Embed Size (px)
Citation preview

H E M A T O P A T H O L O G Y
Original Article
p 5 3 E x p r e s s i o n i n R e e d - S t e r n b e r g C e l l s D o e s
N o t C o r r e l a t e W i t h G e n e M u t a t i o n s i n
H o d g k i n ' s D i s e a s e
KOJO S.J. ELENITOBA-JOHNSON, MD, L. JEFFREY MEDEIROS, MD, JILA KHORSAND, MD, AND THOMAS C. KING, MD, PHD
Immunohistochemically detectable p53 protein expression is common in the Reed-Sternberg and Hodgkin's (RS-H) cells of Hodgkin's disease, but p53 gene mutations have only rarely been identified. The authors found p53 expression in RS-H cells in 16 of 30 cases of Hodgkin's disease (53%), with the percentage of RS-H positive cells ranging from 4% to 85%. In 12 of 30 cases (40%), at least 10% of the RS-H cells were positive for p53. p53 gene mutations were detected in only two cases (7%) using a single-stranded conformational polymorphism assay with a detection sensitivity of between 1% and 5%. The cellular protein, mdm-2, which can stabilize and functionally inactivate wild-type p53
Alterations of the p53 tumor suppressor gene constitute one of the most frequently detected genetic abnormalities in human neoplasms.1'2
The p53 gene is located on chromosome 17pl3 and encodes a 53-kD nuclear phosphoprotein, which is involved in the regulation of transcription, the induction of programmed cell death (apoptosis), and the regulation of cell cycle progression at the GiS checkpoint.3
More than 90% of point mutations in the p53 gene are found in exons 5-9 and are clustered in four mutational "hotspots".4'5
Wild-type p53 protein is rapidly turned over in normal cells and does not accumulate in sufficient amounts to be detected by standard immunohistochemical techniques.
From the Departments of Pathology, Roger Williams Medical Center, Rhode Island Hospital, The Miriam Hospital, and Brown University School of Medicine, Providence, Rhode Island.
Dr. Elenitoba-Johnson is currently at the National Cancer Institute, National Institute of Health, Bethesda, Maryland.
Dr. Medeiros is currently at the City of Hope National Medical Center, Duarte, California.
Manuscript received March 18, 1996; revision accepted July 11, 1996.
Supported by grant #NCI 5P30CA13943-20S1 from Roger Williams Cancer Center.
Address reprint requests to Dr. King: Department of Pathology, The Miriam Hospital, 164 Summit Avenue, Providence, RI 02906.
protein, was expressed in RS-H cells in most of these cases (86%). However, neither case with a p53 gene mutation expressed mdm-2 (P -c.005). The two cases with p53 gene mutations had a higher mean proliferative index than cases without detectable mutations (90% versus 72%; P <.02). p53 expression in RS-H cells may be related to concurrent mdm-2 protein expression and a p53-positive, mdm-2-negative im-munophenotype may be predictive of gene mutations in RS-H cells. (Key words: p53; Hodgkin's disease; mdm-2; mutation; SSCP) Am J Clin Pathol 1996;106:728-738.
Most p53 gene alterations are missense mutations that result in an abnormal p53 protein with a prolonged half-life, which renders it immunohistochemically detectable.6
Because p53 normally functions as homodimer, mutant p53 protein can bind to wild-type p53 protein and sequester it in nonfunctional heterodimers. In this manner, a single mutated allele can result in a dominant negative phenotype.7 p53 gene point mutations can also result in proteins with abnormal regulatory properties and/ or loss of tumor suppressor function. Different point mutations can result in different phenotypes depending on the nature and degree to which protein structure is altered.
Immunohistochemical detection of p53 protein in tumor cells was initially thought to imply the presence of p53 gene mutations.8
However, it is now clear that p53 expression and mutation are not always correlated.9 Nonsense and frame shift mutations tend to produce unstable proteins that are rapidly degraded and can not be detected immunohistochemically. Conversely, wild-type p53 protein can form complexes with a variety of viral and/or cellular proteins that may prolong its half-life and result in its immunohistochemical detection.
The p53 protein is frequently expressed in the Reed-Sternberg and Hodgkin (RS-H) cells of Hodgkin's dis-
728
Dow
nloaded from https://academ
ic.oup.com/ajcp/article/106/6/728/1756733 by guest on 30 Septem
ber 2022

ELENITOBA-JOHNSON ET AL. 729
p53 Mutations in Reed-Sternberg Cells
ease, but p53 gene mutations have only been identified in a few cases.10" Although the failure to detect mutations could result from the low percentage of RS-H cells in many cases of Hodgkin's disease, other mechanisms appear to be responsible for p53 expression in other types of tumors. Viral proteins that bind to p53 can either lead to its rapid degradation or modulate its function and prolong its half-life.12 This latter mechanism could potentially be operative in Hodgkin's disease cases that express p53 protein. The Epstein-Barr virus (EBV) nuclear antigen (EBNA-1) protein has been shown to bind to a 53 kD cellular protein, raising the possibility that p53 protein expression in Hodgkin's disease results from this interac
tion 13 EBV DNA and gene products have been detected in
more than 50% of cases of Hodgkin's disease in some studies,14"16 and the expression of EBNA-1 as well as latent membrane proteins (LMP-1, LMP-2a, and LMP-2b) has been documented.17 LMP-1, a transmembrane phosphoprotein with transforming potential, is expressed in virtually all RS-H cells in EBV-positive cases of Hodgkin's disease.1618
Nonviral cellular proteins can also bind to p53 and regulate its function and modulate its half-life. The human homologue of the murine double minute 2 gene, mdm-2, encodes a zinc finger protein that binds to the activation domain of the p53 protein and regulates its function.61920
The mdm-2 protein has a short half-life and is usually not detectable by conventional immunohistochemis-try." Amplification of the mdm-2 gene occurs frequently in sarcomas that express the mdm-2 protein and permit its immunohistochemical detection. Expression of mdm-2 protein may also result from its stabilization by binding to p53 protein.
In an attempt to better understand the role of p53 in Hodgkin's disease, we have examined 30 cases of Hodgkin's disease for p53 expression and for mutations in ex-ons 5-8 of the p53 gene using a sensitive PCR-SSCP technique.21 We also evaluated RS-H cells for EBV-LMP-1 and mdm-2 expression in order to define their roles in p53 expression. We also evaluated the relationship of the expression of the proliferation marker Ki-67 and clonal immunoglobulin and T cell-antigen receptor y chain gene rearrangement in RS-H cells with p53 expression and gene mutations.
MATERIALS AND METHODS
Case Selection and Pathologic Analysis
Thirty cases of Hodgkin's disease from the Departments of Pathology at Rhode Island Hospital and Roger
Williams Medical Center were selected for study based on the presence of numerous RS-H cells assessed by histologic review of tissue sections and on the availability of formalin-fixed, paraffin tissue blocks, which were adequate for immunohistochemical and DNA analysis. For each case, tissue blocks were selected that had the highest percentage of involvement by Hodgkin's disease and/or the greatest number of RS-H cells. The percentage of RS-H cells was assessed by performing a 250-cell differential count on representative fields selected by scanning at low power.
Immunohistochemical Studies
Immunohistochemical studies were performed using formalin-fixed, paraffin-embedded tissue sections using an avidin-biotin peroxidase complex technique (Vector, Burlingame, CA) and the following of antibodies: LCA (CD45 RB), L26 (CD20), UCHL-1 (CD45 RO), anti-CD3, Ber H2 (CD30), and CS1-4 (Epstein-Barr virus latent membrane protein-1) (Dako, Carpinteria, CA); Leu 22 (CD43) and Leu M1 (CD 15) (Becton-Dickinson, San Jose, CA); MIB-1 (Ki-67) (AMAC, Westbrook, ME); PAb 1801 (anti-p53) and IF-2 (anti-mdm-2) (Oncogene Science, Uniondale, NY). An antigen retrieval method using citrate buffer and microwave heating was used for the MIB-1 and PAb 1801 antibodies as described.22 Positive controls included a normal lymph node for CD antibodies and Ki-67, a p53 positive colon cancer for p53, and a mdm-2 positive leiomyosarcoma for mdm-2. Antigen expression in RS-H cells was judged by comparison with positive controls and appropriate positively and negatively staining background cells in the same tissue sections (eg, T cells, B cells, and endothelial cells).
PCR-SSCP Analysis
DNA was extracted from formalin-fixed, paraffin-embedded tissue blocks as described.23
The oligonucleotide primers used for amplification of p53 exons 5-8 were derived from published sequences and are as follows: p53 exon 5 sense primer-5' -TTC CTC TTC CTG CAG TAC TC-3', exon 5 anti-sense primer 5' -ACC CTG GGC A AC CAG CCC TGT-3'; p53 exon 6 sense primer 5'-ACA GGG CTG GTT GCC CAG GGT-3', exon 6 anti-sense primer 5'- AGT TGC AAA CCA GAA CCT CAG -3'; p53 exon 7 sense primer 5' -GTG TTG TCT CCT AGG TTG GC-3', exon 7 anti-sense primer 5'-GTC AGA GGC AAG CAG AGG CT-3'; p53 exon 8 sense primer 5' -TAT CCT GAG TAG TGG TAA TC -3', exon 8 antisense primer 5' -AAG TGA ATC TGA GGC ATA AC -3'.24
Vol. 106-No. 6
Dow
nloaded from https://academ
ic.oup.com/ajcp/article/106/6/728/1756733 by guest on 30 Septem
ber 2022

730 HEMATOPATHOLOGY
SSCP analysis was performed using a modification of the method reported by Orita and colleagues.25 1 /*g of purified DNA was amplified in a 50 fiL reaction each containing IX GenAmp PCR buffer, 1.5 mM MgCl2, paired sense and anti-sense primers at 1 nM, and 1.25 units of AmpliTaq DNA Polymerase (Perkin Elmer, Norwalk, CT). Reactions included 1-5 fid a32P dCTP with 50 nM dATP, dGTP, and dTTP; 10 fiM dCTP. A "hot start" technique was employed in all assays. Thirty-five cycles of denaturation (96 °C, 1 minute), annealing (59 °C, 1 minute), and extension (72 °C, 1 minute) were performed using a Thermal-Cycler 480 (Perkin-Elmer Cetus). 10 /iL of the PCR products were mixed with 45 uL of 95% formamide, 20 mM EDTA, 0.05% bromophe-nol blue, and 0.05% xylene cyanol. The resulting mixture was then heated at 94 °C for 3 minutes, rapidly chilled in an ice-water bath, and 5-10 /^L was immediately loaded onto a SSCP gel [0.25X MDE (AT Biochem, Malvern, PA) gel cast in a BioRad (Richmond, CA) sequencing gel apparatus]. Electrophoresis was performed in 0.7X TBE, at 8-10 watts constant power, for 5 to 16 hours at room temperature (exons 5 and 7, 75 Watt-hours; exon 6, 65 Watt-hours; and exon 8, 90 Watt-hours). Gels were transferred onto filter paper (3 MM Whatmann, Maidstone Eng., UK), dried, and autoradiographed at - 7 0 °C for 12 to 24 hours with Kodak (Rochester, NY) XAR or XP X-ray film. Putative mutations detected by SSCP were confirmed by DNA sequencing.
To asses the sensitivity of the PCR-SSCP assay employed, DNA from human placenta and two ovarian carcinoma samples with different p53 gene mutations in exon 5 were mixed to achieve different percentages of mutant DNA. One tig of each DNA mixture was then subjected to PCR-SSCP analysis to determine the minimum percentage of mutant DNA detectable with this assay.
DNA Sequencing Analysis
PCR products for the appropriate p53 exon were prepared from tumor DNA as described above except that a32P dCTP was omitted and 200 ^M final concentration of each dNTP was used. PCR products were electropho-resed in an agarose gel and the gel band was excised and melted in water. This material was then used as a template in a second PCR employing a single p53 specific primer (sense or antisense) to generate single-stranded DNA. The resulting single-stranded DNA products were purified using the Qiagen system (Qiagen, Studio City CA) and subjected to DNA sequencing. Both strands of the appropriate exon were sequenced using Sequenase version 2.0 (United States Biochemical, Cleveland, OH)
A.J.C.P..
Article
and the opposite p53 primer pre-labeled with 732P ATP according to the manufacturer's instructions. Samples were denatured and loaded onto standard 6-8% poly-acrylamide/urea sequencing gels.
Alternatively, for case 18, PCR products were prepared from tumor DNA as for standard SSCP analysis except that higher concentrations of dNTPs (200 fiM each) were employed in the reactions. Following SSCP analysis, the altered mobility band was excised using the autoradiograph as a template. DNA was eluted from the gel and re-amplified as described above. For case 26, PCR products were cloned using the TA cloning system (InVitrogen., San Diego, CA), following the manufacturer's instructions. Plasmid clones were screened by SSCP and several that matched the altered mobility pattern of the original tumor sample were further analyzed. Plasmid DNA was directly sequenced as describe above or using an M13 universal primer present in the plasmid vector.
Immunoglobulin Heavy Chain (IgH) and T-Cell Receptor (TCR) y Gene Rearrangement Analysis
IgH gene rearrangement analysis was performed using PCR as described.26 32P dCTP was included in most assays and dried gels were autoradiographed to enhance assay sensitivity. A similar technique was employed for TCR 7 analysis using two joining and two variable region consensus primers.27 In both cases, PCR products were analyzed by electrophoresis in 10% polyacrylamide gels. All assays included appropriate positive (diffuse large B cell lymphoma and T-cell lymphoblastic lymphoma DNA, respectively), negative (normal tonsil and thymus DNA, respectively), and contamination (no added DNA) controls. The assay for IgH gene rearrangement detects 80% of clonally rearranged IgH genes in our laboratory. The assay for TCR y is less sensitive in our hands and is positive in approximately 50% of T-cell lymphomas.
RESULTS
Clinical Data
The patients consisted of 14 males and 16 females ranging in age from 11 to 82 years (median 36 years). Lymph node biopsy sites included submandibular, cervical, supraclavicular, axillary, mediastinal, retroperitoneal, and splenic hilar regions. Using standard histologic criteria, cases were classified as nodular sclerosis (18 cases), mixed cellularity (9 cases), nodular lymphocyte predominance (2 cases), and lymphocyte depletion (1
:r 1996
Dow
nloaded from https://academ
ic.oup.com/ajcp/article/106/6/728/1756733 by guest on 30 Septem
ber 2022

ELENITOBA-JOHNSON ET AL. 731
p53 Mutations in Reed-Sternberg Cells
TABLE 1. HISTOLOGIC, PHENOTYPIC, AND GENOTYPIC PARAMETERS IN HODGKIN'S DISEASE*
atient No.
1 2 3 4 5 6 7 8 9
10 II 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
Subtype
NS NS NS NS NS NS NS NS NS NS NS NS NS NS NS NS NS NS MC MC MC MC MC MC MC MC MC LP LP LD
p53 SSCP+ (exon)
_ ----------------5 -------6 ----
p53 IHC+ RS-H
Cells (%)
0 0 0 0 0 0 0 4 6 7
13 17 30 47 60 71 84 85 0 0 0 0 0
11 31 50 54 0 6 0
p53 IHC+ Total
Cells (%)
0 0 0 0 0 0 0 1 1 5 1 1 5 3 5
30 8
23 0 0 0 0 0 1 2
20 7 0 0 0
mdm-2 IHC+ RS-H
Cells (%)
0 84 30 14 86 ND
0 15 63 65 57 32 52 54 51 37 74 0
70 47 50 23 51 39 20 0
60 30 35 58
EBV IHC+ RS-H Cells
_ ---+ ---+ + --+ ----+ -+ ---+ + + ----
Clonal IgH
_ -----------\V --+ ----w ---+ -----
Ki-67 IHC+ RS-H Cells
68 73 73 85 63 ND 55 53 54 ND 64 66 57 71 51 56 77 90 71 58 75 64 73 50 56 72 55 71 67 50
RS-H
5 ND 14 5
ND 6
ND 12 14 71 11 3
17 7 8
43 10 27 4 6
12 2
ND 8 6
40 14 4 4
87
Age (years)
20 28 38 42 61 62 73 24 42 76 13 34 19 11 26 16 77 33 17 35 58 72 82 38 69 15 78 15 27 29
Sex
F F F F F M F F F F M F F M M F M M F M M F F M M M F M M M
NS = nodular sclerosis: MC = mixed cellularity: LP = lymphocyte predominance: LD = lymphocyte depletion: ND = not done: W = oligoclonal bands.
• Data for cases with ^ 10% p53 + RS-H cells are shown in boldface type.
case). As expected, females accounted for a higher percentage of cases of nodular sclerosis (67%). The percentage of the RS-H cells varied widely, from < 1 % to 71 % (median 8%).
Immunohistochemical Data
In all cases, the RS-H cells had an immunophenotypic profile typical of Hodgkin's disease.28 In the two cases of nodular lymphocytic predominance, the L&H cells were LCA and L26 positive and negative for Leu M1 and T-cell markers. In cases of nodular sclerosis, mixed cellularity and lymphocyte depletion, the RS-H cells were positive for Leu Ml and/or Ber H2, and negative for LCA and T-cell antigens. One case of nodular sclerosis and two cases of mixed cellularity showed L26 expression in RS-H cells. Immunohistochemical results for p53, EBV-LMP, mdm-2, and Ki-67 are detailed below and are listed in Table 1.
p53 Expression
Benign infiltrating lymphocytes and inflammatory cells were consistently negative for p53 protein. In contrast, RS-H cells commonly showed p53 expression (varying intensity, semi-quantitatively graded from 0 to 3+). A 250-differential cell count was performed to assess the overall percentage of RS-H cells and the percentage of RS-H cells that significantly expressed p53. The percentage of p53 positive RS-H cells varied widely between different cases (0% to 85%). Strong (2-3+) immu-noreactivity was found in greater than 10% of the RS-H cells in 12 of 30 (40%) cases of Hodgkin's disease (Table 1 and Fig. 1) and these were operationally classified as p53 positive. In 8 of these 12 cases, the RS-H cells represented greater than 5% of the total cell population. p53 positive cases were evenly divided between nodular sclerosis and mixed cellularity subtypes (44% each). Neither case of lymphocyte predominance nor the single case of
Vol. 106-No. 6
Dow
nloaded from https://academ
ic.oup.com/ajcp/article/106/6/728/1756733 by guest on 30 Septem
ber 2022
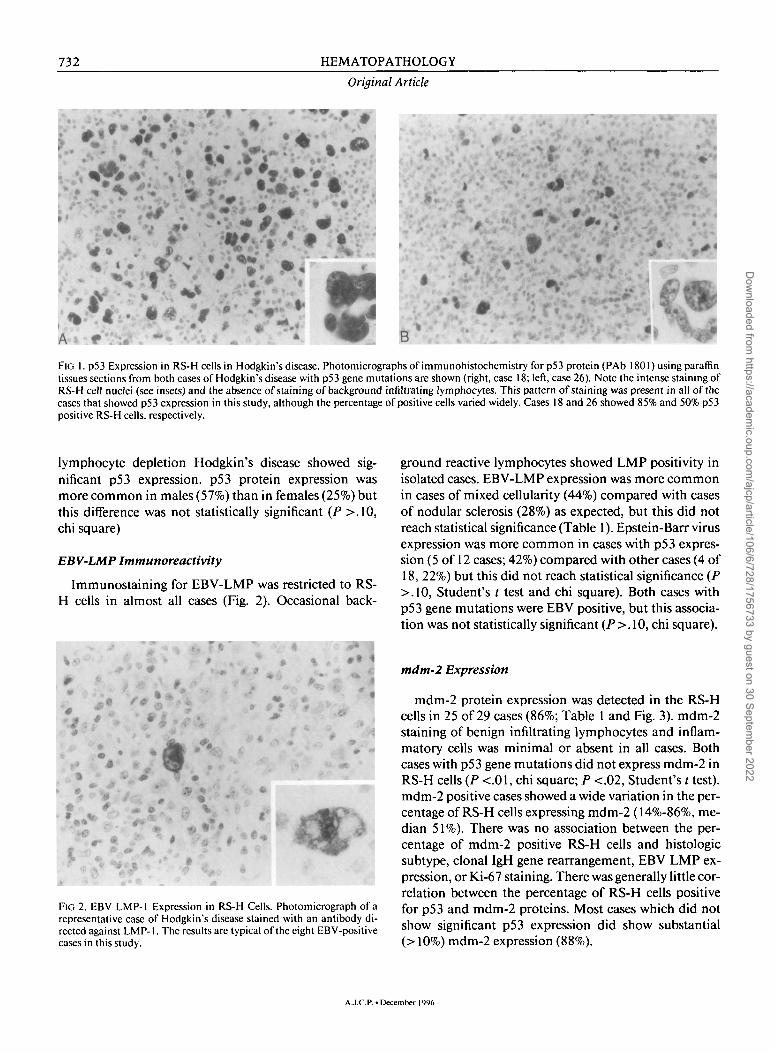
732 HEMATOPATHOLOGY
Original Article
FIG 1. p53 Expression in RS-H cells in Hodgkin's disease. Photomicrographs of immunohistochemistry forp53 protein (PAb 1801) using paraffin tissues sections from both cases of Hodgkin's disease with p53 gene mutations are shown (right, case 18; left, case 26). Note the intense staining of RS-H cell nuclei (see insets) and the absence of staining of background infiltrating lymphocytes. This pattern of staining was present in all of the cases that showed p53 expression in this study, although the percentage of positive cells varied widely. Cases 18 and 26 showed 85% and 50% p53 positive RS-H cells, respectively.
lymphocyte depletion Hodgkin's disease showed significant p53 expression. p53 protein expression was more common in males (57%) than in females (25%) but this difference was not statistically significant (P >.10, chi square)
EBV-LMP Immunoreactivity
Immunostaining for EBV-LMP was restricted to RS-H cells in almost all cases (Fig. 2). Occasional back-
P «
* - % i f F t ' • %
«
/* ~*
«» ,« A.
$ * »* 0 t^iji*- .0
* t'
0 t *
v»
:*» i
"*',»
FIG 2. EBV LMP-1 Expression in RS-H Cells. Photomicrograph of a representative case of Hodgkin's disease stained with an antibody directed against LMP-1. The results are typical of the eight EBV-positive cases in this study.
ground reactive lymphocytes showed LMP positivity in isolated cases. EBV-LMP expression was more common in cases of mixed cellularity (44%) compared with cases of nodular sclerosis (28%) as expected, but this did not reach statistical significance (Table 1). Epstein-Barr virus expression was more common in cases with p53 expression (5 of 12 cases; 42%) compared with other cases (4 of 18, 22%) but this did not reach statistical significance (P >.10, Student's t test and chi square). Both cases with p53 gene mutations were EBV positive, but this association was not statistically significant (P >. 10, chi square).
mdtn-2 Expression
mdm-2 protein expression was detected in the RS-H cells in 25 of 29 cases (86%; Table 1 and Fig. 3). mdm-2 staining of benign infiltrating lymphocytes and inflammatory cells was minimal or absent in all cases. Both cases with p53 gene mutations did not express mdm-2 in RS-H cells (P <.01, chi square; P <.02, Student's t test). mdm-2 positive cases showed a wide variation in the percentage of RS-H cells expressing mdm-2 (14%-86%, median 51%). There was no association between the percentage of mdm-2 positive RS-H cells and histologic subtype, clonal IgH gene rearrangement, EBV LMP expression, or Ki-67 staining. There was generally little correlation between the percentage of RS-H cells positive for p53 and mdm-2 proteins. Most cases which did not show significant p53 expression did show substantial (> 10%) mdm-2 expression (88%).
A.J.C.P.« December 1996
Dow
nloaded from https://academ
ic.oup.com/ajcp/article/106/6/728/1756733 by guest on 30 Septem
ber 2022

ELENITOBA-JOHNSON ET AL. 733
p53 Mutations in Reed-Sternberg Cells
FIG 3. mdm-2 Protein Expression in RS-H cells. Photomicrograph of a representative case of Hodgkin's disease stained with an antibody directed against mdm-2 protein. Note the strong nuclear staining in two RS-H cells similar to the pattern for p53 protein. This case also exhibited p53 protein expression but did not have a detectable p53 gene mutation (data not shown). Background cells showed little or no specific staining.
Ki-67 Expression
Forty-five percent to 90% of the RS-H cells in individual cases exhibited reactivity for the Ki-67 antigen (MIB-1 antibody). There was little correlation between the percentage of p53 positive RS-H cells and the percentage of Ki-67 positive RS-H cells. Both cases with p53 gene mutations showed high level staining for Ki-67. No association between Ki-67 staining and other parameters was identified.
Antigen Receptor Gene Rearrangement
Two cases showed evidence of definite monoclonal IgH gene rearrangement by PCR (Fig. 4 and Table 1). Both of these cases were "classical" Hodgkin's disease (1 NS, 1 MC; Table 1). Each of these cases had a higher percentage cells with p53 protein expression than other cases (16% versus 3%, P <.02, Student's / test). Two additional cases showed an oligoclonal IgH gene rearrangement pattern without a dominant band after gel electrophoresis. None of these four cases had detectable p53 gene mutations. There was no apparent association between monoclonal or oligoclonal IgH gene rearrangement with histologic subtype or with any other parameters evaluated. The frequency of clonal IgH gene rearrangement in this study (7-13%) is less than that reported in two recent series of Hodgkin's disease cases (44% and 50%, respectively) using similar PCR tech-
The frequency of L26 positivity in RS-H cells was also lower in this study (12% of "classical" Hodgkin's disease cases) than in those series and may partially explain the differences observed. Only one of our two cases with clonal IgH gene rearrangement showed L26 expression in RS-H cells. No cases showed evidence of clonal or oligoclonal TCR 7 gene rearrangement (data not shown).
p53 Mutation Analysis
PCR-SSCP analysis detected p53 gene mutations in only two cases (Fig. 5). Case 18 had a transversion (adenine -»• thymidine) at nucleotide 1028 in exon 5 resulting in an amino acid substitution of leucine for methionine at codon 160 (Fig. 6, left panel). Although the functional significance of this particular mutation is unknown, it does not correspond to any previously reported polymorphisms in the p53 coding sequence.31
The presence of this mutation in only a small fraction of tumor DNA eliminates the possibility that it could be a constitutional genetic alteration in the p53 gene. Direct sequencing of PCR products from case 26 showed wild-type sequence in exon 6 up to nucleotide 1151. At this point, the sequencing ladder diverged from the germline
Immunoglobulin Heavy Chain PCR
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 M
— 383
- 1 2 1
niques. 29.30
FlG 4. Immunoglobulin heavy chain gene PCR analysis. DNA isolated from paraffin tissue blocks was amplified using consensus primers for IgH genejoining(J)and variable (V) regions. Positive (lane l)and negative (lanes 2 and 3) control reactions are included. A clonal rearrangement is detected as a discrete band after acrylamide gel electrophoresis (lane 4). Such bands only result if complete VDJ rearrangement has occurred. Other Hodgkin's disease cases in this assay (lanes 5-18) did not show evidence of clonal IgH gene rearrangement. Molecular weight markers are indicated (M) in nucleotides.
Vol. 106. No. 6
Dow
nloaded from https://academ
ic.oup.com/ajcp/article/106/6/728/1756733 by guest on 30 Septem
ber 2022

734 HEMATOPATHOLOGY
Original Article
e x o n 5
l
+
2
wt
3
wt
4
wt
e x o n 6
5
+
6
MM
7
wt
8
wt
FIG 5. Single-stranded conformational polymorphism (SSCP) gel electrophoresis. DNA isolated from paraffin tissue blocks was amplified using primers specific for p53 exon 5 (left panel) or p53 exon 6 (right panel) as described. Following denaturation and gel electrophoresis, gels were dried and autoradiographed. Cases 18 (lane 1) and 26 (lane 5) showed altered mobility bands distinct from the germ line pattern. Negative (wild-type target DNA, lanes 2-4 and 7-8, wt) and no added DNA (lane 6, MM) control reactions gave appropriate results.
pattern (Fig. 6, middle panels) consistent with an insertion or deletion. The SSCP pattern in this case was complex and the altered mobility species in the SSCP gel could not be reliably separated from germline bands (Fig. 5, right panel). Therefore, the mutant PCR product was cloned and sequenced. This demonstrated a deletion of 3 nucleotides between nucleotides 1149 and 1153 (2 cy-tosines and 1 thymidine). This deletion results in the loss of proline at codon 190 of exon 6 without associated amino acid substitutions.
In both cases with p53 gene mutations, the overall percentage of RS-H cells with p53 expression was high compared to cases without gene mutations (23% and 20%, respectively versus 3%; P <.05, Student t test). Because the relatively low percentage of RS-H cells in some cases of Hodgkin's disease could compromise the ability of the
PCR-SSCP assay to detect p53 gene mutations, we evaluated its sensitivity by combining germline DNA with DNA from tumors with different p53 exon 5 gene mutations. The altered mobility bands resulting from two different exon 5 mutations are clearly visible with DNA from each primary tumor which contained little germline DNA (Fig. 7). The addition of germline DNA to these samples reduced the relative intensity of the altered mobility bands as expected. With one mutation (Fig. 7), this band remained clearly visible even when tumor DNA constituted only 1 % of the sample (99% germline p53 DNA sequences present). The altered mobility band resulting from the other p53 mutation was not as well separated from germline bands but could be reliably detected when tumor DNA constituted only 5% of the DNA sample (data not shown).
p53 gene mutations were accurately predicted by a p53 positive, mdm-2 negative immunophenotype (P <.005, chi square). There was a strong association between p53 gene mutation and p53 protein expression by a high percentage of RS-H cells (68% versus 16%; P <.05, Student's t test). There was also a strong association between p53 mutation and a high overall percentage of cells expressing p53 (22% vs. 3%; P <.01, Student's t test). Although cases with p53 gene mutations had a higher percentage of RS-H cells (34% versus 16%), this difference was not statistically significant (P >.10, Student's t test). Ki-67 staining RS-H cells were significantly more abundant in cases with p53 gene mutations (81% vs. 64%; P <.02, Student's t test). There was no significant association between the presence of p53 gene mutations and histologic subtype, clonal IgH gene rearrangement, or EBV-LMP expression. p53 gene mutations occurred in younger patients (16 and 32 years versus a median of 36 years) in this study, but this difference did not reach statistical significance (P >. 10, Student's t test)
DISCUSSION
We observed strong immunohistochemical staining for p53 in more than 10% of the RS-H cell nuclei in 12 of 30 cases (40%) of Hodgkin's disease. This result is consistent with other studies that have reported p53 protein expression in 32% to 70% of cases of Hodgkin's disease.3233
Using a sensitive PCR-SSCP assay, we detected p53 gene mutations in only two of these cases. The low percentage of RS-H cells in some cases of Hodgkin's disease is a possible explanation for the discrepancy between im-munohistochemistry and genetic analysis. We believe that this explanation is unlikely to explain our results given the relatively high sensitivity of the PCR-SSCP as-
A.J.C.P.-December 1996
Dow
nloaded from https://academ
ic.oup.com/ajcp/article/106/6/728/1756733 by guest on 30 Septem
ber 2022

ELENITOBA-JOHNSON ET AL. 735
p53 Mutations in Reed-Sternberg Cells
FIG 6. DNA sequencing gels for p53 exons 5 and 6. PCR products from both cases with evidence of p53 gene mutations by SSCP (cases 18 and 26) were subjected to chain termination DNA sequencing. A single transversion (A -»- T) in exon 5 of case 18 is shown (left panel, arrow). The right-middle panel shows the sequencing pattern derived from PCR products with unfractionated tumor DNA from case 26 for p53 exon 6 using an antisense sequencing primer. The germline pattern for p53 exon 6 is shown for comparison (Fig. 6, left-middle panel). The tumor DNA sequence develops ghost bands after nucleotide 1,151 (arrow). Sequencing of cloned PCR products from case 26 corresponding to the altered mobility bands (right panel) shows a 3 nucleotide deletion relative to the germline sequence corresponding to the point of divergence from the germline pattern (arrows).
p53 exon 5
case #18
say (c.a. 1-5% mutant cells; Fig. 7) and the relatively high percentage of RS-H cells in most of our cases. Many of the p53 positive cases without detectable p53 gene mutations had a percentage of RS-H cells equal to or greater than that in the two cases with p53 gene mutations. Most of these cases (81%) had percentages of RS-H cells and p53 positive RS-H cells in excess of the PCR-SSCP detection limit of 5%. Our results do not exclude the possibility of unusual mutations outside exons 5-8 of the p53 gene in these cases, although less than 10% of p53 gene mutations occur outside these exons in most tumor types.5 Although we included cases of lymphocyte predominance and lymphocyte depletion in this analysis, 90% of these cases are "classical Hodgkin's disease" (nodular sclerosis or mixed cellularity) and the results for "classical Hodgkin's disease" are virtually the same (44% of cases with significant p53 expression and 7% with detectable p53 gene mutations).
Similar results demonstrating the rarity of p53 gene mutations in Hodgkin's disease have been reported by others using different techniques. Using a heteroduplex PCR-mismatch technique, Xerri34 failed to detect p53 gene mutations in ten cases of Hodgkin's disease selected for a high percentage of RS-H cells with p53 protein expression. Inghirami35 found a p53 mutation in 1 of 11 patients that was conserved in geographically distinct disease foci. Trumper and colleagues36 found p53 gene mutations in only one of seven samples using a single RS-H cell isolation method.
In their single positive case, the p53 mutation was present in only a subset of the RS-H cells suggesting that there can be heterogeneity in the occurrence of p53 gene mutations in RS-H cells in a given case. This result is
consistent with the idea that p53 gene alterations may be a late event in the development of Hodgkin's disease. Other studies documenting the presence of p53 gene mutations in a larger percentage of Hodgkin's disease derived cell lines may not accurately reflect in vivo conditions because p53 mutations can confer a selective growth advantage in the establishment of cell lines.37
Wild-type p53 protein can be functionally inactivated by interaction with proteins encoded by some viruses including human papilloma virus and adenovirus.38"40 Although EBNA-1 could potentially interact with p53 in EBV positive cases of Hodgkin's disease to modulate its function and half-life, we found no correlation between EBV-LMP and p53 expression in RS-H cells, in agreement with the results of others.33 Interestingly, both cases with p53 gene mutations were also EBV positive. This is consistent with the idea that EBV is not directly related to p53 expression in Hodgkin's disease.
Some cellular proteins, most notably the mdm-2 protein, can bind to and modulate p53 activity and half-life in a manner analogous to viral proteins.19 The finding of concurrent mdm-2 and p53 expression in the RS-H cells in this study suggests that the two proteins may functionally interact. The mdm-2 protein can bind to and modulate the function of the retinoblastoma gene product (Rb) as well as p53 and could exert potent control over cell cycle progression at this point.41 Available evidence suggests that the interaction between mdm-2 and p53 differs in different tumor cell types. One third of non-Hodgkin's lymphomas that express wild-type p53 protein showed concurrent expression of mdm-2.42 In contrast, testicular germ cell tumors, which rarely if ever have p53 gene mutations, showed a strong correlation
Vol. 106-No. 6
Dow
nloaded from https://academ
ic.oup.com/ajcp/article/106/6/728/1756733 by guest on 30 Septem
ber 2022

736 HEMATOPATHOLOGY
p 5 3 e x o n 5 S S C P
P e r c e n t M u t a n t D N A
FIG 7. Single-stranded conformational polymorphism (SSCP) sensitivity. DNA from an ovarian tumor with a p53 exon 5 gene mutation was analyzed by PCR-SSCP either alone (M) or after dilution with germline DNA to achieve the indicated percentages of mutant DNA (l%-50%). Wild-type DNA (wt) and no added DNA controls (-) are also included. The altered mobility bands are clearly visible when tumor DNA constitutes only 4% of the sample in this autoradiogram.
between mdm-2 and p53 protein expression.43 A similar strong correlation was observed in acute lymphoblastic leukemia cell lines with expression of wild-type p53 protein.44
Discordance for p53 and mdm-2 expression has been observed in other systems. For instance, Barbareschi found that 20% of transitional cell carcinomas with mdm-2 expression co-expressed p53 while only 7% of p53 expressing tumors co-expressed mdm-2.45 p53 gene mutations were not evaluated in this study and the observed discordance could be explained by the relatively high frequency of p53 gene mutations in transitional cell carcinoma. Similar poor correlation between mdm-2 and p53 protein expression was observed in a series of breast carcinomas.46
Adenovirus transformation which induces high level p53 expression in vitro does not result in expression of mdm-2.39
A.J.C.P.-
Article
Taken together, these results from different systems suggest that mdm-2 expression may drive p53 expression in some situations and in other cases p53 expression may promote mdm-2 expression. The consequences of these two scenarios are likely to be different because both of these proteins have other potential binding partners that can influence gene expression, cell cycle progression, and the induction of apoptosis. Our findings in Hodgkin's disease are consistent with the idea that mdm-2 expression may promote p53 expression in RS-H cells without p53 gene mutations.
Studies have shown that RS-H cells exhibit a high proliferative index as judged by Ki-67 staining.47 Paradoxically, mitoses are only rarely observed in RS-H cells consistent with the idea that RS-H cells are arrested in the cell cycle.48 p53 expression correlates closely with growth arrest at G|S induced by DNA damage.49"50
These findings suggest a possible role for p53 protein in the growth arrest of the proliferating RS-H cells. We found a high but variable percentage of Ki-67 positive RS-H cells (45%-90%; median 72%). Interestingly the highest percentage of Ki-67 staining was observed in the two cases with p53 gene mutations. Although there was no correlation between the percentage of Ki-67 and p53 positive RS-H cells, it seems possible that p53 expression may be responsible for the high percentage of Ki-67 staining in the RS-H cells.
Tumors with p53 gene mutations are relatively resistant to most chemotherapeutic agents and radiation. It is not known whether p53 gene mutations and/or protein expression are predictive of poor treatment response in Hodgkin's disease. An unexpected finding in this study was a strong association between the presence of p53 gene mutations and a higher percentage of Ki-67 staining (81% versus 64%; P <.05) in RS-H cells. We also found an association between clonal IgH rearrangement and the percentage of cells over-expressing p53. Both of these features are potential markers of disease aggressiveness.52
In conclusion, we have confirmed that immunohisto-chemical detection of p53 protein in RS-H cells is relatively common in Hodgkin's disease (53% of our cases), whereas p53 gene mutations are infrequent (<10% of p53-positive cases). The sensitivity of the PCR-SSCP assay employed in this study suggests that the low percentage of RS-H cells in many cases of Hodgkin's disease is not the main reason for the observed discordance between immunohistochemical detection of p53 protein and p53 gene mutations, mdm-2 and/or other cellular proteins may be responsible for p53 expression RS-H cells without detectable p53 gene mutations.
Acknowledgments. The authors thank Ms. Anna Chromiak, MS, for her expert technical assistance in the performance of this study.
1996
Dow
nloaded from https://academ
ic.oup.com/ajcp/article/106/6/728/1756733 by guest on 30 Septem
ber 2022

ELENITOBA-JOHNSON ET AL. 737
p53 Mutations in Reed-Sternberg Cells
REFERENCES
1. Hollstein M, Sidransky D, Vogelstein B, HarrisCC. p53 mutations in human cancers. Science 1991;253:49-53.
2. Levine AJ, Momand J, Finlay CA. The p53 tumor suppressor gene. Nature 1991,351:453-456.
3. Martinez J, Georgoff I, Levine AJ. Cellular localization and cell cycle regulation by a temperature-sensitive p53 protein. Genes Dev 1991;5:151-159.
4. Nigro JM, Baker SJ, Preisinger AC, et al. Mutations in the p53 gene occur in diverse human tumour types. Nature 1989;342: 705-708.
5. Caron de Fromentel C, Soussi T. p53 tumor suppressor gene: A model for investigating human mutagenesis. Genes Chrom Cancer 1992;4:1-15.
6. Cordon-Cardo C, Latres E, Drobnjak M, et al. Molecular abnormalities of mdm2 and p53 genes in adult soft tissue sarcomas. Cancer Res 1994;54:794-799.
7. Eliyahu D, Raz A, Gruss P, Givol D, Oren M. Participation of p53 cellular tumour antigen in transformation of normal embryonic cells. Nature 1984; 312:646-649.
8. Iggo R, Gatter K, Bartek J, Lane D, Harris AL. Increased expression of mutant forms of p53 oncogene in primary lung cancer. Lancet 1990;335:675-679.
9. Wynford-Thomas D. p53 in tumor pathology: Can we trust immu-nohistochemistry?7 Pathol 1992; 166:329-330.
10. Pasman PC, Tiesbosch A, Erdkamp FL, Vrints LW, Breed WP, Schouten HC. p53 as a marker of the malignant cell in Hodg-kin's disease. Ann Oncol 1994;5(suppl 7):89-91.
11. Chilosi M, Montagna L, Lestani M, Menestrina F, Mariuzzi GM. Quantitative approach to the immunohistochemical analysis of p53 antioncogene and CD30 antigen in Hodgkin's disease. In Vivo 1993;7:393-394.
12. Hubbert NLSedman SA, Schiller JT. Human papillomavirus type 16 E6 increases the degradation rate of p53 in human keratinoo-cytes. J Virol 1992;66:6237-6241.
13. Luka J, Jornvall H, Klein G. Purification and biochemical characterization of the Epstein-Barr virus-determined nuclear antigen and an associated protein with a 53,000-dalton subunit. J Virol 1980;35:592-602.
14. Weiss LM, Strickler JG, Warnke RA, Purtilo DT, Sklar J. Epstein-Barr viral DNA in tissues of Hodgkin's disease. Am J Pathol 1987;129:86-91.
15. Herbst H, Dallenbach F, Hummel M, et al. Epstein-Barr virus latent membrane protein expression in Hodgkin and Reed-Sternberg cells. Proc Natl Acad Set USA 1991;88:4766-4770.
16. Herbst H, Niedobitek G, Kneba M, et al. High incidence of Epstein-Barr virus genomes in Hodgkin's disease. Am J Pathol 1990;137:13-18.
17. Deacon EM, Pallesen G, Niedobitek G, Brooks L, Rickinson AB, Young LS. Epstein-Barr virus and Hodgkin's disease: Transcriptional analysis of virus latency in the malignant cells. J Exp Med 1993; 177: 339-349.
18. Moorthy RK, Thorley-Lawson D. Biochemical, genetic, and functional analyses of the phosphorylation sites on the EBV-encoded oncogenic latent membrane protein LMP-1. J Virol 1993;67: 2637-2645.
19. Momand J, Zambetti GP, Olson DC, George D, Levine AJ. The mdm-2 oncogene product forms a complex with the p53 protein and inhibits p53-mediated transactivation. Cell 1992;69:1237-1245.
20. Chilosi M, Doglioni C, Menestrina F, et al. Abnormal expression of the p53-binding protein mdm-2 in Hodgkin's disease. Blood 1994;84:4295-4300.
21. Vidal-Puig A, Moller DE. Comparative sensitivity of alternative single-strand conformation polymorphism (SSCP) methods. BioTechniques 1994; 17:490-496.
22. Gown AM, de Wever N, Battifora H. Microwave-based antigenic unmasking. A revolutionary new technique for routine immu-nohistochemistry. Appl Immunohislochem 1993; 1:256-266.
23. Forsthoefel KF, Papp AC, Snyder PJ, Prior TW. Optimization of DNA extraction from formalin fixed tissue and its clinical application in Duchenne muscular dystrophy. Am J Clin Pathol 1992;98:98-104.
24. Matsushima AY, Cesarman E, Chadburn A, Knowles DM. Post-thymic T-cell lymphomas frequently overexpress p53 protein but infrequently exhibit p53 gene mutations. Am J Pathol 1994;144:573-584.
25. Orita M, Suzuki Y, Hayashi K. Rapid and sensitive detection of point mutations and DNA polymorphisms using the polymerase chain reaction. Genomics 1989;5:874-879.
26. Elenitoba-Johnson, K, Medeiros, U, Khorsand, J, KingTC. Lymphoma of the mucosa-associated lymphoid tissue of the lung: A multifocal case of common clonal origin. Am J Clin Pathol 1995;103:341-345.
27. McCarthy KP, Sloane JP, Kabarowsi JHS, Matutes E, Wiedmann LM. A simplified method of detection of clonal rearrangements of the T-cell receptor-gamma chain gene. Diagn Mol Pathol 1992;1:173-179.
28. Chittal SM, Caveriviere P, Schwarting R, et al. Monoclonal antibodies in the diagnosis of Hodgkin's disease: The search for a rational panel. Am J Surg Pathol 1988; 12:9-21.
29. Kamel OW, Chang PP, Grank BS, et al. Clonal VDJ recombination of the immunoglobulin heavy chain gene by PCR in classical Hodgkin's disease. AmJClin Pathol 1995;104:419-423.
30. Orazi A, Jiang B, Lee C-H, et al. Correlation between presence of clonal rearrangements of immunoglobulin heavy chain gene and B-cell antigen expression in Hodgkin's disease. Am J Clin Pathol 1995; 104:413-418.
31. Buchman VL, Chumakov PM, Ninkina NN, Samarina OP, Geor-giev GP. A variation in the structure of the protein coding region of the human p53 gene. Gene 1988;70:245-252.
32. Doglioni C, Pelosio P, Mombello A, Scarpa A, Chilosi M. Immunohistochemical evidence for abnormal expression of the anti-oncogene-encoded p53 phosphoprotein in Hodgkin's disease and CD30+ anaplastic lymphomas. Hematol Pathol 1991;5: 67-73.
33. Niedobitek G, Rowlands DC, Young LS, et al. Overexpression of p53 in Hodgkin's disease: Lack of correlation with Epstein-Barr virus infection. J Pathol 1993; 169:207-212.
34. Xerri L, Pare P, Bouabdallah R, Camerlo J, Hassoun J. PCR-mis-match analysis of p53 gene mutation in Hodgkin's disease. J Pathol 1995; 175:189-194.
35. Inghirami G, Macri L, Rosati S, Zhu BY, Yee HT, Knowles DM. The Reed-Sternberg cells of Hodgkin disease are clonal. Proc Natl Acad Sci USA 1994;91:9842-9846.
36. Trumper LH, Brady G, Bagg A, etal. Single-cell analysis of Hodgkin and Reed-Sternberg cells: Molecular Heterogeneity of gene expression and p53 mutations. Blood 1993;81:3097-3115.
37. Gupta RK, Patel K, Bodmer WF, Bodmer JG. Mutation of p53 in primary biopsy material and cell lines from Hodgkin's disease. Proc Natl Acad Sci USA 1993;90:2817-2821.
38. Yew PR, Berk AJ. Inhibition of p53 transactivation required for transformation by adenovirus early IB protein. Nature 1992;357:82-84.
39. Grand RJ, Lecane PS, Owen D, et al. The high levels of p53 present in adenovirus early region 1-transformed human cells do not caouse up-regulation of MDM2 expression. Virology I995;210: 323-334.
40. Lane DP, Crawford LV. T antigen is bound to a host protein in SV40 transformed cells. Nature 1979;278:261-263.
41. XiaoZX, Chen J, Levine AJ, et al. Interaction between the retinoblastoma protein and the oncoprotein mdm-2. Nature 1995;375:694-698.
Vol. 106-No. 6
Dow
nloaded from https://academ
ic.oup.com/ajcp/article/106/6/728/1756733 by guest on 30 Septem
ber 2022

738 HEMATOPATHOLOGY
Original Article
42. Maestro R, Gloghini A, Boglioni C, et al. MDM2 overexpression does not account for stablization of wild-type p53 in non-Hodg-kin's lymphomas. Blood 1995;85:3239-3246.
43. Riou G, Barrois M, Prost S, Terrier MJ, Theodore C, Levine AJ. The p53 and mdm-2 genes in human testicular germ-cell tumors. Mol Carcinog 1995; 12:124-131.
44. Zhou M, Yearer AM, Smith SD, Findley HW. Overexpression of the MDM2 gene by childhood acute lymphoblastic leukemia cells expressing the wild-type p53 gene. Blood 1995; 85:1608-1614.
45. Barbareschi M, Girlando S, Fellin G, Graffer U, Luciani L, Dalla Palma P. Expression of mdm-2 and p53 protein in transitonal cell carcinoma. Urol Res 1995;22:349-352.
46. McCann AH, Kirley A, Carney DN, et al. Amplificaton of the MDM2 gene in human breast cancer and its associaton with MDM2 and p53 protein status. Br J Cancer 1995,71:981-985.
47. Gerdes J, Van Baarlen J, Pileri S, Schwarting R, Van Unnik JAM, Stein H. Tumor cell growth fraction in Hodgkin's disease. Am J Pathol 1987; 129:390-393.
48. Pecham MJ, Cooper EH. Proliferation characteristics of varying classes of cell in Hodgkin's disease. Cancer 1969;24:135-139.
49. Kuerbitz SJ, Plunkett BS, Walsh WV, Kastan MB. Wild type p53 is a cell cycle checkpoint determinant following irradiation. Proc Natl AcadSci USA 1992;89:7491-7495.
50. Fritsche M, Haessler C, Brandner G. Production of nuclear accumulation of the tumor-suppressor protein p53 by DNA-damag-ingagents. Oncogene 1993;8:307-318.
51. Lin D, Shields MT, Ullrich SJ, Appella E, Mercer WE. Growth arrest induced by wild-type p53 protein blocks cells prior to or near the restriction point in late G1 phase. Proc Natl Acad Sci USA 1992;89:9210-9214.
52. Hummel M, Ziemann K, Lammert H, PilerS, Sabattini E, Stein H. Hodgkin's disease with monoclonal and polyclonal populations of Reed-Sternberg cells. N Engl J Med 1995;333:901-906.
A.J.C.P. -December 1996
Dow
nloaded from https://academ
ic.oup.com/ajcp/article/106/6/728/1756733 by guest on 30 Septem
ber 2022





























