Embed Size (px)
Citation preview

J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te / j p ro t
Venomic analysis and evaluation of antivenom cross-reactivityof South American Micrurus species
Paula H.C. Ciscottoa, b, Breno Ratesa, b, Daniel A.F. Silvaa, b, Michael Richardsonc,Luciano P. Silvad, Helida Andradee, Micheline F. Donatof, Giselle Agostini Cottac,Wany Selena Mariac, Raquel J. Rodriguesc, Eladio Sanchezc,Maria Elena De Limaa, b, Adriano M.C. Pimentaa, b,⁎aLaboratório de Venenos e Toxinas Animais, BrazilbNúcleo de Estudo de Estrutura e Função de Biomoléculas, Departamento de Bioquímica e Imunologia, Instituto de Ciências Biológicas,Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, BrazilcCentro de Pesquisa Professor Carlos R. Diniz, Divisão de Ciências Biomédicas, Fundação Ezequiel Dias, Belo Horizonte, Minas Gerais, BrazildLaboratório de Espectrometria de Massa—LEM, EMBRAPA Recursos Genéticos e Biotecnologia, Brasília, Distrito Federal, BrazileDepartamento de Parasitologia, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, BrazilfBiologia Molecular, Laboratório de Tecnologia Farmacêutica, Universidade Federal da Paraíba, Ceará, Brazil
A R T I C L E I N F O
⁎ Corresponding author at: Laboratório de VenCarlos, 6627, 31270-901, Belo Horizonte, Mina
E-mail address: [email protected] (A
1874-3919/$ – see front matter © 2011 Elsevidoi:10.1016/j.jprot.2011.07.011
A B S T R A C T
Available online 23 July 2011
Coral snakes from Micrurus genus are the main representatives of the Elapidae family inSouth America. However, biochemical and pharmacological features regarding their venomconstituents remain poorly investigated. Here, venomic analyses were carried out aiming ata deeper understanding on the composition of M. frontalis, M. ibiboboca, and M. lemniscatusvenoms. In the three venoms investigated, proteins ranging from 6 to 8 kDa (3FTx) and 12 to14 kDa (PLA2) were found to be the most abundant. Also, the N-terminal sequences of fournew proteins, purified from theM. lemniscatus venom, similar to 3FTx, PLA2 and Kunitz-typeprotease inhibitor from other Micrurus and elapid venoms are reported. Cross-reactivityamong differentMicrurus venoms and homologous or heterologous antivenoms was carriedout by means of 2D-electrophoresis and immunoblotting. As, expected, the heterologousanti-Elapid venom displayed the highest degree of cross-reactivity. Conversely, anti-M.corallinus reacted weakly against the tested venoms. In gel digestions, followed by massspectrometry sequencing and similarity searching, revealed themost immunogenic proteinfamilies as similar to short and long neurotoxins, weak neurotoxins, PLA2, β-bungarotoxin,venom protein E2, frontoxin III, LAO and C-type lectin. The implications of our results for theproduction of Micrurus antivenoms are discussed.© 2011 Elsevier B.V. All rights reserved.
Keywords:Snake venomsMicrurusVenomicsAntivenomCross-reactivity antivenom
1. Introduction
Available epidemiological data concerning envenoming bysnakebites estimate the occurrence of about 5.4 millionaccidents per year, with more than 125,000 deaths worldwide
enos e Toxinas Animais, Departamento de Bioquímica e Imunologia, ICB, UFMG, Av. Antônio
s Gerais, Brazil. Tel.: +55.M.C. Pimenta).er B.V. All rights reserved
[1]. In Brazil, snakebite accidents are considered a public healthproblem and present considerablemedical importance becauseof their high frequency and severity. The latest data released bythe Brazilian Ministry of Health reports that in the year of 2003,25,478 cases of snakes bites envenoming related to the genus of
31 3409 2662.
.

1811J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
higher epidemiological importance, i.e., Bothrops, Crotalus,Lachesis and Micrurus. Although the higher rate of accidents isattributed to Bothrops and Crotalus (87.5% and 9.2%, respective-ly), accidents caused by Micrurus should not be neglected, sincethey account for 0.6% of snakebites, with lethality rate of 0.36%.Recently, it has been observed an increase in the number ofenvenoming cases by Micrurus snakes [2,3].
Snake venoms are composed by a wide variety of proteins,peptides and other organic and inorganicmolecules with diversebiological activities. Such molecules act together to promote theimmobilization, death and digestion of prey, or to serve as adefense mechanism against predators [4,5]. The variability incomposition of snake venoms is a result of multiple factors suchas diet, age, gender and environmental changes [6–10]. Venomsfrom Viperidae and Crotalidae species are known to containproteins that interfere mainly with the blood clotting andhemostatic systems. On the other hand, Elapidae venoms areknown chiefly by their neurotoxic feature [11,12].
Coral snakes, the representative snakes of the Elapidaefamily in the Americas, include the genera Leptomicrurus,Micruroides and Micrurus, being the latter the most diversegenus, with nearly sixty species [13]. Micrurus genus rangesfrom the southern of the United States to the southern of SouthAmerica [11]. In Brazil, envenoming cases by coral snakes arecausedmainly byM. corallinus andM. frontalis, two of the widelyspread species found in the Central, South and Southeastregions. However, there are twenty two species with widegeographic dispersion in Brazil as exemplified byM. lemniscatus(all regions),M. ibiboboca (Northeast) andM. spixii (northern) [14].
Human envenoming by Micrurus species causes palpebralptosis, ophthalmoplegia, paralysis of jaw, larynx and pharynxmuscles, sialorrhea and paralysis of neck and limbmuscles. Insevere cases, respiratory and cardiac arrest may occur andcause death [12,15]. It has been reported that doses rangingfrom 4 to 5 mg of Micrurus venom is lethal to humans [16].Biological activities in Micrurus venoms such as neurotoxicity,myotoxicity, cardiotoxicity and hemolysis are the result froma variety of enzymatic activities [17–20], including phospho-lipase A2 (PLA2), hyaluronidase, phosphodiesterase, acetyl-cholinesterase, L-amino acid dehydrogenase, and L-aminoacid oxidase (LAO) activities [20–23]. Although several biolog-ical activities and clinical manifestations have been described,neurotoxicity is, in fact, the major symptom of Micrurusenvenomation. Micrurus venoms contain neurotoxins thatparalyze the breathing muscles, acting pre (PLA2) or postsyn-aptically (α-neurotoxins—3FTx), representing the main toxinclasses described [15,24,25].
A major obstacle to characterize the components ofMicrurus venoms is theminute quantities of material obtainedfrom specimens milking. Consequently, the biochemical andpharmacological properties of most of their componentsremain unknown or poorly understood. Recently, in order totackle this problem, molecular biology-based approaches andvenomic/anti-venomics studies have been developed and theamount of information in literature is beginning to increase.Micrurus venoms transcriptomes and proteomes characterizedto date are those from M. corallinus, M. surinamensis, M.pirrhocryptus, M. altirostris and M. nigrocinctus [21,26–31].Proteomic studies of M. altirostris and M. corallinus venomsshowed that α-neurotoxins (from the 3FTx family) and PLA2
are the most abundant and diversified toxin families, corre-sponding, respectively, to 79.5% and 13.7% in first venom and81.7% and 11.9% in second venom [27]. In contrast, proteomicanalysis of M. nigrocinctus venom showed that PLA2 ispredominant in this venom [28]. Other protein families wereidentified in Micrurus venoms by proteomic and transcrip-tomic analyses, including natriuretic peptides, ohanin-like,waprin-like, LAO, C-type lectin/lectin-like, metalloproteinase,serine proteinase, nucleotidase, muscarinic-like and bungar-otoxin-like toxins and most recently, lipases [21,26–29].
The treatment of envenoming by Micrurus snakes consistsin the intravenous administration of specific (anti-Micrurusspp) or non-specific (anti-Elapid) antivenom. In Brazil, anti-Elapid antivenom is prepared by the hyperimmunization ofhorses using M. corallinus and M. frontalis venoms. Thispolyvalent antivenom is able to neutralize venoms fromdifferent coral snakes species. However, some toxic compo-nents present in other venoms from different species mightnot be recognized [23,32]. Differences in antivenom recogni-tion among protein families, such as α-neurotoxin, PLA2 andhigh molecular mass proteins, were previously observed inimmunoblot assays using the M. nigrocinctus venom againstthe specific antivenom [29]. Other venomic and antivenomicstudies with Elapidae and Viperidae venoms have shown thatboth mono and polyvalent antivenoms are not able torecognize all the toxins in specific venoms [28,33–37].
Taking these facts into account, a deeper knowledge onvenom composition, with focus on the identification ofstructural families that represent toxic and immunogeniccomponents in Micrurus venoms, is necessary in order toproduce antivenoms bearing higher specificity, reactivity andprotective action. For a higher effectiveness in serologictreatment, it is crucial to assure the presence of antibodiesthat are able to recognize a larger number of protein familiesthat elicit both pharmacological and immunological re-sponses from the specific Micrurus venoms. In light of this, itbecomes necessary to identify the protein families present inthe different venoms, ensuring their presence for the generalanti-Elapid serum production, enhancing its effectiveness.
In this work, we describe a proteomic study, carried out onthe venoms from three coral snakes M. frontalis, M. ibiboboca,M. lemniscatus by 2D (ion-exchange and reverse-phase) high-performance liquid chromatography (HPLC) and mass spec-trometry profiles in order to better know the structuralrichness of these venoms. In addition, we report for the firsttime the N-terminal sequence of four new proteins purifiedfrom the M. lemniscatus venom. Clues on their possiblebiological activities were determined by similarity searchingin protein databanks. The N-terminal sequences of theproteins from these venoms showed a considerable similarityto 3FTx, PLA2 and Kunitz type protease inhibitor fromother Micrurus and elapid venoms. Moreover, by using 2D-electrophoresis and immunoblotting assays, the venoms fromthe abovementioned species, alongwithM. spixii venom,wereused to check the cross-reactivity against homologous andheterologous antivenoms (anti-M. corallinus, anti-M. frontalis,anti-M. ibiboboca and anti-Elapid). By this approach, we alsointended to assess the most immunogenic protein familiesand to correlate their presence in specific venom with theelicited reactivity against a given antiserum.

1812 J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
2. Material and methods
2.1. Venoms and antivenoms
Venoms from M. frontalis and M. lemniscatus were kindlyprovided by Fundação Ezequiel Dias (FUNED, Belo Horizonte,Brazil). Samples from M. frontalis and M. leminiscatus venomswere taken from male and female adults, originated from thesoutheast region of Brazil, pooled andmaintained in FUNED tobe regularly used to antivenom manufacture. M. ibibobocavenom was a pool from 8 specimens, adults from bothgenders, collected at João Pessoa, PB, northeast of Brazil, andkindly provided by Msc. Gentil Alves Pereira Filho, Departa-mento de Sistemática e Ecologia, Universidade Federal da Paraíba.The used specimens were deposited at the HerpetologicalCollection of this Institution. M. spixii venom, from Peru, waskindly provided by Dr. Eládio Sanchez (FUNED) from a pooledvenom from both genders. After collection, all venoms werelyophilized and kept at −20 °C until used. Anti-M. frontalis,anti-M. ibiboboca, anti-M. corallinus and anti-Elapid antivenomswere also provided by FUNED Elapid antivenomwas producedby hyperimmunization of horses with a pool of M. corallinusand M. frontalis venoms (1:1), following the protocol ofimmunization from FUNED. The serum comprise purified F(ab')2 fragments, generated by pepsic digestion. Monospecificantivenoms (anti-M. corallinus, anti-M. frontalis and anti-M.ibiboboca) were produced by horse immunization using pooledvenoms from each species. All the antivenoms were producedunder standard quality control of FUNED and stored at 4 °Cuntil required, as indicated by the manufacturer.
2.2. Two-dimensional liquid chromatography (2D-LC)
All chromatographic steps were performed by using an ÄKTAExplorer 10 system (GEHealthcare, Uppsala, Sweden), controlledby theUNICORN4.11software (GEHealthcare,Uppsala, Sweden).Elution was monitored by absorbance readings at 214 and280 nm. Fractions were collected with a Frac920 automatedfraction collector (GE Healthcare, Uppsala, Sweden).
2D-LC was performed as described previously [38], withmodifications, as described below. The crude venoms ofM. frontalis, M. ibiboboca, and M. lemniscatus were fractionatedby cation-exchange chromatography (CIEX) (first dimension)and the fractions obtained were further subjected to reverse-phase chromatography (RPC) (second dimension). In the firstdimension, pooled venom samples (2 mg) were loaded onto aTSK-Gel CM-SW column, 15 cm×4.6 mm (Tosoh Biosep, Mon-tgomeryville, USA), equilibrated in 20 mM sodium acetatebuffer at pH 5.0 (solvent A). Elution of fractions was carried outusing a linear gradient 0–1 M NaCl in 20 mM sodium-acetatebuffer (solvent B), with a flow rate of 0.75 mL/min at roomtemperature and monitored by absorbance readings at 214and 280 nm. Eluted fractions were collected at fixed volumes(0.4 mL). In the second dimension, the fractions obtained inthe CIEX fractionation step were loaded onto a Source 154.6/100 reverse-phase column (Pharmacia Biotech, Uppsala,Sweden) equilibrated in 0.1% (v/v) trifluoracetic acid (TFA) inwater (solvent A). The components were eluted by means of athree-step gradient of 0.1% (v/v) TFA in acetonitrile (ACN)
(solvent B), at a flow rate of 1.0 mL/min. The first step,consisted of a 2.5 minutes-long linear gradient (0–10% ofsolvent B), the second step consisted of a 69.7 minutes-longlinear gradient (10–80% of solvent B), and the third consisted ofa 1.6 minutes-long linear gradient (80–100% of solvent B).Elution was monitored by absorbance readings at 214 and280 nm. Eluted fractions were collected at fixed volumes(0.5 mL) and subjected to ESI-Q-TOF/MS or MALDI-TOF/TOFanalyses.
2.3. Mass spectrometry analyses
Mass spectrometry (MS) analyses were carried out using anelectrospray ionization quadrupole time-of-flight (ESI-Q-TOF)instrumentation, in a Q-TOF Micro (Micromass, Manchester,UK) operated in positive mode. The Capillary voltage was3000–4000 V and sample cone voltages were 40–60 V. Massspectrometer was calibrated with sodium iodide and caesiumiodide. Samples were introduced using a syringe pump withflow rates of 5–10 μL/min. Prior to interpretation, 20 scans(2.4 s duration) were combined into a single spectrum. Rawcombined spectral data were treated (base line subtraction,smoothing and centering) and manually deconvoluted intomass information.
Alternatively, MS analyses were performed by matrix-assisted laser desorption ionization time-of-flight (MALDI-TOF/TOF) instrumentation, using an Autoflex III or a UltraFlexIII (Bruker Daltonics, Billerica, USA) operated in positive mode.Samples were dissolved in TFA 0.1% and mixed (1:3, v/v) withα-cyano-4-hydroxycinnamic acid matrix solution (saturatedsolution in TFA 0.1%) spotted onto a MALDI target plate(AnchorChip-600; Bruker Daltonics, Billerica, USA) and dried atroom temperature. Monoisotopic masses were obtained inpositive reflected mode. Tandem mass spectrometric spectra(MS/MS) spectra were obtained by LID fragmentation in LIFTmode.Mass spectrometric datawere analyzed bymeans of thesoftware MassLynx 4.0 (Micromass, Manchester, UK) andFlexAnalysis 2.4 (Bruker Daltonics, Bremen, Germany).
2.4. N-terminal amino acid sequencing
Samples of the native proteins (50–200 pmol) were sequencedby Edman degradation using an automated PPSQ-21A proteinsequencer (Shimadzu, Tokyo, Japan) coupled to reverse phaseseparation of PHT-amino acids on a WAKOSIL-PTH(4.6×250 mm) column (Wako, Osaka, Japan).
2.5. 2D-electrophoresis
ImmobilineDryStrips (7 cm; pH3–11NL, GEHealthcare, Upsalla,Sweden) were rehydrated overnight with rehydration buffer(urea 8 M, thiourea 2 M, 4% v/v 3-[(3-cholamidopropyl)dimethy-lammonio]propanesulfonic acid (CHAPS), 1,4-dithio-DL-threitol(DTT), 1% v/v carrier ampholytes) containing approximately50 μg of the venom. Running was performed in an Ettan IPGhor(GE Healthcare, Uppsala, Sweden) system. Strips were thenprepared for first dimension isoelectric focusing gels, by thefollowing protocol: 300 V to 5000 V in 6 h with a final voltage of500 V in 30min. After the first-dimensional run, the IPG gelstrips were sealed in plastic wrap and frozen at −80 °C or

1813J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
incubated at room temperature in equilibration buffer (2-amino-2-hydroxymethyl-propane-1,3-diol (Tris) 50 mM, pH8.8, urea 6 M, 2% v/v sodium dodecyl sulfate (SDS), 30% v/vglycerol, 0.001% v/v bromophenol blue to 0.5% v/v) containing10 mg/mL of DTT and 25 mg/mL of iodoacetamide. Afterincubation in equilibration buffer, the second dimensionelectrophoresis was performed in a vertical system with auniform 12.5% separating gel, at 25 °C. Protein spots in the 2D-electrophoresis were visualized using Coomassie Blue G-250 2%v/v or transblotted onto a nitrocellulosemembrane forWesternblotting. The coomassie stained spots were excised from thepreparative gel and were digested with trypsin.
2.6. Tryptic digestion
Spots of interest were excised from polyacrilamide gels andincubated in ACN 50% and ammonium bicarbonate 25mM, pH8.0. Spots were then dehydrated with ACN 100% and dried in avacuumconcentrator.Digestionswere carriedoutusing sequenc-ing grade modified porcine trypsin (Promega, Madison, USA),following the manufacturers' recommendations. The mixturewas allowed to react for 20 h at 37 °C. The tryptic peptides wereeluted from the gel spots with 50% (v/v) formic acid in ACN 50%(v/v), and dried in a vacuum concentrator. Digests were spottedonto a MALDI-TOF target (AnchorChip 600 (Bruker Daltonics,Billerica, USA)) after desaltingwith ZipTipC18 (Millipore, Billerica,USA) and using α-cyano-4-hydroxycinnamic acid (1:3, v/v) asmatrix. MALDI-TOF MS/MS spectra were obtained by using LIFTmethod in Autoflex III or Ultraflex III (Bruker Daltonics, Billerica,USA) operated in positive and reflector mode.
2.7. Immunoblotting
ForWestern Blot analyses, 2D-electrophoresis usingM. frontalis,M. ibiboboca,M. lemniscatus andM. spixii venoms was performedas described above. Proteins were electro-transferred to anitrocellulose membrane in a transfer system (Mini-Protean®IICell/Bio-Rad). The membrane was then stained with Ponceau Ssolution and the protein markers were labeled. The blottedmembranes were immersed in a blocking solution (polyox-yethylene (20) sorbitan monolaurate (Tween 20) 0.3% v/v andpowdered milk 0.5% w/v in phosphate buffered saline (PBS),pH=7.4), at 25 °C, for 1 h and then washed thoroughly with awashing solution (Tween 20 0.05% v/v in PBS). The blockedmembrane was immersed in anti-Micrurus antivenom (diluted1:1000 in Tween 0.3% v/v and powdered milk 0.5% p/v in PBS,pH=7.4) and incubated at 25 °C for 2 h. After thorough washing,the membrane was incubated with horse anti-IgG/peroxidaseconjugate (1:500). Intermediate washing steps with Tween 0.3%v/v in PBS were followed by addition of the substrate solution(10 mgof 3,3′-diaminobenzidine (DAB) plus 5 mgof chloronaph-tol in28mLof PBScontaining8 μLof30%v/vH2O2and0.55mLofmethanol). Washings with distilled water stopped the colorreaction.
2.8. Sequence similarity search
The search for similar sequences was performed using theFASTA3 [39] and the Swiss-Prot database [40]. Alternatively,BLASTP 2.2.23+ [41] was employed, using the Swiss-Prot and
GeneBank database using the taxonomy filter for snakes.When convenient, the identified retrieved proteins wereselected based on the closest taxonomical level (related toMicrurus genus) and/or the relevant location (venom againstblood or secreted against membrane proteins, for instance).
3. Results and discussion
3.1. Proteomic characterization of Micrurus venoms by2D-LC and MS analyses
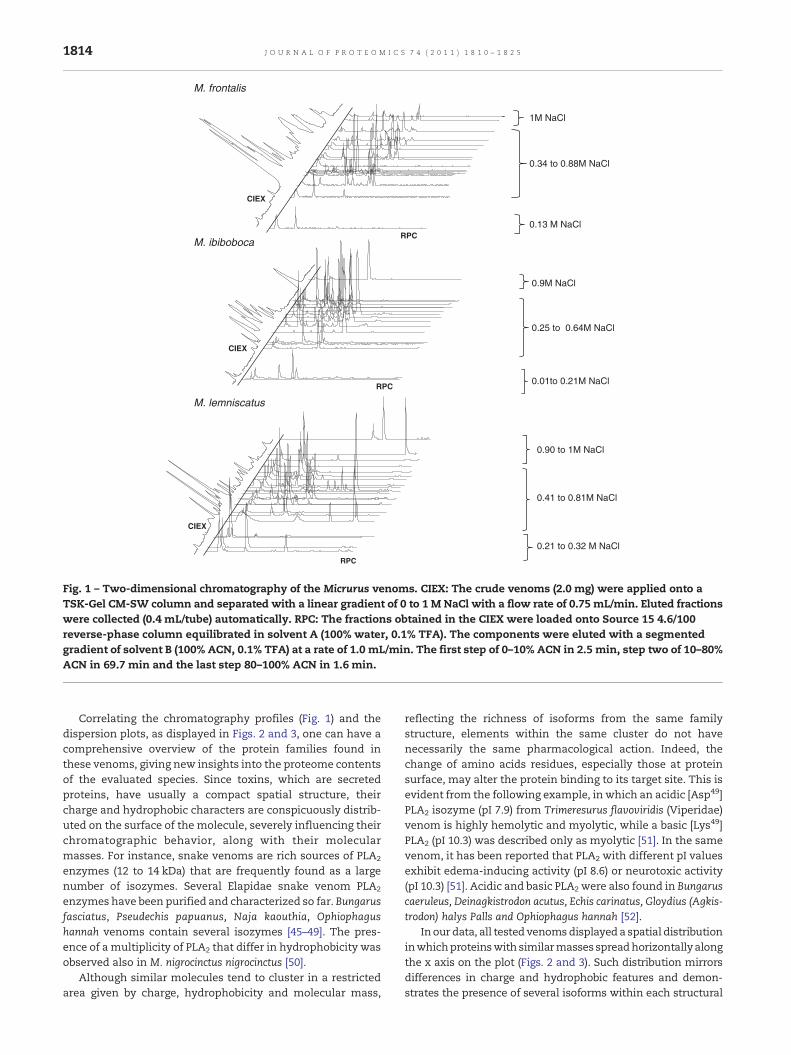
The crude venoms of M. frontalis, M. ibiboboca and M. lemniscatuswere fractionated by CIEX followed by RPC and subjected tomassspectrometry (MS) analyses. The 2D-chromatographic (2D-LC)profile revealed different degrees of complexity for each venom(Fig. 1). TheCIEX separations of these venomsyielded 17 fractionsfor M. frontalis and M. ibiboboca venoms and 24 fractions for theM. leminiscatus venom. M. frontalis and M. lemniscatus venomcontents were eluted within 0.1 to 1 M and 0.2 to 1 M [NaCl]ranges, respectively, whereas the content of M. ibiboboca venomwas eluted within the 0.01–0.9 M [NaCl] range. The fractionsobtained in theCIEX stepwere further separatedbyRPC, inwhichthe contents of M. frontalis and M. ibiboboca venoms elutedbetween 10 and 50% [ACN], whereas the content ofM. lemniscatuseluted within a broader [ACN] range (10–80%), indicating thepresence of more hydrophobic components in this venom.
Proteomic studies on M. alleni, M. multifasciatus, M. surina-mensis, M. nigrocinctus, M. altirostris and M. corallinus venoms,have been accomplished by 1D reverse phase chromatography(1D-LC) [21,27,28,42], each of them using different sets ofchromatographic columns and equipments. Thus, it is difficultto ensure a fair comparison between the chromatographicprofiles of the venoms from species used in the present workwith others Micrurus venoms previously described. However,regarding the hydrophobic behavior, different elution patternscould be observed in other Elapidae venoms. For instance, RPCchromatograms from Naja sub-species showed a major con-centration of proteins with elution peaks in the range of 30 to70% of ACN, indicating that these venoms varied in theirhydrophobic properties [43]. Contrastingly, as verified by Rossoet al. [42], all proteins from M. nigrocinctus nigrocinctus, M. alleniyatesi, and M. multifasciatus have eluted before 35% of ACN [42].
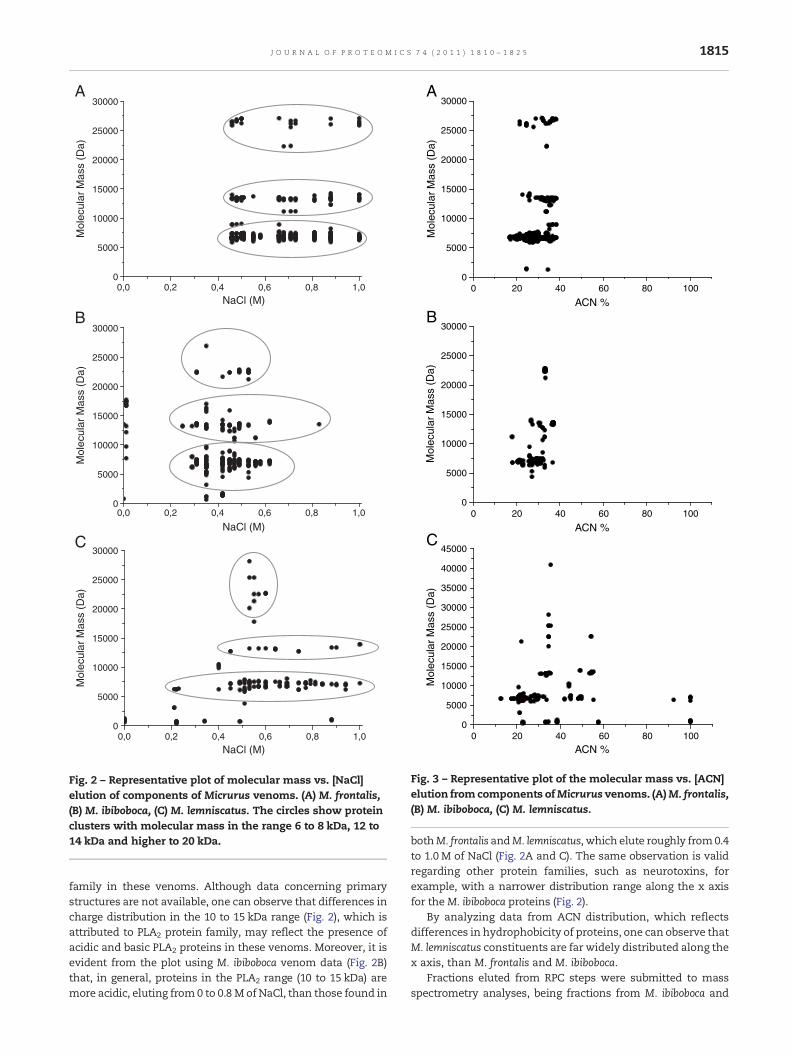
Fig. 2 shows a graphical overview of the CIEX elution profileplotted against the molecular mass of the componentsdetected by means of mass spectrometry. Components withsimilar molecular masses, which indicate possible isoforms,were separated by salt elution, whereas components withdistinct molecular masses were frequently co-eluted. Similarchromatographic behavior has been observed previously byour group in the fractionation of scorpions (Tityus serrulatus,T. stigmurus, T. bahiensis, Leiurus quinquestriatus quinquestriatusand L. q. hebraeus) and centipede (Scolopendra spp) venoms byCIEX [38,44]. Fig. 3 plots the molecular masses of the detectedcomponents in the three assessed venoms against [ACN]elution in RPC. In M. frontalis e M. ibiboboca (Fig. 3A and B)venom, components eluted from 10 to 40% ACN, and thosewith similar molecular masses are frequently clustered in theabscissa. In contrast, venom components in M. lemniscatusdisplayed broader dispersion in both axes (Fig. 3C).
M. frontalis
CIEX
RPC0.13 M NaCl
0.34 to 0.88M NaCl
1M NaCl
M. lemniscatus
0.21 to 0.32 M NaCl
0.41 to 0.81M NaCl
0.90 to 1M NaCl
CIEX
RPC
M. ibiboboca
0.25 to 0.64M NaCl
0.9M NaCl
0.01to 0.21M NaCl
CIEX
RPC
Fig. 1 – Two-dimensional chromatography of the Micrurus venoms. CIEX: The crude venoms (2.0 mg) were applied onto aTSK-Gel CM-SW column and separated with a linear gradient of 0 to 1 M NaCl with a flow rate of 0.75 mL/min. Eluted fractionswere collected (0.4 mL/tube) automatically. RPC: The fractions obtained in the CIEX were loaded onto Source 15 4.6/100reverse-phase column equilibrated in solvent A (100% water, 0.1% TFA). The components were eluted with a segmentedgradient of solvent B (100% ACN, 0.1% TFA) at a rate of 1.0 mL/min. The first step of 0–10% ACN in 2.5 min, step two of 10–80%ACN in 69.7 min and the last step 80–100% ACN in 1.6 min.
1814 J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
Correlating the chromatography profiles (Fig. 1) and thedispersion plots, as displayed in Figs. 2 and 3, one can have acomprehensive overview of the protein families found inthese venoms, giving new insights into the proteome contentsof the evaluated species. Since toxins, which are secretedproteins, have usually a compact spatial structure, theircharge and hydrophobic characters are conspicuously distrib-uted on the surface of the molecule, severely influencing theirchromatographic behavior, along with their molecularmasses. For instance, snake venoms are rich sources of PLA2
enzymes (12 to 14 kDa) that are frequently found as a largenumber of isozymes. Several Elapidae snake venom PLA2
enzymes have been purified and characterized so far. Bungarusfasciatus, Pseudechis papuanus, Naja kaouthia, Ophiophagushannah venoms contain several isozymes [45–49]. The pres-ence of a multiplicity of PLA2 that differ in hydrophobicity wasobserved also in M. nigrocinctus nigrocinctus [50].
Although similar molecules tend to cluster in a restrictedarea given by charge, hydrophobicity and molecular mass,
reflecting the richness of isoforms from the same familystructure, elements within the same cluster do not havenecessarily the same pharmacological action. Indeed, thechange of amino acids residues, especially those at proteinsurface, may alter the protein binding to its target site. This isevident from the following example, in which an acidic [Asp49]PLA2 isozyme (pI 7.9) from Trimeresurus flavoviridis (Viperidae)venom is highly hemolytic and myolytic, while a basic [Lys49]PLA2 (pI 10.3) was described only as myolytic [51]. In the samevenom, it has been reported that PLA2 with different pI valuesexhibit edema-inducing activity (pI 8.6) or neurotoxic activity(pI 10.3) [51]. Acidic and basic PLA2 were also found in Bungaruscaeruleus, Deinagkistrodon acutus, Echis carinatus, Gloydius (Agkis-trodon) halys Palls and Ophiophagus hannah [52].
Inour data, all tested venomsdisplayed a spatial distributioninwhichproteinswith similarmasses spreadhorizontallyalongthe x axis on the plot (Figs. 2 and 3). Such distribution mirrorsdifferences in charge and hydrophobic features and demon-strates the presence of several isoforms within each structural
0,0 0,2 0,4 0,6 0,8 1,00
5000
10000
15000
20000
25000
30000A
B
C
Mol
ecul
ar M
ass
(Da)
NaCl (M)
0,0 0,2 0,4 0,6 0,8 1,00
5000
10000
15000
20000
25000
30000
Mol
ecul
ar M
ass
(Da)
NaCl (M)
0,0 0,2 0,4 0,6 0,8 1,00
5000
10000
15000
20000
25000
30000
Mol
ecul
ar M
ass
(Da)
NaCl (M)
Fig. 2 – Representative plot of molecular mass vs. [NaCl]elution of components of Micrurus venoms. (A) M. frontalis,(B) M. ibiboboca, (C) M. lemniscatus. The circles show proteinclusters with molecular mass in the range 6 to 8 kDa, 12 to14 kDa and higher to 20 kDa.
0 20 40 60 80 1000
5000
10000
15000
20000
25000
30000A
B
C
Mol
ecul
ar M
ass
(Da)
ACN %
0 20 40 60 80 1000
5000
10000
15000
20000
25000
30000
Mol
ecul
ar M
ass
(Da)
ACN %
0 20 40 60 80 1000
5000
10000
15000
20000
25000
30000
35000
40000
45000
Mol
ecul
ar M
ass
(Da)
ACN %
Fig. 3 – Representative plot of the molecular mass vs. [ACN]elution from components ofMicrurus venoms. (A)M. frontalis,(B) M. ibiboboca, (C) M. lemniscatus.
1815J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
family in these venoms. Although data concerning primarystructures are not available, one can observe that differences incharge distribution in the 10 to 15 kDa range (Fig. 2), which isattributed to PLA2 protein family, may reflect the presence ofacidic and basic PLA2 proteins in these venoms. Moreover, it isevident from the plot using M. ibiboboca venom data (Fig. 2B)that, in general, proteins in the PLA2 range (10 to 15 kDa) aremore acidic, eluting from 0 to 0.8 M of NaCl, than those found in
bothM. frontalis andM. lemniscatus, which elute roughly from0.4to 1.0 M of NaCl (Fig. 2A and C). The same observation is validregarding other protein families, such as neurotoxins, forexample, with a narrower distribution range along the x axisfor theM. ibiboboca proteins (Fig. 2).
By analyzing data from ACN distribution, which reflectsdifferences in hydrophobicity of proteins, one can observe thatM. lemniscatus constituents are far widely distributed along thex axis, than M. frontalis and M. ibiboboca.
Fractions eluted from RPC steps were submitted to massspectrometry analyses, being fractions from M. ibiboboca and
1816 J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
M. lemniscatus venoms analyzed by ESI-Q-TOF while those fromM. frontalis were analyzed by MALDI-TOF-TOF. A total of 518molecules were detected in the three venoms, being 217 fromM. frontalis, 181 from M. ibiboboca and 120 from M. lemniscatus.Themolecular species detected inM. frontalis venom span from5874.6 to 27,118.4 Da,whereas those detected inM. ibiboboca andin M. lemniscatus span from 631.3 to 26,907.3 Da and 416.0 to28,169.7 Da, respectively (data not shown).
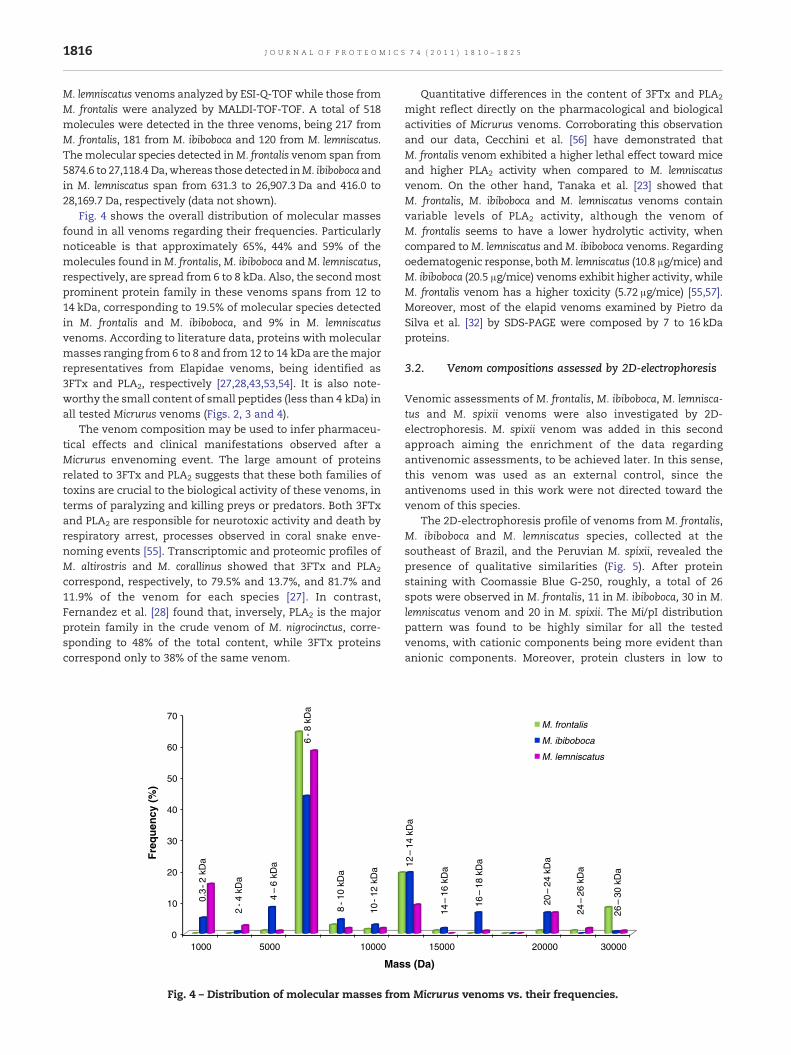
Fig. 4 shows the overall distribution of molecular massesfound in all venoms regarding their frequencies. Particularlynoticeable is that approximately 65%, 44% and 59% of themolecules found inM. frontalis,M. ibiboboca andM. lemniscatus,respectively, are spread from 6 to 8 kDa. Also, the secondmostprominent protein family in these venoms spans from 12 to14 kDa, corresponding to 19.5% of molecular species detectedin M. frontalis and M. ibiboboca, and 9% in M. lemniscatusvenoms. According to literature data, proteins with molecularmasses ranging from 6 to 8 and from12 to 14 kDa are themajorrepresentatives from Elapidae venoms, being identified as3FTx and PLA2, respectively [27,28,43,53,54]. It is also note-worthy the small content of small peptides (less than 4 kDa) inall tested Micrurus venoms (Figs. 2, 3 and 4).
The venom composition may be used to infer pharmaceu-tical effects and clinical manifestations observed after aMicrurus envenoming event. The large amount of proteinsrelated to 3FTx and PLA2 suggests that these both families oftoxins are crucial to the biological activity of these venoms, interms of paralyzing and killing preys or predators. Both 3FTxand PLA2 are responsible for neurotoxic activity and death byrespiratory arrest, processes observed in coral snake enve-noming events [55]. Transcriptomic and proteomic profiles ofM. altirostris and M. corallinus showed that 3FTx and PLA2
correspond, respectively, to 79.5% and 13.7%, and 81.7% and11.9% of the venom for each species [27]. In contrast,Fernandez et al. [28] found that, inversely, PLA2 is the majorprotein family in the crude venom of M. nigrocinctus, corre-sponding to 48% of the total content, while 3FTx proteinscorrespond only to 38% of the same venom.
0.3
-2 k
Da
2 - 4
kD
a
4 –
6 kD
a
6 -8
kD
a
8 -1
0 kD
a
10 -
12 k
Da
0
10
20
30
40
50
60
70
1000 5000 10000
Fre
qu
ency
(%
)
Mas
Fig. 4 – Distribution of molecular masses from
Quantitative differences in the content of 3FTx and PLA2
might reflect directly on the pharmacological and biologicalactivities of Micrurus venoms. Corroborating this observationand our data, Cecchini et al. [56] have demonstrated thatM. frontalis venom exhibited a higher lethal effect toward miceand higher PLA2 activity when compared to M. lemniscatusvenom. On the other hand, Tanaka et al. [23] showed thatM. frontalis, M. ibiboboca and M. lemniscatus venoms containvariable levels of PLA2 activity, although the venom ofM. frontalis seems to have a lower hydrolytic activity, whencompared toM. lemniscatus andM. ibiboboca venoms. Regardingoedematogenic response, bothM. lemniscatus (10.8 μg/mice) andM. ibiboboca (20.5 μg/mice) venoms exhibit higher activity, whileM. frontalis venom has a higher toxicity (5.72 μg/mice) [55,57].Moreover, most of the elapid venoms examined by Pietro daSilva et al. [32] by SDS-PAGE were composed by 7 to 16 kDaproteins.
3.2. Venom compositions assessed by 2D-electrophoresis
Venomic assessments of M. frontalis, M. ibiboboca, M. lemnisca-tus and M. spixii venoms were also investigated by 2D-electrophoresis. M. spixii venom was added in this secondapproach aiming the enrichment of the data regardingantivenomic assessments, to be achieved later. In this sense,this venom was used as an external control, since theantivenoms used in this work were not directed toward thevenom of this species.
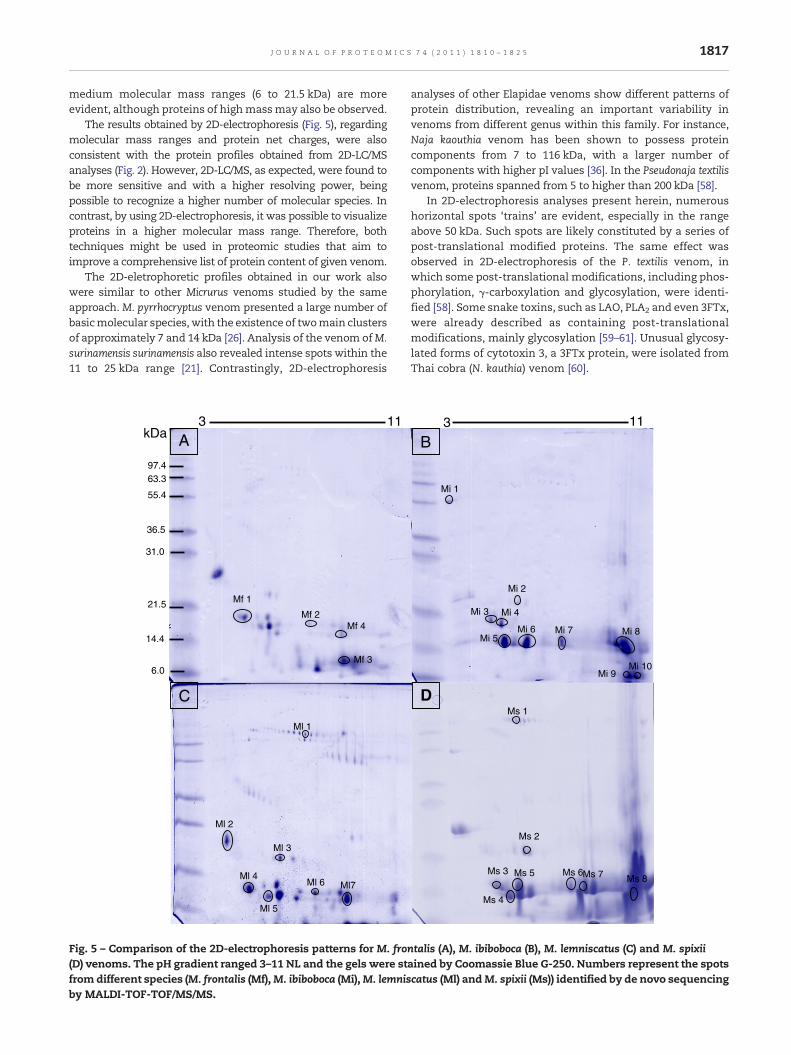
The 2D-electrophoresis profile of venoms from M. frontalis,M. ibiboboca and M. lemniscatus species, collected at thesoutheast of Brazil, and the Peruvian M. spixii, revealed thepresence of qualitative similarities (Fig. 5). After proteinstaining with Coomassie Blue G-250, roughly, a total of 26spots were observed in M. frontalis, 11 in M. ibiboboca, 30 in M.lemniscatus venom and 20 in M. spixii. The Mi/pI distributionpattern was found to be highly similar for all the testedvenoms, with cationic components being more evident thananionic components. Moreover, protein clusters in low to
12 –
14 k
Da
14 –
16 k
Da
16 –
18 k
Da
20 –
24 k
Da
24 –
26 k
Da
26 –
30 k
Da
15000 20000 30000
s (Da)
M. frontalis
M. ibiboboca
M. lemniscatus
Micrurus venoms vs. their frequencies.
1817J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
medium molecular mass ranges (6 to 21.5 kDa) are moreevident, although proteins of highmassmay also be observed.
The results obtained by 2D-electrophoresis (Fig. 5), regardingmolecular mass ranges and protein net charges, were alsoconsistent with the protein profiles obtained from 2D-LC/MSanalyses (Fig. 2). However, 2D-LC/MS, as expected, were found tobe more sensitive and with a higher resolving power, beingpossible to recognize a higher number of molecular species. Incontrast, by using 2D-electrophoresis, it was possible to visualizeproteins in a higher molecular mass range. Therefore, bothtechniques might be used in proteomic studies that aim toimprove a comprehensive list of protein content of given venom.
The 2D-eletrophoretic profiles obtained in our work alsowere similar to other Micrurus venoms studied by the sameapproach. M. pyrrhocryptus venom presented a large number ofbasicmolecular species, with the existence of twomain clustersof approximately 7 and 14 kDa [26]. Analysis of the venom ofM.surinamensis surinamensis also revealed intense spots within the11 to 25 kDa range [21]. Contrastingly, 2D-electrophoresis
Ml 1
Ml 2
Ml 3
Ml 4
Ml 5
Ml 6
C
A
Mf 1
Mf 2
Mf 3
Mf 4
Ml7
6.0
14.4
21.5
31.0
36.5
55.4
63.397.4
3 11kDa
Fig. 5 – Comparison of the 2D-electrophoresis patterns for M. fro(D) venoms. The pH gradient ranged 3–11 NL and the gels were stfrom different species (M. frontalis (Mf),M. ibiboboca (Mi),M. lemnisby MALDI-TOF-TOF/MS/MS.
analyses of other Elapidae venoms show different patterns ofprotein distribution, revealing an important variability invenoms from different genus within this family. For instance,Naja kaouthia venom has been shown to possess proteincomponents from 7 to 116 kDa, with a larger number ofcomponents with higher pI values [36]. In the Pseudonaja textilisvenom, proteins spanned from 5 to higher than 200 kDa [58].
In 2D-electrophoresis analyses present herein, numeroushorizontal spots ‘trains’ are evident, especially in the rangeabove 50 kDa. Such spots are likely constituted by a series ofpost-translational modified proteins. The same effect wasobserved in 2D-electrophoresis of the P. textilis venom, inwhich some post-translational modifications, including phos-phorylation, γ-carboxylation and glycosylation, were identi-fied [58]. Some snake toxins, such as LAO, PLA2 and even 3FTx,were already described as containing post-translationalmodifications, mainly glycosylation [59–61]. Unusual glycosy-lated forms of cytotoxin 3, a 3FTx protein, were isolated fromThai cobra (N. kauthia) venom [60].
DMs 1
Ms 2
Ms 3 Ms 5 Ms 6
B
Mi 1
Mi 2
Mi 3 Mi 4
Mi 5Mi 6 Mi 7 Mi 8
Mi 9Mi 10
Ms 4
Ms 7 Ms 8
3 11
ntalis (A), M. ibiboboca (B), M. lemniscatus (C) and M. spixiiained by Coomassie Blue G-250. Numbers represent the spotscatus (Ml) andM. spixii (Ms)) identified by de novo sequencing
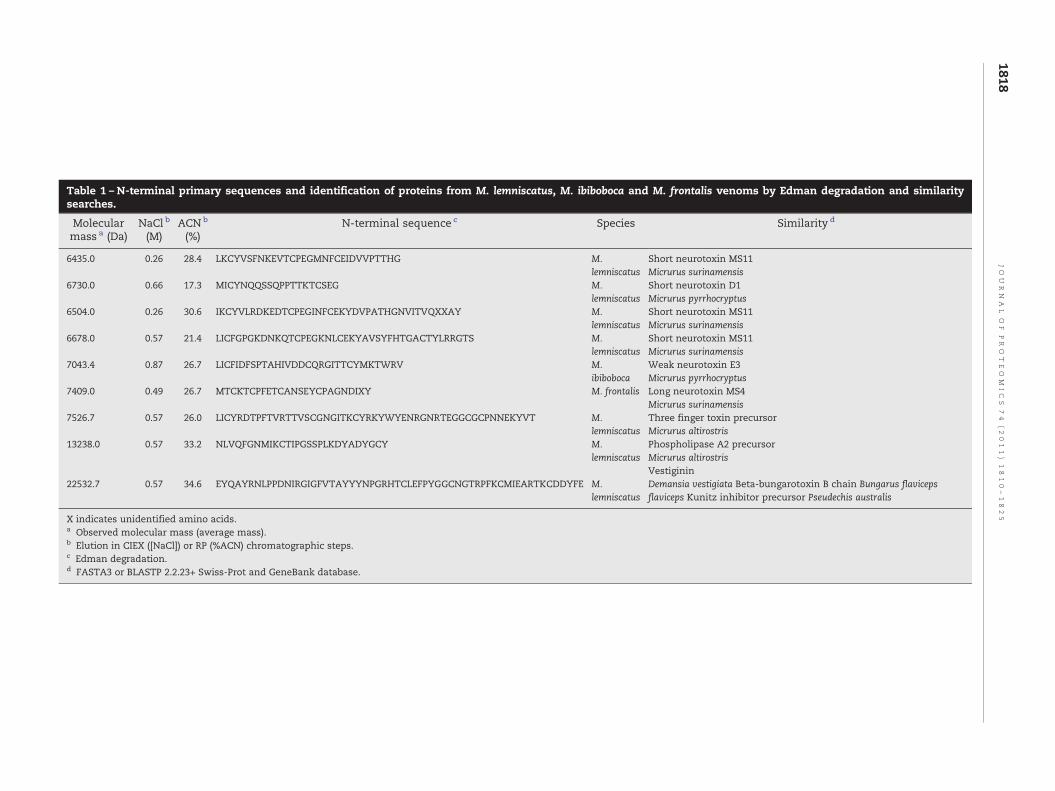
Table 1 – N-terminal primary sequences and identification of proteins from M. lemniscatus, M. ibiboboca and M. frontalis venoms by Edman degradation and similaritysearches.
Molecularmass a (Da)
NaCl b
(M)ACN b
(%)N-terminal sequence c Species Similarity d
6435.0 0.26 28.4 LKCYVSFNKEVTCPEGMNFCEIDVVPTTHG M.lemniscatus
Short neurotoxin MS11Micrurus surinamensis
6730.0 0.66 17.3 MICYNQQSSQPPTTKTCSEG M.lemniscatus
Short neurotoxin D1Micrurus pyrrhocryptus
6504.0 0.26 30.6 IKCYVLRDKEDTCPEGINFCEKYDVPATHGNVITVQXXAY M.lemniscatus
Short neurotoxin MS11Micrurus surinamensis
6678.0 0.57 21.4 LICFGPGKDNKQTCPEGKNLCEKYAVSYFHTGACTYLRRGTS M.lemniscatus
Short neurotoxin MS11Micrurus surinamensis
7043.4 0.87 26.7 LICFIDFSPTAHIVDDCQRGITTCYMKTWRV M.ibiboboca
Weak neurotoxin E3Micrurus pyrrhocryptus
7409.0 0.49 26.7 MTCKTCPFETCANSEYCPAGNDIXY M. frontalis Long neurotoxin MS4Micrurus surinamensis
7526.7 0.57 26.0 LICYRDTPFTVRTTVSCGNGITKCYRKYWYENRGNRTEGGCGCPNNEKYVT M.lemniscatus
Three finger toxin precursorMicrurus altirostris
13238.0 0.57 33.2 NLVQFGNMIKCTIPGSSPLKDYADYGCY M.lemniscatus
Phospholipase A2 precursorMicrurus altirostrisVestiginin
22532.7 0.57 34.6 EYQAYRNLPPDNIRGIGFVTAYYYNPGRHTCLEFPYGGCNGTRPFKCMIEARTKCDDYFE M.lemniscatus
Demansia vestigiata Beta-bungarotoxin B chain Bungarus flavicepsflaviceps Kunitz inhibitor precursor Pseudechis australis
X indicates unidentified amino acids.a Observed molecular mass (average mass).b Elution in CIEX ([NaCl]) or RP (%ACN) chromatographic steps.c Edman degradation.d FASTA3 or BLASTP 2.2.23+ Swiss-Prot and GeneBank database.
1818JO
UR
NA
LO
FPR
OT
EO
MIC
S74
(2011)
1810–1825
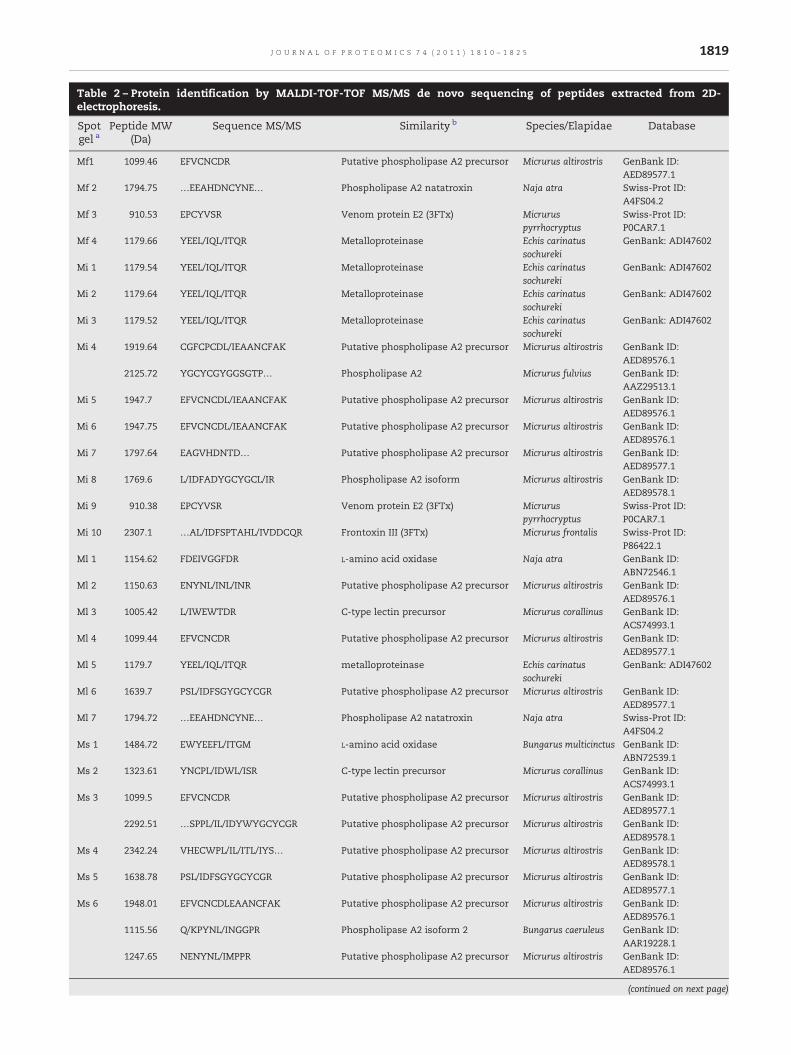
Table 2 – Protein identification by MALDI-TOF-TOF MS/MS de novo sequencing of peptides extracted from 2D-electrophoresis.
Spotgel a
Peptide MW(Da)
Sequence MS/MS Similarity b Species/Elapidae Database
Mf1 1099.46 EFVCNCDR Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89577.1
Mf 2 1794.75 …EEAHDNCYNE… Phospholipase A2 natatroxin Naja atra Swiss-Prot ID:A4FS04.2
Mf 3 910.53 EPCYVSR Venom protein E2 (3FTx) Micruruspyrrhocryptus
Swiss-Prot ID:P0CAR7.1
Mf 4 1179.66 YEEL/IQL/ITQR Metalloproteinase Echis carinatussochureki
GenBank: ADI47602
Mi 1 1179.54 YEEL/IQL/ITQR Metalloproteinase Echis carinatussochureki
GenBank: ADI47602
Mi 2 1179.64 YEEL/IQL/ITQR Metalloproteinase Echis carinatussochureki
GenBank: ADI47602
Mi 3 1179.52 YEEL/IQL/ITQR Metalloproteinase Echis carinatussochureki
GenBank: ADI47602
Mi 4 1919.64 CGFCPCDL/IEAANCFAK Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89576.1
2125.72 YGCYCGYGGSGTP… Phospholipase A2 Micrurus fulvius GenBank ID:AAZ29513.1
Mi 5 1947.7 EFVCNCDL/IEAANCFAK Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89576.1
Mi 6 1947.75 EFVCNCDL/IEAANCFAK Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89576.1
Mi 7 1797.64 EAGVHDNTD… Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89577.1
Mi 8 1769.6 L/IDFADYGCYGCL/IR Phospholipase A2 isoform Micrurus altirostris GenBank ID:AED89578.1
Mi 9 910.38 EPCYVSR Venom protein E2 (3FTx) Micruruspyrrhocryptus
Swiss-Prot ID:P0CAR7.1
Mi 10 2307.1 …AL/IDFSPTAHL/IVDDCQR Frontoxin III (3FTx) Micrurus frontalis Swiss-Prot ID:P86422.1
Ml 1 1154.62 FDEIVGGFDR L-amino acid oxidase Naja atra GenBank ID:ABN72546.1
Ml 2 1150.63 ENYNL/INL/INR Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89576.1
Ml 3 1005.42 L/IWEWTDR C-type lectin precursor Micrurus corallinus GenBank ID:ACS74993.1
Ml 4 1099.44 EFVCNCDR Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89577.1
Ml 5 1179.7 YEEL/IQL/ITQR metalloproteinase Echis carinatussochureki
GenBank: ADI47602
Ml 6 1639.7 PSL/IDFSGYGCYCGR Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89577.1
Ml 7 1794.72 …EEAHDNCYNE… Phospholipase A2 natatroxin Naja atra Swiss-Prot ID:A4FS04.2
Ms 1 1484.72 EWYEEFL/ITGM L-amino acid oxidase Bungarus multicinctus GenBank ID:ABN72539.1
Ms 2 1323.61 YNCPL/IDWL/ISR C-type lectin precursor Micrurus corallinus GenBank ID:ACS74993.1
Ms 3 1099.5 EFVCNCDR Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89577.1
2292.51 …SPPL/IL/IDYWYGCYCGR Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89578.1
Ms 4 2342.24 VHECWPL/IL/ITL/IYS… Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89578.1
Ms 5 1638.78 PSL/IDFSGYGCYCGR Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89577.1
Ms 6 1948.01 EFVCNCDLEAANCFAK Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89576.1
1115.56 Q/KPYNL/INGGPR Phospholipase A2 isoform 2 Bungarus caeruleus GenBank ID:AAR19228.1
1247.65 NENYNL/IMPPR Putative phospholipase A2 precursor Micrurus altirostris GenBank ID:AED89576.1
(continued on next page)
1819J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5

Table 2 (continued)
Spotgel a
Peptide MW(Da)
Sequence MS/MS Similarity b Species/Elapidae Database
Ms 7 2960.44 …L/IDFADYGCYCGYGGSGTPVDEL/IDR
phospholipase A2 I precursor Walterinnesiaaegyptia
GenBank ID:ABX82863.1
Ms 8 2369.16 …CYNDFSYTANTVED… Putative three finger toxin precursor(3FTx)
Micrurus altirostris GenBank ID:AED89559.1
a Mf (M. frontalis), Mi (M. ibiboboca), Ml (M. lemniscatus) and Ms (M. spixii).b FASTA3 or BLASTP 2.2.23+ Swiss-Prot and GeneBank database.
1820 J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
3.3. Identification of protein families from Micrurusvenoms
Identification of proteins from Micrurus venoms in the presentworkwas carried out by determining partial primary structures,using Edman Degradation and de novo sequencing by MALDI-TOF-TOF MS/MS, followed by databank search for similarities.
Firstly, by employing 2D-LC approach, nine proteins fromthe venoms of M. frontalis, M. ibibiboboca and M. lemniscatuswere purified to homogeneity, as determined by MS (MALDI-TOF-TOF or ESI-Q-TOF) analyses, and subjected to N-terminalsequencing by Edman degradation. The partial primarystructures determined in this step are shown in Table 1,which also depicts their observed molecular masses and theirelution times in both CIEX and RPC chromatographic steps.The sequenced proteins display structural similarities to shortand long neurotoxins, weak neurotoxins, PLA2 and β-bungar-otoxin already described in venoms from Micrurus and otherElapidae genera.
Additionally, several spots from 2-D electrophoresis (usingM. frontalis,M. ibiboboca,M. lemniscatusandM. spixiivenoms)wereselected for identificationby in geldigestions followedbydenovosequencing byMALDI-TOF-TOFMS/MSof tryptic fragments. Theidentified proteins from M. frontalis, M. ibiboboca, M. lemniscatusand M. spixii venoms were similar to PLA2, venom proteins E2,Frontoxin III, a weak neurotoxin, LAO, metalloproteinase and aC-type lectin from other Micrurus and Elapidae snakes (Table 2).
In a recent paper, Fernandez et al. [28] have used N-terminaland MS/MS de novo peptide sequencing to identify differentproteins similar to PLA2, 3FTx, LAO, C-type lectin/ lectin-like,metalloproteinase, serine proteinase, ohanin, and nucleotidasein M. nigrocinctus venom. Also, transcriptomic and proteomicanalyses of both M. altirostris and M. corallinus venoms showedthe occurrence of the several toxin families such as PLA2, 3FTx,Kunitz-type proteinase inhibitor, C-type lectin-like, natriureticpeptide, metalloproteinase, veficolin, lysosomal acid lipase A,ohanin-like and waprin-like [27].
In the present study, themajority of proteins identifiedwerefound to be similar to PLA2. Protein identification indicated thepresenceof bothacidicandbasicPLA2 inM. frontalis,M. ibiboboca,M. lemniscatus andM. spixii, as canbeobservedby awide range ofCIEX dispersion and pI values (Figs. 2 and 5 and Tables 1 and 2).Consistently, the presence of both basic and acidic PLA2 in thevenom of M. frontalis had been previously reported [62]. Ingeneral, basic PLA2 enzymes are more toxic and exhibit higherpharmacological potency than their neutral and acidic coun-terparts, being the basic residues responsible for such potencyand lethality [63].
Aside from displaying enzymatic activities, some vPLA2
possess a wide range of toxic effects, including neurotoxicity,myotoxicity, cardiotoxicity, cytotoxicity, and may provokeconvulsion and hypotension or affect blood coagulation andplatelet aggregation.
Table 2 shows the N-terminal sequence obtained from a22,532.7 Da protein fromM. lemniscatus, which is similar to theB chain of β-bungarotoxin from Bungarus flaviceps flaviceps.β-bungarotoxin is a 21.8 kDa heterodimeric basic protein,which exerts its neurotoxic effects presynaptically. In thisheterodimer, the A-chain has phospholipase activitywhile theB-chain is similar to bovine pancreatic trypsin inhibitor,regarding its primary structure [64]. However, the threedimensional structure of the B-chain is significantly differentfrom native bovine pancreatic trypsin inhibitors and, there-fore, could account for the lack of such inhibitory activity [65].
Several proteins herein identified from the four assessedvenoms possess significant similarity with neurotoxins charac-terized from other Micrurus species and/or other Elapid snakes.Several α-neurotoxin isoforms from Elapid venoms have beendescribed previously [66] and they are divided, based on theirprimary structures, into long chain α-neurotoxin group (65–72 residues with five disulfide bridges) and short chain α-neurotoxin group (60–62 residues with four disulfide bridges)[67]. Neurotoxins block nervous transmission by binding toacetylcholine receptors at the postsynaptic membranes ofskeletal muscles or neurons [68]. Cobrotoxin, a short-chainpost-synaptic α-neurotoxin isolated from Naja atra, has beenreported to show analgesic activity [69].
Few neurotoxins from other Micrurus venoms have beencharacterized to date [30,42]. Recently, Moreira et al. [70]isolated six new α-neurotoxins (Fontoxin I–VI) fromM. frontalisvenom. These toxins produce neuromuscular blockage byinteraction with postsynaptic nicotinic acethylcholine recep-tors in frog neuromuscular junctions.
In this study, a peptide sequence, identified in bothM. frontalis and M. ibiboboca venoms, has shown similarity tovenom protein E2 (Swiss-Prot ID: P0CAR7.1) from M. pyrrhocryp-tus (Table 2). Venom protein E2 does not show significantsimilarity to other Elapidae toxins and, for that reason, wasdescribed as representative from a novel structural family inElapidae venom [26]. The biologic activity of this proteinremains elusive.
In both venoms of M. lemniscatus and M. spixi, it waspossible to identify high molecular mass proteins (≈55.4 kDa)similar to LAOs (Table 2). LAOs are flavoenzymes, whichcatalyze the oxidative deamination of L-amino acids withsubsequent production of α-keto acids, H2O2 and ammonia.

C
D
A
(I) (II) (III) (IV)
B
6.0
14.4
21.5
36.5
55.4
6.0
14.4
21.5
36.5
55.4
6.0
14.4
21.5
36.5
55.4
6.0
14.4
21.5
36.5
55.4
3 11 3 11 3 11 3 11
PLA 2
3FTX
PLA2
3FTX
LAO
PLA2C-type lectin
LAO
PLA2C-type lectin
kDa
Fig. 6 – 2D-immunoblot ofMicrurus venoms. (A)M. frontalis venom (B)M. lemniscatus venom (C)M. ibiboboca venom (D)M. spixiivenom (I) anti- M. corallinus, (II) anti-M. frontalis,(III) anti-Elapid, (IV) anti-M. ibiboboca. Arrows indicate the Mr of the indicatedstructural families.
1821J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
Moreover, LAOs are also involved in different activitiesincluding the induction or inhibition of platelet aggregation[71–73], anticoagulant activity [74], stimulation of edemaformation [72,75], besides hemorrhagic [72], antibacterial,and leishmanicidal activities by means of apoptotic mecha-nisms [59,75,76]. Although the apoptosis mechanism is notcompletely clear, it seems to involve the production of H2O2,achieved by oxidation processes of some plasma membraneproteins [77]. This protein family is better characterized inViperidae venoms and, although it has been also identified inMicrurus venoms [21,78], further characterization in Elapidaevenoms is still lacking.
3.4. Cross-reactivity between different Micrurus venomsand Micrurus anti-venoms
The cross reactivity between different Elapid antivenoms(anti-M. corallinus, anti-M. frontalis, anti-M. ibiboboca andheterologous anti-Elapid) and M. frontalis, M. ibiboboca,M. lemniscatus and M. spixii venoms was investigated bymeans of 2D-immunoblotting. Among all the tested antive-noms, heterologous anti-Elapid was the serum that best
reacted against all venoms (Fig. 6, column III). This heterolo-gous antivenom recognized venom protein families withmolecular mass spanning from 6 to 15 kDa, 21.5 to 36.5 kDaand above 55 kDa. By correlating the results of immunoblot-tingwith the identification of proteins byMS/MS, it is observedthat anti-Elapid serum presented high recognition rate ofproteins such as PLA2, 3FTx, LAO and C-type lectin. Morerecently, Tanaka et al. [23] demonstrated that the heterolo-gous anti-Elapid (produced by Butantan Institute, Brazil) couldrecognize several but not all components in Micrurus sp.venoms. Analysis by 1D-immunoblotting showed that the M.corallinus and M. ibiboboca venoms had a higher number ofcomponents recognized among those tested venoms and thatproteins, with molecular mass above 64 kDa, were notrecognized.
In general terms, anti-M. frontalis and anti-M. ibiboboca havealso shown good ability in recognizing all tested venoms (Fig. 6columns II and IV), although M. lemniscatus venom was poorlyrecognized by anti-M. ibiboboca antivenom (Fig. 6B-IV). Proteinfamilies, as PLA2, 3FTx, LAO and C-type lectin, were alsorecognized by both anti-M. frontalis and anti-M. ibibobocaantivenoms. On the other hand, anti-M. corallinus antivenom

1822 J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
reactedweakly against proteins fromM. frontalis,M. lemniscatus,M. ibiboboca and M. spixii venoms. This antivenom recognizedonly a few components with molecular mass above 55 kDa,corresponding to LAO proteins as identified by MS/MS, andsome low molecular mass components. Although, these lowmolecular mass components have not been identified specifi-cally in gel carried outwithM. spixii venom, theywere identifiedin other venoms as 3FTx proteins (Fig. 6 column I).
M. frontalis venom has presented the highest score ofproteins recognized by anti-M. corallinus, anti-M. frontalis, anti-M. ibiboboca and anti-Elapid antivenoms (Fig. 6 lane A).
In some cases, immunoblotting revealed weak recognitionof proteins with molecular mass around 6 kDa, identified as3FTx. This was the case of anti-M. corallinus, anti-M. frontalisand anti-Elapid antivenoms (Fig. 6 column I to III) that havenot recognized adequately such proteins from M. frontalis andM. lemniscatus venoms (Fig. 6 lane A and B). The weakrecognition of proteins from this structural family by specificand heterologous antivenomsmight be of concern, since theseproteins represent one of the most toxic components ofMicrurus and other Elapidae venoms.
Previous studieshavealso demonstrated the low recognitionpower against 3FTx and other neurotoxins [28,37]. The immu-noreactivity profile of heterologous anti-Elapid antivenom,produced by Butantan Institute—achieved by mixing M. coralli-nus andM. frontalisvenoms, againstM.altirostris venom, showedweak recognition of themajor lowmolecularmass toxins (3FTxand PLA2) compared to higher molecular mass molecules [27].The immunoblotting carried out using M. nigrocinctus venomagainst equineM. nigrocinctusmonospecific antivenom revealeda weak recognition of the 3FTx components, a slightly higherrecognition of PLA2 and a stronger recognition of the highmolecular mass proteins [28]. In another study, cross-reactivityof the M. frontalis, M. corallinus, M. ibiboboca, M. hemprichii,M. spixii, M. fulvius, M. altirostris, M. surinamensis, and M.lemniscatusvenomsrevealed thathomologousandheterologousantivenoms (anti-M. corallinus, anti-M. frontalis, anti-M. spixiiiand anti-Elapid) have recognized venom components in therange of 14.8 to 64.2 kDa [23].
From our data, one may notice that anti-M. ibibobocaantivenom displayed a strong recognition power againstproteins with low molecular mass, corresponding to 3FTx, asmay be observed in 2-D immunoblotting against M. frontalisvenom (Fig.6A-IV).
Currently, in Brazil, anti-Elapid antivenom is manufac-tured by hyperimmunizing horses with equal amounts ofM. corallinus and M. frontalis venoms. Our findings show thatchanging from M. corallinus to M. ibiboboca venom in immuni-zation protocols used to produce anti-Elapid serum mightconstitute an alternative path to enhance the recognitionability of this heterologous antivenom. Such enhancementwould be achieved firstly due the ability of anti-M. ibiboboca torecognize, with higher sensitivity when compared to anti-M.corallinus, 3FTx-like proteins. Secondly, anti-M. corallinus anti-venom itself has showed weaker ability in recognizing a widerange of protein families in venoms from several species [23].It is worth mentioning, however, that despite these observa-tions, the use of M. ibiboboca venom to antivenom manufac-ture is realistically unlike to be achieved due the scarcity ofthis species, and therefore its venom, in nature and captivity.
4. Conclusions
Proteomic profiles of M. frontalis, M. ibiboboca and M. lemnisca-tus venoms carried out by 2D-cromatography andMS analyseshave shown that these venoms contain a large number ofcomponents, which may be clustered regarding their molec-ular masses, although differences related to their charge andhydrophobicity suggest the presence of different structuralfamilies composed by a variety of isoforms. Twomajor proteinfamilies observed inM. frontalis,M. ibiboboca andM. lemniscatusvenoms havemolecularmass spanning from 6 to 8 kDa and 12to 14 kDa, corresponding to 3FTx and PLA2, respectively. Ourdata are in accordance with previous studies using otherMicrurus and Elapidae venoms, in which these proteins are themain representatives in terms of structural families[27,28,43,53]. Proteins from 3FTx and PLA2 families mayaccount for the major biological effects, being responsiblemainly for neurotoxicity and death by respiratory arrest[53,54]. 3FTx are non-enzymatic polypeptides containing 60to 74 amino acid residues, which present similar tertiarystructure composed by three-stranded loops extending from acentral core constrained by four conserved disulphide bridges[79]. PLA2 enzymes, in turn, hydrolyze glycerophospholipids atthe sn-2 position of the glycerol backbone releasing lysopho-spholipids and fatty acids. PLA2 induce a wide spectrum ofpharmacological effects such as neurotoxic, cardiotoxic,myotoxic, hemolytic, convulsive, anticoagulant, antiplateletactivity, edema inducing and tissue damaging effects [52,54].
N-terminal and MALDI-TOF-TOF/MS/MS de novo peptidesequencing identified proteins similar to PLA2, venom proteinE2, frontoxin III, short neurotoxins, long neurotoxins, weakneurotoxin, LAO, C-type lectin, metalloproteinase and β-bungarotoxin B chain. Also, we reported for the first time theN-terminal sequence of four new proteins purified fromM. leminiscatus venom. It is worth noting that these proteinsmay act synergistically to promote respiratory and cardiacparalysis in injured patients.
The effective treatment of systemic snake envenomation isthe intravenous administration of antivenom, which is pre-pared by thehyperimmunizationofhorses. Antivenomsmaybemono or polyvalent, [80,81] (homologous or heterologous,respectively), taking into account theuse of one ormore specificvenoms to produce the hyperimmune sera. Polyvalent anti-venoms, therefore, are usually more effective because theirability to neutralize venom from more than one species [82].
Unfortunately, in many cases the heterologous antivenommay not effectively protect patients, since antibodies thatrecognize a specific toxic component present in specificvenom may be absent. The best antivenom must be thatable to recognize and neutralize the wider range of toxins invenoms from different species as possible [83]. Heterologousanti-Elapid antivenomsmanufactured by FUNED and InstitutoButantan (São Paulo, Brazil) – two references regardingantivenom production in Brazil by appointment of theMinistry of Health – are generated by hyperimmunizing horseswith M. corallinus and M. frontalis venoms (1:1) [84]. Bothspecies M. corallinus and M. frontalis, which occur in the highlypopulated Central, Southern and Southeastern region ofBrazil, are responsible for the majority of coral snake

1823J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
envenoming cases [3], which explains the choice of thesevenoms to the use in immunization protocol, so far. However,in vivo neutralization assays have shown that anti-Elapid(Instituto Butantan) antivenom is able to neutralize M. spixii,M. frontalis and M. corallinus venoms, but poorly neutralizesM. altirostris and M. lemniscatus venoms [23].
It is worth mentioning that some factors render theproduction of heterologous anti-Elapid or homologous anti-Micrurus antivenom a complex task. Firstly, the maintenance ofMicrurus snakes incaptivity is notably difficult,mainlydue to thehighmortality rates usually observed. Besides, togetherwith thefact that Micrurus specimens are rarely observed in nature, lowvenomyields uponmilking renders small venomavailability forboth antivenomproduction and for scientific researchpurposes.Considering the above, it is crucial to develop alternativeways toimprove the production and yield of the heterologous anti-Elapid serum. Further investigations on protein structural andfunctional content of Micrurus spp venoms is, therefore, anecessary step to pave this alternative way.
In our view, the structural and immunological analysescarried out in this work contribute to enhance our knowledgeon these two fronts and providing subsidies to improve theanti-Elapid antivenom production. The observed cross-reac-tivity between venoms and antivenoms confirmed the impor-tance of usingM. frontalis venom in the current immunizationprotocol for the production of the heterologous anti-Elapidserum. On the other hand, it was also observed that M.corallinus venom is not the most effective venom to be used insuch protocols. According to our data, the replacement ofM. corallinus venom by M. ibiboboca venom in the currentheterologous anti-Elapid serum production protocols couldresult in enhanced antivenom, since anti-M. ibiboboca anti-venom was able to cross-react more efficiently with othervenom proteins, when compared to anti-M. corallinus. Proteinfamilies, such as PLA2, 3FTx, LAO and C-type lectin/lectin-likewere recognized by anti-M. ibiboboca, anti-M. frontalis and anti-Elapidic but were poorly recognized by anti-M. corallinus.
However,M. ibiboboca is a rare species and this replacementis, therefore, unviable due the scarcity of its venom. Also, sincethe majority of immune responses revealed by 2D immuno-blotting analyses were achieved against 3FTx and PLA2 pro-teins, it is reasonable to believe that an antiserum raisedagainst fractionated venom containing such proteins, couldyield better protection.
Acknowledgments
This work was supported by the FAPEMIG, CNPq, CAPES, MCT-FINEP and INCTTOX-Fapesp.
R E F E R E N C E S
[1] Stock RP, Massougbodji A, Alagón A, Chippaux JP. Bringingantivenoms to sub-SaharanAfrica. Nat Biotechnol 2007;25:173–7.
[2] Brasil, Saúde dd. Guia de vigilância epidemiológica. Ministérioda Saúde 2005.
[3] Md Saúde. Manual de diagnóstico e tratamento de acidentespor animais peçonhentos. Ministério da Saúde: FundaçãoNacional de Saúde; 2001.
[4] Kochva E, Nakar O, Ovadia M. Venom toxins: plausibleevolution from digestive enzymes. Am Zool 1983;23:427.
[5] Mebs D. Snake venom composition and evolution of Viperidae.Kaupia 1999;8:145–8.
[6] Andrade DV, Abe AS. Relationship of venom ontogeny anddiet in Bothrops. Herpetologica 1999;55:200–4.
[7] Saldarriaga MM, Otero R, Núñez V, Toro MF, Díaz A, GutiérrezJM. Ontogenetic variability of Bothrops atrox and Bothrops aspersnake venoms from Colombia. Toxicon 2003;42:405–11.
[8] Pimenta DC, Prezoto BC, Konno K, Melo RL, Furtado MF,Camargo A, et al. Mass spectrometric analysis of theindividual variability of Bothrops jararaca venom peptidefraction. Evidence for sex based variation among thebradykinin potentiating peptides. Rapid Commun MassSpectrom 2007;21:1034–42.
[9] Zelanis A, Travaglia-Cardoso SR, De Fátima DominguesFurtado M. Ontogenetic changes in the venom of Bothropsinsularis (Serpentes: Viperidae) and its biological implication.South Am J Herpetol 2008;3:43–50.
[10] Monteiro RQ, Yamanouye N, Carlini CR, Guimarães JA, Bon C,Zingali RB. Variability of bothrojaracin isoforms and othervenom principles in individual jararaca (Bothrops jararaca)snakes maintained under seasonally invariant conditions.Toxicon 1998;36:153–63.
[11] Campbell JA, Lamar WW. The venomous reptiles of LatinAmerica. Comstock Pub. Associates; 1989.
[12] Rosenfeld G. Symptomatology, pathology, and treatment ofsnake bites in South America. Venomous animals and theirvenoms: venomous vertebrates; 1971. p. 345.
[13] da Silva NJ, Sites JW. Phylogeny of South American triad coralsnakes (Elapidae: Micrurus) based on molecular characters.Herpetologica 2001;57:1–22.
[14] Silva Jr N, Bucaretchi F, Cardoso J, França F, Fan H, Málaque C,et al. Mecanismo de ação do veneno elapídico e aspectosclínicos dos acidentes. Cardoso JLC, França FOS, Fan HW,Málaque CMS, Haddad Jr V Animais peçonhentos no BrasilBiologia, clínica e terapêutica dos acidentes São Paulo: Sarvier2003:99–107.
[15] Vital Brazil O, Fontana. Açôes pré-juncionais e pós-juncionaisda peçonha da cobra coral Micrurus corallinus na junçãoneuromuscular. Mem Inst Butantan 1984;47:1984.
[16] Fix JD. Venom yield of the North American coral snake and itsclinical significance. South Med J 1980;73:737.
[17] Gutiérrez JM, Arroyo O, Chaves F, Lomonte B, Cerdas L.Pathogenesis ofmyonecrosis induced by coral snake (Micrurusnigrocinctus) venom in mice. Br J Exp Pathol 1986;67:1–12.
[18] Gutiérrez JM, Rojas G, da Silva Jr NJ, Nunez J. Experimentalmyonecrosis induced by the venoms of South AmericanMicrurus (coral snakes). Toxicon 1992;30:1299–302.
[19] Ramsey HW, Taylor WJ, Boruchow IB, Snyder GK. Mechanismof shock produced by an elapid snake (Micrurus f. fulvius)venom in dogs. Am J Physiol 1972;222:782.
[20] Tan NH, Ponnudrai G. The biological properties of venoms ofsome American coral snakes (genusMicrurus). Comp BiochemPhysiol Part B: Comp Biochem 1992;101:471–4.
[21] Olamendi-Portugal T, Batista CV, Restano-Cassulini R, PandoV, Villa-Hernandez O, Zavaleta-Martinez-Vargas A, et al.Proteomic analysis of the venom from the fish eating coralsnake Micrurus surinamensis: novel toxins, their function andphylogeny. Proteomics 2008;8:1919–32.
[22] Aird SD, da Silva Jr NJ. Comparative enzymatic composition ofBrazilian coral snake (Micrurus) venoms. Comp BiochemPhysiol Part B: Comp Biochem 1991;99:287–94.
[23] Tanaka GD, de Fátima DF Maria, Portaro FCV, Sant'Anna OA,Tambourgi DV. Diversity of Micrurus snake species related to

1824 J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
their venom toxic effects and the prospective of antivenomneutralization. PLoS Negl Trop Dis 2010;4:1–60.
[24] Serafim FG, Reali M, Cruz-Hofling MA, Fontana MD. Action ofMicrurus dumerilii carinicauda coral snake venom on themammalian neuromuscular junction. Toxicon 2002;40:167–74.
[25] Moraes FV, Sousa-e-Silva MC, Barbaro KC, Leitao MA, FurtadoMF. Biological and immunochemical characterization ofMicrurus altirostris venom and serum neutralization of itstoxic activities. Toxicon 2003;41:71–9.
[26] Dokmetjian JC, Del Canto S, Vinzon S, de Jimenez Bonino MB.Biochemical characterization of the Micrurus pyrrhocryptusvenom. Toxicon 2009;53:375–82.
[27] Corrêa-Netto C, Junqueira-de-Azevedo ILM, Silva DA, Ho PL,Leitão-de-Araújo M, Alves MLM, et al. Snake venomics andvenom gland transcriptomic analysis of Brazilian coral snakes,Micrurus altirostris and M. corallinus. J Proteom in press.
[28] Fernandez J, Alape-Giron A, Angulo Y, Sanz L, Gutirrez JM,Calvete JJ, et al. Venomic and antivenomic analyses of theCentral American coral snake,Micrurus nigrocinctus (Elapidae). JProteom Res 2011;10:1816–27.
[29] Ho PL, Soares MB, Maack T, Gimenez I, Puorto G, Furtado MF,et al. Cloning of an unusual natriuretic peptide from theSouth American coral snake Micrurus corallinus. Eur J Biochem1997;250:144–9.
[30] Silveira de Oliveira J, Rossan de Brandao Prieto da Silva A,Soares MB, Stephano MA, de Oliveira Dias W, Raw I, et al.Cloning and characterization of an alpha-neurotoxin-typeprotein specific for the coral snake Micrurus corallinus.Biochem Biophys Res Commun 2000;267:887–91.
[31] de Oliveira UC, Assui A, da Silva AR, de Oliveira JS, Ho PL.Cloning and characterization of a basic phospholipase A2homologue from Micrurus corallinus (coral snake) venomgland. Toxicon 2003;42:249–55.
[32] da Silva AR Prieto, Yamagushi IK, Morais JF, Higashi HG, Raw I,Ho PL, et al. Cross reactivity of different specific Micrurusantivenom sera with homologous and heterologous snakevenoms. Toxicon 2001;39:949–53.
[33] Lomonte B, Escolano J, Fernandez J, Sanz L, Angulo Y, JMGutierrez, et al. Snake venomics and antivenomics of thearboreal neotropical pitvipers Bothriechis lateralis and Bothrie-chis schlegelii. J Proteome Res 2008;7:2445–57.
[34] Calvete JJ, Borges A, Segura Á, Flores-Díaz M, Alape-Girón A,Gutiérrez JM, et al. Snake venomics and antivenomics ofBothrops colombiensis, a medically important pitviper of theBothrops atrox asper complex endemic to Venezuela: contributingto its taxonomy and snakebite management. J Proteomics2009;72:227–40.
[35] Nuftez V, Cid P, Sanz L, De La Torre P, Angulo Y, Lomonte B,et al. Snake venomics and antivenomics of Bothrops atroxvenoms from Colombia and the Amazon regions of Brazil,Peru and Ecuador suggest the occurrence of geographic variationof venom phenotype by a trend towards paedomorphism. JProteomics 2009;73:57–78.
[36] Kulkeaw K, Chaicumpa W, Sakolvaree Y, Tongtawe P,Tapchaisri P. Proteome and immunome of the venom of theThai cobra, Naja kaouthia. Toxicon 2007;49:1026–41.
[37] Correa-Netto C, Teixeira-Araujo R, Aguiar AS, Melgarejo AR,De-Simone SG, Soares MR, et al. Immunome and venome ofBothrops jararacussu: a proteomic approach to study themolecular immunology of snake toxins. Toxicon 2010;55:1222–35.
[38] RatesB, BemquererMP, RichardsonM,BorgesMH,Morales RAV,De Lima ME, et al. Venomic analyses of scolopendra viridicornisnigra and scolopendra angulata (Centipede, Scolopendromorpha):shedding light on venoms from a neglected group. Toxicon2007;49:810–26.
[39] Pearson WR. Rapid and sensitive sequence comparison withFASTP and FASTA. Methods Enzymol 1990;183:63–98.
[40] Bairoch A, Apweiler R. The SWISS-PROT protein sequencedatabase and its supplement TrEMBL in 2000. Nucleic AcidsRes 2000;28:45.
[41] Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, MillerW, et al. Gapped BLAST and PSI-BLAST: a new generation ofprotein database search programs. Nucleic Acids Res 1997;25:3389.
[42] Rosso JP, Vargas-Rosso O, Gutierrez JM, Rochat H, Bougis PE.Characterization of alpha-neurotoxin and phospholipase A2activities from Micrurus venoms. Determination of the aminoacid sequence and receptor-binding ability of the majoralpha-neurotoxin from Micrurus nigrocinctus nigrocinctus. Eur JBiochem 1996;238:231–9.
[43] Nawarak J, Sinchaikul S, Wu CY, Liau MY, Phutrakul S, ChenST. Proteomics of snake venoms from Elapidae and Viperidaefamilies by multidimensional chromatographic methods.Electrophoresis 2003;24:2838–54.
[44] Nascimento DG, Rates B, Santos DM, Verano-Braga T,Barbosa-Silva A, Dutra AAA, et al. Moving pieces in ataxonomic puzzle: venom 2D-LC/MS and data clusteringanalyses to infer phylogenetic relationships in somescorpions from the Buthidae family (Scorpiones). Toxicon2006;47:628–39.
[45] Liu CS, Chen JM, Chang CH, Chen SW, Teng CM, Tsai IH. Theamino acid sequence and properties of an edema-inducingLys-49 phospholipase A2 homolog from the venom ofTrimeresurus mucrosquamatus. Biochim Biophys Acta(BBA)-Protein Struct Mol Enzym 1991;1077:362–70.
[46] Liu CS, Kuo PY, Chen JM, Chang CH, Tseng CC, TzengMC, et al.Primary structure of an inactive mutant of phospholipase A2in the venom of Bungarus fasciatus (banded krait). J Biochem1992;112:707.
[47] Laing GD, Kamiguti AS, Wilkinson MC, Lowe GM, TheakstonRDG. Characterisation of a purified phospholipase A2 fromthe venom of the Papuan black snake (Pseudechis papuanus).Biochim Biophys Acta (BBA)-Protein Struct Mol Enzym1995;1250:137–43.
[48] Doley R, Mukherjee AK. Purification and characterization ofan anticoagulant phospholipase A2 from Indian monocledcobra (Naja kaouthia) venom. Toxicon 2003;41:81–91.
[49] Chiou JY, Chang LS, Chen LN, Chang CC. Purification andcharacterization of a novel phospholipase A 2 from king cobra(Ophiophagus hannah) venom. J Protein Chem 1995;14:451–6.
[50] Alape-Girón A, Stiles B, Schmidt J, Girón-Cortes M, ThelestamM, Jörnvall H, et al. Characterization of multiple nicotinicacetylcholine receptor-binding proteins and phospholipasesA2 from the venom of the coral snake Micrurus nigrocinctusnigrocinctus. FEBS Lett 1996;380:29–32.
[51] Kihara H, Uchikawa R, Hattori S, Ohno M. Myotoxicity andphysiological effects of three Trimeresurus flavoviridisphospholipases A 2. IUBMB Life 1992;28:895–903.
[52] Kini RM. Structure-function relationships and mechanism ofanticoagulant phospholipase A2 enzymes from snakevenoms. Toxicon 2005;45:1147–61.
[53] Kini RM, Doley R. Structure, function and evolution ofthree-finger toxins-mini proteins with multiple targets.Toxicon 2010.
[54] Kini RM. Venom phospholipase A2 enzymes: structure,function, and mechanism. John Wiley & Son Ltd; 1997.
[55] Casais-e-Silva L, Furtado M. Biochemical and biologicalcharacterization of venomsofMicrurus sp. (Serpentes; Elapidae)from the state of Bahia, Brazil. Toxicon 1996;34:15–6.
[56] Cecchini AL, Marcussi S, Silveira LB, Borja-Oliveira CR,Rodrigues-Simioni L, Amara S, et al. Biological and enzymaticactivities of Micrurus sp. (Coral) snake venoms. CompBiochem Physiol A Mol Integr Physiol 2005;140:125–34.
[57] Sanchez E, Freitas T, Ferreira-Alves D, Velarde D, Diniz M,Cordeiro M, et al. Biological activities of venoms from SouthAmerican snakes. Toxicon 1992;30:95–103.

1825J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 1 8 1 0 – 1 8 2 5
[58] Birrell GW, Earl S, Masci PP, De Jersey J, Wallis TP, Gorman JJ,et al. Molecular diversity in venom from the Australian BrownSnake, Pseudonaja textilis. Mol Cell Proteom 2006;5:379.
[59] Ciscotto P, Machado de Avila R, Coelho E, Oliveira J, Diniz C,Farías L, et al. Antigenic, microbicidal and antiparasiticproperties of an l-amino acid oxidase isolated from Bothropsjararaca snake venom. Toxicon 2009;53:330–41.
[60] Osipov AV, Kasheverov IE, Makarova YV, Starkov VG,Vorontsova OV, Ziganshin RK, et al. Naturally occurringdisulfide-bound dimers of three-fingered toxins. J Biol Chem2008;283:14571.
[61] de Albuquerque Modesto JC, Spencer PJ, Fritzen M, ValençaRC, Oliva MLV, da Silva MB, et al. BE-I-PLA2, a novel acidicphospholipase A2 from Bothrops erythromelas venom: isolation,cloning and characterization as potent anti-platelet andinductor of prostaglandin I2 release by endothelial cells.Biochem Pharmacol 2006;72:377–84.
[62] Francis BR, da Silva Jr NJ, Seebart C, Casais e Silva LL, SchmidtJJ, Kaiser II. Toxins isolated from the venom of the Braziliancoral snake (Micrurus frontalis frontalis) include hemorrhagictypephospholipasesA2andpostsynapticneurotoxins. Toxicon1997;35:1193–203.
[63] Kini RM, Evans HJ. Structure-function relationships ofphospholipases. The anticoagulant region of phospholipasesA2. J Biol Chem 1987;262:14402.
[64] Kondo K, Toda H, Narita K. Characterization of phospholipasea activity of β-bungarotoxin from Bungarus multicinctusvenom: I. Its enzymatic properties and modification withp-bromophenacyl bromide. J Biochem 1978;84:1291.
[65] Sharma S, Karthikeyan S, Betzel C, Singh T. Isolation,purification, crystallization and preliminary X-ray analysis of1-bungarotoxin from Bungarus caeruleus (Indian commonkrait). Acta Crystallogr D Biol Crystallogr 1999;55:1093–4.
[66] Endo T, Tamiya N. Current view on the structure-functionrelationship of postsynaptic neurotoxins from snake venoms.Pharmacol Ther 1987;34:403–51.
[67] Harvey A. Structure-function relationships of postsynapticneurotoxins from snake venoms. Snake toxins 1991:165.
[68] Tsetlin V. Snake venom neurotoxins and other ‘three finger’proteins. Eur J Biochem 1999;264:281–6.
[69] Chen Z, Zhang H, Zhen-lun GU, Chen B, Rong HAN, Reid PF,et al. A long form neurotoxin from cobra venom producespotent opioid independent analgesia. Acta Pharmacol Sin2006;27:402–8.
[70] Moreira KG, Prates MV, Andrade FA, Silva LP, Beirao PS,Kushmerick C, et al. Frontoxins, three-finger toxins fromMicrurus frontalis venom, decrease miniature endplatepotential amplitude at frog neuromuscular junction. Toxicon2010;56:55–63.
[71] Li ZY, Yu TF, Lian ECY. Purification and characterization ofL-amino acid oxidase from king cobra (Ophiophagus hannah)
venomand its effects on human platelet aggregation. Toxicon1994;32:1349–58.
[72] Stabeli RG, Marcussi S, Carlos GB, Pietro RCLR, Selistre-de-AraújoHI. Platelet aggregation and antibacterial effects of anl-aminoacid oxidase purified from Bothrops alternatus snake venom.Bioorg Med Chem 2004;12:2881–6.
[73] Wei XL, Wei JF, Li T, Qiao LY, Liu YL, Huang T, et al.Purification, characterization and potent lung lesion activityof an L-amino acid oxidase from Agkistrodon blomhoffiiussurensis snake venom. Toxicon 2007;50:1126–39.
[74] Sakurai Y, Shima M, Matsumoto T, Takatsuka H, Nishiya K,Kasuda S, et al. Anticoagulant activity of M-LAO,-amino acidoxidase purified from Agkistrodon halys blomhoffii, throughselective inhibition of factor IX. Biochim Biophys Acta(BBA)-Proteins Proteom 2003;1649:51–7.
[75] Izidoro LFM, Ribeiro MC, Souza GRL, Sant'Ana CD, HamaguchiA, Homsi-Brandeburgo MI, et al. Biochemical and functionalcharacterization of an L-amino acid oxidase isolated fromBothrops pirajai snake venom. Bioorg Med Chem 2006;14:7034–43.
[76] Zhang H, Teng M, Niu L, Wang Y, Liu Q, Huang Q, et al.Purification, partial characterization, crystallization andstructural determination of AHP-LAAO, a novel L-amino-acidoxidase with cell apoptosis-inducing activity fromAgkistrodonhalys pallas venom. Acta Crystallogr D Biol Crystallogr 2004;60:974–7.
[77] Torii S, NaitoM, Tsuruo T. Apoxin I, a novel apoptosis-inducingfactor with L-amino acid oxidase activity purified fromWestern diamondback rattlesnake venom. J Biol Chem1997;272:9539.
[78] Leão L, Ho P, de Junqueira-de-Azevedo I. Transcriptomic basisfor an antiserum against Micrurus corallinus (coral snake)venom. BMC Genomics 2009;10:112.
[79] Ménez A. Functional architectures of animal toxins: a clue todrug design? Toxicon 1998;36:1557–72.
[80] Chippaux JP, Goyffon M. Venoms, antivenoms andimmunotherapy. Toxicon 1998;36:823–46.
[81] Espino-Solis G, Riaño-Umbarila L, Becerril B, Possani L.Antidotes against venomous animals: state of the art andprospectives. J Proteomics 2009;72:183–99.
[82] Theakston R, Warrell D, Griffiths E. Report of a WHOworkshop on the standardization and control of antivenoms.Toxicon 2003;41:541–57.
[83] Gutiérrez JM, Lomonte B, León G, Alape-Girón A, Flores-Díaz M,Sanz L, et al. Snake venomics and antivenomics: proteomictools in the design and control of antivenoms for the treatmentof snakebite envenoming. J Proteomics 2009;72:165–82.
[84] Raw IG, R., Higashi HG, Kelen EM. Antivenins in Brazil:preparation. In: AT T, editor. Handbook of natural toxinsreptile venoms and toxins. New York: Marcel Dekker; 1991.p. 557–81.





























