Embed Size (px)
Citation preview

Cytologie and Cytotoxic Changes Induced by Nucleic Acids*
ALBERTG. SMITH(Department of Pathology, Duke University Medical Center, and Veterans Administration Hospital, Durham, North Carolina)
SUMMARY
Nucleic acids and protein prepared from Sarcoma 180, from Ehrlich ascites tumor,from NCTC 929 cells derived from strain L, and from normal mouse liver, wheninjected subcutaneously in mice with cultured viable NCTC 929 strain L cells, occasionally induce cytologie atypia in the NCTC 929 cells, characterized by giantcells, abnormal cytoplasmic shapes and staining, large nuclei, and prominence ofnuclear chromatin patterns.
Cytologie atypia was not induced in mouse host cells adjacent to the site of nucleicacid injection. Histones without nucleic acids did not induce atypia in concomitantlyinjected viable NCTC 929 cells in the mouse. Injections of NCTC 929 cells alonedid not present the same atypia as was seen when nucleic acids were injected withthe NCTC 929 cells.
Ascites tumor, mouse liver, and Sarcoma 180 nucleic acids and nucleic acids-proteinwere cytotoxic to NCTC 929 cells in tissue cultures at varying concentrations above250 milligrams per milliliter of culture fluid. Cytotoxicity was produced by deoxy-ribonucleic acid, ribonucleic acid, or a mixture of both deoxyribonucleic acid and ri-bonucleic acid.
The atypia induced in NCTC 929 cells in mice by nucleic acid-protein and thecytotoxicity produced by similar nucleic acid preparations acting upon NCTC 929cells in tissue culture may be related.
This study was undertaken in an attempt to determinewhether nucleic acids have any morphologic or cytologieeffect discoverable by observation of injected animals,their treated tissues, or cells that might bear some relationto the development of tumors or to greatly altered metabolism in the affected cells.
MATERIALS AND METHODSCells of NCTC clone 929 derived from strain L (14),
free of pleuro-pneumonia-like organisms, were maintainedin continuous culture in Roux bottles or Porter flasks bymultiple trypsinations and transfers. These cells were alsofree from human cell contamination as tested by Coomb's
mixed hemagglutination technic (9). Tissue culture medium was 80 per cent Eagle HeLa medium (12) (Difco);20 per cent horse serum (Difco); penicillin, 100 units/ml;and streptomycin, 50 units/ml. This medium waschanged every other day. L cells generally had overgrown the Roux flask and required transfer every 4 days.For use in animal injections L cells were obtained fromtrypsinated cultures suspended in tissue culture medium.A cell count of the suspended cells was then made, withappropriate dilutions and a clinical hemocytometer counting chamber.
* Supported by Grant C-3894 from the National Cancer Institute and in part by American Cancer Society institutional grant,Duke University.
Received for publication August 20, 1963.
Ehrlich ascites tumor and Sarcoma 180 (S-180) weremaintained by repeated intraperitoneal and subcutaneoustransfers in Swiss mice. These Swiss mice were also usedfor the nucleic inoculation experiments and were inbred atthe laboratories of the North Carolina State Board ofHealth from mice originally obtained from the Swiss-Webster colony of the Rockefeller Institute. Samples oftissue cultures, culture media, and tissue for nucleic acidpreparation were cultured at frequent intervals for bacteria to exclude bacterial contamination.
Nucleic acid with nucleoprotein was prepared as follows:All steps were done in an ice bath as quickly as possible.Ehrlich ascites tumor cells were obtained by centrifugingperitoneal fluid from tumor-bearing mice. S-180 tumorcells were obtained from solid viable tumors in mice.Liver, lungs, heart, kidney, and spleen of normal mice wereobtained, minced, and mixed. Multiple large flasks of 929cells were grown in tissue cultures, the cells scraped fromglass, and the resulting cells and culture medium centri-fuged; the culture medium was then decanted, and the remaining cells were employed for further processing. Eachof these tissues was processed as follows: The tissues wereput into an all-glass Ten Broeck homogenize! and homogenized in 5 ml. 0.1 M sodium citrate for 3 minutes,centrifuged at 2000 r.p.m. for 15 minutes, and the supernatant was discarded. This procedure was repeated 2more times, yielding about 1 ml. of packed cell residue.
603
Research. on February 18, 2018. © 1964 American Association for Cancercancerres.aacrjournals.org Downloaded from

604 Cancer Research Vol. 24, May 1964
The residue was homogenized in 9 ml. 0.14 sodium chloride,0.015 M sodium citrate buffer, and the homogenate waspoured into a 50-ml. centrifuge tube. The amount wasmeasured, and 3 volumes of cold 95 per cent ethanol wasgradually added, with stirring. A stringy, white precipitate appeared in the opalescent homogenate. This stringyprecipitate was collected on a glass rod, removed from theopalescent protein homogenate, placed in 95 per centalcohol, and packed by centrifuging at 2000 r.p.m. for 20minutes. Additional protein can be left with the nucleicacids by centrifuging and packing the precipitate in theopalescent solution without removal on the glass rod. Theamount of precipitate was noted and the supernatant discarded. A small portion of the precipitate was homogenized with an equal amount of 0.14 M sodium chloride,0.015 M sodium citrate buffer for 3 minutes. A drop ofthis homogenate was examined on a slide previouslycoated with methyl-green-pyronin stain. Smears of thehomogenate were air-dried, fixed in formalin, and stainedwith hematoxylin and eosin or by the Feulgen technic forexamination for the presence of intact cells. No intactcells were found. Eagle HeLa (Difco), 40 per cent horseserum (Difco) tissue culture medium was added in anamount 5 times the amount of the remaining white precipitate, and the mixture was homogenized for 3 minutes.The DNA preparation was used immediately, or storedtightly stoppered at 4°C. in a refrigerator for brief periods.
This crude nucleic acid preparation gave negative reactions to biuret, ninhydrin, sulfosalicylic acid, xanthopro-teic, and Millon's tests for protein. The preparation had
a peak absorption at 260 m/n,and from 3 to 5 per cent of itsdry weight was phosphorus, indicating that about 40 percent of the preparation was nucleic acids. Analysis bypaper chromatography and by the amino acid analyzer revealed contamination with amino acids, with significantamounts of arginine, lycine, and histidine being specificallyidentified. Hall and Gale (20) found that their bacterialpreparations treated by a similar method contained about30 per cent carbohydrates.
Purified ascites tumor and S-180 nucleic acids were prepared by the chloroform-amyl alcohol method of McCartyand Avery (32) with protein extractions proceeding untilno cloudy material remains (over ten extractions). Thesepreparations had a peak absorption at 260 m^. From 5 to6 per cent of dry weight was phosphorus, indicating, inproportion to theoretical phosphorus in nucleic acids(about 9 per cent), that about 60 per cent of the preparations was nucleic acid. The solution was clear and viscid,in contrast to the opalescent fluid yielded by the nucleicacids-protein procedure described above. Analysis bypaper chromatography revealed much less amino acid contamination than in the nucleic acids-protein preparation, but some amounts of an amino acid that was probably glycine were still detectable.
Approximately 55 per cent of the nucleic acid in thepreparations were ribonucleic acid and 45 per cent deoxy-ribonucleic acid, as determined by phosphorus determinations on acid-soluble and acid-insoluble fractionsaccording to the Berenblum-Chain-Heatly procedure asgiven by Schmidt (42). Eight per cent of the preparationswas nitrogen, and N/P ratios were 1.5-1.6. The approxi
mate 40 per cent of the preparations not accounted for asphosphorus- and nitrogen-containing substances (i.e.,nucleic acids) was possibly carbohydrate, such as deoxy-ribose and ribose. Colorimetrie determinations of deoxy-ribonucleic acid by the diphenylamine and cysteine methods, and of ribonucleic acid by the orcinol technic, all asoutlined by Bische (10), gave optical densities for ourpreparations greater than the theoretical optical densityvalues for our DNA and RNA as calculated from the optical densities of commercial deoxyribonucleic acids of 8 percent phosphorus content (about 89 per cent purity), andribonucleic acid of 5.2 per cent phosphorous content (58per cent purity), in both the deoxyribonucleic acid andribonucleic acid colorimetrie tests. In each case the percent increase in optical density over theoretical values wasapproximately 50 per cent. Since few substances otherthan DNA and deoxyribose produce the required colorand absorbance in the diphenylamine and cysteine reactions, the increase in color over theoretical values produced by our preparations is probably due to deoxyribose.The increase in color in the orcinol reaction may be due toexcess ribose or to the deoxyribonucleic acid present. Byoptical density determinations on the Beckman DU spec-trophotometer, our preparations generally satisfy (within90 per cent) the rule that 45 Mg-of nucleic acids produces anoptical density of 1.0 at 260 imt for a 1-cm. light path (24).Our preparations generally gave a 24 per cent rise in O.D.following sodium hydroxide denaturation (5). Phenolextractions of nucleic acids were not employed because ofpossible contamination of the nucleic acids injected intoanimals or added to tissue culture medium with phenol,which might in itself produce atypical cells or be toxic tocells in culture.
To insure potency of the DNase and RNase (Worth-ington) employed, these enzymes were tested upona mixed DNA and RNA preparation by treatment ofthese solutions under conditions as closely resemblingthe experimental treatment of the culture medium withDNA and RNA as possible. Each enzyme was tested for24-37 per cent increase in optical density that occurs at260 m/* following depolymerization of DNA (27) or RNA(30) with the DNase or RNase according to the method ofHotchkiss (24).
Experiments were performed as follows:Animal injection experiments.—Animal injection experi
ments were performed as listed in Table 1, with the methods of preparation, substances, and mice as describedabove. In Experiment 16 the Ehrlich ascites tumor cellswere injected in the mouse peritoneal fluid in which theywere removed from the mouse. In Experiment 17, about0.5 cc. of Sarcoma 180 tissue was injected after removal assolid tumor from the donor mouse and mincing.
Tissue culture experiments.—A. The effect of Ehrlich ascites tumor nucleic acids-
protein upon NCTC 929 cells: Ehrlich ascites tumor nucleic acids-protein was added to tissue cultures of NCTC929 cells in amounts of 0.2 mg., 0.5 mg., l mg., and 5 mg.per milliliter of culture fluid. Five flasks of 929 cells weretested at each concentration, and five control flasks containing only culture medium were maintained. Cultureswere examined twice daily, and final interpretation of
Research. on February 18, 2018. © 1964 American Association for Cancercancerres.aacrjournals.org Downloaded from

SMITH—Cytologieand Cytotoxic Changes Induced by Nucleic Acids 605
TABLE 1HISTOLOGICEFFECTSOF NUCLEICACIDSANDCONTROLSUBSTANCESIN MICE
PreparationinjectedS-180
nucleic acids-protein & 929cellsAscites
tumor nucleic acids-protein &929cells929
cell nucleic acids-protein & 929cellsLiver
nucleic acids-protein & 929cellsHistones
& 929cellsAscites
tumor nucleic acids, purified, &929cellsS-180
nucleicacids-proteinAscites
nucleicacids-proteinAscites
nucleic acids-protein (4 injections) at weeklyintervals929
cells nucleicacids-proteinLiver
nucleicacids-proteinMixed
organ nucleic-acidsproteinAscites
nucleic acids,purifiedS-180
nucleic acids,purified929
cells &bufferAscites
cellsS-180
cellsAmount
nucleic acidpreparationor histoneinjected
(mg.)55555555555555000Amount
cellsinjected
in 0.1 ml.Eagle-HeLamedium500,000500,000500,000500,000500,000500,00000000000500,0001,000,000.5Amount
NaCl-Nacitratebuffer
injected.06.06.06.06.5.06.5.5.5.5.5.5.5.5.060Amount
Eagle-HeLa
mediuminjected.29.29.29.290.2900000000.29.5pH7.67.67.67.67.67.67.67.67.67.67.67.67.67.67.6Volume
of injection
(ml.).5.5.5.5.5.5.5.5.5.5.5.5.5.5.5.1.5No.
micegiveninjections1020510205161161010131010201010Days
followinginjectionanimals
sacrificed3
and73
and73
and73
and53
and73
and710-90773
and73
and77-60777107Histologie
demonstration of marked atypia,
cytologie variationwith giant cells,giant nuclei andpyknoticforms++
++
++
inhibitory effect was made 3 days following the additionof the nucleic acids-protein. The 929 cells were grown oncoverslips in Porter flasks, and at the end of 3 days thecoverslips were removed and the cells fixed and stained.
B. Effect of Ehrlich ascites tumor nucleic acids uponNCTC 929 cells: Ehrlich ascites tumor nucleic acids wereadded to tissue cultures of 929 cells in amounts of 0.05,0.1, 0.25, 0.5, 1, 2, 2.5, 3 and 5 mg/ml, under conditionsotherwise similar to experiment A.
C. Effect of Sarcoma 180 nucleic acids-protein uponNCTC 929 cells: Under conditions similar to those inExperiment A, the effect of Sarcoma 180 nucleic acids-protein upon 929 cells was tested at a concentration of5 mg. of nucleic acids-protein/ml of culture fluid.
D. Effect of mouse liver nucleic acids upon NCTC 929cells : Under conditions similar to those of Experiment A,the effect of mouse liver nucleic acids upon 929 cells wastested in concentrations of 0.123, 0.25, 0.5, 1.0, 1.5, 2.0,3.0, and 5.0 mg/ml of culture fluid.
E. Effect of nucleic acid derivatives upon NCTC 939cells: Under conditions similar to those of experiment A,the following derivatives were tested for effect upon 929cell growth at the following concentrations: adenine, 0.25mg/ml; guanine, 0.25 mg/ml; deoxycytidine, 0.25 and5 mg/ml; and deoxyguanosine, 0.25 and 5 mg/ml.
F. Effect of DNase and RNase upon the effect of Ehrlichascites tumor and mouse liver nucleic acids upon NCTC929 cells: To test the effect of DNase upon the effect ofEhrlich ascites tumor and mouse liver nucleic acids upon929 cells, 60 Mg-of DNase (Worthington) and 0.3 mg. ofMgS04 were added to each milliliter of culture fluid oftest flasks and control cultures. Test flasks contained 2.0,2.5, 3.0, and 4.0 mg. of ascites nucleic acids and 0.125,0.5, 1.0, 1.5, 2.0, and 3 mg. of liver nucleic acid per ml. ofculture fluid. In some experiments the nucleic acids werepretreated with DNase in prescribed buffer, and thetreated nucleic acid preparations then added to test flasks.Similarly, nucleic acids were treated with RNase by the
Research. on February 18, 2018. © 1964 American Association for Cancercancerres.aacrjournals.org Downloaded from

606 Cancer Research Vol. 24, May 1964
addition of a small amount of RNase to the nucleic acids.The nucleic acids were pretreated with RNase in prescribed buffer, and the remaining DNA re-precipitatedwith ethanol and added to the culture medium.
G. Effect of histones (Worthington) upon NCTC 929cells: Three days following implantation of NCTC 929cells, 0, 0.5 mg., l mg., 5 mg., and 10 mg. of histones(Worthington) per ml. of culture medium were added tofresh culture medium in each test flask. Several flasksat each concentration were tested, with the flasks with noadded histone serving as controls.
RESULTS
Results of these various experiments with mice are summarized in Table 1. For the purpose of the morphologicobservations desired in these experiments, it was foundthat the nucleic acids-protein was required, since thispreparation yielded a proteinaceous material that precipitated as an amorphous mass at the site of injection(Figs. 1, 2) and contained Feulgen-positive material presumed to be deoxyribonucleic acid. Nucleic acid preparations made by the chloroform-amyl alcohol protein extraction procedure did not remain localized at the site,leaving little to be observed. In mice given injections ofthe purified nucleic acids, a small nodule was palpable for3 or 4 days following injection; but this subsequentlydisappeared, and only minimal microscopic changes offibrosis were apparent.
Large vesicular nuclei, mitotic figures, and prominentchromatin network are characteristic of NCTC 929 cells,Ehrlich ascites tumor cells, and Sarcoma 180 cells. Theascites tumor, S-180, and in some instances NCTC 929cells produced spherical masses of tumor when injectedsubcutaneously. The variation categorized as "marked
atypia, cytologie variation with giant cells and giantnuclei and pyknotic forms" was found only in preparationsin which NCTC 929 cells and nucleic acids-protein wereinjected (Figs. 1, 3-6). The altered cells were present atthe junction zone between the mass of amorphous nucleo-protein material and more normal fibroblast, inflammatorycell or 929 cell growth and did not appear in great abundance. Liver nucleic acids-protein also produced themarkedly atypical changes (Fig. 5) produced by S-180,ascites tumor (Fig. 1), or 929 cell nucleic acids-protein(Fig. 6), indicating that this effect is not specific to tumornucleic acids but is a property of nucleic acids in general.
The control experiments in which 929 cells alone wereinjected into Swiss mice did not produce any of thesemarkedly atypical cells, although some injections of 929cells did result in small viable tumors composed of 929cells that did not grow beyond 2-5 mm. in size. Nogrossly obvious tumors or progressive tumors ever developed when strain L-929 cells were injected into Swissmice. NCTC mouse fibroblast cultures vary considerablyin their capacity to produce tumors (41). NCTC 929strain L cells have been found to produce tumors only inC3H mice (40). Regressive growth of 929 cells, similarto our results with mice, have been produced in unconditioned hamsters given doses of not less than 1,000,000cells (18). Control experiments in which histone and Lcells were injected produced amorphous masses similar to
the masses produced by nucleic acids and nucleoprotein.However, markedly atypical cells were not seen in thesehistone and 929 cell injection experiments. Histones(Worthington) were chosen for this test, because they arethe most likely form of protein contaminant that could bepresent in the nucleic acids-protein preparations. Nonucleic acid preparation, whether with protein or purified,produced marked atypia in the normal mouse connectivetissues. Only inflammation and reactive fibrosis withamorphous nucleoprotein masses were produced.
Various mice and additional newborn mice given different nucleic acid preparations were followed for periodsranging up to 6 months. No gross evidence of tumor,either by observation of the animal or by autopsy, wasever found, except where viable ascites tumor, S-180, orlarge doses of L cells were given. The liver, lungs, kidneys, and spleen from some mice given S-180 nucleic acidsand nucleoprotein were examined microscopically, but nounusual nuclear changes were found. Some Swiss micewere sensitized to S-180 nucleic acids and nucleoprotein,as well as mixed organ nucleic acids and nucleoprotein, byinjection of 5 mg. of sensitizing nucleic acid preparations 4times at weekly intervals, and these mice were challenged6 weeks later with injections of 0.5 cc. of minced S-180tumor only. Control mice were given 0.5 cc. of mincedS-180 tumor at the time of challenge. The challenge andcontrol groups of mice both developed S-180 tumors, andno differences between the tumors or tumor cells of eithergroup were noted.
Cytotoxicity was noted in cultures containing each ofthe types of nucleic acids (Ehrlich ascites tumor nucleicacids-protein, Ehrlich ascites tumor nucleic acids, Sarcoma 180 nucleic acids-protein, and mouse liver nucleicacids) and was usually first evident after 12 hours' in
cubation with increasing detrimental effects up to 3 daysof incubation (Figs. 7-11). As previously described (43),this toxic effect usually manifests itself within the first 12hours by rounding and shrinkage of cells. Three daysfrom the time the nucleic acids were added, no cells oronly a few pyknotic cells are usually observable (Fig. 11).The Ehrlich ascites tumor nucleic acids-protein was cyto-toxic to some degree at a concentration of .5 mg/ml ofculture fluid and was markedly cytotoxic at all greaterconcentrations. Slight cytotoxicity was noted in culturesto which Ehrlich ascites tumor nucleic acids were added ina concentration of .1 mg/ml, and marked cytotoxicity atall greater concentrations. Drying and freezing did notaffect the cytotoxicity of Ehrlich ascites tumor nucleicacids. Sarcoma 180 nucleic acids and nucleoprotein weremarkedly cytotoxic at the concentrations tested. Mouseliver nucleic acids were markedly cytotoxic at concentrations of 0.25 mg/ml of culture fluid and above. Adenine,guanine, thymidine, deoxyadenosine, and deoxycytidinedid not affect 929 cells cultures in the concentrationstested. Deoxyguanosine had no effect at 0.25 mg/ml, butwas cytotoxic at 5 mg/ml.
Under proper conditions, DNase partially preventedthe cytotoxic action of both ascites tumor nucleic acidsand mouse liver nucleic acids at all concentrations tested(Fig. 9). RNase also partially prevented the cytotoxicaction of both types of nucleic acids, perhaps to a slightly
Research. on February 18, 2018. © 1964 American Association for Cancercancerres.aacrjournals.org Downloaded from

SMITH—Cytologieand Cytotoxic Changes Induced by Nucleic Acids 607
greater degree than the prevention of cytotoxicity byDNase (Fig. 10). The use of combined DNase and RNasealmost completely prevented the cytotoxic action of bothEhrlich ascites tumor and liver nucleic acids (Fig. 8).Since the depolymerized nucleic acids were left on the 929cell cultures, absence of cytotoxicity under these conditionsimplies that the depolymerized elements of nucleic acidsare not cytotoxic at the concentrations tested. Controlcultures, including controls with culture medium only,those with MgS04 added, and those with DNase andRNase, showed little or no growth inhibition or cytotoxicity. The nuclease experiments sometimes requiredseveral repeated performances because of variation inresults. Titrations for cytotoxicity of nucleic acids occasionally required several re-verification performances.
Study of the effect of histones (Worthington) uponNCTC 929 cells indicates that no cytotoxicity was observable when 0.5 mg. of histones was added to the culturemedium. Increase in groups of shrunken cells of a moderate degree was observed when 1 mg. of histones was employed. Five mg. of histones produced a heavy deposit ofmaterial over the cells and adherent to the cells ; there wasan increase in numbers of shrunken cells, but there weremany good fusiform viable 929 cells. At 10 mg. of histones the precipitate was very dense, there was a definitereduction in viable cells, but many fusiform viable 929cells remained. These effects were observed for 1-5 daysfollowing the addition of histones. These findings oftoxicity to histones (Worthington) are in sharp contrastto the cytotoxic effect of nucleic acids upon 929 cells.Twelve hours following the addition of nucleic acids tothe medium, vacuolization and an increase in shrunkencells were noted, with complete destruction of all cellswithin 24-48 hours, with concentrations of nucleic acidsas low as 0.24 or 0.5 mg/ml of culture medium.
An effort was made to evaluate increases in mitoses,giant nuclei, multiple nuclei, or bizarre cellular forms of929 cells in these tissue culture experiments. The tissuecultures and their fixed and stained slides were each examined, and a qualitative impression of each was determined. No significant difference between control culturesand the variously treated cultures was found, except forsome possible increase in the giant cell forms in culturesto which DNase or RNase had been added.
DISCUSSION
Under certain conditions nucleic acids induced an atypiain NCTC 929 cells injected into mice that did not appearwhen either 929 cells or nucleic acid preparations wereinjected alone into mice. Incorporation of nucleic acidsor nucleoprotein into cells in vitro and in vivo has beendemonstrated (7, 8, 11, 23, 37). Nucleic acid materialsused in our experiments may have been incorporatedsimilarly into the viable injected 929 cells that subsequently became histologically atypical. These atypicalchanges found in 929 cells injected with nucleic acids maypossibly bear some relation to nuclear atypia found incells adjacent to tumor in histologie sections of human oranimal tumors. The NCTC strain L cell or 929 cell canproduce grossly recognizable tumors in C3H mice undercertain conditions (13, 401. Tt,is understandable that 929
cells after trypsination and exposure as single cells tonucleic acids may be in a more delicate physiologic statehan normal mouse cells, serving as an indicator of the
presence of foreign nucleic acids where normal host cellswill not show any effect.
Prior reports indicate that the cells, nuclei, mitotic rates,or nucleic acid metabolism in a host may be affected bythe presence of a tumor or by injection of nucleic acids.Griffin (19) finds that mammary carcinoma, melanoma,and lymphosarcoma in mice cause an increase in DNAmetabolism of adjacent muscle tissues. Anderson (1)et al. find the turnover of DNA and RNA to be higher inlivers of tumor-bearing mice than in normal mice. Increased mitotic rates in livers of tumor-bearing mice andrats have been reported (2, 31). Breast cancers of micemay cause the appearance of abnormally large cells withlarge nuclei and periodic acid-Schiff-positive nucleoli inthe liver (29). Deoxyribonucleic acid from herring spermhas been used by Hewer and Meek (22) to induce cancerof the intestine in mice. Lacour et al. (28) have injectedRNA prepared from lymph nodes of humans with leukemiainto the abdomen of newborn Swiss mice, producing bizarretumors. Injection of DNA from one strain of duck issaid to produce pigment changes in another strain, presumably by somatic mutation (6). Some of these effectsare most difficult to reproduce (4).
High concentrations of nucleic acids were lethal toNCTC 929 cells under certain conditions in tissue culturein concentrations greater than 200 us/ml of culture medium. The nature of this cytotoxic action of nucleic acidsupon cells in tissue culture (43) is unknown. A similarcytotoxic effect has been reported by Floersheim (17),who found DNA-RNA preparations made by the phenolmethod of Kirby from mouse spleens were cytotoxic toisologous bone marrow cells after 2 hours' incubation in
concentrations of 0.35 mg nucleic acids/ml with subsequent reinjection of the treated marrow cells into miceinjured by dimethyl-myleran. Fahmy and Fahmy (16)report a significant increase in the incidence rate of sex-linked recessive lethals when 1 per cent DNA from adultDrosophilia melanogaster was injected into the hemocoel ofother Drosophilia melanogaster.
As originally demonstrated by Avery, MacLeod, andMcCarty (3), pneumococcal-transforming principle iscapable of inducing transformation in amounts rangingfrom 0.02 to 0.003 Mg-in 2.25 ml. of medium. The toxicdose of nucleic acids in our studies is effective in minimumconcentrations varying from 200 to 2000 Mg/ml of culturefluid. The cytotoxic effect is manifest within 12 hours byvacuolation of the cultured cells, followed by rounding upof the cells, pyknosis, and cell death. All the cells in aculture may be killed in from 24 to 48 hours. Avery,MacLeod, and McCarty produced pneumococcal transformation after incubation periods of from 18 to 24 hours.Although these incubation times are similar, the findingsthat lO^lO5 times as much nucleic acid is required forcytotoxicity as compared with transformation effects uponbacteria mean that this cytotoxic effect is not the sametype of effect as the usual transforming effect. Cytotoxicity may take place partially at mitosis, since from 24to 48 hours are usually required before nearly all cells are
Research. on February 18, 2018. © 1964 American Association for Cancercancerres.aacrjournals.org Downloaded from

608 Cancer Research Vol. 24, May 1964
dead. The original strains of NCTC strain L cells roughlydouble in number in from 24 to 48 hours (39), indicatingthat most cells have entered mitosis in this period. However, vacuolization observed in our cultures sometimeswithin 4-6 hours and the cytotoxic effect produced byFloersheim after 2 hours' incubation indicate that mitosis
may not be required for the cytotoxicity.Morgan (33) found adenine, guanine, hypoxanthine,
xanthine, thymine, uracil, adenosine-5-phosphate, andadenosine triphosphate inhibitory to strain L cells in tissueculture in unspecified amounts. Waymouth (47) reported guanine inhibitory (36) and adenine and adenosinetoxic to lymphocytes at 2 niM (270 Mg/ml) (46). Adenosine, adenylic acids, and adenosine triphosphate have beenreported to cause pre-prophasic inhibition of mitosis inchick fibroblasts in concentrations as low as 0.6 mM withnucleolar fragmentation, without apparent damage tointerphase cells (25). Harris (21) found some nucleotidesand nucleosides (1-100 mg/100 ml) and purine andpyrimidine bases at concentrations of 1-10 mg/100 ml tobe inhibitory. Evans, Fioramanti, Randolph, and Earle(15) reported that thymidine (1 mg/100 ml), deoxyguano-sine (1 mg/100 ml), and 5-methylcytosine (0.01 mg/100 ml) inhibit the growth of tissue cells in vitro. Theinhibitory action of thymidine has been shown to be prevented by deoxycytidine (34). The inhibitory effect ofdeoxyadenosine has been found to be an inhibitory effectupon DNA synthesis, and this inhibitory effect has beenfound to be abolished by adenosine (26). It has beenassumed that these inhibitory effects by deoxyadenosineand thymidine and their prevention may play a part hinatural nucleic acid synthesis (26). Vaccinia virus multiplication in mouse and chick cells in tissue culture has beengreatly inhibited by the addition of 50 Mg-of nucleic acidof heterologous species per ml. of culture medium (38).This latter effect was produced by the treatment of cellswith nucleic acid prior to the addition of vaccinia virusand has been postulated to be due to induced productionof interferon in the test cells by the heterologous nucleicacid. Here again, the effect was produced by small concentrations of nucleic acid and probably does not representthe mechanism by which cytotoxicity is produced bynucleic acids. Whether the inhibitory effect of nucleicacids upon cell growth which we report is a metabolicallyuseful mechanism in vivo and whether the effect is accomplished without some breakdown of the nucleic acidsare unknown.
Syzbalska and Syzbalski (44,45) have altered an inosinicacid pyrophosphory lase-negative strain of human cell line
D98, incapable of hypoxanthine utilization as a sole purinesource (D98/Ah-2), into a strain inosinic acid pyrophos-phorylase-positive, capable of hypoxanthine utilization,with the use of DNA prepared from an originally inosinicacid pyrophosphorylase-positive, hypoxanthine-utilizingstrain of D98 cells in concentrations of 1-100 Mg/ml ofmedium. Niu, Cordova, and Niu (35) treated Nelsonmouse ascites cells with RNA prepared from mouse andcalf liver hi concentrations of approximately 0.6-2 mg/ml(O.D., 25-100/ml. at 260 m^) and reduced the tumor-producing capacity of such cells from 97 to 10-20 per cent.This reduction in tumor-producing capacity was accompanied by an induced biosynthesis by the treated cellsof an albumin-like protein. Thus, DNA can induce transmissible alterations in cell metabolism in concentrationsof 1-100 Mg/ml, and RNA can induce alterations hicell metabolism in treated cells in concentrations of 600-2000 Mg/ml. The concentration required for the RNAeffect upon Nelson mouse ascites cells (35) is approximately the same we find required for cytotoxicity.Although cytotoxicity in our systems can be producedalmost uniformly, certain variations in the system, suchas the employment of heterologous cells, variation inserum content, or other unknown factors may result infailure of the system to exhibit cytotoxicity. The experiments of Floersheim (17) and the work reported hereindicate that nucleic acids are cytotoxic when homologouscells and nucleic acids are used. All the factors concerned in the production of cytotoxicity are not known.It is possible that the induction of cellular atypia by nucleic acids in 929 cells injected into the mouse and thecytotoxicity of nucleic acids to 929 cells in tissue culturemay be related phenomena.
ACKNOWLEDGMENTS
The author wishes to acknowledge the help received fromHertha R. Cress, Ph.D., and Mrs. Johanna Eiring in this work.
REFERENCES
1. ANDERSON,E. P.; YEN, C. Y.; MANDEL, H. G.; AND SMITH,P. K. Ureidosuccinic Acid as a Precursor of Nucleic AcidPyriraidines in Normal and Tumor-bearing Mice. J. Biol.Chem., 213:625-33, 1955.
2. ANNAU, E.; MANQENILLI,A.; ANDROTH, A. Increased Weightand Mitotic Activity in the Liver of Tumor-bearing Rats andMice. Cancer Res. 11:304-7, 1951.
3. AVERT, O. T.;MACLEOD, C.M.; ANDMCÃœARTT,M. Studies onthe Chemical Nature of the Substance Inducing Transformation of Pneumococcal Types. J. Exp. Med., 79:137-57, 1944.
4. BEARN, J. G. An Attempt To Produce Malignant Change withDeoxyribonucleic Acid from Rat Sarcoma and Hepatoma.Nature, 184:824, 1959.
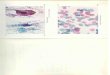
FIG. 1.—Border of injected NCTC 929 strain L cells and amorphous ascites tumor nucleic acids-protein preparation (right) andreaction zone of cells (left) in the mouse. Note large, bizarre,multinucleated cells with prominent nucleoli. X 245.
FIG. 2.—Injection site of ascites tumor nucleic acids-proteinonly. The amorphous mass of injected material is at upper right.The surrounding reaction zone is comprised of fibroblasts and inflammatory cells, but no bizarre cells. X 271.
FIGS. 3, 4.—Higher-power views of the bizarre cells in the reaction zone near the injected ascites nucleic acid preparation andNCTC 929 cells. Double nuclei, prominent nucleoli, and clumpednuclear chromatin are evident. X 660; X 730.
Research. on February 18, 2018. © 1964 American Association for Cancercancerres.aacrjournals.org Downloaded from

w*nîw• •-V ' ••:•*»v . ^* -.',
* VA-V¿V, -IV;
-^ÄSl
r * \.- . > *V
$&&M609
Research. on February 18, 2018. © 1964 American Association for Cancercancerres.aacrjournals.org Downloaded from

FIG. 5.—Injection area of mouse liver nucleic acids-protein andNCTC 929 cells. The amorphous injected material is at lowerleft. Three cells with large bizarre nuclei are at the margin of thereaction zone of host fibroblasts and NCTC 929 cells and the amorphous material. X 679 (reduced).
FIG. 6.—Proliferating NCTC 929 cells at the margin of site ofinjection of NCTC 929 cells and 929 cell nucleic acids-protein.There is greater nuclear variability and pyknosis as compared withcontrols. X 271.
FIG. 7.—Control tissue culture of NCTC 929 strain L cells 5 daysfollowing transfer. X 228.
FIG. 8.—NCTC 929 cells, G days after treatment with 3 mg. ofascites nucleic acids that had been treated with RNase and DNase.X228.
FIG. 9.—NCTC 929 cells, 6 days after treatment with 2.5 mg. ofascites nucleic acids that had been treated with DNase. X 228.
FIG. 10.—NCTC 929 cells, 6 days after treatment with 3 mg. ofascites nucleic acids that had been treated with RNase. X 228.
FIG. 11.—NCTC 929 cells, Gdays after treatment with 3 mg. ofascites nucleic acids. X 228.
Research. on February 18, 2018. © 1964 American Association for Cancercancerres.aacrjournals.org Downloaded from

611
Research. on February 18, 2018. © 1964 American Association for Cancercancerres.aacrjournals.org Downloaded from

SMITH—Cytologieand Cytotoxic Changes Induced by Nucleic Acids 613
5. BEAVEN, G. H.; HOLIDAY,E. R.; ANDJOHNSON,E. A. In: E.Chargaff, and J. N. Davidson (eds.), Optical Properties ofNucleic Acids and Their Components in the Nucleic Acids,1:519. New York: Academic Press, Inc., 1955.
6. BENOIT, J.; LEROY, P.; VENDRELY, R.; AND VENDRELY, C.Modifications induites chez des canards pekin par le DNA decanard khaki campbell injectéeaprèsla naissance. Press. Med.,65:1623-24,1957.
7. BENSCH, K. G., AND KING, D. W. Incorporation of Heterol-ogous Deoxyribonucleic Acid into Mammalian Cells. Science,133:381-82, 1961.
8. BORENFREUND,E., ANDBENDICH,A. A Study of the Penetration of Mammalian Cells by Deoxyribonucleic Acids. J. Bi-ophys. Biochem. Cytol., 9:81-91,1961.
9. COOMBS,R. R. A. The Mixed Agglutination Reactions in theStudy of Normal and Malignant Cells. Cancer Res., 21:1198-1202, 1961.
10. DISCHB, Z. Color Reactions of Nucleic Acid Components. In:E Chargaff and J. N. Davidson, (eds.), The Nucleic Acids,1:285-305. New York: Academic Press, Inc.. 1955.
11. DUMONT, A. E.; AYVAZIAN,J. H.; AND MCÃœLUSKEY,R. T.Incorporation of Host Nuclear Materials into TransplantedTumor Cells in Surgical Wounds. Nature, 194:193-94, 1962.
12. EAGLE, H. Nutrition Needs of Mammalian Cells in TissueCulture. Science, 122:501-4, 1955.
13. EARLE, W. R.,; NETTLESHIP, A.; SCHILLING,E. L.; STARK,T.H.; STRAUS,N. R.; BROWN,M. F.; ANDSHELTON,E. Production of Malignancy in Vitro. V. Results of Injections of Cultures into Mice. J. Nati. Cancer Inst., 4:213-27, 1943.
14. EVANS, V. J., BRYANT,J. C.; McQuinciN, W. T.; FIORAMONTI,M. C.; SANFORD,K. K.; WESTFALL,B. B.; ANDEARLE, W. R.Studies of Nutrient Media for Tissue Cells in Vitro. II. AnImproved Protein-free Chemically Defined Medium for Long-Term Cultivation of Strain L-929 Cells. Cancer Res., 16:87-94,1956.
15. EVANS, V. J.; F'OHAMONTI, M. C.; RANDOLPH, L. K.; ANDEARLE, W. R. Studies of Nutrient Media for Tissue Cells inVitro. VI. The Effects of the Nucleic Acid Derivatives Mixtureof Medium NCTC 109 on NCTC Strain 2071, and NCTC Strain2981. Am. J. Hyg., 71:168-75, 1960.
16. FAHMY, O. G., ANDFAHMY,M. J. Induction of Mutations byDeoxyribonucleic Acid in Drosophila melanogaster. Nature,191:776-79, 1961.
17. FLOERSHEIM,G. L. Cytotoxic Action of Nucleic Acids. Expe-rientia, 18:328-29, 1962.
18. FOLEY, G. E., ANDHANDLER, A. H. Differentiation of "Normal" and Neoplastic Cells Maintained in Tissue Culture by
Implantation into Normal Hamsters. Proc. Soc. Exp. Biol.Med., 94:661-64, 1957.
19. GRIFFIN, A. The Effect of Malignancy upon Nucleic AcidMetabolism in Normal Tissues. Texas Rep. Biol. Med., 15:161-68, 1957.
20. HALL, R. H., ANDGALE, G. O. New Observations of Pneumo-coccal Transformations in Vivo. Proc. Soc. Exp. Biol. Med.,101:487-91, 1959.
21. HARRIS, M. Use of Dialyzed Media for Studies in Cell Nutrition. J. Cell Comp. Physiol., 40:279-301, 1952.
22. HEWER, T. F., ANDMEEK, E. S. Intestinal Carcinoma in MiceFollowing Injection of Herring-Sperm Deoxyribonucleic Acid.Nature, 181:990-91, 1958.
23. HILL, M., ANDJAKUBICKOVA,J. Intercellular Passage of DNAas Revealed in Bone Marrow Autoradiographs. Exp. Cell.Res.,26:541-51, 1962.
24. HOTCHKISS, R. D. Methods for Characterization of NucleicAcid. In: Methods in Enzymology, 3:708-15. New York:Academic Press, Inc., 1957.
25. HUGHES, A. The Effect of Purines and Related Substancesupon Cells in Chick Tissue Cultures. Exp. Cell. Res., 3:108-20, 1952.
26. KLENOW, H. Further Studies on the Effect of Deoxyadenosineon the Accumulation of Deoxyadenosine Triphosphate and
Inhibition of Deoxyribonucleic Acid Synthesis in EhrlichAscites Tumor Cells in Vitro. Biochim. Biophys. Acta, 61:885-96, 1962.
27. KUNITZ, M. Crystalline Desoxyribonuclease I. Isolation andGeneral Properties, Spectrophotometric Method for theMeasurement of Desoxyribonuclease Activity. J. Gen.Physiol., 33:349-77, 1950.
28. LACOUR,F.; LACOUH,J.; ANDHAREL, J. Transplantable Malignant Tumors in Mice Induced by Preparations ContainingRibonucleic Acid Extracted from Human and Mouse Tumors.J. Nati. Cancer Inst., 24:201-327, 1960.
29. LEUCHTENBERGER,C.; LEUCHTENBERGBH,R.; ANDUYEKI, E.Cytological and Cytochemical Changes in Liver of White MiceFollowing Intraperitoneal Injections of DNA Preparationsfrom Breast Cancers of Agouti C3H Mice. Proc. Nati. Acad.Sci., 44:700-705, 1958.
30. MAGASANIK,G., ANDCHAHGAFF,E. Studies on the Structure ofRibonucleic Acids. Biochim. Biophys. Acta, 7:396-112, 1951.
31. MALMGREN,R. A. Observation on a Liver Mitotic StimulantPresent in Tumor Tissue. Cancer Res., 16:232-36, 1956.
32. MCCARTY, M., AND AvERY, O. T. Studies on the ChemicalNature of the Substance Inducing Transformation of Pneumo-coccal Types III. An Improved Method for the Isolation of theTransforming Substance and Its Application to PneumococcusTypes II, III, VI. J. Exp. Med., 83:97-105, 1946.
33. MORGAN,J. F. Tissue Culture Nutrition. Bact. Rev., 22:20-45, 1958.
34. MORRIS, N. R.; REICHARD, P.; AND FISCHER, G. A. StudiesConcerning the Inhibition of Cellular Reproduction byDeoxyribonucleosides. Biochim. Biophys. Acta, 68:93-99,1963.
35. Niu, M. C.; CORDOVA,C. C.; ANoNlu, L. C. Ribonucleic Acid-induced Changes in Mammalian Cells. Proc. Nati. Acad. Sci.,47:1689-1700, 1961.
36. REREBEK, E., ANDREREBEK, J. Wirkung freier Nukleinbasenauf das Wachstum der Fibroelasten. Zellforsch, mikroscop.Anat., 36:503-21, 1952.
37. RIEKE, W. O. The in Vivo Reutilization of Lymphocytic andSarcoma DNA by Cells Growing in the Peritoneal Cavity. J.Cell Biol., 13:205-16, 1962.
38. ROTEM, Z.; Cox, R. A.; AND ISAACS,A. Inhibition of VirusMultiplication by Foreign Nucleic Acid. Nature, 197:564-66,1963.
39. SANFORD,K. K.; EARLE, R. R.; ANDLIKELY, G. D. The Growthin Vitro of Single Isolated Tissue Cells. J. Nati. Cancer Inst.,9:229-46, 1948.
40. SANFORD,K. K.; HOBBS, G. L.; ANDEARLE, W. R. The Tumor-producing Capacity of Strain L Mouse Cells after 10 Years inVitro. Cancer Res., 16:162-70, 1956.
41. SANFORD,K. K.; MERWIN, R. M.; HOBBS, G. L.; ANDYOUNG,J. M. Clonal Analysis of Variant Cell Lines Transformed toMalignant Cells in Tissue Culture. J. Nati. Cancer Inst., 23:1035-60, 1959.
42. SCHMIDT,G. Determination of Nucleic Acids by PhosphorusAnalysis. In: S. P. Colowick, and N. O. Kaplan (eds.),Methods in Enzymology, 3:671-79. New York: Academic Press,Inc., 1957.
43. SMITH, A. G., ANDCRESS, H. R. Effect of Tumor Deoxyribonucleic Acid upon Tissue Culture of Fibroblasts. Lab. Investigation, 10:898-907, 1961.
44. SZYBALSKA,E. H., ANDSzYBALSKi,W. Genetics of Human CellLines. IV. DNA-mediated Heritable Transformation of a Biochemical Trait. Proc. Nati. Acad. Sci., 48:2026-34, 1962.
45. SZYBALSKI,W.; SZYBALSKA,E. H.; AND RAGNI, G. GeneticStudies with Human Cell Lines, in Analytic Cell Culture. Nati.Cancer Inst. Monograph, No. 7, 1962.
46. TROWELL, O. A. The Action of Cortisone on Lymphocytes inVitro. J. Physiol., 119:274-85, 1953.
47. WAYMOUTH,C. The Nutrition of Animal Cells. Internat. Rev.Cytol., 3:1-68, 1954.
Research. on February 18, 2018. © 1964 American Association for Cancercancerres.aacrjournals.org Downloaded from

1964;24:603-613. Cancer Res Albert G. Smith Cytologic and Cytotoxic Changes Induced by Nucleic Acids
Updated version
http://cancerres.aacrjournals.org/content/24/4_Part_1/603
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/24/4_Part_1/603To request permission to re-use all or part of this article, use this link
Research. on February 18, 2018. © 1964 American Association for Cancercancerres.aacrjournals.org Downloaded from