Embed Size (px)
Citation preview

Ri
M*Va
R
pcremHdraoteatmmpps
aa
pbtccwi(
bbAc
Biochemical and Biophysical Research Communications 273, 272–281 (2000)
doi:10.1006/bbrc.2000.2931, available online at http://www.idealibrary.com on
0CA
estructured Transactivation Domainn Hamster AH Receptor
erja Korkalainen,* Jouko Tuomisto,* and Raimo Pohjanvirta*,†,‡Laboratory of Toxicology, National Public Health Institute, P.O. Box 95, FIN-70701 Kuopio, Finland; †Nationaleterinary and Food Research Institute, Regional Laboratory of Kuopio, Kuopio, Finland; and ‡Department of Foodnd Environmental Hygiene, Faculty of Veterinary Medicine, University of Helsinki, Helsinki, Finland
eceived May 19, 2000
molecules of the chaperone hsp90, certain immunophi-lirdaceiavdv
urTslptTv
Tos(WttitcmTpfih
Hamsters and Han/Wistar (Kuopio; H/W) rats showeculiarly selective responsiveness to 2,3,7,8-tetra-hlorodibenzo-p-dioxin (TCDD). They are extremelyesistant to its acute lethality but sensitive to, e.g.,nzyme induction. The biological effects of TCDD areediated by the AH receptor (AHR). Recent studies on/W rat AHR discovered a remodelled transactivationomain which appears to be critical for the TCDDesistance of these animals. Here, molecular cloningnd sequencing of hamster AHR reveals another typef restructured transactivation domain. In hamsters,he functionally pivotal Q-rich region is substantiallyxpanded and enriched in glutamine compared withll other AHRs cloned to date. By contrast, the amino-erminal end is highly conserved, which is in agree-ent with the H/W rat AHR. Because of the additionalaterial in the transactivation domain, hamster AHR
rotein is larger than that in rats or mice, but theattern of AHR mRNA expression in tissues isimilar. © 2000 Academic Press
Key Words: 2,3,7,8-tetrachlorodibenzo-p-dioxin; TCDD;ryl hydrocarbon receptor; hamster; cloning; trans-ctivation; species differences.
2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) is therototype for a class of halogenated aromatic hydrocar-ons that are widespread and persistent environmen-al contaminants. Most of the biological effects of theseompounds are mediated by the aryl hydrocarbon re-eptor (AHR), a ligand-activated transcription factor,hich structurally belongs to a newly discovered fam-
ly of regulatory proteins: basic helix-loop-helixbHLH)/PAS transcription factors (1).
The molecular mechanism of AHR action has so fareen only elucidated for CYP1A1 induction, but this iselieved to be a general mode of gene regulation by theHR. In an inactive state, the AHR is located in the
ytoplasm in a protein conglomerate along with two
272006-291X/00 $35.00opyright © 2000 by Academic Pressll rights of reproduction in any form reserved.
ins and c-src. Binding of ligand such as TCDD resultsn translocation of the AHR into the nucleus and dis-uption of the complex. Inside the nucleus, the AHRimerizes with a related bHLH/PAS protein, ARNT,nd then binds to DNA at specific sites containing aonsensus hexanucleotide core. These dioxin responselements act as enhancers for genes regulated by diox-ns. Since the enhancer sites are usually situated rel-tively far upstream of the gene promoter, gene acti-ation by dioxins probably involves nucleosomalisruption and interaction with transcriptional coacti-ators and/or corepressors (2).The AHR protein consists of distinct functional mod-
les. The bHLH domain located in the N terminus isesponsible for DNA binding and heterodimerization.he PAS motif flanking the bHLH structure affordspecificity to dimerization and also contains most of theigand-binding domain. The C terminus comprises aotent transactivation domain composed of several in-eracting subdomains, one of which is a Q-rich subunit.his has been shown to be essential for the transacti-ation function of the AHR in vitro (3, 4).Hamsters are highly resistant to the acute toxicity of
CDD. Compared with guinea pigs, they tolerate anver 1000-fold higher dose (5). The difference of theame magnitude exists between a TCDD-sensitiveLong-Evans, Turku AB; L-E) and resistant (Han/
istar Kuopio; H/W) rat strain (6). However, despiteheir resistance hamsters and H/W rats are susceptibleo certain biochemical and even toxic effects of TCDDncluding enzyme induction, thymus atrophy and feto-oxicity (7). Recent cloning of H/W rat AHR revealedhanges in the architecture of the transactivation do-ain, which appear to be the principal reason forCDD resistance in H/W rats (8, 9). Therefore, therimary structure of hamster AHR was determined tond out if TCDD resistance has a similar basis inamsters and to gain better understanding of the role

os
M
wTfgs
hiAtwCGGGCAFGGRCAC
tEcrOcpw(tmaffit
wPpaw
wwfTptOw1wsSopw(APG
cCwrmAspnssr
wt1mefsbr4rlFrM
2sc
h
Vol. 273, No. 1, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
f the AHR structure as a determinant of dioxin sen-itivity.
ATERIALS AND METHODS
Animals. Two golden Syrian hamsters, a male and a female,ere purchased from Harlan Nederland (Horst, The Netherlands).he hamsters were killed by decapitation at the age of 4 weeks. The
ollowing tissues were rapidly removed, flash-frozen in liquid nitro-en, and stored at 280°C until analysis: liver, lung, heart, kidney,pleen, thymus, hypothalamus, testis and ovary.
Primers. The first primers (F137 and R1096) were selected fromighly conserved regions of the AHR. Thereafter, one of the primers
n subsequent PCR reactions was always chosen from the segment ofHR cDNA already resolved. The primers are shown in relation to
he full-length AHR cDNA in Fig. 1. Their primary structures (59-39)ere as follows: Anchor-RT, CCA-CCT-ACC-CAC-TAC-CCC-TCT-AT-T(22)-V; Anchor-R, CCA-CCT-ACC-CAC-TAC-CCC-TCT-CA;SP1, GCT-TGT-TAA-TAA-CAT-CTT-GTG; GSP2, TGT-CTC-TAT-TC-GCT-TAG-AAG; GSP3, GGA-CTG-GCT-TCA-CTG-TTT-TCT-CA; Biotseq, CTT-GCT-TAG-GAA-CGC-CTG-GGA; Biotcomp,CC-AGG-CGT-TCC-TAA-GCA-A; F2406, CCC-GGC-GTT-CCA-GT-GTG-CAC-A; F2501, TGT-TTA-CAA-GTT-CCT-GAA-AAC-C;2584, CAT-GTC-CAT-GTA-YCA-GTG-CCA; F137, GCT-GAA-GA-ATY-AAG-TCA-AA; F820, GAA-CAA-GAA-GGG-GAG-GAT-GA; R1096, CAT-GCC-ACT-TTC-TCC-AGT-CTT-AAT-CAT;1248, CAG-CGT-CAT-ACT-CCG-CTT-CTG; F1752, CAA-ACG-AT-GCA-GAG-TGA-GGA; R1826, CAC-GTA-AGT-GAG-GAT-TTC-TC-TG; R2904, TGG-ACA-GAC-AGT-CCC-GGT-ATC; R3023,TC-TCC-CTA-TGC-TCC-TGC-AGA.
RT-PCR for cloning. The liver samples were homogenized andotal RNA was isolated using Trizol reagent (Life Technologies,ggenstein, Germany). Protein was isolated simultaneously (10).DNA was synthesized with enhanced avian myeloblastosis viruseverse transcriptase (AMV-RT) (Sigma, St. Louis, MO) or withmniscript reverse transcriptase (Qiagen, Hilden, Germany). For
DNA synthesis, either oligo-dT or random hexamers were used asrimers in all cases except for cloning of the ends (see below). PCRas performed with DyNAzyme EXT DNA polymerase blend
Finnzymes, Espoo, Finland) and with either Uno II or TGradienthermocycler (Biometra, Gottingen, Germany). The “touchdown”ethod was applied throughout all reactions: the annealing temper-
ture was set at about 5°C above the melting temperature of primersor the first cycle and then decreased by 1°C/cycle down to the desirednal temperature (determined with the aid of the gradient feature ofhe TGradient thermocycler). A total of 35 cycles were usually run.
To obtain the 39 end of the cDNA, an anchored primer (Anchor-RT)as employed in the RT phase followed by a total of 3 seminestedCR reactions (Anchor-R as reverse primer with 3 different forwardrimers: F2406, F2501 and F2584). The 59 end was first pursued bymodified ligation-based method (11). In brief, RT was conductedith a gene specific primer (GSP1). cDNA was purified of the primer
FIG. 1. Location of primers used in PCR for cloning the hamelix-loop-helix region (bHLH), PAS A and B repeat segments and t
273
ith the Qiaquick PCR Purification Kit (Qiagen). The RNA templateas degraded by alkaline treatment (0.3N NaOH for 30 min at 50°C
ollowed by neutralization with 0.3 M acetic acid for 30 min at 50°C).he single-stranded cDNA was concentrated with Pellet Paint co-recipitant (Novagen, Madison, WI). A special 29-base oligonucleo-ide (Biotseq) with a phosphate group at its 59 end and the 39 endH-group blocked with biotin was ligated to the 39 end of the cDNAith T4 RNA ligase (MBI Fermentas, Vilnius, Lithuania) (22°C for6 h, 25°C for 6 h). Two seminested PCR reactions were then runith a constant forward primer (Biotcomp; complementary to Biot-
eq) and two nested gene specific reverse primers (GSP1 and GSP2).ince this method yielded a product suggestive of alternative splicingf exon 1 (see below), the result was verified with a different ap-roach. A homopolymeric dA tail was added to the cDNA generatedith the GSP1 primer using terminal deoxynucleotidyl transferase
MBI Fermentas). The first PCR reaction was performed with thenchor-RT and GSP1 primers. This was followed by two seminestedCR reactions with Anchor-R as the forward primer and GSP2 orSP3 as the reverse primer.
Cloning and sequencing. PCR products were cloned by blunt-endloning into pCR-Script SK(1) Amp plasmid (Stratagene, La Jolla,A) or pT7Blue-2 plasmid (Novagen). XLB-1 supercompetent cellsere used in transformations (Stratagene). The plasmids were pu-
ified by Wizard Plus SV Minipreps DNA Purification System (Pro-ega, Madison, WI). The inserts were sequenced with an A.L.F. or.L.Fexpress DNA sequencer (Amersham Pharmacia Biotech, Upp-ala, Sweden) using either Thermo Sequenase fluorescent labelledrimer cycle sequencing kit or Thermo Sequenase CY5 Dye Termi-ator Kit (Amersham Pharmacia Biotech). All ambiguities were re-olved by auxiliary clonings. The entire coding region was cloned andequenced from both the female and male hamster with identicalesults.
Northern blotting. Poly(A)1 -RNA was purified from total RNAith oligo dT-coated latex beads (Oligotex midi, Qiagen) and concen-
rated using Pellet Paint Co-Precipitant (Novagen). 2 ml 10 3 MOPS,0 ml formaldehyde and 3.3 ml formamide were added to 4.7 mlRNA sample, denatured for 10 min at 70°C and chilled on ice. After
lectrophoresis in a 1.2% formaldehyde gel, the samples were trans-erred onto a positively charged nylon membrane (Hybond1, Amer-ham Pharmacia Biotech) by capillary diffusion overnight. Prehy-ridization and hybridization were accomplished in bottles in aotisserie-equipped hybridization oven (Hybaid, Teddington, UK) at2°C in ULTRAhyb hybridization solution (Ambion, Austin, TX). Aat oligo showing 100% homology to hamster AHR was 39 end-abelled using digoxigenin-ddUTP and terminal transferase (MBIermentas). Detection was performed with the colorimetric detectioneagents NBT/BCIP according to manufacture’s instructions (Roche,annheim, Germany).
Tissue specific expression. RT-PCR was carried out from 60 ng or00 ng of total RNA by RobusT RT-PCR Kit (Finnzymes). Sequencepecific primers from exon 10 (F1752) and exon 11 (R2904) of theoding region of hamster AHR were used (spanning of intron 10
r AHR (F, forward, and R, reverse). The positions of the basicglutamine-rich subdomain (Q) are indicated.
stehe

padEc927
s
ctaoumft
h
ot
Vol. 273, No. 1, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
revented amplification of genomic DNA). The 12.5 ml RT-PCR re-ction contained 4 ml template, 2.5 pmoles of primers, 200 mM eachNTP, 1.25 units AMV reverse transcriptase, 0.5 units DyNAzymeXT DNA polymerase, 1.5 mM MgCl2 in RobusT reaction buffer. The
ycling conditions were: 46°C for 60 min, 94.5°C for 2 min 25 sec;4.5°C for 35 sec, 66°C (1°C decrement per cycle) for 40 sec; 72°C formin 3 7; 94.5°C for 35 sec, 59°C for 40 sec, 72°C for 2 min 3 25;
2°C for 10 min.
In vitro protein synthesis. The full-length hamster AHR con-tructed by PCR was subcloned into a mammalian expression vector
FIG. 2. Nucleotide and deduced amino acid sequence of the hamsn the right. The termination codon is indicated with an asterisk (*)ext) are underlined.
274
ontaining the expression cassette from pCI plasmid (Promega) inhe polylinker of pCR-Script SK(1) AMP plasmid (Stratagene) withkozak sequence incorporated immediately upstream the first ATG
f the insert. In vitro transcription and translation were carried outsing Single Tube Protein System3 (Novagen). 0.5 mg or 1 mg plas-id DNA was added to T7 transcription mix and incubated at 30°C
or 30 min. Methionine and translation mix were added directly toranscription reaction and incubation was continued for 2 hours.
Western blotting. The in vitro-generated proteins as well as theamster lung and liver protein fraction were analyzed by immuno-
AHR cDNA. Amino acids are numbered on the left and nucleotidespetitive DNA sequences and the possible alternative start site (see
ter. Re

bpR
nua
Vol. 273, No. 1, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
lotting after sodium dodecylsulfate–polyacrylamide gel electro-horesis (SDS–PAGE). A 6% polyacrylamide minigel was run (Bio-ad, Hercules, CA). The proteins were transfered onto a
FIG. 2—
275
itrocellulose membrane and then subjected to Western blot analysissing anti-AHR, rabbit polyclonal antibody (1:50,000) as a primaryntibody (BioMol Research Laboratories, Plymouth Meeting, PA)
ntinued
Co
ab
R
c
awwtiw
idN
Vol. 273, No. 1, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
nd an alkaline phosphatase-labelled anti-rabbit IgG as a second anti-ody (Roche). The bands were visualized with NBT/BCIP (Roche).
ESULTS
The coding region of hamster AHR cDNA proved toontain 2763 nucleotides encoding a protein of 921
FIG. 3. Comparison of hamster, rat, mouse, and human AHRnserted to improve alignment. The basic helix-loop-helix region, PASomain and LXCXE motif by a double line (20). The sequence aligumbers of the AHR sequences used are: Sprague-Dawley rat (U09
276
mino acids (Fig. 2). The calculated molecular weightas 7.1 kDa larger than the wild-type rat receptor,hich was due to auxiliary material in the C-terminal
ransactivation domain. The cDNA contained 6 repet-tive sequences of satellite-like DNA incorporated to-ards the 39 end of the receptor (underlined in Fig. 2).
uences. Dots indicate identities while dashed lines indicate gapsmain and glutamine (Q) rich domain are indicated by lines over the
ent was performed using Clustal W (1.7). The GenBank Accession), C57BL/6 mouse (M94623), and human (L19872).
seqdo
nm000

Thcsso4s2otaatatta
Nlcboas
Titlact6
DNNHDpT
lb
Vol. 273, No. 1, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
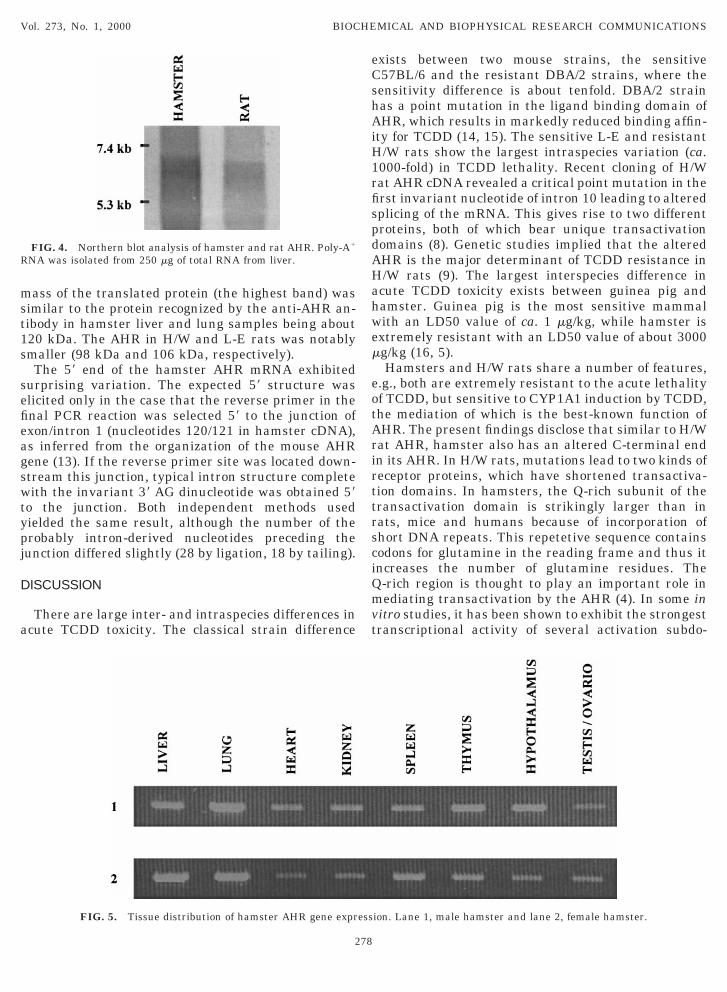
hus the C-terminus of the translated protein wouldave some 60 extra amino acids compared with theorresponding region in the rat AHR (Fig. 3). Morepecifically, this change affected the Q-rich subdomaino that this region was much larger in hamster than inther AHRs cloned to date. The Q-rich region contains9 glutamine residues in hamster, while the corre-ponding figure for rat, mouse, and human receptors is8, 27, and 25, respectively. The amino acid sequencesf rat, mouse and human AHRs were compared withhe corresponding hamster sequence (Fig. 3). Thisnalysis revealed a high degree of conservation in themino terminal half of the hamster receptor includinghe basic helix-loop-helix and PAS domains. The aminocid identity was lowest in the C-terminal transactiva-ion domain (Table 1). The overall amino acid identityo the rat, mouse and human receptors was 70%, 82%nd 61%, respectively.The mRNA size of hamster AHR was determined byorthern blot analysis (Fig. 4). It turned out to be
arger than in rat, approximately slightly over 7 kb asompared with 6.6 kb in rat (12). Based on Northernlot data, the UTR we resolved by 39 race-PCR wasnly partial (expected size ca. 4.2 kb), probably due toshort stretch of successive adenines in the UTR re-
ulting in unspecific annealing of the anchor primer.
FIG. 3—
277
he tissue distribution of the hamster AHR was stud-ed by the RT-PCR method (Fig. 5). Liver, lung andhymus expressed the highest levels of AHR, withower levels observed in hypothalamus, heart, kidneynd spleen. To demonstrate the ability of hamster AHRDNA to encode protein, we used in vitro transcription/ranslation system to make translation products (Fig.). Although the yield was low, the apparent molecular
ntinued
TABLE 1
Identities of AHR Functional Domains
Domain
Hamstervs
SD rat%
Hamstervs
C57 mouse%
Hamstervs
human%
NA binding (9-20, 27-39, 220) 100 100 100LS (13-39) 100 96.3 96.3ES (55-75) 100 95.2 100SP90 binding (27-79, 185-378) 92.3 92.7 87.4imerization (40-79, 120-293) 93.9 92.5 88.8RB binding (;330; LXCXE) 100 100 100ransactivation (493-920) 60.0 52.1 51.2
Note. Numbering is based on the hamster sequence. NLS, nuclearocalization signal; NES, nuclear export signal (35); pRb binding,inding site for the retinoblastoma tumor suppressor protein (20).
Co
mst1s
sefieagswtypj
D
a
exists between two mouse strains, the sensitiveCshAiH1rfispdAHahwem
eotArirttrsciQmvt
R
Vol. 273, No. 1, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
ass of the translated protein (the highest band) wasimilar to the protein recognized by the anti-AHR an-ibody in hamster liver and lung samples being about20 kDa. The AHR in H/W and L-E rats was notablymaller (98 kDa and 106 kDa, respectively).The 59 end of the hamster AHR mRNA exhibited
urprising variation. The expected 59 structure waslicited only in the case that the reverse primer in thenal PCR reaction was selected 59 to the junction ofxon/intron 1 (nucleotides 120/121 in hamster cDNA),s inferred from the organization of the mouse AHRene (13). If the reverse primer site was located down-tream this junction, typical intron structure completeith the invariant 39 AG dinucleotide was obtained 59
o the junction. Both independent methods usedielded the same result, although the number of therobably intron-derived nucleotides preceding theunction differed slightly (28 by ligation, 18 by tailing).
ISCUSSION
There are large inter- and intraspecies differences incute TCDD toxicity. The classical strain difference
FIG. 4. Northern blot analysis of hamster and rat AHR. Poly-A1
NA was isolated from 250 mg of total RNA from liver.
FIG. 5. Tissue distribution of hamster AHR gene expr
278
57BL/6 and the resistant DBA/2 strains, where theensitivity difference is about tenfold. DBA/2 strainas a point mutation in the ligand binding domain ofHR, which results in markedly reduced binding affin-
ty for TCDD (14, 15). The sensitive L-E and resistant/W rats show the largest intraspecies variation (ca.000-fold) in TCDD lethality. Recent cloning of H/Wat AHR cDNA revealed a critical point mutation in therst invariant nucleotide of intron 10 leading to alteredplicing of the mRNA. This gives rise to two differentroteins, both of which bear unique transactivationomains (8). Genetic studies implied that the alteredHR is the major determinant of TCDD resistance in/W rats (9). The largest interspecies difference incute TCDD toxicity exists between guinea pig andamster. Guinea pig is the most sensitive mammalith an LD50 value of ca. 1 mg/kg, while hamster isxtremely resistant with an LD50 value of about 3000g/kg (16, 5).Hamsters and H/W rats share a number of features,
.g., both are extremely resistant to the acute lethalityf TCDD, but sensitive to CYP1A1 induction by TCDD,he mediation of which is the best-known function ofHR. The present findings disclose that similar to H/Wat AHR, hamster also has an altered C-terminal endn its AHR. In H/W rats, mutations lead to two kinds ofeceptor proteins, which have shortened transactiva-ion domains. In hamsters, the Q-rich subunit of theransactivation domain is strikingly larger than inats, mice and humans because of incorporation ofhort DNA repeats. This repetetive sequence containsodons for glutamine in the reading frame and thus itncreases the number of glutamine residues. The-rich region is thought to play an important role inediating transactivation by the AHR (4). In some in
itro studies, it has been shown to exhibit the strongestranscriptional activity of several activation subdo-
ion. Lane 1, male hamster and lane 2, female hamster.
ess
mIQhofDcntctmQttcvps
sbhettpti7wtNnttAHrslt
pgemaecspepaballAm
tTtbboshdeahdmio
A
RooE
Vol. 273, No. 1, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
ains in the C-terminal half of the AHR sequence (17).n other studies, the transactivation potency of the-rich subdomain as such has proved to be low, but itas been able to remarkably augment functions ofther subunits in this domain (18). A completely dif-erent function is exerted in the Kruppel protein ofrosophila, where the Q-rich region is reported to be
rucial for transactivation repression (19). It is alsooteworthy that an important regulator of cell cycle,he retinoblastoma tumor suppressor protein, has re-ently been found to be associated with the AHR, andhe Q-rich subdomain is one of the two sites at which itakes contact with the AHR (20). Interestingly, the-rich structure is lacking in the AHR of some ex-
remely TCDD-sensitive teleost fish such as rainbowrout (21). In contrast, a fish species, Fundulus hetero-litus, which is sensitive to dioxins but capable of de-eloping dioxin resistance after multigenerational ex-osure, has been reported to possess a Q-richubdomain in one of its two AHRs (22).The evidence obtained of alternative splicing of ham-
ter AHR mRNA at its 59 end was totally unexpectedecause it is unprecedented in all other species studieditherto. Since two independent methods relying onntirely different primer structures provided essen-ially the same result, it is unlikely to be due to con-amination with genomic DNA. The more probable ex-lanation is skipping of exon 1 in mRNA processing. Ifhe alternative transcript is translated, the first ATGn frame with the wild-type start codon is at nucleotide61. Usage of this codon as the translational start siteould generate a protein 134 amino acids smaller than
he full-length AHR and devoid of the critical-terminal bHLH structure. However, the neighborucleotides of this codon do not favor efficient transla-ion (23). Furthermore, although hamsters are resis-ant to acute lethality of TCDD, they exhibit the typicalHR-mediated phenomenon, P450 induction (24).amster AHR is also fully able to bind to its cognate
esponse element in DNA (25). Therefore, alternativeplicing of AHR mRNA at the 59 end may be a pecu-iarity specific for hamster but of little, if any, func-ional consequence.
FIG. 6. Western
279
There are differences in molecular weights of AHRrotein among species and also among strains in aiven species. The hamster receptor is one of the larg-st studied (26, 27). Although the calculated molecularass is 103.3 kDa, Western analysis yielded a size of
pproximately 120 kDa, which is in accordance witharlier findings (26). This difference between the cal-ulated size and the mobility of protein has been ob-erved previously for rat, mouse and human AHR andossibly reflects aberrant mobility during SDS–PAGElectrophoresis or post-translational modifications ofroteins (8, 28, 29). The mRNA of hamster AHR waslso larger than that of the rat (;7 kb vs ;6.6 kb)ased on Northern analysis. Provided that we man-ged to clone the full leader sequence, the 39 untrans-ated region constitutes the bulk of the mRNA paral-eling the situation in the rat. The tissue distribution ofHR gene expression resembled that in rats (12, 30),ice (31) and humans (28).The N-terminal end of the hamster AHR containing
he bHLH and the PAS domains is highly conserved.his high degree of homology is not surprising sincehe bHLH domain has been shown to mediate DNAinding and the PAS region is important in ligandinding and AHR/ARNT dimerization (32, 33). It isbvious that the reason for dioxin resistance in ham-ters cannot be found in this region. In contrast, theamster AHR stands out from all other AHRs cloned toate for its C-terminal structure, especially for thexpanded Q-rich subdomain. This could conceivablyccount for the peculiarly selective responsiveness ofamsters to TCDD if one assumes promoter context-ependent interplay between the transactivation do-ains of AHR and ARNT (34). However, further stud-
es are needed to elucidate the functional significancef the structural alteration found.
CKNOWLEDGMENTS
This study was financially supported by grants from the Finnishesearch Program for Environmental Health (SYTTY, the Academyf Finland, Research Council for Health, Grant 42386), the Academyf Finland, Research Council for Health (Grant 43984), and theuropean Commission (Contract QLK4-1999-01446).
analysis of AHR.
blot
REFERENCES
1
1
1
1
1
1
1
dibenzo-p-dioxin (TCDD) in the hamster. Toxicol. Appl. Pharma-
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
Vol. 273, No. 1, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
1. Hoffman, E. C., Reyes, H., Chu, F. F., Sander, F., Conley, L. H.,Brooks, B. A., and Hankinson, O. (1991) Cloning of a factorrequired for activity of the Ah (dioxin) receptor. Science 252,954–958.
2. Rowlands, J., and Gustafsson, J.-Å. (1997) Aryl hydrocarbonreceptor-mediated signal transduction. CRC Crit. Rev. Toxicol.27, 109–134.
3. Fukunaga, B. N., Probst, M. R., Reisz-Porszasz, S., and Hankin-son, O. (1995) Identification of functional domains of the arylhydrocarbon receptor. J. Biol. Chem. 270, 29270–29278.
4. Jain, S., Dolwick, K. M., Schmidt, J. V., and Bradfield, C. A.(1994) Potent transactivation domains of the Ah receptor and theAh receptor nuclear translocator map to their carboxyl termini.J. Biol. Chem. 269, 31518–31523.
5. Henck, J. M., New, M. A., Kociba, R. J., and Rao, K. S. (1981)2,3,7,8-Tetrachlorodibenzo-p-dioxin: acute oral toxicity in ham-sters. Toxicol. Appl. Pharmacol. 59, 405–407.
6. Pohjanvirta, R., Unkila, M., and Tuomisto, J. (1993) Compara-tive acute lethality of 2,3,7,8-tetrachlorodibenzo-p-dioxin(TCDD), 1,2,3,7,8-pentachlorodibenzo-p-dioxin and 1,2,3,4,7,8-hexachlorodibenzo-p-dioxin in the most TCDD-susceptible andthe most TCDD-resistant rat strain. Pharmacol. Toxicol. 73,52–56.
7. Pohjanvirta, R., and Tuomisto, J. (1994) Short-term toxicity of2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in laboratory ani-mals: Effects, mechanisms, and animal models. Pharmacol. Rev.46, 483–549.
8. Pohjanvirta, R., Wong, J. M. Y., Li, W., Harper, P. A., Tuomisto,J., and Okey, A. B. (1998) Point mutation in intron structurecauses altered carboxyl-terminal structure in the aryl hydrocar-bon receptor of the most 2,3,7,8-tetrachlorodibenzo-p-dioxin-resistant rat strain. Mol. Pharmacol. 54, 86–93.
9. Tuomisto, J. T., Viluksela, M., Pohjanvirta, R., and Tuomisto, J.(1999) The Ah receptor and a novel gene determine acute toxicresponses to TCDD: Segregation of the resistant alleles to differ-ent rat lines. Toxicol. Appl. Pharmacol. 155, 71–81.
0. Chomczynski, P. (1993) A reagent for the single-step simulta-neous isolation of RNA, DNA and proteins from cell and tissuesamples. Biotechniques 15, 532–537.
1. Dumas, J. B., Edwards, M., Delort, J., and Mallet, J. (1991)Oligodeoxyribonucleotide ligation to single-stranded cDNAs: Anew tool for cloning 59 ends of mRNAs and for constructingcDNA libraries by in vitro amplification. Nucleic Acids Res. 19,5227–5232.
2. Elferink, C. J., and Whitlock, J. P., Jr. (1994) Dioxin-dependent,DNA sequence-specific binding of a multiprotein complex con-taining the Ah receptor. Receptor 4, 157–173.
3. Schmidt, J. V., Carver, L. A., and Bradfield, C. A. (1993) Molec-ular characterization of the murine Ahr gene. Organization,promoter analysis, and chromosomal assignment. J. Biol. Chem.268, 22203–22209.
4. Ema, M., Ohe, N., Suzuki, M., Mimura, J., Sogawa, K., Ikawa, S.,and Fujii-Kuriyama, Y. (1994) Dioxin binding activities of poly-morfic forms of mouse and human aryl hydrocarbon receptors.J. Biol. Chem. 269, 27337–27343.
5. Poland, A., Palen, D., and Glover, E. (1994) Analysis of the fouralleles of the murine aryl hydrocarbon receptor. Mol. Pharmacol.46, 915–921.
6. Olson, J. R., Gasiewicz, T. A., and Neal, R. A. (1980) Tissuedistribution, excretion, and metabolism of 2,3,7,8-tetrachloro-
280
col. 56, 78–85.7. Sogawa, K., Iwabuchi, K., Abe, H., and Fujii-Kuriyama, Y. (1995)
Transcriptional activation domains of the Ah receptor and Ahreceptor nuclear translocator. J. Cancer Res. Clin. Oncol. 121,612–620.
8. Ma, Q., Dong, L., and Whitlock, J. P., Jr. (1995) Transcriptionalactivation by the mouse Ah receptor. Interplay between multiplestimulatory and inhibitory functions. J. Biol. Chem. 270, 12697–12703.
9. Licht, J. D., Hanna-Rose, W., Reddy, J. C., English, M. A., Ro,M., Shaknovich, R., and Hansen, U. (1994) Mapping and mu-tagenesis of the amino-terminal transcription repression domainof the Drosophila Kruppel protein. Mol. Cell. Biol. 14, 4057–4066.
0. Ge, N.-L., and Elferink, C. J. (1998) A direct interaction betweenthe aryl hydrocarbon receptor and retinoblastoma protein.J. Biol. Chem. 273, 22708–22713.
1. Abnet, C. C., Tanguay, R. L., Hahn, M. E., Heideman, W., andPeterson, R. E. (1999) Two forms of aryl hydrocarbon receptortype 2 in rainbow trout (Oncorhynchus mykiss). J. Biol. Chem.274, 15159–15166.
2. Karchner, S. I., Powell, W. H., and Hahn, M. E. (1999) Identifi-cation and functional characterization of two divergent aryl hy-drocarbon receptors (AHR1 and AHR2) in the teleost Fundulusheteroclitus. J. Biol. Chem. 274, 33814–33824.
3. Kozak, M. (1991) Structural features in eukaryotic mRNAs thatmodulate the initiation of translation. J. Biol. Chem. 266,19867–19870.
4. Henry, E. C., and Gasiewicz, T. A. (1987) Changes in thyroidhormones and thyroxine glucuronidation in hamsters comparedwith rats following treatment with 2,3,7,8-tetrachlorodibenzo-p-dioxin. Toxicol. Appl. Pharmacol. 89, 165–174.
5. Bank, P. A., Yao, E. F., Phelps, C. L., Harper, P. A., and Denison,M. S. (1992) Species-specific binding of transformed Ah receptorto a dioxin responsive transcriptional enhancer. Eur. J. Phar-macol. 228, 85–94.
6. Poland, A., and Glover, E. (1987) Variation in the molecularmass of the Ah receptor among vertebrate spacies and strains ofrats. Biochem. Biophys. Res. Commun. 146, 1439–1449.
7. Poland, A., Glover, E., and Bradfield, C. A. (1990) Characteriza-tion of polyclonal antibodies to the Ah receptor prepared byimmunization with a synthetic peptide hapten. Mol. Pharmacol.38, 20–26.
8. Dolwick, K. M., Schmidt, J. V., Carver, L. A., Swanson, H. I., andBradfield, C. A. (1993) Cloning and expression of a human Ahreceptor cDNA. Mol. Pharmacol. 44, 911–917.
9. Burbach, K. M., Poland, A., and Bradfield, C. A. (1992) Clon-ing of the Ah-receptor cDNA reveals a distinctive ligand-activated transcription factor. Proc. Natl. Acad. Sci. USA 89,8185– 8189.
0. Carver, L. A., Hogenesch, J. B., and Bradfield, C. A. (1994)Tissue specific expression of the rat Ah-receptor and ARNTmRNAs. Nucleic Acids Res. 22, 3038–3044.
1. Li, W., Donat, S., Dohr, O., Unfried, K., and Abel, J. (1994) Ahreceptor in different tissues of C57BL/6J and DBA/2J mice: Useof competitive polymerase chain reaction to measure AH-receptor mRNA expression. Arch. Biochem. Biophys. 315, 279–284.
2. Dolwick, K. M., Swanson, H. I., and Bradfield, C. A. (1993) Invitro analysis of Ah receptor domains involved in ligand-activated DNA recognition. Proc. Natl. Acad. Sci. USA 90, 8566–8570.
3. Whitelaw, M. L., Gottlicher, M., Gustafsson, J.-Å., and Poel-

linger, L. (1993) Definition of a novel ligand binding domain of a
3
dioxin receptor and its basic helix-loop-helix/PAS partner factor
3
Vol. 273, No. 1, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
nuclear bHLH receptor: Co-localization of ligand and hsp90binding activities within the regulable inactivation domain ofthe dioxin receptor. EMBO J. 12, 4169–4179.
4. Whitelaw, M. L., Gustafsson, J.-Å., and Poellinger, L. (1994)Identification of transactivation and repression functions of the
281
ARNT: Inducible versus constitutive modes of regulation. Mol.Cell. Biol. 14, 8343–8355.
5. Ikuta, T., Eguchi, H., Tachibana, T., Yoneda, Y., and Kawajiri,K. (1998) Nuclear localization and export signals of the humanaryl hydrocarbon receptor. J. Biol. Chem. 273, 2895–2904.





























