Embed Size (px)
Citation preview

The Veterinary Journal 189 (2011) 72–76
Contents lists available at ScienceDirect
The Veterinary Journal
journal homepage: www.elsevier .com/ locate / tv j l
Apoptosis in postweaning multisystemic wasting syndrome (PMWS) hepatitisin pigs naturally infected with porcine circovirus type 2 (PCV2)
Ana R. Resendes a,b, Natàlia Majó a,b, Ted S.G.A.M. van den Ingh c, Enric Mateu a,b, Mariano Domingo a,b,Maria Calsamiglia b, Joaquim Segalés a,b,⇑a Centre de Recerca en Sanitat Animal (CReSA), Facultat de Veterinària, Universitat Autònoma de Barcelona, 08193 Bellaterra (Barcelona), Spainb Departament de Sanitat i d’Anatomia Animals, Facultat de Veterinària, Universitat Autònoma de Barcelona, 08193 Bellaterra (Barcelona), Spainc Department of Pathobiology, Faculty of Veterinary Medicine, Utrecht University, 3508 TD Utrecht, The Netherlands
a r t i c l e i n f o a b s t r a c t
Article history:Accepted 25 June 2010
Keywords:Porcine circovirus type 2 (PCV2)Postweaning multisystemic wastingsyndrome (PMWS)ApoptosisCleaved caspase-3Immunohistochemistry
1090-0233/$ - see front matter � 2010 Elsevier Ltd. Adoi:10.1016/j.tvjl.2010.06.018
⇑ Corresponding author at: Centre de Recerca enDepartament de Sanitat i d’Anatomia Animals, FacultAutònoma de Barcelona, 08193 Bellaterra (Barcelona63; fax: +34 93 581 44 90.
E-mail address: [email protected] (J.
The degree of apoptosis in the livers of pigs with hepatitis due to naturally-occurring postweaning mul-tisystemic wasting syndrome (PMWS) was evaluated semi-quantitatively by immunohistochemicaldetection of the apoptotic marker cleaved caspase-3 (CCasp3). The amount and distribution of porcinecircovirus type 2 (PCV2) virus in the liver was evaluated using in situ hybridisation.
Livers with mild, stage I hepatitis exhibited similar degrees of apoptosis to controls; those with stage IIlesions had variable apoptotic rates, ranging from mild to high, and in livers with more severe, stage IIIhepatitis, high levels of hepatocyte apoptosis was a feature. Statistical analyses indicated a positive asso-ciation between the rate of apoptosis, the severity of the hepatitis and the amount of PCV2 DNA in theliver. Double immunolabelling for CCasp3 and PCV2 DNA revealed a predominance of cells labelling onlyfor PCV2, followed by fewer cells labelling only for CCasp3, and the least number labelling for both. Thefindings suggest that apoptosis, possibly triggered by PCV2 infection and/or hepatic inflammation, plays akey role in the pathogenesis of hepatitis in pigs with naturally-occurring PMWS.
� 2010 Elsevier Ltd. All rights reserved.
Introduction
Porcine circovirus type 2 (PCV2) belongs to the genus Circovirusof the Circoviridae family (small non-enveloped viruses) with a cir-cular single-stranded DNA genome of approximately 1.76 kb (Prin-gle, 1999). PCV2 is the primary causative agent of postweaningmultisystemic wasting syndrome (PMWS), a multifactorial diseaseof swine (Krakowka et al., 2000; Bolin et al., 2001).
PMWS affects nursery and fattening pigs of 5–18 weeks of ageand clinical signs include weight loss, anaemia, dyspnoea and,occasionally, diarrhoea and jaundice (Clark, 1997; Rosell et al.,1999). Mortality rates vary from 1% to 30%, and the disease hashad a significant economic impact on pig production worldwide(Harding, 2004; Segalés et al., 2005). Gross lesions include non-col-lapsed, tan-mottled lungs, enlarged lymph nodes, and less fre-quently, multifocal necrosis of lymph nodes, jaundice, hepaticatrophy and multiple white foci in kidney cortices (Rosell et al.,1999; Segalés et al., 2004). Microscopic hallmarks of PMWS includelymphocyte depletion and histiocytic infiltration of lymphoid or-
ll rights reserved.
Sanitat Animal (CReSA) andat de Veterinària, Universitat), Spain. Tel.: +34 93 581 45
Segalés).
gans, occasionally accompanied by multinucleated giant cells andbasophilic cytoplasmic inclusion bodies, interstitial pneumonia,hepatitis and nephritis (Ellis et al., 1998; Rosell et al., 2000; Dar-wich et al., 2003; Segalés et al., 2004).
PCV2 is a relatively recently recognised cause of infectious hep-atitis in pigs (Rosell et al., 2000), occurring in up to 44% of PMWS-affected animals. Four stages of hepatic damage have been de-scribed based on the amount of lymphohistiocytic infiltrationand hepatocyte injury (cytoplasmic swelling and vacuolation,necrosis, karyomegaly and apoptotic bodies), as well as on the ex-tent of the disorganisation of the hepatic plates and perilobularfibrosis (Harding and Clark, 1997; Rosell et al., 1999, 2000).
Hepatitis is classified as severe (lesion stages III and IV) in 15%of all cases of hepatitis associated with PMWS, and appears to de-velop late in the course of disease (Rosell et al., 2000; Segalés et al.,2004). Other viral causes of hepatitis in pigs include swine hepati-tis E virus (HEV), porcine reproductive and respiratory syndromevirus (PRRSV), porcine parvovirus (PPV) and Aujeszky’s diseasevirus (ADV), although HEV, PRRSV or PPV do not cause severe hep-atitis under natural or experimental infections (Krakowka et al.,2000; Halbur et al., 2001; Paul et al., 2003; de Deus et al., 2007;Lee et al., 2009).
Four patterns of PCV2 distribution are associated with PMWShepatitis and infected cells include hepatocytes, Kupffer-cells,sinusoidal endothelial cells, histiocytes and mononuclear cells

A.R. Resendes et al. / The Veterinary Journal 189 (2011) 72–76 73
(Rosell et al., 2000). Given that the severity of the hepatitis hasbeen associated with the amount and distribution of PCV2 present,and that PCV2 can replicate in hepatocytes and macrophages(Pérez-Martín et al., 2007), it has been suggested that the extentof hepatocyte infection is directly attributable to lesion severity(Rosell et al., 2000; Hirai et al., 2003; Krakowka et al., 2005).
A number of viruses encode pro-apoptotic genes, activatingtheir transcription to facilitate the release of viral progeny duringacute infection (O’Brien, 1998; Benedict et al., 2003; Irusta et al.,2003). The potential role of virus-induced apoptosis in the patho-genesis of the hepatitis associated with PMWS remains unclear.Hirai et al. (2003) demonstrated apoptosis in cases of severe hep-atitis in pigs experimentally infected with PCV2 by both electronmicroscopy and using the TUNEL-assay, whereas Krakowka et al.(2004) did not associate hepatocyte loss with apoptosis in gnotobi-otic pigs with PMWS. Furthermore, Kiupel et al. (2001) found evi-dence of apoptosis in lymphoid tissues of mice infected with PCV2.
The objective of the present study was to investigate the possi-ble role of apoptosis in the development of hepatitis in pigs thathad naturally developed PMWS using an in situ detection systemthat identifies early apoptotic events (Hengartner, 2000; Eckleet al., 2004).
Materials and methods
Case selection and sampling
Liver samples from 16 pigs, 2–3.5 months of age and affected by PMWS withdifferent degrees of hepatitis, were selected from archived formalin-fixed, paraf-fin-embedded material at the Veterinary Pathology Service of the Veterinary Schoolof Barcelona. PMWS had been diagnosed in these animals using established criteria(Segalés and Domingo, 2002) and all were negative for PRRSV, ADV and swine influ-enza virus (SIV) by immunohistochemistry, although tissues had not been evalu-ated for infection with PPV or HEV.
Five livers from clinically normal 3.5 month-old pigs used in a previous studywere used as controls (Resendes et al., 2004). PCV2 was not detected in these ani-mals using PCR on serum and in situ hybridisation (ISH) on tissues, respectively(Quintana et al., 2002). The control pigs came from a high health status farm freeof PMWS and seronegative to PRRSV, ADV, PPV, SIV and Mycoplasma hyopneumo-niae, and were euthanased using IV pentobarbital in line with European guidelineson the welfare of experimental animals (Directive 86/609 EEC).
A complete necropsy was performed on each pig, a blood sample was collectedand the tissues were fixed by immersion in 10% neutral-buffered formalin forapproximately 24 h. Tissues were then processed routinely and stained using hae-matoxylin and eosin for histopathological examination.
Histopathological examination and in situ hybridisation for PCV2
PCV2 DNA was detected using ISH (Rosell et al., 1999, 2000). In brief, thehybridisation procedure used a 40 nucleotide-long probe (50-CCTTCCTCAT-TACCCTCCTCGCCAACAATAAAATAATCAAA-30) that was complementary to the Repgene sequence (ORF1). The positive control tissue was a lymph node exhibiting his-topathological evidence of PMWS and positive for PCV2 on PCR. Lymph node tissuefrom a pig negative for PCV2 on PCR was used as a negative control.
The number and distribution of PCV2-positive cells in the liver was categorisedas previously described (Rosell et al., 2000), as follows: (1) small amounts of virus(labelled hepatocytes rarely observed, small numbers of labelled Kupffer-cells witha diffuse distribution and small numbers of positive mononuclear cells in periportalareas); (2) moderate amounts of virus (diffuse cell labelling with positive Kupffer-cells, intermediate numbers of labelled hepatocytes throughout all lobules and po-sitive mononuclear cell infiltrates in perilobular areas); (3) large amounts of virus(diffuse cell labelling with large numbers of positive hepatocytes and Kupffer-cellsin all lobules, and periportal labelling of mononuclear cells).
Hepatic injury was categorised according to the severity and distribution of:periportal and parenchymal mononuclear inflammatory infiltration; hepatocyteswelling, vacuolation, necrosis, karyomegaly, shrinkage with chromatin condensa-tion and apoptosis; and disorganisation of hepatic plates and periportal fibrosis (Ro-sell et al., 2000). These evaluations were carried out by two pathologists withoutprior knowledge of the sample group. Three stages of lesion severity were identi-fied: stage I (mild hepatitis), characterised by mild, multifocal lymphohistiocyticinfiltrates in the portal tracts and irregularly distributed in the parenchyma; stageII (moderate hepatitis), characterised by intermediate to intense lymphohistiocyticinflammation of the portal tracts often accompanied by foci of mononuclear inflam-matory cells in the parenchyma and scattered hepatocyte necrosis in some lobules,and stage III (severe hepatitis), where intense, multifocal to coalescing infiltrations
of mononuclear cells were observed in the parenchyma along with a variable de-gree of hepatic plate disorganisation and frequent hepatocyte necrosis and apopto-sis in all lobules.
Immunohistochemical examination and semi-quantitative scoring
Cleaved caspase-3 (CCasp3) was detected using a polyclonal rabbit anti-cleavedcaspase-3 antibody (anti-Asp175) diluted 1/50 (Cell Signaling Inc.) (Resendes et al.,2004). Briefly, 4 lm thick sections of liver on silane-coated slides (3-[trietoxysilyl]-propylamine), were dewaxed in xylene, rehydrated through graded alcohols, placedin distilled water and processed using the routine immunohistochemical avidin–biotin peroxidase (ABC) method. Lymphoid tissue and murine liver where apoptosishad been induced by ischaemia were used as positive controls. An isotype-matchedantibody was used in the negative control. Positive labelling was counted in 10, ran-domly-selected microscopic fields at 200� magnification. Positive hepatocytes,apoptotic bodies, endothelial cells, leucocytes and cells of undetermined type werecounted. Positive labelling within inflammatory aggregates in portal areas was notcounted.
This semi-quantitative analysis resulted in four categories: (1) no or minimalapoptosis (1–2 positive figures in some lobules); (2) low level of apoptosis (2–6 po-sitive figures in some lobules); (3) moderate level of apoptosis (6–15 positive fig-ures in most lobules), and (4) high level of apoptosis (>15 positive figures inmost/all lobules).
Double-immunostaining for CCasp3 and PCV2
Double-immunostaining to detect PCV2 DNA and CCasp3 simultaneously wascarried out by adjusting the described ISH (Rosell et al., 1999, 2000) and CCasp3immunolabelling (Resendes et al., 2004) protocols. Two cases with severe hepatitisand large amounts of both PCV2 infected and CCasp3-positive hepatocytes were as-sessed. Tissue sections were first tested using the ISH protocol and, after stainingwith nitroblue tetrazolium, were intensively washed once with 10 mM Tris (pH8) buffer (Sigma, Trizma base T-1503), twice with 1 mM EDTA and twice withTris-buffered saline (TBS) (pH 7.4). Immunohistochemical examination for CCasp3was then carried out on the sections as described above, using a routine avidin–bio-tin peroxidase protocol and detection method (0.05% diaminobenzidine, 3% hydro-gen peroxide in TBS for 10 min).
Tissue sections were counterstained with fast green prior to dehydration andmounting for examination. Positive and negative controls for PCV2 and CCasp3were included in the protocols, respectively. Semi-quantitative assessment of posi-tive labelling for CCasp3, PCV2, and for both components was expressed as low,moderate and high.
Statistical analysis
Associations between rates of apoptosis (minimal, mild, moderate and high),the severity of the hepatitis (stages I–III), amounts of PCV2 in the liver (negative,mild, moderate and high), were assessed using the v2 test. Differences were consid-ered significant when P < 0.05. Statistical analyses were performed using the StatsDirect v.2.5.4 program.
Results
The results of virus detection and its association with hepatitisseverity are summarised in Table 1. Nine pigs had stage I hepatitis,and all had small amounts of PCV2 DNA. Three animals had stage IIhepatitis and moderate amounts of viral DNA, and four pigs hadstage III hepatitis with moderate to large amounts of PCV2. All fivecontrols were negative for PCV2 DNA and none had liver lesions.
Apoptosis was minimal in the control livers (Table 1). Althoughvery occasional labelling was scattered throughout the paren-chyma, most lobules had no positive staining (Fig. 1A). CCasp3-la-belled figures were leucocytes within sinusoids, hepatocytes (somewith marginated chromatin), apoptotic bodies outside or withinhepatocytes (both chromatin-containing and chromatin-free) andendothelial cells. Similar proportions for each cell type were de-tected. All labelled cells generally displayed intense, diffuse cyto-plasmic and/or nuclear immunostaining. These apoptotic ratesand the distribution of CCasp3-labelling were taken as the normal‘background’ expression of this protein in normal pig liver.
Apoptotic labelling associated with each stage of hepatitis isalso detailed in Table 1. In cases of stage I hepatitis (n = 9) therewere minimal to low levels of apoptosis. Labelled figures and theirdistribution were similar to those detected in controls. Variable
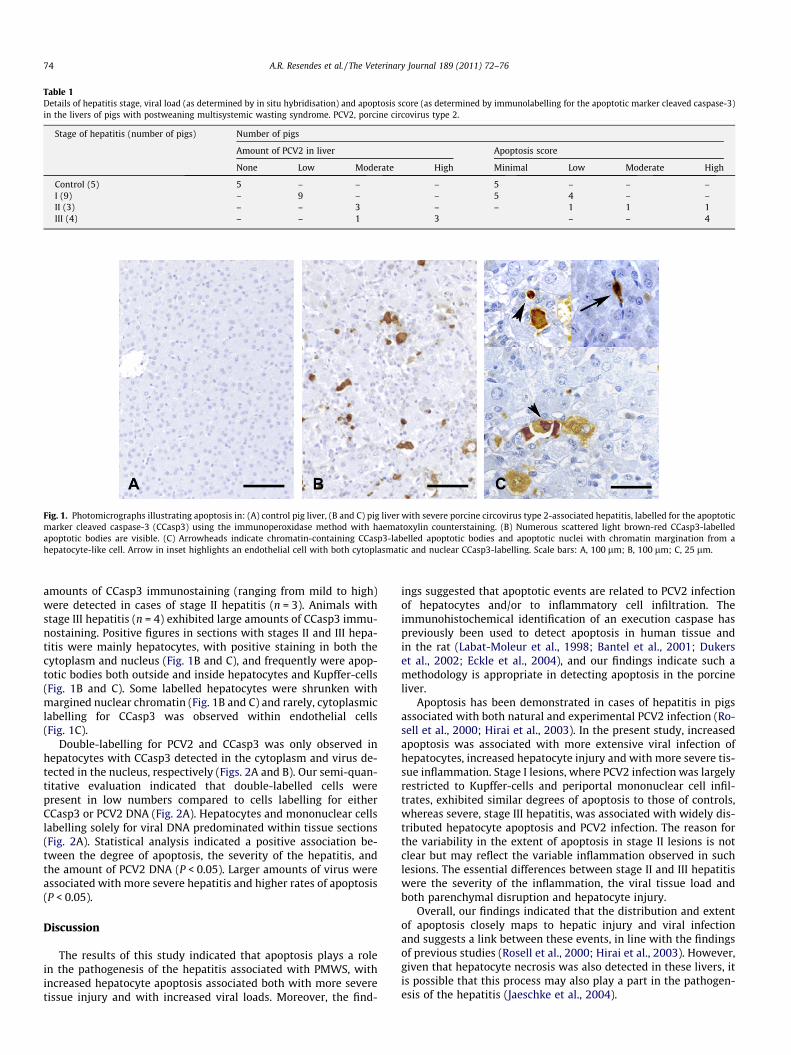
Table 1Details of hepatitis stage, viral load (as determined by in situ hybridisation) and apoptosis score (as determined by immunolabelling for the apoptotic marker cleaved caspase-3)in the livers of pigs with postweaning multisystemic wasting syndrome. PCV2, porcine circovirus type 2.
Stage of hepatitis (number of pigs) Number of pigs
Amount of PCV2 in liver Apoptosis score
None Low Moderate High Minimal Low Moderate High
Control (5) 5 – – – 5 – – –I (9) – 9 – – 5 4 – –II (3) – – 3 – – 1 1 1III (4) – – 1 3 – – 4
Fig. 1. Photomicrographs illustrating apoptosis in: (A) control pig liver, (B and C) pig liver with severe porcine circovirus type 2-associated hepatitis, labelled for the apoptoticmarker cleaved caspase-3 (CCasp3) using the immunoperoxidase method with haematoxylin counterstaining. (B) Numerous scattered light brown-red CCasp3-labelledapoptotic bodies are visible. (C) Arrowheads indicate chromatin-containing CCasp3-labelled apoptotic bodies and apoptotic nuclei with chromatin margination from ahepatocyte-like cell. Arrow in inset highlights an endothelial cell with both cytoplasmatic and nuclear CCasp3-labelling. Scale bars: A, 100 lm; B, 100 lm; C, 25 lm.
74 A.R. Resendes et al. / The Veterinary Journal 189 (2011) 72–76
amounts of CCasp3 immunostaining (ranging from mild to high)were detected in cases of stage II hepatitis (n = 3). Animals withstage III hepatitis (n = 4) exhibited large amounts of CCasp3 immu-nostaining. Positive figures in sections with stages II and III hepa-titis were mainly hepatocytes, with positive staining in both thecytoplasm and nucleus (Fig. 1B and C), and frequently were apop-totic bodies both outside and inside hepatocytes and Kupffer-cells(Fig. 1B and C). Some labelled hepatocytes were shrunken withmargined nuclear chromatin (Fig. 1B and C) and rarely, cytoplasmiclabelling for CCasp3 was observed within endothelial cells(Fig. 1C).
Double-labelling for PCV2 and CCasp3 was only observed inhepatocytes with CCasp3 detected in the cytoplasm and virus de-tected in the nucleus, respectively (Figs. 2A and B). Our semi-quan-titative evaluation indicated that double-labelled cells werepresent in low numbers compared to cells labelling for eitherCCasp3 or PCV2 DNA (Fig. 2A). Hepatocytes and mononuclear cellslabelling solely for viral DNA predominated within tissue sections(Fig. 2A). Statistical analysis indicated a positive association be-tween the degree of apoptosis, the severity of the hepatitis, andthe amount of PCV2 DNA (P < 0.05). Larger amounts of virus wereassociated with more severe hepatitis and higher rates of apoptosis(P < 0.05).
Discussion
The results of this study indicated that apoptosis plays a rolein the pathogenesis of the hepatitis associated with PMWS, withincreased hepatocyte apoptosis associated both with more severetissue injury and with increased viral loads. Moreover, the find-
ings suggested that apoptotic events are related to PCV2 infectionof hepatocytes and/or to inflammatory cell infiltration. Theimmunohistochemical identification of an execution caspase haspreviously been used to detect apoptosis in human tissue andin the rat (Labat-Moleur et al., 1998; Bantel et al., 2001; Dukerset al., 2002; Eckle et al., 2004), and our findings indicate such amethodology is appropriate in detecting apoptosis in the porcineliver.
Apoptosis has been demonstrated in cases of hepatitis in pigsassociated with both natural and experimental PCV2 infection (Ro-sell et al., 2000; Hirai et al., 2003). In the present study, increasedapoptosis was associated with more extensive viral infection ofhepatocytes, increased hepatocyte injury and with more severe tis-sue inflammation. Stage I lesions, where PCV2 infection was largelyrestricted to Kupffer-cells and periportal mononuclear cell infil-trates, exhibited similar degrees of apoptosis to those of controls,whereas severe, stage III hepatitis, was associated with widely dis-tributed hepatocyte apoptosis and PCV2 infection. The reason forthe variability in the extent of apoptosis in stage II lesions is notclear but may reflect the variable inflammation observed in suchlesions. The essential differences between stage II and III hepatitiswere the severity of the inflammation, the viral tissue load andboth parenchymal disruption and hepatocyte injury.
Overall, our findings indicated that the distribution and extentof apoptosis closely maps to hepatic injury and viral infectionand suggests a link between these events, in line with the findingsof previous studies (Rosell et al., 2000; Hirai et al., 2003). However,given that hepatocyte necrosis was also detected in these livers, itis possible that this process may also play a part in the pathogen-esis of the hepatitis (Jaeschke et al., 2004).

Fig. 2. Photomicrographs illustrating double immunolabelling of a liver with severe porcine circovirus type 2-associated hepatitis for viral nucleic acid and for the apoptoticmarker cleaved caspase-3 (CCasp3): (A) large amounts of virus (blue) and a small number of brown-staining CCasp3-positive apoptotic bodies (arrows) visible; (B) arrow ininset highlights an apoptotic hepatocyte surrounded by PCV2-positive inflammatory cells. Arrowhead points to a PCV2-positive hepatocyte that does not label for CCasp3.Scale bars: A, 50 lm; B, 25 lm.
A.R. Resendes et al. / The Veterinary Journal 189 (2011) 72–76 75
Although double-immunostaining found that a proportion ofhepatocytes exhibited both PCV2 infection and evidence of apopto-sis, the majority of hepatocytes demonstrated either one or otherof these processes suggesting a potentially more complex associa-tion between these processes, perhaps dependent on the stage ofthe cell cycle and/or on the stage of viral replication. PCV2 can rep-licate in and activate the transcription of pro-apoptotic proteinssuch as the ORF3 in both murine and porcine hepatocytes (Liuet al., 2005; Karuppannan et al., 2009). It is also possible that im-mune-mediated mechanisms are involved as exemplified by theexpression of pro-apoptotic Fas-ligand by cytotoxic T-lymphocytesin hepatitis B in humans (Mochizuki et al., 1996; Hayashi and Mita,1999).
It must be considered that co-infections with swine HEV, PPVand/or bacteria could have contributed to the development of thehepatitis in these cases, as both HEV and PPV infection have beenlinked to mild hepatic lesions in pigs (Krakowka et al., 2000; Hal-bur et al., 2001; Paul et al., 2003; Lee et al., 2009), and dual exper-imental infections with PPV and PCV2 result in severe hepaticlesions (Krakowka et al., 2000). However, evidence against such arole for co-infections include (1) the fact that severe hepatitis hasbeen induced in PCV2-inoculated gnotobiotic and conventionalpigs that did not receive any other pathogen (Krakowka et al.,2001; Fenaux et al., 2002; Ladekjaer-Mikkelsen et al., 2002); (2)pigs with PMWS infected with HEV usually have mild stage I andII hepatitis and this lesion exists in 48.2% of HEV-negative animalswith PMWS (Martin et al., 2007), and (3) PMWS has only sporadi-cally been linked to infection with PPV infection under field condi-tions (Larochelle et al., 2003). It remains possible that other, non-infectious factors are involved in the pathogenesis of this hepatitis.
Conclusions
This study suggests that apoptosis, possibly triggered by PCV2infection and/or hepatic inflammation, plays a key role in the path-ogenesis of hepatitis in pigs with naturally-occurring PMWS.
Conflict of interest statement
None of the authors of this paper has a financial or personalrelationship with other people or organisations that could inappro-priately influence or bias the content of the paper.
Acknowledgements
The authors would like to thank Merche Mora, Mónica Perez,Blanca Perez, Marina Sibila and Alex Olvera from CReSA and Unitatd’ Histologia i Anatomia Patològica, Universitat Autònoma de Bar-celona for their excellent technical assistance. The first author wasgranted a scholarship for mobility in research from Agencia de Ges-tió d’Ajuts Universitaris i de Recerca (AGAUR), Generalitat deCatalunya (Spain) and the work was funded by QLRT-PL-199900307 from European Commission’s Fifth Framework Pro-gramme (1998–2002) and 2-FEDER-1997-1341 of the Plan Nacion-al I+D (Spain).
References
Bantel, H., Ruck, P., Gregor, M., Schulze-Osthoff, K., 2001. Detection of elevatedcaspase activation and early apoptosis in liver diseases. European Journal of CellBiology 80, 230–239.
Benedict, C.A., Banks, T.A., Ware, C.F., 2003. Death and survival: viral regulation ofTNF signaling pathways. Current Opinion in Immunology 15, 59–65.
Bolin, S.R., Stoffregen, W.C., Nayar, G.P., Hamel, A.L., 2001. Postweaningmultisystemic wasting syndrome induced after experimental inoculation ofcesarean-derived, colostrum-deprived piglets with type 2 porcine circovirus.Journal of Veterinary Diagnostic Investigation 13, 185–194.
Clark, E.G., 1997. Post-weaning multisystemic wasting syndrome. In: Proceedings ofthe Annual Meeting of the American Association of Swine Practitioners. QuebecCity, Quebec, Canada, pp. 499–501.
Darwich, L., Pie, S., Rovira, A., Segalés, J., Domingo, M., Oswald, I.P., Mateu, E., 2003.Cytokine mRNA expression profiles in lymphoid tissues of pigs naturallyaffected by postweaning multisystemic wasting syndrome. Journal of GeneralVirology 84, 2117–2125.
de Deus, N., Seminati, C., Pina, S., Mateu, E., Martín, M., Segalés, J., 2007. Detection ofhepatitis E virus in liver, mesenteric lymph node, serum, bile and faeces ofnaturally infected pigs affected by different pathological conditions. VeterinaryMicrobiology 119, 105–114.
Dukers, D.F., Oudejans, J.J., Vos, W., ten Berge, R.L., Meijer, C.J., 2002. Apoptosis in B-cell lymphomas and reactive lymphoid tissues always involves activation ofcaspase 3 as determined by a new in situ detection method. Journal ofPathology 196, 307–315.
Eckle, V.S., Buchmann, A., Bursch, W., Schulte-Hermann, R., Schwarz, M., 2004.Immunohistochemical detection of activated caspases in apoptotic hepatocytesin rat liver. Toxicological Pathology 32, 9–15.
Ellis, J., Hassard, L., Clark, E., Harding, J., Allan, G., Willson, P., Strokappe, J., Martin, K.,McNeilly, F., Meehan, B., Todd, D., Haines, D., 1998. Isolation of circovirus fromlesions of pigs with postweaning multisystemic wasting syndrome. CanadianVeterinary Journal 39, 44–51.
Fenaux, M., Halbur, P.G., Haqshenas, G., Royer, R., Thomas, P., Nawagitgul, P., Gill, M.,Toth, T.E., Meng, X.J., 2002. Cloned genomic DNA of type 2 porcine circovirus isinfectious when injected directly into the liver and lymph nodes of pigs:characterization of clinical disease, virus distribution, and pathologic lesions.Journal of Virology 76, 541–551.
Halbur, P.G., Kasorndorkbua, C., Gilbert, C., Guenette, D., Potters, M.B., Purcell, R.H.,Emerson, S.U., Toth, T.E., Meng, X.J., 2001. Comparative pathogenesis of

76 A.R. Resendes et al. / The Veterinary Journal 189 (2011) 72–76
infection of pigs with hepatitis E viruses recovered from a pig and a human.Journal of Clinical Microbiology 39, 918–923.
Harding, J.C., 2004. The clinical expression and emergence of porcine circovirus 2.Veterinary Microbiology 98, 131–135.
Harding, J.C.S., Clark, E.G., 1997. Recognizing and diagnosing postweaningmultisystemic wasting syndrome (PMWS). Swine Health and Production 5,201–203.
Hayashi, N., Mita, E., 1999. Involvement of Fas system-mediated apoptosis inpathogenesis of viral hepatitis. Journal of Viral Hepatitis 6, 357–365.
Hengartner, M.O., 2000. The biochemistry of apoptosis. Nature 407, 770–776.Hirai, T., Nunoya, T., Ihara, T., Kusanagi, K., Kato, T., Shibuya, K., 2003. Acute hepatitis
in a piglet experimentally inoculated with tissue homogenates from pigs withpostweaning multisystemic wasting syndrome. Journal of Veterinary MedicalScience 65, 1041–1045.
Irusta, P.M., Chen, Y.-b., Hardwick, J.M., 2003. Viral modulators of cell death providenew links to old pathways. Cell Biology 15, 700–705.
Jaeschke, H., Gujral, J.S., Bajt, M.L., 2004. Apoptosis and necrosis in liver disease.Liver International 24, 85–89.
Karuppannan, A.K., Jong, M.H., Lee, S.-H., Zhu, Y., Selvaraj, M., Lau, J., Jia, Q., Kwang,J., 2009. Attenuation of porcine circovirus 2 in SPF piglets by abrogation of ORF3function. Virology 383, 338–347.
Kiupel, M., Stevenson, G.W., Choi, J., Latimer, K.S., Kanitz, C.L., Mittal, S.K., 2001. Viralreplication and lesions in BALB/c mice experimentally inoculated with porcinecircovirus isolated from a pig with postweaning multisystemic wasting disease.Veterinary Pathology 38, 74–82.
Krakowka, S., Ellis, J.A., Meehan, B., Kennedy, S., McNeilly, F., Allan, G., 2000. Viralwasting syndrome of swine: experimental reproduction of postweaningmultisystemic wasting syndrome in gnotobiotic swine by coinfection withporcine circovirus 2 and porcine parvovirus. Veterinary Pathology 37, 254–263.
Krakowka, S., Ellis, J.A., McNeilly, F., Ringler, S., Rings, D.M., Allan, G., 2001.Activation of the immune system is the pivotal event in the production ofwasting disease in pigs infected with porcine circovirus-2 (PCV-2). VeterinaryPathology 38, 31–42.
Krakowka, S., Ellis, J., McNeilly, F., Meehan, B., Oglesbee, M., Alldinger, S., Allan, G.,2004. Features of cell degeneration and death in hepatic failure and systemiclymphoid depletion characteristic of porcine circovirus-2-associatedpostweaning multisystemic wasting disease. Veterinary Pathology 41, 471–481.
Krakowka, S., Ellis, J., McNeilly, F., Waldner, C., Allan, G., 2005. Features of porcinecircovirus-2 disease: correlations between lesions, amount and distribution of virus,and clinical outcome. Journal of Veterinary Diagnostic Investigation 17, 213–222.
Labat-Moleur, F., Guillermet, C., Lorimier, P., Robert, C., Lantuejoul, S., Brambilla, E.,Negoescu, A., 1998. TUNEL apoptotic cell detection in tissue sections: criticalevaluation and improvement. Journal of Histochemistry and Cytochemistry 46,327–334.
Ladekjaer-Mikkelsen, A.S., Nielsen, J., Stadejek, T., Storgaard, T., Krakowka, S., Ellis, J.,McNeilly, F., Allan, G., Botner, A., 2002. Reproduction of postweaningmultisystemic wasting syndrome (PMWS) in immunostimulated and non-immunostimulated 3-week-old piglets experimentally infected with porcinecircovirus type 2 (PCV2). Veterinary Microbiology 89, 97–114.
Larochelle, R., Magar, R., D’Allaire, S., 2003. Comparative serologic and virologicstudy of commercial swine herds with and without postweaning multisystemicwasting syndrome. Canadian Journal of Veterinary Research 67, 114–120.
Lee, Y.H., Ha, Y., Ahn, K.K., Chae, C., 2009. Localisation of swine hepatitis E virus inexperimentally infected pigs. The Veterinary Journal 179, 417–421.
Liu, J., Chen, I., Kwang, J., 2005. Characterization of a previously unidentified viralprotein in porcine circovirus type 2-infected cells and its role in virus-inducedapoptosis. Journal of Virology 79, 8262–8274.
Martin, M., Segales, J., Huang, F.F., Guenette, D.K., Mateu, E., de Deus, N., Meng, X.J.,2007. Association of hepatitis E virus (HEV) and postweaning multisystemicwasting syndrome (PMWS) with lesions of hepatitis in pigs. VeterinaryMicrobiology 122, 16–24.
Mochizuki, K., Hayashi, N., Hiramatsu, N., Katayama, K., Kawanishi, Y., Kasahara, A.,Fusamoto, H., Kamada, T., 1996. Fas antigen expression in liver tissues ofpatients with chronic hepatitis B. Journal of Hepatology 24, 1–7.
O’Brien, V., 1998. Viruses and apoptosis. Journal of General Virology 79, 1833–1845.Paul, P.S., Halbur, P., Janke, B., Joo, H., Nawagitgul, P., Singh, J., Sorden, S., 2003.
Exogenous porcine viruses. Current Topics in Microbiology and Immunology278, 125–183.
Pérez-Martín, E., Rovira, A., Calsamiglia, M., Mankertz, A., Rodríguez, F., Segalés, J.,2007. A new method to identify cell types that support porcine circovirus type 2replication in formalin-fixed, paraffin-embedded swine tissues. Journal ofVirological Methods 146, 86–95.
Pringle, C.R., 1999. Virus taxonomy at the XIth international congress of virology,Sydney, Australia, 1999. Archives of Virology 144, 2065–2070.
Quintana, J., Balasch, M., Segalés, J., Calsamiglia, M., Rodriguez-Arrioja, G.M., Plana-Duran, J., Domingo, M., 2002. Experimental inoculation of porcine circovirusestype 1 (PCV1) and type 2 (PCV2) in rabbits and mice. Veterinary Research 33,229–237.
Resendes, A.R., Majo, N., Segalés, J., Espadamala, J., Mateu, E., Chianini, F., Nofrarias,M., Domingo, M., 2004. Apoptosis in normal lymphoid organs from healthynormal, conventional pigs at different ages detected by TUNEL and cleavedcaspase-3 immunohistochemistry in paraffin-embedded tissues. VeterinaryImmunology and Immunopathology 99, 203–213.
Rosell, C., Segalés, J., Domingo, M., 2000. Hepatitis and staging of hepatic damage inpigs naturally infected with porcine circovirus type 2. Veterinary Pathology 37,687–692.
Rosell, C., Segalés, J., Plana-Duran, J., Balasch, M., Rodriguez-Arrioja, G.M., Kennedy,S., Allan, G.M., McNeilly, F., Latimer, K.S., Domingo, M., 1999. Pathological,immunohistochemical, and in-situ hybridization studies of natural cases ofpostweaning multisystemic wasting syndrome (PMWS) in pigs. Journal ofComparative Pathology 120, 59–78.
Segalés, J., Domingo, M., 2002. Postweaning multisystemic wasting syndrome(PMWS) in pigs. A review. Veterinary Quarterly 24, 109–124.
Segalés, J., Rosell, C., Domingo, M., 2004. Pathological findings associated withnaturally acquired porcine circovirus type 2 associated disease. VeterinaryMicrobiology 98, 137–149.
Segalés, J., Allan, G.M., Domingo, M., 2005. Porcine circovirus diseases. AnimalHealth Research Reviews 6, 119–142.





























