Embed Size (px)
Citation preview

1475
Activation of Platelets in Blood PerfusingAngioplasty-Damaged Coronary Arteries
Flow Cytometric Detection
Riidiger E. Scharf, Aaron Tomer, Ulla M. Marzec, Paul S. Teirstein,Zaverio M. Ruggeri, and Laurence A. Harker
Fluorescence-activated flow cytometry has been used to investigate platelet activation in blood flowingthrough atherosclerotic coronary arteries after sustaining mechanical damage induced by percutaneoustransluminal angioplasty (PTCA). For flow cytometry, platelets and platelet-derived microparticles wereidentified by biotinylated anti-glycoprotein (GP) Ib monoclonal antibody (mAb) and a fluorophore,phycoerythrin-streptavidin. Activated platelets were detected by using a panel of fluoresceinated mAbsspecific for activation-dependent platelet epitopes, including 1) activated GPIIb-IIIa complex (PAC1); 2)fibrinogen bound to platelet GPIIb-IIIa (9F9); 3) ligand-induced binding sites on GPIIIa (anti-LJBSl);and 4) P-selectin, an or-granule membrane protein expressed on the platelet surface after secretion (S12).The binding of antibodies to platelets was determined in blood that was sampled continuously viaheparin-coated catheters from the coronary sinus in 1) patients before, during, and for 30 minutes afterPTCA and 2) control patients undergoing coronary angiography without PTCA. Platelets in coronarysinus blood showed significant binding of mAbs that specifically detect activation epitopes associated withthe GPIIb-IIIa complex (PAC1, anti-LJBSl, and 9F9) during and for 30 minutes after angioplasty in fourof the five patients. The relative proportion of platelets positive for PAC1 and anti-LJBSl increased frombaseline values of 2.0±0J% (mean±SD) and 2.0±0.5% to 18±14% and 28±14%, respectively, duringPTCA or 30 minutes after PTCA (p<0.01 in both cases). Binding with 9F9 was less prominent. Theexpression of P-selectin was detected in one of the five patients. By contrast, activation-specific mAbsfailed to bind detectably with platelets obtained from 1) the peripheral blood during coronary angiographyin eight patients or 2) coronary sinus blood obtained via catheter throughout control catheterizationprocedures in three patients or before PTCA in five. We conclude that circulating platelets becomeactivated while flowing through PTCA-damaged stenotic coronary arteries and that this process of plateletactivation is readily demonstrated by measuring the expression of activation-specific membrane GPepitopes by flow cytometric analysis. (Arteriosclerosis and Thrombosis 1992;12:1475-1487)
KEY WORDS • activated platelets • platelet membrane glycoproteins • activation-specificantiplatelet monoclonal antibodies • platelet receptors • flow cytometry • percutaneous transluminalcoronary angioplasty
Successful percutaneous transluminal coronary an-gioplasty (PTCA) mechanically disrupts stenos-ing atheromatous plaques.1 The resultant expo-
sure of flowing blood to the highly thrombogenicconstituents that compose subendothelial structuresand atherosclerotic lesions gives rise to the adhesionand activation of platelets.2 Concurrently, thrombin isproduced at the site of vascular injury via sequential
From the Roon Research Center for Cardiovascular Diseaseand Thrombosis (R.E.S., P.S.T., Z.M.R.), Department of Molec-ular and Experimental Medicine, The Scripps Research Institute,La Jolla, Calif., and the Division of Hematology-Oncology (A.T.,U.M.M., L.A.H.), Department of Medicine, Emory UniversitySchool of Medicine, Atlanta, Ga.
Supported in part by grants HL-41619 and HL-31950 from theNational Institutes of Health. R.E.S. is the recipient of a Heisen-berg Scholarship (Scha 358/2-1) of the Deutsche Forschungsge-meinschaft, Bonn-Bad Godesberg, FRG.
Address for reprints: Laurence A. Harker, MD, Division ofHematology and Oncology, Emory University School of Medicine,PO Drawer AR, Atlanta, GA 30322.
Received May 13, 1992; revision accepted August 31, 1992.
zymogen activation of serine proteases comprising bothextrinsic and intrinsic pathways of coagulation.3-5 Lo-cally deposited platelets further amplify thrombin gen-eration.6 Thus, PTCA-initiated thrombus formationmay lead to thrombo-occlusion in some patients7"9 andmay give rise to subsequent intimal proliferative lesionformation, both directly by liberating platelet-derivedgrowth factor and indirectly by amplifying the genera-tion of thrombin, an important mitogenic factor at sitesof vascular injury.1011
The sequence of events underlying platelet recruit-ment includes 1) adhesion of nonactivated platelets withsubendothelial cytoadhesive proteins (collagen, vonWillebrand factor [vWF], fibronectin, laminin, thrombo-spondin, and vitronectin), 2) activation of adherentplatelets and their release of proaggregatory ADP andthromboxane A2 (TxA2), 3) thrombin production, 4)activation (by thrombin, ADP, and TxA2) and receptorexpression for adhesive molecules by ambient platelets,5) platelet accumulation through platelet-ligand inter-actions (primarily binding of fibrinogen and vWF to
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

1476 Arteriosclerosis and Thrombosis Vol 12, No 12 December 1992
TABLE 1. Expression of Activation-Specific PlaUlet Epitopes and Giycoprotein Ilb-IIIa in Whole Blood From PatienU UndergoingPTCA, Control Patients, and Normal Subjects, as Evaluated by Flow Cytomerric Measurement
Patient No.
1
2
3
4
5
Total
Control patients (n=3)t
Normal subjects (n=5)
Coronaryintervention
BeforeDuring/AfterBeforeDuring/AfterBeforeDuring/AfterBeforeDuring/AfterBeforeDuring/AfterBeforeDuring/AfterBeforeDuring/After
PAC1 (act.GPIIb-IIIa)
2
2
8
23
9
30
8
14
7
19
6.8±2.8t17.6*10.55.7±1.2f6.0±2.0
5.5±1.9t
Mean fluorescence
S12(GMP-140)
2
1
2
39
2
3
2.5
2
2
2
2.1±0.29.4±16.62.5±0.52.3±0.62.1 ±0.5
intensity (arbitrary units)*mAbs bound per platelet
of FITC-conjugated
9F9 Anti-LIBSl (LIBS(platelet-bound Fg)
7
8
14
36
12
22
19
26
17
29
13.8±4.724.2±10.412.0±2.714.0±2.612.4±4.0
on GPIIIa)
12
18
17
44
11
42
4.5
14
15
44.511.9±4.832.5±15.213.3±4.014.7±4.211.4+2.1
U-P4(GPIIb-IIIa)
230
245
343
361
314
390
254
282
285
323
285 ±45320±58300±45307±49302±47
PTCA, percutaneous transluminal coronary angioplasty; FITC, fluorescein isothiocyanate; mAbs, monoclonal antibodies; act., activated;GP, giycoprotein; GMP, a-granule membrane protein; LIBS, ligand-induced binding sites; Fg, fibrinogen.
'Listed are baseline and peak levels of log platelet membrane immunofluorescence intensity, as obtained for the different mAbs in serialwhole-blood samples that were collected before, during, and after termination of coronary intervention procedures, including coronaryangiography and PTCA (patients 1-5) or coronary angiogTaphy only (control patients).
tMean±SD.tPatients for whom PTCA was considered but not performed.
giycoprotein [GP] Ilb-IIIa), and 6) stabilization ofplatelet thrombus by thrombin-catalyzed fibrin forma-tion.6 Adhesion of circulating platelets to thrombogenicsurfaces is mediated by the interaction of several plate-let functional receptors, including GPIb-IX, GPIa-IIa,GPIc-IIa, GPIIIa (IV), and the vitronectin receptorcomplexes; platelet aggregate formation depends pri-marily on the GPIIb-IIIa complex.612 Under high-shearflow conditions that are characteristic of stenosed cor-onary arteries,13 the binding of platelets to subendothe-lial vWF via the GPIb-IX complex may be important inmediating platelet accumulation.12 Since platelet GPIbalso binds with thrombin,14 GPIb may have an addi-tional role in the activation of circulating platelets.15
The recruitment of platelets after denuding vascularinjury depends on the expression by activated plateletsof a functional receptor for adhesive ligand molecules inthe GPIIb-IIIa complex.16 GPIIb-IIIa, a member of theintegrin superfamily of adhesion receptors,17 interactsvia the recognition sequence Arg-Gly-Asp common to avariety of adhesive molecules, including fibrinogen,fibronectin, vitronectin, and vWF.18-19 Other events mayoccur at the platelet surface on activation, includingconformational changes in receptors occupied by li-gands, as well as in platelet-bound ligands themselves,and expression of membrane constituents after granulesecretion, with the fusion of granule membranes to theplatelet plasma membrane.20"23 The changes at theactivated platelet surface may be evaluated by specificmurine monoclonal antibodies (mAbs) raised againstactivation-dependent platelet epitopes and bound li-gands by flow cytometry.20 Fluorescence-activated flowcytometry (FAFC) detects platelet activation by identi-
fying qualitative or quantitative changes of membraneGPs on small samples of whole blood.24-27 Using flowcytometric analysis, we now report that platelets un-dergo detectable activation during PTCA procedures inhumans.
MethodsPatients
We studied five patients undergoing PTCA, threepatients for whom PTCA was considered but not per-formed, and five normal control subjects. Mean patientage was 64 (range, 50-71) years. All patients undergoingPTCA suffered from coronary artery disease and dem-onstrated at least one lesion under consideration fordilation with a diameter of stenosis of 70% or more, asestimated by visual examination. Written informed con-sent according to the guidelines of the Human SubjectsCommittee of Scripps Clinic and Research Foundationwas obtained from all individuals participating in thisstudy.
Drug Regimens and Angioplasty ProceduresPatients received standard periprocedural medica-
tions, including aspirin (325 mg administered orally themorning of the procedure), heparin (intravenous bolusinjection of 10,000 units before PTCA and an additionalbolus of 5,000 units after 1 hour when the procedureswere still ongoing), low-molecular-weight dextran (500ml of 10% dextran 40), verapamil (5 mg), and lidocaine(75 mg). Nitroglycerin (200 /xg) was given intravenouslyimmediately before dilation.
The dilation procedure was performed using thesteerable technique.28 In all patients a successful dila-
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Scharf et al Platelet Activation During Angioplasty 1477
CUU
CUO00CU
CUcCO
.a
CU
10*
10Ji
101-
10'
A '
101 101 102 101 1Q«10
10° 101 10* 1Q1 10*
Membrane ImmunofluorescenceFIGURE 1. Quantification of an activated-platelet subpopulation by flow cytometric analysis in a patient undergoing percutaneoustransluminal coronary angioplasty (PTCA). Platelets in whole blood were doubly labeled with biotinylated PACl monoclonalantibody (mAb; reacts with activated gfycoprotein [GP] Ilb-IIIa), followed by phycoerythrin-streptavidin and a fluoresceinisothiocyanate (FITC) -conjugated anti-GPIb mAb, LJ-P3. Panel A: Bidimensional contour plot representing baseline bindinglevels of PACl and LJ-P3. An analytical marker (vertical line) was set in the red fluorescence channel to define 2% of the plateletpopulation with the highest membrane fluorescence (quadrant 2, upper right). Panel B: Contour plot analysis of a blood samplecollected during PTCA. Note the increase in fluorescence intensity (quadrant 2), corresponding to activated platelets.Quantification of this subpopulation revealed a sevenfold increase in particle number and a fivefold increase in PACl bindingcompared with baseline data. Binding of FITC-LJ-P3 to this platelet subpopulation remained unchanged, as determined by meanfluorescence intensity (115 arbitrary units in panel A and 112 in panel B). Contour lines represent numbers of platelets, with theinnermost contours corresponding to the highest numbers of platelets.
tion of the diseased coronary vessel(s) was achieved(reduction of stenosis to 50% diameter or less), asassessed visually from coronary angiograms obtainedimmediately after PTCA. The time required for vascu-lar intervention therapy and coronary angiographyranged from 20 to 60 minutes. A heparin-coated cath-eter was placed in the coronary sinus before vascularintervention therapy and remained in situ for 30 min-utes after termination of the dilation procedure to allowfurther collection of blood samples after PTCA. Theseheparin-coated catheters were a gift from Baxter Lab-oratories, Irvine, Calif.
mAbsLJ-P4, an immunoglobulin G murine mAb specific for
the GPIIb-IIIa complex on resting and activated plate-lets, was prepared and characterized as the previouslyreported anti-GPIIb-IIIa mAbs, LJ-P5 and LJ-P9.29
LJ-P4 is of the "complex-specific" variety and does notinhibit platelet aggregation. LJ-P3, a murine immuno-globulin G anti-GP Iba tnAb, which was obtained andcharacterized as described,30-31 partially inhibits bindingof vWF to platelets.30
To identify and quantify activated circulating plate-lets, a panel of murine mAbs directed to distinct acti-vation-dependent epitopes on the platelet membranewas used, including 1) PACl, specific for the activatedGPIIb-IIIa complex21; 2) 9F9, specific for surface-boundfibrinogen32; 3) anti-LIBSl for ligand (e.g., fibrinogen)-
induced binding sites on GPIIIa33; and 4) S12 forP-selectin (GMP-140), a 140-kd a-granule membraneprotein expressed on the platelet surface after secre-tion.22 These mAbs were generously provided by Dr.Sanford J. Shattil, Hospital of the University of Penn-sylvania, Philadelphia, Pa. (PACl); Dr. Andrei Z.Budzynski, Department of Biochemistry, Temple Uni-versity School of Medicine, Philadelphia, Pa. (9F9);Drs. Mark H. Ginsberg and Edward F. Plow, TheScripps Research Institute, La Jolla, Calif. (anti-LIBSl); and Dr. Rodger P. McEver, Oklahoma MedicalResearch Foundation, Oklahoma City, Okla. (S12).Antibody preparations were pure, as judged by sodiumdodecyl sulfate-polyacrylamide gel electrophoresis(data not shown).
mAbs were conjugated to fluorescein isothiocyanate(FITC, Calbiochem, La Jolla, Calif.) by standard tech-niques24-34 at a fluorescein to protein molar ratio of 7 to1. PACl, S12, and LJ-P3 were also prepared as biotiny-lated conjugates by standard techniques.34 Phycoery-thrin-streptavidin (PE-SA, Becton-Dickinson, San Jose,Calif.) was used to quantify biotinylated antibody thatwas bound to platelets. mAbs were used at saturatingconcentrations determined previously.21-22'29-31-33
Specimen CollectionBlood was continuously withdrawn via a heparin-
coated catheter (Baxter Laboratories) placed into thecoronary sinus, and serial samples (n=8) were obtained
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
1478 Arteriosclerosis and Thrombosis Vol 12, No 12 December 1992
2,000
uM
1.500-
500-
« • •
10*-
10*
1
ft-
f
B'
i——
RBCrtWBC
r\V
to* IO1 w 10*FSC
10*
10'-
10*
-
•
RfC
0r
A"ID--
10*
•
•ttnMblUM.
010" 10' 101 10* 10*
FSC
10» 10' W 10* 10*
FSC
2.5 7.5 10 15 20
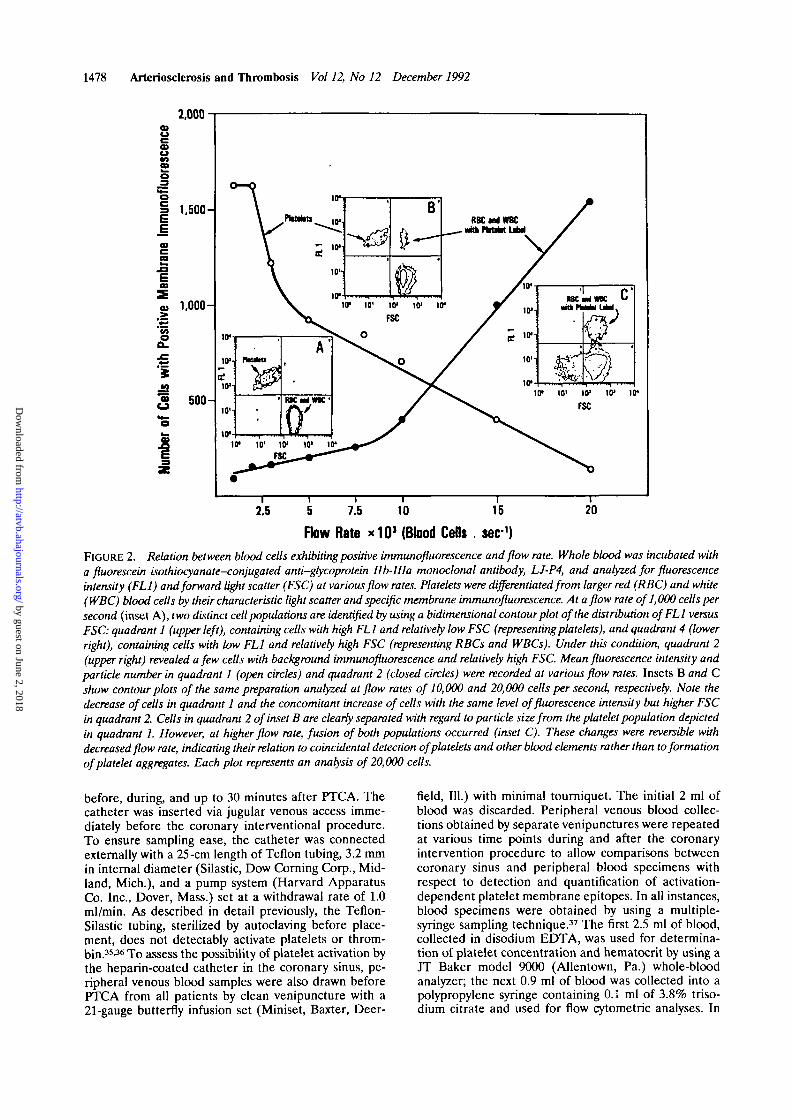
Flow Rate * 1 0 J (Blood Cefls . sec-1)FIGURE 2. Relation between blood cells exhibiting positive immunofluorescence and flow rate. Whole blood was incubated witha fluorescein isothiocyanate-conjugated anti-glycoprotein Ilb-IIIa monoclonal antibody, LJ-P4, and analyzed for fluorescenceintensity (FL1) and forward light scatter (FSC) at various flow rates. Platelets were differentiated from larger red (RBC) and white(WBC) blood cells by their characteristic light scatter and specific membrane immunofluorescence. At a flow rate of 1,000 cells persecond (inset A), two distinct cell populations are identified by using a bidimensional contour plot of the distribution ofFLl versusFSC: quadrant 1 (upper left), containing cells with high FL1 and relatively low FSC (representingplatelets), and quadrant 4 (lowerright), containing cells with low FL1 and relatively high FSC (representing RBCs and WBCs). Under this condition, quadrant 2(upper right) revealed a few cells with background immunofluorescence and relatively high FSC. Mean fluorescence intensity andparticle number in quadrant 1 (open circles) and quadrant 2 (closed circles) were recorded at various flow rates. Insets B and Cshow contour plots of the same preparation analyzed at flow rates of 10,000 and 20,000 cells per second, respectively. Note thedecrease of cells in quadrant 1 and the concomitant increase of cells with the same level of fluorescence intensity but higher FSCin quadrant 2. Cells in quadrant 2 of inset B are clearly separated with regard to particle size from the platelet population depictedin quadrant 1. However, at higher flow rate, fusion of both populations occurred (inset C). These changes were reversible withdecreased flow rate, indicating their relation to coincidental detection of platelets and other blood elements rather than to formationof platelet aggregates. Each plot represents an analysis of 20,000 cells.
before, during, and up to 30 minutes after PTCA. Thecatheter was inserted via jugular venous access imme-diately before the coronary interventional procedure.To ensure sampling ease, the catheter was connectedexternally with a 25-cm length of Teflon tubing, 3.2 mmin internal diameter (Silastic, Dow Corning Corp., Mid-land, Mich.), and a pump system (Harvard ApparatusCo. Inc., Dover, Mass.) set at a withdrawal rate of 1.0ml/min. As described in detail previously, the Teflon-Silastic tubing, sterilized by autoclaving before place-ment, does not detectably activate platelets or throm-bin.35-36 To assess the possibility of platelet activation bythe heparin-coated catheter in the coronary sinus, pe-ripheral venous blood samples were also drawn beforePTCA from all patients by clean venipuncture with a21-gauge butterfly infusion set (Miniset, Baxter, Deer-
field, 111.) with minimal tourniquet. The initial 2 ml ofblood was discarded. Peripheral venous blood collec-tions obtained by separate venipunctures were repeatedat various time points during and after the coronaryintervention procedure to allow comparisons betweencoronary sinus and peripheral blood specimens withrespect to detection and quantification of activation-dependent platelet membrane epitopes. In all instances,blood specimens were obtained by using a multiple-syringe sampling technique.37 The first 2.5 ml of blood,collected in disodium EDTA, was used for determina-tion of platelet concentration and hematocrit by using aJT Baker model 9000 (Allentown, Pa.) whole-bloodanalyzer; the next 0.9 ml of blood was collected into apolypropylene syringe containing 0.1 ml of 3.8% triso-dium citrate and used for flow cytometric analyses. In
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Scharf et al Platelet Activation During Angioplasty 1479
TABLE 2. Plasma Levels of Platelet-Specific Proteins and Fibnnopeptide A in Patients Undergoing PTCA, ControlPatients, and Normal Subjects
Patient No.
1
2
3
4
5
Total
Control patients (n=3)$
Normal subjects (n°5)
Coronaryintervention
BeforeDuringAfterBeforeDuringAfterBeforeDuringAfterBeforeDuringAfterBeforeDuringAfterBeforeDuringAfterBeforeDuringAfter
0TG
1.5
16.388.0
1.6
34.422.726.186.1
113.023.030.541.09.3
17.439.7
12.3±11.7t36.9±28.66O.9±37.912.0±7.2t13J±8.113.7±7.5
3.8±1.0t
Plasma levels (ng/ml)* of
PF4
72.076.068.019.461.660.056.050.040.058.074.089.038.038.048.0
48.7+20.559.9+16.161.0+19.048.3 + 13.748.7+10.049.0+19.11.5+0.5
FPA
4.8
4.3
4.3
3.3
3.8
2.4
3.5
5.0
4.5
1.1
2.1
2.0
1.8
2.5
2.0
2.9±1.53.5 + 1.23.0±1.32.4±0.93.3 + 1.03.3±0.71.4±0.4
PTCA, percutaneous transluminal coronary angioplasty; 0TG, /J-thromboglobulin; PF4, platelet factor 4; FPA,fibrinopeptide A.
•Listed are baseline and peak levels of serial determinations performed in each individual patient before, during, andafter termination of coronary intervention procedures, including coronary angiography and PTCA (patients 1-5) orcoronary angiography only (control patients).
tMean±SD.•Patients for whom PTCA was considered but not performed.
addition, serial coronary sinus blood samples werecollected and processed for radioimmunologic determi-nation of plasma levels of /J-thromboglobulin (/3TG),platelet factor 4 (PF4), and fibrinopeptide A (FPA) asdescribed previously.37-41
Preparation of Whole-Blood Samples forFlow Cytometric Analysis
Immunolabeling of platelets with mAbs was per-formed using minor modifications of previously pub-lished procedures24 that allow either one- or dual-coloranalysis by flow cytometry. Briefly, immediately afterblood collection, 5-/xl aliquots were added to polypro-pylene tubes (Becton-Dickinson) preloaded with 45 jxlphosphate-buffered saline (PBS) and saturating concen-trations of one of the following FITC-conjugated mAbs(final concentrations: PAC1, 30 Mg/ml; 9F9, 40 tiglm\;anti-LIBSl, 25 /xg/ml; S12, 10 /ig/ml; LJ-P4, 5 /ng/ml;and LJ-P3, 5 Mg/ml). Nonspecific membrane immuno-fluorescence was determined by using an anti-humanTG mAb. Autofluorescence of platelets was determinedin an antibody-unlabeled whole-blood sample processedsimilarly. Samples were incubated in the dark, withoutstirring, for 15 minutes at room temperature, thendiluted with 1 ml PBS and analyzed by flow cytometrywithin 2 hours of collection. Immunolabeling for dual-color analysis was performed in one of two ways. First,
in initial experiments (patients 1 and 2; Table 1),FITC-LJ-P3 was used to identify platelets and platelet-derived micToparticles, whereas PE-SA conjugate wasused to detect binding of biotinylated mAbs to activa-tion-dependent epitopes (Figure 1). Second, in patients3-5, a combination of biotinylated LJ-P3 (followed byPE-SA) and FITC-conjugated activation-specific mAbswas used to correlate functional changes in membraneGP epitopes in activated platelet subpopulations, asdetermined by one-color analysis. In both cases, 5-/xlaliquots of whole blood were incubated with the bio-tinylated primary mAbs for 15 minutes at room temper-ature, followed by the simultaneous addition of 20 ylPE-SA and FITC-conjugated mAb at a saturating con-centration. After another incubation for 15 minutes atroom temperature, samples were diluted with 1 ml PBSand analyzed.
Flow Cytometric Analysis of Platelets in Whole BloodBlood samples were analyzed with a FACStar flow
cytometer (Becton-Dickinson), equipped with a 2-Wcoherent argon-ion laser set at an excitation wavelengthof 488 nm. The flow cytometer was calibrated withstandard fluorescent microbeads (Flow CytometryStandards Corp., Research Triangle Park, N.C.). Aneutral-density filter (optical density of 0.65, transmis-sion of 22.39%) was placed in front of the forward-side
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
1480 Arteriosclerosis and Thrombosis Vol 12, No 12 December 1992
PAC1
0 5 10 - 15 20 25 W 35 40 45 60 55 (0
Time (nin)
0 5 10 15 20 25 30 35 40 45
Time (min)
c w20
I -
10
0
2D
10
•E 40C9
0J
i PAC1
Dhtka
h— Conaar Angfagrapfcr—H
- o D 9F9
*UBS1
10 15 20 25 30 35 40 45
Tana (min)
D
?
30-
20-
10-
-
CO
30-
20 -
10-
50
40-
30
2 0 -
10-
40
20-
0
i PAC1
10 15 20 25 30 35 40 45
Tone (mm)
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Scharfetal Platelet Activation During Angioplasty 1481
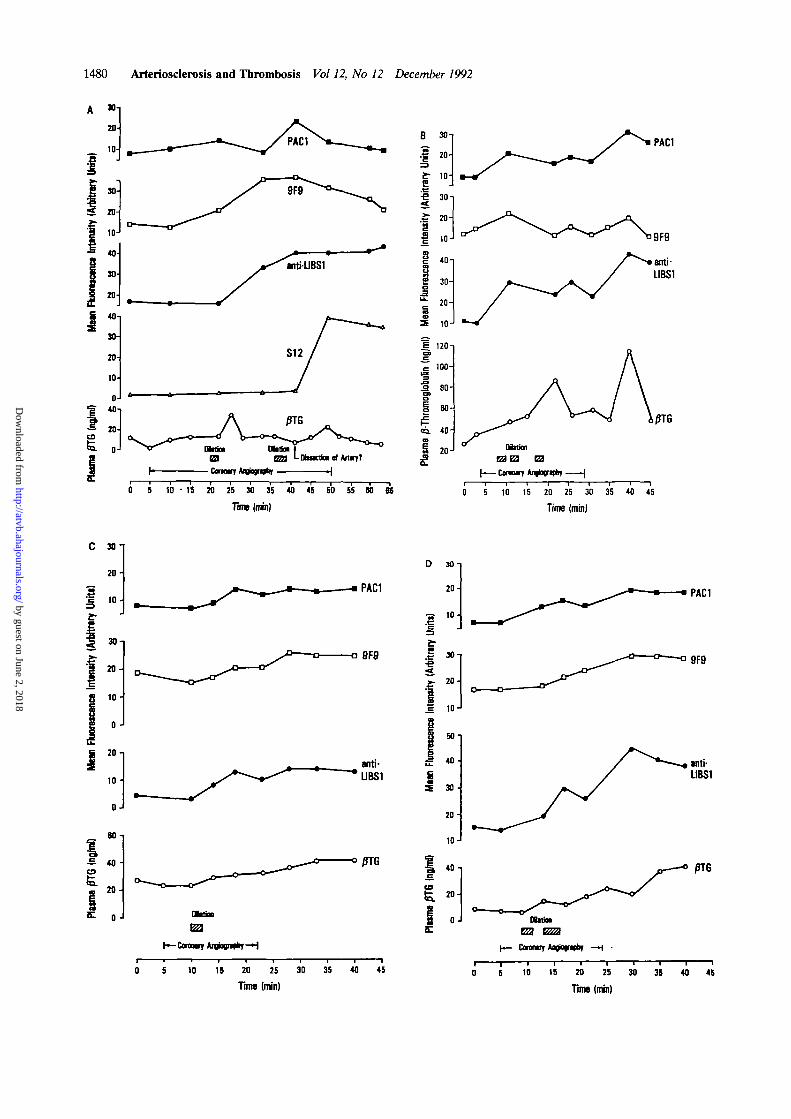
FIGURE 3. Time course of changes in the expression ofactivation-dependent platelet epitopes and plasma levels ofp-thromboglobulin (fiTG) in patients undergoing percutane-ous transluminal coronary angioplasty. Binding of activation-specific antibodies, prepared as fluorescein isothiocyanate-conjugates, to the total platelet population was evaluated byone-color flow cytometric analysis in whole blood and ex-pressed as log mean fluorescence intensity. Plasma levels ofPTG were determined by radioimmunoassay. Panels A-Dshow data of patients No. 2-5 (Table 1). As indicated in panelA, S12-positive platelets were detected in several blood samplesconcurrently with the angiographically suspected dissection ofa dilated coronary artery. Note that the expression of P-selec-tin (GMP-140) was associated with a minor and only tran-sient increase in plasma /3TG.
scatter detector to optimize resolution. FITC and PEfluorescence was detected with a 530/30- and 575/26-nm band-pass filter, respectively, and overlap in theemission spectra of FITC (green) and PE (red) wascorrected. Blood samples were analyzed at a flow rate ofless than 2,000 cells per second as detailed in "Results."Platelets and platelet-related microparticles were dif-ferentiated from red and white blood cells by theircharacteristic light scatter and specific membrane im-munofluorescence. Data from 5,000 to 10,000 plateletsand microparticles were recorded in a logarithmic modeand analyzed on a Hewlett-Packard H-P 217 computer(Palo Alto, Calif.) using a Consort 30 program(Becton-Dickinson).
Analysis of Activated Platelet Subpopulations andPlatelet-Derived Microparticles
Antibody binding was expressed as the mean fluores-cence intensity in arbitrary fluorescence units of theanalyzed platelet population. To increase resolution ofplatelet activation, an analytical marker was set withinthe fluorescence profile for each antibody at baselinelevel (before PTCA) to define a 2% and 10% plateletsubpopulation with the highest membrane immunoflu-orescence (Figure 1). This marker was used as a thresh-old to determine the proportion of platelets exhibitingimmunofluorescence above these levels in all subse-quent samples. Unless otherwise stated, data obtainedby one-color analysis were used to quantify plateletactivation. Platelet-derived microparticles were de-tected on the basis of their size, as assessed by forwardlight scatter and distinct immunofluorescence.32-42-43
Statistical AnalysesStatistical comparisons of the data were performed
with the Wilcoxon matched-pairs signed-rank test. Alldata are given as the mean±SD. Differences wereconsidered significant atp<0.05.
ResultsStandardization of FAFC
In initial experiments, several potentially confound-ing variables for the FAFC analysis were evaluated,including 1) fixation of blood samples with paraformal-dehyde; 2) collection of blood in the presence of pros-taglandin E, (PGE,); 3) time intervals between bloodsampling, incubation with activation-dependent mAbs,
and FAFC analysis; 4) drugs administered to patientsundergoing PTCA; and 5) effects of flow rate on FAFCanalysis of platelets in whole blood.
The effect of in vitro blood fixation on flow cytomet-ric measurements was evaluated. Binding of LJ-P4,anti-LIBSl, and PAC1 to both resting and stimulatedplatelets (10 fiM ADP or 1 /iM phorbol myristateacetate [PMA]) in blood that was fixed with 1%paraformaldehyde before incubation with FITC-con-jugated mAbs was significantly reduced compared withunfixed platelets. This inhibition of binding was ob-served at paraformaldehyde concentrations as low as0.1%. Similar observations have been reported byothers24-32 studying the binding of PAC1 and S12 toactivated platelets fixed with paraformaldehyde. Con-sequently, fixation of blood samples was avoided in thepresent study.
Spontaneous activation of platelets in vitro was stud-ied in citrated whole-blood samples collected in thepresence or absence of 10 or 100 /*M PGE,. Thesamples were stored at room temperature and at varioustimes up to 90 minutes; aliquots were incubated withfluoresceinated LJ-P4, anti-LIBSl, or PAC1 for 15minutes at room temperature, diluted 20-fold with 1 mlPBS, and analyzed by flow cytometry. At time zero, nosignificant difference in mAb binding was observedbetween the samples. In blood samples collected with-out PGE, there was a continuous increase in binding ofFITC-LJ-P4 and in the percentage of platelets positivefor binding of anti-LIBSl or PAC1. The increasedbinding showed a bimodal distribution curve, with slowincreases between time zero and 60 minutes (up to1.2-fold only) and relatively rapid increases between 60and 90 minutes (up to twofold). PGE, at either concen-tration eliminated this effect. Moreover, PGE! reversedspontaneous in vitro platelet activation (data notshown). Thus, PGE, was not added to the anticoagu-lated blood samples from patients undergoing PTCA.Instead, blood aliquots were immediately mixed withantibody containing buffer, incubated for 15 minutes atroom temperature, and then diluted 20-fold with PBS.In addition, throughout the study, control samples fromhealthy volunteers were always run in parallel to assessthe possibility of spontaneous platelet activation duringthe labeling procedure. Increased incubation times to 30minutes did not result in increased binding of anti-LIBSl or PAC1. Membrane immunofluorescence levelsremained stable for 6 hours, as shown by FAFC analysesperformed at various time intervals after processing ofblood samples. The relative coefficient of variationobtained for measurements of mean immunofluores-cence was 4% for LJ-P4, 5% for anti-LIBSl, and 7% forPAC1. These data demonstrate that under the condi-tions employed, platelets maintain their initial bindinglevels of the specific mAbs.
We also assessed the possibility that either heparin ordextran administered periprocedurally to patients un-dergoing PTCA might interfere with binding of mAbs toplatelets. The binding of mAbs to platelets in peripheralblood samples collected before and after the adminis-tration of heparin and dextran was unchanged. More-over, in vitro additions of heparin and dextran to a levelcomparable to fivefold the estimated in vivo plasmapeak levels showed no effect on binding of mAbs to
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

1482 Arteriosclerosis and Thrombosis Vol 12, No 12 December 1992
resting platelets or platelets stimulated with 10ADP or 1 jtM PMA.
The effects of flow rate on the FAFC analysis ofplatelets were also evaluated in preliminary studies withwhole-blood samples by recording fluorescence inten-sity and forward light scatter at increasing flow rates(Figure 2). At flow rates exceeding 2,000 cells persecond, the number of platelets identified by the posi-tive membrane immunofluorescence and their typicallight-scatter profiles decreased with reciprocal increasesin the number of cells with a light-scatter profile of redblood cells and white blood cells exhibiting similarfluorescence levels. This effect indicated coincidentaldetection of platelets, red blood cells, and white bloodcells at higher flow rates. Similar observations havebeen made by Shattil et al,24 who reported that about5% of the total platelet population may be associatedwith leukocytes by using an anti-GPIb mAb to identifyplatelets. However, since in these experiments flowrates of 10,000 cells per second were used,24 flow-coincidence artifact cannot be excluded.
Detection of Activated Platelets During PTCATo examine whether platelet activation is directly
detectable, five patients undergoing PTCA were ana-lyzed by whole-blood FAFC and mAbs specific foractivation-dependent platelet membrane epitopes. Be-fore angioplasty, platelets bound minimal amounts ofPAC1, S12, 9F9, and anti-LIBSl, similar to those foundin platelets of control patients undergoing catheteriza-tion without PTCA and normal volunteers (Table 1).Moreover, no increased binding of these antibodies wasobserved with platelets in blood obtained by peripheralvenipuncture (data not shown), indicating that theheparin-coated catheter used in this study did notinduce detectable platelet activation. Similarly, plasmalevels of platelet-secreted proteins (/3TG and PF4) andFPA were comparable among specimens of blood ob-tained via catheter from the coronary sinus beforePTCA and peripheral venipuncture (data not shown).Before PTCA, basal plasma levels of /3TG or FPA weredetectably elevated in three patients; a concomitantincrease in plasma /3TG and FPA was observed in onesubject only (patient 3; Table 2). Although bolus hepa-rin transiently mobilizes PF4 from its endothelial bind-ing sites into the circulation over about 5 minutes,44 thesustained elevation in plasma levels of PF4 ranging from19.4 to 72.0 ng/ml found in all five patients probablyreflects platelet activation as opposed to the effect ofheparin.
After angioplasty, activated platelets were identi-fied in the coronary sinus blood obtained from four ofthe five patients, as evidenced by a mean increase inbinding of PAC1 (2.6-fold) and anti-LIBSl (2.7-fold).PTCA was associated with lesser increases in 9F9binding (1.5-fold to twofold); in one patient increasedbinding with S12 was detected. Baseline and peaklevels of the mean platelet membrane immunofluores-cence intensity obtained with the different FITC-conjugated mAbs by one-color flow cytometric analysisare shown for each patient in Table 1. Evaluation ofthe GPIIb-IIIa complex using mAb LJ-P4 that binds toboth resting and activated platelets revealed a con-comitant mean increase of approximately 12% in thefive patients undergoing PTCA. This increase in LJ-P4
binding may reflect a translocation of GPIIb-IIIamolecules from the a-granule membrane to the plate-let surface on activation.45
Mean platelet membrane immunofluorescence inten-sity of binding of the different activation-specific mAbsto platelets in whole blood, plasma levels of /3TG andPF4, and their course in relation to coronary angiogra-phy and angioplasty are depicted in Figure 3. Nocorrelation was found between the expression of activa-tion-dependent platelet epitopes and platelet secretion,as determined by /3TG and PF4 release, as shown inTables 1 and 2 and Figure 3. For example, an approxi-mately 10-fold increase in binding of S12 observed afterPTCA in several blood specimens over a time period of20 minutes was associated with a minor and onlytransient increase in plasma /3TG (Figure 3A). Inanother patient, even at peak levels of plasma /3TG, noS12-positive platelets were detectable in coronary sinusblood samples after PTCA (Figure 3B and Table 1).This discrepancy in results was not obviously attribut-able to concurrently administered medications capableof modifying platelet function, although drug effectscould not be excluded.
Detection of activated circulating platelets afterPTCA, as documented by FAFC analysis, was restrictedto the blood specimens obtained from the coronarysinus. In peripheral venous blood samples that wereobtained and analyzed in parallel, no activated circulat-ing platelets could be identified.
Subpopulations of Activated Platelets andPlatelet-Derived Microparticles
To quantify the relative proportion of activated plate-lets, for each antibody an analysis marker within thefluorescence profile was set at baseline level (beforePTCA) to define a 2% and 10% platelet subpopulationwith the brightest membrane immunofluorescence (Fig-ure 4). This procedure permitted the analysis of adistinctly activated platelet subpopulation for increasesin relative number and fluorescence intensity. In addi-tion, a separate proportion of GPIb-positive micropar-ticles before and after PTCA was quantified.
In the five patients undergoing PTCA, the relativeproportion of platelets positive for PAC1 and anti-LIBSl increased from 2.0±0.3% and 2.0±0.5% at base-line to 18.2±14.2% and 28.0±14.4%, respectively(p<0.01 in both cases). A lesser increase (2.0±0.2% atbaseline to 6.7±2.5%) was observed in the relativeproportion of platelets that had expressed the epitope inplatelet-bound fibrinogen recognized by 9F9 (Figures5A and 5B). Compared with the mean fluorescencelevels obtained with the different mAbs for the totalplatelet population, the analysis of a 2% and 10%subpopulation with the highest fluorescence levels atbaseline did not improve sensitivity for detecting acti-vated platelets but increased the resolution of plateletactivation results. This is illustrated for the individualpatients by the data depicted in Figure 6A, demonstrat-ing quantitative functional changes in membrane GPepitopes of activated platelet subpopulations afterPTCA. The improved resolution of activation obtainedby analysis of platelet subpopulations was also evidentwhen baseline and peak levels of fluorescence intensityfrom all five patients undergoing PTCA were averaged(Figure 6B). For example, by comparing pre-PTCA and
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
Scharf et al Platelet Activation During Angioplasty 1483
2% Panel 10% Panel
PAC1
9F9CO
.52cu
*tr 10* 101 10' 10' 10
ami-UBS1
S12
10* 10' 10' 101 10' 10* 10' 101 10' 104
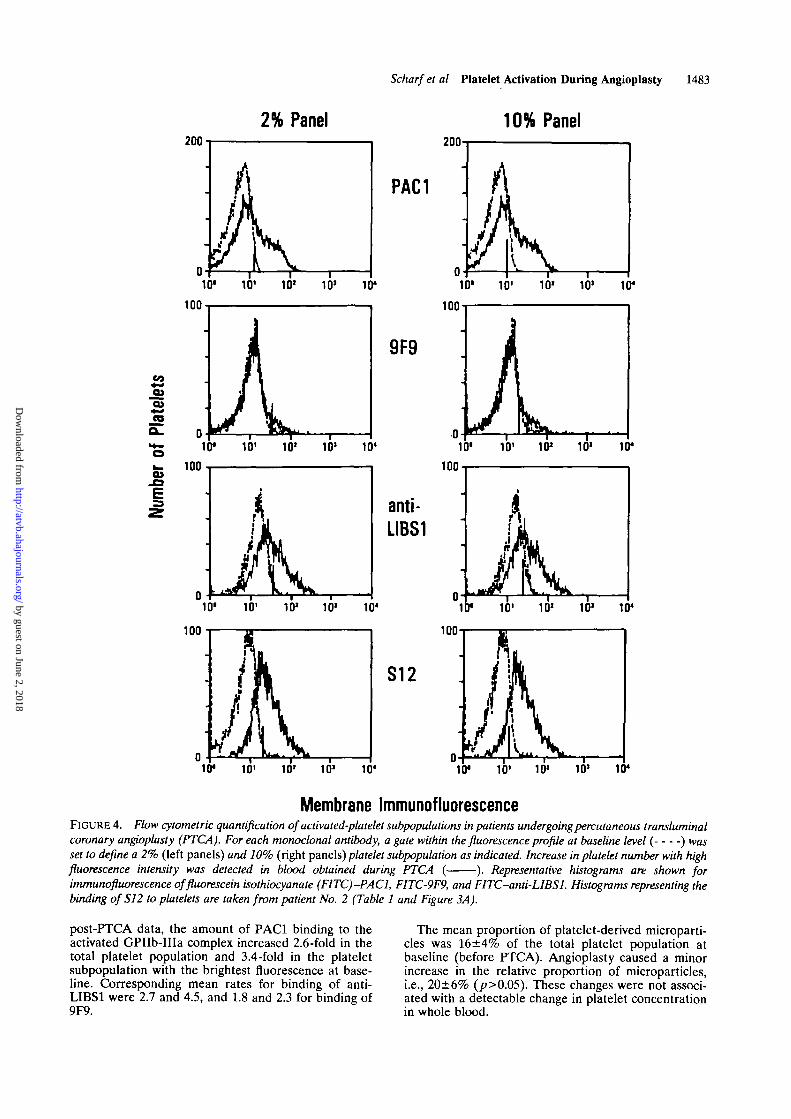
Membrane ImmunofluorescenceFIGURE 4. Wow cytometric quantification of activated-platelet subpopulations in patients undergoing percutaneous transluminalcoronary angioplasty (PTCA). For each monoclonal antibody, a gate within the fluorescence profile at baseline level ( ) wasset to define a 2% (left panels) and 10% (right panels) platelet subpopulation as indicated. Increase in platelet number with highfluorescence intensity was detected in blood obtained during PTCA ( ). Representative histograms are shown forimmunofluorescence offluorescein isothiocyanate (FITC)^ACl, FITC-9F9, and FITC-anti-LIBSl. Histograms representing thebinding of S12 to platelets are taken from patient No. 2 (Table 1 and Figure 3A).
post-PTCA data, the amount of PAC1 binding to theactivated GPIIb-IHa complex increased 2.6-fold in thetotal platelet population and 3.4-fold in the plateletsubpopulation with the brightest fluorescence at base-line. Corresponding mean rates for binding of anti-LIBS1 were 2.7 and 4.5, and 1.8 and 2.3 for binding of9F9.
The mean proportion of platelet-derived microparti-cles was 16±4% of the total platelet population atbaseline (before PTCA). Angioplasty caused a minorincrease in the relative proportion of microparticles,i.e., 20±6% (p>0.05). These changes were not associ-ated with a detectable change in platelet concentrationin whole blood.
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
1484 Arteriosclerosis and Thrombosis Vol 12, No 12 December 1992
Before During
PAC1
Befor* During
9F9
Before During
anti-LIBS1
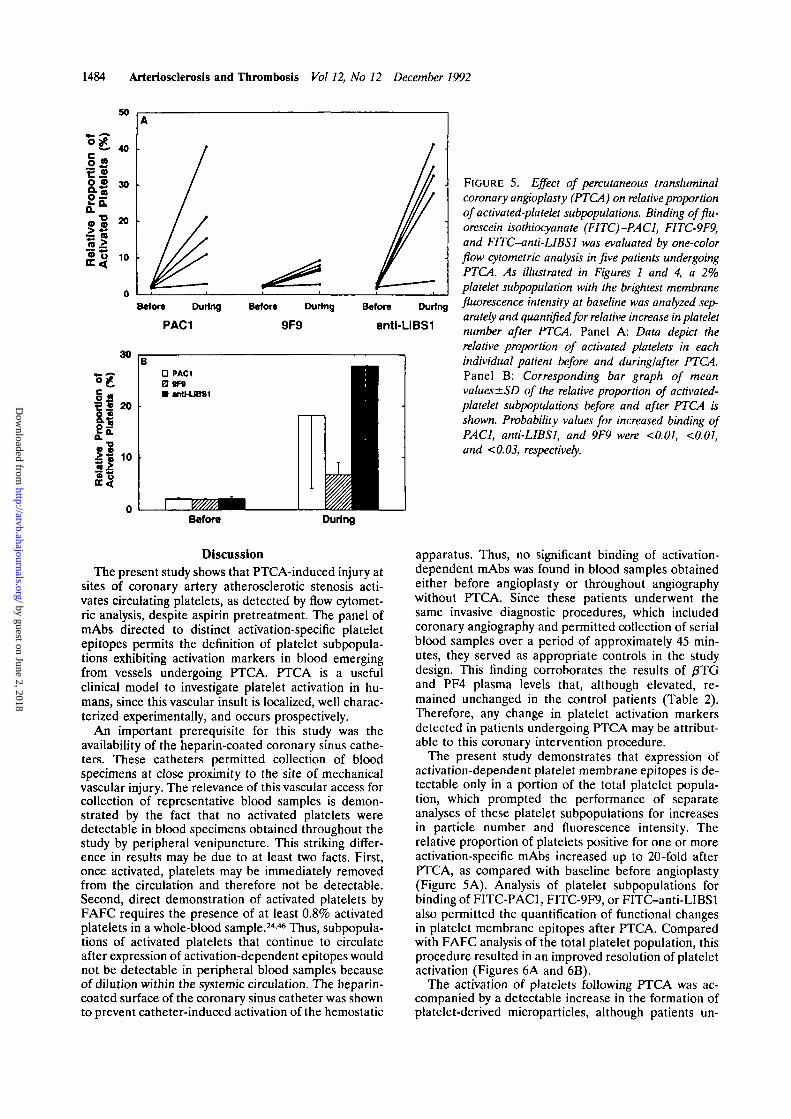
FIGURE 5. Effect of percutaneous transluminalcoronary angioplasty (PTCA) on relative proportionof activated-platelet subpopulations. Binding offlu-orescein isothiocyanate (FITC)-PACl, FITC-9F9,and FITC-anti-LIBSl was evaluated by one-colorflow cytometric analysis in five patients undergoingPTCA. As illustrated in Figures 1 and 4, a 2%platelet subpopulation with the brightest membranefluorescence intensity at baseline was analyzed sep-arately and quantified for relative increase in plateletnumber after PTCA. Panel A: Data depict therelative proportion of activated platelets in eachindividual patient before and during/after PTCA.Panel B: Corresponding bar graph of meanvalues ±SD of the relative proportion of activated-platelet subpopulations before and after PTCA isshown. Probability values for increased binding ofPAC1, anti-LIBSl, and 9F9 were <0.01, <0.01,and <0.03, respectively.
Before During
DiscussionThe present study shows that PTCA-induced injury at
sites of coronary artery atherosclerotic stenosis acti-vates circulating platelets, as detected by flow cytomet-ric analysis, despite aspirin pretreatment. The panel ofmAbs directed to distinct activation-specific plateletepitopes permits the definition of platelet subpopula-tions exhibiting activation markers in blood emergingfrom vessels undergoing PTCA. PTCA is a usefulclinical model to investigate platelet activation in hu-mans, since this vascular insult is localized, well charac-terized experimentally, and occurs prospectively.
An important prerequisite for this study was theavailability of the heparin-coated coronary sinus cathe-ters. These catheters permitted collection of bloodspecimens at close proximity to the site of mechanicalvascular injury. The relevance of this vascular access forcollection of representative blood samples is demon-strated by the fact that no activated platelets weredetectable in blood specimens obtained throughout thestudy by peripheral venipuncture. This striking differ-ence in results may be due to at least two facts. First,once activated, platelets may be immediately removedfrom the circulation and therefore not be detectable.Second, direct demonstration of activated platelets byFAFC requires the presence of at least 0.8% activatedplatelets in a whole-blood sample.244* Thus, subpopula-tions of activated platelets that continue to circulateafter expression of activation-dependent epitopes wouldnot be detectable in peripheral blood samples becauseof dilution within the systemic circulation. The heparin-coated surface of the coronary sinus catheter was shownto prevent catheter-induced activation of the hemostatic
apparatus. Thus, no significant binding of activation-dependent mAbs was found in blood samples obtainedeither before angioplasty or throughout angiographywithout PTCA. Since these patients underwent thesame invasive diagnostic procedures, which includedcoronary angiography and permitted collection of serialblood samples over a period of approximately 45 min-utes, they served as appropriate controls in the studydesign. This finding corroborates the results of /3TGand PF4 plasma levels that, although elevated, re-mained unchanged in the control patients (Table 2).Therefore, any change in platelet activation markersdetected in patients undergoing PTCA may be attribut-able to this coronary intervention procedure.
The present study demonstrates that expression ofactivation-dependent platelet membrane epitopes is de-tectable only in a portion of the total platelet popula-tion, which prompted the performance of separateanalyses of these platelet subpopulations for increasesin particle number and fluorescence intensity. Therelative proportion of platelets positive for one or moreactivation-specific mAbs increased up to 20-fold afterPTCA, as compared with baseline before angioplasty(Figure 5A). Analysis of platelet subpopulations forbinding of FITC-PAC1, FITC-9F9, or FITC-anti-LIBSlalso permitted the quantification of functional changesin platelet membrane epitopes after PTCA. Comparedwith FAFC analysis of the total platelet population, thisprocedure resulted in an improved resolution of plateletactivation (Figures 6A and 6B).
The activation of platelets following PTCA was ac-companied by a detectable increase in the formation ofplatelet-derived microparticles, although patients un-
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Scharf et al Platelet Activation During Angioplasty 1485
Totn PtetetotPopuiMkwi
10% PtateletSubpopulation
2% PlttetM8ubpoputaUon
BD BD BD B D B D BO BD BD BDPACT m M - PACT an • « - PACI «n antb
• Tott Plrtilit PopuUtJona 10% PUttlll SubpopulrtonD2% Ptmirt gubpoputtton
Before During Before During Before During
B PACI 9F9 antWJBSI
FIGURE 6. Effect of percutaneous transluminal coronaryangioplasty (PTCA) on the expression of activation-depen-dent platelet epitopes. Binding of fluorescein isothiocyanate(FITC) -conjugated monoclonal antibodies specific for glyco-protein (GP) Ilb-IIIa (PACI), platelet-bound fibrinogen(9F9), and ligand-induced binding sites on GPIIla (anti-LIBS1) was evaluated in coronary sinus whole-blood samplesfrom five patients before (B) and during/after (D) PTCA andexpressed as log mean fluorescence intensity. Panel A:Changes in fluorescence intensity detected by one-color flowcytometric analysis of the total platelet population are shownfor each patient in the left panel. Baseline and peak levels ofantibody binding are depicted. To increase resolution ofplatelet activation, a 10% and 2% platelet subpopulation(center and right panels) with the brightest fluorescence atbaseline was analyzed separately, as illustrated in Figure 4,and quantified for increase in fluorescence intensity afterPTCA. Panel B: Corresponding mean values of fluorescenceintensity for binding of the different antibodies are shownwhen the total platelet population, a 10%, or a 2% plateletsubpopulation was analyzed. The differences in the totalplatelet population and the 10% and 2% subpopulations werestatistically significant for PACI, 9F9, and anti-LIBSl, eachat p<0.05. Bars indicate SD.
dergoing cardiopulmonary bypass exhibit greater rela-tive increases in platelet-derived microparticles.32 Thisdifference in results may largely reflect the difference inthe magnitude of the stimulus for platelet activationrather than a difference in the mechanism of plateletactivation by the cardiopulmonary bypass apparatus.
The mAbs employed in the present study are directedto distinct activation-dependent epitopes on the plateletsurface and could, therefore, reflect different aspects ofthe platelet activation process. The affinity of the anti-
body for its platelet epitope is another variable that willaffect the amount of binding. After PTCA, binding ofFITC-PAC1, FITC-9F9, and FITC-anti-LIBSl in-creased concordantly in four of the five patients, asevidenced by an increase in mean platelet membraneimmunofluorescence (Table 1 and Figures 3 and 6). Theincrease in binding of these mAbs to the platelet surfaceindicates expression of the activated GPIIb-IIIa com-plex, receptor occupancy by ligands, and expression ofligand-induced binding sites on GPIIla.
Unlike anti-LIBSl and PACI, detection of an epitopeexpressed on platelet-bound fibrinogen turned out to beof lower sensitivity, as evidenced by the minor increaseof activated platelets positive for binding of 9F9 afterPTCA (Figures 5A and 5B). The reason for this findingis unclear at present; however, several possible expla-nations may be considered. First, the affinity of 9F9 toits epitope appears to be significantly lower than that ofPACI or anti-LIBSl.2432 Second, 9F9 specifically reactswith fibrinogen bound to its activated receptor on theplatelet surface. Since binding of fibrinogen to GPIIb-IIIa is initially reversible,19 only a proportion of acti-vated platelets may be recognized by 9F9. Conversely,activated platelets that have irreversibly bound fibrino-gen may interact with each other at the site of vascularinjury to form a platelet aggregate or be removed fromthe circulation. Finally, it is possible that ligands otherthan fibrinogen are bound to the functional receptor. Inany event, an FAFC assay with 9F9 would fail to detectthis fraction of activated platelets.
Increased binding of antibody S12 was obtained inone of five patients only. This antibody is directed to agranule membrane GP, P-selectin (GMP-140), ex-pressed on the platelet membrane surface only aftera-granule release. S12, therefore, allows monitoring ofplatelets that have undergone secretion. The FAFCresults obtained with this antibody appear to be incon-sistent with the increase in plasma /3TG after PTCA(Table 2 and Figure 3A). This discrepancy in resultscannot simply be attributed to the variety of medica-tions, although a drug-related effect cannot be excluded.Absent or inconsistent expression of P-selectin has alsobeen observed in patients undergoing cardiopulmonarybypass,27-32 a clinical setting in which secretion of plate-let-specific proteins has been demonstrated.39 This dis-crepancy may result from the fact that activated plate-lets, which have undergone a-granule release andthereby expressed P-selectin, are only transiently pres-ent in the circulation. Alternatively, redistribution orendocytosis of P-selectin may occur,47 thereby reducingthe accessibility to S12 antibodies.46 If in fact P-selectinis rapidly removed from the platelet surface, then anyassays that are dependent on its detection may haveinsufficient sensitivity. Indeed, it has been suggestedthat plasma assays for secreted platelet-specific proteinsmay be more sensitive than are direct flow cytometricanalyses.32-46
In contrast to our findings, it has recently beenreported that P-selectin-positive platelets were detect-able in peripheral venous and coronary sinus bloodfrom patients undergoing angioplasty.48-49 These obser-vations suggest that platelets that have undergone se-cretion and thereby expressed P-selectin on the plateletsurface may not be rapidly removed from the circula-tion. Interestingly, the highest levels of P-selectin ex-
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

1486 Arteriosclerosis and Thrombosis Vol 12, No 12 December 1992
pression after PTCA were observed in those patientswho developed acute vascular occlusion and/or myocar-dial infarction within 24 hours.48-50 It remains to be seenwhether this platelet activation marker has any predic-tive value with regard to acute thrombotic complicationsafter PTCA. A 1-year follow-up of the five patients ofthe present study revealed that among the four individ-uals in whom activated circulating platelets were detect-able after PTCA, three had no evidence of restenosis atlate angiographic study and one had developed a myo-cardial infarction remote from the procedure. Theremaining patient who had no evidence of activatedplatelets by FAFC analysis was not available for angio-graphic reevaluation.
In summary, the data of this limited study demon-strate that it is feasible to evaluate platelet activation byflow cytometric analysis. By using specific mAbs thatreact with distinct activation-dependent membraneepitopes, platelet subpopulations can be detected andquantified for functional changes in platelet membraneGPs. However, additional observations are needed toevaluate the clinical significance and the therapeuticimplication of this finding.
AcknowledgmentsWe wish to thank Mark Rehse for expert technical assis-
tance; Drs. Andrei Z. Budzynski, Mark H. Ginsberg, RodgerP. McEver, Edward F. Plow, and Sanford J. Shattil forgenerously providing the mAbs used in this study, and Dr.James A. Koziol for statistical advice.
References1. Block PC, Myler RK, Stertzer S, Fallon JT: Morphology after
transluminal angioplasty in human beings. N Engl J Med 1981;305:382-385
2. Baumgartner HR, Muggli R: Adhesion and aggregation: Morpho-logical demonstration and quantitation in vivo and in vitro, inGordon JL (ed): Platelets in Biology and Pathology, volume 2. NewYork, Elsevier, 1976, pp 23-40
3. Majerus PW, Miletich JP, Kane WP, Hoffmann SL, Stanford N,Jackson CM: The formation of thrombin on platelet surface, inMann KG, Taylor FB (eds): The Regulation of Coagulation. NewYork, Elsevier, 1980, pp 215-224
4. Nesheim ME, Hibbard LS, Tracy PB, Bloom JW, Myrmel KH,Mann KA: Participation of factor Va in prothrombinase, in MannKG, Taylor FB (eds): The Regulation of Coagulation. New York,Elsevier, 1980, pp 145-159
5. Wilcox JN, Smith KM, Schwartz S, Gordon D: Localization oftissue factor in the normal vessel wall and in the atheroscleroticplaque. Proc Nad Acad Set U S A 1989;86:2839-2843
6. Harker LA, Mann KG: Thrombosis and fibrinoh/sis, in Fuster V,Verstraete M (eds): Thrombosis in Cardiovascular Disorders. Phil-adelphia, WB Saunders Co, 1992, pp 1-16
7. Holmes DR, Vlietstra RE, Smith HC, Vetrovec GW, Kent KM,Cowley MJ, Faxon DP, Gruentzig AR, Kelsey SF, Detre KM:Restenosis after percutaneous transluminal coronary angioplasty(PTCA): A report from the PTCA registry of the National Heart,Lung and Blood Institute. Am J Cardiol 1984;53:77C-81C
8. Fuster V, Badimon L, Badimon J, Adams PC, Turitto V, ChesebroJH: Drugs interfering with platelet functions: Mechanisms andclinical relevance, in Verstraete M, Vermylen J, Lignen R, ArnoutJ (eds): Thrombosis and Haemostasis. Leuven, Belgium, LeuvenUniversity Press, 1987, pp 349-418
9. Fuster V, Badimon L, Cohen M, Ambrose JA, Badimon JJ, Chese-bro JH: Insights into the pathogenesis of acute ischemic syn-dromes. Circulation 1988;77:1213-1220
10. Fuster V, Badimon L, Badimon JJ, Chesebro JH: Mechanisms ofdisease: The pathogenesis of coronary artery disease and the acutecoronary syndromes. N Engl J Med 1992;326:242-250
11. Wilcox JN, Hanson SR, Rodriguez J, Subramanian R, OllerenshawJ, Zhong C, Hayzer DJ, Lumsden A, Salem T, Kelly AB, HarkerLA, Runge M: Thrombin receptor expression in vascular lesionformation. J Ctin Invest (in press)
12. Ruggeri ZM: Receptor-specific antiplatelet therapy. Circulation1989;80:192O-1922
13. Badimon L, Badimon JJ, Galvez A, Chesebro JH, Fuster V: Influ-ence of arterial wall damage and wall shear rate on platelet depo-sition: Ex vivo study in a swine model. Arteriosclerosis 1986;6:312-320
14. Harmon JT, Jamieson GA: The glycocalicin portion of plateletglycoprotein Ib expresses both high and moderate affinity receptorsites for thrombin: A soluble radioreceptor assay for the interac-tion of thrombin with platelets. J Biol Chem 1986;261:13224-13229
15. Vermylen J, Verstraete M, Fuster V: Role of platelet activationand fibrin formation in thrombogenesis. J Am Coll Cardiol 1986;8:2B-9B
16. Coller BS: Platelets and thrombolytic therapy. N Engl J Med 1990;322:33-42
17. Pytela R, Pierschbacher MD, Ginsberg MH, Plow EF, Ruoslahti E:Platelet membrane glycoprotein Ilb-IIIa: Member of a family ofArg-Gly-Asp-specific adhesion receptors. Science 1986;231:1559-1562
18. Ruggeri ZM: Inhibition of platelet-vessel wall interaction: Plateletreceptors, monoclonal antibodies, and synthetic peptides. Circula-tion 1990;81(suppl I):I-35-I-39
19. Bennett JS, Vilaire G: Exposure of platelet fibrinogen receptors byADP and epinephrine. J Oin Invest 1979;64:1393-1401
20. Shattil SJ: Expression, regulation and detection of fibrinogenreceptors on activated human platelets, in Jamieson GA (ed):Platelet Membrane Receptors: Biochemistry, Immunology andPathology. New York, Alan R Liss, 1988, pp 345-380
21. Shattil SJ, Hoxie JA, Cunningham MC, Brass LF: Changes in theplatelet membrane glycoprotein Ilb-IIIa complex during plateletactivation. J Biol Chem 1985;260:l 1107-11114
22. McEver RP, Martin MN: A monoclonal antibody to a membraneglycoprotein binds only to activated platelets. J Biol Chem 1984;259:9799-9804
23. Nieuwenhuis HK, van Oosterhout JJG, Rozemuller E, van Iwaar-den F, Sixma JJ: Studies with a monoclonal antibody against acti-vated platelets: Evidence that a secreted 53,000 molecular weightlysosome-like granule protein is exposed on the surface of acti-vated platelets in the circulation. Blood 1987;70:838-845
24. Shattil SJ, Cunningham M, Hoxie JA: Detection of activated plate-lets in whole blood using activation-dependent monoclonal anti-bodies and flow cytometry. Blood 1987;70:307-315
25. Adelman B, Michelson HD, Handin RI, Ault KA: Evaluation ofplatelet glycoprotein Ib by fluorescence flow cytometry. Blood1985;66:423-427
26. Jennings LK, Ashmun RA, Wang WC, Dockter ME: Analysis ofhuman platelet glycoproteins Ilb-IIIa and Glanzmann's thrombas-thenia in whole blood by flow cytometry. Blood 1986;68:173-179
27. George JN, Pickett EB, Saucerman S, McEver RP, Kunicki TJ,Kieffer N, Newman PJ: Platelet surface glycoproteins: Studies onresting and activated platelets in patients during adult respiratorydistress syndrome and cardiac surgery. J Clin Invest 1986;78:340-348
28. Block PC: Percutaneous transluminal coronary angioplasty, inEagle KA, Haber E, DeSanctis RW, Austen WG (eds): The Prac-tice of Cardiology Boston, Little Brown, 1989, pp 545-561
29. Trapani-Lombardo V, Hodson E, Roberts JR, Kunicki TJ, Zim-merman TS, Ruggeri ZM: Independent modulation of von Wille-brand factor and fibrinogen binding to the platelet membraneglycoprotein Ilb-IIIa complex as demonstrated by monoclonal anti-body./ Clin Invest 1985;76:1950-1958
30. Ruggeri ZM, Hudson E: Use of murine monoclonal antibodies todetermine platelet membrane glycoprotein structure and function,in Kunicki TJ, George JN (eds): Platelet Immunobiology: Molecularand Clinical Aspects. Philadelphia, JB Uppincott, 1989, pp 235-254
31. Handa M, Titani K, Holland LZ, Roberts JR, Ruggeri ZM: Thevon Willebrand factor-binding domain of platelet membrane gly-coprotein Ib: Characterization by monoclonal antibodies and par-tial amino acid sequence analysis of proteorytic fragments. J BiolChem 1986;261:12579-12585
32. Abrams CS, Ellison N, Budzynski AZ, Shattil SJ: Direct detectionof activated platelets and platelet-derived microparticles inhumans. Blood 1990;75:128-138
33. Frelinger AL III, Cohen I, Plow EF, Smith MA, Roberts J, LamSC, Ginsberg MH: Selective inhibition of integrin function byantibodies specific for ligandoccupied receptor conformers. J BiolChem 1990;265:6346-6352
34. Goding JW: Monoclonal Antibodies: Principles and Practice —Production and Application of Monoclonal Antibodies m Cell Biol-
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Scharf et al Platelet Activation During Angioplasty 1487
ogy, Biochemistry and Immunology. London, Academic Press,1986, pp 255-280
35. Hanson SR, Kotze HF, Savage B, Harker LA: Platelet interactionswith Dacron vascular grafts: A model of acute thrombosis inbaboons. Arteriosclerosis 1985;5:595-603
36. Hanson SR, Harker LA, Bjornsson TO: Effects of platelet-modifying drugs on arterial thromboembolism in baboons: Aspirinpotentiates the antithrombotic actions of dipyndamole andsulfinpyrazone by mechanism(s) independent of platelet cyclooxy-genase inhibition. J Clin Invest 1985;75:1591-1599
37. Scharf RE: Thrombozyten und Mikrozirkulationsstorungen: Kli-nische und experimentelle Untersuchungen zum Sekretionsver-hallen und Arachidonsaurestoffwechsel der BlutplSttchen. (thesis)Stuttgart, FRG, Schattauer, 1986
38. Files JC, Malpass TW, Yee EK, Ritchie JL, Harker LA: Studies ofhuman platelet alpha-granule release in vivo. Blood 1981;58:607-618
39. Harker LA, Malpass TW, Branson HE, Hessel I, Slichter SJ:Mechanism of abnormal bleeding in patients undergoing cardio-pulmonary bypass: Acquired transient platelet dysfunction associ-ated with selective alpha granule release. Blood 1980,56:824-834
40. Scharf RE, Tsamaloukas A, Schneider W: Inhibition of plateleta-granule release in vitro by forskolin. Thromb Res 1984;33:661-664
41. Hanson SR, Harker LA: Baboon models of acute arterial throm-bosis. Thromb Haemost 1987^8:801-805
42. Sims PJ, Faioni EM, Wiedmer T, Shattil SJ: Complement proteinsC5b-9 cause release of membrane vesicles from the platelet surfacethat are enriched in the membrane receptor for coagulation factorVa and express prothrombinase activity. J BkA Chem 1988;263:18205-18212
43. Sims PJ, Wiedmer T, Esmon CT, Weiss HJ, Shattil SJ: Assembly ofthe platelet prothrombinase complex is linked to vesiculation ofthe platelet plasma membrane. J Bid Chem 1989;264:17049-17057
44. Dawes J, Smith RC, Pepper DS: The release, distribution andclearance of human ^-thromboglobulin and platelet factor 4.Thromb Res 1978;12:851-861
45. Niiya K, Hodson E, Bader R, Byers-Ward V, Koziol JA, Plow EF,Ruggeri ZM: Increased surface expression of the membrane gh/-coprotein I Ib/111 a complex induced by platelet activation: Rela-tionship to the binding of fibrinogen and platelet aggregation.Blood 1987;70:475-483
46. Abrams C, Shattil SJ: Immunological detection of activated plate-lets in clinical disorders. Thromb Haemost 1991;65:467-473
47. McEver RP: Leukocyte interactions mediated by selectins. ThrombHaemost 1991;66:80-87
48. Palabrica TM, Smith JJ, Aronovitz MJ, Kimmelstiel D, Golden JS,Scott D, Haik BJ, Salem DN, Konstam MA: Flow cytometricanalysis of platelet PADGEM expression during percutaneoustransluminal coronary angioplasty. (abstract) Circulation 1990;82(suppl III):III-655
49. Nurden AT, Lacaze D, Macchi L, Pintigny D, Durrieu C, Besse P,Sanchez G, Chevaleyre J, Ferrer AM, Vezon G, Hourdille P:Platelet activation in two different clinical conditions: (i) Patientswith severe burns and (ii) after angioplasty in coronary arterydisease, (abstract) Thromb Haemost 1991;65:679
50. Tschoepe D, Schultheiss HP, Kolarov P, Nieuwenhuis K, DannehK, Kehrel B, Strauer B, Gries FA: Platelet activation markers arepredictive for an increased PTCA risk, (abstract) Thromb Haemost1991;65:772
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

R E Scharf, A Tomer, U M Marzec, P S Teirstein, Z M Ruggeri and L A Harkercytometric detection.
Activation of platelets in blood perfusing angioplasty-damaged coronary arteries. Flow
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 1992 American Heart Association, Inc. All rights reserved.
Avenue, Dallas, TX 75231is published by the American Heart Association, 7272 GreenvilleArteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/01.ATV.12.12.14751992;12:1475-1487Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/12/12/1475World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document.Permissions and Rights Question and AnswerFurther information about this process is available in theis being requested is located, click Request Permissions in the middle column of the Web page under Services.Clearance Center, not the Editorial Office. Once the online version of the published article for which permission
can be obtained via RightsLink, a service of the CopyrightArteriosclerosis, Thrombosis, and Vascular Biology Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from





























