Embed Size (px)
Citation preview

UNIVERZITET U NIŠU
PRIRODNO-MATEMATIČKI FAKULTET
DEPARTMAN ZA BIOLOGIJU I EKOLOGIJU
Hemotaksonomska karakterizacija podsekcija Pinus i
Pinaster (Pinus, Pinaceae) na osnovu terpenskih markera
MASTER RAD
Kandidat: Mentor:
Jelena Stevanović 152 Dr Zorica S. Mitić
Niš, 2016.

UNIVERSITY OF NIŠ
FACULTY OF SCIENCE AND MATHEMATICS
DEPARTMENT OF BIOLOGY AND ECOLOGY
Chemotaxonomic characterisation of subsections Pinus
and Pinaster (Pinus, Pinaceae) based on terpene markers
MASTER THESIS
Candidate: Mentor:
Jelena Stevanović 152 Dr. Zorica S. Mitić
Niš, 2016.

BIOGRAFIJA KANDIDATA
Jelena Stevanović, roĎena je 16. decembra 1991. godine u Nišu. Završava osnovnu
školu ”Jovan Jovanović Zmaj” u Malči i Prehrambeno-hemijsku školu u Nišu. Prirodno-
matematički fakultet u Nišu upisuje 2011. godine na Departmanu za biologiju i ekologiju.
Osnovne akademske studije završava 2014. godine i iste godine upisuje Master akademske
studije, smer biologija, koje u predviĎenom roku završava.

ZAHVALNICA
Najsrdačnije se zahvaljujem svom mentoru dr Zorici Mitić sa Departamana za
biologiju i ekologiju, Prirodno-matematičkog fakulteta u Nišu na izboru teme, neizmernoj
saradnji, razumevanju, pomoći i usmeravanju tokom celokupne izrade ovog master rada.
Veliku zahvalnost dugujem i prof. dr Bojanu Zlatkoviću sa Departamana za biologiju
i ekologiju, Prirodno-matematičkog fakulteta u Nišu kao i prof. dr Petru Marinu sa Katedre
za morfologiju i sistematiku biljaka, Biološkog fakulteta u Beogradu na dragocenoj pomoći u
sakupljanju biljnog materijala.
Posebnu zahvalnost dugijem prof. dr Gordani Stojanović, Snežani Jovanović i Ivani
Zrnzević sa Departmana za hemiju, Prirodno-matematičkog fakulteta u Nišu, na pomoći u
realizaciji hemijskih analiza i interpretaciji dobijenih rezultata.
Najveću zahvalnost na bezgraničnoj podršci i pomoći bez koje ne bih završila studije
dugujem svojim roditeljima i sestrama Karolini i Mileni.

SAŽETAK
Hemijski sastav etarskog ulja četina 24 taksona sekcije Pinus, uključujući 17 taksona
podsekcije Pinus i 7 taksona podsekcije Pinaster, uporeĎen je u cilju utvrĎivanja
hemotaksonomskog značaja terpena na mezosistematskom (infrageneričnom nivou). Sastav
etarskog ulja P. nigra ssp. pallasiana sa područja Srbije, Makedonije i Grčke analiziran je u
ovom radu, a osnovni kriterijumi za izbor literaturnih podataka bili su način izolacije etarskog
ulja (hidrodestilacija) i vrsta organa (četine). Na ovaj način, u analizu je uključeno 57 uzoraka
etarskog ulja dobijenih metodom hidrodestilacije iz četina 24 taksona sect. Pinus. Prema
rezultatima analize varijanse (ANOVA) moţe se videti da 15, od ukupno 34 ispitivana
terpenska karaktera, poseduje visok nivo statističke značajnosti, što ukazuje na to da postoji
statistički značajna razlika izmeĎu ispitivanih podsekcija. Najveći nivo statističke značajnosti
(P<0.001) pokazuju γ-kadinen i kamfen, a zatim slede α-pinen, δ-kadinen, germakren D, α-
kadinol, gvajol i (E)-β-ocimen (P<0.01). Najmanji nivo statističke značajnosti (P<0.05)
poseduju feniletil-3-metil butanoat, mircen, biciklogermakren, γ-terpinen, kembren, α-
terpineol i bornil acetat. Klaster analiza (CA) ukazala je na izdvajanje 5 grupa taksona, pri
čemu prve 3 grupe čine predstavnici subsect. Pinaster (sa izuzetkom P. merkusii), a druge
dve članovi subsect. Pinus (sa izuzetkom P. canariensis). Ovakvi rezultati se u velikoj meri
poklapaju sa dosadašnjim morfološkim, biohemijskim i molekularnim istraţivanjima koji
jasno ukazuju na postojanje dve sestrinske klade u okviru sekcije Pinus uključujući "prave"
mediteranske borove sa jedne (subsect. Pinaster) i evroazijske borove sa druge strane
(subsect. Pinus). TakoĎe, rezultati ovog rada potvrĎuju značaj terpena kao
hemotaksonomskih markera na infrageneričnom nivou.
Ključne reči: subsect. Pinus, subsect. Pinaster, terpeni, hemotaksonomija

ABSTRACT
The chemical composition of needle essential oils of 24 taxa from the section Pinus,
including 17 and 7 taxa of subsections Pinus and Pinaster, respectively, were compared in
order to determine chemotaxonomic significance of terpenes on mesosystematic
(infrageneric) level. The composition of essential oils in P. nigra ssp. pallasiana from Serbia,
Macedonia and Greece was analyzed in this paper, while the basic criteria for selection of
literature data included the method of essential oil isolation (hydrodistillation) and the type of
conifer organs (needles). In this way, 57 samples of essential oil obtained by hydrodistillation
from needles of 24 taxa of sect. Pinus were analyzed. According to the results of analysis of
variance (ANOVA), out of 34 examined terpene characters 15 had a high level of statistical
significance, indicating statistically significant difference between the examined subsections.
The highest level of statistical significance was shown by γ-cadinene and camphene
(P<0.001), followed by α-pinene, δ-cadinene, germacrene D, α-cadinol, guaiol and (E)-β-
ocimene (P<0.01). The lowest levels of statistical significance (P<0.05) were recorded for
phenyl ethyl-3-methyl butanoate, myrcene, bicyclogermacrene, γ-terpinene, cembrene, α-
terpineol, and bornyl acetate. Cluster analysis (CA) has shown separation of five groups of
taxa, where first three groups included representatives of subsect. Pinaster (with the
exception of P. merkusii), and the other two members of subsect. Pinus (with the exception of
P. canariensis). These results are largely consistent with previous morphological,
biochemical and molecular studies that clearly indicated presence of two clades in the section
Pinus, including "proper" Mediterranean pines on one side (subsect. Pinaster) and Eurasian
pines on the other side (subsect. Pinus). In addition, the results of this study confirm the
significance of terpenes as chemotaxonomic markers on infrageneric level.
Keywords: subsect. Pinus, subsect. Pinaster, terpenes, chemotaxonomy

SADRŽAJ
1. UVOD ............................................................................................................................................. 1
1.1. Opšte karakteristike roda Pinus L. .......................................................................................... 1
1.2. Opšte karakteristike Pinus sect. Pinus .................................................................................... 2
1.3. Opšte karakteristike vrste Pinus nigra Arnold ........................................................................ 3
1.4. Opšte karakteristike etarskih ulja ............................................................................................ 6
1.4.1. Fizičke osobine etarskih ulja ........................................................................................... 8
1.4.2. Hemijski sastav etarskih ulja ........................................................................................... 9
1.4.3. Taksonomski značaj terpena ......................................................................................... 10
1.4.4. Upotreba i biološke aktivnosti etarskog ulja borova ..................................................... 12
2. CILJEVI RADA ............................................................................................................................ 13
3. MATERIJAL I METODE ............................................................................................................ 14
3.1. Biljni material ............................................................................................................................ 14
3.2. Izolovanje etarskog ulja ............................................................................................................. 14
3.3. GC-FID i GC-MS analiza etarskog ulja ..................................................................................... 16
3.4. Literaturni podaci ....................................................................................................................... 16
3.5. Statistička analiza ....................................................................................................................... 19
4. REZULTATI I DISKUSIJA ......................................................................................................... 21
4.1. Hemijski sastav etarskog ulja četina P. nigra ssp. pallasiana ................................................... 21
4.2. Hemijski sastav etarskog ulja četina podsekcije Pinus .............................................................. 28
4.3. Hemijski sastav etarskog ulja četina podsekcije Pinaster .......................................................... 31
4.4. Analiza varijanse (ANOVA) ...................................................................................................... 35
4.5. Klaster analiza (CA) .................................................................................................................. 38
5. ZAKLJUČCI ................................................................................................................................. 41
6. LITERATURA ............................................................................................................................. 43

Jelena Stevanović Master rad
1
1. UVOD
"Within the wealth of data available for Pinus, it is perhaps surprising that key
phylogenetic relationships remain unresolved" (Syring et al., 2005).
1.1. Opšte karakteristike roda Pinus L.
Rod Pinus, sa oko 110 opisanih vrsta, predstavlja najveći recentni rod četinara.
Prirodna distribucija roda ograničena je na severnu hemisferu, izuzev jedne populacije P.
merkusii koja se nalazi juţno od ekvatora na Sumatri (Mirov, 1967, prema Šarac, 2014).
Borovi su obilno zastupljeni u Severnoj i Centralnoj Americi, Evropi i Aziji, a nekoliko
taksona proteţe se i do severne Afrike. Javljaju se na raznovrsnim staništima, od nivoa mora
do 3700 m nadmorske visine, a vrlo često su i edifikatori različitih zajednica. Po pravilu,
borovi odsustvuju u toplim i vlaţnim regionima tropskog pojasa, gde su slabi kompetitori sa
ostalim taksonima (Mirov, 1967, prema Šarac, 2014). U okviru tropskog pojasa javljaju se
isključivo u planinskim ili semi-aridnim područjima.
Najšire prihvaćenu klasifikaciju roda predloţili su Little i Critchfield (1969), a u
kasnijim revizijama izmenjen je broj vrsta i modifikovana taksonomska organizacija na nivou
sekcija i podsekcija (Farjon i Styles, 1997; Price et al., 1998; Gernandt et al., 2003; 2005,
prema Šarac, 2014). Naime, rod Pinus se deli deli na dva podroda: Pinus (= Diploxylon ili
trvdi borovi) i Strobus (= Haploxylon ili meki borovi). Subgenus Pinus odlikuju dva sudovna
snopića u centru lista i trajni rukavac koji obavija četine u čuperku (osim kod vrsta P.
leiophylla i P. lumholtzii). Plodne ljuspe su deblje i rigidnije u poreĎenju sa plodnim
ljuspama Strobus. Ostale karakterisitke podroda, koje navodi Farjon (1984), su veoma
polimorfne ili imaju malu dijagnostičku vrednost: broj iglica u čuperku varira od 2-8, poloţaj
smolnih kanala polimorfan (unutrašnji, spoljašnji, medijalni ili septalni), krilce spojeno ili
sraslo sa semenom, zrele šišarke se otvaraju odmah ili nakon odreĎenog vremena, subgenus
je rasprostranjen širom severne hemisfere i na različitim tipovima staništa. Sa druge strane,
subgenus Strobus odlikuje jedan sudovni snopić u centru lista i rukavci četina koji brzo
opadaju (osim kod vrste P. nelsonii). Plodne ljuspe su tanje i fleksibilnije, broj iglica u

Jelena Stevanović Master rad
2
čuperku varira od 1-5 (retko 6), smolni kanali su medijalni ili spoljašnji, krilca semena veoma
polimorfna (odsutno, rudimentirano, spojeno ili sraslo), zrele šišarke se obično odmah
otvaraju. Strobus je, takoĎe, rasprostranjen širom severne hemisfere, na različitim tipovima
staništa, osim u subtropskim nizijama. Dalja podela na sekcije i podsekcije se razlikuje
izmeĎu pojedinih autora. PoreĎenjem morfologije, anatomije, citologije, sekundarnih
metabolita, proteina i DNK, Price et al. (1998) izdvajaju 111 vrsta u 2 podroda, 4 sekcije i 17
podsekcija, dok su Gernandt et al. (2005) na osnovu analize hloroplastne DNK (matK i rbcL)
smanjili broj podsekcija na 11 (tabela 1.1) (prema Šarac, 2014).
Tabela 1.1. Klasifikacija roda Pinus prema Gernandt et al. (2005).
Podrod Pinus (Diploxylon) Sekcija Pinus, Podsekcija Pinus: P. densata, densiflora, hwangshanensis, kesiya, luchuensis,
massoniana, merkusii, mugo*, nigra*, resinosa, sylvestris*, tabuliformis, taiwanensis, thunbergii,
tropicalis, uncinata, yunnanensis; Podsekcija Pinaster: P. brutia, canariensis, halepensis, heldreichii*,
pinaster, pinea, roxburghii.
Sekcija Trifoliae, Podsekcija Contortae: P. banksiana, clausa, contorta, virginiana;
Podsekcija Australes: P. attenuata, caribaea, cubensis, echinata, elliottii, glabra, greggii, herrerae,
jaliscana, lawsonii, leiophylla, lumholtzii, muricata, occidentalis, oocarpa, palustris, patula,
praetermissa, pringlei, pungens, radiata, rigida, serotina, taeda, tecunumanii, teocote; Podsekcija
Ponderosae: P. cooperi, coulteri, donnell-smithii, devoniana, douglasiana, durangensis, engelmannii,
hartwegii, jeffreyi, maximinoi, montezumae, nubicola, ponderosa, pseudostrobus, sabineana, torreyana,
washoensis.
Podrod Strobus (Haploxylon) Sekcija Parrya, Podsekcija Balfourianae: P. aristata, balfouriana, longaeva; Podsekcija
Cembroides: P. cembroides, culminicola, discolor, edulis, johannis, maximartinezii, monophylla,
pinceana, quadrifolia, remota, rzedowskii; Podsekcija Nelsoniae: P. nelsonii
Sekcija Quinquefoliae, Podsekcija Gerardianae: P. bungeana, gerardiana, squamata;
Podsekcija Krempfianae: P. krempfii; Podsekcija Strobus: P. albicaulis, armandii, ayacahuite,
bhutanica, cembra, chiapensis, dabeshanensis, dalatensis, fenzeliana, flexilis, koraiensis, lambertiana,
monticola, morrisonicola, parviflora, peuce*, pumila, sibirica, strobus, wallichiana, wangii.
*Autohtone vrste u flori Srbije (Jovanović, 1992).
1.2. Opšte karakteristike Pinus sect. Pinus
Sekciju Pinus odlikuje trajni rukavac, broj iglica u čuperku od dve do tri, obično
spoljašnji ili medijalni poloţaj smolnih kanala, ali ponekad i unutrašnji (P. merkusii) ili
septalni (P. tropicalis). Zrele šišarke se otvaraju nakon odreĎenog vremena (osim kod P.
pinea) i imaju debele plodne ljuspe sa dorzalnim poloţajem štita (umbo). Krilce je labavo
vezano za seme kod većine vrsta, osim kod P. canariensis i P. roxburghii gde je krilce čvrsto
vezano za seme. Sekcija je rasprostranjena širom Evroazije i Mediterana sa samo dve
američke vrste (P. resinosa iz istočne Severne Amerike i P. tropicalis sa zapadne Kube).

Jelena Stevanović Master rad
3
Na osnovu varijabilnosti hloroplastnih DNA sekvenci (matK i rbcL), Gernandt et al.
(2005) ukazali su na postojanje dve klade u okviru sect. Pinus (tabela 1.1):
subect. Pinus koja uključuje evroazijske borove i dve severnoameričke vrste
(P. resinosa i P. tropicalis).
subsect. Pinaster koja obuhvata "prave" mediteranske borove zajedno sa
himalajskom vrstom P. roxburghii.
Sa druge strane, analiza matriksa kvalitativnih morfoloških karaktera nije otkrila
nijedan diskriminišući karakter izmeĎu podsekcija Pinus i Pinaster (Gernandt et al., 2005).
1.3. Opšte karakteristike vrste Pinus nigra Arnold
Pinus nigra Arnold (crni ili austrijski bor) je visoko, srednje ili nisko drvo do 30, reĎe
40 (50) m, u zavisnosti od staništa, sa širokokupastom do amrelastom krošnjom (slike 1.1 i
1.2). Kora na odraslom stablu debela, tamnosiva; rano ispuca nepravilnim, kasnije dubokim
uzduţnim i poprečnim brazdama. Kora se u boji i debljini malo razlikuje u donjem i gornjem
delu (razlika od P. sylvestris). Mlade grančice gole, sjajne, pokrivene sitnim rombičastim
ljuspama. Pupoljci valjkasti pri vrhu, oštro zašiljeni, smolasti. Četine po dve, tamnozelene,
krupne, duge 8-18 cm, široke, 1.5-2.1 mm, pravo pruţene ili blago uvijene, traju obično 4-5
godina, pri bazi su obuhvaćene zajedničkim rukavcem. Smoni kanali u četinama medijalno
postavljeni; rukavac trajan, dug 10-12 mm, obavija po dve četine (Vidaković, 1982;
Jovanović, 1992).
Crni bor je jednodoma vrsta sa jednopolnim strobilusima. Muški i ţenski strobilusi se
pojavljuju od aprila do maja meseca. Muški strobilusi (mikrostrobilusi) podsećaju na rese,
zbijeni su pri osnovi mladih grančica, ţutozeleni, gotovo sedeći i koţasti. Ţenski strobilusi
(makro-, megastrobilusi) su 5-7.5 cm dugi, i 2-3.5 (4) cm široki, skoro sedeći, smešteni pri
vrhu mladih izdanaka. Razvijen ţenski strobilus (šišarka, conus) je izgraĎen od kompaktno
rasporeĎenih, odrvenelih plodnih ljuspi koje su, sa donje strane, crnosmeĎe, poprečno
ispucale, a na vrhu poseduju štitoliki izraštaj, apofizu. Zreli semeni zameci su izduţeno
jajasti, 4-6 mm dugi, sivi, sa izduţenim, krilastim, poluprozirnim dodatkom 3-4 puta duţim
od samog semenog zametka. Sazrevanje šišarke i semenih zametaka traje skoro dve sezone
nakon oploĎenja (Zlatković i Šarac, 2016).

Jelena Stevanović Master rad
4
Slika 1.1. P. nigra ssp. pallasiana: A) Olimp, Grčka (foto: Rajčević, 2015);
B) Stumica, Makedonija (foto: Zlatković, 2013)
A
B

Jelena Stevanović Master rad
5
Crni bor je jedan od najšire rasprostranjenih i najpolimorfnijih četinara Evrope.
Diskontinuirano je rasprostranjen oko severnog Mediterana, od istočne Španije i juţne
Francuske, preko Korzike i Kalabrije,
Austrije i Balkanskog poluostrva do Male
Azije i Krima. U Flori Evrope Gaussen et
al. (1993) razlikuju pet alopatričnih
podvrsta crnog bora (slika1.2): ssp. nigra
(od Austrije do centralne Italije i Grčke),
ssp. dalmatica (Vis.) Franco (priobalni
region i ostrva Dalmacije - Hrvatska), ssp.
pallasiana (Lamb.) Holmboe (Balkansko
poluostrvo, juţni Karpati i Krim), ssp.
salzmannii (Dun.) Franco (Ceven i Pirineji
- Francuska, centralna i istočna Španija), i
ssp. laricio (Poir.) Maire (Kalabrija i
Sicilija - Italija i Korzika - Francuska).
TakoĎe, afričke populacije P. nigra
opisane su kao posebna podvrsta: ssp.
mauretanica (Maire i Peyerimh) Heywood
(Greuter et al., 1984).
Palasov ili krimski crni bor (ssp.
pallasiana) (slike 1.1 i 1.2) se od tipične
podvrste odvaja, pre svega, duţim lisnim
rukavcima (18-26 mm) (Jovanović, 1992).
Ovogodišnje grančice su ţute do
svetlosmeĎe, sjajne. Ljuspe na grančicama
svetloţute, po duţini jednake rukavcima
četina ili duţe. Ljuspe su tri puta duţe od
svoje širine, sa paralelnim ivicama; na
leĎima imaju usko, istaknuto rebro i
zašiljen, povijen vrh. Četine prave do malo
savijene, krute, duge 8-12 (-18) cm i 1.6-Slika 1.2. P. nigra ssp. pallasiana na planini Crnook
(Jarešnik), jugoistočna Srbija (foto: Zlatković, 2013)

Jelena Stevanović Master rad
6
2.1 mm široke, tamnozelene. Hipodermalne ćelije sa 2-5 slojeva. Smonih kanala u parenhimu
obično manje od 11. Šišarice tamnosmeĎe i sjajne, duge 5-10 cm, 2-4 cm široke sa tupim
rebrom na zrakasto naboranim apofizama gornjih i srednjih ljuspi; grbice crvenkaste do sive.
Na teritoriji Srbije krimski crni bor zabeleţen je jedino u jugoistočnoj Srbiji
(Rajevski, 1950) na planini Crnook (Jarešnik) u okolini Bosilegrada, na nadmorskoj visini
oko 1300 m, na kristalastim škriljcima (slika 1.2). Takson je zakonom zaštićen kao prirodna
retkost na području Srbije (Sl. gl. R. Srbije br. 66/91, 83/92), dok je stanište na Crnooku
proglašeno za strogi rezervat prirode.
1.4. Opšte karakteristike etarskih ulja
Etarska ulja su mirisne smese isparljivih, lipofilnih sastojaka (sekundarnih biljnih
metabolita), koje se sintetišu i lokalizuju u specijalnim sekretornim strukturama biljaka i
mogu predestilovati vodenom parom (Petrović et al., 2009).
Etarska ulja su uglavnom lako pokretljive, mirisne tečnosti; pojedina su na sobnoj
temperaturi viskozna ili čvrste konzistencije. Glavni sastojci etarskih ulja su monoterpeni,
seskviterpeni i u manjoj meri derivati fenilpropana. To su različiti ugljovodonici (zasićeni ili
nezasićeni), alkoholi, fenoli, estri, aldehidi, ketoni, epoksidi ili karboksilne kiseline. Etarska
ulja su smese oko 20-200 sastojaka. Ovi sastojci su, na osnovu koncentracije u etarskom ulju,
podeljeni na: glavne (20-95%), sporedne (1-20%) i zastupljene u tragovima (ispod 1%).
Etarska ulja najčešće sadrţe svega nekoliko glavnih komponenti (npr. karvakrol (30%) i
timol (27%) u etarskom ulju Origanum compactum, a mentol (59%) i menton (19%) u ulju
Mentha x piperita), a nekada je to samo jedan sastojak (npr. 1,8-cineol (50%) u etarskom ulju
Cinnamomum camphora ili linalol (68%) u ulju Coriandrum sativum).
Etarska ulja su uglavnom produkti kopnenih biljaka (Embryophyta). Ove biljke su
rasporeĎene u preko pedeset familija, a najpoznatije su aromatične biljke familija Pinaceae,
Cupressaceae, Lauraceae, Zingiberaceae, Myrtaceae, Rutaceae, Lamiaceae, Apiaceae i
Asteraceae. Etarska ulja se mogu naći u svim delovima biljke (korenu, rizomu, stablu, kori,
semenu, listovima, cvetovima i plodovima), ravnomerno rasporeĎena u svim biljnim
organima ili samo u odreĎenom biljnom organu - najčešće i u najvećem procentu su
zastupljena u listu i cvetu. Interesantno je da etarska ulja lokalizovana u različitim delovima
iste biljke, mogu biti sličnog sastava, ali mogu se i značajno razlikovati (etarsko ulje zelenog

Jelena Stevanović Master rad
7
i zrelog ploda, listova i korena peršuna razlikuju se po dominantnim sastojcima) (Kovačević,
2004). Prinosi etarskih ulja se obično kreću izmeĎu 0.2 i 2.0%, mada postoje znatna
odstupanja. U zavisnosti od sadrţaja etarskog ulja koji moţe da varira (prinos izuzetno
cenjenog ruţinog ulja često ne prelazi 0.03%, dok prinos nekih drugih ulja moţe dostizati i
do 20%, npr. etarsko ulje karanfilića) biljke su podeljene u red bogatih ili siromašnih uljem
(Gašić, 1985).
Etarska ulja se najčešće nalaze u slobodnom stanju i to egzogeno: u ţlezdama,
ţlezdanim ljuspama i ţlezdanim dlakama, i endogeno: u sekretornim ćelijama,
intercelularnim šizogenim šupljinama i intercelularnim šizogenim i lizogenim rezervoarima.
Postoje izuzeci kada su ulja vezana u obliku heterozida (seme crne slačice, seme gorkog
badema). U vezanom obliku ulja su bez mirisa, a pod uticajem specifičnih enzima i u
prisustvu vode, nastaju isparljivi mirisni sastojci (aglikoni).
Biosinteza etarskih ulja se odvija u sekretornim ćelijama sa gustom citoplazmom i
većim brojem plastida, mitohondrija i diktiozoma (Simić, 2000). U posebnim delovima
biljaka dolazi do akumulacije ulja, najčešće u neposrednoj blizini ili na samim mestima gde i
nastaju. Postoji više vrsta sekretornih struktura, u zavisnosti od samih biljaka, i to:
1. ulje se sakuplja na mestu nastanka u ćeliji - modifikovane parenhimske ćelije
(idioblasti). Ovo je slučaj kod biljaka familija Lauraceae, Piperaceae, Zingiberaceae, Araceae
itd.;
2. meĎućelijski sekretorni prostori - okrugle šupljine ili cilindrični kanalići koji se
prema postanku dele na šizogene (Myrtaceae), lizigene (u plodovima Citrus voća) i
šizolizigene (kod Rutaceae); kod predstavnika familije Pinaceae unutar mezofila listova često
se nalaze smolni kanali šizogenog porekla, okruţeni parenhimskim ţlezdanim ćelijama koje
produkuju smolu i etarska ulja (slika 1.3.)
3. ţlezde i ţlezdane dlake (peltatne i kapitatne dlake) koje se nalaze na površini
zeljastih delova biljaka (Asteraceae i Lamiaceae).

Jelena Stevanović Master rad
8
Biološka funkcija etarskih ulja povezana je sa atraktantnim delovanjem na insekte,
čime je obezbeĎeno je prenošenje polena i oprašivanje biljaka. Sa druge strane, veoma je
značajna i uloga etarskih ulja u zaštiti biljaka od napada insekata i ţivotinja, herbivora.
TakoĎe, gotovo sva ulja više ili manje intenzivno sprečavaju razvoj mikroorganizama, i tako
suzbijaju širenje infekcija na biljkama u kojima nastaju. Interesantni su i podaci o specifičnim
efektima ulja jedne biljke, koje prelazi u podlogu i inhibira klijanje semena drugih biljaka
(alelopatija). Neka etarska ulja sprečavaju klijanje sopstvenih semena i obezbuĎuju ţivotni
prostor svake individue. Stara je teorija, koja samo delimično moţe biti prihvaćena, da biljke
sintezom i odavanjem ovih isparljivih produkata stvaraju specifičnu mikroklimu, koja ih štiti
od prekomerne transpiracije u različitim nepovoljnim uslovima (Kovačević, 2004).
1.4.1. Fizičke osobine etarskih ulja
Etarska ulja su uglavnom bezbojne, ţute ili ţutozelene tečnosti; pojedina su specifično
obojena (npr. etarsko ulje cimeta i karanfilića smeĎecrveno). Droge sa proazulenima daju
destilacijom vodenom parom plavo ili zeleno obojena etarska ulja (npr. plava etarska ulja
daju Matricariae flos, Absinthii herba i Millefolii herba). Proazuleni su neisparljivi, bezbojni,
gorki seskviterpenski laktoni. Tokom destilacije vodenom parom prelaze u obojene isparljive
azulene. Miris etarskih ulja potiče od sastojaka sa kiseoničnim funkcionalnim grupama. On je
intenzivan čak i kada su ovi sastojci zastupljeni u niskom procentu (etarsko ulje limuna). U
koncentrovanom obliku etarska ulja uglavnom imaju oštar, nadraţujući ukus, koji pali.
Slika 1.3. Poprečni presek kroz list (četinu) P. nigra
smolni kanali

Jelena Stevanović Master rad
9
Razblaţivanjem ukus uglavnom postaje prijatan. Izuzeci su npr. etarsko ulje anisa i morača
koja su slatkog ukusa (Petrović et al., 2009).
Etarska ulja su lipofilna, rastvaraju se u nepolarnim organskim rastvaračima, etanolu,
mastima i masnim uljima. Generalno, etarska ulja se ne rastvaraju u vodi. U stvari, samo mali
broj sastojaka ulja je rastvorljiv; ovakvi proizvodi nazivaju se aromatične vode.
Kao tečnosti, etarska ulja su definisana nekim fizičkim konstantama. Relativna
gustina ulja moţe biti manja ili veća od 1; ipak neuporedivo je mani broj etarskih ulja čija je
relativna gustina veća od 1. Kao parametar kvaliteta, značajne su vrednosti indeksa refrakcije
i ugla skretanja ravni polarizovane svetlosti. Kao specifična karakteristika definisana je i
rastvorljivost etarskog ulja u etanolu odreĎenog razblaţenja.
Specifičnost ove grupe biljnih proizvoda je da nemaju tačno definisanu temperaturu
ključanja. Pošto su to smeše različitih jedinjenja, svako od njih isparava na svojoj temperaturi
ključanja. Tako, etarska ulja frakciono k - C (Kovačević,
2004).
1.4.2. Hemijski sastav etarskih ulja
Etarska ulja su sloţene mešavine dve različite klase jedinjenja: terpena i
fenilpropanskih derivata. U okviru terpenske frakcije, zastupljeni su isparljivi mono- i
seskviterpeni.
Monoterpeni se javljaju u obliku acikličnih (linearnih), mono- i bicikličnih, alifatičnih
i aromatičnih struktura. Na osnovu uvedenih funkcionalnih grupa, sastojci ulja mogu biti
uljovodonici, alkoholi, aldehidi, ketoni, kiseline, estri, fenoli, etri, oksidi, peroksidi, epoksidi.
U okviru frakcije seskviterpena, formirane su još raznovrsnije strukture osnovnog
skeleta; zbog duţine lanaca C atoma veća je mogućnost različitih ciklizacija. I ova grupa
terpena javlja se u obliku ugljovodonika, oksida, alkohola, ketona i estara.
U etarskim uljima su aromatični, fenilpropanski sastojci (C6-C3) zastupljeni u manjim
količinama. Postoje i izuzeci, kada glavninu ulja čine upravo ovakva jedinjenja. Najčešće su
to aril- ili propenilfenoli (eugenol, anetol) i aldehidi (cimetni aldehid). Ovakva jedinjenja su
uglavnom prisutna u etarskom ulju biljaka iz familija Apiaceae i Myrtaceae. Kao sastojci
etarskih ulja sreću se i C6-C1 jedinjenja, kao što je vanilin i derivati antranilne kiseline.

Jelena Stevanović Master rad
10
Tokom destilacije, iz biljne sirovine mogu predestilisati i različita alifatična jedinjenja
manje molekulske mase (zasićeni i nezasićeni ugljovodonici, alkoholi, aldehidi, estri, laktoni)
koji se rastvaraju u etarskom ulju i postaju njegov sastavni deo. TakoĎe, kod nekih droga koje
su izloţene fermentaciji ili torefakciji, na primer, male količine prisutnog etarskog ulja
postaju obogaćene sumpornim i azotnim jedinjenjima koja nastaju tokom tih specifičnih
procesa; time se formira specifična aroma ovakvih droga (seme kafe, kakaovca) (Kovačević,
2004).
1.4.3. Taksonomski značaj terpena
Upotreba terpena u sistematici biljaka ima veoma dugu tradiciju, a zasniva se na
činjenici da je sastav terpena pod genetičkom kontrolom i u velikoj meri nezavisan od uticaja
faktora spoljašnje sredine (Incerti et al., 2013). Kod mnogih vrsta sastav terpena je povezan
sa poreklom biljnog materijala, tako da terpeni mogu biti korišćeni kao markeri u istraţivanju
genetičkog variranja na geografskom ili ekološkom nivou (Nikolić et al., 2007; 2008; 2009)
kao i u verifikaciji materijala nepoznatog porekla (Cenard et al., 1997). Hanover (1992) je
istakao da su terpeni odlični markeri za istraţivanje biodiverziteta, geografske varijabilnosti,
evolucije i sistematike, naročito kod četinara. Do danas je objavljen veliki broj radova o
terpenima kod četinara i ostalih vaskularnih biljaka, a meĎu njima ima najviše onih koji se
bave proučavanjem sastava terpena i njihovim geografskim varijabilitetom. TakoĎe, dosta je
uraĎeno u oblasti taksonomije, biosinteze terpena i ispitivanja uticaja terpena na otpornost
biljaka prema bolestima, štetočinama i spoljašnjim faktorima (Šarac, 2014).
I pored neosporne činjenice da su sastav i produkcija terpena pod genetičkom
kontrolom, kod terpena (kao i kod drugih sekundarnih metabolita biljaka) uočava se odreĎena
varijabilnost, kao posledica uticaja različitih ekoloških faktora (temperatura, vlaţnost i
aeracija zemljišta, mineralne soli u podlozi, nadmorska visina, duţina izloţenosti svetlu,
kvalitet svetlosti, vlaţnost vazduha i vazdušna strujanja). Zbog toga je pre analize
taksonomskog značaja neophodno istraţiti intra- i interpopulacionu varijabilnost terpena
odreĎenog taksona. TakoĎe, pravilno uzorkovanje i doslednost u postupcima branja,
skladištenja i izolacije etarskih ulja su preduslovi validne interpretacije dobijenih rezultata.
Niži nivoi klasifikacije. Iako je hemotaksonomski značaj terpena kod četinara
potvrĎen na nivou rodova (Picea i Pinus, Nikloić et al., 2011) i sekcija (Juniperus i Sabina,

Jelena Stevanović Master rad
11
Adams, 1991; Tsuga i Hesperopeuce, Lagalante i Montgomery, 2003), u najvećem broju
radova terpeni su uspešno korišćeni (prema Šarac, 2014):
1. kao hemotaksonomski markeri na nivou vrsta:
• Pinus halepensis, P. brutia, P. nigra, P. pinea i P. canariensis (Roussis et al.,
1995),
• Pinus attenuata, P. heldreichii, P. peuce, P. pinaster i P. radiata, (Petrakis et al.,
2001),
• P. pinea, P. halepensis, P. pinaster i P. nigra (Macchioni et al., 2003), itd.
2. u identifikaciji infraspecijskih taksona i proceni geografske varijabilnosti vrsta iz
rodova:
• Pinus (P. nigra, Arbez et al., 1974; Kubeczka i Schultze, 1987; Gerber et al.,
1995; Bojović, 1995; Chalchat i Gorunovic, 1995a; 1995b; Vidrich et al., 1996; Rafii et al.,
1996; Rezzi et al., 2001; Bojovic et al., 2005; Sezik et al., 2010; P. sylvestris, Manninen et
al., 2002; Naydenov et al., 2002; 2005b; Semiz et al., 2007; P. heldreichii, Nikolić et al.,
2007; P. peuce, Nikolić et al., 2008; P. contorta var. latifolia, Pauly i von Rudloff, 1971; P.
pinea, De Simón et al., 2001; P. pinaster, Arrabal et al., 2005; itd.),
• Picea (P. glauca, von Rudloff, 1972; P. abies, Persson et al., 1996; P. omorica,
Nikolić et al., 2009; itd.),
• Abies (A. lasiocarpa, Zavarin et al., 1970; A. alba, Zeneli et al., 2001; A.
nephrolepsis, Li et al., 2005; itd.),
• Pseudotsuga (P. taxifolia var. menziesii i var. glauca, von Rudloff, 1973)
• Thuja (T. plicata, von Rudloff et al., 1988),
• Juniperus (J. squamata var. fargesii, Adams et al., 1996; J. communis, Mastelić
et al., 2000; J oxycedrus, Medini et al., 2010; J. deltoides, Rajčević et al., 2013; itd.)
2. u proučavanju i detekciji hibrida: Picea glauca x P. engelmannii (Schaefer i
Hanover, 1986), Pinus brutia x P. halepensis (Gallis i Panetsos, 1977), Pinus nigra x P.
sylvestris i P. densiflora x P. thunbergiana (Idţojtić et al., 2005) itd.
3. u proučavanju prelaznih oblika: izmeĎu Pinus discolor i P. johannis (Zavarin i
Snajberk, 1986), Pinus discolor i P. cembroides (Bailey et al., 1982).
Viši nivoi klasifikacije. Obimni literaturni podaci ukazuju da je korišćenje terpena u
sistematici biljaka, pre svega korisno na niţim nivoima klasifikacije (najviše do nivoa roda).

Jelena Stevanović Master rad
12
Njihov značaj na suprageneričnim nivoima se smanjuje povećavanjem taksonomnskog nivoa,
što je očekivano s obzirom na njihovu funkciju u biljci, njihovu biosintezu i na strukturne
karakteristike (Marin, 2003).
Jedan od interesantnih primera značaja terpena na višim nivoima klasifikacije se
odnosi na familiju Lamiaceae. Naime, po nekim starijim klasifikacijama, Lamiaceae se dele
na dve podfamilije: Lamioideae i Nepetoideae. Nepetoideae su, generalno, bogate etarskim
uljima, za razliku od Lamioideae. Sa druge strane, kod Lamioideae se javljaju iridoidni
glukozidi koji su, po pravilu, odsutni kod predstavnika Nepetoideae. Postoji još nekoliko
hemijskih parametara koji diferenciraju ove dve velike grupe unutar familje Lamiaceae.
1.4.4. Upotreba i biološke aktivnosti etarskog ulja borova
Prema Dervendzi (1992), etarsko ulje borova (Pini aetheroleum) najviše se koristi u
narodnoj medicini za lečenje respiratornih infekcija praćenih kašljem, bronhitisom,
bronhijalnom astom, efizemom, traheitisom, sinuzitisom, laringitisom, faringitisom i
influencom. Pored toga, etarska ulja borova široko se koriste kao aditivi u prehrambenoj i
industriji alkoholnih pića. U mnogim proizvodima namenjenim za čišćenje, ulja borova
poznata su kao osveţavajuća i dezinfekciona sredstva. TakoĎe, ova ulja intenzivno se koriste
u farmaceutskoj, kozmetičkoj i parfimerijskoj industriji.
Na osnovu pregleda literaturnih podataka, moţemo reći da etarsko ulje borova čitav
niz bioloških aktivnosti:
• antibakterijsku i antifungalnu (Toroglu, 2007; Kolayli et al., 2009; Politeo et
al., 2011; Karapandzova et al., 2011; Šarac et al., 2014);
• akaricidnu (Macchioni et al., 2002);
• larvicidnu i repelentnu aktivnost protiv Aedes albopictus (Koutsaviti et al.,
2015);
• antiagregantnu (antitrombocitnu) (Tognolini et al., 2006).
• antioksidativnu (Ustun et al., 2012);
• herbicidnu aktivnost protiv Phalaris canariensis, Trifolium campestre i
Sinapis arvesis (Amri et al., 2014).

Jelena Stevanović Master rad
13
2. CILJEVI RADA
Osnovni naučni ciljevi ovog istraţivanja mogu se predstaviti u nekoliko teza:
1. Upoznavanje varijabilnosti sastava i sadrţaja etarskog ulja četina u prirodnim
populacijama P. nigra ssp. pallasiana sa područja Srbije, Makedonije i Grčke.
2. PoreĎenje hemijskog sastava etarskog ulja četina dve sestrinske podsekcije (Pinus
i Pinaster) u cilju utvrĎivanja hemotaksonomskog značaja terpena na
mezosistematskom (infrageneričnom) nivou.
3. Definisanje potencijalnih hemotipova istraţivanih podsekcija.
4. PoreĎenje dobijenih rezultata sa morfološkim, biohemijskim i molekularnim
karakterima koji su do sada korišćeni u taksonomiji i sistematici sect. Pinus.

Jelena Stevanović Master rad
14
3. MATERIJAL I METODE
3.1. Biljni material
Odabirom populacija P. nigra ssp. pallasiana, sprovedenim tokom 2013. i 2015.
godine, obuhvaćene su 4 populacije sa teritorije centralnog i juţnog Balkana: 1. Jarešnik
(Dukat, Srbija), 2. Mreţičko-Roţden (Koţuf, Makedonija), 3. Tri Vodi (Strumica,
Makedonija), 4. Kalamaras (Olimp, Grčka). Geografski poloţaj populacija je odreĎen uz
pomoć GPS ureĎaja (GPS Garmin eTrex Vista® C). Orografske karakteristike staništa pre
svega nadmorska visina i ekspozicija su preuzeti iz SRTM (Shuttle Radar Topography
Mission) (Reuter et al., 2007) digitalnog elevacionog modela rezolucije 90 x 90 m (Farr et al.,
2007) pomoću softvera ArcGIS 10 (ESRI, 2011) (tabela 3.1). Herbarski primerci deponovani
su u Herbarijumu Departama za biologiju i ekologiju, Prirodno-matematičkog fakulteta u
Nišu (HMN).
U svakoj od odabranih populacija crnog bora izvršeno je sakupljanje grančica sa
dvogodišnjim četinama sa većeg broja odraslih stabala. U ručnom friţideru ili u specijalnim
kesama sa ledom uzorci su sa mesta sakupljanja prenošeni do zamrzivača (- 20°C) na
skladištenje do momenta upotrebe.
3.2. Izolovanje etarskog ulja
Hidrodestilacija. Iz 100 g sveţih i iseckanih četina P. nigra ssp. pallasiana
(uklonjenih sa grančica 20 različitih individua iz svake ispitivane populacije) destilovano je
etarsko ulje u aparatu tipa Clavenger u toku 2.5 h. Ulje je ekstrahovano etrom, a etarski
ekstrakt sušen anhidrovanim Mg2SO4 (European Pharmacopoeia, 1983). Posle odvajanja
sredstva za sušenje filtracijom, etar je uklonjen destilacijom na rotacionom-vakuum
uparivaču bez zagrevanja, a etarsko ulje čuvano u zatvorenim hromatografskim vijalama na
4°C do analize. Prinos etarskog ulja je izraţen preko zapreminskih procenata (%) tj. v/w,
obračunato u odnosu na 100 g biljnog materijala. 40 µL svakog ulja razblaţeno je etrom do
1mL za dalje hromatografske analize.
Jelena Stevanović Master rad
15
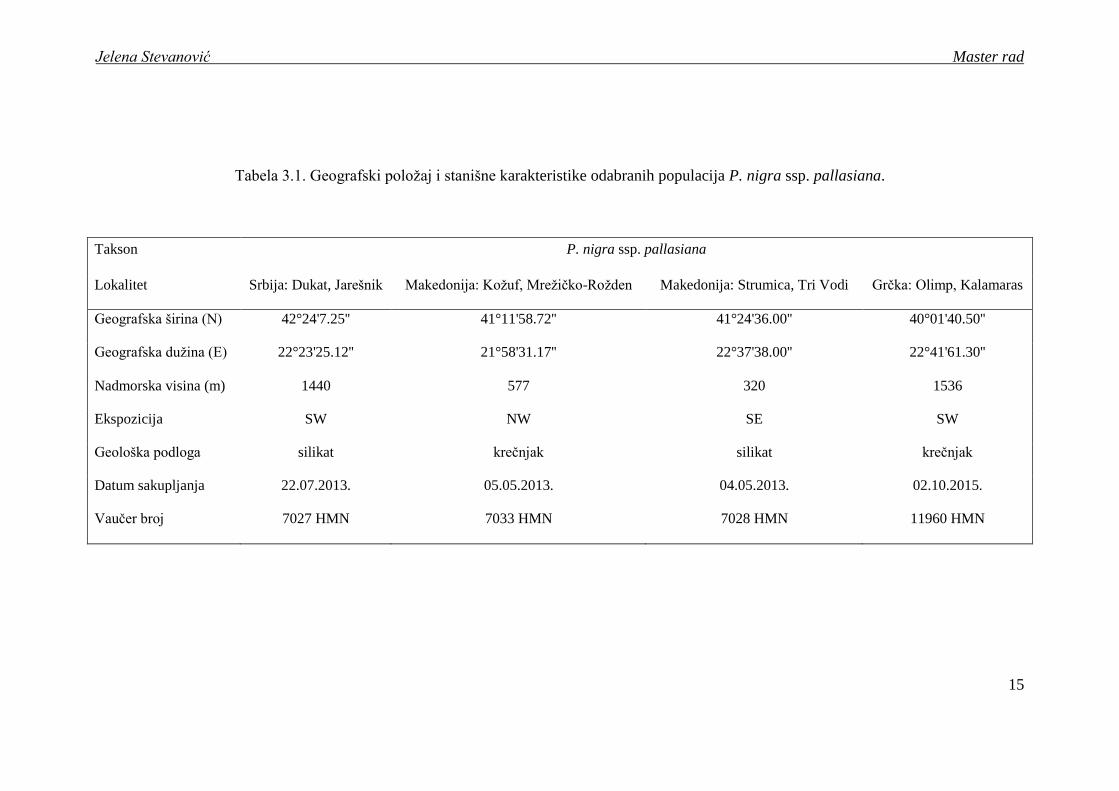
Tabela 3.1. Geografski poloţaj i stanišne karakteristike odabranih populacija P. nigra ssp. pallasiana.
Takson P. nigra ssp. pallasiana
Lokalitet Srbija: Dukat, Jarešnik Makedonija: Koţuf, Mreţičko-Roţden Makedonija: Strumica, Tri Vodi Grčka: Olimp, Kalamaras
Geografska širina (N) 42°24'7.25'' 41°11'58.72'' 41°24'36.00'' 40°01'40.50''
Geografska duţina (E) 22°23'25.12'' 21°58'31.17'' 22°37'38.00'' 22°41'61.30''
Nadmorska visina (m) 1440 577 320 1536
Ekspozicija SW NW SE SW
Geološka podloga silikat krečnjak silikat krečnjak
Datum sakupljanja 22.07.2013. 05.05.2013. 04.05.2013. 02.10.2015.
Vaučer broj 7027 HMN 7033 HMN 7028 HMN 11960 HMN

Jelena Stevanović Master rad
16
3.3. GC-FID i GC-MS analiza etarskog ulja
Svaki uzorak etarskog ulja rastvoren u etru je sniman u triplikatu na 7890/7000B
gasnom hromatografu (Agilent Technologies, USA) sa Combi PAL auto-semplerom i HP-5
MS kolonom (5% fenil metil siloksan, 30 m x 0.25 mm i.d. i 0.25 µm debljina filma)
kuplovan sa plamenim jonizacionim detektorom 7890A (FID) i masenim detektorom 5977A
(MS). Temperaturni program: izotermalno prvih 2.25 minuta 50 °C, zatim linearni porast
temperature (gradijent 4 °C/min) od 50 do 290 °C. Temperatura injektora 250 °C, interfejsa
300 °C; protok nosećeg gasa – helijuma 1.0 mL/min. Injektovana zapremina ulja 1µL, split
40:1; injektovana zapremina HS isparljivih 500 μL, split 10:1. Bekfleš 1.89 min, na 280 °C,
sa helijumom pod pritiskom od 50 psi. MS uslovi (EI): jonizacija -70 eV, opseg masa 35 –
435 amu (3 skena u sekundi), vreme skenova 0.2 sekunde. GC-FID analiza je izvedena pod
istim eksperimentalnim uslovima kao što je već navedeno za GC-MS analizu. Procentni
sastav je preračunat na osnovu površine GC-FID pikova sa hromatograma snimljenih pod
istim uslovima kao GC-MS, bez korekcionih faktora.
Identifikacija komponenti. Komponente etarskih ulja identifikovane su pomoću
AMDIS softvera (Automated Mass Spectral Deconvolution and Identification System, Ver.
2.1, DTRA/NIST, 2011), na osnovu lineranih retencionih indeksa (RI), koji su odreĎeni na
osnovu retencionih vremena standardne serije n-alkana (C8-C40), snimljene na istom aparatu i
pod istim uslovima kao i uzorak ulja. RI su uporeĎivani sa literaturnim vrednostima (Adams,
2007 i NIST Chemistry WebBook).
3.4. Literaturni podaci
Hemijski sastav etarskog ulja četina 24 taksona sect. Pinus, uključujući 17 taksona
subsect. Pinus i 7 taksona subsect. Pinaster, uporeĎeno je u cilju utvrĎivanja
hemotaksonomskog značaja terpena na mezosistematskom (infrageneričnom nivou). Sastav
etarskog ulja P. nigra ssp. pallasiana sa područja Srbije, Makedonije i Grčke analiziran je u
ovom radu, a osnovni kriterijumi za izbor literaturnih podataka bili su način izolacije etarskog
ulja (hidrodestilacija) i vrsta organa (četine). Na ovaj način u analizu je uključeno 57 uzoraka
etarskog ulja, dobijenih medotom hidrodestilacije iz četina 24 taksona sect. Pinus (tabela
3.2).

Jelena Stevanović Master rad
17
Tabela 3.2. Pregled istraţivanih taksona subsect. Pinus i subsect. Pinaster.
R. br.
uzorka
Takson1) Referenca
Podsekcija Pinus
1 P. densiflora Ioannou et al. (2014)
2 P. massoniana Ioannou et al. (2014)
3 P. merkusii Kurose et al. (2007)
4 P. mugo Ioannou et al. (2014)
5 P. mugo var. prostrata Ioannou et al. (2014)
6 P. mugo var. pumilio Ioannou et al. (2014)
7 P. nigra ssp. nigra_1 Krstić (2016)
8 P. nigra ssp. nigra_2 Krstić (2016)
9 P. nigra ssp. nigra_3 Krstić (2016)
10 P. nigra ssp. nigra_4 Krstić (2016)
11 P. nigra ssp. nigra_5 Krstić (2016)
12 P. nigra ssp. nigra_6 Ioannou et al. (2014)
13 P. nigra ssp. pallasiana_1 ova studija
14 P. nigra ssp. pallasiana_2 ova studija
15 P. nigra ssp. pallasiana_3 ova studija
16 P. nigra ssp. pallasiana_4 ova studija
17 P. nigra ssp. caramanica Ioannou et al. (2014)
18 P. nigra ssp. dalmatica Politeo et al. (2011)
19 P. nigra ssp. laricio_1 Rezzi et al. (2001)
20 P. nigra ssp. laricio_2 Rezzi et al. (2001)
21 P. nigra ssp. laricio_3 Ioannou et al. (2014)
22 P. nigra ssp. laricio_4 Amri et al. (2014)
23 P. nigra ssp. salzmannii Ioannou et al. (2014)
24 P. nigra_1 Koutsaviti et al. (2015)
25 P. nigra_2 Koutsaviti et al. (2015)
26 P. nigra_3 Macchioni et al. (2003)
27 P. nigra_4 Ustun et al. (2012)
28 P. sylvestris_1 Ustun et al. (2012)
29 P. sylvestris_2 Ioannou et al. (2014)
30 P. sylvestris ssp. scotica Ioannou et al. (2014)
31 P. tabuliformis Ioannou et al. (2014)

Jelena Stevanović Master rad
18
32 P. taiwanensis Ioannou et al. (2014)
33 P. thunbergii Ioannou et al. (2014)
Podsekcija Pinaster
34 P. brutia_1 Koutsaviti et al. (2015)
35 P. brutia_2 Ustun et al. (2012)
36 P. brutia_3 Ioannou et al. (2014)
37 P. brutia_4 Lahlou (2003)
38 P. canariensis_1 Koutsaviti et al. (2015)
39 P. canariensis_2 Ioannou et al. (2014)
40 P. halepensis_1 Koutsaviti et al. (2015)
41 P. halepensis_2 Macchioni et al. (2003)
42 P. halepensis_3 Fekih et al. (2014)
43 P. halepensis_4 Ustun et al. (2012)
44 P. halepensis_5 Ioannou et al. (2014)
45 P. halepensis_6 Lahlou (2003)
46 P. halepensis_7 Dob et al. (2005)
47 P. heldreichii_1 Petrakis et al. (2001)
48 P. heldreichii_2 Ioannou et al. (2014)
49 P. pinaster_1 Koutsaviti et al. (2015)
50 P. pinaster_2 Macchioni et al. (2003)
51 P. pinaster_3 Ioannou et al. (2014)
52 P. pinaster_4 Lahlou (2003)
53 P. pinea_1 Macchioni et al. (2003)
54 P. pinea_2 Ustun et al. (2012)
55 P. pinea_3 Ioannou et al. (2014)
56 P. roxburghii_1 Ioannou et al. (2014)
57 P. roxburghii_2 Satyal et al. (2013)
1) Lista istraţivanih taksona prema klasifikaciji roda Pinus od strane
Gernandt et al. (2005). Taksoni za koje postoji veći broj literaturnih
podataka o terpenskom sastavu četina su numerisani.

Jelena Stevanović Master rad
19
3.5. Statistička analiza
Deskriptivna statistika. Za komponente koje su detektovane u koncentraciji većoj od
0.5% u etarskom ulju istraţivanih populacija P. nigra ssp. pallasiana, kao i za komponente
pristune u koncentraciji većoj od 2% u minimalno jednom od 57 analiziranih uzoraka sect.
Pinus, uraĎena je osnovna deskriptivna statistika (srednja vrednost, maksimum, minimum,
standardna devijacija i koeficijent varijacije) na osnovu koje je uraĎena analiza varijabilnosti
pojedinačnih hemijskih karaktera. Ovi rezultati su grafički prikazani uz pomoć whisker box
plotova gde je za svaki karakter prikazana njegova hemijska varijabilnost na nivou ispitivane
podvrste, podsekcija i sekcije. Koeficijent varijacije (CV%) je korišćen za opisivanje
hemijske varijabilnosti karaktera i u odnosu na njegove vrednosti karakteri su okarakterisani
kao nisko varijabilni (CV% < 10), umereno varijabilni (10 < CV% < 50), visoko varijabilni
hemijski karakteri (50 < CV% < 100), oni karakteri kod kojih je CV% >100 su okarakterisani
kao jako visoko varijabilni.
Jednofaktorska analiza varijanse (ANOVA) je uraĎena da bi se utvrdilo da li
postoji statistički značajna razlika u prosečnim vrednostima hemijskih karaktera izmeĎu
analiziranih podsekcija (subsect. Pinus i subsect. Pinaster). Osnovni postupak na kome se
zasniva ova tehnika jeste izvoĎenje dve različite ocene varijanse podsekcija iz podataka, za
koje se zatim računa količnik. Jedna od ovih ocena (varijansa izmeĎu grupa) predstavlja meru
efekata nezavisne promenljive udruţene sa varijansom greške. Druga ocena (varijansa unutar
grupa) je mera samo varijanse greške. Količnik varijansi izmeĎu i unutar grupa označava se
kao F-vrednost ili F-količnik. Statistički značajnan F-količnik ukazuje da su prosečne
vrednosti posmatranog karaktera za date podsekcije veoma različite. Za ovu analizu
izdvojene su 34 komponente etarskog ulja koje su detektovane u koncentraciji većoj od 2% u
minimalno jednom od istraţivanih uzoraka.
Klaster analiza. Klaster analiza omogućava grupisanje analiziranih varijabli u grupe
ili klastere tako se da slični uzorci nalaze u istoj klasi (klasteru). U ovoj analizi je korišćen
algomerativni hijerarhijski metod (UPGMA) koji kao krajnji rezultata produkuje fenogram
koji u suštini, moţe da se shvati kao grafički prikaz sličnosti uporeĎivanih grupa. Kako bi se
izračunala distanca analiziranih jedinica posmatranja, korišćena je Euklidova distanca. Ova
analiza je korišćena da bi se utvrdila sličnost izmeĎu prvobitno neklasifikovanih objekata tj.
taksona subsect. Pinus i subsect. Pinaster u ovom slučaju.

Jelena Stevanović Master rad
20
Statistička obrada podataka je raĎena u statističkom programu Statistica, version 5.1
(StatSoft Inc., 1996, Tulsa, OK, USA).

Jelena Stevanović Master rad
21
4. REZULTATI I DISKUSIJA
4.1. Hemijski sastav etarskog ulja četina P. nigra ssp. pallasiana
Biljni materijal P. nigra ssp. pallasiana prikupljen je sa četiri lokaliteta na području
Srbije, Makedonije i Grčke tokom 2013. i 2015. godine (tabela 3.1). Iz sveţih četina,
postupkom hidrodestilacije u aparatu po Clevenger-u izolovana su etarska ulja čiji se prinos
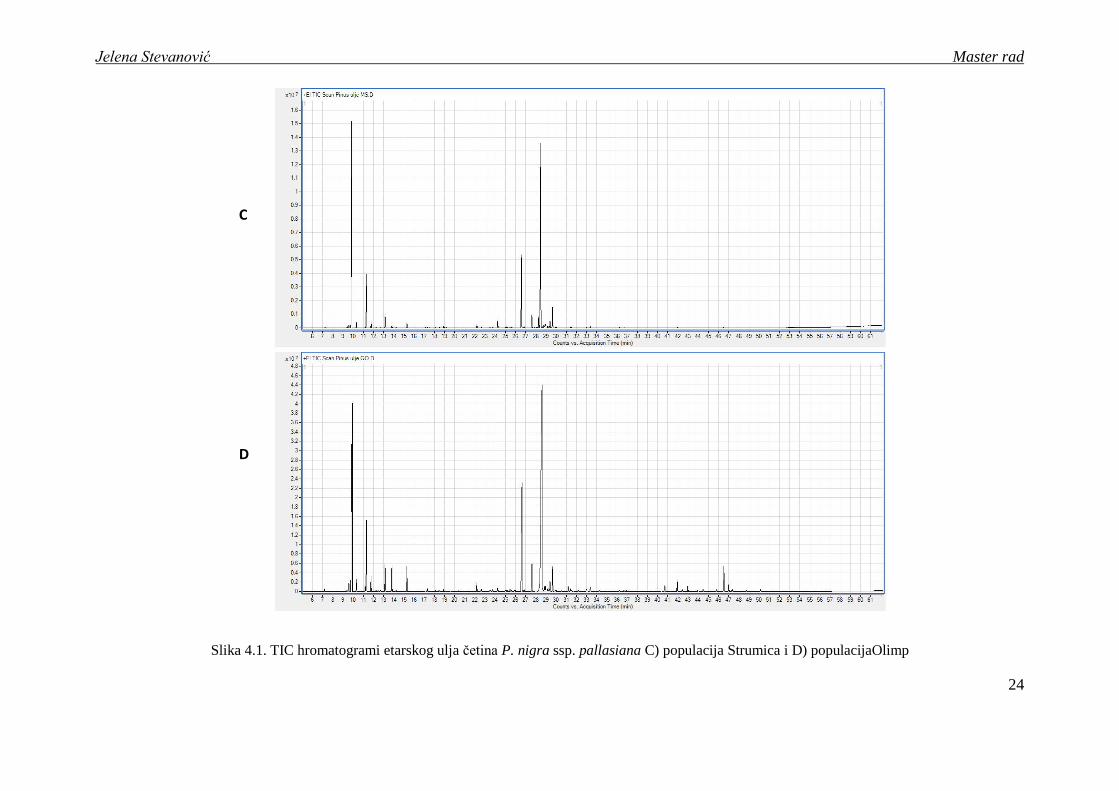
nalazi u opsegu 0.16 - 0.29 (v/w) (tabela 4.1). Kombinacijom GC i GC/MS analize u
etarskom ulju istraţivanih populacija detektovano je 114 jedinjenja, od kojih je 96
identifikovano. Kvlatitativni i kvantitativni sastav ulja je dat u tabeli 4.1, pri čemu su
prikazana samo jedinjanja prisutna u koncentraciji većoj od 0.5%, minimalno u jednoj od
ispitivanih populacija. Rezultati deskriptivne statistike za izabrane hemijske karaktere na
nivou podvrste predstavljeni su u tabeli 4.2 i na slici 4.2. Dobijeni hromatogrami za sve
ispitivane populacije su prikazani na slici 4.1.
U etarskom ulju četina ispitivanih populacija P. nigra ssp. pallasiana utvrĎeno je
prisustvo 16 komponenti u koncentraciji većoj od 0.5%: 8 monoterpenskih ugljovodonika, 1
oksidovani monoterpen, 6 seskviterpenskih ugljovodonika i 1 oksidovani diterpen. Prosečno
ove komponete čine 90.7% etarskog ulja, pri čemu su seskviterpeni nešto više zastupljeni
(48.6%) u odnosu na monoterpenske komponente (41.8%) (tabela 4.2).
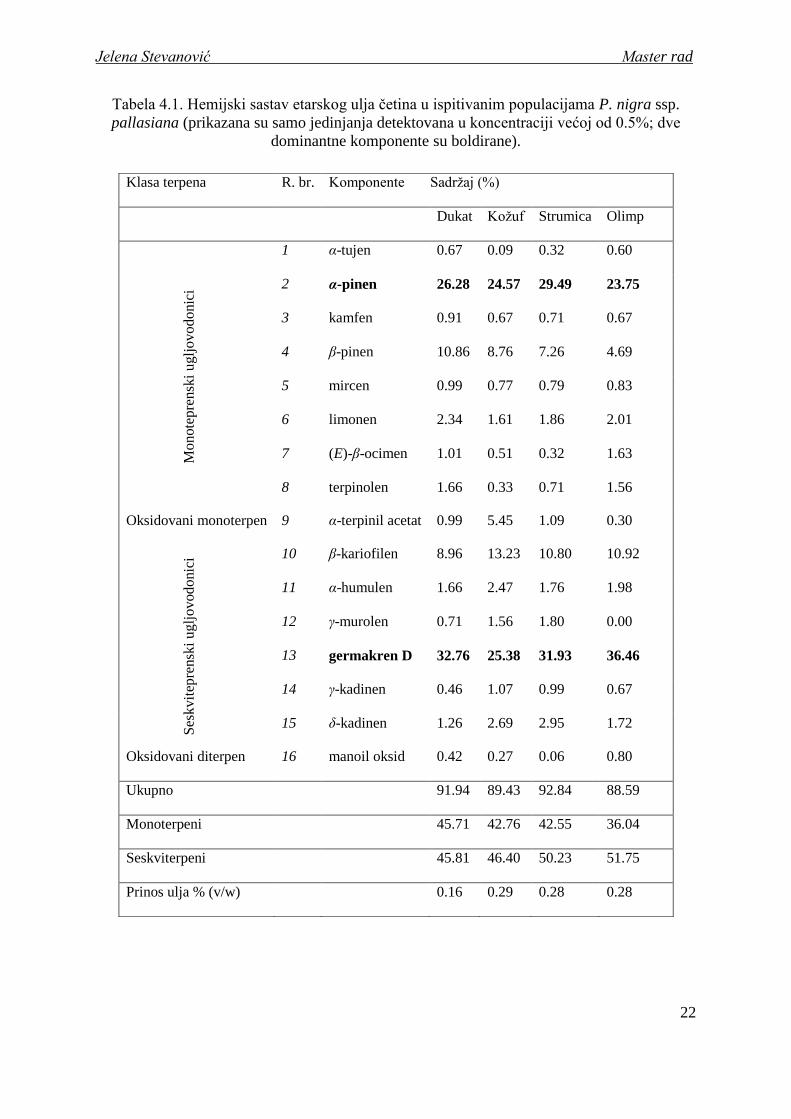
Dve dominantne komponente u etarskom ulju P. nigra ssp. pallasiana čine zajedno
57.7% ulja: seskviterpenski ugljovodonik, germakren D (13)1 (prosečna zastupljenost 31.6%)
i monoterpenski ugljovodonik, α-pinen (2) (prosečna zastupljenost 26.0%). Za preostale
prikazane komponente moţemo reći da imaju srednje visok sadrţaj (0.5 – 10.0%, prema von
Rudloff, 1975; Lieutier et al., 1991), osim α-tujena i manoil oksida čiji je prosečni sadrţaj
manji od 0.5% na nivou podvrste.
1 Italic brojevi se odnose na redne brojeve u tabeli 4.1.
Jelena Stevanović Master rad
22
Tabela 4.1. Hemijski sastav etarskog ulja četina u ispitivanim populacijama P. nigra ssp.
pallasiana (prikazana su samo jedinjanja detektovana u koncentraciji većoj od 0.5%; dve
dominantne komponente su boldirane).
Klasa terpena R. br. Komponente Sadrţaj (%)
Dukat Koţuf Strumica Olimp
Monote
pre
nsk
i u
glj
ov
od
onic
i
1 α-tujen 0.67 0.09 0.32 0.60
2 α-pinen 26.28 24.57 29.49 23.75
3 kamfen 0.91 0.67 0.71 0.67
4 β-pinen 10.86 8.76 7.26 4.69
5 mircen 0.99 0.77 0.79 0.83
6 limonen 2.34 1.61 1.86 2.01
7 (E)-β-ocimen 1.01 0.51 0.32 1.63
8 terpinolen 1.66 0.33 0.71 1.56
Oksidovani monoterpen 9 α-terpinil acetat 0.99 5.45 1.09 0.30
Ses
kvit
epre
nsk
i uglj
ovodonic
i 10 β-kariofilen 8.96 13.23 10.80 10.92
11 α-humulen 1.66 2.47 1.76 1.98
12 γ-murolen 0.71 1.56 1.80 0.00
13 germakren D 32.76 25.38 31.93 36.46
14 γ-kadinen 0.46 1.07 0.99 0.67
15 δ-kadinen 1.26 2.69 2.95 1.72
Oksidovani diterpen 16 manoil oksid 0.42 0.27 0.06 0.80
Ukupno 91.94 89.43 92.84 88.59
Monoterpeni 45.71 42.76 42.55 36.04
Seskviterpeni 45.81 46.40 50.23 51.75
Prinos ulja % (v/w) 0.16 0.29 0.28 0.28
Jelena Stevanović Master rad
23
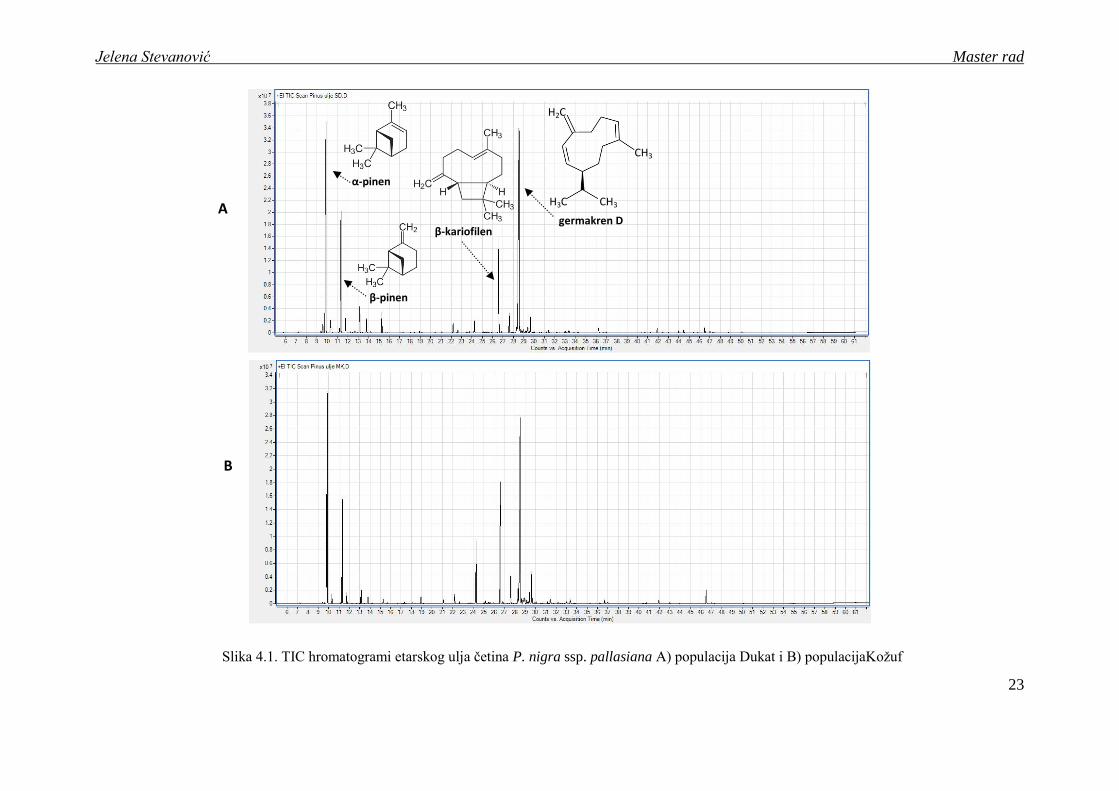
Slika 4.1. TIC hromatogrami etarskog ulja četina P. nigra ssp. pallasiana A) populacija Dukat i B) populacijaKoţuf
A
B
β-pinen
α-pinen
β-kariofilen germakren D
H2C
H3C CH3
CH3
Jelena Stevanović Master rad
24
Slika 4.1. TIC hromatogrami etarskog ulja četina P. nigra ssp. pallasiana C) populacija Strumica i D) populacijaOlimp
C
D

Jelena Stevanović Master rad
25
Tabela 4.2. Rezultati deskriptivne statistike za 16 komponenti etarskog ulja P. nigra ssp.
pallasiana (dve dominantne komponente su boldirane; Avg - srednja vrednost, Min -
minimalna vrednost, Max maksimalna vrednost, St.Dev - standardna devijacija, CV –
koeficijent varijacije).
R. br. Karakter Avg Min Max St.Dev. CV%
1 α-tujen 0.42 0.09 0.67 0.27 63.56
2 α-pinen 26.02 23.75 29.49 2.54 9.76
3 kamfen 0.74 0.67 0.91 0.11 15.53
4 β-pinen 7.89 4.69 10.86 2.60 32.89
5 mircen 0.85 0.77 0.99 0.10 11.81
6 limonen 1.96 1.61 2.34 0.31 15.61
7 (E)-β-ocimen 0.87 0.32 1.63 0.59 67.52
8 terpinolen 1.07 0.33 1.66 0.65 60.98
9 α-terpinil acetat 1.96 0.30 5.45 2.35 120.29
10 β-kariofilen 10.98 8.96 13.23 1.75 15.93
11 α-humulen 1.97 1.66 2.47 0.36 18.33
12 γ-murolen 1.02 0.00 1.80 0.82 80.97
13 germakren D 31.63 25.38 36.46 4.61 14.57
14 γ-kadinen 0.80 0.46 1.07 0.28 35.57
15 δ-kadinen 2.16 1.26 2.95 0.80 37.01
16 manoil oksid 0.39 0.06 0.80 0.31 80.55
monotepreni 41.77 36.04 45.71 4.08 9.77
seskviterpeni 48.55 45.81 51.75 2.90 5.97

Jelena Stevanović Master rad
26
Prosečan hemijski profil glavnih terpenskih komponenti (preko 5%) u etarskom ulju
P. nigra ssp. pallasiana moţemo predstaviti na sledeći način:
Germakren D >> α-Pinen >> ß-Kariofilen > ß-Pinen
(> označava razliku 1.1-5.0%, >> razliku 5.1-15.0%, prema Petrakis et al., 2001).
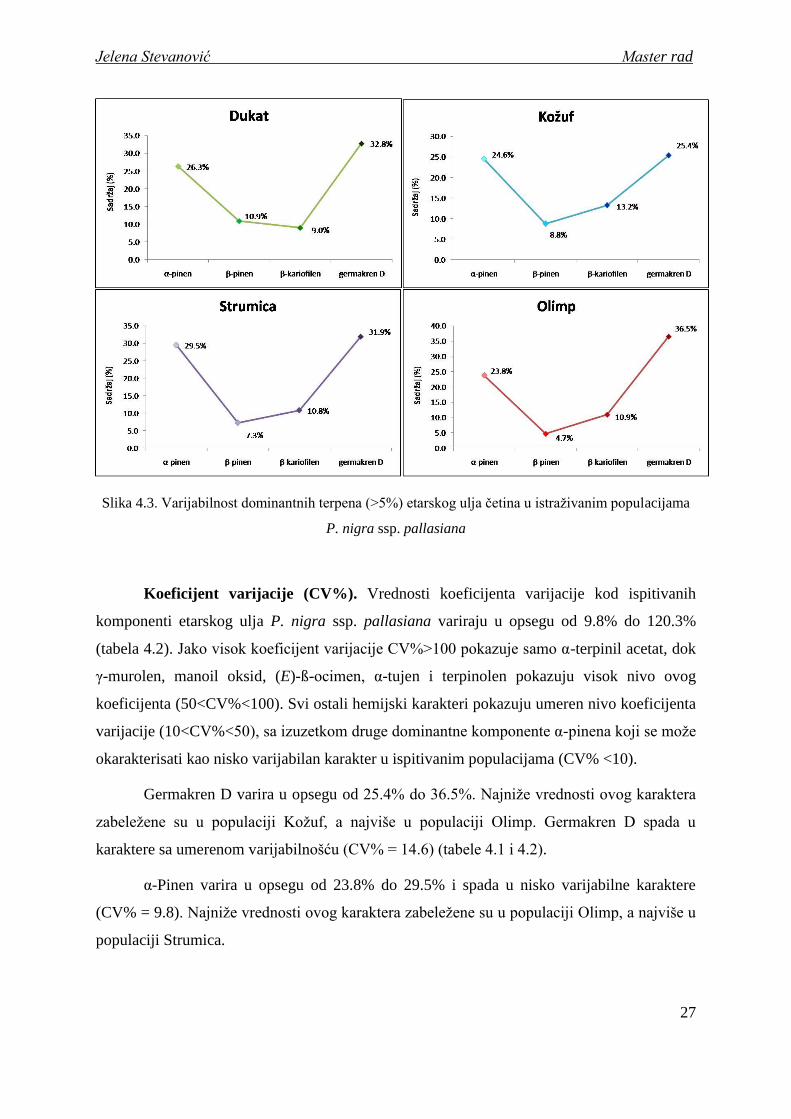
Ako uporedimo redosled dominantnih terpena izmeĎu ispitivanih populacija, moţemo
primetiti da je u svim populacijama germakren D prva, a α-pinen druga dominantna
komponenta, dok je redosled treće i četvrte komponente promenjiv. Naime, jedino je u
populaciji sa područja Srbije (Dukat) ß-pinen treća, a ß-kariofilen četvrta dominantna
komponenta, dok je u svim ostalim populacijama situacija obrnuta (slika 4.3).
Slika 4.2. Whisker plot za 16 komponenti etarskog ulja P. nigra ssp. pallasiana
Jelena Stevanović Master rad
27
Koeficijent varijacije (CV%). Vrednosti koeficijenta varijacije kod ispitivanih
komponenti etarskog ulja P. nigra ssp. pallasiana variraju u opsegu od 9.8% do 120.3%
(tabela 4.2). Jako visok koeficijent varijacije CV%>100 pokazuje samo α-terpinil acetat, dok
γ-murolen, manoil oksid, (E)-ß-ocimen, α-tujen i terpinolen pokazuju visok nivo ovog
koeficijenta (50<CV%<100). Svi ostali hemijski karakteri pokazuju umeren nivo koeficijenta
varijacije (10<CV%<50), sa izuzetkom druge dominantne komponente α-pinena koji se moţe
okarakterisati kao nisko varijabilan karakter u ispitivanim populacijama (CV% <10).
Germakren D varira u opsegu od 25.4% do 36.5%. Najniţe vrednosti ovog karaktera
zabeleţene su u populaciji Koţuf, a najviše u populaciji Olimp. Germakren D spada u
karaktere sa umerenom varijabilnošću (CV% = 14.6) (tabele 4.1 i 4.2).
α-Pinen varira u opsegu od 23.8% do 29.5% i spada u nisko varijabilne karaktere
(CV% = 9.8). Najniţe vrednosti ovog karaktera zabeleţene su u populaciji Olimp, a najviše u
populaciji Strumica.
Slika 4.3. Varijabilnost dominantnih terpena (>5%) etarskog ulja četina u istraţivanim populacijama
P. nigra ssp. pallasiana

Jelena Stevanović Master rad
28
ß-Kariofilen varira u opsegu od 9.0% do 13.2%. Najniţe vrednosti ovog karaktera su
zabeleţene su u populaciji Dukat, a najviše u populaciji Koţuf. Kao i germakren D, spada u
karaktere sa umerenom varijabilnošću (CV% = 15.9).
ß-Pinen varira u opsegu od 4.7% do 10.9% i takoĎe spada u karaktere sa umerenom
varijabilnošću (CV% = 32.9). Najniţe vrednosti ovog karaktera zabeleţene su u populaciji
Olimp, a najviše u populaciji Dukat.
4.2. Hemijski sastav etarskog ulja četina podsekcije Pinus
Za komponente etarskog ulja koje su detektovane u koncentraciji većoj od 2% u
minimalno jednom od 33 ispitivana uzorka subsect. Pinus (tabela 3.2), uraĎena je osnovna
deskriptivna statistika (srednja vrednost, maksimum, minimum, standardna devijacija i
koeficijent varijacije) na osnovu koje je uraĎena analiza varijabilnosti pojedinačnih hemijskih
karaktera (tabela 4.2). Ovi rezultati su grafički prikazani uz pomoć whisker box plota gde je
za svaki karakter prikazana njegova hemijska varijabilnost na nivou ispitivane podsekcije
(slika 4.4).
U etarskom ulju subsect. Pinus utvrĎeno je prisustvo 33 komponente detektovane u
koncentraciji većoj od 2% u minimalno jednom od ispitivanih uzoraka. Prosečno ove
komponete čine 86.6% etarskog ulja, pri čemu su monotepreni nešto više zastupljeni (45.4%)
u odnosu na seskviterpenske komponente (38.8%). Diterpeni su, kao što je i očekivano,
najslabije zastupljena klasa terpenoidnih jedinjenja u analiziranim etarskim uljima (2.5%)
(tabela 4.3).
Dve dominantne komponente u etarskom ulju subsect. Pinus čine zajedno 41.01%
ulja: monoterpenski ugljovodonik, α-pinen (1)2 (prosečna zastupljenost 21.8%) i
seskviterpenski ugljovodonik, germakren D (22) (prosečna zastupljenost 19.2%). Srednje
visok sadrţaj (0.5 – 10.0%, prema von Rudloff, 1975; Lieutier et al., 1991) ima 20
komponenti, dok je preostalih 11 prikazanih jedinjenja zastupljeno u koncentraciji ispod
0.5% na nivou podsekcije (tabela 4.3).
2 Italic brojevi se odnose na redne brojeve u tabeli 4.3.

Jelena Stevanović Master rad
29
Tabela 4.3. Rezultati deskriptivne statistike za 33 komponente etarskog ulja subsect. Pinus
(dve dominantne komponente su boldirane; Avg - srednja vrednost, Min - minimalna
vrednost, Max maksimalna vrednost, St.Dev - standardna devijacija, CV – koeficijent
varijacije).
R. br. Karakter Avg Min Max St.Dev CV%
1 α-pinen 21.84 2.10 44.20 10.21 46.74
2 kamfen 1.38 0.00 6.50 1.42 103.43
3 sabinen 0.33 0.00 4.80 0.86 258.05
4 β-pinen 7.88 0.40 21.30 5.89 74.81
5 mircen 2.24 0.00 7.70 2.05 91.42
6 δ-3-karen 1.77 0.00 16.10 4.17 235.68
7 limonen 2.62 0.00 12.50 2.71 103.29
8 β-felandren 1.66 0.00 15.60 3.75 226.27
9 (E)-β-ocimen 0.99 0.00 3.00 0.80 80.33
10 γ-terpinen 0.11 0.00 0.80 0.18 157.12
11 terpinolen 1.19 0.00 5.60 1.22 103.21
12 terpinen-4-ol 0.05 0.00 0.60 0.12 218.11
13 α-terpineol 0.42 0.00 2.30 0.61 145.69
14 timol metil etar 0.19 0.00 2.50 0.54 284.83
15 linalil acetat 0.21 0.00 2.00 0.44 206.20
16 bornil acetat 1.61 0.00 14.10 2.84 176.83
17 α-terpinil acetat 0.90 0.00 5.45 1.22 135.12
18 β-kariofilen 10.95 2.60 43.10 7.11 64.95
19 β-kopaen 0.24 0.00 3.90 0.67 276.03
20 α-humulen 1.97 0.00 9.00 1.58 80.35
21 γ-murolen 0.51 0.00 2.58 0.75 147.07
22 germakren D 19.17 0.00 38.21 11.35 59.20
23 feniletil-2-metil
butanoat 0.14 0.00 2.60 0.46 331.89
24 biciklogermakren 0.86 0.00 6.00 1.79 209.21
25 feniletil-3-metil
butanoat 0.22 0.00 2.30 0.57 262.66
26 δ-amorfen 0.21 0.00 2.50 0.44 213.85
27 γ-kadinen 0.81 0.00 3.80 0.78 95.75
28 δ-kadinen 2.40 0.00 9.00 1.87 77.85
29 kariofilen oksid 0.61 0.00 11.90 2.06 337.70
Jelena Stevanović Master rad
30
30 α-kadinol 1.01 0.00 4.30 1.32 130.30
31 kembren 0.08 0.00 1.20 0.27 353.93
32 manoil oksid 2.38 0.00 38.00 7.57 318.60
33 abietadien 0.02 0.00 0.22 0.06 284.51
monoterpeni 45.38 11.60 74.50 13.51 29.77
seskviterpeni 38.75 14.40 70.80 13.70 35.35
diterpeni 2.47 0.00 38.00 7.55 305.55
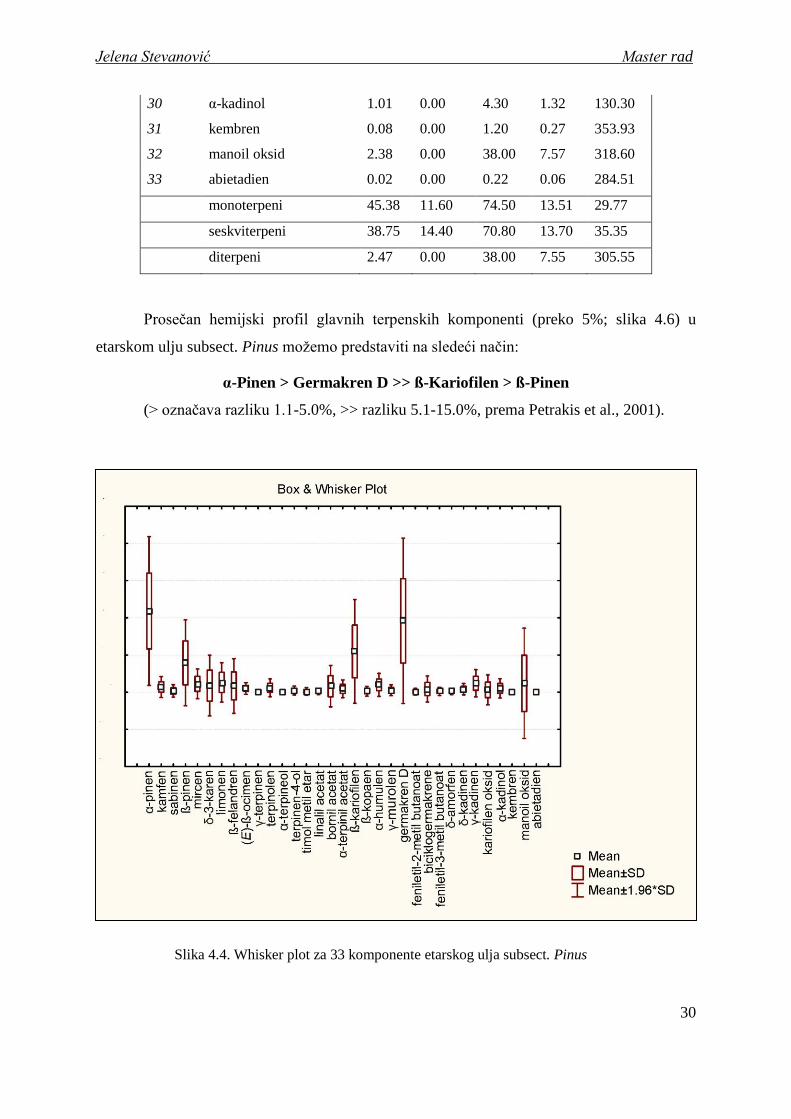
Prosečan hemijski profil glavnih terpenskih komponenti (preko 5%; slika 4.6) u
etarskom ulju subsect. Pinus moţemo predstaviti na sledeći način:
α-Pinen > Germakren D >> ß-Kariofilen > ß-Pinen
(> označava razliku 1.1-5.0%, >> razliku 5.1-15.0%, prema Petrakis et al., 2001).
Slika 4.4. Whisker plot za 33 komponente etarskog ulja subsect. Pinus

Jelena Stevanović Master rad
31
Koeficijent varijacije (CV%). Vrednosti koeficijenta varijacije kod ispitivanih
komponenti etarskog ulja subsect. Pinus variraju u opsegu od 46.7% do 353.9% (tabela 4.3).
Jako visok koeficijent varijacije (CV%>300) pokazuju 4 komponente: kembren, kariofilen
oksid, feniletil-2-metil butanoat i manoil oksid, dok visok nivo koeficijenta varijacije
(200<CV%<300) ispoljava čak 11 komponenti etarskog ulja. U kategorijama umereno
(100<CV%<200) i nisko (10<CV%<100) varijabilnih karaktera nalazi se po 9 komponenti
etarskog ulja.
Prva dominantna komponenta u etarskom ulju subsect. Pinus, α-pinen, spada u
kategoriju nisko varijabilnih karaktera, sa najniţom vrednosti koeficijenta varijacije u odnosu
na sve ostale komponente (CV% = 46.7). Preostale tri dominantne komponente (germakren
D, β-kariofilen i β-pinen), takoĎe, spadaju u nisko varijabilne karaketere (10<CV%<100).
4.3. Hemijski sastav etarskog ulja četina podsekcije Pinaster
U etarskom ulju subsect. Pinaster utvrĎeno je prisustvo 33 komponente, detektovane
u koncentraciji većoj od 2% u minimalno jednom od ispitivanih uzoraka (tabela 4.4 i slika
4.5). Prosečno ove komponete čine 79.8% etarskog ulja, pri čemu monotepreni dominiraju
(50.05%) u odnosu na seskviterpenske komponente (28.19%). Diterpeni su, kao što je i
očekivano, najslabije zastupljena klasa terpenoidnih jedinjenja u analiziranim etarskim uljima
(1.5%).
Iako je isti broj komponenti detektovan i u Pinus uljima (tabel 4.3), ineteresanto je
napomenuti da je biciklogermakren komponenta koja odlikuje isključivo Pinus, a gvajol
Pinaster ulja. Odnos mono- i seskviterpena je sličan u obe istraţivane podsekcije, iako je
monoterpenska frakcija nešto jače zastupljena u Pinaster, a seskviterpenska u Pinus uljima
(monoterpeni/seskviterpeni = 45.4%/38.8%, tabela 4.3).
Dve dominantne komponente u etarskom ulju subsect. Pinaster čine zajedno 27.3%
ulja: seskviterpenski ugljovodonik, β-kariofilen (18)3 (prosečna zastupljenost 13.9%) i
monoterpenski ugljovodonik, α-pinen (1) (prosečna zastupljenost 13.4%). Srednje visok
sadrţaj (0.5 – 10.0%, prema von Rudloff, 1975; Lieutier et al., 1991) ima 17 komponenti, dok
3 Italic brojevi se odnose na redne brojeve u tabeli 4.4.
Jelena Stevanović Master rad
32
je preostalih 14 prikazanih jedinjenja zastupljeno u koncentraciji ispod 0.5% na nivou
podsekcije (tabela 4.3).
Tabela 4.4. Rezultati deskriptivne statistike za 33 komponente etarskog ulja subsect. Pinaster
(dve dominantne komponente su boldirane; Avg – srednja vrednost, Min – minimalna
vrednost, Max maksimalna vrednost, St.Dev – standardna devijacija, CV – koeficijent
varijacije).
R. br. Karakter Avg Min Max St.Dev CV%
1 α-pinen 13.40 0.40 28.90 7.25 54.09
2 kamfen 0.34 0.00 1.50 0.38 112.17
3 sabinen 1.21 0.00 9.40 2.37 194.85
4 β-pinen 12.00 0.00 47.50 16.77 139.76
5 mircen 5.59 0.20 27.90 7.84 140.32
6 δ-3-karen 2.25 0.00 18.60 4.76 211.22
7 limonen 7.53 0.00 58.90 14.69 195.11
8 β-felandren 0.70 0.00 6.70 1.39 197.33
9 (E)-β-ocimen 0.44 0.00 1.70 0.55 123.30
10 γ-terpinen 0.41 0.00 2.80 0.75 182.97
11 terpinolen 2.23 0.00 10.50 3.50 156.55
12 terpinen-4-ol 1.82 0.00 30.10 6.16 338.20
13 α-terpineol 1.07 0.00 6.30 1.70 158.72
14 timol metil etar 0.05 0.00 1.20 0.24 489.90
15 linalil acetat 0.08 0.00 1.20 0.27 358.74
16 bornil acetat 0.36 0.00 5.30 1.06 293.28
17 α-terpinil acetat 0.56 0.00 5.30 1.27 224.98
18 β-kariofilen 13.86 0.00 40.31 9.86 71.16
19 β-kopaen 0.04 0.00 0.40 0.09 246.33
20 α-humulen 2.80 0.50 7.92 2.18 77.85
21 γ-murolen 0.44 0.00 6.80 1.38 311.71
22 germakren D 8.60 0.00 55.80 13.99 162.56
23 feniletil-2-metil
butanoat
0.11 0.00 1.00 0.25 225.96
24 feniletil-3-metil
butanoat 1.15 0.00 7.70 2.13 184.98
25 δ-amorfen 0.06 0.00 0.60 0.15 244.31
26 γ-kadinen 0.18 0.00 1.50 0.41 228.64
Jelena Stevanović Master rad
33
27 δ-kadinen 1.01 0.00 4.20 1.25 123.47
28 kariofilen oksid 0.37 0.00 1.20 0.38 101.36
29 gvajol 0.67 0.00 4.00 1.24 184.65
30 α-kadinol 0.15 0.00 1.30 0.29 192.62
31 kembren 0.80 0.00 6.50 1.89 237.19
32 manoil oksid 0.03 0.00 0.30 0.09 275.05
33 abietadien 0.69 0.00 14.80 3.02 439.25
monoterpeni 50.05 6.32 81.30 24.80 49.55
seskviterpeni 28.19 10.80 68.20 15.42 54.71
diterpeni 1.52 0.00 14.80 3.41 224.62
Prosečni hemijski profil glavnih terpenskih komponenti (preko 5%; slika 4.6) u
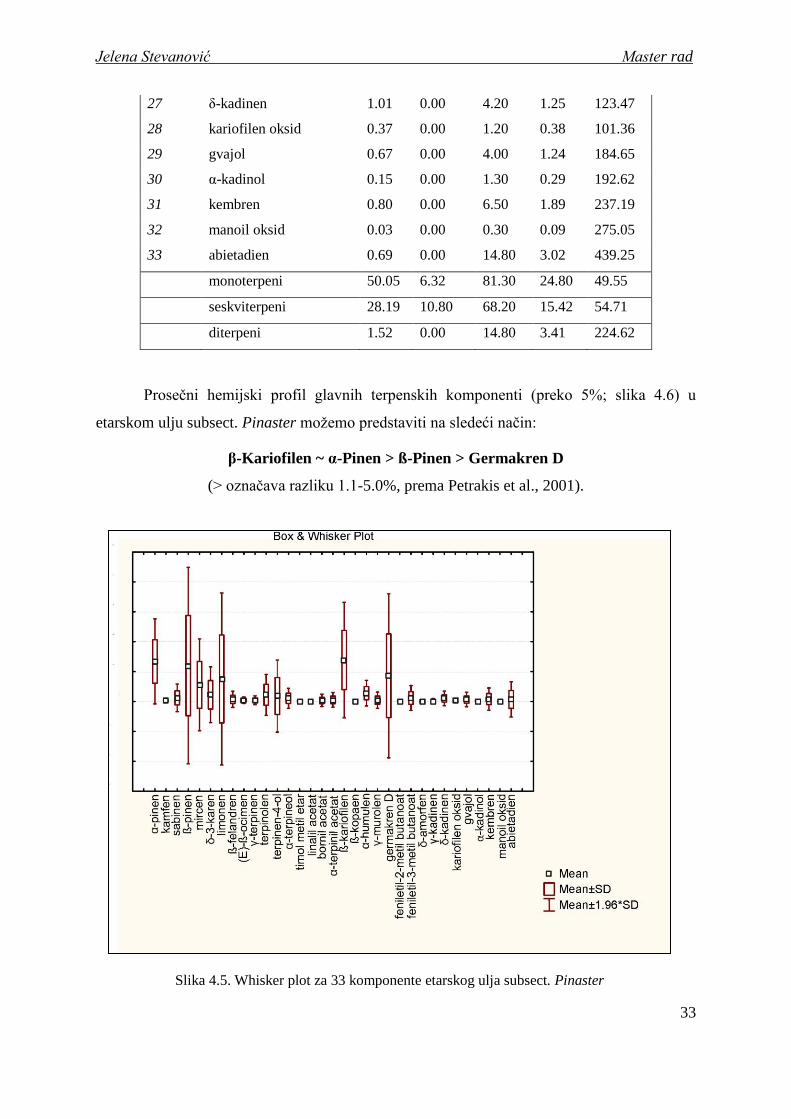
etarskom ulju subsect. Pinaster moţemo predstaviti na sledeći način:
β-Kariofilen ~ α-Pinen > ß-Pinen > Germakren D
(> označava razliku 1.1-5.0%, prema Petrakis et al., 2001).
Slika 4.5. Whisker plot za 33 komponente etarskog ulja subsect. Pinaster
Jelena Stevanović Master rad
34
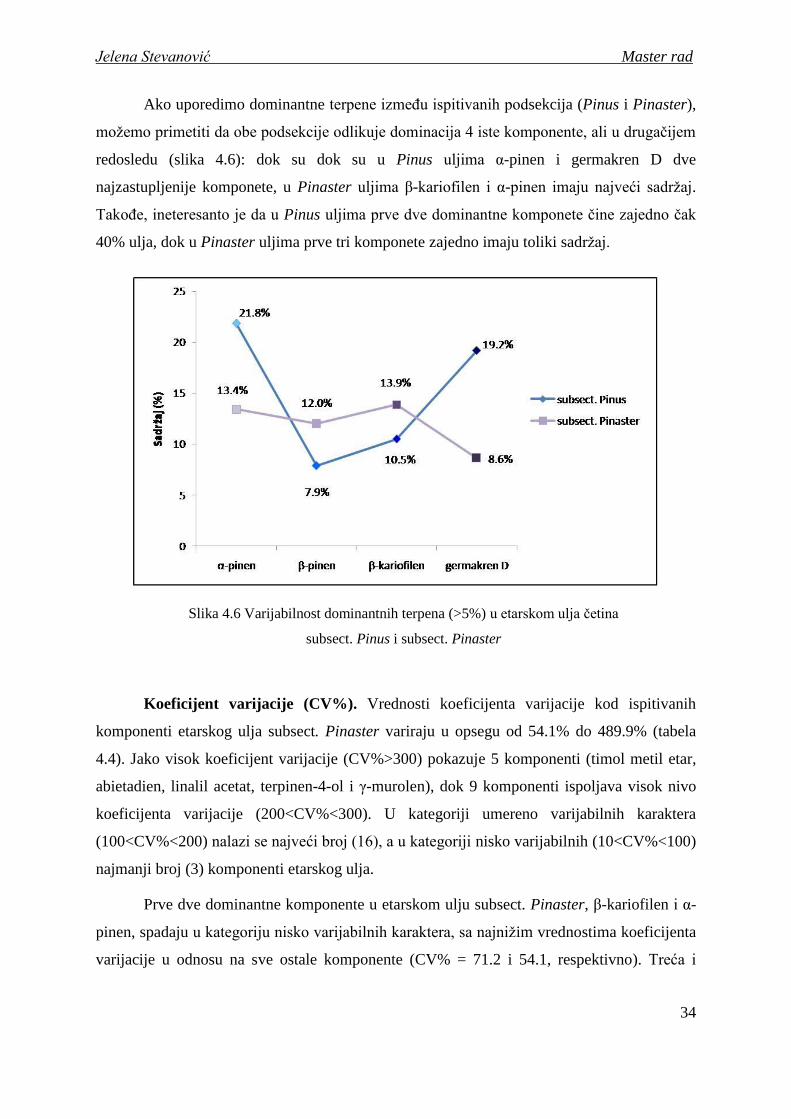
Ako uporedimo dominantne terpene izmeĎu ispitivanih podsekcija (Pinus i Pinaster),
moţemo primetiti da obe podsekcije odlikuje dominacija 4 iste komponente, ali u drugačijem
redosledu (slika 4.6): dok su dok su u Pinus uljima α-pinen i germakren D dve
najzastupljenije komponete, u Pinaster uljima β-kariofilen i α-pinen imaju najveći sadrţaj.
TakoĎe, ineteresanto je da u Pinus uljima prve dve dominantne komponete čine zajedno čak
40% ulja, dok u Pinaster uljima prve tri komponete zajedno imaju toliki sadrţaj.
Koeficijent varijacije (CV%). Vrednosti koeficijenta varijacije kod ispitivanih
komponenti etarskog ulja subsect. Pinaster variraju u opsegu od 54.1% do 489.9% (tabela
4.4). Jako visok koeficijent varijacije (CV%>300) pokazuje 5 komponenti (timol metil etar,
abietadien, linalil acetat, terpinen-4-ol i γ-murolen), dok 9 komponenti ispoljava visok nivo
koeficijenta varijacije (200<CV%<300). U kategoriji umereno varijabilnih karaktera
(100<CV%<200) nalazi se najveći broj (16), a u kategoriji nisko varijabilnih (10<CV%<100)
najmanji broj (3) komponenti etarskog ulja.
Prve dve dominantne komponente u etarskom ulju subsect. Pinaster, β-kariofilen i α-
pinen, spadaju u kategoriju nisko varijabilnih karaktera, sa najniţim vrednostima koeficijenta
varijacije u odnosu na sve ostale komponente (CV% = 71.2 i 54.1, respektivno). Treća i
Slika 4.6 Varijabilnost dominantnih terpena (>5%) u etarskom ulja četina
subsect. Pinus i subsect. Pinaster

Jelena Stevanović Master rad
35
četvrta dominantna komponenta (β-pinen i germakren D) spadaju u umereno varijabilne
karaktere (10<CV%<100), kao i najveći broj komponeti Pinaster ulja.
4.4. Analiza varijanse (ANOVA)
U cilju utvrĎivanja da li postoji statistički značajna razlika u prosečnim vrednostima
hemijskih karaktera (terpena) izmeĎu analiziranih podsekcija (subsect. Pinus i subsect.
Pinaster) uraĎena je analiza varijnse (ANOVA). ANOVA ili disperziona analiza, predstavlja
analitički model za testiranje značajnosti razlika izmeĎu srednjih vrednosti ispitivanih grupa,
koji omogućava izdvajanje karaktera koji su statistički značajni za izdvajanje i diferencijaciju
ispitivanih grupa (Petz, 1985; Bojović i Mitrović, 2010).
Za ANOVA izdvojene su 34 komponente etarskog ulja koje su detektovane u
koncentraciji većoj od 2% u minimalno jednom od istraţivanih uzoraka (tabela 4.5). Prema
rezultatima ANOVA analize moţe se videti 15, od ukupno 34 ispitivana karaktera, poseduje
visok nivo statističke značajnosti, što ukazuje na to da postoji statistički značajna razlika
izmeĎu ispitivanih podsekcija. Najveći nivo statističke značajnosti (P<0.001) pokazuju γ-
kadinen i kamfen, a zatim slede α-pinen, δ-kadinen, germakren D, α-kadinol, gvajol i (E)-β-
ocimen (P<0.01). Najmanji nivo statističke značajnosti (P<0.05) poseduju feniletil-3-metil
butanoat, mircen, biciklogermakren, γ-terpinen, kembren, α-terpineol i bornil acetat.
Tabela 4.5. Koeficijenti dobijeni ANOVA analizom (karakteri koji poseduju statističku
značajnost su boldirani; nivoi statističke značajnosti: *P<0.05; **P<0.01; ***P<0.001).
R. br. Karakter F P
1 α-pinen 11.99 **
2 kamfen 12.11 ***
3 sabinen 3.91
4 β-pinen 1.71
5 mircen 5.54 *
6 δ-3-karen 0.17
7 limonen 3.54
8 β-felandren 1.40
9 (E)-β-ocimen 8.52 **
10 γ-terpinen 4.80 *

Jelena Stevanović Master rad
36
11 terpinolen 2.55
12 terpinen-4-ol 2.74
13 α-terpineol 4.19 *
14 timol metil etar 1.38
15 linalil acetat 1.87
16 bornil acetat 4.16 *
17 α-terpinil acetat 1.04
18 β-kariofilen 1.68
19 β-kopaen 2.21
20 α-humulen 2.78
21 γ-murolen 0.05
22 germakren D 9.90 **
23 feniletil-2-metil butanoat 0.06
24 biciklogermakren 5.46 *
25 feniletil-3-metil butanoat 5.79 *
26 δ-amorfen 2.35
27 γ-kadinen 13.19 ***
28 δ-kadinen 9.95 **
29 kariofilen oksid 0.32
30 gvajol 9.75 **
31 α-kadinol 9.87 **
32 kembren 4.70 *
33 manoil oksid 2.29
34 abietadien 1.62
Ukoliko izdvojimo prvih osam komponenti etarskog ulja, koje pokazuju najveće
nivoe statističke značajnost (P<0.001 i P<0.01), hemotipovi istraţivanih podsekcija se mogu
predstaviti na sledeći način:
1. Hemotip subsect. Pinus – etarska ulja sadrţe veći procenat γ-kadinena (0.8%),
kamfena (1.4%), α-pinena (21.8%), δ-kadinena (2.4%), germakrena D (19.2%), α-
kadinola (1.0%) i (E)-β-ocimena (1.0%), dok guaiol nije detektovan.
2. Hemtip subsect. Pinaster – etarska ulja sadrţe manji procenat γ-kadinena (0.2%),
kamfena (0.3%), α-pinena (13.4%), δ-kadinena (1.0%), germakrena D (8.6%), α-

Jelena Stevanović Master rad
37
kadinola (0.2%) i (E)-β-ocimena (0.4%), dok je guaiol detektovan u prosečnoj
koncentraciji od 0.7%.
Lista istraţivanih taksona subsect. Pinus i subsect. Pinaster prikazana je u poglavlju
3.4. "Literaturni podaci", a prati klasifikaciju roda Pinus od strane Gernandt et al. (2005).
Naime, Gernandt et al. (2005) su na osnovu varijabilnosti hloroplastnih DNA sekvenci (matK
i rbcL) ukazali na postojanje dve grupe vrsta u okviru sect. Pinus (tabela 1.1). S obzirom da
je ova dihotomija takoĎe potvrĎena i od strane Wang et al. (1999), na osnovu analize 4
hloroplastne DNA sekvence, moţemo zaključiti da molekularni markeri jasno ukazuju na
postojanje dve sestrinske klade u okviru sect. Pinus uključujući "prave" mediteranske borove
sa jedne (subsect. Pinaster) i evroazijske borove sa druge strane (subsect. Pinus).
MeĎutim, Gernandt et al. (2005) navode da analiza matriksa kvalitativnih morfoloških
karaktera nije otkrila nijedan diskriminišući karakter izmeĎu navedenih podsekcija, s obzirom
da vrste obe podsekcije odlikuje trajni rukavac, broj iglica u čuperku od dve do tri, spoljašnji
ili medijalni poloţaj smolnih kanala, zrele šišarke se otvaraju nakon odreĎenog vremena,
debele plodne ljuspe sa dorzalnim poloţajem štita (umbo) i krilce koje je labavo vezano za
seme.
Kaundun i Leberton (2010) su po prvi put uspešno okarakterisali podsekcije Pinus i
Pinaster na osnovu morfometrije četina i količine metilisanog flavonola izoramnetina.
Podsekciju Pinus odlikuju relativno kraće četine (88 mm vs. 126 mmm), manja količina
metilisanog flavonola izoramnetina (IRh: 17% vs. 28%) kao i manji sadrţaj ukupnih
metilisanih flavonola - izoramnetina, laricitrina i siringetina zajedno (MeO indeks: 17 vs. 36).
S obzirom da izoramnetin nastaje od kvercetina nepovratnom reakcijom O-metil-transferaze,
taksoni koji nemaju sposobnost sinteze ovog metilisanog flavonola mogu se smatrati
ancestralnim (primitivnijim) i obrnuto. Na ovaj način, subsect. Pinus se moţe smatrati
primitivnijom u odnosu na subsect. Pinaster, meĎutim umesto zaključivanja o direktnoj vezi,
autori smatraju da je prikladnije predloţiti postojanje dve odvojene grupe na različitim
nivoima evolucije.

Jelena Stevanović Master rad
38
4.5. Klaster analiza (CA)
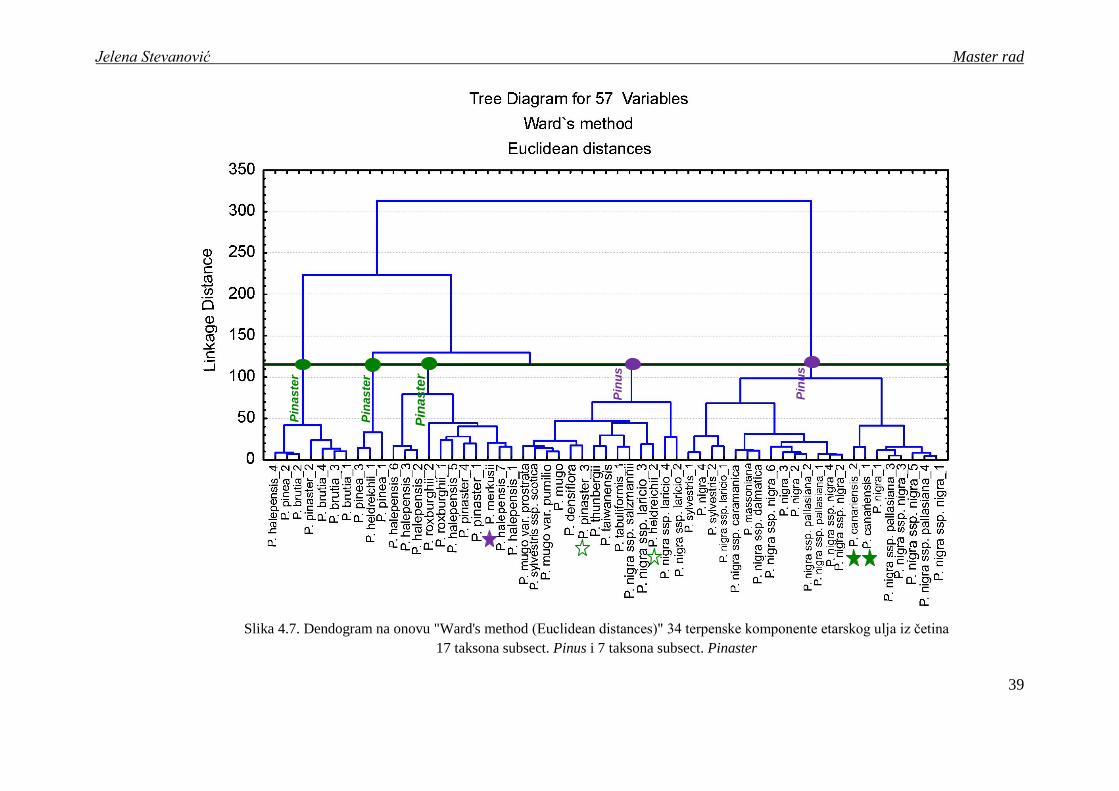
Klaster analiza na osnovu 57 uzoraka etarskog ulja (dobijenih metodom
hidrodestilacije) iz četina 24 taksona sect. Pinus ukazala je na izdvajanje 5 grupa, pri čemu
prve 3 grupe čine predstavnici podsekcije Pinaster (sa izuzetkom P. merkusii), a druge dve
članovi podsekcije Pinus (sa izuzetkom P. canariensis). Ovakvi rezultati se u velikoj meri
poklapaju sa dosadašnjim morfološkim, biohemijskim i molekularnim istraţivanjima koji
takoĎe jasno ukazuju na postojanje dve sestrinske klade u okviru sect. Pinus uključujući
"prave" mediteranske borove sa jedne (subsect. Pinaster) i evroazijske borove sa druge strane
(subsect. Pinus) (Wang et al., 1999; Gernandt et al., 2005; Kaundun i Leberton, 2010).
Na prikazanom dendogramu (slika 4.7) moţemo uočiti da prve tri grupe taksona čine
isključivo predstavnici podsekcije Pinaster sa izuzetkom vrste P. merkusii. Pinus merkusii je
azijska vrsta čija je prirodna distribucija ograničena na Vijetnam, KamboĎţu, Filipine,
Maleziju i Indoneziju, pri čemu se jedna populacija nalazi juţno od ekvatora na Sumatri. Na
ovaj način, P. merkusii predstavlja jedinu vrstu bora koja se prirodno javlja na juţnoj
Zemljinoj polulopti. Iako je ova vrsta na osnovu varijabilnosti dve cpDNK sekvence (matK i
rbcL) svrstana u subsect. Pinus (Gernandt et al., 2005) sa ostalim azijskim Diploxylon
borovima, nekoliko dosadašnjih morfoloških i fitohemijskih studija je već ukazalo na
distinktan poloţaj P. merkusii (Cooling, 1968; Weissmann i Lange, 1987; Szmidt et al.,
1996). Pored toga, udaljenost od ostalih azijskih predstavika subect. Pinus na molekularnom
nivou objavili su Wang i Szmidt (1993) na osnovu cpDNA restrikcionih mesta, Liston et al.
(1999) na osnovu jedarnog ITS regiona i Wang et al. (1999) na osnovu 4 cpDNA sekvence.
Pretpostavljeno je da je distinktan karakter P. merkusii rezultat rane separacije i duge
izolacije ove vrste u odnosu na ostale azijske predstavnike. Prema Mirov (1967), tokom
perioda Jure i Krede tropski borovi su bili prisutni u jugoisočnoj Aziji. Moguće je da je vrsta
P. merkusii nastavila svoj nezavistan razvoj u ovom regionu još od tog perioda, dok su ostale
azijske vrste migrirale u severoistočnu Aziju početkom Tercijara (Mirov, 1967). Uzimajući u
obzir sve ove činjenice kao i rezultate ovog rada, vrsta P. merkusii bi mogla biti isključena iz
subsect. Pinus. Sličan predlog dao je i Frankis (1993), koji je na osnovu sličnosti šišarki P.
merkusii i P. brutia, azijsku vrstu P. merkusii svrstao u subsect. Pinaster zajedno sa ostalim
mediteranskim borovima. Ipak, uzimajući u obzir nekonzistentnost karaktera dostupnih
morfoloških i molekularnih dokaza, dodatne studije su neophodne za svrstavanje ove vrste u
jednu ili drugu podsekciju.
Jelena Stevanović Master rad
39
Slika 4.7. Dendogram na onovu "Ward's method (Euclidean distances)" 34 terpenske komponente etarskog ulja iz četina
17 taksona subsect. Pinus i 7 taksona subsect. Pinaster
Pin
as
ter
Pin
as
ter
Pin
as
ter
Pin
us
Pin
us

Jelena Stevanović Master rad
40
Četvrtu grupu taksona čine predstavnici subsect. Pinus i po jedan uzorak P. pinaster i
P. heldreichii, dok se ostali uzorci ovih vrsta nalaze u okviru prve tri Pinaster grupe. Pinus
heldreichii (munika) je tercijarni relikt i subendemit Balkanskog poluostrva, sa malim
dopunskim arealom u juţnoj Italiji (Mirov, 1967). Taksonomski poloţaj munike je dugo
vremena bio nejasan i kontraverzan: Shaw (1914) je opisuje kao varijetet P. nigra; Mirov
(1967) je prvi ukazao na mogućnost da se radi o posebnoj vrsti s obzirom na potpuno
drugačiji terpenski sastav četina u odnosu na P. nigra; Klaus (1989) je zatim tretira kao
nezavisnu vrstu čiji su najbliţi srodnici P. nigra, P. sylvestris i azijski Diploxylon borovi
(subsect. Pinus); Schirone et al. (1991), na osnovu analize proteina semena, ukazuju na
intermedijarni poloţaj P. heldreichii izmeĎu mediteranskih (subsect. Pinaster) i evropskih
Diploxylon borova ("serija" sylvestres u okviru subsect. Pinus). MeĎutim, najnovija
molekularna istraţivanja jasno potvrĎuju distinktan taksonomski status ove retke i ugroţene
vrste kao i činjenicu da se njeni najbliţi srodnici ipak nalaze u okviru grupe "pravih"
mediteranskih borova (subsect. Pinaster) (Wang et al., 1999; Gernandt et al., 2005). S
obzirom da su ovom radu analizirana samo dva uzorka etaskog ulja P. heldreichii, pri čemu
se jedan nalazi u okviru Pinaster, a drugi u okviru Pinus grupe, za donošenje definitinvog
zaključka o poloţaju ovog kontraverznog taksona na osnovu terpenskih markera, trebalo bi
povećati broj ispitivanih uzoraka. Za sada moţemo reći da naši rezultati najpribliţniji gledištu
Schirone et al. (1991).
Petu grupu, kao i prethodnu čine predstavnici subsect. Pinus zajedno sa P.
canariensis, vrstom koja je endemična za Kanarska ostrva u Atlanskom okeanu. Iako su sve
dosadašnje studije nedvosmilsno svrstale ovu vrstu u subsect. Pinaster, izolovanost ostvrskih
populacija P. canariensis u odnosu na ostale mediteranske borove mogla se odraziti i na
drugačiji sastav etarskih ulja, po čemu je ona očigledno bliţa subsect. Pinus.

Jelena Stevanović Master rad
41
5. ZAKLJUČCI
1. U etarskom ulju četina iz četiri autohtone populacije P. nigra ssp. pallasiana sa područja
Srbije, Makedonije i Grčke detektovano je 114 jedinjenja, od kojih je 96 indentifikovano.
Od toga je samo 16 jedinjenja zastupljeno u koncentraciji većoj od 0.5%, pri čemu ove
komponente, u proseku, čine oko 90.7% ulja. Seskviterpenske komponente su nešto više
zastupljene (48.6%) u odnosu na monoterpene (41.8%), dok oksidovani diterpen (manoil
oksid) učestvuje sa svega 0.4%. Prosečan hemijski profil glavnih terpenskih kompinenti
(preko 5%) se moţe predstaviti na sledeći način:
Germakren D >> α-Pinen >> ß-Kariofilen > ß-Pinen
(> označava razliku 1.1-5.0%, >> razliku 5.1-15.0%, prema Petrakis et al., 2001).
2. U etarskom ulju subsect. Pinus utvrĎeno je prisustvo 33 komponente detektovane u
koncentraciji većoj od 2% u minimalno jednom od 33 ispitivana uzoraka. Prosečno ove
komponete čine 86.6% etarskog ulja, pri čemu su monotepreni nešto više zastupljeni
(45.4%) u odnosu na seskviterpenske komponente (38.8%). Diterpeni su, kao što je i
očekivano, najslabije zastupljena klasa terpenoidnih jedinjenja u analiziranim etarskim
uljima (2.5%). Prosečan hemijski profil glavnih terpenskih komponenti (preko 5%) u
etarskom ulju subsect. Pinus moţemo predstaviti na sledeći način:
α-Pinen > Germakren D >> ß-Kariofilen > ß-Pinen
3. U etarskom ulju subsect. Pinaster utvrĎeno je prisustvo 33 komponente, detektovane u
koncentraciji većoj od 2% u minimalno jednom od 24 ispitivana uzorka. Prosečno ove
komponete čine 79.8% etarskog ulja, pri čemu monotepreni dominiraju (50.05%) u
odnosu na seskviterpenske komponente (28.19%). Diterpeni su, kao što je i očekivano,
najslabije zastupljena klasa terpenoidnih jedinjenja u analiziranim etarskim uljima
(1.5%). Iako je isti broj komponenti detektovan i u uljima subsect. Pinus, ineteresanto je
napomenuti da je biciklogermakren komponenta koja odlikuje isključivo Pinus, a guaiol
Pinaster ulja. Odnos mono- i seskviterpena je sličan u obe istraţivane podsekcije, iako je
monoterpenska frakcija nešto jače zastupljena u u Pinaster, a seskviterpenska u Pinus

Jelena Stevanović Master rad
42
uljima. Prosečni hemijski profil glavnih terpenskih komponenti (preko 5%) u etarskom
ulju subsect. Pinaster moţemo predstaviti na sledeći način:
β-Kariofilen ~ α-Pinen > ß-Pinen > Germakren D
4. Prema rezultatima ANOVA moţe se videti 15, od ukupno 34 ispitivana karaktera,
poseduje visok nivo statističke značajnosti, što ukazuje na to da postoji statistički
značajna razlika izmeĎu ispitivanih podsekcija. Najveći nivo statističke značajnosti
(P<0.001) pokazuju γ-kadinen i kamfen, a zatim slede α-pinen, δ-kadinen, germakren D,
α-kadinol, guaiol i (E)-β-ocimen (P<0.01). Najmanji nivo statističke značajnosti (P<0.05)
poseduju feniletil-3-metil butanoat, mircen, biciklogermakren, γ-terpinen, kembren, α-
terpineol i bornil acetat. Na ovaj način, hemotipovi istraţivanih podsekcija se mogu
predstaviti na sledeći način:
Hemotip subsect. Pinus – etarska ulja sadrţe veći procenat γ-kadinena (0.8%), kamfena
(1.4%), α-pinena (21.8%), δ-kadinena (2.4%), germakrena D (19.2%), α-kadinola
(1.0%) i (E)-β-ocimena (1.0%), dok guaiol nije detektovan.
Hemtip subsect. Pinaster – etarska ulja sadrţe manji procenat γ-kadinena (0.2%),
kamfena (0.3%), α-pinena (13.4%), δ-kadinena (1.0%), germakrena D (8.6%), α-
kadinola (0.2%) i (E)-β-ocimena (0.4%), dok je guaiol detektovan u prosečnoj
koncentraciji od 0.7%.
5. Klaster analiza (CA) na osnovu 57 uzoraka etarskog ulja (dobijenih metodom
hidrodestilacije) iz četina 24 taksona sect. Pinus ukazala je na izdvajanje 5 grupa, pri
čemu prve 3 grupe čine predstavnici podsekcije Pinaster (sa izuzetkom P. merkusii), a
druge dve članovi podsekcije Pinus (sa izuzetkom P. canariensis). Ovakvi rezultati se u
velikoj meri poklapaju sa dosadašnjim morfološkim, biohemijskim i molekularnim
istraţivanjima koji jasno ukazuju na postojanje dve klade u okviru sekcije Pinus
uključujući "prave" mediteranske borove sa jedne (subsect. Pinaster) i evroazijske borove
sa druge strane (subsect. Pinus). TakoĎe, rezultati ovog rada potvrĎuju značaj terpena kao
hemotaksonomskih markera na infrageneričnom nivou.

Jelena Stevanović Master rad
43
6. LITERATURA
Adams R. P. 1991. Analysis of juniper and other forest tree oils. In: Linskins H. F., Jackson J.
F., editors. Modern methods of plant analysis. Springler, Berlin, pp. 131-157.
Adams R. P. 2007. Identification of Essential Oil Component by Gas
Chromatography/Quadrupole Mass Spectrometry, Allured Publishing Corporation,
Carol Stream, IL.
Amri I., Hanana M., Jamoussi B., Hamrouni L. 2014. Essential oils of Pinus nigra J.F.
Arnold subsp. laricio Maire: Chemical composition and study of their herbicidal
potential. Arabian J. Chem. doi:10.1016/j.arabjc.2014.05.026.
Cooling E. N. G. 1968. Pinus merkusii. Commonwealth Forestry In- stitute, Department of
Forestry, Oxford University, Oxford.
Dervendzi V. 1992. Sovremeno lekuvanje so lekoviti bilki, Tabernakul, Skopje.
Dob T., Berramdane T., Chelgoum C. 2005. Chemical composition of essential oil of Pinus
halepensis Miller growing in Algeria. C. R. Chim. 8, 1939-1945.
European Pharmacopoeia. 1997: Biological tests. 3th ed. Paris: Council of Europe, 123–7.
Farr T. G., Rosen P. A., Caro E., Crippen R., Duren R., Hensley S., Kobrick M., Paller M.,
Rodriguez E., Roth L., Seal D., Shaffer S., Shimada J., Umland J., Werner M.,Oskin
M., Burbank D., Alsdorf D. 2007. The shuttle radar topography mission. Rev.
Geophys. 45, RG2004. 10.1029/2005RG000183.
Fekih N., Allali H., Merghache S., Chaib F., Merghache D., Dib M. A., Djabou N., Muselli
A., Tabti B., Costa J. 2014. Chemical composition and antibacterial activity of Pinus
halepensis Miller growing in West Northern of Algeria. Asian Pac. J. Trop. Dis. 4,
97-103.
Frankis M. P. 1993. Morphology and affinities of Pintis brutia. In International Symposium
on Pinus brutia Ten., 11-18. Ministry of Forestry, Ankara.
Gašić M. J. 1985. Etarska ulja. IHTM, Beograd.

Jelena Stevanović Master rad
44
Gaussen H., Heywood V. H., Charter A. O. 1993. Pinus L.. In: Tutin T. G., Heywood V. H.,
Burges N. A., Valentine D. H., Walters S. M., Webb B. A., editors. Flora Europea.
Vol. 1, 2nd edn. Cambridge University Press, Cambridge, pp. 40-44.
Gernandt D. S., Geada López G., Ortiz García S., Liston A. 2005. Phylogeny and
classification of Pinus. Taxon 54, 29-42.
Hanover J. W. 1992. Applications of terpene analysis in forest genetics. New Forest. 6, 159-
178.
Ioannou E., Koutsaviti A., Tzakou O., Roussis V. 2014. The genus Pinus: a comparative
study on the needle essential oil composition of 46 pine species. Phytochem. Rev.
DOI 10.1007/s11101-014-9338-4.
Jovanović B. 1992. P. nigra Arn. In: Sarić M, editor. Flora Srbije 1. Srpska akademija nauka
i umetnosti, Beograd, pp. 200-202.
Karapandzova M., Stefkova G., Cvetkovikj I., Trajkovska-Dokik E., Kaftandzieva A.,
Kulevanova S. 2014. Chemical composition and antimicrobial activity of the essential
oils of Pinus peuce (Pinaceae) growing wild in R. Macedonia. Nat. Prod. Commun. 9,
1623-1628.
Kaundun S. S., Lebreton Ph. 2010. Taxonomy and systematic of the genus Pinus based on
morphological, biogeographical and biochemical characters. Plant Syst. Evol. 284, 1-
15.
Klaus W. 1989. Mediterranean pines and their history. Plant Syst. Evol. 162, 133-163.
Kolayli S., Ocak M., Aliyazicioglu R., Karaoglu S. 2009. Chemical analysis and biological
activities of essential oils from trunk-barks of 8 trees. Asian J. Chem. 21, 2684-2694.
Koutsaviti K., Giatropoulos A., Pitarokili D., Papachristos D., Michaelakis A., Tzakou O.
2015. Greek Pinus essential oils: larvicidal activity and repellency against Aedes
albopictus (Diptera: Culicidae). Parasitol. Res. 114, 583-592.
Kovačević N. 2004. Osnovi farmakognozije. Treće izdanje. Srpaska školska knjiga, Beograd.
Krstić N. 2016. Vrijabilnost terpena u prirodnim populacijama Pinus nigra Arnold: uticaj
ekoloških parametara i načina izolacije. Master rad. Prirodno-matematički fakultet,
Univerzitet u Nišu.

Jelena Stevanović Master rad
45
Kurose K., Okamura D., Yatagai M. 2007. Composition of the essential oils from the leaves
of nine Pinus species and the cones of three of Pinus species. Flavour Frag. J. 22, 10-
20.
Lagalante A. F., Montgomery M. E. 2003. Analysis of terpenoids from Hemlock (Tsuga)
species by solid-phase microextraction/gas chromatography/ion-trap mass
spectrometry. J. Agric. Food Chem. 51, 2115-2120.
Lahlou M. 2003. Composition and molluscicidal properties of essential oils of five Moroccan
Pinaceae. Pharm. Biol. 41, 207-210.
Liston A., Robinson W. A., Pinero D., Alvarez-Buylla E. R. 1999. Phylogenetics of Pinus
(Pinaceae) based on nuclear ribosomal DNA internal transcribed spacer region
sequences. Mol. Phylogenet. Evol. 11, 95-109.
Little E. L., Critchfield W. B. 1969. Subdivisions of the genus Pinus (Pines). USDA
Miscellaneous Publication 1144, Washington, DC.
Macchioni F., Cioni P. L., Flamini G., Morelli I., Maccioni S., Ansaldi M. 2003. Chemical
composition of essential oils from needles, branches and cones of Pinus pinea, P.
halepensis, P. pinaster and P. nigra from central Italy. Flavour Fragr. J. 18, 139-143.
Macchioni F., Cioni P. L., Flamini G., Morelli I., Perrucci S., Franceschi A., Macchioni G.,
Ceccarini L. 2002. Acaricidal activity of pine essential oils and their main
components against Tyrophagus putrescentiae, a stored food mite. J. Agric. Food
Chem. 50, 4586-4588.
Marin P. D. 2003. Biohemijska i molekularna sistematika biljaka. NNK international,
Beograd.
Mirov N. T. 1967. The genus Pinus. Ronald Press Company, New York, NY.
Nikolić B., Ristić M., Bojović S., Marin P. D. 2007. Variability of the needle essential oils of
Pinus heldreichii from different populations in Montenegro and Serbia. Chem.
Biodiversity 4, 905-916.
Nikolić B., Ristić M., Bojović S., Marin P. D. 2008. Variability of the needle essential oils of
Pinus peuce from different populations in Montenegro and Serbia. Chem.
Biodiversity 5, 1377-1388.

Jelena Stevanović Master rad
46
Nikolić B., Ristić M., Tešević V., Marin P. D., Bojović S. 2011. Terpene chemodiversity of
relict conifers Picea omorika, Pinus heldreichii, and Pinus peuce, endemic to Balkan.
Chem. Biodiversity 8, 2247-2260.
Nikolić B., Tešević V., ĐorĎević I., Marin P. D., Bojović S. 2009. Essential oil variability in
natural populations of Picea omorika, a rare European conifer. Chem. Biodiversity 6,
193-203.
Petrakis P. V., Tsitsimpikou C., Tzakou O., Couladis M., Vagias C., Roussis V. 2001. Needle
volatiles from five Pinus species growing in Greece. Flavour Fragr. J. 16, 249-252.
Petrović S., Maksimović Z., Kundaković T. 2009. Analiza sastojaka biljnih droga. Priručnik
za teorijsku i praktičnu nastavu iz predmeta Farmakognozija. Farmaceutski fakultet,
Beograd.
Politeo O., Skocibusic M., Maravic A., Ruscic M., Milos M. 2011. Chemical composition
and antimicrobial activity of the essential oil of endemic Dalmatian black pine (Pinus
nigra ssp. dalmatica). Chem. Biodiversity 8, 540-547.
Rajevski L. 1950: Nalazište Pinus nigra Arn. var. pallasiana (Lamb.) u okolini Bosilegrada.
Zbornik radova Instituta za ekologiju i biogeografiju, Srpska Akademija Nauka i
Umetnosti, Beograd.
Reuter H. I., Nelson A., Jarvis, A. 2007. An evaluation of void filling interpolation methods
for SRTM data. Int. J. Geogr. Inf. Sci. 21, 983-1008.
Rezzi S., Bighelli A., Mouillot D., Casanova J. 2001. Composition and chemical variability
of the needle essential oil of Pinus nigra subsp. laricio from Corsica. Flavour Fragr. J.
16, 379-383.
Satyal P., Paudel P., Raut J., Deo A., Dosoky N. S., Setzer W. N. 2013. Volatile constituents
of Pinus roxburghii from Nepal. Pharmacognosy Res. 5, 43-48.
Schirone B., Piovesan G., Bellarosa R., Pelosi C. 1991. A taxonomic analysis of seed proteins
in Pinus spp. (Pinaceae). Plant Syst. Evol. 178, 43-53.
Shaw G. R. 1914. The genus Pinus. Publication of Arnold Arboretum Number 5. Riverside
Press, Cambridge, MA.
Simić N. 2000. Hemijsko i mikrobiološko ispitivanje domaćih vrsta roda Achillea L..
Doktorska disertacija. Prirodno-matematički fakultet, Univerzitet u Nišu.

Jelena Stevanović Master rad
47
Syring J., Willyard A., Cronn R., Liston A. 2005. Evolutionary relationships among Pinus
(Pinaceae) subsections inferred from multiple low-copy nuclear loci. Am. J. Bot. 92,
2086-2100.
Szmidt A. E., Wang X.-R., Changtragoon S. 1996. Contrasting patterns of genetic diversity in
two tropical pines: Pinus kesiya (Royle ex Gordon) and P. merkusii (Jungh et De
Vriese). Theor. Appl. Genet. 92, 436-441.
Šarac Z. 2014. UtvrĎivanje varijabilnosti i diferencijacije prirodnih populacija Pinus nigra
Arnold u Srbiji korišćenjem fitohemijskih i molekularnih markera. Doktorska
disertacija. Biološki fakultet, Univerzitet u Beogradu.
Šarac Z., Matejić J. S., Stojanović-Radić Z. Z., Veselinović J. B., Dţamić A. M., Bojović S.,
Marin P. D. 2014. Biological activity of Pinus nigra terpenes – evaluation of FtsZ
inhibition by selected compounds as contribution to their antimicrobial activity.
Comput. Biol. Med. 54, 72-78.
Tognolini M., Barocelli E., Ballabeni V., Bruni R., Bianchi A., Chiavarini M., Impicciatore
M. 2006. Comparative screening of plant essential oils: phenylpropanoid moiety as
basic core for antiplatelet activity. Life Sci. 78, 1419-1432.
Toroglu S. 2007. In vitro antimicrobial activity and antagonistic effect of essential oils from
plant species. J. Environ. Biol. 28, 551-559.
Ustun O., Senol F. S., Kurkcuoglu M., Erdogan Orhan I., Kartal M., Baser K. H. C. 2012.
Investigation on chemical composition, anticholinesterase and antioxidant activities of
extracts and essential oils of Turkish Pinus species and pycnogenol. Ind. Crops. Prod.
38, 115-123.
Vidaković M. 1982. Četinjače. Morfologija i varijabilnost. JAZU i Sveučilišna naklada Liber,
Zagreb.
Wang X. R., Tsumura Y., Yoshimaru H., Nagasaka K., Szmidt A. E. 1999. Phylogenetic
relationships of Eurasian pines (Pinus, Pinaceae) based on chloroplast rbcL, matK,
rpl20-rps18 spacer, and trnV intron sequences. Am. J. Bot. 86, 1742-1753.
Wang X.-R., Szmidt A. E. 1993. Chloroplast DNA-based phy- logeny of Asian Pinus species
(Pinaceae). Plant Syst. Evol. 188, 197-211.

Jelena Stevanović Master rad
48
Weissmann V. G., Lange W. 1987. Zusammensetzung der Neutralteile des
Balsamkolophoniums von Pinus massoniana Lamb., Pinus merkusii Jungh. und Pinus
luchuensis Mayr. Holzforschung 41, 147-154.
Zlatković B., Šarac Z. 2016. Praktikum iz sistematike biljaka. Prirodno-matematički fakultet,
Univerzitet u Nišu.

Прилог5/1
ПРИРОДНO-MАТЕМАТИЧКИ ФАКУЛТЕТ
НИШ
КЉУЧНА ДОКУМЕНТАЦИЈСКА ИНФОРМАЦИЈА
Редни број, РБР:
Идентификациони број, ИБР:
Тип документације, ТД: Монографска
Тип записа, ТЗ: текстуални / графички
Врста рада, ВР: мастер рад
Аутор, АУ: Јелена Стевановић
Ментор, МН: Зорица С. Митић
Наслов рада, НР: Хемотаксономска карактеризација подсекција Pinus и Pinaster
(Pinus, Pinaceae) на основу терпенских маркера Језик публикације, ЈП: Српски
Језик извода, ЈИ: Енглески
Земља публиковања, ЗП: Р. Србија
Уже географско подручје, УГП: Р. Србија
Година, ГО: 2016.
Издавач, ИЗ: ауторски репринт
Место и адреса, МА: Ниш, Вишеградска 33.
Физички опис рада, ФО: (поглавља/страна/ цитата/табела/слика/графика/прилога)
48 страна; 9 слика; 8 табела
Научна област, НО: Биологија
Научна дисциплина, НД: Ботаника
Предметна одредница/Кључне речи, ПО: subsect. Pinus, subsect. Pinaster, терпени, хемотаксономија УДК 582.475 : 581.573
Чува се, ЧУ: Библиотека Важна напомена, ВН:
Извод, ИЗ: Хемијски састав етарског уља четина 24 таксона sect.
Pinus, укључујући 17 таксона subsect. Pinus и 7 таксона subsect.
Pinaster, упоређен је у циљу утврђивања хемотаксономског
значаја терпена на мезосистематском (инфрагенеричном
нивоу). Састав етарског уља P. nigra ssp. pallasiana са подручја
Србије, Македоније и Грчке анализиран је у овом раду, а
основни критеријуми за избор литературних података били су
начин изолације етарског уља (хидродестилација) и врста
органа (четине). На основу анализе варијансе (ANOVA) може се
видети да 15, од укупно 34 испитивана терпенска карактера,
поседује висок ниво статистичке значајности, што указује на то
да постоји статистички значајна разлика између испитиваних
подсекција. Кластер анализа (CA) указала је на издвајање 5
група таксона, при чему прве 3 групе чине представници
subsect. Pinaster (са изузетком P. merkusii), а друге две чланови
subsect. Pinus (са изузетком P. canariensis). Датум прихватања теме, ДП: 31.08.2016.
Датум одбране, ДО:
Чланови комисије,
КО:
Председник: Др Гордана Стојановић
Члан: Др Бојан Златковић
Члан, ментор: Др Зорица Митић
Образац Q4.09.13- Издање 1

Прилог 5/2
ПРИРОДНО - МАТЕМАТИЧКИ ФАКУЛТЕТ
НИШ
KEY WORDS DOCUMENTATION
Accession number, ANO:
Identification number, INO:
Document type, DT: monograph
Type of record, TR: textual / graphic
Contents code, CC: master thesis
Author, AU: Jelena Stevanović
Mentor, MN: Zorica S. Mitić
Title, TI: Chemotaxonomic characterisation of subsections Pinus and Pinaster (Pinus,
Pinaceae) based on terpene markers
Language of text, LT: Serbian
Language of abstract, LA: English
Country of publication, CP: Republic of Serbia
Locality of publication, LP: Serbia
Publication year, PY: 2016
Publisher, PB: author’s reprint
Publication place, PP: Niš, Višegradska 33.
Physical description, PD: (chapters/pages/ref./tables/pictures/graphs/a
ppendixes)
48 p.; 9 pictures; 8 tables
Scientific field, SF: Biology
Scientific discipline, SD: Botany
Subject/Key words, S/KW: subsect. Pinus, subsect. Pinaster, terpenes, chemotaxonomy
UC 582.475 : 581.573
Holding data, HD: library
Note, N:
Abstract, AB: The chemical composition of needle essential oils of 24 taxa from the section
Pinus, including 17 and 7 taxa of subsections Pinus and Pinaster, respectively,
were compared in order to determine chemotaxonomic significance of terpenes
on mesosystematic (infrageneric) level. The composition of essential oils in P.
nigra ssp. pallasiana from Serbia, Macedonia and Greece was analyzed in this
paper, while the basic criteria for selection of literature data included the
method of essential oil isolation (hydrodistillation) and the type of conifer organs
(needles). According to the results of analysis of variance (ANOVA), out of 34
examined terpene characters 15 had a high level of statistical significance,
indicating statistically significant difference between the examined subsections.
Cluster analysis (CA) has shown separation of five groups of taxa, where first
three groups included representatives of subsect. Pinaster (with the exception
of P. merkusii), and the other two members of subsect. Pinus (with the
exception of P. canariensis).
Accepted by the Scientific Board
on, ASB:
31.08.2016.
Defended on, DE:
Defended
Board,
DB:
President: Dr. Gordana Stojanović
Member: Dr. Bojan Zlatković
Member, Mentor: Dr. Zorica Mitić
ОбразацQ4.09.13- Издање 1