ity and soil moisture. In situ measurements of soil moistureat the time of sampling were proportional to seasonally ob-served amounts of soil moisture at different topographic po-sitions, which indicated that soil infectious capacity declinedwith greater mean soil moisture percentages in these soils.This was particularly true for Shepherdia-infective Frankia,where the greatest infectious capacity was found in plotswith a moisture content of less than 10%. Maximum-valueborder plots of soil moisture (Fig. 3) illustrate that soils withgreater than 20% moisture content had lower maximumShepherdia-infectious capacities. Maximum infectious ca-pacities for Myrica- and Alnus-infective soils remained sta-ble compared with Shepherdia infectious capacities, whichdeclined with increasing soil moisture. Relatively low infec-tious capacities were observed for all species in soils with20%–50% soil moisture at the time of sampling. Soil sub-merged beneath more than 21 cm of water had no detectableFrankia infectious capacity for any of the species studied.
Discussion
Our research revealed differential distribution and abun-dance of host-specific, soilborne, infective Frankia, indica-tive of different Frankia nodular-strain ecotypes. Thepresence of Frankia in a plot was not dependent on the pres-ence of specific host-plant species. However, there was apositive relationship between Frankia presence and the pres-
ence of actinorhizal plant species in general. It is thereforepossible that host signaling and stimulation of infectivitymay operate independently of host recognition.
Nonactinorhizal genera in the same family as actinorhizalgenera are known to stimulate Frankia infectious capacity ofsoils for their hosts (Smolander 1990; Paschke and Dawson1992a; Gauthier et al. 2000). Thus stimulation of Frankia in-fectious capacity may be a generalized and indiscriminantprocess in actinorhizal plants and their relatives. This furthersuggests the possible independence of genetic and physio-logical mechanisms that regulate Frankia nodulation capac-ity and those that confer host specificity.
In a previous study at this site by Huguet et al. (2001),PCR-RFLP (polymerase chain reaction – restriction frag-ment length polymorphism) of the 16S–23S intergenicspacer (IGS) found that A. incana subsp. rugosa- andM. gale-specific Frankia collected from host nodules inproximity to our sampling location were genetically distinctbut closer genetically to one another than to Shepherdia-in-fectious Frankia. The nodular Frankia strains specific to thethree actinorhizal hosts described in the present studyexhibited ecological differences according to differences inrelatedness indicated by the genetic analysis. There was sim-ilarity in distribution and nodulation capacity betweenAlnus- and Myrica-infective Frankia and distinct separationof Shepherdia-infective Frankia from the other genotypesaccording to the bioassay.
Shepherdia-infective Frankia were more abundant in soilsamples from drier and earlier successional sites, whereasAlnus- and Myrica-infective Frankia were more abundantin soils from moister, later successional sites. HenceShepherdia-infective Frankia apparently differed ecologi-cally from the Frankia strains that infect A. incana subsp.rugosa and M. gale. The nodular Frankia from the presentstudy were delineated using PCR-RFLP of the 16S–23S IGS(Huguet et al. 2004). Within the diverse delineated group ofShepherdia-infective nodular strains, one distinct strain wasdominant in drier, bare soils of the earliest stages of sanddune succession, soils that also promoted the greatestamount of Shepherdia nodulation. Another nodulating Fran-kia strain dominated in soils from later successional stages,particularly from nodules formed in soils sampled nearShepherdia host plants. Results suggest that soilborne infec-
© 2004 NRC Canada
706 Can. J. Bot. Vol. 82, 2004
Soil (ridge No.,topoposition)
Soil moisture(%)a
Organicmatter (%)
Total N(g/kg) P (mg/kg) K (mg/kg) pH
CECb
(mmol·L–1·100 g–1)
Beach 20.97 (0.8) 1.05 (0.7) 0.023 (0.0) 12.25 (3.7) 13.17 (2.1) 7.47 (0.1) 6.26 (0.47)Dune 15.78 (0.4) 1.01 (0.4) 0.032 (0.0) 6.41 (2.2) 10 (1.5) 7.42 (0.0) 7.08 (0.53)1, foreslope 11.66 (0.4) 1.13 (0.4) 0.093 (0.0) 6.37(1.8) 13.67(1.8) 7.25 (0.1) 6.66 (0.37)1, top 12.69 (3.0) 1.01 (0.4) 0.101 (0.0) 5.75 (0.9) 15.91 (1.7) 7.47 (0.0) 6.51 (0.48)1, afterslope 16.11 1.35 (0.3) 0.040 (0.0) 7.08 (2.3) 13.25 (1.6) 7.35 (0.0) 7.42 (0.41)1–2, swale 34.90 (2.9) 3.64 (0.4) 0.062 (0.0) 6.45 (0.8) 11.27 (0.7) 7.49 (0.0) 9.25 (0.21)2, top 9.25 (0.8) 5.26 (0.3) 0.074 (0.0) 5.58 (1.2) 14.58 (1.4) 7.08 (0.1) 10.08 (0.8)2–3, swale 32.72 (0.7) 11.87 (2.4) 4.969 (1.0) 5.58 (1.2) 14.58 (1.4) 7.28 (0.1) 12.15 (1.2)8, top 11.87 (1.7) 13.66 (4.2) 0.065 (0.0) 6.12 (1.0) 38.08 (4.8) 6.13 (0.2) 13.45 (2.3)8–9, swale 79.80 (8.9) 57.39 (7.1) 8.750 (2.1) 10.30 (2.1) 39.30 (5.9) 6.48 (0.0) 29.78 (1.8)
Note: Values are means (±SE); means are significantly different at p < 0.001 within columns. N, nitrogen; P, phosphorus; K, potassium.an = 6–12 for all variables, except total n and soils from Moonlight Bay, where n = 3.bCation-exchange capacity.
Table 1. Soil chemical properties of samples collected from the Ridges Sanctuary on ridges 1, 2, and 8.

tive Frankia are not only host specific, but are also specificin their nodulation response along ecological gradients.
Frankia infectivity varied significantly with a set of soilmoisture levels used as an index of relative seasonal mois-ture content (Fig. 3), but not with other ecological variables.Moisture levels vary interactively with such factors as sea-son, topography, position of water table, soil texture, andsoil organic matter. Saturated and submerged soils had theleast infectious capacity. Shepherdia-infective Frankiaseemed to predominate at drier soil topographic positions ofa chronologically younger age, whereas Alnus- and Myrica-infective Frankia were more abundant in moister, but notsaturated, soils of dune slopes. Owing to their sandy nature,the soils of the ridge tops varied more in moisture contentseasonally than soils of ridge slopes. In a similar studyin Australia with different endemic actinorhizal associates,Dawson et al. (1989) found that Frankia-infective Casuarinaand Allocasuarina units were most abundant in moist, well-aerated sands above the water table in an alluvial floodplainand least abundant in dry, finer textured soils of the adjacentslopes. Thus there may be ecotypic adaptation in strains ofFrankia to soil moisture conditions. Frankia ecotypic adap-tations to soil moisture conditions seemed to follow thesame trends as their host taxa, as suggested by Fig. 2, withShepherdia being adapted to drier site conditions than theAlnus and Myrica species.
Although host adaptations to soil moisture conditions co-incided generally with those of their specific micro-symbionts, soil infectious capacity was not closelyassociated with host-plant cover, which was patchy and scat-tered. Shepherdia canadensis cover was found to be mostabundant in the communities surrounding ridge 1. Alnusincana subsp. rugosa vegetative cover was greatest in thecommunities surrounding ridge 2, while M. gale was re-
stricted in occurrence to the swale between ridges 8 and 9.In contrast with the nodulation trend for Shepherdia, the in-fectious capacity of soils for Myrica and Alnus increasedgenerally with chronological age and successional stage.These trends highlight the possibility that ecological factorsdifferentially influence the infectious capacity of Frankiaecotypes.
The foreslope soils of ridge 1 had a Shepherdia-infectiouscapacity double that of soils from other communities in ourstudy area. This slope supports the vanguard of woody plantcolonizers. Although only a few S. canadensis were foundgrowing nearby, increased rooting of other plants could pro-vide increased soil stabilization and create suitable rhizo-sphere microsites for Frankia. In addition, scattered shrubscould attract birds that may disperse Frankia ingested withsoil invertebrates (Burleigh and Dawson 1995; Paschke andDawson 1993). The adjacent ridge tops had the greatest rela-tive percent cover by S. canadensis of any of the communi-ties sampled and also could have increased the infectiouscapacity of soils downslope through the movement of itsFrankia propagules in water or on soil particles. In a studywhere nodulated Alnus were planted in plots on peat soils,Frankia was detected outside the planting areas only in plotsthat were irrigated (Arveby and Huss-Danell 1988), suggest-ing the movement of Frankia by water. No Frankia wasdetected in lake water or filtered swale water, and less than1 IU Frankia/cm3 was found in unfiltered swale water. NoFrankia was detected in soil of the swales between ridges ifit was submerged to a depth of greater than 21 cm at thetime we sampled. Huss-Danell et al. (1997) found infectiveFrankia in silty river water and superficial river and lakesediments from Sweden and Alaska, but not in 100- to 3000-year-old lake sediments. These results suggest transport ofFrankia in sediments originating recently from proximal
© 2004 NRC Canada
Batzli et al. 707
Fig. 3. Border plot distribution of maximum mean infectious capacities by host plant species in infectious units as a function of per-centage of soil moisture at time of sampling (July 1996). Lines border the highest infectious capacity for a given percent moisture(Shepherdia canadensis, solid line; Myrica gale and Alnus incana subsp. rugosa, broken line).

soils harboring populations of this actinomycete. The mas-sive water volume and low sediment loads of Lake Michiganwaters precludes abundant sediment with associated Fran-kia, but sediment in the waters of interdunal swales adjacentto infective soils did contain infective propagules ofFrankia.
The variability of infectious capacity among soil samplesrepresenting the 12 plots sampled in the community on theforeslope of ridge 1 was high. Burleigh and Dawson (1994)as well as others (Oremus 1980; Paschke and Dawson1992a) found localized variation to be high among popula-tions of infective Frankia in otherwise uniform soils. Thisvariation in infectious capacity may reflect patchy distribu-tion of favorable soil microsites or patchy patterns of disper-sion and growth of infective forms of Frankia.
Frankia strains that nodulate S. canadensis are ecologi-cally distinct from those that nodulate M. gale and A. incanasubsp. rugosa (Huguet et al. 2001, 2004). Evidence suggeststhat ecological factors, particularly those related to succes-sional stage and soil moisture, differentially interact with in-fectious populations of host-specific Frankia genotypes toinfluence the nodulation capacity of a given soil from a LakeMichigan sand dune ecosystem.
Acknowlegements
Thanks are extended to the board and staff of The RidgesSanctuary, Inc., Baileys Harbor, Wisconsin, for permissionto conduct research within the nature preserve and for assis-tance with logistics and partial funding. Research was sup-ported in part by the Illinois Agricultural Experiment Stationthrough USDA McIntire-Stennis Project 875-319.
References
Arveby, A.S., and Huss-Danell, K. 1988. Presence and dispersal ofinfective Frankia in peat and meadow soils in Sweden. Biol.Fertil. Soils, 6: 39–44.
Becking, J.H. 1970. Plant–endophyte symbiosis in non-leguminousplants. Plant Soil, 32: 611–654.
Bermudez de Castro, F., Miguel, C., and Rodrigues-Barrueco, C.1976. A study of the capacity of soil to induce nodules in Alnusglutinosa (L.) Gaertn., and Myrica gale (L.) with special refer-ence to the specificity of the endophytes. Ann. Inst. Pasteur(Paris), 127: 307–315.
Bonham, C.D. 1989. Measurements for terrestrial vegetation. JohnWiley & Sons, New York.
Booth, R.K., and Jackson, S.T. 2000. Development and dynamicsof Lake Superior coastal wetlands [online]. Available fromhttp://asuwlink.uwyo.edu/~rkbooth/michigan/GTB.htm [accessed4 June 2002].
Burleigh, S.H., and Dawson, J.O. 1994. Occurrence of Myrica-nodulating Frankia in Hawaiian volcanic soils. Plant Soil, 164:283–289.
Burleigh, S.H., and Dawson, J.O. 1995. Spores of Frankia strainHFPCcI3 nodulate Casuarina equisetifolia after passage throughthe digestive tracts of captive parakeets (Melopsittacus undu-latus). Can. J. Bot. 75: 1527–1530.
Carson, P.L. 1980. Recommended potassium test. In Recom-mended chemical test procedures for the north central region.Bulletin No. 499 (revised). Edited by W.C. Dahnke. North Da-kota Agricultural Experiment Station, Fargo, N.D. pp. 17–18.
Chapin, F.S., Walker, L.R., Fastie, L.C., and Sharman, L.C. 1994.Mechanisms of primary succession following deglaciation atGlacier Bay, Alaska. Ecol. Monogr. 64: 149–175.
Curtis, J.T. 1959. The vegetation of Wisconsin: an ordination ofplant communities. The University of Wisconsin Press, Madi-son.
Dalling, J.W. 1994. Vegetation colonization of landslides in theBlue Mountains, Jamaica. Biotropica, 26: 392–399.
Dawson, J.O., and Klemp, M.T. 1987. Variation in the capacity ofblack alder to nodulate in central Illinois soils. In Central Hard-wood Forest Conference VI, Knoxville, Tenn., 24–26 February1987. Edited by R.L. Hay, F.W. Woods, and H. DeSelm. Depart-ment of Forestry, Wildlife and Fisheries, University of Tennes-see, Knoxville, Tenn. pp. 255–260.
Dawson, J.O., Kowalski, D.G., and Dart, P.J. 1989. Variation withsoil depth, topographic position and host species in the capacityof soils from an Australian locale to nodulate Casuarina andAllocasuarina seedlings. Plant Soil, 118: 1–11.
Elo, S., Maunuksela, L., Salkinoja-Salonen, M., Smolander, A, andHaahtela, K. 2000. Humus bacteria of Norway spruce stands:plant growth-promoting properties of birch, red fescue, and al-der colonizing capacity. FEMS Microbiol. Ecol. 31: 143–152.
Gauthier, D., Jaffre, T., and Prin, Y. 2000. Abundance of Frankiafrom Gymnostoma spp. in the rhizosphere of Alphitonia neoca-ledonica, a non-nodulated Rhamnaceae endemic to New Caledo-nia. Eur. J. Soil Biol. 36: 169–175.
Huguet, V., Batzli, J.M., Zimpfer, J.F., Normand, P., Dawson, J.O.,and Fernandez, M.P. 2001. Diversity and specificity of Frankiastrains in nodules of sympatric Myrica gale, Alnus incana, andShepherdia canadensis determined by rrs gene polymorphism.Appl. Environ. Microbiol. 67: 2116–2122.
Huguet, V., Batzli, J.M., Zimpfer, J.F., Gourbière, F., Dawson, J.O.,and Fernandez, M.P. 2004. Nodular symbionts of Shepherdia,Alnus and Myrica from a sand dune ecosystem: trends in occur-rence of soilborne Frankia genotypes. Can. J. Bot. 82: 691–700.
Huss-Dannell, K. 1978. Nitrogenase activity measurements in in-tact plants of Alnus incana. Physiol. Plant. 43: 372–376.
Huss-Dannell, K., and Frej, A.K. 1986. Distribution of Frankia insoils from forest and afforestation sites in northern Sweden.Plant Soil, 90: 407–418.
Huss-Danell, K., Uliassi, D., and Renberg, I. 1997. River and lakesediments as sources of infective Frankia (Alnus). Plant Soil,197: 35–39.
Kirsten, W.J. 1983. Rapid automatic high capacity Dumas determi-nation of nitrogen. Microchem. J. 28: 529–547.
Knudsen, D. 1980. Recommended phosphorus tests. In Recom-mended chemical test procedures for the north central region.Bulletin No. 499 (revised). Edited by W.C. Dahnke. North Da-kota Agricultural Experiment Station, Fargo, N.D. pp. 14–16.
Lanyon, L.E., and Heald, W.H. 1982. Magnesium, calcium, stron-tium, and barium. In Methods of soil analysis. Part 2. Chemicaland microbiological properties. 2nd ed. Edited by A.L. Page.Agronomy 9(2): 247–262.
Lawrence, D.B., Schoenike, R.E., Quispel, A., and Bond, G. 1967.The role of Dryas drummondii in vegetation developmentfollowing ice recession at Glacier Bay, Alaska, with special ref-erence to its nitrogen fixation by root nodules. J. Ecol. 55: 793–813.
Lichter, J. 1998. Primary succession and forest development oncoastal Lake Michigan sand dunes. Ecol. Monogr. 68: 487–510.
Link, E.G., Elmer, S.L., and Vanderveen, S.A. 1977. Soil survey ofDoor County, Wisconsin. USDA Soil Conservation Service incooperation with Research Division of the College of Agricul-tural and Life Sciences, University of Wisconsin, Madison.
© 2004 NRC Canada
708 Can. J. Bot. Vol. 82, 2004

Mclean, E.O. 1980. Recommended pH and lime requirement tests.In Recommended chemical test procedures for the north centralregion. Bulletin No. 499 (revised). Edited by W.C. Dahnke.North Dakota Agricultural Experiment Station, Fargo, N.D.pp. 14–16.
Oremus, P.A.I. 1980. Occurrence and infective potential of theendophyte of Hippophae rhamnoides L. ssp. rhamnoides incoastal sand dune areas. Plant Soil, 56: 123–139.
Paschke, M.W., and Dawson, J.O. 1992a. Frankia abundance insoils beneath Betula nigra and other non-actinorhizal woodyplants. Acta Oecol. 13: 407–415.
Paschke, M.W., and Dawson, J.O. 1992b. The occurrence of Fran-kia in tropical forest soils of Costa Rica. Plant Soil, 142: 63–67.
Paschke, M.W., and Dawson, J.O. 1993. Avian dispersal of Fran-kia. Can. J. Bot. 71: 1128–1131.
Rhoades, J.D. 1982. Cation exchange capacity. In Methods of soilanalysis. Part 2. Chemical and microbiological properties. 2nded. Edited by A.L. Page. Agronomy 9(2): 247–262.
Righetti, T.L., Chard, C.H., and Backhaus, R.A. 1986. Soil andenvironmental factors related to nodulation in Cowania andPurshia. Plant Soil, 91: 147–160.
Rodriguez-Barrueco, C. 1968. The occurrence of the root noduleendophytes of Alnus glutinosa and Myrica gale in soils. J. Gen.Microbiol. 52: 189–194.
Sherrill, M.G. 1978. Geology and ground water in Door County,Wisconsin, with emphasis on contamination potential in theSilurian Dolomite. US Geological Survey Water-Supply Paper2047. US Government Printing Office, Washington, D.C.
Smolander, A. 1990. Frankia populations in soils under differenttree species — with special emphasis on soils under Betulapendula. Plant Soil, 121: 1–10.
Smolander, A., and Sundman, V. 1987. Frankia in acid soils of for-ests devoid of actinorhizal plants. Physiol. Plant. 70: 297–303.
Smolander, A., Nurmiaho-Lassila, E., and Sundman, V. 1988. Ef-fect of a clay mineral (montmorillonite) on the nodulation ofAlnus and on the nitrogenase activity of Frankia in pure culture.Symbiosis, 6: 37–52.
Thompson, T.A., and Baedke, S.J. 1997. Strandplain evidence forlate Holocene lake-level variations in Lake Michigan. GSA Bul-letin, 109: 666–682.
Tortosa, R.D., and Cusato, M. 1991. Effective nodulation ofrhamnaceous actinorhizal plants induced by air dry soils. PlantSoil, 131: 229–233.
Trick, J.A. 1983. Preliminary report on the vascular flora of theRidges Sanctuary.Available from the Ridges Sanctuary, Inc.,Baileys Harbor, Wisc.
Van Dijk, C. 1979. Endophyte distribution in the soil. In Symbioticnitrogen fixation in the management of temperate forests. Editedby J.C. Gordon, C.T. Wheeler, and D.A. Perry. Oregon StateUniversity Press, Corvallis, Oreg. pp. 84–94.
Vitousek, P.M., and Walker, L.R. 1989. Biological invasion byMyrica faya in Hawaii: plant demography, nitrogen fixation,ecosystem effects. Ecol. Monogr. 59: 247–265.
Walker, L.R. 1993. Nitrogen fixers and species replacements in pri-mary succession. In Primary succession on land. Edited by J.Miles and D.W.H. Walton. British Ecological Society, SpecialPublication 12. Blackwell Scientific Publications, Boston, Mass.pp. 249–272.
Whitney, D. 1980. Micronutrient soil tests — zinc, iron, manga-nese and copper. In Recommended chemical test procedures forthe north central region. Bulletin No. 499 (revised). Edited byW.C. Dahnke. North Dakota Agricultural Experiment Station,Fargo, N.D. pp. 18–21.
Young, D.R., Sande, E., Peters, G.A. 1992. Spatial relationships ofFrankia and Myrica cerifera on a Virginia, USA barrier island.Symbiosis, 12: 209–220.
Zimpfer, J.F., Smyth, C.A., and Dawson, J.O. 1997. The capacityof Jamaican mine spoils, agricultural and forest soils to nodulateMyrica cerifera, Leucaena leucocephala, and Casuarina cun-ninghamiana. Physiol. Plant. 99: 664–672.
Zimpfer, J.F., Kennedy, G.J., Smyth, C.A., Hamelin, J., Navarro,E., and Dawson, J.O. 1999. Localization of Casuarina-infectiveFrankia near Casuarina cunninghamiana trees in Jamaica. Can.J. Bot. 77: 1248–1256.
© 2004 NRC Canada
Batzli et al. 709





























