-
V. Ranđelović, 2017 - FITOGEOGRAFIJA
9
Poglavlje 2
Areali biljnih taksona
Areal ili geografski opseg
Biljne vrste su na različite načine rasprostranjene na Zemlji.
One zauzimaju
manju ili veću površinu, pri čemu uvek postoji deo prostora na
kojem nisu
pronađene (Gaston, 2009). Geografska oblast koju zauzima vrsta
svojom
distribucijom naziva se areal ili geografski opseg (rang) vrste
(Rapoport,
1982). Areal je osnovna jedinica u biogeografiji (Brown et al.,
1996).
Površina koju zauzima određena biljna vrsta određena je
sveukupnošću
podataka o njenom rasprostranjenju, a
dobija se nanošenjem tačaka, koje
odgovaraju geografskim koordinatama svih
lokaliteta na kojima je ona pronađena, na
geografske karte. Spajanjem graničnih
tačaka ovako istačkane površine dobija se
oblast rasprostranjenja, odnosno areal te
vrste (Walter, 1979). Detaljno upoznavanje
suštine areala vrste, odnosno određivanje
njegovih granica, veličine, oblika i
strukture, preduslov je za poznavanje njene
biologije, ekologije i istorije (Janković,
1976). Do podataka o rasprostranjenju vrsta
dolazi se na osnovu florističkih spiskova
(lokalne i opšte flore, floristički radovi,
katalozi i druga literatura), herbarskih zbirki
i terenskih istraživanja, što predstavlja
ogroman posao velikog broja istraživača.
Razvojem kompjuterske tehnologije, pre
svega softvera za izradu baza podataka i
automatsko kartiranje, u mnogome je olakšano određivanje areala
vrsta.
Osim areala vrste, u biogeografiji se često analiziraju i
geografske oblasti
koje zauzimaju više taksonomske kategorije, tako da se može
govoriti o
arealima rodova, familija i redova, ali i niže taksonomske
kategorije, odnosno
areali podvrsta i varijeteta. Zbog toga se u fitogeografiji
često koristi termin
areal taksona.
Termin areal se ne primenjuje samo na taksone, već i na
sintaksone,
odnosno vegetacijske jedinice, jer i one imaju odgovarajuće
rasprostranjenje,
odnosno zauzimaju određenu površinu na Zemlji (Polunin,
1960).
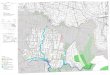
Slika 7. Areal vrste Ophrys helenae (Ranđelović
et al., 2015)
-
Areali biljnih taksona
10
Fitohorioni ili floristički horioni
Termin areal taksona ne bi trebalo poistovetiti sa terminom
floristički areal,
koji se takođe koristi u fitogeografiji. Floristički areal
predstavlja geografsku
oblast koja je naseljena karakterističnom, manje-više uniformnom
florom. U
literaturi se češće koriste sinonimi za ovaj termin, a to su
fitohorion ili
floristički horion (Takhtajan, 1986, Cox, 2001). Horion (grč.
χωρίον -
naselje) kao opšti biogeografski termin predložio je Turrill
(1958), a uporedo
sa njim i termine fitohorion i zoohorion.
Fitohorioni mogu biti različitog ranga (floristička carstva,
oblasti,
regioni, provincije), a u florističkom sistemu su klasifikovani
hijerarhijski
(Тахтаджян, 1978), o čemu će više reči biti u poglavlju
„Regionalna
fitogeografija“. Rang fitohoriona se označava terminom
fitogeografska
jedinica (Pedrotti, 2013).
Fitohorion je okarakterisan pre svega endemičnim biljnim vrstama
ili
višim taksonima, koji naseljavaju teritoriju koju on zauzima,
i
karakterističnim spektrom familija. Spektar familija
predstavlja
procentualno učešće pojedinih familija u izgradnji flore nekog
područja.
Horološko kartiranje
Horološke karte prikazuju areale vrsta i
drugih taksonomskih jedinica. Svrha
horološkog kartiranja je upoznavanje
prostorne distribucije taksona bez njihove
interakcije sa drugim vrstama. Horološke
karte su geografske karte na kojima je
prikazano rasprostranjenje taksona
različitim grafičkim tehnikama (Pedrotti,
2013).
Areal se može prikazati na različite
načine, ali u osnovi postoje dve metode –
tačkasta ili punktuirajuća i konturna metoda
(Sl. 8) (Janković, 1990).
Tačkasta ili punktuirajuća metoda
kartiranja areala podrazumeva prikazivanje
rasprostranjenja pomoću tačaka. Ovaj način
prikazivanja omogućava jasan pregled
gustine areala, koja se obično smanjuje od
centra prema periferiji. Nedostatak ove
metode je taj što se ne vide jasno granice
areala taksona.
Konturna metoda kartiranja areala
podrazumeva spajanje graničnih tačaka
rasprostranjenja zatvorenom krivom linijom
i senčenje tako dobijene površine.
Primenom ove metode ne postoji mogućnost
uočavanja gustine rasporeda taksona u
okviru areala.
Slika 8. Tačkasto (A), kombinacija tačkastog i
konturnog (B) i konturno (C) predstavljanje areala
roda Taxus (Taxaceae, Pinophyta)
-
Fitogeografija
11
Kombinacijom ovih metoda (kombinovana metoda) izbegavaju se
njihovi nedostaci.
Horološke karte mogu da prikažu deo ili
ceo areal vrste. Prema preciznosti i
metodama prikazivanja, Pedrotti (2013)
razlikuje sledeće tipove horoloških karti:
- Karta lokaliteta ili floristička karta - podaci o tačnim
mestima na kojima se
vrsta javlja prikazani su simbolima na
topografskim kartama (Sl. 9). Ove
karte se koriste prilikom sistematske
revizije vrsta i rodova, reviziju
prisustva vrste na datoj teritoriji ili za
prikazivanje novih lokaliteta, za izradu
crvenih knjiga i atlasa zaštićenih
biljnih vrsta, za kariološka i
citotaksonomska istraživanja i za
mnoge druge namene. One mogu da
prikazuju rasprostranjenje vrste na
celokupnom arealu ili na određenom
delu teritorije kao što je kontinent,
ostrvo, poluostrvo, država, region,
planina i slično.
- Mrežne karte - podaci o rasprostranjenju taksona se nanose
u
unapred definisana polja na mreži
kojom je određena teritorija podeljena
na jednake kvadrate. Uobičajeno se koristi UTM mreža (UTMG -
Universal Transverse Mercator Grid) (Sl. 10);
- Karte homogenih teritorija - horološki podaci se unose u polja
različite veličine koja ne grade mrežu, a definišu se na osnovu
njihovih
homogenih morfoloških i botaničkih karakteristika;
- Horoplet karte (grč. horo (χώρο) - prostor ili region, i
pletos (πλήθος) – mnoštvo) teritorija sa fiksnim internim granicama
(na primer, granice
administrativnih jedinica) koje prikazuju da li je takson
rasprostranjen
ili nije u okviru fiksnog podpodručja ili regiona;
- Apsolutne karte areala prikazuju areal zatvorenim krivim
linijama koje predstavljaju granice rasprostranjenja vrste (Sl.
8C);
- Kvantifikovane horološke karte se baziraju ne samo na
prisustvu/odsustvu ili nekom drugom kvalitativnom kriterijumu, već
i
na kvantitativnoj proceni jednog ili više karaktera, kao što su
horološki
elementi, životne forme ili ekološki pokazatelji.
Osim naučnog značaja, horološko kartiranje ima veliku primenu
u
očuvanju biodiverziteta. Znanje o tačnim lokalitetima vrsta je
od velikog
značaja za njihovu zaštitu i uspostavljanje zaštićenih područja.
Horološki
podaci su od suštinskog značaja za izradu "crvenih knjiga" i
procenu i
vrednovanje biljne raznovrsnosti određenih geografskih područja
(Pedrotti,
2013). Mnogobrojni drugi aspekti primene kartiranja areala
ukazuju na
njegovu izuzetnu važnost. U te aspekte primene spadaju
upravljanje
staništima i njihova restauracija, planiranje očuvanja staništa,
analiza
održivosti populacija, procena širenja invazivnih vrsta i
njihovih zajednica,
predviđanje efekata globalne promene životne sredine na vrste i
ekosisteme.
Slika 9. Areal vrste Xylorhiza orcuttii
Endemična vrsta pustinje Borrego Badlands u južnoj
Kaliforniji i njen areal prikazan na karti lokaliteta
(http://tchester.org/bd/species/asteraceae/xylorhiza_orcuttii.h
tml)
-
Areali biljnih taksona
12
Za predikciju promena životne sredine na osnovu podataka o
rasprostranjenju
vrsta u sadašnjosti i prošlosti, kao i prognoziranje promene
areala tih vrsta,
koriste se često statistički modeli koji omogućavaju rad sa
velikim brojem
podataka. Modeliranje rasprostranjenja vrsta (SDM, od engleskog
termina
species distribution modeling) je samo jedan od primera primene
modela koji
koriste lokalna merenja za praćenje i prognoziranje promena u
životnoj
sredini na regionalnom i globalnom nivou (Franklin, Miller,
2010).
Struktura, granice i forme areala
Areal vrste ne predstavlja površinu u okviru koje su lokalne
populacije
ravnomerno raspoređene u svim njegovim delovima i koju vrsta u
potpunosti
ispunjava. Raspored lokalnih populacija zavisi od mnogih
faktora, a pre svega
od istorijskog razvoja vrste i ekoloških uslova kojima se ona
prilagođava.
Janković (1990) smatra da je neravnomerna naseljenost vrste u
okviru areala
rezultat kombinovanog delovanja nehomogenosti teritorije areala,
pre
svega u pogledu variranja ekoloških uslova, i ekološke
ograničenosti date
vrste, koja nije adaptirana na sve tipove staništa u okviru
areala. Neke vrste
se pojavljuju češće u okviru areala, dok se neke pojavljuju
ređe, bez obzira
što im je areal približno isti. U tom slučaju govorimo o
različitoj gustini
areala vrste. Vrste koje se pojavljuju češće, naseljavaju veći
broj različitih
staništa i prilagođene su uslovima koji na njima vladaju. Takve
vrste obično
imaju široku ekološku valencu (eurivalentne vrste) za različite
ekološke
faktore. One vrste koje se ređe pojavljuju u okviru areala,
prilagođene su
uslovima koji vladaju na realativno malom broju različitih
staništa, često
samo jednom tipu staništa. Te vrste imaju usku ekološku
valencu
(stenovalentene vrste) makar za jedan ekološki faktor.
Osim toga, brojnost (abundanca) i gustina lokalnih populacija
za
određenu vrstu nije podjednaka u svim delovima areala. Najčešće
je gustina
Slika 11. Rasprostranjenje vrste Pinus nigra u
Evropi i Mediteranu prikazano na horiplet karti
(zelenom bojom prikazan prirodni deo areala)
Slika 10. Areal vrste Ramonda serbica,
endemične vrste Balkanskog poluostrva
prikazan na karti sa UTM mrežom
-
Fitogeografija
13
veća u središnjem delu areala, dok se prema periferiji sve više
smanjuje
(Brown, 1984). Ovaj koncept u biogeografiji se naziva hipoteza
centralnog
obilja (the abundant center hypothesis) (Sagarin, Gaines, 2002,
Sakulich,
2011).
Granice areala predstavljaju zatvorene krive linije povučene
između
krajnjih, graničnih tačaka (lokaliteta) do kojih je
rasprostranjena određena
vrsta. U osnovi, granice areala određuju ekološki uslovi kojima
se vrsta
prilagođava. Ipak, mnogi autori izdvajaju posebno mehaničke
uticaje, kao
što su planinski lanci i morska prostranstva. Takođe, značajan
uticaj na
formiranje i granice areala imaju antropogeni uticaji. Ekološki
uslovi koji
utiču na formiranje areala su najčešće
klimatski faktori. Vrsta zauzima onu
površinu čije joj karakteristike klime
(temperatura, vlažnost i druge) odgovaraju
i izvan te površine se ne širi (Алехин,
1944). Janković (1990) razlikuje stvarne i
iznuđene granice areala. Stvarne granice
areala su uslovljene nemogućnošću vrste
da van tih granica opstane usled
nepovoljnih ekoloških uslova, dok su
iznuđene granice areala uslovljene
prisustvom neke mehaničke prepreke, pri
čemu bi vrsta, da nije nje, mogla živeti i
izvan postojećih granica. Na primer,
kaktusi iz roda Opuntia, koji su prirodno
rasprostranjeni u pustinjama i na drugim
kserotermnim staništima američkih
kontinenata, sa najvećim brojem vrsta u
Meksiku (Anderson, 2001) veoma
uspešno rastu i na području Mediterana
(Sl. 12), ali i na drugim sličnim staništima
širom sveta, gde ih je preneo čovek nakon 15. veka. Nemogućnost
širenja
njihovog areala na evropski kontinent uslovljena je mehaničkim
uticajem,
odnosno velikim okeanskim prostranstvom.
Ukoliko su sve populacije u granicama jedne zatvorene linije,
areal je
kompaktan. Međutim, mnoge vrste imaju areale koji pokrivaju dve
ili više
značajno odvojenih površina. Takvi areali su disjunktni. Granice
areala
prikazane kao zatvorene krive linije ne oslikavaju pravu prirodu
areala vrste
i predstavljaju njegovu veštačku karakteristiku. Naime, biljke u
okviru areala
nisu ravnomerno raspoređene po celoj njegovoj površini, već
naseljavaju
odgovarajuća staništa, koja su odvojena jedna od drugih
(Стоянов, 1950). Ta
odvojenost je sve veća kako se ide ka granici areala, odnosno
gustina areala
se smanjuje prema njegovoj periferiji. Centar areala može da
bude relativno
kontinuirano naseljen, a prema periferiji se mogu javiti veće
ili manje
odvojene površine u obliku ostrva unutar granica areala (Brown
et al., 1996)
Ipak, dok god su rastojanja između pojedinih lokalnih populacija
dovoljno
mala da se na geografskoj karti uočavaju kao kontinuirani delovi
areala, one
se nalaze u granicama areala vrste. Periferijski delovi areala
se odlikuju
lokalnim populacijama koje su udaljene od kontinuiranog dela,
odnosno,
nalaze se van njegovih granica. To su periferijske disjunkcije
areala.
Međutim, kod određenih vrsta se pojavljuju i lokalne populacije
koje su
značajno udaljene od kontinuiranog dela areala, a površina koju
one
Slika 12. Opuntia sp. na stenovitoj obali
Mediteranskog mora u gradu Nici u Francuskoj
-
Areali biljnih taksona
14
zauzimaju naziva se eksklava areala (Walter, 1979). Zbog toga se
može
govoriti o strukturi granice areala vrste (Sl. 13) (Gorodkov,
1986, Gaston
2003). Gorodkov (1986) razlikuje
više graničnih linija u okviru
areala:
1) granicu zone kontinuirane distribucije - u čijim okvirima
su
klimatski i drugi uticaji povoljni
za nesmetano održavanje vrste;
2) granicu zone disjunktne distribucije - u čijim okvirima
je
održavanje vrste moguće samo na
onim delovima prostora gde su
klimatski i drugi uticaji istovetni
onima u granicama kontinuirane
distribucije, a razlikuju se od
okolnih;
3) granicu pulsiranja areala, odnosno zone povremenog
izumiranja - u čijim okvirima se
lokalne populacije vrste
pojavljuju povremeno usled
trenutnih povoljnih sezonskih
uticaja;
4) granicu zone sterilne invazije - u čijim okvirima se
javljaju
jedinke ili lokalne populacije koje
ne kompletiraju životni ciklus
usled nepovoljnih klimatskih
uticaja;
5) granicu zone pojavljivanja uginulih delova tela, odnosno
potpunog odsustva živih individua - u čijim okvirima je moguće
pronaći uginule
ostatke biljaka, koji su tu dospeli nošeni vodom, vetrom ili
životinjama.
Granice areala nisu stalne i one se vremenom menjaju, što zavisi
od
dinamike areala. Kod nekih vrsta one se menjaju sporo, gotovo
neprimetno,
u dužim vremenskim periodima, te se čini da su
nepromenljive.
Granicama areala određen je i njegov oblik. Teoretski, areal
u
dvodimenzionalnom prostoru bi imao oblik kruga, čiji centar je
centar
nastanka taksona. Međutim, u stvarnosti, oblik areala zavisi od
više faktora:
oblika kontinenata, klimatskih i mikroklimatskih uticaja,
razđenosti terena
(prisustvo planinskih venaca, rečnih dolina, klisura, kanjona i
drugih
elemenata reljefa), osobina zemljišta i stena, prethodnih
geoloških događaja i
prisustva drugih vrsta (interspecijska kompeticija) (Rapoport,
1982).
Nastanak i dinamika areala Razumevanje promena oblika i
veličine, kao i položaja areala vrsta
predstavlja ključno pitanje u ekologiji i evolucionoj biologiji
pojedinih
taksona (Gaston, 2009). Današnje granice areala razultat su
dugotrajnog
procesa formiranja areala. Sasvim je jasno da formiranju areala
vrste prethodi
Slika 13. Struktura granice areala vrste (prema Gorodkov,
1986, Gaston, 2003)
-
Fitogeografija
15
njen nastanak na nekom delu Planete, odnosno proces specijacije.
Deo
prostora koji naseljava novonastala vrsta naziva se primarni
areal
(Толмачов, 1974, Груев, Кузманов, 1994), odnosno centar
nastanka
taksona (Cain, 1944). Odatle se vrsta širi zahvaljujući nekom od
načina
rasprostiranja i rasejavanja, odnosno disperzije. Disperzija
obično nastupa
kada gustina populacije premaši mogućnosti životne sredine,
odnosno kada
više nema dovoljno resursa za povećavanje brojnosti populacije.
Povećavanje
areala vrste zavisi od njene mogućnosti prilagođavanja uslovima
sredine,
odnosno njenih ekološko-bioloških osobina, ali i od ekoloških
pogodnosti
koje im sredina pruža (Janković, 1990). Areal vrste se povećava
sve dok joj
ekološki uslovi sredine, a pre svega klimatski faktori,
odgovaraju, ili dok ne
naiđe na mehaničku barijeru koju ne može da savlada (mora i
okeani,
planinski lanci). Kako je proces formiranja
areala izuzetno dugotrajan, nakon dužeg
vremenskog perioda, vrsta ili rod može
potpuno da nestane sa mesta nastanka, a
svojim arealom da zahvati neka druga
prostranstva. Drugim rečima, areal može
da promeni položaj. Takav je slučaj sa
nekim pleistocenskim glacijalnim
reliktima rasprostranjenim u planinskim
područjima Evroazije i Severne Amerike,
čiji je primarni areal (ili areal neposredne
predačke vrste) bio na severu Holarktika.
Ova pojava je u skladu pre svega sa
klimatskim promenama koje su se
odigravale od momenta nastanka vrste do
danas. Areali mnogih alpskih endemičnih
biljaka su nastali nakon glacijacije u
pleistocenu. Ove biljke su naselile Alpe povlačeći se ispred
ledene kalote, da
bi se nakon Ledenog doba zadržale na najvišim planinskim
vrhovima (Sl. 14).
Povećavanje areala može da bude i naglo, ali do takvih promena
dolazi
uglavnom pod uticajem antropogenog faktora. Naime, pod uticajem
čoveka,
areali mnogih vrsta su se u kratkom vremenskom periodu proširili
uprkos
mehaničkim barijerama.
Sa druge strane, kod nekih vrsta granice se menjaju naglo, u
kraćim
vremenskim periodima i bez uticaja
čoveka. Nagle promene areala
uglavnom dovode do njegovog
smanjivanja. Do tih naglih promena
dolazi usled smanjivanja vitalnosti
populacija ili postepenih slabljenja
mogućnosti za održavanje u ranijim
granicama (Gaston, 2009).
Poslednja faza u dinamici areala
vrste je nestanak areala, odnosno
izumiranje vrste. Nestanku areala
vodi niz faktora kao što su nagle
klimatske promene, prirodne
katastrofe (na primer, erupcija
vulkana), konkurencija drugih vrsta,
a sve češće i antropogeni uticaji.
Slika 14. Papaver alpinum, endemit Alpa
glacijalnog porekla
Slika 15. Campanula calycialata
Endemit Stare planine u Srbiji; nestala je
ubrzo nakon što je opisana kao nova za
nauku (Ranđelović, Zlatković 1998)