Embed Size (px)
Citation preview

ehled metod molekulární biologievyužívaných v patofyziologii

• Molekulárn biologická diagnostikaproteomu, genomu a transkriptomu
• Co umož uje diagnostika na úrovninukleových kyselin
• Principy základních metodik• Praktické poznámky
Molekulárn biologická diagnostika

Molekulárn biologická diagnostika
• Diagnostika na úrovni DNA, RNA aprotein
• Analýza genomu, transkriptomu,proteomu, p íp. metabolomu
• Analýza kvalitativní i kvantitativní

Proteomika
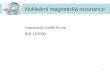
ProteomProteom – poprvé zaveden v roce 1994 Marcem R. Wilkinsempro ozna ení všech protein kódovaných genomem
GenyGeny mRNAmRNA ProteinyProteiny
Bu kaBu ka
Vnit ní parametryVnit ní parametry Vn jší parametryVn jší parametry
StárnutíStárnutíGenetickéGenetickédispozicedispozice Onemocn níOnemocn ní Lé ivaLé iva
ŽŽivotníivotníprost edíprost edí
Genová analýzaGenová analýza•• SNPSNP••MutaceMutace••SekvenaceSekvenace
Analýza expreseAnalýza exprese••mRNAmRNA••ProteinProtein
Analýza interakcíAnalýza interakcí•• ProteinProtein--proteinprotein••AntigenAntigen--protilátkaprotilátka••EnzymEnzym--substrátsubstrát••ProteinProtein--DNADNA••LigandLigand--receptorreceptor

Studium p ítomnosti protein v bu káchStudium p ítomnosti protein v bu kách(analýza(analýza proteomuproteomu))
Metody pro stanovení fyzické p ítomnosti protein :
• polyakrylamidová gelová elektroforéza (PAGE)• westernový p enos• imunoprecipitace• imunohistochemie• izoelektrická fokusace• dvourozm rná elektroforéza v polyakrylamidovém
gelu• chromatografie• fluorescen ní a elektronová mikroskopie• pr toková cytometrie


Nejpoužívan jší techniky - proteiny
• Dvoudimenzionální ELFO• Rentgenová krystalografie• NMR• Hmotnostní spektroskopie• PAGE• Chromatografie (HPLC)• ELISA

Metody využívané sou asnouMetody využívané sou asnou proteomikouproteomikoupro studium proteinpro studium protein
1. Dvoudimenzionální elektroforéza
Nejd íve je vzorek protein rozd len izoelektrickou fokusací zleva dopravaa pak elektroforézou podle své hmotnosti shora dol .

2. Rentgenová krystalografie
Rentgenové zá ení, stejn jako sv tlo, je druh elektromagnetického zá ení o velmi malé vlnovédélce. Jestliže zam íme svazek paralelních rentgenových paprsk na dob e vyvinutý krystal istéhoproteinu, n které paprsky budou rozptýleny ur itým zp sobem atomy v krystalu a projeví sejakour itý obrazec difrak ních skvrn na vhodném detektoru. Poloha a intenzita skvrn v obrazciobsahuje informaci o poloze atom v krystalu proteinu, ze kterého obrazec vznikl.

3. NMR - spektroskopie
Roztok proteinu se umístí do silného magnetického pole, kde je vystaven pulz m rádiové frekvence.Signály, v tomto p ípad z atomových jader vodíku v r zných aminokyselinách lze identifikovat aur it tak vzdálenosti mezi r znými ástmi proteinové molekuly.V ásti (A) je nazna eno dvourozm rné NMR-spektrum odvozené z karboxylového konce enzymucelulázy . Skvrny p edstavují interakce mezi sousedními atomy H. Výsledná struktura odpovídajícírozmíst ní skvrn je v ásti (B).

1 5 5 0 1 5 5 5 1 5 6 0 1 5 6 5 1 5 7 0
0
2 0
4 0
6 0
8 0
1 0 0
1 5 5 7 .7
1 5 5 8 .6
1 5 5 9 .61 5 6 0 .6
1 5 6 3 .8
1 5 6 2 .6
1 5 6 1 .6
Inte
nsity
[%]
M a s s /C h a rg e
Hmotnostní spektroskopie je fyzikáln -chemická metoda pro ur ování hmotnosti molekul a jejichástí. Hmotností spektrometr je iontov optické za ízení, které ze sm si molekul a iont separujenabité ástice podle jejich efektivní hmotnosti m/z (m je hmotnost, z je náboj) a umož uje je
stanovit. Dále poskytuje údaje o relativním zastoupení iont stejné hmotnosti v celkovém množstvíiont ve sm si. Záznam molekulárních a fragmentových iont je charakteristický pro danou látku adává cenné informace o její struktu e a na jeho základ lze v tšinou strukturu látky odvodit nebopotvrdit. Hmotností spektrometrie je metoda citlivá a umož uje analyzovat látky v množství kolem
10 9 g.
4. Hmotnostní spektroskopie

SDS-PAGE(= sodium dodecylsulphate-
polyacrylamide gel electrophoresis)
-metoda pro separaci protein dle jejichvelikosti
5. SDS-PAGE elektroforéza

SDS-Polyacrylamide GelElectrophoresis (SDS-PAGE)
•SDS (Sodium Dodecyl Sulfate) aniontovýdetergent
–solubilizuje a denaturuje proteiny
–dodává protein m negativní náboj
tšina protein váže SDS ve stejnémpom ru, asi 1,4 g SDS/g proteinu
•Zvýšená teplota denaturuje proteiny
O
S
O
O
O
-
CH2
CH2
CH2
CH2
CH2
CH2
CH2
CH2
CH2
CH2
CH2
CH3
SDS

-proteiny jsou tepeln denaturovány vevzorkovém pufru obsahujícím dodecylsulfát sodný (SDS)- zachována je takpouze jejich primární struktura-denaturované proteiny mají vláknitýtvar a stejný záporný náboj danývazbou SDS – migrace tedy závisí pouzena jejich molekulové hmotnosti

Pohyb takto upravených protein velektrickém poli:
-proteiny mající negativní náboj se pohybujísm rem ke kladn nabité elektrod

Jak funguje SDS-PAGE?
•Negativn nabitéproteiny se pohybují kekladné elektrod
•Menší proteiny sepohybují rychleji
•Proteiny se rozd lujípodle velikosti(molekulovéhmotnosti)
-
+
s-s
SDS, zah átí
Proteiny sSDS

Pro používat akrylamidový gel kseparaci protein ?
• Akrylamidový gel: pevná a inertní matrix
• Ideální pro separaci protein
• Menší velikost pór než u agarosy
• Proteiny jsou menší než
intaktní chromoz. DNA
Gel je vytvo en polymerací aktylamidu a N´,N´-methylenbisakrylamidu zahájenou volnýmí radikályvzniklými p i rozkladu persíranu amonného.

Akrylamidový gel
akrylamid
N,N‘ - methylenbisakrylamid

Akrylamidový gel

Separace protein v akrylamidovém gelu

íprava gelu

Denaturované vzorky jsounaneseny na gel

Podmínky proelektroforézu:
•15 min, 50 V
•15 min, 100 V
•1 – 1,5 hod 150 V

Zviditeln ní protein separovaných PAGE
- nespecificky: obarvení všech protein ve vzorku proteinovýmibarvivy- specificky: westernový p enos (detekce specifických proteinprotilátkami)
Nespecifické barvení protein v gelech:- st íbro, „coomassie brilliant blue“
Specifické barvení protein p i westernovém p enosu:- p enos rozd lených protein z gelu na pevný filtr (elektroblotting)- navázání protilátky na p íslušný antigen p i promývání filtru
v roztoku specifické primární protilátky (protilátka musí rozeznatlineární epitop – protein je denaturován)- zviditeln ní protilátky na filtru nap . radioaktivní sondou nebo
sekundární protilátkou konjugovanou s ur itým enzymem

K vizualizaci protein dochází v roztokuCoomasie blue.

Zviditeln ní protein westernovým p enosem

Blotting protein a nukleových kyselinPro další charakterizaci nebo p i mikropreparaci je nutnéextrahovat proteiny i nukleové kyseliny z gelu, kteý jenevhodný pro manipulaci i pro provád ní detek níchchemických reakcí. Je p itom nutné zabránit smíseníseparovaných biomakromolekul.
Nej ast ji se používají techniky tzv. blottingu (angl. blot =skvrna, ka ka, esky p enos) . P i blottingu se pásyseparované elektroforézou p enášejí na ur itou membránu,kde jsou uchyceny adsorpcí nebo kovalentní vazbou. Povazb na membránu je pak možné provád t analýzu
znými zp soby.
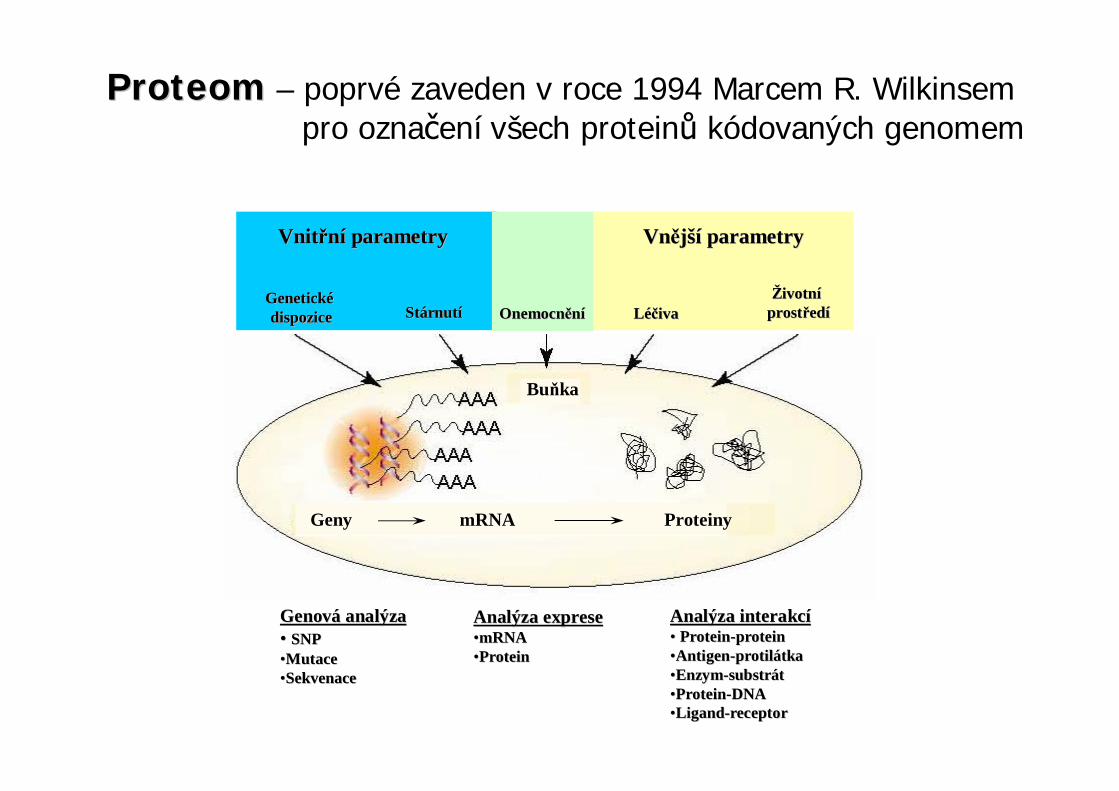
DNA analýza (Southern blotting, Southern 1975)RNA analýza (Northern blotting, Alwine et al., 1977)PROT analýza (Western blotting, Towbin et al., 1979)

Southern blottingDNA je z gelu p enesena na nitrocelulózovou nebonylonovou membránu. ítomnost ur itéjedno et zcové DNA je pak potvrzena hybridizací -vazbou komplementárního DNA et zce (sonda iproba), který je zna en radioaktivn , biotinylací nebovazbou antigenu pro imunodetekci. Použití nap . pro
azení fragment restrik ního št pení genomové DNAjednotlivým gen m.
Northern blottingSeparovaná RNA je p enesena na membránu. Specifickádetekce se d je hybridizací s komplementární DNAsondou, která je zna ena.
Ob metody mají velký význam pro molekulární biologii,ale v sou asné dob jsou ast ji zastupovány PCRtechnikami.
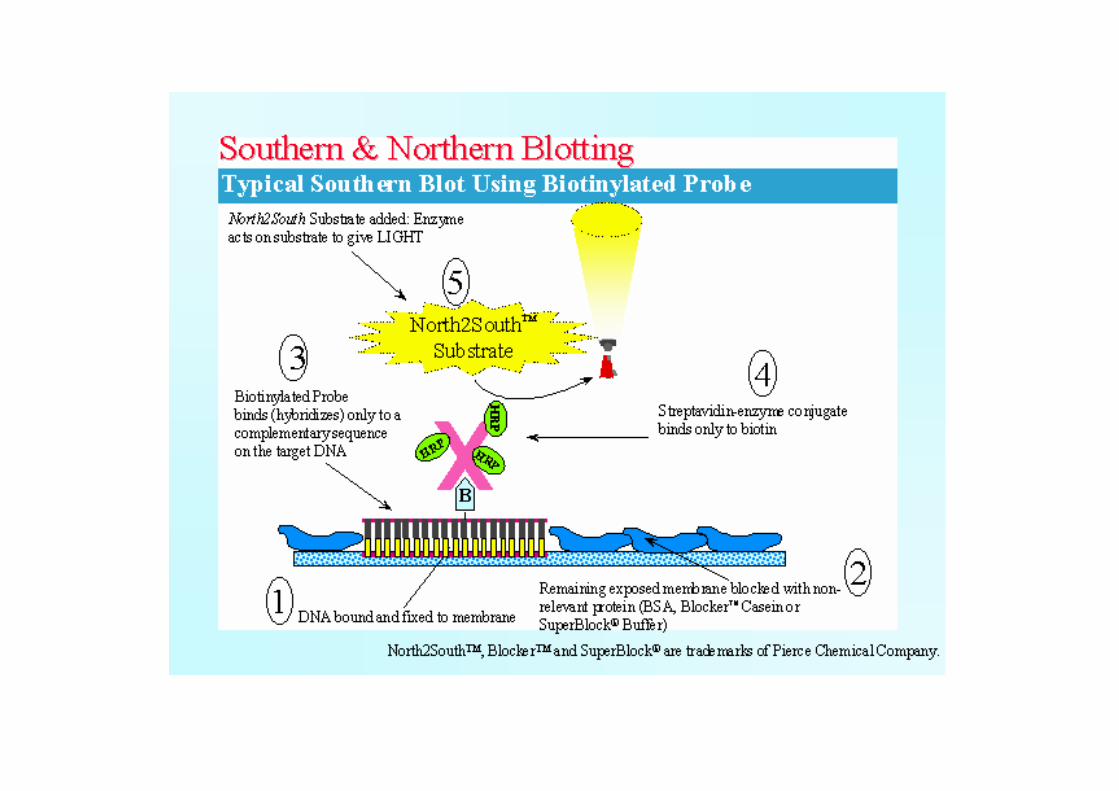

Western blottingProteiny jsou separovány SDS-PAGE a následn p enesenyna nitrocelulózovou nebo polyvinylidendifluoridovou(PVDF) membránu. Barvení na membrán se provádípomocí Ponceau S, Fast Green nebo Amido erni10 B.
Specifická detekce protein se provádí použitímprimárních protilátek; vzniklý imunokomplex se visualizujevazbou zna ené sekundární protilátky nebo zna enéhoproteinu A i G (zna ení nej ast ji vazbou enzymu - nap .alkalické fosfatasy a peroxidasy, ale i koloidním zlatem,radioaktivn , luminiscen i fluorescen . Detekceglykoprotein se d je modifikovanou Schiffovou reakcí,alcianovou mod í, vazbou lektin . N které enzymy lzeprokázat b žnou reak ní sm sí nebo p i blottingu astodochází k renaturaci již denaturovaných bílkovin.


BlokováníVzhledem k tomu, že v tšina používaných detek níchsystém po blottingu obsahuje jako indika ní složkyproteiny, je nutné po vlastním p enosu blokovat zbývajícívazebná místa na membrán , aby zde nedocházelo knespecifické vazb t chto bílkovin. Nesmí však dojít kvyt sn ní vzorku, jeho modifikaci nebo k interferenci sdetek ní reakcí. Používají se: inertní proteiny BSA,kasein, hemoglobin, želatina p íp. nízkotu né mléko.
Nespecifické vazb protein na membránu p i detekcibrání v n kterých aplikacích použití neionogenníchdetergent .
Obdobn je nutné inaktivovat volné reaktivní skupinydiazotovaných membrán a membrán aktivovaných CNBr.To se d lá použitím 10% roztoku ethanolaminu.

Rozd lení metod blottingu podle provedení
Difuzní blotting - prostá difuze v tanku s p enosovým pufrem,dlouhá doba 36-48 hodin, velkou nevýhodou difuze do stran -zhoršení rozlišení vlastní elektroforetické separace.
Kapilární blotting - blotovací membrána umíst na na povrchugelu, který leží na porézní podložce v nádob s p enosovýmpufrem, na ní pak vrstva vlhkého filtra ního papíru a ada vrstevsuchých filtra ních papír . Celá jednotka je zatížena. Suchý papírnasává kapilárními silami pufr, vzorek je tak tažen z gelu namembránu. Trvá zhruba 12 hodin.
Vakuový blotting - obdobné uspo ádání jako u kapilárníhoblottingu, místo kapilárních sil vzorek tažen vakuem. Podstatnrychlejší (30 min). Výhodou též ost ejší pásy (potla ení difuze).

Rozd lení metod blottingu podle provedení
Elektroblotting - nejrychlejší a nejú inn jší metoda. Hnací silouje v tomto p ípad síla elektrického pole. Podle provedení serozlišuje tzv. „tankový (tank, wet)“ blotting a „polosuchý (semi-dry)“ blotting. Jediný pro vysokomolekulární biopolymery.
Hlavními výhodami polosuchého blottingu v porovnání s tankovýmjsou:
1) homogenní elektrické pole, 2) možnost vyššího nap ovéhogradientu , 3) menší spot eba p enosového pufru, 4) možnostsimultánního p enosu z n kolika gel
Volba p enosového pufru je podmínkou pro úsp šný blotting.ležitý pro výb r složení je zejména typ použité membrány, u
elektroblottingu p istupuje ješt pH pufru a iontová síla.

Elektroblotting


Nukleové kyseliny

• Detekce p ítomnosti nukleové kyseliny(detekce p ítomnosti specifické sekvence)
• Analýza struktury (sekvence) nukleovékyseliny
• Kvantifikace nukleové kyseliny sespecifickou sekvencí

Molekulárn biologická diagnostika
• Interdisciplinární– Biochemie– Patologie (in situ diagnostika)– Léka ská genetika– Onkologie– Hematologie– Nukleární medicína

Nejpoužívan jší metodologické postupy
• Detekce p ítomnosti DNA specifické prour itý živo išný druh / bun ný klon
• Detekce jednonukleotidových zám n• Detekce inzercí/delecí• Detekce po tu repetitivních sekvencí• Sekvenace

Požadavky na izolaci nukleových kyselin
• Prvním krokem p i práci s nukleovými kyselinamije jejich izolace v nativním stavu z p irozenéhomateriálu, v dostate ném množství a istot .
• Nukleové kyseliny je t eba zbavit všech látek,které se po lyzi bun k nebo virových ástic stávajísou ástí hrubých lyzát a jejichž p ítomnost bybránila ú innému a specifickému p sobeníenzym používaných k jejich analýze a úpravám.

Materiál pro izolaci nukleových kyselin
• Výchozím materiálem mohou být:– Jednotlivé bu ky (nap . prokaryotických organizm
nebo kvasinek)• Genomová• Plazmidová
– Tkán a orgány eukaryot, které jsou nejd ívehomogenizovány
– Virové ástice purifikované centrifuga nímitechnikami
– Nukleové kyseliny v elektroforetických gelech– Produkty PCR reakcí

Metodické principy využívané p i izolacinukleových kyselin
• Rozrušení bun ných st n nebo virovýchnukleokapsid sobením– Enzym (lysozym a celulázy)– Detergent (dodecylsulfát sodný)
• Enzymatické kroky pro odstran ní kontaminant– Proteináza K– RNáza nebo DNáza
• Kroky využívající r znou rozpustnost– Fenolová extrakce– Adsorpce na pevný podklad
• Centrifugace• Precipitace

Typy metod pro izolaci nukleových kyselin
1. Metody využívající rozdílné rozpustnosti• Zahrnují fenolové extrakce a etanolové nebo
izopropanolové precipitace (srážení)• Obecn rozší ené, široké aplikace• Vhodné pro vysokomolekulární genomové NK
2. Metody adsorp ní• DNA se váže na k emi ité sklo v p ítomnosti
chaotropní látky• Vhodné zejména pro rychlou purifikaci plazmid a
malých fragment3. Centrifugace v hustotním gradientu
• Izopyknické centrifugace v gradientu CsCl• Vhodné p i velkém množství a pro vysokou istotu• Možnost frakcionace podle velikosti v sacharózových
gradientech

Typická fenolová extrakce• Promíchání lyzátu bun k s roztokem fenolu, p ípadn se sm sí
fenolu a chloroformu. Fenol je organické rozpoušt dlopoužívané k odd lení protein od nukleových kyselin.Proteiny jsou hydrofobní a z stávají v organické fázi, zatímco NKjsou vysoce nabité a p echázejí do vodné fáze. Chloroformdenaturuje proteiny, rozpouští tuky a napomáhá odd leníjednotlivých fází získaných v následujícím kroku.
• Centrifugace, p i níž dojde k odd lení spodní organické fáze,tvo ené fenolem (p ípadn sm sí fenolu a chloroformu), mezifáze,tvo ené denaturovanými proteiny a zbytky bun k, a horní vodnéfáze, v níž jsou rozpušt ny nukleové kyseliny.
• Vysrážení nukleových kyselin etanolem,ípadn izopropanolem.innému vysrážení nukleových kyselinítomných v nízkých koncentracích se
napomáhá snížením teploty a p ídavkem solí.• Shromážd ní precipitátu nukleových kyselin
centrifugací a rozpušt ní získaného sedimentu ve vhodném roztoku.

Izolace RNA• Izolace RNA fenolovou extrakcí je podobná
izolaci DNA s následujícími rozdíly:– Inhibitory RNáz, sterilní boxy, rukavice!– Extrakce v guanidinových solích– Fenolové extrakce p i pH 5-6– Odstran ní DNA DNázami bez RNáz– Selektivní srážení RNA pomocí LiCl– Afinitní chromatografie na oligo-dT kolonách pro
izolaci mRNA

Adsorp ní metody• Využívají schopnost nukleových kyselin adsorbovat se na
povrchy z oxidu k emi itého (kolonky do centrifuga níchmikrozkumavek) v p ítomnosti chaotropní soli (Vogelsteinand Gillespie, 1979)– jodid sodný (NaI)– guanidin thiokyanát– guanidin hydrochlorid
• Síla vazby závisí na– Typu nukleové kyseliny (DNA nebo RNA)– Iontové síle– pH roztoku
• Promývání pro odstran ní protein a dalších kontaminant• Eluce DNA pufrem s nízkou koncentrací solí nebo H2O• Rychlá metoda, vysoký výt žek (malé fragmenty), vysoká
istota

Mechanismus interakce DNA s oxidememi itým

Izolace mRNA• mRNA tvo í pouze malý podíl z celkové RNA,
proto je její izolace obtížná• Pro izolaci se využívají
– Tradi ní metody, kdy se nejprve izoluje celkováRNA, která je následn separovaná na mRNA, rRNAa tRNA
– Metody využívající afinitu poly(A) konce u mRNA abiotinem zna ené oligo(dT) sondy• Sonda se v lyzátu selektivn váže na mRNA, aniž by
interagovala s DNA nebo jinými RNA• Hybridní molekuly biotinylované dT-A mRNA jsou
imobilizovány na pevném podkladu pokrytémstreptavidinem
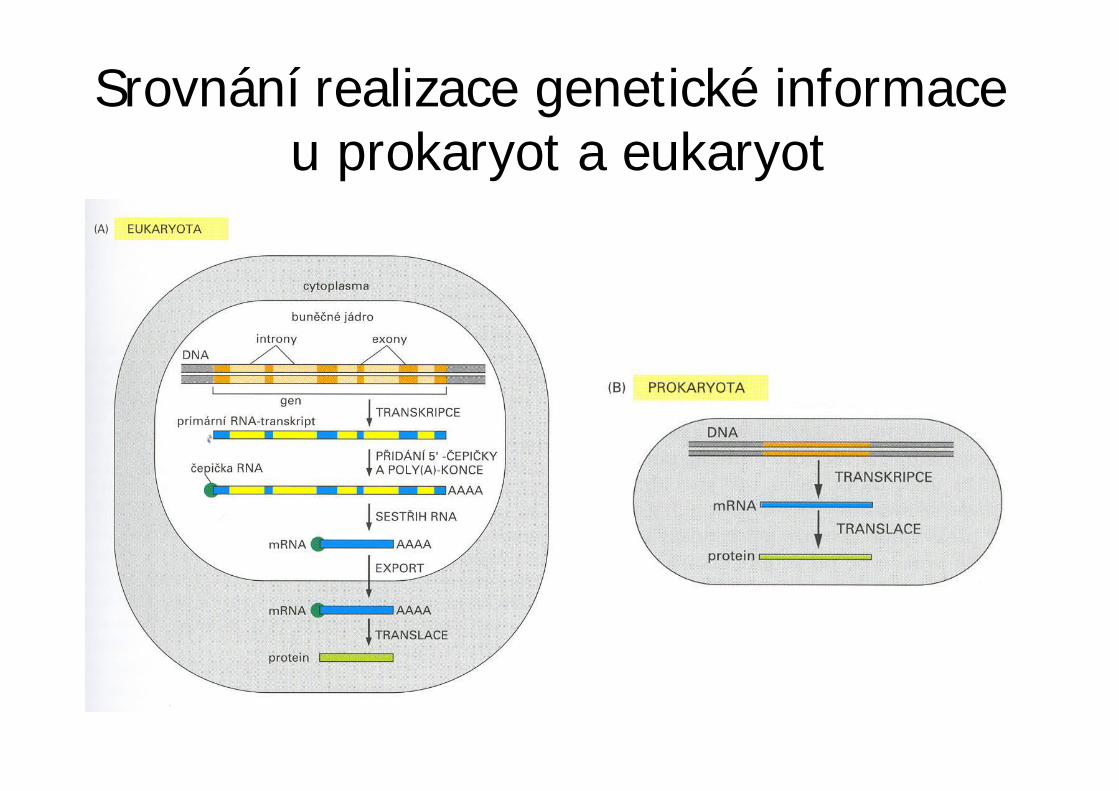
Srovnání realizace genetické informaceu prokaryot a eukaryot

Analýza a kvantifikace• SPEKTROFOTOMETRICKÁ METODA
– Je to vhodná metoda pro m ení vzork , které jsou dostate isté bezvýznamného množství kontaminant.
– Nukleové kyseliny absorbují UV zá ení s maximem absorbance v oblasti vlnovédélky okolo 260 nm.
– Z hodnot optické hustoty lze koncentraci a istotuvzorku stanovit podle empirických vztah . Stupeistoty nukleových kyselin se stanovuje z pom ru
absorbance p i 260 a 280 nm. P i zne išt ní vzorkuproteiny, jejichž absorp ní maximum leží okolo280 nm, bude vypo tený pom r výrazn nižší astanovení koncentrace NK nebude p esné.
• FLUORESCEN NÍ METODA– Je vhodná u vzork s nízkou koncentrací nukleových kyselin nebo zne išt ných
ítomností jiných látek.– Využívá se fluorescence, k níž dochází po ozá ení komplexu etidiumbromidu
navázaného na DNA UV sv tlem. Vzniká viditelné ervenooranžové sv tlo ovlnové délce 590 nm, jehož intenzitu lze srovnat s intenzitou standardu oznámé koncentraci bu vizuáln , nebo po po ízení fotografického záznamu.Stanovení se provádí na agarózových gelech obsahujících nízkou koncentracietidiumbromidu, na n ž se testované vzorky bu nakapou, nebo podrobíelektroforéze, která umožní áste nou purifikaci nukleových kyselin.

Nejpoužívan jší techniky - NK
• Elektroforéza• Restrik ní št pení• Klonování• Sekvenace• PCR• RFLP• Chromatografie (HPLC)• Hmotnostní spektroskopie

lící techniky nukleových kyselin
• Metody– Centrifuga ní– Elektromigra ní– Chromatografické
• Vlastnostivyužívané pro
leníbiomakromolekul– Molekulová
hmotnost– Konformace a tvar– Náboj– Hustota

Metody elektroforetické
• Princip– Pohyb nabitých molekul nukleových kyselin
v elektrickém poli– Hlavním nositelem náboje nukleových
kyselin jsou záporn nabité fosfátovéskupiny
• Cíl– D lení molekul na základ rozdílných
elektroforetických pohyblivostí
Elektroforéza pat í v molekulární biologii knejpoužívan jím separa ním technikám p i izolaci a
analýze nukleových kyselin a protein

Gelová elektroforéza• Základ
• Z praktických d vod se elektroforéza neprovádí p ímo v roztoku, ale navhodném nosi i
• V p ípad nukleových kyselin bývá nosi em nej ast ji gel• Pro p ípravu gelu jsou výhodné látky, které samy gelují a mají r znou
porozitu v závislosti na koncentraci gelu
• Výhody• Elektroforetické metody nahradily ultracentrifugaci• Aparatury jsou levné, asto je lze vyráb t svépomocí• D lit lze všechny d ležité biomakromolekuly• Zm nou podmínek lze d lit podle r zných hledisek• Rychlost• Lze pracovat s mikrokvanty nebo preparativn v mikrogramových
množstvích• Rozd lené molekuly lze snadno prokázat a ve funk ní form izolovat
z gelu

Používané nosi e• Elektroforetické gely používané pro separaci
nukleových kyselin jsou nej ast ji tvo eny– agarózou– polyakrylamidem
• Vytvá ejí složitou sí ovou strukturu polymerníchmolekul s póry
• Velikost pór lze ovlivnit složením roztoku akoncentrací polymeru
• Optimální velikost separovaných molekul– agarózové gely 100 bp až 50 000 bp– polyakrylamidové 10 až 1000 bp
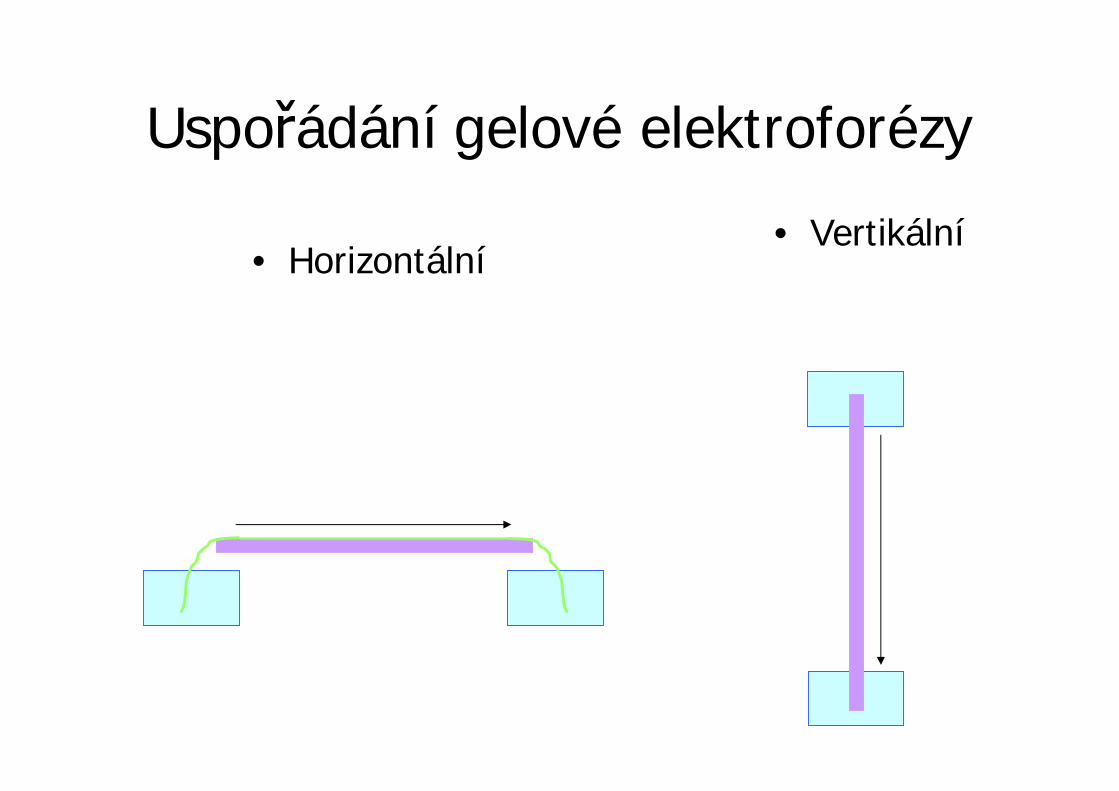
Uspo ádání gelové elektroforézy
• Horizontální• Vertikální
ì
î
ìî
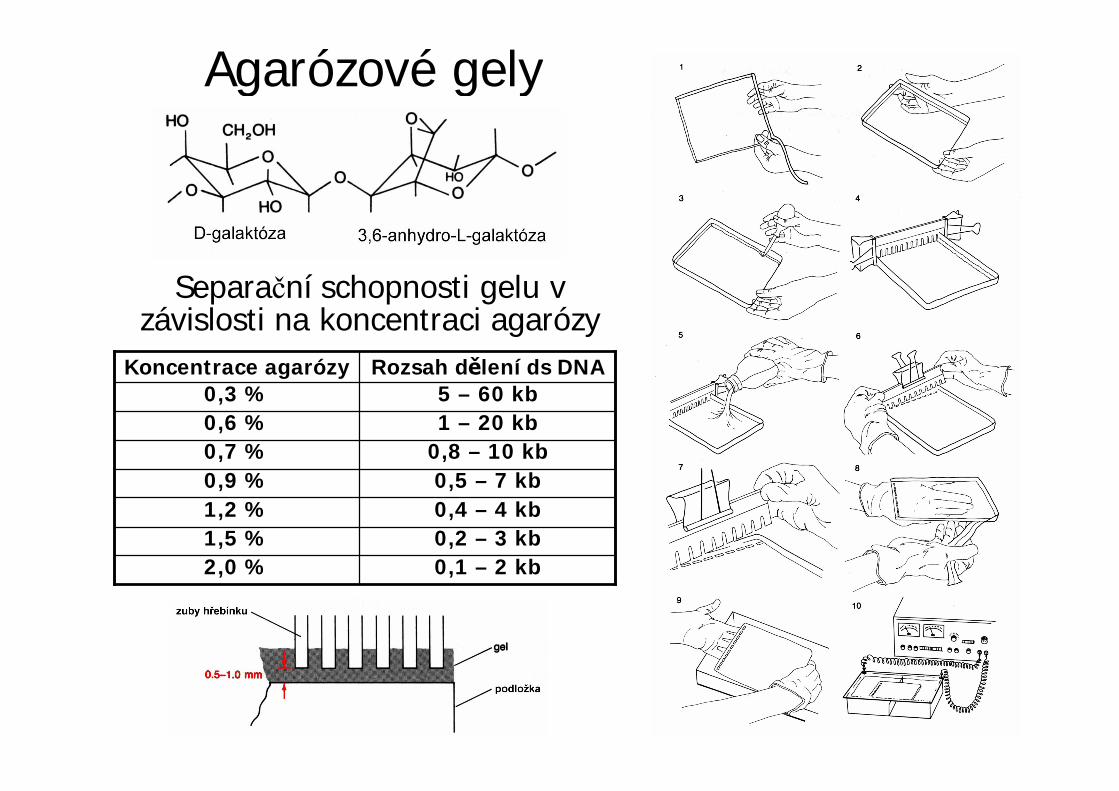
Agarózové gely
0,1 – 2 kb2,0 %0,2 – 3 kb1,5 %0,4 – 4 kb1,2 %0,5 – 7 kb0,9 %
0,8 – 10 kb0,7 %1 – 20 kb0,6 %5 – 60 kb0,3 %
Rozsah d lení ds DNAKoncentrace agarózy
Separa ní schopnosti gelu vzávislosti na koncentraci agarózy

íprava agarózového gelu

Provedení elektroforézy

Test zm ny pohyblivosti v gelu• Gelová elektroforéza je rovn ž vhodným
prost edkem pro studium interakcí mezinukleovými kyselinami a proteiny,zejména t mi, které se vážou naspecifické sekvence DNA– transkrip ní faktory– represory
• Metoda se ozna uje jako test zm nypohyblivosti v gelu. Vychází z poznatku,že po vazb protein na DNA se jejíelektroforetická pohyblivost obvyklesníží.
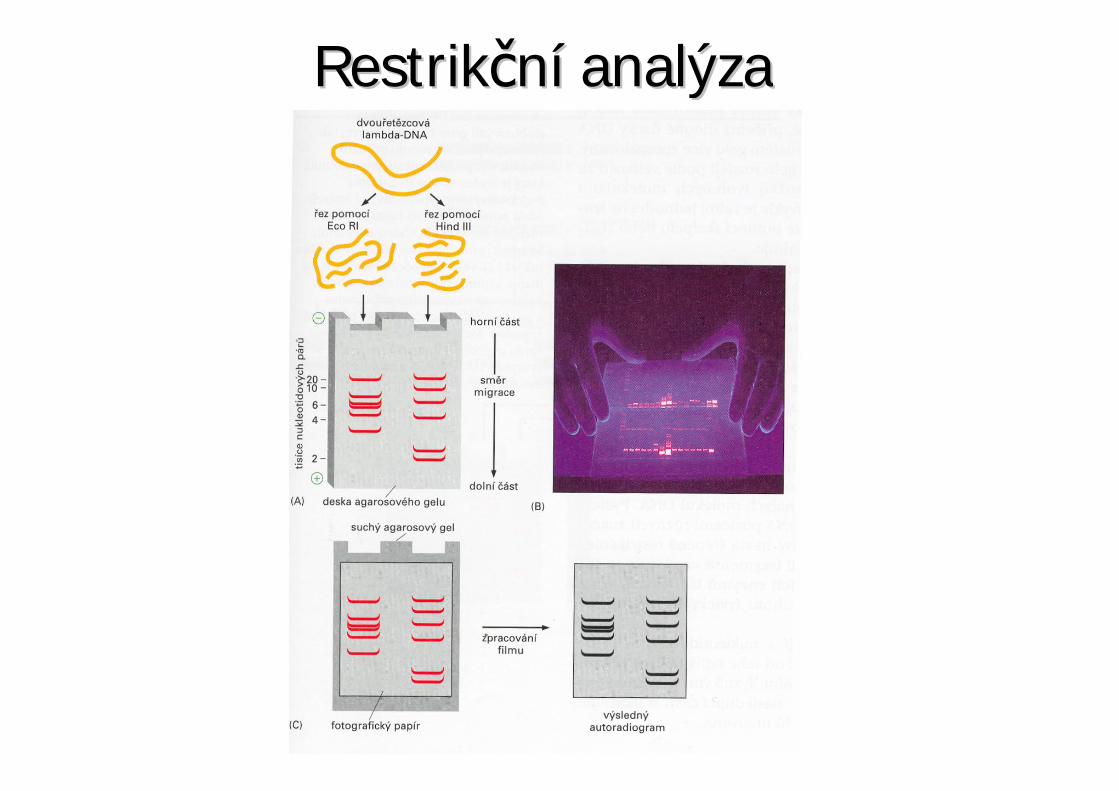
Restrik ní analýzaRestrik ní analýza

Sekvenování DNA

Klonování DNA

PCR
1983 Kary Mullis1993 Nobelova cena
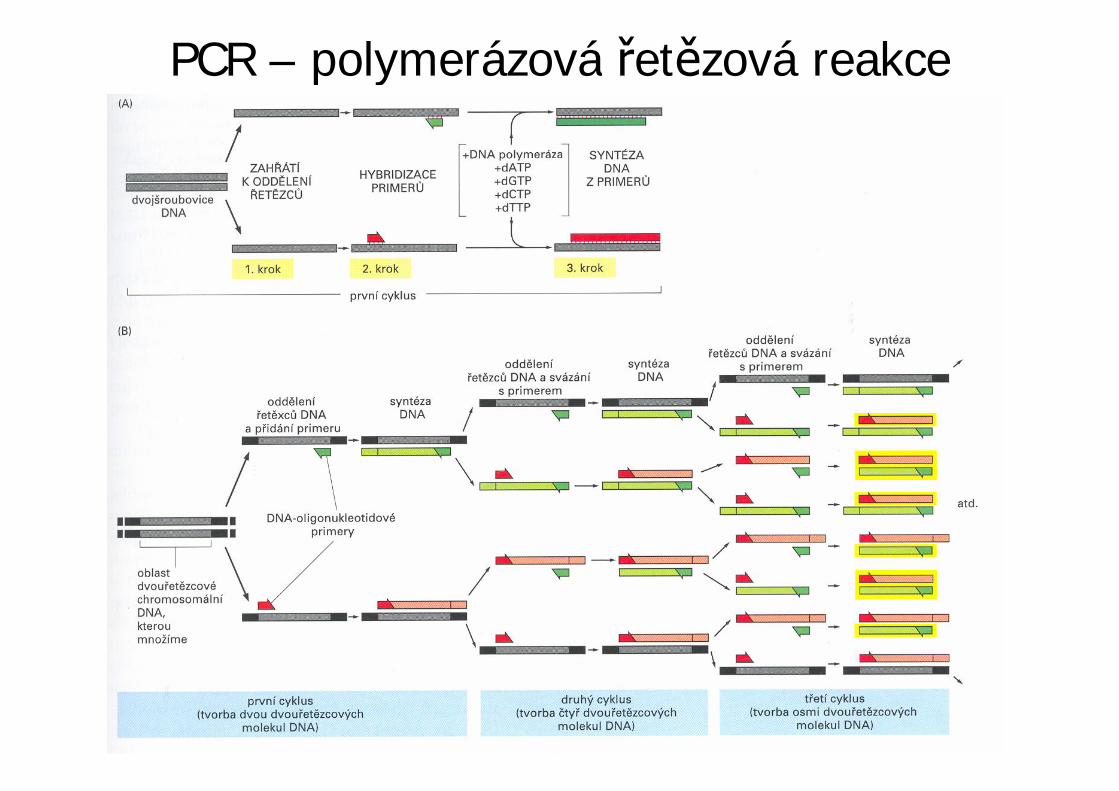
PCR – polymerázová et zová reakce

Základní komponenty reakce
• voda• pufr• dNTP (dATP+dTTP+dCTP+dGTP)• MgCl2• Taq polymeráza
• primers• templát

Princip PCR
• Metoda pro mnohonásobné zmnožení (amplifikaci)specifického úseku DNA in vitro založená na principureplikace.
• K opakující se enzymové syntéze komplementárníchet zc vybraných úsek dvou et zcové DNA dochází
po p ipojení dvou primer vázajících se na protilehléet zce DNA tak, že jejich 3'-OH-konce sm ují proti
sob .• PCR umož uje získat požadovanou zcela specifickou
sekvenci bez klonování.

• PCR využívá základních rys replikace(enzymatické syntézy) DNA:– Jako templát slouží ssDNA, podle níž je syntetizován
komplementární et zec.– K zahájení reakce je zapot ebí primer, který se
ipojuje na komplementární úseky DNA. Tím jezárove vymezen úsek DNA, který bude amplifikován.
– Jako templáty pro syntézu mohou sloužit oba et zcedsDNA, po p edchozí denaturaci.
• Primery se vybírají tak, aby se p ipojovaly kmíst m ohrani ujícím z obou stran amplifikovanýúsek.
• Teoreticky lze získat 2n et zc (kopií).


Teplotní režim
• 96ºC Iniciální denaturace• 96ºC denaturace• 40-72ºC annealing• 72ºC elongace• 72ºC záv re ná elongace• 4ºC zchlazení
30x

Replikace DNA in vivo vyžadujemnoho enzym

Replikace DNA in vitro vyžaduje pouzejeden enzym
• Reak ní sm s obsahuje:– Templátovou nukleovou kyselinu (DNA nebo RNA).
• Množství DNA jako výchozího materiálu je velmi nízké: obvykleposta uje mén než 1 mg genomové DNA, teoreticky posta uje jednamolekula.
– Kvalita templátu ovliv uje výsledek PCR.– Velké množství RNA m že vázat ionty Mg2+– zne išt ný templát m že obsahovat inhibitory PCR
• Zdroj: mikroorganizmy, bu ky z tká ových kultur, t lní tekutiny,bioptické vzorky, st ry, vlasy, atd…
• Primery. Chemicky syntetické ologonukleotidy o velikost10 - 30 nukleotid .
• dNTP ve form Na+ nebo Li+ solí• Mg2+ ionty tvo í rozpustný komplex s dNTP a vytvá ejí
substrát, který rozpoznává DNA polymeráza.• Termostabilní DNA polymerázu, která odolává
teplotám až 98 °C.

Reak ní podmínky PCR1. Po áte ní denaturace DNA. D ležitá je kompletní denaturace templátu,
obvykle posta uje zah átí sm si na 2 – 5 min / 95 °C. V p ípad , že dojdepouze k áste né denaturaci, molekuly DNA velice rychle renaturují a to vede knespecifické vazb primer („self-priming“) a možným falešným výsledk m.
Vlastní et zová reakce:2. Denatura ní krok (separace et zc ) : 94 – 95 °C / 20 – 45 s, záleží na objemu
reakce, tlouš ce st n zkumavek. Nedostate denaturovaná DNA neumož ujeístup primer m, naopak p íliš dlouhá denaturace snižuje aktivitu DNA polymerázy
(stabilní cca 2 hod / 98 °C).3. P ipojení primer (55 - 65 °C / 30 – 90 s) teplota ur uje specifi nost a závisí na Tm
primeru a templátu. Pro oba primery by m la být Tm podobná. Ta se optimalizuje vteplotním gradientu nebo se stanovuje empiricky.
4. Prodlužování primeru (polymera ní reakce) p i standardní PCR probíhá p i 72 °C/45 – 90 s. Taq DNA polymeráza syntetizuje DNA p i této teplot rychlostí cca 60bází/s. Nov syntetizované et zce slouží jako templáty pro další cyklus.
Cyklické st ídání teplot jednou založené sm si zajiš uje pr h reakce PCR, provádí se vtermocyklerech, v tšinou se provádí 25 - 35 cykl .
5. Záv re ná extenze se provádí obvykle po posledním cyklu (72°C / 5 min) aslouží k dokon ení syntézy a renaturaci jedno et zcových produkt .

Návrh primer pro standardní PCR
– 18 – 25 bází dlouhé
– neobsahují vnit ní sekundární struktury
– obsahují 40 – 60 % G+C
– mají rovnom rnou distribuci oblastí bohatých na G/C aA/T páry
– nejsou komplementární navzájem na 3’-koncích, takženevytvá ejí navzájem nebo samy se sebou duplexy
– na matricové DNA nemají falešná vazebná místa
– mají Tm teplotu 55 – 65 °C

Detekce amplifikovaného produktu PCR1. Stanovení fyzikální velikosti produktu
gelovou elektroforézou (agaróza,polyakrylamid). Detekce obarvením apozorování pod UV sv tlem.
2. Št pení produktu restrik ními enzymya posouzení spektra vznikajícíchrestrik ních fragment (PCR-RFLP)
3. Elektroforetická separace produkt zaspecifických podmínek pro detekcisekven ních polymorfizm– DGGE – denatura ní gradientová gelová
elektroforéza– SSCP – analýza polymorfizmu konformace
jedno et zcových forem
íklad standardnígelové elektroforézys produkty PCRv agarózovém gelu.
1 = hmotnostní st.2 - 4 = produkty PCR

4. hybridizací s neradioaktivn zna enou sondoukomplementární k ásti sekvence amplifikovanéhoúseku a detekcí enzymoimunoanalýzou vmikrotitra ní desti ce (PCR-EIA/ELISA),hybridiza ní sonda m že být následnamplifikována (PCR-OLA)
5. imobilizací PCR-produkt na membrán a te kovouhybridizací se zna enými alelov -specifickýmioligonukleotidy (ASO) nebo zp tnou te kovouhybridizací s r znými ASO imobilizovanými namembrán ,
6. hybridizací na DNA- ipech,7. stanovením sekvence DNA.

Elektroforéza v agarovém gelu

+
-

PCR-RFLP• Amplifikace známé sekvence se dv ma
specifickými primery– Cílová sekvence (obvykle ur itého genu)
o délce 1 až 2 kb je amplifokována p ivysoce stringentních podmínkách.
– Výsledkem amplifikace jsou amplikony(PCR produkty o stejné délce) detekovanéelektroforeticky
• Amplikony jsou št peny restrik níendonukleázou se 4 bp rozpoznávacímmístem a poté op t analyzoványpomocí elektroforézy
• Separace fragment DNA vagarózovém nebo polyakralamidovémgelu.
• Srovnání restrik ních fragmentamplifikované DNA u r zných vzork .

Využití metody PCR1. Základní výzkum
– izolace gen nebo jejich ástí– sekvencování DNA– mutageneza in vitro– modifkace konc DNA– analýza (selekce) klon z
genových knihoven– p íprava zna ených sond
2. Aplikovaný genetickývýzkum– prenatální diagnostika
(d di ných chorob)– detekce mutací v genech– studium polymorfizmu gen
(nap . RAPD)– popula ní genetika
3. Využití v klinickýchdisciplinách– detekce patogenních
mikroorganism (baktérií,vir , prvok , hub)
– identifikace onkogen– typizace nádor– stanovení pohlaví
4. Využití v praxi– archeologie– soudnictví– kriminalistika

Praktická ást –detekce I/D polymorfismu v genu
pro ACE

Inzer /dele ní polymorfismus v genu pro angiotensinkonvertující enzym je d ležitým genetickým markeremvzniku hypertenze a ischemické choroby srde ní.
Hypertenze je jedním z nej ast jších onemocn ní, kterése nemusí zejména zpo átku projevovat žádnýminápadnými p íznaky.
Pozd ji se ale mohou dostavit závažné komplikace:srde ní selhání, urychlení aterosklerózy, ischemická chorobasrde ní (ICHS), cévní mozkové p íhody, postižení ledvin

Jedním z genetických aspekt vzniku hypertenze je polymorfismusangiotensin konvertujícího enzymu (ACE).

ACE je lokalizován v membránách cév hlavn v plicích a št píangiotensin I na angiotensin II, který zp sobuje zužování cév(vazokonstrikce).
Hladina ACE je závislá na p ítomnosti inzerce nebo delece v genuACE.
Genotyp D/D je spojen s vyšší hladinou ACE v séru a rizikemvzniku ischemické choroby srde ní.
Indikace k vyšet ení· Vysoký krevní tlak· Srde ní selhání· Cévní mozkové p íhody· Renální selhání