Embed Size (px)
Citation preview

The elevation gradient of lichen species richness in Nepal
Chitra Bahadur BANIYA, Torstein SOLHØY, Yngvar GAUSLAAand Michael W. PALMER
Abstract: This study of elevation gradients of lichen species richness in Nepal aimed to comparedistribution patterns of different life-forms, substratum affinities, photobiont types, and Nepaleseendemism. Distribution patterns of lichens were compared with elevational patterns shown by a widerange of taxonomic groups of plants along the Nepalese Himalayan elevational gradient between200–7400m. We used published data on the elevation records of 525 Nepalese lichen species tointerpolate presence between the maximum and minimum recorded elevations, thereby givingestimates of lichen species richness at each 100-m elevational band. The observed patterns werecompared with previously published patterns for other taxonomic groups. The total number of lichensas well as the number of endemic species (55 spp.) showed humped relationships with elevation. Theirhighest richness was observed between 3100–3400 and 4000–4100m, respectively. Almost 33% of thetotal lichens and 53% of the endemic species occurred above the treeline (>4300m). Non-endemicrichness had the same response as the total richness. All growth forms showed a unimodal relationshipof richness with elevation, with crustose lichens having a peak at higher elevations (4100–4200m) thanfruticose and foliose lichens. Algal and cyanobacterial lichen richness, as well as corticolous lichenrichness, all exhibited unimodal patterns, whereas saxicolous and terricolous lichen richness exhibitedslightly bimodal relationships with elevation. The highest lichen richness at mid altitudes concurredwith the highest diversity of ecological niches in terms of spatial heterogeneity in rainfall, temperature,cloud formation, as well as high phorophyte abundance and diversity implying large variation in barkroughness, moisture retention capacity, and pH. The slightly bimodal distributions of saxicolous andterricolous lichens were depressed at the elevational maximum of corticolous lichens.
Key words: altitude, endemism, Himalaya
Introduction
The Himalayas are the highest mountainrange in the world and show a rich diversityof eco-climatic zones. In such a landscape,elevational gradients are of particular interestin applied as well as in theoretical ecology(Wolf 1993). The pattern of changes inspecies-richness with elevation characterizes
the vegetation in a simple but powerful way.For example, Yoda (1967) found graduallydecreasing tree species richness with increas-ing altitude. Later, Hunter & Yonzon (1993)reported a similar pattern for Nepalese birdsand mammals. A number of studies haveproduced macro-scale elevational gradientsfor the whole of Nepal, using interpolationmethods on data from secondary sources.Several groups of organisms have beenstudied and exhibit maximum richness atgroup-specific intermediate elevations, re-ferred to as a unimodal, or humped, eleva-tion pattern. For example, Himalayanvascular plants showed their highest totalrichness at altitudes between 1500 to 2500mand their highest endemic richness at 4000m(Grytnes & Vetaas 2002; Vetaas & Grytnes2002), while fern richness peaked at 2000m(Bhattarai et al. 2004). However, orchids
C. B. Baniya (corresponding author) and T. Solhøy:Department of Biology, University of Bergen, Allégaten41, P.O. Box 7803, N-5020 Bergen, Norway. Email:[email protected] address: Central Department of Botany,Tribhuvan University Kirtipur, Kathmandu, Nepal.Y. Gauslaa: Department of Ecology and NaturalResource Management, Norwegian University of LifeSciences, P.O. Box 5003, NO-1432 Ås, Norway.M. W. Palmer: Department of Botany, OklahomaState University, 104 LSE Stillwater OK 74078USA 405-744-7717.
The Lichenologist 42(1): 83–96 (2010) © British Lichen Society, 2009doi:10.1017/S0024282909008627

showed their maximum richness at 1600mwhereas the highest orchid endemic richnesswas bimodal with two separate peaks at 1200and 3300m (Acharya 2008). Liverworts andmosses had their maximum richness at 2800and 2500m, respectively, and the peak forendemic liverwort richness occurred at3300m (Grau et al. 2007).
Relatively few studies on elevational rich-ness patterns for lichens have been under-taken (Kessler 2000; Bruun et al. 2006;Grytnes et al. 2006) and among terrestrialphotosynthetic organisms they are the onlymajor group that has not yet been investi-gated in this respect in Nepal. They areamong the most successful organisms in ex-treme environments such as cold arctic andalpine environments where few other plantscan grow (e.g. Schroeter et al. 1994; Kappenet al. 1996). Lichens also show a high diver-sity as eipiphytes and thus may benefit fromhigh diversity of trees and shrubs withspecies-specific bark chemistry, texture andstability (Kessler 2000; Bruun et al. 2006;Grytnes et al. 2006). This paper describesand interprets the Himalayan elevational dis-tribution of lichen species richness in Nepal.
Material and Methods
Study area
Nepal is a mountainous country in the centralHimalayas between 26°22# to 30°27# N and 80°04# to88°12# E with an area of 147 181 km2. The elevationranges from 60m above sea level to 8848m (Mt. Everest).The country is surrounded by the Indo-Gangetic Plainin the east, west and south, and the Tibetan Plateautowards the north. Parallel mountain ranges run south-east and north-west dissecting the country into north-south running river valleys that are associated with fertileplains, inner valleys, lakes, and the outer flood plains.
The Indian monsoon beginning in May and ending inSeptember is the main source of precipitation (morethan 80%). Monsoon rain forms after condensation ofwater vapour arising from the Bay of Bengal, and con-sequently there is a spatial gradient of monsoon rainfalldecreasing from east to west. Precipitation also occurs inwinter formed from central Asian water vapour anddiminishes eastwards.
Physiographically, Nepal has been divided into fivezones (Hagen 1969; Upreti 1999). 1) A 30–40km widesouthernmost plain is called Terai between 60 and 300m.It is the most fertile, densely populated zone with a tropi-cal climate (between 22–27°C during winter and above
35°C during the warmest summer months); it is mainlycultivated but has still some tropical forests. 2) TheSiwalik located from 700 to 1500m in the foothills ofthe Mahabharat Range. 3) The Mahabharat Range liesbetween 1500 and 3000m; it belongs to the Midlands orlesser Himalaya and includes the principal inner valleyssuch as Kathmandu and Pokhara. The climate is mildthroughout the year and receives the highest annual rain-fall (above 4000mm) south of Annapurna, central Nepal.It includes the lower cloud zone at 2000 to 2500m (Miehe1989, 1990). 4) The Greater-Himalaya extends above2700 m. Here the climate varies from dry summers to coldwinters with snow, and subzero temperatures on themountain tops. This zone includes the uppermost limit ofcloud-zone forest in Nepal at 4000m (Miehe 1989, 1990).5) Trans-Himalaya lies towards the northern side of theGreater-Himalaya with elevations above 3000m. It is inthe rain-shadow zone with an average annual rainfallbetween 200 and 400mm.
Vegetation is one of the prime determinants of lichendiversity and distribution (Awasthi 2007). In Nepal,vegetation has been studied and classified into 8 eleva-tional forest zones (Yoda 1967; Stainton 1972, 2001;Dobremez & Jest 1969; Dobremez 1976; Miehe 1982,1989). 1) Tropical forest between 60 and 1000 m(Stainton 1972) dominated by Shorea robusta Gaertn.,Dalbergia sissoo Roxb., Adina cordifolia (Willd. ex Roxb.)Benth. & Hook. f. ex Brandis, Terminalia spp., Lager-stroemia spp., Michelia champaca L., and Bombax ceiba L.2) Subtropical forest occurring between 1000 to 2000mwith Pinus roxburghii Sarg. as the dominant species onsouth-facing slopes in the Midlands. 3) Lower temper-ate broad-leaved forest at 1700–2400m in the eastand 2000–2700m in the west. Dominant trees includeAlnus nitida (Spach.) Endel., Castanopsis tribuloides A.DC., Castanopsis hystrix A. DC., Lithocarpus pachyphylla(Kurz.) Rehder., and Quercus spp. Moister slopes be-tween 1700–2200m are dominated by Cinnamomumspp. and represent the lower temperate mixed broad-leaved forest. Upper temperate broad-leaved forests arerepresented by various types. Drier southern slopes at2200–3000m are dominated by Schima wallichii (DC.)Korth., Castanopsis indica (Roxb.) Miq., and Quercussemecarpifolia J. E. Smith. Moist, north and westslopes between 2500–3500m from central to eastNepal are dominated by Acer spp., Rhododendron spp.,Aesculus spp. and Juglans spp. 4)Temperate coniferousforests between 2000 and 3700m dominated by Pinuswallichiana A. B. Jack. Other conifers include Cedrusdeodara (Roxb.) G. Don, Picea smithiana (Wall.) Boiss.,Juniperus indica Bertol., Larix himalaica W. C. Cheng &L. K. Fu, Larix griffithiana Carriere, Cupressus torulosa D.Don, and Tsuga dumosa (D. Don) Eichler. 5) Sub-alpineforests occurring between 3000 and 4100m (Stainton1972) dominated by Betula utilis D. Don, Abies spectabilis(D. Don) Mirb., and Rhododendron spp. This forestincludes the climatic treeline at 4100–4300m. 6) Open,low-alpine shrub communities including Caragana spp.,Lonicera spp., Rosa spp., Sophora spp., Rhododendronanthopogon D. Don, R. lepidotum Wall. ex G. Don,Ephedra gerardiana Wall. ex C. A. Meyer, and Hippophaetibetana Schldl. 7) Mid-alpine zone with dwarf-shrubs
84 THE LICHENOLOGIST Vol. 42

but largely dominated by herb communities. 8) Nivalzone begins above the mid-alpine zone with morelichens and fewer mosses and angiosperms. Within thenival zone, there are vascular plants of open wind ex-posed areas as well as those tolerant of snow-lie.
Data sources
The main source of data for this study is the Lichens ofNepal (Sharma 1995). However, many species reportedin Sharma (1995) lack elevation data and since thenmany new species have been described. Other relevantand easily accessible literature as well as the recent keysfor both microlichens and macrolichens of Nepal,India and Sri Lanka by Awasthi (1991; 2007) have alsobeen checked. In addition, the following literature onNepalese lichens (Lamb 1966; Poelt 1966a, b, 1974;Bystrek 1969; Poelt & Reddi 1969; Abbayes 1974; Jahns& Seelen 1974; Kurokawa 1974; Mitchell 1974;Schmidt 1974; Vezda & Poelt 1975; Hellmich & Poelt1977; Awasthi 1986; Vitikainen 1986; Awasthi &Mathur 1987; Miehe 1990; Awasthi 1991, 2007;Esslinger & Poelt 1991; Poelt & Hinteregger 1993;Sharma 1995; Baniya 1996; Jørgensen 2001) have pro-vided information on the elevation ranges of Nepaleselichens. These sources have provided elevational distri-bution data for 525 species, including 55 Nepaleseendemics (Appendix 1). Among the 525 species, 172,including 29 endemics, are reported from above thetreeline (> 4300m). In addition, data about life-forms(crustose, foliose and fruticose), photobiont types(cyanobacteria and green algae) and five substratumtypes (tree bark, wood, mosses, rocks and soil) wereobtained. Tripartite lichens were treated as cyanobacte-rial. There were 50 species of lichens for which eleva-tional data or other important information are eitherincomplete or lacking; these species were excluded fromthe analysis. Lignicolous and muscicolous substrata hadbeen reported for a few species (5 and 17, respectively)and were thus also excluded during the analyses involv-ing substrata. Lichen nomenclature follows Awasthi(1991; 2007).
The altitudinal range of lichens in Nepal, 200–7400m, was divided into 73 bands of 100m each and acomplete data matrix for all species was assembled.Presence of a species indicates that the species occurs in,or has been collected in the past, from that elevationband and absence means either that the species does notoccur or has previously not been collected from thatelevation. A species is assumed to be present in allpossible 100m bands between its upper and lower eleva-tion limits. For example, a lichen that has elevationaloccurrences between 210 to 451m in the literature fallsbetween the 200- and 500-m bands. A list of the lichensincluded in this study together with their elevationranges is provided in Appendix 1.
Species richness is an estimate of the total number oflichen species occurring in each 100m elevation band.This is a macro-scale study (gamma diversity, sensuWhittaker 1972) that covers the entire elevational rangeof Nepal. Fifty-six taxa, varieties, forms or subspecies,were treated as species (Appendix 1).
Data analysis
Patterns related to total lichen species richness andtheir sub-sets (life forms, algal partners, and substrata)as responses and their elevations as a predictor vari-able were extracted by using a cubic smooth spline (s)within the framework of Generalized Additive Models(GAM, Hastie & Tibshirani 1990; Heegaard 2004) witha default of c. 4 degrees of freedom. Response variablesare counts; thus, the variance changes with the mean andnegative predictions are meaningless. We found over-dispersion in our data. We thus applied a Quasi-poissonfamily error distribution with a logarithmic link function(Crawley 2006). We confirmed our assumption ofnormal distribution of error after the Q-Q diagnosticplots plotted against residuals. The change in deviancefollows the F-distribution. We used R 2.7.0 (R Devel-opment Core Team 2008) to analyse our data andsmoothers were fitted with library GAM (Hastie &Tibshirani 1990). GAM was used because it is anon-parametric approach that does not make a prioriassumptions about the species-elevational relationship.
One of the main purposes of this study is to contrastobserved lichen richness patterns with other studies.Biogeographic data have biases and constraints, such asa focus on specific taxa and/or a restriction to easilyaccessible landscapes. However, these problems areshared among data sets with various taxonomic groups(Grytnes & Vetaas 2002; Vetaas & Grytnes 2002;Bhattarai et al. 2004; Grau et al. 2007). Thus, a com-parison of empirical patterns among available studies ispossible.
Results
The 525 lichen species recorded (Appendix1) represented 40 families and 121 genera.Among these lichens, 35·4% were crustose,46·3% were foliose and 18·3% were fruti-cose. A total of 12% had a cyanobacterialphotobiont and 88% had a green algal photo-biont. Lichens endemic to Nepal (n=55) rep-resented 10% of the total Nepalese lichenflora. There were 172 species reported fromabove the treeline (R 4300m) and 29 ofthese were endemics. Carbonea vorticosa inNepalese Himalaya at 7400m was theworld’s highest reported lichen. Heterodermiapseudospeciosa represented one of the lichensoccurring at the lowest elevation ranges inNepal (150–2100m).
The total lichen species richness showeda unimodal relationship with elevation. Themaximum modelled total richness occurredat 3100–3400m (Fig. 1A), whereas theobserved maximum richness (144 species)
2010 Elevation gradients of lichens in Nepal—Baniya et al. 85

was at approximately 4000m. Endemiclichens also had a unimodal relationship withelevation, but their maximum modelledrichness occurred at 3900–4400 m (Fig. 1B)approximately 800m higher than the peakfor total species. Non-endemic richnessmatched closely the elevational patterns ofthe total species richness (data not shown).
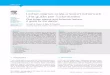
Species richness of all three growth formsof lichens had unimodal responses to eleva-tion (Fig. 2A–C). Among the three morpho-logical types, crustose lichens peaked at thehighest altitude (4100–4200m) while folioselichens had their maximum richness atconsistently lower altitudes (2400–2500m).Fruticose lichens peaked at an intermediateelevation (3200m). With respect to photo-biont type, cyanolichens exhibited maximumrichness at lower altitudes (2900–3000m;Fig. 3A) than green algal lichens (3300–3500m; Fig. 3B).
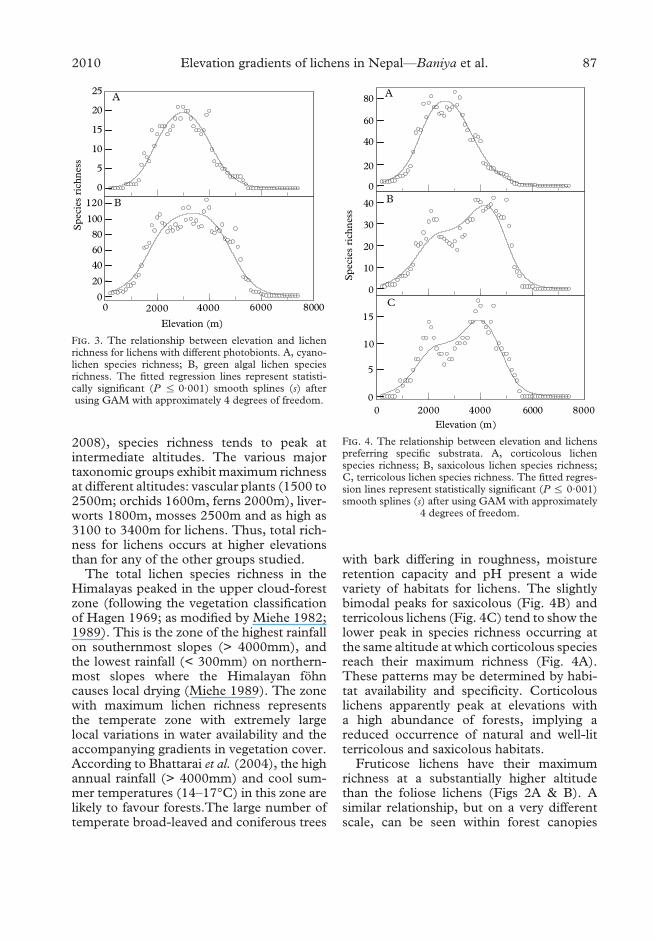
Among the five substratum categoriesstudied, corticolous lichens had a clear uni-modal relationship to elevation, with thehighest species richness occurring between2500 and 2700m (Fig. 4A) in the lower partof the temperate forests. Saxicolous and ter-ricolous lichens had slightly bimodal patterns
with a prominent hump at 3900–4200m(Fig. 4B & C).
The regression analysis results showingthe best selected model for each responsevariable are recorded in Appendix 2.
Discussion
Total lichen species richness in Nepal variesstrongly with elevation (Figs 1A & B) in linewith previous findings for vascular plants(Grytnes & Vetaas 2002; Vetaas & Grytnes2002; Bhattarai & Vetaas 2003; Bhattaraiet al. 2004) including orchids (Acharya2008), and for bryophytes (Grau et al. 2007).In all these studies, as well as studies dealingwith lichens from other countries (Wolseley& Aguirre-Hudson 1997; Negi 2000, 2003;Wolf & Alejandro 2003; Pinokiyo et al.
F!". 1. Relationship between elevation and lichenspecies richness in Nepal. A, total lichen species rich-ness; B, total endemic lichen species richness. Thefitted regression lines represent statistically significant(P % 0·001) smooth spline (s) after using GAM with
approximately 4 degrees of freedom.
F!". 2. Relationship between elevation and lichenspecies life-form richness in Nepal. A, foliose lichenspecies richness; B, fruticose lichen species richness; C,crustose lichen species richness. The fitted regressionlines represent statistically significant (P % 0·001)smooth spline (s) after using GAM with approximately 4
degrees of freedom.
86 THE LICHENOLOGIST Vol. 42
2008), species richness tends to peak atintermediate altitudes. The various majortaxonomic groups exhibit maximum richnessat different altitudes: vascular plants (1500 to2500m; orchids 1600m, ferns 2000m), liver-worts 1800m, mosses 2500m and as high as3100 to 3400m for lichens. Thus, total rich-ness for lichens occurs at higher elevationsthan for any of the other groups studied.
The total lichen species richness in theHimalayas peaked in the upper cloud-forestzone (following the vegetation classificationof Hagen 1969; as modified by Miehe 1982;1989). This is the zone of the highest rainfallon southernmost slopes (> 4000mm), andthe lowest rainfall (< 300mm) on northern-most slopes where the Himalayan föhncauses local drying (Miehe 1989). The zonewith maximum lichen richness representsthe temperate zone with extremely largelocal variations in water availability and theaccompanying gradients in vegetation cover.According to Bhattarai et al. (2004), the highannual rainfall (> 4000mm) and cool sum-mer temperatures (14–17°C) in this zone arelikely to favour forests.The large number oftemperate broad-leaved and coniferous trees
with bark differing in roughness, moistureretention capacity and pH present a widevariety of habitats for lichens. The slightlybimodal peaks for saxicolous (Fig. 4B) andterricolous lichens (Fig. 4C) tend to show thelower peak in species richness occurring atthe same altitude at which corticolous speciesreach their maximum richness (Fig. 4A).These patterns may be determined by habi-tat availability and specificity. Corticolouslichens apparently peak at elevations witha high abundance of forests, implying areduced occurrence of natural and well-litterricolous and saxicolous habitats.
Fruticose lichens have their maximumrichness at a substantially higher altitudethan the foliose lichens (Figs 2A & B). Asimilar relationship, but on a very differentscale, can be seen within forest canopies
F!". 3. The relationship between elevation and lichenrichness for lichens with different photobionts. A, cyano-lichen species richness; B, green algal lichen speciesrichness. The fitted regression lines represent statisti-cally significant (P % 0·001) smooth splines (s) afterusing GAM with approximately 4 degrees of freedom.
F!". 4. The relationship between elevation and lichenspreferring specific substrata. A, corticolous lichenspecies richness; B, saxicolous lichen species richness;C, terricolous lichen species richness. The fitted regres-sion lines represent statistically significant (P % 0·001)smooth splines (s) after using GAM with approximately
4 degrees of freedom.
2010 Elevation gradients of lichens in Nepal—Baniya et al. 87

where the biomass of fruticose lichens in-creases and that of foliose lichens decreaseswith height above the ground (Goward 1998;Campbell & Coxson 2001; Gauslaa et al.2008). In well-lit and open high altitude for-ests and the upper canopies of trees, fruticoselichens may have the advantage in being ableto utilise light from all directions, whereasmany foliose and flat lichens maximize theharvest of more or less unidirectional light inshady positions in dense forests and on lowcanopy branches (Gauslaa et al. 2009). Thefoliose lichen richness peaks in the same zoneas the highest richness for mosses (Grau et al.2007), taxa that are also relatively shadetolerant. Furthermore, finely dissected fruti-cose lichens on twigs lack thick boundarylayers. Thus, they are more closely coupledto ambient air than flat foliose lichens andabsorb water vapour more readily from theair as discussed by Jonsson et al. (2008). Bycontrast, flat lichens and bryophyte carpetsare more dependent on rainwater.
The elevation range between 4000 to4300m is the sub-alpine zone of Nepal(Stainton 1972). This humid upper-cloudzone (Miehe 1989) is densely colonized byepiphytic mosses reflecting the high humidity(Grau et al. 2007). This humidity may alsofacilitate high lichen richness associated withopen canopies with more light facilitatinglichen growth (Gauslaa et al. 2007).
A high degree of isolation creates ende-mism (Cox & Moore 2000), and isolationmight increase with elevation. High moun-tain peaks with an exceptionally cold andharsh climate act as islands (Grau et al.2007), and phytogeographical isolationcan lead to localized speciation events. Theoccurrence of 29 endemic species (17%) outof the 172 total species above the treeline(R 4300 m) emphasises the link betweenelevation and endemism. The Nepaleselichen endemism rate is higher than that formosses (3 out of 480 spp. i.e., < 1%) andliverworts (33 out of 368 spp. i.e., 9%), butslightly less than that for vascular plants (303out of 1957 spp. i.e., 16%). Above 4500mboth total and endemic lichen diversity (Figs1A & B) decline until 6100m, which is thehighest reported elevation for lichens en-
demic to Nepal. Cold winds leading to rapidfreezing and drying may reduce competitionfrom surviving vascular plant species (Grime1977). The harsh climate might lead to vari-ous specializations and thus facilitate forma-tion of endemic species (Aptroot & Bungartz2007).
Land area might impose a unimodal rich-ness pattern. In general, the larger the areasampled, the greater will be the numberof species encountered (Rosenzweig 1995;Qian et al. 2007). A decreasing trend in landarea in each 500m band occurs with increas-ing elevation (Vetaas & Grytnes 2002) asestimated from the Digitized Map by theInternational Centre for Integrated Moun-tain Development (ICIMOD, Kathmandu).This is not always a common pattern (Körner2007). If we consider species-area relation-ships, more species should be expected to-wards lower elevations than we observed.However, lichen diversity can be greatlylimited by lack of long ecological continuityin forests (Rose 1976) and lichens may beless prevalent at low altitudes because ofdense forest canopies and intensive land-use.Furthermore, as lichen species richness alsotends to be universally greater in cool or coldclimates (Mattick 1953), we believe that thestrong reduction in lichen richness at lowelevations is real and not a sampling artefact.
In conclusion, lichens and various sub-groups of lichens exhibit unimodal patternssimilar to those found in other major taxo-nomic groups, but the highest total lichenrichness peaks at higher altitudes than inany other group. The maximum lichen rich-ness occurred at the altitude with the highestdiversity of ecological niches in terms ofspatial heterogeneity, rainfall, temperature,cloud formation, as well as high phorophyteabundance and diversity implying large vari-ation in bark roughness, moisture retentioncapacity, and pH.
We are grateful to the Norwegian State Education LoanFund (Lånekassen) for providing funding and theCentral Department of Botany, Tribhuvan University,Nepal, for providing study leave for CBB. Thanks aredue to John Birks for his valuable comments, WalterObermayer and Per Magnus Jørgensen for provision ofliterature on lichens and to Govind Ghimire and Bijaya
88 THE LICHENOLOGIST Vol. 42

Kattel for their feedback. Thanks are also due to LouiseOlley, Teuvo Ahti and Tor Tönsberg for their help inrevising the species list and to the Senior Editor, PeterCrittenden and two anonymous referees for their usefulcomments and suggestions.
R#$#%#&'#(
Abbayes, H. D. (1974) Cladonia du Nepal. In KhumbuHimal 6 (W. Hellmich & J. Poelt, eds): 111–116.München: Universitätsverlag Wagner Ges.M. B. H. Innsbruck.
Acharya, K. P. (2008) Orchid species richness along aHimalayan elevation gradient. M. Sc. thesis, BergenUniversity.
Aptroot, A. & Bungartz, F. (2007) The lichen genusRamalina on the Galapagos. Lichenologist 39: 519–542.
Awasthi, G. (1986) Lichen genus Usnea in India. Journalof the Hattori Botanical Laboratory 61: 333–421.
Awasthi, D. D. (1991) A key to the microlichens ofIndia, Nepal and Sri Lanka. Bibliotheca Licheno-logica 40: 1–337.
Awasthi, D. D. (2007) A Compendium of the Macrolichensfrom India, Nepal and Sri Lanka. Dehra Dun: BishenSingh & Mahendra Pal Singh.
Awasthi, D. D. & Mathur, R. (1987) Species of thelichen genera Bacidia, Badimia, Fellhanera andMycobilimbia from India. Proceedings of Indian Acad-emy of Sciences, Bangalore. 97: 481–503.
Baniya, C. B. (1996) Floristic composition of lichens ofSikles (Kaski) and Shivapuri (Kathmandu) andtheir ecology. M. Sc. thesis, Tribhuvan University,Kathmandu, Nepal.
Bhattarai, K. R. & Vetaas, O. R. (2003) Variation inplant species richness of different life forms along asubtropical elevation gradient in the Himalayas,east Nepal. Global Ecology and Biogeography 12:327–340.
Bhattarai, K. R., Vetaas, O. R. & Grytnes, J. A. (2004)Fern species richness along a central Himalayanelevational gradient, Nepal. Journal of Biogeography31: 389–400.
Bruun, H. H., Moen, J., Virtanen, R., Grytnes, J. A.,Oksanen, L. & Angerbjörn, A. (2006) Effects ofaltitude and topography on species richness ofvascular plants, bryophytes and lichens in alpinecommunities. Journal of Vegetation Science 17:37–46.
Bystrek, J. L. (1969) Die Gattung Alectoria. In KhumbuHimal 6 (W. Hellmich & J. Poelt, eds): 17–24.München: Universitätsverlag Wagner Ges.M. B. H. Innsbruck.
Campbell, J. & Coxson, D. S. (2001) Canopy micro-climate and arboreal lichen loading in subalpinespruce-fir forest. Canadian Journal of Botany 79:537–555.
Cox, C. B. & Moore, P. D. (2000) Biogeography: anEcological and Evolutionary Approach. London:Blackwells.
Crawley, M. J. (2006) Statistics: an Introduction using R.London: John Wiley & Sons, Ltd.
Dobremez, J. F. (1976) Le Nepal: Écologie et Bio-geographie. Paris: Centre National de la RechercheScientifique.
Dobremez, J. F. & Jest, C. (1969) Carte Écologique de laRegion Annapurna Dhaulagiri. Paris: CentreNational de la Recherche Scientifque.
Esslinger, T. L. & Poelt, J. (1991) Parmelia masonii,a new lichen species (Ascomycota) from theHimalayas. Bryologist 94: 203–206.
Gauslaa, Y., Palmqvist, K., Solhaug, K. A., Holien, H.,Hilmo, O., Nybakken, L., Myhre, L. C. & Ohlson,M. (2007) Growth of epiphytic old forestlichens across climatic and successional gradients.Canadian Journal of Forest Research 37: 1832–1845.
Gauslaa, Y., Lie, M. & Ohlson, M. (2008) Epiphyticlichen biomass in a boreal Norway spruce forest.Lichenologist 40: 257–266.
Gauslaa, Y., Palmqvist, K., Solhaug, K. A., Hilmo, O.,Holien, H., Nybakken, L. & Ohlson, M. (2009) Sizedependent growth in two old-growth associatedmacrolichen species. New Phytologist 18: 683–692.
Goward, T. (1998) Observations on the ecology ofthe lichen genus Bryoria in high elevation coniferforests. Canadian Field Naturalist 112: 496–501.
Grau, O., Grytnes, J.-A. & Birks, H. J. B. (2007) Acomparison of altitudinal species richness patternsof bryophytes with other plant groups in Nepal,Central Himalaya. Journal of Biogeography 34:1907–1915.
Grime, J. P. (1977) Evidence for the existence of threeprimary strategies in plants and its relevance toecological and evolutionary theory. AmericanNaturalist 11: 1169–1194.
Grytnes, J. A., Heegaard, E. & Ihlen, P. G. (2006)Species richness of vascular plants, bryophytes, andlichens along an altitudinal gradient in westernNorway. Acta Oecologica 29: 241–246.
Grytnes, J. A. & Vetaas, O. R. (2002) Species richnessand altitude: a comparison between null modelsand interpolated plant species richness along theHimalayan altitudinal gradient, Nepal. AmericanNaturalist 159: 294–304.
Hagen, T. (1969) Report on the Geological Survey ofNepal: Preliminary Reconnaissance. Zurich: OrellFüssli Arts Graphiques.
Hastie, T. J. & Tibshirani, R. J. (1990) GeneralisedAdditive Models. London: Chapman & Hall.
Heegaard, E. (2004) Trends in aquatic macrophytespecies turnover in Northern Ireland – which factorsdetermine the spatial distribution of local speciesturnover? Global Ecology and Biogeography 13:397–408.
Hellmich, W. & Poelt, J. (eds) (1977). Khumbu Himal 6.München: Universitätsverlag Wagner Ges.M. B. H. Innsbruck.
Hunter, M. L. & Yonzon, P. (1993) Altitudinal distri-butions of birds, mammals, people, forests, andparks in Nepal. Conservation Biology 7: 420–423.
Jahns, H. M. & Seelen, E. J. R. (1974) Baeomyces-Fundeaus dem Himalaya. In Khumbu Himal 6 (W.Hellmich, & J. Poelt, eds): 101–108. München:Universitätsverlag Wagner Ges. M. B. H. Innsbruck.
2010 Elevation gradients of lichens in Nepal—Baniya et al. 89

Jonsson, A. V., Moen, J. & Palmqvist, K. (2008) Pre-dicting lichen hydration using biophysical models.Oecologia 156: 259–273.
Jørgensen, P. M. (2001) The lichen genus Erioderma(Pannariaceae) in China and Japan. AnnalesBotanici Fennici 38: 259–264.
Kappen, L., Schroeter, B., Scheidegger, C.,Sommerkorn, M. & Hestmark, G. (1996) Coldresistance and metabolic activity of lichens below0°C. In Life Sciences: Space and Mars Recent Results18: 119–128. Oxford: Pergamon Press Ltd.
Kessler, M. (2000) Altitudinal zonation of Andeancryptogam communities. Journal of Biogeography27: 275–282.
Körner, C. (2007) The use of ‘altitude’ in ecologicalresearch. Trends in Ecology and Evolution 22:569–574.
Kurokawa, S. (1974) Anaptychia of the Nepal-Himalaya. In Khumbu Himal 6 (W. Hellmich, & J.Poelt, eds): 109–110. München: UniversitätsverlagWagner Ges. M. B. H. Innsbruck.
Kuusinen, M. (1996) Epiphyte flora and diversity onbasal trunks of six old-growth forest tree species insouthern and middle boreal Finland. Lichenologist28: 443–463.
Lamb, L. M. (1966) Die Gattung Stereocaulon. InKhumbu Himal 1 (W. Hellmich, ed): 349–352.München: Universitätsverlag Wagner Ges.M. B. H. Innsbruck.
Mattick, F. (1953) Lichenologisches Notizen. Berichteder Deutschen Botanischen Gesellschaft 66: 263–276.
Miehe, G. (1982) Vegetationsgeographische Untersuc-hungen im Dhaulgiri und Annapurna Himalaya,Dissertationes Botanicæ. Journal Cramer, Vaduz 66:66.
Miehe, G. (1989) Vegetation patterns in Mount Everestas influenced by monsoon and föhn. Vegetatio 79:21–32.
Miehe, G. (1990) Langtang himal-flora und vegetation alsklimazeiger und-zeugen im Himalaya. Ph. D. thesis,Universität Göttingen.
Mitchell, M. (1974) Die Gattung Leptogium sect. Mal-lotium im Himalaya. In Khumbu Himal 6 (W.Hellmich, & J. Poelt, eds): 121–126. München:Universitätsverlag Wagner Ges. M. B. H.Innsbruck.
Negi, H. (2000) On the patterns of abundance anddiversity of macrolichens of Chopta-Tunganath inthe Garhwal Himalaya. Journal of Biosciences 25:367–378.
Negi, H. (2003) Lichens: a valuable bioresource forenvironmental monitoring and sustainable develop-ment. Resonance 8: 51–58.
Pant, G. & Awasthi, D. D. (1989a) Caliciales from Indiaand Nepal. Biovigyanam 15: 3–27.
Pant, G. & Awasthi, D. D. (1989b) Lichen genus Cati-llaria s. lat. in India. Proceedings of Indian Academy ofSciences (Plant Science). 99: 369–384.
Pinokiyo, A., Singh, K. P. & Singh, J. S. (2008) Diver-sity and distribution of lichens in relation to altitudewithin a protected biodiversity hotspot, north-eastIndia. Lichenologist 40: 47–62.
Poelt, J. (1966a) Die Gattung Ochrolechia. In KhumbuHimal 1 (W. Hellmich, ed): 251–261. München:Universitätsverlag Wagner Ges. M. B. H.Innsbruck.
Poelt, J. (1966b) Die Lobaten Arten der SammelgattungLecanora, Lecanoraceae. In Khumbu Himal 3 (W.Hellmich, ed): 187–202. München: Universitäts-verlag Wagner Ges. M. B. H. Innsbruck.
Poelt, J. (1974) Die Gattungen Physcia, Physciopsis undPhysconia. In Khumbu Himal 6 (W. Hellmich, & J.Poelt, eds): 57–99. München: UniversitätsverlagWagner Ges. M. B. H. Innsbruck.
Poelt, J. & Hinteregger, E. (1993) Beiträge zur Kenntnisder Flechtenflora des Himalaya. Bibliotheca Licheno-logica 50: 1–247.
Poelt, J. & Reddi, B. V. (1969) Candelaria und Cande-lariella. In Khumbu Himal 6 (W. Hellmich, & J.Poelt, eds): 1–16. München: UniversitätsverlagWagner Ges. M. B. H. Innsbruck.
Qian, H., Fridley, J. D. & Palmer, M. W. (2007) Thelatitudinal gradient of species-area relationshipsfor vascular plants of North America. AmericanNaturalist 170: 690–701.
R Development Core Team (2008) R: A language andenvironment for statistical computing version 2.7.0.
Rose, F. (1976) Lichenological indicators of age andenvironmental continuity in woodlands. InLichenology: Progress and Problems (D. H. Brown,D. L Hawksworth & R. H. Bailey, eds): 279–307.London: Academic Press.
Rosenzweig, M. L. M. L. (1995) Species Diversity inSpace and Time. Cambridge: Cambridge UniversityPress.
Schmidt, A. (1974) Chaenotheca und Coniocybe. InKhumbu Himal 6 (W. Hellmich, & J. Poelt, eds):133–143. München: Universitätsverlag WagnerGes. M. B. H. Innsbruck.
Schroeter, B., Green, T. G. A., Kappen, L. & Seppelt,R. D. (1994) Carbon dioxide exchange rate at sub-zero temperatures. Field measurements on Umbili-caria aprina in Antarctica. Cryptogamic Botany 4:233–241.
Sharma, L. R. (1995). Enumeration of Lichens ofNepal. Euroconsult, Biodiversity Profiles Project,Publication No. 3. Nepal Government, 1–109.
Stainton, J. D. A. (1972) Forests of Nepal. London: JohnMurray Publisher, Ltd.
Stainton, A. (2001) Flowers of the Himalaya: a Supple-ment. Oxford: Oxford University Press.
Upreti, B. N. (1999) An overview of the stratigraphy andtectonics of the Nepal Himalaya. Journal of AsianEarth Sciences 17: 577–606.
Vetaas, O. R. & Grytnes, J. A. (2002) Distribution ofvascular plant species richness and endemic rich-ness along the Himalayan elevation gradient inNepal. Global Ecology and Biogeography 11:291–301.
Vezda, A. & Poelt, J. (1975) Die Gattungen Dimerellaund Pachyphiale. In Khumbu Himal 6 (W. Hellmich,& J. Poelt, eds): 127–132. München: Universitäts-verlag Wagner Ges. M. B. H. Innsbruck.
90 THE LICHENOLOGIST Vol. 42

Vitikainen, O. (1986) Peltigera dolichospora, a newHimalayan-western Chinese lichen. Lichenologist18: 387–390.
Whittaker, R. H. (1972) Evolution and measurement ofspecies diversity. Taxon 21: 213–251.
Wolf, J. H. D. (1993) Diversity patterns and biomass ofepiphytic bryophytes and lichens along an altitudi-nal gradient in the Northern Andes. Annals of theMissouri Botanical Garden 80: 928–960.
Wolf, J. H. D. & Alejandro, F.-S. (2003) Patterns inspecies richness and distribution of vascular epi-phytes in Chiapas, Mexico. Journal of Biogeography30: 1689–1707.
Wolseley, P. A. & Aguirre-Hudson, B. (1997) The ecol-ogy and distribution of lichens in tropical deciduousand evergreen forests of northern Thailand. Journalof Biogeography 24: 327–343.
Yoda, K. (1967) A preliminary survey of the forestvegetation of eastern Nepal II. General description,structure and floristic composition of sample plotschosen from different vegetation zones. Journal ofthe College of Arts and Sciences, Chiba UniversityNational Science Series 5: 99–140.
Accepted for publication 28 March 2009
Appendix 1. Elevation ranges of lichenspecies of Nepal.
Name of lichen species Altitudinalrange (m)
Alectoria ochroleuca 4600–5100Allocetraria flavonigrescens* 4600–4800A. globulans 3700–4700A. oakesiana 3900–4200A. sinensis 4830A. stracheyi 3000–5000Anthracothecium himalayense 1650A. leucostomum 2400–2600Arctocetraria nigricascens 3800Arctoparmelia subcentrifuga 4600Aspicilia cinerea 4300Awasthia melanotricha* 4350–4510Bacidia millegrana 1800–3200B. nigrofusca 3100B. personata 300–1800B. rubella 300B. spadicea 2600Baeomyces pachypus 2000–4200B. roseus 2000–2500Bryonora castanea var. castanea 3750B. castanea var. euryspora 4420B. curvescens 3500–5000B. pulvinar var. microspora* 4720–5070B. pulvinar var. pulvinar* 4830–5100B. pulvinar* 4720–5390B. reducta 3200B. rhypariza var. cyanotropha 3750–3900B. rhypariza var. lamaina 5100–5200B. rhypariza var. rhypariza 4000–4100B. selenospora* 3800–4400B. stipitata* 3000–5080B. yeti 4190–5200Bryoria bicolor 3000–5050B. confusa 3450
Appendix 1. Continued
Name of lichen species Altitudinalrange (m)
B. furcellata 3000B. himalayana 3900–4000B. implexa 3700–4000B. lactinea 3300–3450B. lanestris 3800–3900B. levis 3450B. nadvornikiana 3800–3900B. nepalensis* 3900–4000B. nitidula 3900B. perspinosa 3800–4500B. poeltii 4400B. smithii 3000–4511B. variabilis 3800–4000Buellia geophila 4200B. granularis 2100B. inornata 700–900B. papillata 4650Bulbothrix isidiza 900–2180B. meizospora 2100–2300B. setschwanensis 1260–2300Calicium abietinum 3000–3300C. lenticulare 2250C. viride 3000–3300Caloplaca arnoldii 3500–4000C. aureosora 2500–3000C. borealis var. borealis 3650C. borealis var. oligosperma 4000C. castellana 4000–5540C. cerina var. cerina 3200C. cerina var. chloroleuca 4950–5050C. cerinopsis* 1800–3200C. cirrochroa 3900–4500C. cirrochroopsis* 1480C. citrina 3750C. cupreobrunnea* 2700
2010 Elevation gradients of lichens in Nepal—Baniya et al. 91

Appendix 1. Continued
Name of lichen species Altitudinalrange (m)
C. cupulata* 3200–3800C. epiphyta 4400–4500C. epithallina 4350–4450C. exsecuta var. aphanes* 2060C. farinosa* 3000–3500C. grimmiae 3500C. holocarpa 3700–5000C. holochracea 1700–2300C. insularis 4600–5540C. isabellina* 4100–5200C. leptocheila 5000C. lithophila 3200–4850C. lobulascens* 1500–1800C. lypera* 2000–3000C. maura* 4950–5000C. obliterans 3200–3700C. ochroplaca 1800–3000C. phoenicopta* 3200C. praeruptorum* 3200–3650C. procerispora* 4100–5000C. rinodinopsis 2000–3000C. sancta* 4000–4340C. saxicola var. chamaeleon 3200–4000C. saxicola var. saxicola 2900–5000C. saxifragarum 4950–5000C. tetraspora 4400–5000C. ulcerata* 3500–3750C. variabilis 2000–3500Calvitimela aglaea 5000C. armeniaca 5000Candelaria crawfordii 1300–3000C. sphaerobola 3000–3900Candelariella aurella 1600–3500C. coralliza 3800–5000C. grimmiae 4250–5000C. himalayana 3700–5100C. nepalensis 5000–5400C. sorediosa 3700–5200C. vitellina var. glacialis 4900–5400C. vitellina var. vitellina 1600–5540Canomaculina subsumpta 1200–1500C. subtinctoria 1500Canoparmelia aptata 1500–2000C. ecaperata 1300–2350C. eruptens 5000Carbonea vorticosa 5000–7400Catapyrenium cinereum 4300–5080C. daedaleum 3900–5080Catillaria leptocheiloides 2100Catolechia wahlenbergii 4500Cetraria ambigua 3400–5390C. nepalensis* 4500Cetrelia braunsiana 3150
Appendix 1. Continued
Name of lichen species Altitudinalrange (m)
C. cetrarioides 2000–3000C. olivetorum 2850Cetreliopsis rhytidocarpa ssp. langtangi 2880Chaenotheca brunneola 3000–3556†C. chrysocephala 2700–3879†C. furfuracea 3000C. hispidula †2947–3250C. phaeocephala 3000–3869†C. stemonea 3000Chrysothrix chlorina 3100–3200Cladia aggregata 3900–4000Cladonia amaurocraea 4530–5230C. awasthiana 950–5100C. calyciformis 1500–2700C. cariosa 3100C. carneola 3900–4100C. cartilaginea 1100–2000C. ceratophyllina 3000–4600C. chlorophaea 2000–5350C. ciliata 4480C. coccifera 2000–4000C. corniculata 1650–2300C. corymbescens 1500–5100C. delavayi 2200–5230C. fenestralis 3900–5100C. fimbriata 3900–4250C. fruticulosa 1500–5100C. furcata 1700–3900C. humilis 1200–4250C. laii 2700–4600C. luteoalba 3200–3660C. macilenta 1500–3800C. macroptera 2700–3900C. mongolica 1600–4500C. nitida 3900C. ochrochlora 1500–3500C. pocillum 2000–3600C. pyxidata 4000–4100C. ramulosa 2000–3366C. rangiferina 3400–4500C. scabriuscula 2845–3000C. singhii 750–2700C. squamosa 1800–4000C. stellaris 4400–4500C. stricta 3300–4000C. subconistea 2500–3500C. subulata 2200–2700C. yunnana 3300–4000Coccocarpia erythroxyli 3150C. palmicola 1800Collema nepalense* 3900–4000C. poeltii* 3900–4000C. pulcellum 1350–1500
92 THE LICHENOLOGIST Vol. 42

Appendix 1. Continued
Name of lichen species Altitudinalrange (m)
C. rugosum 1800C. subconveniens 1260C. substipitatum 3000–4000Dermatocarpon miniatum var. miniatum 3200D. vellereum 2000–3600Dibaeis sorediata 2500Dimelaena oreina 3200–4884Dimerella lutea 3000–4000Diploicia canescens 4650Diploschistes muscorum 900–2160D. muscorum subsp. bartlettii 900–2100D. muscorum subsp. muscorum 1500–2160D. nepalensis 900D. scruposus 2000–3000Dirinaria aegialita 1800D. applanata 600D. consimilis 600Erioderma meiocarpum 3000–3050Eumitria pectinata 400–2000Evernia mesomorpha 3962–4572Everniastrum cirrhatum 1900–3300E. nepalense 1410–3600E. rhizodendroideum 2490–3800Flavocetraria cucullata 3600–4500F. nivalis 3800–5300Flavocetrariella leucostigma 3500–4500F. melaloma 3900–4200Flavoparmelia caperata 2250–2743Flavopunctelia flaventior 2250–2800Fuscopannaria poeltii* 3500F. praetermissa †3805–4000Glyphis cicatricosa 1600Graphis scripta 1600–3366G. subglauconigra 2000–3200Haematomma puniceum 1800Heterodermia angustiloba 1800–2300H. awasthii 1900H. boryi 1500–3000H. comosa 1500–2500H. dactyliza 2100H. dactyliza f. serpens 3800–5150H. dendritica 1800H. diademata 410–3807†H. dissecta var. dissecta 1500–3000H. firmula 1200–2200H. flabellata 3100H. himalayensis 1500–2000H. incana 1800–2100H. isidiophora 2100–2250H. japonica 3000–4000H. obscurata 1400–3000H. pellucida 2500–3992H. propagulifera 1500–3000
Appendix 1. Continued
Name of lichen species Altitudinalrange (m)
H. pseudospeciosa 150–2100H. punctifera 2250H. rubescens 1800–2800H. speciosa 1650–2100H. togashii 3000–4000H. tremulans 1500Hyperphyscia granulata 1200H. minor 1200–1400Hypogymnia delavayi 1800–4080H. hypotrypa 3600–4050H. vittata 2800–4200Hypotrachyna adducta 2100H. crenata 2500–2800H. exsecta 1800–2300H. flexilis 2160H. imbricatula 1400–2600H. infirma 1000–2100H. koyaensis 1500–2100H. majoris 1650H. neodissecta 2250–2550H. osseoalba 1100–2200H. revoluta 2000–2250H. rhabdiformis 2500–2800H. scytophylla 2250–3250H. sinuosa 3500H. sublaevigata 2250Immersaria athroocarpa 3900–4340Ingvariella bisporus 3750Ioplaca pindarensis 2900–5200Lasallia freyana 3800–3900L. pertusa 3800–5050Lecanora adolfii 5540L. amorpha* 5000L. chlarotera 200L. chondroderma 3800–5400L. demissa 2700–4000L. emodi* 5540L. formosa 4850–5860L. garovaglii 3000–5050L. hellmichiana* 1800–3500L. himalayae* 3900–4850L. kirra* 4400–4500L. lesleyana* 5029–5334L. meridionalis 1600–3366L. muralis var. dubyi 4930L. muralis var. muralis 3500–4340L. phaeodrophthalma 4100–4500L. rubina var. australis 3300–4250L. rugosella 2732–3200L. sherparum* 3300–4600L. somervellii 3700–5500L. sulphurea 4300–5639L. terestiuscula 4500–5200
2010 Elevation gradients of lichens in Nepal—Baniya et al. 93

Appendix 1. Continued
Name of lichen species Altitudinalrange (m)
L. tschomolongmae* 3000–5000Lecidea advena 5000–5540L. auriculata 5000–5200L. bella* 4950–5000L. brachyspora 4300–5900L. bucculenta 5000–5540L. diducens 4720L. epiiodiza 4850L. fuscoatra var. indecora 4340L. haerjedalica var. gyrodisca 5150–5200L. himalaica 5150–5200L. khumbuensis 5540L. lactea 5460L. leptoboloides 4340–5200L. molybdochroa 3900–4000L. poeltii 4500–5000L. secernens 5000L. silacea 5000L. steineri 5150–5200L. tessellata 5080–5639Lecidella carpathica 3900–4000L. dimelaenophila 3800–3900L. stigmatea 4520–5200Leprocaulon arbuscula 1900–3100Leproplaca chrysodeta 3300–4850Leptogium asiaticum 3900–4100L. askotense 1500–1800L. azureum 1410L. brebissonii 1800–3200L. burnetiae var. burnetiae 1500–2250L. cochleatum 1800L. delavayi 3000–4100L. delavayi f. fuliginosulum 3900–4000L. isidiosellum 1500L. javanicum 1410–1800L. pedicellatum 1500–3000L. phyllocarpum 2000–3100L. resupinas 1600L. saturninum 1500–2100L. trichophorum 1450–1500Lethariella cladonioides 4724Letrouitia domingensis 240Lobaria discolor 3150L. isidiosa 2800–3962L. kurokawae 1800–3400L. pindarensis 2700–4000L. pseudopulmonaria 2550–4050L. retigera 1600–3650L. subretigera 2800Lobothallia alphoplaca 4400L. praeradiosa 4000–4500Melanelia tominii 3200–3600Melanohalea poeltii* 4500–4600
Appendix 1. Continued
Name of lichen species Altitudinalrange (m)
Menegazzia terebrata 1300–4000Mycobilimbia hunana 1650Myelochroa aurulenta 1410–2250M. entotheiochroa 2000–2200M. subaurulenta 1950Nephroma helveticum var. helveticum 2160–2800N. nakaoi 2800Nephromopsis ahtii 4090–4530N. isidioidea 2900–3100N. nephromoides 2400–3600N. pallescens 2400–3048N. stracheyi 2700–3300Ochrolechia bryophaga 3900–4000O. glacialis 5000–5200O. margarita 5000O. rosella f. sorediascens 3500O. subviridis 2900O. trochophora 3000–4000Pachyphiale himalayensis 3900–4000Parmelaria subthomsonii 1800–3200P. thomsonii 1800–3366Parmelia adaugescens 3300–3600P. erumpens 1900–2200P. latissima var. marmariza 2900P. masonii* 3000–6100P. meiophora 1800–3350P. omphalodes 3500–4500P. ricasolioides 2700P. squarrosa 3000–3800P. submutata 2800P. sulcata 3000–3366Parmelina tiliacea 2100Parmelinella simplicior 2000–2900P. wallichiana 1800–2400Parmelinopsis expallida 1800P. minarum 1500Parmotrema austrosinense 750–3000P. cooperi 1500–2100P. hababianum 1500–2000P. maclayanum 1500P. melanothrix 2400–3200P. mellissii 1500P. nilgherrensis 1080–2800P. praesorediosum 1410–1800P. pseudonilgherrense 1800–3530P. pseudotinctorum 2900–3366P. rampoddense 900–2800P. ravum 1500–2500P. reticulatum 1500–2500P. sancti–angelii 1410–2100P. stuppeum 1800P. tinctorum 240–2250P. ultralucens 1500
94 THE LICHENOLOGIST Vol. 42

Appendix 1. Continued
Name of lichen species Altitudinalrange (m)
P. yodae* 2300Peltigera canina 3150–3300P. dolichorrhiza f. dolichorrhiza 1800–2250P. dolichospora* 3000–4100P. elisabethae 1350P. malacea 2700–3300P. membranacea 1900–2400P. polydactylon 1950–2920P. pruinosa 1800P. rufescens 2100P. scabrosa 3800Pertusaria hemisphaerica 2400–3366Phaeographina pyrrhochroa 2100Phaeophyscia endococcina 3800–3900P. endococcina var. khumbuensis 3900P. endococcinoides var. megalospora 3900–4000P. hispidula var. exornatula 365P. hispidula var. hispidula 3800–3900P. lygaea* 4500–4600P. primaria* 3800–3900P. pyrrhophora* 1420P. sciastra 3900–5200Phlyctella indica 1800Physcia aipolia 1800–2200P. caesia 3900–5000P. clementei 3500–4000P. dilatata 1600–5000P. dubia 5000P. phaea 4340–5000P. stellaris ssp. intestiniformis* 4250–4340P. tribacia 3800–4600P. tribacioides 1400Physciella nepalensis* 1400Physconia distorta 3000–4000P. enteroxantha 2740P. grisea 1400–4200P. muscigena 4250–5540Physma byrsaenum 1350Placidium squamulosum 4400–4500Pleopsidium chlorophanum* 3500–4000Porpidia aerolotera 5150–5200P. crustulata 2800–5000P. elegantior 4700P. hydrophila 3900–4000P. macrocarpa 2950–4000Protoparmelia badia var. badia 3750–4000P. effigurans* 3750Punctelia borreri 1600–3200P. rudecta 1500–3200P. subrudecta 1500–2500Pyrenula cayennensis 3000–3200P. immersa 2000Pyxine coccifera 750
Appendix 1. Continued
Name of lichen species Altitudinalrange (m)
P. meissnerina 300P. philippina 900–1800P. sorediata 1410–1800Ramalina conduplicans 2133R. farinacea 2700R. flabelliformis* 3200R. hossei var. hossei 1200–2500R. sinensis 2700–4267R. subfarinacea 1610Rhizocarpon geographicum 4500Rhizoplaca chrysoleuca var. chrysoleuca 2100–5150R. melanophthalma var. obscura 4270R. peltata 3660Sagema potentillae 4000Sclerophora coniophaea 3000Solorina bispora 3600–4200Stereocaulon claviceps 4000S. foliolosum var. foliolosum 3300–4000S. foliolosum var. botryophorum 3600–4600S. foliolosum var. strictum 2400–3600S. glareosum 4200–4400S. himalayense 2500–5400S. myriocarpum 3900–5303S. paradoxum 2000–3366S. piluliferum 2160–5150S. pomiferum 2500–4700S. sasaki var. sasaki 3600–4200Sticta henryana 3657–3962S. nylanderiana 1800–3600S. platyphylloides 1900–3150S. praetextata 2100–3500S. weigelii var. weigelii 800–2250Sulcaria sulcata 3150–3300S. virens 3000–3600Tephromela aglaea 5000–5200T. armeniaca 5000–5200T. glacialis* 4500T. siphulodes var. siphulodes* 4830Thamnolia vermicularis 3869–5455Thelenella luridella 1410Tremolecia atrata 5000–5860Tuckneraria laureri 3200–4900Tylophoron moderatum 1800Umbilicaria badia 2550U. cinereorufescens 4250–5200U. decussata var. decussata 5100–5500U. decussata var. rhizinata* 4950–5000U. indica var. indica 1800–3150U. indica var. nana* 4200–4300U. krascheninnikovii 3700–5100U. leiocarpa 5150U. nenella 5100U. nepalensis 3600–5100
2010 Elevation gradients of lichens in Nepal—Baniya et al. 95

Appendix 1. Continued
Name of lichen species Altitudinalrange (m)
U. thamnodes 3600–5050U. trabeculata 5000–5100U. vellea 3600–5050U. yunnana 2550Usnea aciculifera 1200–2250U. baileyi 1500–2400U. compressa 2400–2700U. dendritica 1200–3000U. galbinifera var. subfibrillosa* 2200U. himalayana 1800–3300U. longissima 2700–3750U. montisfuji 2200–3900U. nepalensis 3800–4000U. norkettii* 3000–3500U. pectinata 3962U. pseudomontisfuji 3200U. robusta 2500–3000
Appendix 1. Continued
Name of lichen species Altitudinalrange (m)
U. rubicunda 2700U. splendeus 1800–3300U. thomsonii 1800–3150Xanthoparmelia coreana 3400–3850X. dentata 4480–4572Xanthoparmelia isidiosa 2800–3200X. mexicana 3070–3700X. nepalensis* 3900Xanthoria borealis 3900–4000X. elegans 3750–6000X. fallax 3200–3400X. fulva 2700–3200X. sorediata 3200–5000X. ulophyllodes var. ulophyllodes 3200–4850†
* species endemic to Nepal†Louise Olley’s unpublished personal observation
Appendix 2. The elevational gradient of lichen species richness regression analysisresults modelled after different species richness as response variables and theirelevation as predictor variable. The Quasi-poisson family of error fitted in theGAM model after the cubic smooth spline (s) with approximately 4 degrees of
freedom. (P%0·05)
Response variables Null df Res. df D2 Deviance F Pr (>F)
Total species 72 68 0·945 3598·1 304·5 <0·001Endemic species 72 68 0·915 362·3 205 <0·001Crustose species 72 68 0·914 1008 181·4 <0·001Foliose species 72 68 0·941 1764·9 288·4 <0·001Fruticose species 72 68 0·961 1252·2 560·9 <0·001Cyanolichens 72 68 0·944 632·2 338·4 <0·001Green algal lichens 72 68 0·941 3047·7 280·9 <0·001Corticolous 72 68 0·970 2374·3 588 <0·001Saxicolous 72 68 0·908 1075·7 190·2 <0·001Terricolous 72 68 0·895 422·3 192·8 <0·001
Df = degree of freedom, Res. = residual, D2 = regression coefficient of determination
96 THE LICHENOLOGIST Vol. 42