Embed Size (px)
Citation preview

1
Quantifying Crosstalk in Biochemical SystemsEnoch Yeung⇤, Jongmin Kim†, Ye Yuan�, Jorge Goncalves�, and Richard M. Murray⇤,†
Abstract— Recent work has introduced biocircuit architec-
tures that exhibit robust oscillatory behavior in organisms rang-
ing from cyanobacteria to mammals. Complementary research
in synthetic biology has introduced oscillators in vivo and in
vitro suggesting that robust oscillation can be recapitulated
using a small number of biochemical components. In this
work, we introduce signaling crosstalk in biocircuits as a
consequence of enzyme-mediated biochemical reactions. As a
motivating example, we consider an in vitro oscillator with
two types of crosstalk: crosstalk in production and degradation
of RNA signals. We then pose a framework for quantifying
crosstalk and use it to derive several dynamical constraints
and suggest design techniques for ameliorating crosstalk in in
vitro biochemical systems. As an example, we show that the
balance between production and degradation crosstalk plays a
key role in determining system stability, potentially leading to
loss of oscillatory behavior. We demonstrate that the effects of
crosstalk can attenuated through the effective tuning of two key
parameters in order to recover desired system dynamics.
I. INTRODUCTION
High-throughput technologies have led to the era ofsystems biology as an integrative approach to connectmolecular-level mechanisms to cell-level behavior [1]. Whilethe systems biology approach generated complex wiringdiagrams for analyzing biological systems on a large scale,a complementary approach in synthetic biology focused oncreating networks to elucidate design principles to achievedesired functions [2]. However, it has often been over-looked that biological systems are under continuous selec-tion pressure to maximize performance while minimizingcost, i.e., maintaining only the minimal amount of criticalmachinery [3], [4]. Consequently, there exists a potential tooverload important pathways during times of stress and toplace unanticipated burden on central cellular enzymes [5],[6]. Therefore, it is critical to have a detailed understandingof the underlying molecular mechanisms that capture theloading effects on different components — indeed, suchunderstanding is a necessary first step in further developmentof systems and synthetic biology.
Within the context of synthetic biology, a biocircuit can beconceptually divided into two (intertwined) layers — a layerthat constitutes molecular architecture reflecting designedinteractions and a layer that consists of uncertain or hiddeninteractions, referred to as crosstalk, arising from eitherbackground processes or byproducts of circuit processes.This crosstalk layer is not easily avoided and often appears
For correspondence, please contact [email protected] [email protected]. Affiliations: ⇤ Department of Controland Dynamical Systems, † Department of Bioengineering Departmentat California Institute of Technology; � Control Group, Department ofEngineering, University of Cambridge, Cambridge, UK
fundamental to circuit operation [7], ways have been sug-gested to overcome crosstalk in the form of retroactivity [8].
In engineering, crosstalk is traditionally viewed as theinteraction between two signal relays. Old electronic cir-cuits suffered various problematic effects such as ringing,crosstalk, and noise. Thus, essentially all modern circuitsmust be designed with signal integrity in mind [9]. However,in biology, crosstalk seems to be a ubiquitous feature takingthe form of, e.g., hidden reactions [10], non-specific bind-ing [11], waste management [12], and enzyme loading [5]possibly with different types of impact on system dynamics.In this work, we consider crosstalk arising from enzymeloading, i.e., the saturation effects that occur from substratescompeting for the same enzyme.
We organize this work as follows: In Section II weconsider the simplified oscillator model for a circadian clockproposed by Barkai and Leibler which is robust to stochasticnoise in simulation studies [13], [14] — unlike other cir-cadian clock models [15]. We introduce an in vitro modelreconstructing the Barkai-Leibler oscillator and analyze theimpact of the saturation effects of competing substrates forproduction and degradation enzymes. We show how thesecompetitive effects lead to undesired interactions betweensystem components — a dynamical effect we call crosstalk.In Section III we introduce the mathematical backgroundfor studying crosstalk in biochemical systems and propose ameasure for quantifying crosstalk between components andfor the system as a whole. Finally, in Section IV we introduceconceptual approaches towards designing systems that arerobust to crosstalk. In particular, we draw conclusions onthe impact of crosstalk on system stability, individual statetrajectories and weak competition between states. Further,we discuss two ways to ameliorate crosstalk in the contextof robust biochemical system design and illustrate how thesetechniques can be used to recover oscillations in the Barkai-Liebler oscillator.
II. MOTIVATING EXAMPLE
A. In Vitro Barkai-Liebler Oscillator
The model for circadian clock proposed by Barkai andLeibler take the form of Figure 1 inset [13], [14]. It in-volves two genes, DA and DR, coding for an activatorA and a repressor R, which are transcribed into mRNAand subsequently translated into protein. The activator Abinds to the promoters for genes DA and DR to increasetheir transcription rate, while R acts as the negative elementby sequestering the activator A. It is noteworthy that theoscillation does not rely on a cooperative binding of A tothe promoters of genes or cooperative binding of R to A; the
Submitted, 2012 Conference on Decision and Control (CDC)http://www.cds.caltech.edu/~murray/papers/yeu+12-cdc.html

2
ON
AD
D
OFF
ON
AD
D
OFF
RNAP
R
D RD A
A
A
RA
R
A
RNAP
RNaseHRNaseH
R
RNaseR
RNaseR
RNaseR
C
Fig. 1. A schematic diagram of the Barkai-Liebler oscillator (top left)and the detailed molecular reactions for in vitro genelet oscillator. Thereare two synthetic templates, DA and DR, producing an activator A anda repressor R using RNAP. The activator A binds to the promoter sitesof the two templates to enhance transcription rates, while the repressor Rsequesters A into a complex C. The RNA signals A, R, and C are degradedselectively by RNase R, while A bound to DNA templates are degraded byRNase H. The sequence domains are color-coded to indicate identical orcomplementary sequences.
activation mechanism is a simple binding and the repressionmechanism is a stochiometric sequestering. On the otherhand, a key requirement for oscillation was the time-scaleseparation for A and R dynamics — the production anddegradation of A are on a much faster time scale than thoseof R.
Here, we propose a model that captures the essentialfeatures of Barkai-Leibler oscillator in a simplified in vitrosetting (Figure 1). We first introduce a synthetic in vitroswitch design and lay groundwork for general circuit con-struction methods. The OFF state of the switch consistsof a double-stranded (ds) DNA template (“D’’) with apartially single-stranded (ss) and thus incomplete T7 RNApolymerase (RNAP) promoter region. The switch is turnedON by the binding of a ssDNA or ssRNA activator (“A’’)that completes the RNAP promoter region. The resultingtemplate (“DA’’) has a nicked promoter but still transcribeswell [12]. (Experimental characterization of genelets usedDNA activator [12]; RNA activator would likely be lessefficient than its DNA counterpart [16].) To provide a sharpthreshold of activation, an inhibitor strand (ssDNA or ssRNA,“I’’) can bind to a complementary free-floating activator A,resulting in a functionally inert activator-inhibitor complex“AI.’’ (This reaction is not explicitly implemented for thecircuit presented in this paper.) Using these design motifs forswitches and signals, networks with arbitrary connectivitycan be constructed modularly, in principle, as continuous-time analog neural networks [17]. In a typical reactionnetwork, RNA outputs will be produced by RNAP fromupstream switches using NTP as fuel, and these outputswill serve as inputs for downstream switches. At the sametime, the degradation of RNA signals by Escherichia coliribonuclease H (RNase H) and/or ribonuclease R (RNase R)
removes RNA signal, thereby undoing the regulatory effectof the RNA inputs.
The synthetic in vitro transcrptional circuits (‘genelets’)have been used to create bistable circuits, oscillators, anddrive other molecular machines using modular architecturewith programmable connectivity [12], [18], [19]. Thus, itwould be possible to experimentally understand and char-acterize elementary reactions and system dynamics for ourproposed circuit in the near future. A notable difference fromthe models of Barkai and Leiber is that, for our proposedbiochemical circuit, the RNA signals act directly as theactivator and repressor of the system, obviating the needfor translation machinery. The deterministic dynamics of themodel is given by the set of reaction rate equations below.
DOFFA = �kONADOFF
A + kOFFDONA (1)
DOFFR = �kONADOFF
R + kOFFDONR
A = kp,ADONA + ↵kp,AD
OFFA � kONADOFF
A
�kONADOFFR � kANNAR� �AA
R = kp,RDONR + ↵kp,RD
OFFR � kANNAR
+�AC � �RR
C = kANNAR� �AC
where DA and DR are synthetic DNA templates coding RNAsignals A and R, while C is the complex of A and R. Thetemplates DA and DR have incomplete promoter regions forRNAP such that the transcription rates are slow. When theRNA signal A binds and completes the incomplete promoterregion, the transcription rate increases. Therefore, DON
A isthe ON-state switch, i.e., DAA complex, while DOFF
A issimply DA.
To effectively achieve time-scale separation for the dy-namics of A and R, kp,A and �A should be much largerthan kp,R and �R, respectively. To adjust production rates,one possibility is tuning the catalytic rate of RNAP fortemplates by changing the consensus sequence right afterthe promoter region; a simpler solution would be usinghigh concentration of templates coding for A. To adjustdegradation rates, we can utilize the property of E. coliRNase R that can by itself degrade single-stranded RNAswith extensive secondary structure provided that a single-stranded 30 overhang more than 4-base long is present [20].By incorporating extensive secondary structures for 30 end ofR while leaving 30 end of A structure-free, we can achievethe desired differential degradation for A and R. We observedoscillation with experimentally plausible parameter values(Figure 2).
To gain insight into the essential elements required foroscillations, we simplified the deterministic rate equationsby making quasi-steady-state assumptions for templates DA,DR, and activator A. The resulting two-variable system withthe repressor R and the complex C as the two slow variables:
R = kp,RDtotR
↵koff + konA(R)
koff + konA(R)
�kANN A(R)R+ �AC � �RR
C = kANN A(R)R� �AC
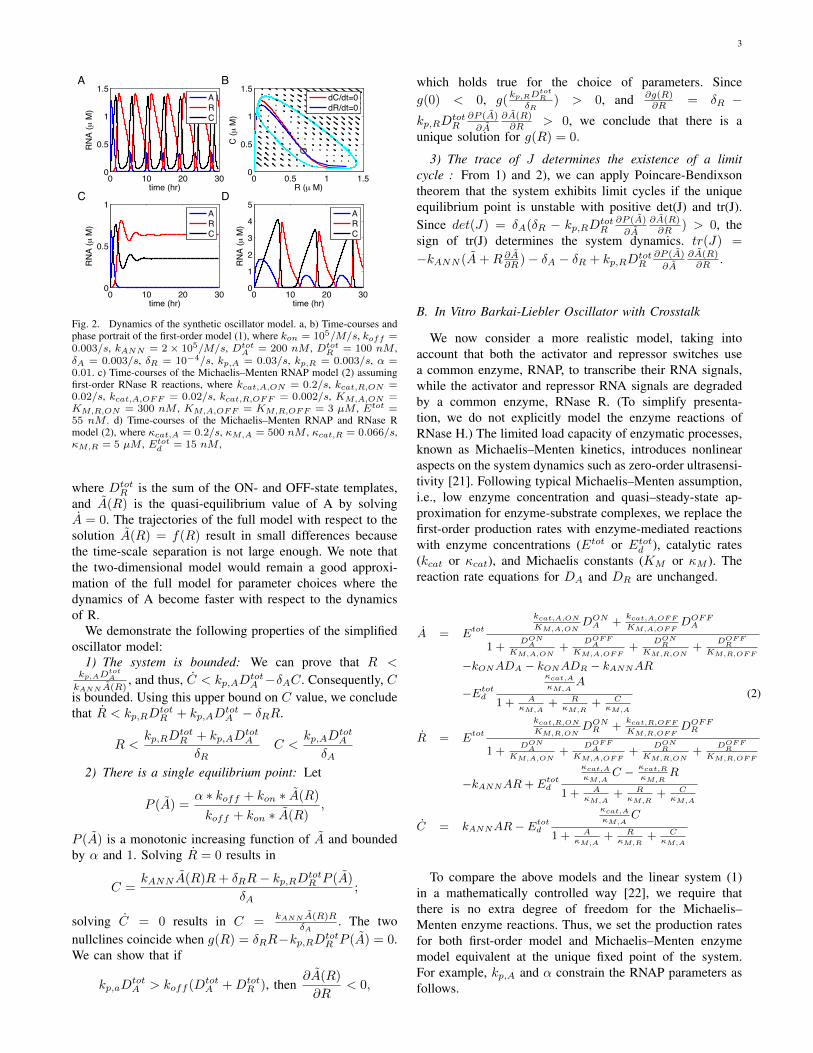
3
0 0.5 1 1.50
0.5
1
1.5
R (µ M)
C (µ
M)
B
dC/dt=0dR/dt=0
0 10 20 300
0.5
1
1.5
time (hr)
RN
A (µ
M)
A
ARC
0 10 20 300
0.5
1
time (hr)
RN
A (µ
M)
C
ARC
0 10 20 300
1
2
3
4
5
time (hr)
RN
A (µ
M)
D
ARC
Fig. 2. Dynamics of the synthetic oscillator model. a, b) Time-courses andphase portrait of the first-order model (1), where kon = 105/M/s, koff =0.003/s, kANN = 2 ⇥ 105/M/s, Dtot
A = 200 nM , DtotR = 100 nM ,
�A = 0.003/s, �R = 10�4/s, kp,A = 0.03/s, kp,R = 0.003/s, ↵ =0.01. c) Time-courses of the Michaelis–Menten RNAP model (2) assumingfirst-order RNase R reactions, where kcat,A,ON = 0.2/s, kcat,R,ON =0.02/s, kcat,A,OFF = 0.02/s, kcat,R,OFF = 0.002/s, KM,A,ON =KM,R,ON = 300 nM , KM,A,OFF = KM,R,OFF = 3 µM , Etot =55 nM . d) Time-courses of the Michaelis–Menten RNAP and RNase Rmodel (2), where cat,A = 0.2/s, M,A = 500 nM , cat,R = 0.066/s,M,R = 5 µM , Etot
d = 15 nM ,
where DtotR is the sum of the ON- and OFF-state templates,
and A(R) is the quasi-equilibrium value of A by solvingA = 0. The trajectories of the full model with respect to thesolution A(R) = f(R) result in small differences becausethe time-scale separation is not large enough. We note thatthe two-dimensional model would remain a good approxi-mation of the full model for parameter choices where thedynamics of A become faster with respect to the dynamicsof R.
We demonstrate the following properties of the simplifiedoscillator model:
1) The system is bounded: We can prove that R <kp,ADtot
A
kANN A(R), and thus, C < kp,AD
totA ��AC. Consequently, C
is bounded. Using this upper bound on C value, we concludethat R < kp,RD
totR + kp,AD
totA � �RR.
R <kp,RD
totR + kp,AD
totA
�RC <
kp,ADtotA
�A
2) There is a single equilibrium point: Let
P (A) =↵ ⇤ koff + kon ⇤ A(R)
koff + kon ⇤ A(R),
P (A) is a monotonic increasing function of A and boundedby ↵ and 1. Solving R = 0 results in
C =kANN A(R)R+ �RR� kp,RD
totR P (A)
�A;
solving C = 0 results in C = kANN A(R)R�A
. The twonullclines coincide when g(R) = �RR�kp,RD
totR P (A) = 0.
We can show that if
kp,aDtotA > koff (D
totA +Dtot
R ), then@A(R)
@R< 0,
which holds true for the choice of parameters. Sinceg(0) < 0, g(kp,RDtot
R
�R) > 0, and @g(R)
@R = �R �kp,RD
totR
@P (A)
@A
@A(R)@R > 0, we conclude that there is a
unique solution for g(R) = 0.
3) The trace of J determines the existence of a limitcycle : From 1) and 2), we can apply Poincare-Bendixsontheorem that the system exhibits limit cycles if the uniqueequilibrium point is unstable with positive det(J) and tr(J).Since det(J) = �A(�R � kp,RD
totR
@P (A)
@A
@A(R)@R ) > 0, the
sign of tr(J) determines the system dynamics. tr(J) =
�kANN (A+R @A@R )� �A � �R + kp,RD
totR
@P (A)
@A
@A(R)@R .
B. In Vitro Barkai-Liebler Oscillator with Crosstalk
We now consider a more realistic model, taking intoaccount that both the activator and repressor switches usea common enzyme, RNAP, to transcribe their RNA signals,while the activator and repressor RNA signals are degradedby a common enzyme, RNase R. (To simplify presenta-tion, we do not explicitly model the enzyme reactions ofRNase H.) The limited load capacity of enzymatic processes,known as Michaelis–Menten kinetics, introduces nonlinearaspects on the system dynamics such as zero-order ultrasensi-tivity [21]. Following typical Michaelis–Menten assumption,i.e., low enzyme concentration and quasi–steady-state ap-proximation for enzyme-substrate complexes, we replace thefirst-order production rates with enzyme-mediated reactionswith enzyme concentrations (Etot or Etot
d ), catalytic rates(kcat or cat), and Michaelis constants (KM or M ). Thereaction rate equations for DA and DR are unchanged.
A = E
tot
kcat,A,ON
KM,A,OND
ONA +
kcat,A,OFF
KM,A,OFFD
OFFA
1 +DON
AKM,A,ON
+DOFF
AKM,A,OFF
+DON
RKM,R,ON
+DOFF
RKM,R,OFF
�kONADA � kONADR � kANNAR
�E
totd
cat,A
M,AA
1 + AM,A
+ RM,R
+ CM,A
(2)
R = E
tot
kcat,R,ON
KM,R,OND
ONR +
kcat,R,OFF
KM,R,OFFD
OFFR
1 +DON
AKM,A,ON
+DOFF
AKM,A,OFF
+DON
RKM,R,ON
+DOFF
RKM,R,OFF
�kANNAR+ E
totd
cat,A
M,AC � cat,R
M,RR
1 + AM,A
+ RM,R
+ CM,A
C = kANNAR� E
totd
cat,A
M,AC
1 + AM,A
+ RM,R
+ CM,A
To compare the above models and the linear system (1)in a mathematically controlled way [22], we require thatthere is no extra degree of freedom for the Michaelis–Menten enzyme reactions. Thus, we set the production ratesfor both first-order model and Michaelis–Menten enzymemodel equivalent at the unique fixed point of the system.For example, kp,A and ↵ constrain the RNAP parameters asfollows.

4
kp,A =
kcat,A,ON
KM,A,ONE
tot
1 +DON
AKM,A,ON
+DOFF
AKM,A,OFF
+DON
RKM,R,ON
+DOFF
RKM,R,OFF
↵ =kcat,A,OFF
KM,A,OFF/
kcat,A,ON
KM,A,ON
For the Michaelis–Menten RNAP reaction equations, wecan follow the model reduction procedure for the first-ordermodel because A is still a function of R only. On the otherhand, the Michaelis–Menten RNase R reaction equationsare functions of A, R, and C, making the model reductionapproach difficult. We focused our attention to numericallyprobing system behavior as the saturation levels for RNAPand RNase change. Using the above constraints, the locationof equilibrium point and molecular flux (production anddegradation rates) at the equilibrium point remain identi-cal for different enzyme parameter choices. The moderatesaturation of RNAP significantly deteriorated the oscillation(Figure 2c), while the moderate saturation of RNase Rcould recover the oscillation albeit at slower time-scales(Figure 2d). If the RNase reactions are completely saturated,the system would accumulate RNA signals without bound.
III. TOWARDS QUANTIFYING CROSSTALK
Crosstalk arises from species competing for commonlyshared enzymes. Enzymes drive two types of reactions inbiochemical systems, reactions involving production anddegradation. On the other hand, there are other reactionsthat do not employ enzymes that can be viewed as ’inter-action’ reactions, i.e. a reaction involving components of thebiochemical system interacting in accordance with designprinciples and system function. We decompose the dynamicsof the system accordingly:
x = fc(x) = Pc(x) + I(x)�Dc(x), x(0) = x0 (3)
where x 2 Rn , Pc(x) (Dc(x)) describes the rates ofproduction (degradation) from enzyme-mediated reactionsand I(x) describes the dynamics of interaction reactions.We will suppose (3) satisfies the Michaelis Menten as-sumption, namely all enzyme-substrate complexes are inchemical equilibrium. If xi is not produced (degraded) byan enzyme-mediated reaction, then the corresponding entryin Pc,i(x) (Dc,i(x)) is 0. For notational convenience, wewill suppose that there are np substrate species indexedas x1, ..., xns that combine with enzymes to produce np
products: x↵(1), ...., x↵(np). For example, if x↵(i) is producedby substrates x1, ..., xnk
in competition with other enzyme-sharing substrates x1, ..., xnc
, we will suppose that Pc,↵(i)(x)can be written
Pc,↵(i)(x) = EtotnkX
j=1
kcat,jgj(x1, ..., xnc) (4)
⌘ EtotnkX
j=1
kcat,j
xj
KM,j
1 +Pnc
l=1xl
KM,l
Similarly, we let xd(1), ..., xd(nd) denote the list of systemstates that are degraded commonly by one or more enzymes
and suppose that for each xi degraded by an enzyme sharedby competing substrates xd(1), ..., xd(nc) , Dc,i(x) can bewritten as
Dc,i(x) = Etotd cat,iui(xd(1), ..., xd(nc)) (5)
⌘ Etotd cat,i
xi
M,i
1 +Pd(nc)
l=d(1)xl
M,l
(6)
The processes of production and degradation rate functionsare fundamental to system function: without productionreactions, a system cannot synthesize the various compo-nents required to achieve intended dynamics and withoutdegradation, a system quickly saturates and in certain casestends towards instability. Nonetheless, any production ordegradation reaction involving competing substrates will pro-duce crosstalk effects. Thus, crosstalk arising from enzyme-mediated reactions is a fundamental feature of any suffi-ciently complex biochemical system. To quantify crosstalk insuch systems, we need to compare the dynamics of system(3) against the dynamics of a reference system that is freeof crosstalk. Such a system, that reflects the complexityand function of the original system may be impossible torealize physically. However, there are simple systems wherea production (or degradation) enzyme may uniquely bind toa single substrate. Though these systems are trivial in design(and perhaps functionality), they provide a motivation fordefining the following conceptual crosstalk-free system :
Definition 1: [Alternative System] Given the system (3)with equilibrium point xe, define the alternative productionfunction P (x) : Rn !2 Rn, where for each nonzero entryP↵(i)(x), i = 1, ..., np, if x↵(i) is produced from substratesx1, ...., xnk
2 {x1, ..., xns} in system (3), then P↵(i)(x) is
defined as
P↵(i)(x) ⌘ EtotnkX
j=1
kcat,jhj(xj) (7)
⌘ EtotnkX
j=1
kcat,j
xj
KM,j
1 + xj
KM,j
. (8)
Implicitly, we posit the presence of E1, ..., Ensenzymes
that combine respectively with x1, ..., xns, with identical
concentration totals Etot1 = . . . = Etot
ns= Etot and
identical Michaelis Menten constants. Similarly, we defineD(x) : Rn !2 Rn to be a vector of functions describingthe degradation dynamics for nd species each producedby nd different enzymes. If xd(1), ..., xd(nc) are substratescompeting for degradation enzyme Ed then we write thedegradation rate of xd(i) as
Dc,d(i)(x) ⌘ Etotd cat,ivi(xd(i)) (9)
⌘ Etotd cat,i
xi
M,i
1 + xi
M,i
(10)
We then define the alternative system to be
x = fa(x) = P (x) + I(x)�D(x), x(0) = x0 (11)The alternative system is constructed to function with the
same interaction reactions as the original system and thus,

5
sequestration, activation and inhibition, or branch displace-ment reactions are all conserved. Wherever production ordegradation of a component of the system is concerned,however, the dynamics of the alternative system are modifiedto remove competition effects between substrates. Note thatthe effects of nonlinear saturation are still retained throughthe use of Michaelis Menten functions.
Remark 1: The alternative system is a conceptual refer-ence system to compare against the crosstalk system (3).For the comparison to be fair, it is important that (11) satisfyinternal equivalence [22], namely that we hold fixed any dy-namics or parameters unassociated with crosstalk, e.g. I(x)or x(0). The remaining degrees of freedom are the functionalforms of P (x) and D(x) and their respective parameters.Following the process in [22], we impose constraints onthe external behavior to achieve external equivalence of thealternative system: 1) the enzymes satisfy the MichaelisMenten assumption and 2) in the absence of competingspecies, the production (degradation) rates of Pc,↵(i)(x) andP↵(i)(x) (Dc,d(i)(x) and Dd(i)(x)) coincide. We can thinkof P↵(i)(x) as the limit of Pc,↵(i)(x) when exactly oneof x1, ..., xs are nonzero. These constraints imply that theMichaelis Menten and enzyme parameters for P↵(i)(x) andPc,↵(i)(x) are identical. Thus, as we compare the behavior ofboth systems, any differences in production or degradationrates reflect only the effects of crosstalk.
With the definition of an alternative system in place, wecan now quantify the amount of crosstalk that occurs betweena pair of species in system (3).
Definition 2 (Pairwise Crosstalk): Let xi and xj be twodistinct species in the biochemical system (3). ExpandingPc,i(x) as in equation (4), we define the directed pairwiseproduction crosstalk of xj on xi as
cp(xi, xj) =
(Pi(x)� Pc,i(x), xj 2 {x1, ..., xnc
}0, otherwise.
(12)
Expanding Dc,i(x) as in equation (5), we define the directedpairwise degradation crosstalk of xj on xi as
cd(xi, xj) =
(Di(x)�Dc,i(x), xj 2 {x1, ..., xnc}0, otherwise.
(13)
We define the undirected pairwise production crosstalkbetween xi and xj as Cp(xi, xj) ⌘ cp(xi, xj) +cp(xj , xi) and the undirected pairwise degradation crosstalkas Cd(xi, xj) ⌘ cd(xi, xj) + cd(xj , xi). We refer toC(xi, xj) ⌘ Cp(xi, xj) + Cd(xi, xj) as the pairwisecompetitive crosstalk between xi and xj .
Remark 2: By construction, Cp(xi, xj) � 0 andCp(xi, xj) = Cp(xj , xi). Cp does not, in general, definea metric. However, if we restrict the domain of Cp to a P-class, defined as a set consisting of a single product statex↵(i) and substrates x1, ..., xnc competing for the enzymethat produces x↵(i), then it follows immediately from thedefinitions that Cp is a pseudometric.
Similarly, if we consider any set of species D degradedby a single enzyme, Cd(xi, xj) then defines a metric on theassociated discrete state space D = Vs of degradable species.
In particular, the 2-tuple (D, Cd) is a metric space and if fis a contraction mapping on (X , Cd), then the contractionmapping theorem applies.
Remark 3: If K is the number of distinct enzymes in thesystem and x1 competes for enzyme E1, x2 for E2,..., xK forEK , then by Definition 2, defining �p(x) = Pc(x) � P (x)and �d(x) = Dc(x)�D(x) we get
fc(x)� fa(x) = Pc(x)�Dc(x)� P (x) +D(x)
= �p(x)��d(x)
=KX
k=1
�cp(x, xk) + cd(x, xk),
where cp(x, xk), cd(x, xk) is the vector of associated pair-wise crosstalk terms. This result shows that the productioncrosstalk and degradation crosstalk vectors can be recoveredthrough an appropriate collection of states that represen-tatively span the set of enzymes for which competitivecrosstalk occurs.
Definition 3 (System Crosstalk): The total systemcrosstalk for system (3), denoted F is defined asCF ⌘
Pi<j C(xi, xj)
IV. CROSSTALK AND ROBUST DESIGN
Since crosstalk is fundamental to system function, it isimportant to understand how to design a system in a waythat reduces crosstalk. Intuitively, if we want productionof a particular state to be reliable, we should impose theconstraint that the populations of all other competing speciesare relatively small in comparison with the population of thestate we wish to produce. On the other hand, if we wanta state is to follow a reference trajectory, it is importantthat we understand how the relative (dynamic) populationsof each species drive the crosstalk that our target stateexperiences. In this section, we present two types of results:criteria for rendering the effects of crosstalk as negligibleby imposing constraints on the dynamics of the systemand criteria for attenuating crosstalk by effectively alteringsystem parameters.
A. Dynamic Constraints
At times it is important that a biochemical system maintainan asymptotically stable equilibrium point. For example,we may want to linearize the nonlinear model to applyresults from linear system theory, see for instance [23]—the alternative system is nice in this regard, as the absenceof competitive substrates in its production and degradationfunctions leads to a simpler linearization. Naturally, wewould want system (3) to have the same equilibrium point.
Proposition 1 (Robust-to-Crosstalk Stability): If the alter-native system (11) has an asymptotically stable equilibriumpoint xe in the positive orthant characterized by a Lyapunovfunction satisfying dV
dx > 0 for all x on the positiveorthant, then the system (3) with crosstalk maintains the sameasymptotically stable equilibrium point xe if �p(x+ xe) <�d(x+ xe) for all x in the positive orthant.

6
Proof: The result is a straightforward application ofLyapunov’s stability theorem.The key condition for ensuring V (x) is a Lyapunov functionfor both systems (11) and (3) is that the inner productdVdx (�p(x+xe)+�d(x+xe) < 0. If we restrict our attentionto the class of Lyapunov functions with dV
dx > 0, e.g. V (x) =Pi xi, then we obtain a particular design constraint on the
crosstalk terms. Recall that �d(x) = Dc(x) � D(x) 0and �p(x) = Pc(x)� P (x) 0. Intuitively, if we want theequilibrium point to be unaltered when we add crosstalk inthe system, then the amount of degradation crosstalk shouldbe less than the production crosstalk so that degradationprocesses drive the system towards equilibrium unhampered,while production processes are attenuated by productioncrosstalk and degradation.
Our next result shows that by effectively bounding com-peting species, we can make the dynamics of a particularstate robust.
Proposition 2 (Designing robust x↵(i)): Suppose x↵(i) isproduced by a single substrate xi in competition withx1, ..., xi�1, xi+1, ..., xnc
for enzyme E. Further, supposex↵(i) is degraded by enzyme Ed, in competition withxd(1), ..., xd(nd). If for a given x0, there exists ⇢ > 0 and� > 0 such that the solution of the crosstalk system (3),xc(t), lies in both the set
Si⇢ = {x 2 Rn
>0|ncX
j=1,j 6=i
xj
KM,j⇡ ⇢} (14)
and the set
Si� = {x 2 Rn
>0|d(nd)X
j=d(n1),j 6=↵(i)
xj
M,j⇡ �} (15)
and xi(t) � ⇢, � for all t � 0, then the magnitude of thecrosstalk
|Cp(x↵(i), xl) + Cd(x↵(i),xq)| ⇡ O(1
xi),
for any l 6= i and q 6= i, and for any interval of time [t, t+ ⌧ ]
|xi,c � xi,a| ⇡ O(⌧
xi,a),
where xi,a is the ith component of the solution to thealternative system (11).
Proof: We write
Pc,↵(i)(x) ⇡xi
kM,i
1 + xi
KM,i+ ⇢
and P↵(i)(x) as before. The first result follows directly fromcomputing the magnitude of the difference and the secondfrom integrating over the time interval.In practice it may be difficult to force the trajectories ofall other competing states to be small. Likewise, it maybe difficult to increase the Michaelis Menten constants ofall other competing species while maintaining the sameMichaelis constant for x↵(i). In particular, such a constraintwill be difficult to satisfy with an oscillator featuring two orthree states coupled by the same production enzyme. In order
for (out-of-phase) oscillations to occur, one state must bepresumably be abundant or another state relatively depleted.When competing for the same enzyme, this presents a diffi-culty for the depleted state as it returns from the minimumin its oscillatory trajectory to its peak, since it must out-compete the more abundant state for production enzyme.Analogous complications arise from a shared degradationenzyme. Conceptually, one solution to avoid this unwantedcrosstalk coupling is to provide two or three independentenzymes that facilitate the production and degradation of theoscillatory states, but the number of enzymes are limited sothis approach is not scalable.
B. Tuning Crosstalk
In this section, we take a different approach. Rather thanconsidering the dynamical constraints that must be imposedfor the crosstalk system to match the alternative system, weconsider effectively tuning certain aspects of the system sothat the crosstalk can be rendered negligible.
While it may be difficult to introduce n independentenzymes to drive and regulate the trajectories of n differentstates, it is certainly possible to incorporate extra substratesthat reliably maintain a steady state and exhibit populationsizes that wash out the fluctuations of native system species.We make emphasize this result, for its conceptual importancebut more especially for its practical significance.
Proposition 3: Let x1, ..., xs be the set of system func-tional substrates and let xs+1, ..., xs+b be a set of extrasubstrates that compete for the same enzyme as x1, ..., xs.Similarly, let xd(nd)+1, ..., xd(nd)+w be a set of extra sub-strates that compete for degradation enzyme. Suppose theseextra substrates maintain a steady state. If the trajectory ofxc(t) satisfies
sX
j=1
xj
KM,j⌧
s+bX
j=s+1
xj , e
KM,j
andd(nd)X
j=1
xj
M,j⌧
d(nd)+wX
j=d(nd)+1
xj , e
M,j
then fc(x) is robust to crosstalk and can be approxi-mated with linear production and degradation functionsPL(x), DL(x), scaled to reflect the dominant populations ofthe extra substrates.
Proof: The result follows immediately from approxi-mating the denominators in each entry of Pc(x) and Dc(x)with the dominant steady state terms of the extra substrates.
Since the denominators in the production and degrada-tion functions are approximately constant, production anddegradation become linear processes. Again, since the extrasubstrates are at steady state, this is effectively the sameas increasing the Michaelis Menten constants of competingsubstrates. It is these Michaelis Menten constants that dictatethe effects of production and degradation crosstalk.
The next theorem reveals a second parameter that is keyto attenuating crosstalk: the total production and degradation

7
enzyme concentration. Intuitively, if we want to reduce com-petition among the species for enzyme, then by increasingthe amount of enzyme, each substrate will be degraded orproduced as if degradation or production were a first orderprocess — the availability of substrate dictates the progressof the reaction.
Theorem 1: Suppose xq are bounded for q = 1, ..., ns andxl are bounded for l = d(1), ..., d(nd)). Then as Etot andEtot
d ! 1 then Cp(xi, xj) ! 0 and Cd(xi, xj) ! 0 forall i 6= j and taking Etot ! 1, Etot
d ! 1 simultaneously,then CF ! 0.
Proof: We consider a single production enzyme first,E, as the argument is identical from enzyme to enzyme.Suppose xi is produced by E in combination with substratesx1, ..., xk that form complexes (xl : E), l = 1, ..., k complexin competition with x1, ..., xnc and Etot is large, we insteadwrite xl = xfree
l + (xl : E) since there will be a nontrivialamount of enzyme-substrate complex formed. Suppose xj 2{x1, ..., xnc
}, we will show
limEtot!1
cp(xi, xj) = 0,
(otherwise, c(xi, xj) = 0 already). First, in the crosstalksystem (3), detailed balance yields
(xi : E) =xfreei Efree
kM,i
and by conservation of mass, we get
Efree = Etot �ncX
l=1
(xl : E)
= Etot �ncX
l=1
(xl � xfreel )
� Etot �ncX
l=1
xl
Following the derivation in [18], the modified productionrates can then be written as
Pc,i =kX
q=1
kcat,qxfreeq Efree
kM,q
and substituting for xfreeq = xq
1+Efree
kM,q
we get
Pc,i =kX
q=1
kcat,qxqE
free
kM,q + Efree(16)
Similarly, for the alternative system, we can derive
Pi =kX
q=1
kcat,qxqE
freeq
kM,q + Efreeq
.
By definition of the alternative system, Etotq = Etot for q =
1, ..., k. For each q, we have
E
freeq +E
freeq
xq
kM,q + E
freeq
= E
free+E
freencX
l=1
xl
kM,l + E
free.
and
cp(xi, xj) =kX
q=1
kcat,q
xqE
freeq
kM,q + E
freeq
� xqEfree
kM,q + E
free
!
=kX
q=1
kcat,q
0
@E
free � E
freeq +
X
l 6=q
E
freexl
kM,l + E
free
1
A
and as Etot approaches infinity, since xl are bounded, allfree xl are sequestered by enzyme, so that the free enzymecan be expressed as Efree = Etot �
Pnc
l=1 xl. Similarly,Efree
q = Etot � xq, q = 1..., k. Thus, by direct substitutionlimEtot!1 c(xi, xj) =
limEtot!1
kX
q=1
kcat,q
0
@�ncX
l=1
xl + xq +X
l 6=q
Efreexl
kM,l + Efree
1
A
= 0.
Since xi and xj were arbitrary, then the undirected crosstalkCp(xi, xj) must also approach 0 for all i and j as Etot
increases. A similar proof shows the degradation rates satisfy
limEtot
d !1Cd(xi, xj) ! 0
Thus, by the definition of CF and the algebraic propertiesof limits,
limEtot,Etot
d !1CF = lim
Etot,Etotd !1
X
i,j,i<j
C(xi, xj)
=X
i,j,i<j
limEtot!1
Cp(xi, xj)
+X
i,j,i<j
limEtot
d !1Cd(xi, xj) = 0
= 0
This theorem suggests that by increasing the total productionand degradation enzyme, the production and degradationcrosstalk correspondingly decrease. Indeed, this is made clearby examining the alternate expressions for production anddegradation in equation (16) and substituting the expressionfor Efree = Etot�
Pnc
l=1 xl. As we increase the total amountof enzyme, both the alternative and crosstalk production ratesbecome approximately linear, with the alternative systemapproaching slightly faster than the crosstalk system. Thissuggests a mode of adjusting system performance by simplyattenuating the production or degradation crosstalk. Again,this theorem is practically significant in in vitro systemswhere total enzyme concentrations can be highly controlled.As a final example, we illustrate how tuning enzyme con-centrations and the effective Michaelis Menten constantsin the Barkai-Liebler system attenuate crosstalk to recoveroscillatory dynamics.
Example 1 (Tuning the Barkai-Liebler System): In theBarkai-Liebler system (2), when RNAP is saturated,the templates DA and DR vie for the common RNAP,resulting in crosstalk: they effectively down-regulate eachother’s transcription rates. This crosstalk interferes withA promoting its own production through DA. Because

8
0 5 10 15 20 25 300
0.2
0.4
0.6
0.8
1
1.2
1.4
time (hr)
RNA
(µ M
)
ARC
Fig. 3. A simulation of the Barkai-Liebler system using identical parame-ters as Figure 2c.), but with four times the amount of RNAP and 1.08µMof background template.
the production and degradation of A must be on a fasttime-scale to achieve oscillation, we see that RNAP-mediated crosstalk deteriorates oscillation. On the otherhand, RNA signals A, R, and C experience degradationcrosstalk as they each compete for saturated RNase R, i.e.,through crosstalk the three RNA signals become mutuallyactivating. Except for R activating A (which is weak whenM,R > M,A), these mutual activation would fortify theexisting activating reaction cascades — A promotes creationof R and formation of C, R promotes formation of C, andC returns to R. Therefore, qualitatively, the saturation ofRNase R helps the circuit oscillate. Nevertheless, saturationin RNAP (production crosstalk) must be removed inorder for the beneficial effects of RNase R to be seen. InFigure 1, we see that by holding RNase R at its currentconcentration and increasing RNAP, we effectively reducethe production crosstalk and we recover oscillations in theBarkai-Liebler system (2). We note that by simultaneouslyincreasing all enzyme concentrations, the first orderproduction-degradation model (1) becomes valid again, butover a different set of parameters. Thus, models with linearproduction and degradation are not only instrumental incharacterizing simplified system dynamics, but representthe limit for large enzyme concentrations or small substrateconcentrations.
V. FUTURE WORK & CONCLUSIONS
In this work, we introduced signaling crosstalk as afundamental phenomenon of enzyme-mediated reactions inbiochemical systems. As a motivating example, we intro-duced an in vitro version of the Barkai-Liebler oscillatorand demonstrated that under simplifying assumptions, thesystem would oscillate. We posed a more realistic modelthat introduced crosstalk in the production and degradationfunctions and consequently, the system did not oscillate. Wethen posed a theoretical framework for quantifying crosstalkand used it to derive several dynamical constraints and the-oretical results that suggest design strategies for attenuatingcrosstalk. Finally, we showed that by effectively changing theMichaelis Menten constants or by changing the total amountof enzyme, the Barkai-Liebler system (2) was able to recoveroscillatory behavior.
We note that in this work we did not model degradationproducts . Because the chemical identity of degradation
products for a single enzyme-substrate pair can vary fromone instance of a degradation reaction to another, the re-sulting number of states required to describe all potentialdegradation products can be staggering. Furthermore, degra-dation products can result in another type of crosstalk. Futureresearch will address crosstalk from degradation products,hidden regulatory interactions from background gene expres-sion, and develop a framework for quantifying crosstalk invivo.
REFERENCES
[1] H. Kitano, “Systems biology: A brief overview,” Science, vol. 295, pp.1662-1664, 2002.
[2] M. Elowitz and W. A. Lim, “Build life to understand it,” Nature, vol.468, pp. 889-890, 2010.
[3] M. Scott, C. W. Gunderson, E. M. Mateescu, Z. Zhang, and T. Hwa,“Interdependence of cell growth and gene expression: Origins andconsequences,” Science, vol. 330, pp. 1099-1102, 2010.
[4] A. Zaslaver, et al., ”Invariant distribution of promoter activities inEscherichia coli,” PLoS Comput Biol, vol. 5, pp. e1000545, 2009.
[5] N. A. Cookson, et al., “Queueing up for enzymatic processing: corre-lated signaling through coupled degradation,” Mol Syst Biol, vol. 7, pp.561, 2011.
[6] Y. Rondelez, “Competition for catalytic resources alters biologicalnetwork dynamics,” Phys Rev Lett, vol. 108, pp. 018102, 2012.
[7] P. Marguet, et al., ”Oscillations by minimal bacterial suicide circuitsreveal hidden facets of host-circuit physiology,” PLoS One, vol. 5, pp.e11909, 2010.
[8] D. Del Vecchio, A. J. Ninfa, and E. D. Sontag, ”Modular cell biology:retroactivity and insulation,” Mol. Syst. Biol., vol. 4, pp. 161, 2008.
[9] L. K. Scheffer, L. Lavagno, and G Martin, Electronic design automationfor integrated circuits handbook, Boca Raton, Florida, CRC/Taylor &Francis, 2006.
[10] J. Liu, et al., ”Modelling and experimental analysis of hormonalcrosstalk in Arabidopsis,” Mol Syst Biol, vol. 6, pp. 373, 2010.
[11] P. H. Von Hippel, A. Revzin, C. A. Gross, and A. C. Wang, “Non-specific DNA binding of genome regulating proteins as a biologicalcontrol mechanism: 1. The lac operon: Equilibrium aspects,” Proc NatlAcad Sci USA, vol. 71, pp. 4808–4812, 1974.
[12] J. Kim, K. S. White, and E. Winfree, “Construction of an in vitrobistable circuit from synthetic transcriptional switches,” Mol Syst Biol,vol. 2, pp. 68, 2006.
[13] N. Barkai and S. Leibler, “Circadian clocks limited by noise,” Nature,vol. 403, pp. 267-268, 2000.
[14] J. M. G. Vilar, H. Y. Kueh, N. Barkai, and S. Leibler, “Mechanisms ofnoise-resistance in genetic oscillators,” Proc Natl Acad Sci USA, vol.99, pp. 5988-5992, 2002.
[15] A. Goldbeter, Biochemical Oscillations and Cellular Rhythms, Cam-bridge, UK: Cambridge University Press, 1999.
[16] K. E. McGinness and G. F. Joyce, “Substitution of ribonucleotides inthe T7 RNA polymerase promoter element,” J Biol Chem, vol. 277, pp.2987-2991, 2002.
[17] J. Kim, J. J. Hopfield, and E. Winfree, “Neural network computationby in vitro transcriptional circuits,” Advances in Neural InformationProcessing Systems, vol. 17, pp. 681-688, 2004.
[18] J. Kim and E. Winfree, “Synthetic in vitro transcriptional oscillators,”Mol Syst Biol, vol. 7, pp. 465, 2011.
[19] E. Franco, et al., “Timing molecular motion and production with asynthetic transcriptional clock,” Proc Natl Acad Sci USA, vol. 108, pp.E784-E793, 2011.
[20] H. A. Vincent and M. P. Deutscher, “Substrate recognition andcatalysis by the exoribonuclease RNase R,” J Biol Chem, vol. 281,pp. 20769-29775, 2006.
[21] J. J. Tyson, K. C. Chen, and B. Novak, ”Sniffers, buzzers, toggles andblinkers: dynamics of regulatory and signaling pathways in the cell,”Curr Opin Cell Biol, vol. 15, pp. 221–231, 2003.
[22] M. A. Savageau, “Design principles for elementary gene circuits:Elements, methods, and examples,” Chaos, vol. 11, pp. 142-159, 2001.
[23] Y. Yuan, G. Stan, S. Warnick and J . Goncalves, “Robust dynamicalnetwork structure reconstruction,”, Automatica, 2011.





























