Embed Size (px)
Citation preview

MOLECULAR
Molecular Phylogenetics and Evolution 33 (2004) 428–439
PHYLOGENETICSANDEVOLUTION
www.elsevier.com/locate/ympev
Phylogenetic relationships of the African bush-shrikesand helmet-shrikes (Passeriformes: Malaconotidae)
Jerome Fuchsa,b,*, Rauri C.K. Bowiec,d, Jon Fjeldsae, Eric Pasqueta,b
a FRE 2695 Origine, Structure et Evolution de la Biodiversite, Departement Systematique et Evolution, Museum National d�Histoire
Naturelle, 55 Rue Buffon, 75005 Paris, Franceb Service Commun de Systematique Moleculaire, IFR CNRS 101, Museum National d�Histoire Naturelle, 43, rue Cuvier, 75005 Paris, France
c Department of Zoology, University of Stellenbosch, Private Bag X1, Matieland 7602, South Africad Department of Zoology, Division of Birds, Field Museum of Natural History, 1400 S. Lake Shore Dr, Chicago, IL 60605, USA
e Zoological Museum, University of Copenhagen, DK-2100 Copenhagen, Denmark
Received 28 January 2004; revised 10 June 2004
Abstract
The African bush-shrikes and helmet-shrikes (Malaconotidae sensu [A Complete Checklist of the Birds of the World, third ed.,
Helm Editions, London, 2003]) include 10 genera and 52 species of predatory passerine birds for which monophyly, sister-group,
and inter-generic relationships are disputed. To resolve their relationships, we analyzed 2313bp of sequence data obtained from
two nuclear introns (myoglobin intron-2, b-fibrinogen intron-5) and a mitochondrial protein-coding gene (ND2) using parsimony,
maximum likelihood, and Bayesian inference. A strongly supported clade that included representatives of the Malaconotidae, Platy-
steiridae, and Vangidae was found in all analyses. Three main groups emerged within this clade but relationships between these three
groups were always poorly supported. The first group included the helmet-shrikes (Prionops), flycatcher-shrikes (Bias and Megab-
yas), and vangas (Cyanolanius and Pseudobias), currently placed in the families Malaconotidae, Platysteiridae, and Vangidae,
respectively. The second group consisted of four Platysteiridae genera (Lanioturdus, Batis, Platysteira, and Dyaphorophyia), with
the remaining Malaconotidae genera (�core malaconotids�) forming the last group. Two main clades emerged within the �core mala-
conotids,� with the position of the genus Nilaus being variable. The first clade includedMalaconotus, Dryoscopus, Bocagia, and Tcha-
gra and the second Chlorophoneus, Laniarius, Rhodophoneus, and Telophorus. Monophyly of the genus Chlorophoneus was never
recovered, a result that is consistent with morphological data.
� 2004 Elsevier Inc. All rights reserved.
Keywords: Bush-shrikes; Helmet-shrikes; ND2; Myoglobin intron-2; b-Fibrinogen intron-5
1. Introduction
The African bush-shrikes and helmet-shrikes (Mala-
conotidae sensu Howard and Moore, 2003) include 10
genera and 52 species that are variable in size and
plumage, some being drab and other brightly colored.
These predatory birds mainly feed on large insects
(Coleoptera, Hymenoptera, and Orthoptera) and even
1055-7903/$ - see front matter � 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.ympev.2004.06.014
* Corresponding author. Fax: +33-1-40-79-30-63.
E-mail address: [email protected] (J. Fuchs).
small mammals (rodents). Bush-shrikes and helmet-shrikes have traditionally been grouped with all Old
World shrikes in the family Laniidae (Morony et al.,
1975; Rand, 1960). The Laniidae is a poorly defined
assemblage that is only supported by the presence of
two adaptive characters, a hooked bill and the reduced
state of the Musculus flexor perforatus digiti III muscle
(Raikow et al., 1980). Results based on biochemical,
behavioral, and morphological characters (Harris andArnott, 1988; Harris and Franklin, 2000; Wolters,
1975–1982) or on DNA–DNA hybridization data

J. Fuchs et al. / Molecular Phylogenetics and Evolution 33 (2004) 428–439 429
(Sibley and Ahlquist, 1990) suggest that the Malaco-
notidae are only distantly related to ‘‘true’’ shrikes
(genera Corvinella, Eurocephalus, and Lanius) and the
Bornean Bristlehead (Pityriasis gymnocephala). Instead,
they suggest that the Malaconotidae are more closely
related to the families Platysteiridae or Vangidae.However, due to insufficient taxon sampling or analyt-
ical problems, these studies failed to resolve the rela-
tionships of bush-shrikes and helmet-shrikes. In
particular, monophyly, sister-group, and inter-generic
relationships remain controversial. The Malaconotidae,
Platysteiridae, and Vangidae are biogeographically
interesting as they represent African and Malagasy
clades of a larger group of corvids that apparentlyoriginated in Australia, subsequently radiating
throughout the Oriental region and finally colonizing
Africa (Barker et al., 2002; Ericson et al., 2002a).
The putative closely related family Platysteiridae
(batises, flycatcher-shrikes, and wattle-eyes) is endemic
to Africa and comprises six genera and 28 species
(Howard and Moore, 2003). All these species share
some distinctive morphological traits, e.g., a flat broadbill and bristles around the nostrils and gape, adapted
for flycatching behavior. Despite these differences in
foraging behavior and overall similarity, the Platysteir-
idae share many biochemical and morphological char-
acters with bush-shrikes and helmet-shrikes (Harris
and Arnott, 1988; Harris and Franklin, 2000; Pocock,
1966; Traylor, 1970; Wolters, 1975–1982). The family
Vangidae (15 genera and 22 species sensu Howardand Moore, 2003), endemic to Madagascar (with one
subspecies in the Comoros), is difficult to define due
to the considerable morphological divergence exhibited
within the family. A molecular study of Vangidae rela-
tionships suggests that these taxa are closely related to
a bush-shrike (Laniarius luehderi) (Schulenberg, 1993).
Further, similarity in vocalizations between the Vangi-
dae and some Malaconotidae (especially Malaconotus
alius and Telophorus kupeensis) lead Harris and Frank-
lin (2000) to propose these as potential sister-groups.
Monophyly and inter-generic relationships within the
Vangidae are still debated (Howard and Moore,
2003; Schulenberg, 1993; Yamagishi et al., 2001) and
resolving these relationships are beyond the scope of
our study.
There are also some phylogenetic problems withinthe Malaconotidae. First, the limits and monophyly
of the family are unclear. The position of the genus
Prionops (helmet-shrikes) in the Malaconotidae is par-
ticularly controversial due to the helmet-shrikes differ-
ing from other bush-shrikes by several morphological
and ethological characters, which has lead many
authors to treat them as relatively distant from other
bush-shrikes (Harris and Arnott, 1988; Raikow et al.,1980; Sibley and Monroe, 1990; Urban et al., 1997;
Wolters, 1975–1982). Some other genera, Nilaus—due
to its particular nest building and plumage—and Dry-
oscopus—due to its reproductive and communication
behavior—are thought to belong to the Platysteiridae
(Benson et al., 1971; Mayr, 1943; Traylor, 1970; Wol-
ters, 1975–1982). Conversely, the genera Bias, Megab-
yas, and Lanioturdus (Platysteiridae) are believed tobe closer to the Malaconotidae (Morony et al., 1975;
Rand, 1960; Sibley and Monroe, 1990; Traylor,
1970). Finally, limits of some genera (Bocagia, Chloro-
phoneus, Malaconotus, Rhodophoneus, Tchagra, and
Telophorus) are unclear. For example, the species La-
nius cruentus Ehrenberg, 1828 has been assigned to
the genera Tchagra, Telophorus or Rhodophoneus,
depending on the authors (Harris and Franklin, 2000;Howard and Moore, 2003; Morony et al., 1975; Rand,
1960; Sibley and Monroe, 1990; Urban et al., 2000;
Wolters, 1975–1982).
Here, we use 2313bp of aligned sequence data ob-
tained from two nuclear introns (myoglobin intron-2
and b-fibrinogen intron-5) and a mitochondrial pro-
tein-coding gene (ND2) to address the following ques-
tions: (1) Are the Malaconotidae sensu Howard andMoore (2003) monophyletic? (2) What is the sister-
group of the Malaconotidae? (3) What are the phyloge-
netic relationships among Malaconotidae genera?
2. Materials and methods
2.1. Taxon sampling
The DNA–DNA hybridization work by Sibley and
Ahlquist (1990) suggests a rapid mid-Tertiary radiation
of bush-shrike-like birds, with densely packed inter-
nodes. We selected a suite of species that encompass
the scope of this radiation, but avoids multiple represen-
tation from groups of closely related species. Resolving
the associations of the Malaconotidae, Platysteiridae,and Vangidae with various Asiatic taxa (Philentoma,
Tephrodornis, Aegithinidae, Artamidae, and Cractici-
dae, see Sibley and Ahlquist, 1990; Yamagishi et al.,
2001; and Barker et al., 2002) was beyond the scope of
our present study.
We obtained samples from all recognized Malaconot-
idae genera (19 species from 10 genera) as well as sam-
ples from putative closely related taxa in thePlatysteiridae (six species representing all six genera),
and Vangidae (two species). Trees were rooted with
the Tropical Kingbird (Tyrannus melancholichus, Tyran-
nidae, Suboscines) which is outside the Infraorder Os-
cines (see Ericson et al., 2003 for a review). Alternative
rooting with GenBank sequences from a stem Oscines,
i.e., Menura novaehollandiae (ND2: AY064754, Myogli-
bin: AY064744, Ericson et al., 2002b) did not affect thetopologies. Sample origins and GenBank accession
numbers are reported in Table 1.

Table 1
List of taxa studied (following Howard and Moore, 2003), voucher number, and GenBank accession numbers
Species Family Sample No. Origin Myoglobin b-Fibrinogen ND2
Ingroup taxa
Batis poensis Platysteiridae MNHN CG 1998-783 Cameroon AY529907 AY529974 AY529941
Bias musicus Platysteiridae MNHN 3-23 Cameroon AY529908 AY529975 AY529942
Bocagia minuta Malaconotidae ZMUC 128785 Congo AY529909 AY529976 AY529943
Campephaga flava Campephagidae AY165803a
Campephaga flava Campephagidae RCKB613 Kenya AY529977 AY529944
Chlorophoneus dohertyi Malaconotidae FMNH 358005 Burundi AY529910 AY529978 AY529945
Chlorophoneus nigrifrons Malaconotidae ZMUC 120151 Tanzania AY529911 AY529979 AY529946
Chlorophoneus sulfureopectus Malaconotidae MNHN CG 1998-823 Malawi AY529912 AY529980 AY529947
Coracina melaschistos Campephagidae MNHN 6-69 Laos AY529913 AY529981 AY529948
Corvus corone Corvidae MNHN 13-16 France AY529914 AY529982 AY529949
Cyanolanius madagascarinus Vangidae MNHN E117 Comoros AY529915 AY529983 AY529950
Dicrurus paradiseus Dicruridae MNHN 5-57 Laos AY529916 AY529984 AY529951
Dryoscopus cubla Malaconotidae ZMUC 116780 Kenya AY529917 AY529985 AY529952
Dryoscopus gambensis Malaconotidae ZMUC 124320 Kenya AY529918 AY529986 AY529953
Dyaphorophyia chalybea Platysteiridae MNHN 3-19 Cameroon AY529919 AY529987 AY529954
Laniarius aethiopicus Malaconotidae FMNH 356738 Tanzania AY529920 AY529988 AY529955
Laniarius barbarus Malaconotidae ZMUC 116792 Cage-bird AY529921 AY529989 AY529956
Laniarius funebris Malaconotidae ZMUC 123466 Tanzania AY529922 AY529990 AY529957
Laniarius luehderi Malaconotidae ZMUC 119044 Uganda AY529923 AY529991 AY529958
Lanioturdus torquatus Platysteiridae US-001 Namibia AY529924 AY529992 AY529959
Lanius collaris Laniidae MNHN 2-26 Cameroon AY529925 AY529993 AY529960
Malaconotus blanchoti Malaconotidae ZMUC 116824 Kenya AY529926 AY529994 AY529961
Megabyas flammulatus Platysteiridae MNHN CG 1968-1160 Kenya AY529927 AY529995 AY529962
Nilaus afer Malaconotidae FMHN uncatalogued Bostwana AY529928 AY529996 AY529963
Oriolus xanthornus Oriolidae MNHN 4-10D Thailand AY529929 AY529997 AY529964
Platysteira cyanea Platysteiridae MNHN 2-22 Cameroon AY529930 AY529998 AY529965
Prionops retzii Malaconotidae ZMUC 119500 Kenya AY529931 AY529999 AY529966
Prionops scopifrons Malaconotidae ZMUC 117528 Kenya AY529932 AY530000 AY529967
Pseudobias wardi Vangidae FMHN 356702 Madagascar AY529933 AY530001 AY529968
Rhipidura albicollis Rhipiduridae MNHN 5-48 Laos AY529934 AY530002 AY529969
Rhodophoneus cruentus Malaconotidae US-002 Kenya AY529935 AY530003 AY529970
Tchagra australis Malaconotidae ZMUC 116831 Kenya AY529936 AY530004 AY529971
Tchagra senegala Malaconotidae ZMUC 116834 Kenya AY529937 AY530005 AY529972
Telophorus zeylonus Malaconotidae FMNH 390107 South Africa AY529938 AY530006 AY529973
Terpsiphone viridis Monarchidae MNHN 2-20 Cameroon AY529939 AY530007
Terpsiphone viridis Monarchidae AF407058b
Outgroup taxa
Tyrannus melancholicus Tyrannidae MNHN 28-56 South America AY529940 AY530008
Empidonax flaviventris Tyrannidae AF447629c
Acronyms are: FMNH, Field Museum of Natural History, Chicago, USA; MNHN, Museum National d�Histoire Naturelle, Paris, France; RCKB,
Rauri C.K. Bowie; US, University of Stellenbosch, South Africa; and ZMCU, Zoological Museum, University of Copenhagen, Denmark.a Johanson and Ericson (2003).b Sorenson and Payne (2001).c Cicero and Johnson (2002).
430 J. Fuchs et al. / Molecular Phylogenetics and Evolution 33 (2004) 428–439
2.2. Laboratory procedures
Genomic DNA was extracted from either alcohol-
preserved tissues, frozen tissues (muscle, blood) or from
skin of museum specimens (0.5–1cm2) using a CTAB
buffer with Proteinase K (0.1mgml�1) or a Puregene
DNA isolation kit (Gentra Systems, Minneapolis, Min-
nesota). Myoglobin intron-2 was amplified with primers
Myo 2 and Myo 3F (Heslewood et al., 1998; Slade et al.,1993) and in a few cases with internal primers (Myo2int:
50 TRA GAC CCA TAA AAC TAA GTG 30; Myo3int:
50 TGA TCT GCT TCA TGA CCT T 30). The ND2
gene was amplified with primers L5219 Met andH6313 Trp (Sorenson et al., 1999). b-Fibrinogen in-
tron-5 was amplified using the primers Fib5 (50 CGC
CAT ACA GAG TAT ACT GTG ACA T 30) and
Fib6 (50 GCC ATC CTG GCG ATT CTG AA 30).
The thermocycling conditions included a hotstart at
94 �C, an initial denaturation at 94 �C for 3min, fol-
lowed by 35–40 cycles at 94 �C for 40s, 52–56 �C for
30s, and 72 �C for 50s, and was completed by a finalextension at 72 �C for 5min. Three microliters of the
amplification products was electrophoresed on 1.5%
agarose gel and visualized under UV light with ethidium

J. Fuchs et al. / Molecular Phylogenetics and Evolution 33 (2004) 428–439 431
bromide to check for the correct fragment size and to
control for the specificity of the amplifications. For b-fi-brinogen intron-5, amplicons of the appropriate length
were cut out of the gel and purified using GELase (Epi-
centre Technologies, Madison, Wisconsin). Purified
PCR products were cycle-sequenced using the ‘‘CEQDye Terminator Cycle Sequencing’’ kit (Beckman) or
the Big Dye terminator chemistry (Applied Biosystems
[ABI]) in both forward and reverse direction with the
same primers used for PCR amplifications and run on
an automated CEQ2000 DNA Analysis System sequen-
cer (Beckman) or on an ABI 3100 automated DNA se-
quencer. Sequences were always obtained from both
DNA strands. All b-fibrinogen intron-5 sequences werescreened for the presence of conserved exon sequences at
both the 30 and 50 ends, to ensure that the whole intron
was sequenced and that any length variation was due to
the presence of insertions or deletions (indels).
Allelic variation within individuals, i.e., heterozygos-
ity, was apparent in the two nuclear loci. Heterozygosity
could occur in two different ways: (1) length variation
between alleles and (2) single nucleotide polymorphism(SNP). Length variation between alleles was only appar-
ent for b-fibrinogen intron-5. PCR products from such
individuals were cloned using the TOPO cloning kit
(Inviatron) and multiple clones (6–10) were cycle-se-
quenced. A total of four species were cloned. Length
variation was present in two taxa (Tyrannus melancholi-
cus and Lanius collaris) with the alleles differing by 1 or
2bp. Both alleles were included in initial parsimonyanalyses. In all instance the two alleles from a single
individual clustered together with high bootstrap sup-
port and we therefore used the consensus sequences of
the two alleles obtained with Sequencher 4.1 (Genes
Codes, Ann Arbor, Michigan) for further analysis.
Occurrence of SNP in some b-fibrinogen intron-5 and
myoglobin intron-2 sequences was suggested by the
presence of double peaks. These double peaks were trea-ted using the appropriate IUPAC code.
2.3. Sequences alignment
Multiple alignment of intron sequences was accom-
plished by hand modification after an initial alignment
by Sequencher 4.1. Final alignment indicated the pres-
ence of several insertions–deletions events in the two nu-clear introns (18 in the myoglobin intron-2 and 39 in the
b-fibrinogen intron-5). Alignment of these indels was
fairly straightforward, yielding no region of significant
ambiguity, i.e., that led to topological differences be-
tween different alignments, in the myoglobin intron-2 se-
quences. Indels in the b-fibrinogen intron-5 sequences
could not be unambiguously aligned in one zone, mainly
due to the presence of multiple insertions–deletionsevents within an A–T-rich zone which made hypothesis
about homology questionable. We deleted this particu-
lar zone from our analyses (nucleotides 77–90 in our
b-fibrinogen intron-5 alignment). After removing align-
ment ambiguities, the number of potential informative
indels were 4 and 12 for myoglobin intron-2 and b-fi-brinogen intron-5, respectively. Gaps were treated as
missing data.
2.4. Phylogenetic analyses
2.4.1. Search parameters
Phylogenetic trees were estimated using parsimony,
maximum likelihood, and Bayesian inferences as imple-
mented in PAUP* 4.0b10 (Swofford, 2001) and MrBa-
yes v.3.0 (Huelsenbeck et al., 2001). Parsimony (P)analyses were performed under the heuristic search op-
tion with 10 random addition replicates with the
branch-swapping option tree bisection and reconnection
(TBR) implemented. Support for nodes was estimated
using the non-parametric bootstrap (Felsenstein, 1985)
with 1000 replicates. Likelihood models and parameters
were estimated using Modeltest v.3.04 (Posada and
Crandall, 1998) and the Hierarchical Likelihood RatioTest criterion. We also tested the applicability of the
molecular clock for all individual data set as well as
for the combined data set by using the Likelihood Ratio
Test (LRT) (�2 lnA = 2(lnk1 � lnk2), where k1 is the
likelihood of the restricted model) (Huelsenbeck and
Rannala, 1997), which follows a v2 distribution with
n � 2 degrees of freedom where n is the number of taxa.
Tree searches were estimated by heuristic TBR branch-swapping with 10 random addition replicates. Nodal
support was calculated using the non-parametric boot-
strap (100 replicates). For comparing different a priori
and a posteriori topologies, we use the non-parametric
Shimodaira–Hasegawa test (SH test) (Shimodaira and
Hasagawa, 1999) implemented in PAUP. Scores of the
constrained trees were obtained by instructing PAUP
to find the best tree under a specific constraint. TheSH test between the best tree and the constrained tree
was then performed using the RELL option with 1000
bootstrap replicates.
For the Bayesian analyses, the optimal model param-
eterization as determined by ModelTest was assumed
for each data set. Bayesian analyses for the combined
data set were performed freeing different parameters to
vary between the three partitions (ND2, myoglobin in-tron-2, and b-fibrinogen intron-5) (using the prset and
unlink options). Each analysis was initiated from a ran-
dom starting tree. Analyses were conducted using
Metropolis coupling chains with four incrementally
heated Markov chains. Chains were run for 4 · 106 gen-
erations, and sampled every 100. We determined the
minimum number of generations to discard as ‘‘burn-
in’’ by plotting likelihood scores against generationnumber. Posterior probabilities for clades were esti-
mated by a 50% majority-rule consensus tree based on
432 J. Fuchs et al. / Molecular Phylogenetics and Evolution 33 (2004) 428–439
the remaining saved trees. Four independent Bayesian
runs were performed for each data set. Log-likelihood
values and posterior probabilities were also checked to
confirm that using a different starting tree did not alter
our results significantly.
2.4.2. Evaluation of congruence
Significant incongruences between the individual gene
trees were detected by comparing the topologies and no-
dal support obtained under different analytical methods
(parsimony, maximum likelihood, and Bayesian infer-
ences). Criterions for incongruence were set at 70% for
the bootstrap values, according to Hillis and Bull
(1993), and at 0.95 for posterior probabilities; these lat-ter are argued to have a straightforward statistical inter-
pretation (Huelsenbeck et al., 2001). The support or
conflict of individual data sets with particular nodes
recovered in the shortest parsimony trees was also eval-
uated using the Partitioned Bremer Support (Baker and
DeSalle, 1997; Baker et al., 1998; Bremer, 1994), calcu-
lated with TreeRot v. 2 (Sorenson, 1999).
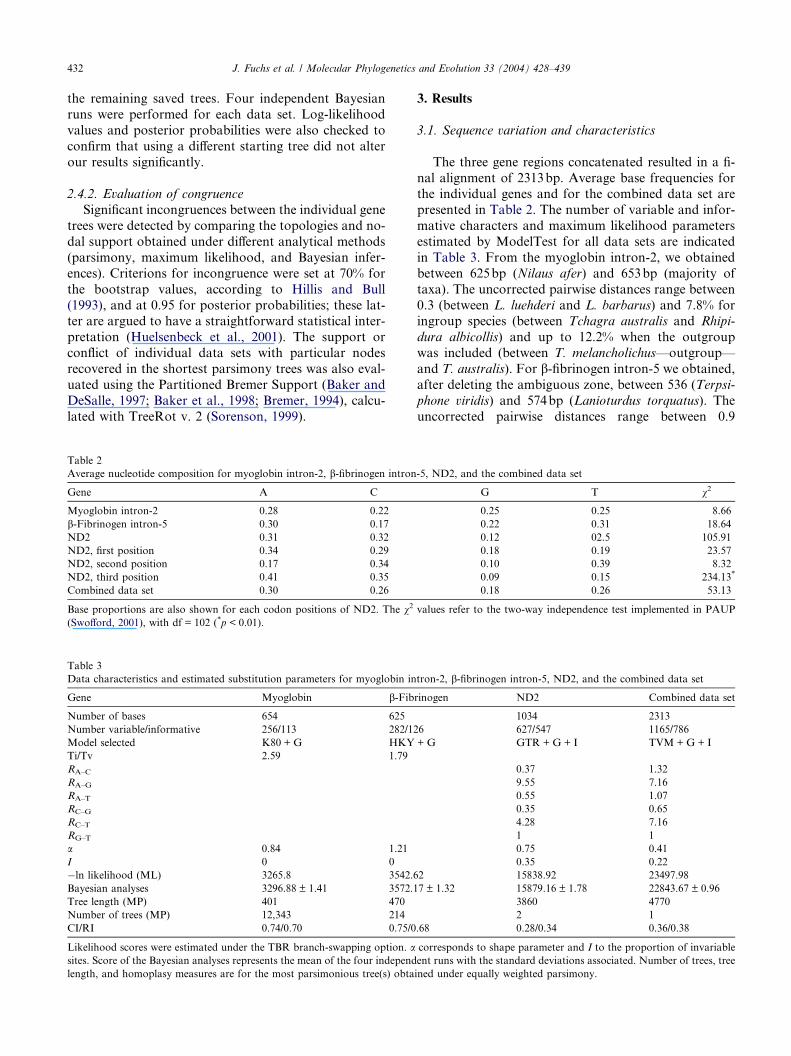
Table 2
Average nucleotide composition for myoglobin intron-2, b-fibrinogen intron
Gene A C
Myoglobin intron-2 0.28 0.22
b-Fibrinogen intron-5 0.30 0.17
ND2 0.31 0.32
ND2, first position 0.34 0.29
ND2, second position 0.17 0.34
ND2, third position 0.41 0.35
Combined data set 0.30 0.26
Base proportions are also shown for each codon positions of ND2. The v2
(Swofford, 2001), with df = 102 (*p < 0.01).
Table 3
Data characteristics and estimated substitution parameters for myoglobin in
Gene Myoglobin b-Fib
Number of bases 654 625
Number variable/informative 256/113 282/12
Model selected K80 + G HKY
Ti/Tv 2.59 1.79
RA–C
RA–G
RA–T
RC–G
RC–T
RG–T
a 0.84 1.21
I 0 0
�ln likelihood (ML) 3265.8 3542.6
Bayesian analyses 3296.88 ± 1.41 3572.1
Tree length (MP) 401 470
Number of trees (MP) 12,343 214
CI/RI 0.74/0.70 0.75/0
Likelihood scores were estimated under the TBR branch-swapping option. asites. Score of the Bayesian analyses represents the mean of the four independ
length, and homoplasy measures are for the most parsimonious tree(s) obta
3. Results
3.1. Sequence variation and characteristics
The three gene regions concatenated resulted in a fi-
nal alignment of 2313bp. Average base frequencies forthe individual genes and for the combined data set are
presented in Table 2. The number of variable and infor-
mative characters and maximum likelihood parameters
estimated by ModelTest for all data sets are indicated
in Table 3. From the myoglobin intron-2, we obtained
between 625bp (Nilaus afer) and 653bp (majority of
taxa). The uncorrected pairwise distances range between
0.3 (between L. luehderi and L. barbarus) and 7.8% foringroup species (between Tchagra australis and Rhipi-
dura albicollis) and up to 12.2% when the outgroup
was included (between T. melancholichus—outgroup—
and T. australis). For b-fibrinogen intron-5 we obtained,
after deleting the ambiguous zone, between 536 (Terpsi-
phone viridis) and 574bp (Lanioturdus torquatus). The
uncorrected pairwise distances range between 0.9
-5, ND2, and the combined data set
G T v2
0.25 0.25 8.66
0.22 0.31 18.64
0.12 02.5 105.91
0.18 0.19 23.57
0.10 0.39 8.32
0.09 0.15 234.13*
0.18 0.26 53.13
values refer to the two-way independence test implemented in PAUP
tron-2, b-fibrinogen intron-5, ND2, and the combined data set
rinogen ND2 Combined data set
1034 2313
6 627/547 1165/786
+ G GTR + G + I TVM + G + I
0.37 1.32
9.55 7.16
0.55 1.07
0.35 0.65
4.28 7.16
1 1
0.75 0.41
0.35 0.22
2 15838.92 23497.98
7 ± 1.32 15879.16 ± 1.78 22843.67 ± 0.96
3860 4770
2 1
.68 0.28/0.34 0.36/0.38
corresponds to shape parameter and I to the proportion of invariable
ent runs with the standard deviations associated. Number of trees, tree
ined under equally weighted parsimony.

J. Fuchs et al. / Molecular Phylogenetics and Evolution 33 (2004) 428–439 433
(between L. luehderi and L. aethiopicus) and 10.9% for
ingroup species (between Dyaphorophyia chalybea and
Rhipidura abicollis) and up to 15.4% with the inclusion
of the outgroup (between T. melancholichus and Lanius
collaris).
For ND2, we obtained a total of 1034bp, corre-sponding to positions 5246–6279 of the mtDNA chicken
sequence (Desjardins and Morais, 1990). We could not
obtain a clearly resolved chromatogram for T. melan-
cholichus. Thus, we used the ND2 sequence of Empido-
nax flaviventris (GenBank Accession No. AF447626,
Cicero and Johnson, 2002) that also belong to the family
Tyrannidae (Suboscines) as an outgroup. The v2 test
implemented in PAUP detected no base compositionbias (p = 0.38) among the whole gene. Homogeneity
across the taxa was not rejected for first and second co-
don positions (Table 2), however, the test detected sig-
nificant heterogeneity across all taxa for the third
codon position, with a deficiency in guanine. Deficiency
in guanine at third codon position, as well as the absence
of insertions, deletions, and stop-codon in the reading
frame and the absence of abnormal amino acid substitu-tion pattern in closely related taxa are typical of mito-
chondrial DNA of birds (Barker, 2004; Bates et al.,
1999; Hackett, 1996; Sorenson and Quinn, 1998), sug-
gesting that we had not sequenced nuclear pseudogenes.
The numbers of parsimony informative sites per codon
position were: 159 (29%) at first codon position, 64
(12%) at second position, and 324 (59%) at third posi-
tion. The uncorrected pairwise distances range between8.8 (between L. funebris and L. aethiopicus) and 25.4%
for ingroup species (between Bias musicus and Telopho-
rus zeylonus) and up to 30.75% when including the out-
group (between E. flaviventris and T. zeylonus).
3.2. Evaluation of congruence between the three genes
Only two significant topological incongruences, as in-ferred from bootstrap and Partitioned Bremer Support
values, were found between the individual genes trees
(Table 4). The first one involves the positions of the gen-
era Batis and Lanioturdus relative to Platysteira and
Dyaphorophyia (node E on Fig. 1). The three molecular
markers are all in siginificant conflict for this node. b-Fi-brinogen intron-5 indicated a sister-group relationships
between Batis and Lanioturdus while myoglobin in-tron-2 suggested that Lanioturdus is more closely related
to Platysteira and Dyaphorophyia than to Batis. Finally,
ND2 strongly supported the basal position of Laniotur-
dus in relation to Batis, Platysteira, and Dyaphorophyia.
The second incongruent node involves the position of
the genera Dryoscopus and Malaconotus relative to Boc-
agia and Tchagra (node I on Fig. 1). ML and Bayesian
analyses of ND2 and all analysis undertaken with thefibrinogen sequences supported a sister-group relation-
ships between Malaconotus and Dryoscopus, while the
unweighted MP analysis of the mitochondrial data set
suggested that Dryoscopus is closer to Bocagia and
Tchagra than to Malaconotus, but with bootstrap values
close to 70%, i.e., the threshold of significant
incongruence.
The Molecular Clock model was rejected for themitochondrial gene, suggesting rate variation among lin-
eages, but not for the two nuclear introns (Table 5).
3.3. Phylogenetic results—combined data set
Parsimony analysis resulted in a single most parsimo-
nious tree (Fig. 1; length = 4770 steps, CI = 0.36,
RI = 0.38) with the Malaconotidae, Platysteiridae, andVangidae forming a well-supported clade (Boot-
strap = 91%; node A on Fig. 1); a result supported in
both the ML (�ln = 23497.98, TVM + I + G model,
see Table 3 for parameters) and BI (GTR + G + I model
for ND2, a K80 + G model for myoglobin intron-2, and
a HKY85 + G model for b-fibrinogen intron-5) analyses
(Fig. 2, Table 4). A molecular clock was rejected
(�2 lnA = 87.44; df = 33; p = <0.001) when the threedata sets were combined. No significant variations in
the posterior probabilities among the four independent
runs and log-likelihood value were detected (Table 5).
In all three analyses the Malaconotidae (sensu How-
ard and Moore, 2003) are not monophyletic due to Pri-
onops being more closely related to the vangas
(Cyanolanius and Pseudobias, Vangidae) and flycatch-
er-shrikes (Bias and Megabyas, Platysteiridae) than totraditional Malaconotidae genera (Figs. 1 and 2; Table
4). Monophyly of the Malaconotidae was rejected by
the Shimodaira–Hasegawa test (dL = 39.14, p = 0.017).
The Platysteiridae genera (Lanioturdus, Batis, Platy-
steira, and Dyaphorophyia) are united in a single clade
in all three analyses (node D on Fig. 1, hereafter called
the �core platysteirids�) with considerable support (Figs.
1 and 2, Table 4). In the MP analyses the �core platy-steirids� are sister to the �core malaconotids,� whereasin the ML and BI analyses this clade is sister to the �corevangids.� However, neither hypothesis is strongly sup-
ported (Figs. 1 and 2, Table 4) and the SH test did
not reject the MP hypothesis (dL = 1.06 p = 0.42).
In all three analyses two main clades emerged within
the �core malaconotids� (node F on Fig. 1). The first in-
cluded Malaconotus, Dryoscopus, Bocagia, and Tchagra
(clade H) and the second Chlorophoneus, Laniarius,
Rhodophoneus, and Telophorus (clade J). Position of
the genus Nilaus was variable and not well supported
in any of the three analyses. Monophyly of the genus
Chlorophoneus was never recovered as C. dohertyi is
more closely related to Telophorus and Rhodophoneus
than to other Chlorophoneus species. Monophyly of
the genera Dryocopus, Laniarius, and Tchagra were allstrongly supported (Figs. 1 and 2, Table 4). Slight re-ar-
rangements among taxa occurred within clade H (see
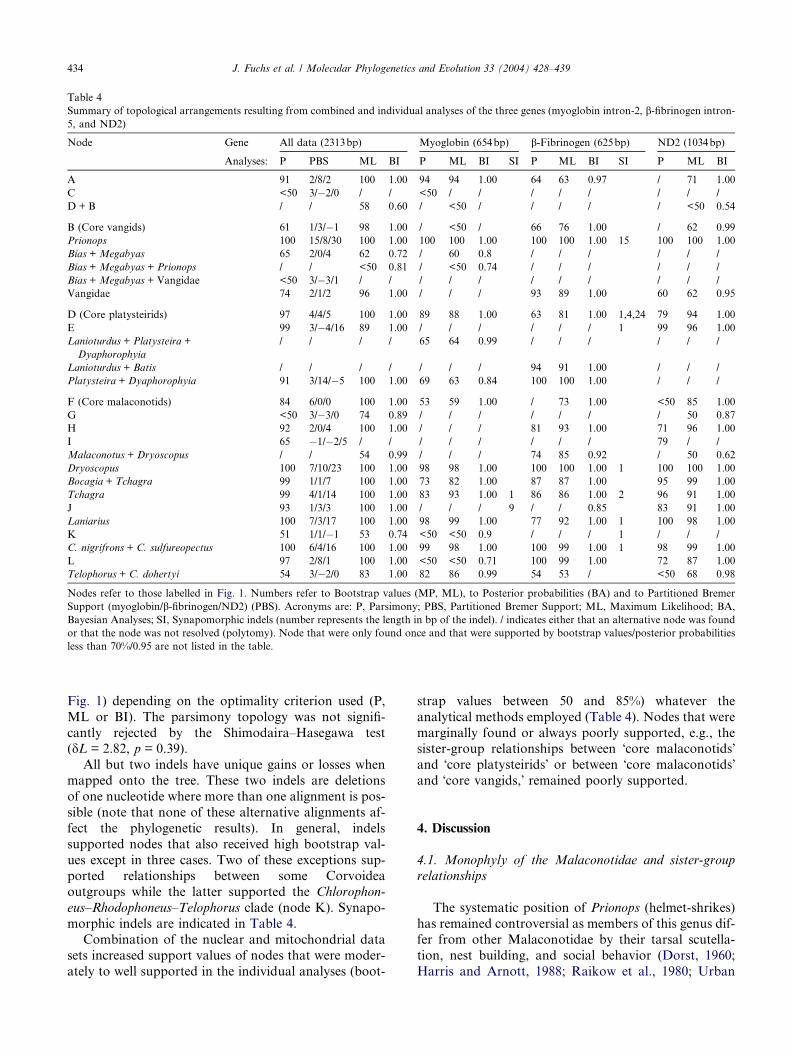
Table 4
Summary of topological arrangements resulting from combined and individual analyses of the three genes (myoglobin intron-2, b-fibrinogen intron-
5, and ND2)
Node Gene All data (2313bp) Myoglobin (654bp) b-Fibrinogen (625bp) ND2 (1034bp)
Analyses: P PBS ML BI P ML BI SI P ML BI SI P ML BI
A 91 2/8/2 100 1.00 94 94 1.00 64 63 0.97 / 71 1.00
C <50 3/�2/0 / / <50 / / / / / / / /
D + B / / 58 0.60 / <50 / / / / / <50 0.54
B (Core vangids) 61 1/3/�1 98 1.00 / <50 / 66 76 1.00 / 62 0.99
Prionops 100 15/8/30 100 1.00 100 100 1.00 100 100 1.00 15 100 100 1.00
Bias + Megabyas 65 2/0/4 62 0.72 / 60 0.8 / / / / / /
Bias + Megabyas + Prionops / / <50 0.81 / <50 0.74 / / / / / /
Bias + Megabyas + Vangidae <50 3/�3/1 / / / / / / / / / / /
Vangidae 74 2/1/2 96 1.00 / / / 93 89 1.00 60 62 0.95
D (Core platysteirids) 97 4/4/5 100 1.00 89 88 1.00 63 81 1.00 1,4,24 79 94 1.00
E 99 3/�4/16 89 1.00 / / / / / / 1 99 96 1.00
Lanioturdus + Platysteira +
Dyaphorophyia
/ / / / 65 64 0.99 / / / / / /
Lanioturdus + Batis / / / / / / / 94 91 1.00 / / /
Platysteira + Dyaphorophyia 91 3/14/�5 100 1.00 69 63 0.84 100 100 1.00 / / /
F (Core malaconotids) 84 6/0/0 100 1.00 53 59 1.00 / 73 1.00 <50 85 1.00
G <50 3/�3/0 74 0.89 / / / / / / / 50 0.87
H 92 2/0/4 100 1.00 / / / 81 93 1.00 71 96 1.00
I 65 �1/�2/5 / / / / / / / / 79 / /
Malaconotus + Dryoscopus / / 54 0.99 / / / 74 85 0.92 / 50 0.62
Dryoscopus 100 7/10/23 100 1.00 98 98 1.00 100 100 1.00 1 100 100 1.00
Bocagia + Tchagra 99 1/1/7 100 1.00 73 82 1.00 87 87 1.00 95 99 1.00
Tchagra 99 4/1/14 100 1.00 83 93 1.00 1 86 86 1.00 2 96 91 1.00
J 93 1/3/3 100 1.00 / / / 9 / / 0.85 83 91 1.00
Laniarius 100 7/3/17 100 1.00 98 99 1.00 77 92 1.00 1 100 98 1.00
K 51 1/1/�1 53 0.74 <50 <50 0.9 / / / 1 / / /
C. nigrifrons + C. sulfureopectus 100 6/4/16 100 1.00 99 98 1.00 100 99 1.00 1 98 99 1.00
L 97 2/8/1 100 1.00 <50 <50 0.71 100 99 1.00 72 87 1.00
Telophorus + C. dohertyi 54 3/�2/0 83 1.00 82 86 0.99 54 53 / <50 68 0.98
Nodes refer to those labelled in Fig. 1. Numbers refer to Bootstrap values (MP, ML), to Posterior probabilities (BA) and to Partitioned Bremer
Support (myoglobin/b-fibrinogen/ND2) (PBS). Acronyms are: P, Parsimony; PBS, Partitioned Bremer Support; ML, Maximum Likelihood; BA,
Bayesian Analyses; SI, Synapomorphic indels (number represents the length in bp of the indel). / indicates either that an alternative node was found
or that the node was not resolved (polytomy). Node that were only found once and that were supported by bootstrap values/posterior probabilities
less than 70%/0.95 are not listed in the table.
434 J. Fuchs et al. / Molecular Phylogenetics and Evolution 33 (2004) 428–439
Fig. 1) depending on the optimality criterion used (P,
ML or BI). The parsimony topology was not signifi-
cantly rejected by the Shimodaira–Hasegawa test(dL = 2.82, p = 0.39).
All but two indels have unique gains or losses when
mapped onto the tree. These two indels are deletions
of one nucleotide where more than one alignment is pos-
sible (note that none of these alternative alignments af-
fect the phylogenetic results). In general, indels
supported nodes that also received high bootstrap val-
ues except in three cases. Two of these exceptions sup-ported relationships between some Corvoidea
outgroups while the latter supported the Chlorophon-
eus–Rhodophoneus–Telophorus clade (node K). Synapo-
morphic indels are indicated in Table 4.
Combination of the nuclear and mitochondrial data
sets increased support values of nodes that were moder-
ately to well supported in the individual analyses (boot-
strap values between 50 and 85%) whatever the
analytical methods employed (Table 4). Nodes that were
marginally found or always poorly supported, e.g., thesister-group relationships between �core malaconotids�and �core platysteirids� or between �core malaconotids�and �core vangids,� remained poorly supported.
4. Discussion
4.1. Monophyly of the Malaconotidae and sister-group
relationships
The systematic position of Prionops (helmet-shrikes)
has remained controversial as members of this genus dif-
fer from other Malaconotidae by their tarsal scutella-
tion, nest building, and social behavior (Dorst, 1960;
Harris and Arnott, 1988; Raikow et al., 1980; Urban

Fig. 1. Parsimony tree (4770 steps; CI = 0.36; RI = 0.38) obtained with
the unweighted combined data set. Numbers below branches represent
bootstrap values greater than 50% (1000 replicates). The Malaconot-
idae (sensu Howard and Moore, 2003) are in bold type.
Fig. 2. Maximum likelihood tree (�ln = 23497.98) and Bayesian tree
(mean log-likelihood value = �22843.67, SD = 0.96) obtained with the
combined data set under a TVM + I + G model for the ML search and
with the combined data set with the best-fit models estimated by
Modeltest 3 assigned to each partition for the Bayesian analysis (a
GTR + G + I model for ND2, a K80 + G model for myoglobin intron-
2, and a HKY85 + G model for b-fibrinogen intron-5). ML parameters
estimated from Modeltest 3.04: bases frequencies = (0.3155, 0.2711,
0.1602, 0.2532), Rmat = (1.32, 7.16, 1.07, 0.65, 7.16), shape = 0,41, and
Pinvar = 0.22. Four heated Markov chains were run for 4 · 106
generations for the Bayesian analysis. The first 2000 trees (200,000
generations) were discarded (�burn-in�). The three gamma shape
distributions were specified as unlinked. Symbols */* next to branches
represent bootstrap values/posterior probabilities greater than 95%/
0.95. The Malaconotidae (sensu Howard and Moore, 2003) are in bold
type.
J. Fuchs et al. / Molecular Phylogenetics and Evolution 33 (2004) 428–439 435
et al., 2000). Interestingly, DNA–DNA hybridization
data suggested a placement of Prionops closer to the
Vangidae and Platysteiridae than to other Malaconoti-
dae (Sibley and Ahlquist, 1990), but its exact relation-
ships remained uncertain due to short internodes, the
fact that only four species were used as �drivers� and that
tests of robustness were lacking. Our analyses confirmedthe tentative result obtained by the DNA–DNA hybrid-
izations by providing strong support for a clade com-
prising Prionops, Bias, and Megabyas (Platysteiridae),
and Cyanolanius and Pseudobias (as representatives of
the Vangidae). Several authors have proposed a close
link between Prionops and the Vangidae because they
share characters such as tarsal scutellation, jaw muscula-
ture, bill structure, and gregarious behavior (Beecher,1953; Benson, 1985; Dorst, 1960). Our analyses are the
first to provide strong support for this hypothesis.
Table 5
Likelihood Ratio Test on the applicability of the molecular clock to the dat
Gene �lnka �lnkb
Myoglobin intron-2 3277.80 3265.80
b-Fibrinogen intron-5 3561.96 3542.62
ND2 15877.02 15838.92
Combined data set 23541.70 23497.98
a The log likelihood of the restricted model (application of the molecularb The log likelihood of the non-restricted model (non-application of the mc H0 accepted.d H0 rejected.
Previous studies that tackle themonophyly of the Van-
gidae only included one representative of the supposed
closest genus (Laniarius), providing thus only a weak test
of Vangidae monophyly (Schulenberg, 1993; Yamagishiet al., 2001). In our present study, the two Vangidae
a
�2 lnA df p
24 33 0.87c
38.68 33 0.77c
76.20 33 <0.001d
87.44 33 <0.001d
clock).
olecular clock).

436 J. Fuchs et al. / Molecular Phylogenetics and Evolution 33 (2004) 428–439
genera (Cyanolanius andPseudobias) clustered together in
all combined analyses. As we only sampled two vangid
species, we were unable to test the monophyly of the Van-
gidae. This needs to be further explored with representa-
tives of all Vangidae genera. Here, we show that an
appropriate taxonomic sampling for this test should in-clude a substantial number of genera of the �core malaco-
notids,� �core platysteirids,� and �core vangids.�Bias and Megabyas have sometimes been merged in
Bias due to their similar skull, plumage (black upper-
parts with green sheen in males, rufous in females), size,
and proportions (Erard, 1987; Traylor, 1970). Our anal-
yses failed to provide strong support for a sister-group
relationship between these two genera, although theyform a sister-group in the phylogeny. Molecular diver-
gence as well as differences in ecology, morphology, so-
cial organization, and general behavior are sufficiently
high for retaining two genera (Harris and Franklin,
2000). The position of Bias and Megabyas within the
�core vangids� is quite surprising because they were sus-
pected to be related to the �core platysteirids,� despitesome morphological differences (Erard, 1987). Never-theless, their relationships to other birds have remained
obscure mainly due to the lack of comparative data
(Harris and Arnott, 1988; Harris and Franklin, 2000;
Traylor, 1970).
The systematic affinities of Lanioturdus have for
many years been uncertain, as its name suggests. It has
been considered an atypical member of the �core malaco-
notids� (Morony et al., 1975; Rand, 1960; Sibley andMonroe, 1990) or a close relative of the helmet-shrikes
(Wolters, 1975–1982). All our analyses included it in
the �core platysteirids� with strong support (high boot-
strap values and three synapomorphic indels). Its posi-
tion within the �core platysteirids� is also supported by
jaw musculature and plumage pattern (Beecher, 1953;
Harris and Arnott, 1988; Harris and Franklin, 2000; Ur-
ban et al., 1997). Thus, Lanioturdus represents an aber-rant platysteirid adapted to a mostly terrestrial life in the
Namib desert.
The position of Lanioturdus as the first offshoot in the
�core platysteirids� was a significant source of incongru-
ence between our data sets. Analyses undertaken with
the b-fibrinogen intron-5 sequences placed it as the
sister-group of the genus Batis, myoglobin intron-2 se-
quences placed it between Batis and the Dyaphoro-
phyia–Platysteira clade, and the ND2 sequences placed
it at the same position as the combined data set. The
SH test did not reject a sister-group relationship between
Batis and Lanioturdus (dL = 12.05, p = 0.12) nor did it
reject a sister-group relationship between Lanioturdus
and the Dyaphorophyia–Platysteira clade (dL = 13.49,
p = 0.09). A synapomorphic insertion of one nucleotide
within the large 24bp insertion that supports the mono-phyly of the �core platysteirids� seems to confirm the
monophyly of the Batis–Platysteira–Dyaphorophyia
clade but it could alternatively be interpreted as an auta-
pomorphic deletion in Lanioturdus. The placement of
Lanioturdus as the basal branch in the �core platysteirids�needs to be confirmed by further studies including addi-
tional representatives of the genera Batis, Dyaphoro-
phyia, and Platysteira.Species from these two latter genera were tradition-
ally placed together in Platysteira (Sibley and Monroe,
1990; Traylor, 1970) but Erard (1987) and Harris and
Franklin (2000) pointed out sufficient behavioral and
morphological differences for retaining two genera.
These clustered together with high bootstrap values in
most of our analyses but our taxonomic sampling does
not allow us to clarify the generic limits.Monophyly of the �core malaconotids� was well sup-
ported in all our analyses and is in agreement with the
insertion pattern of the Musculus flexor digitorum longus
muscle (Raikow et al., 1980). The other synapomorphy
proposed by the authors (presence of fluffy back feath-
ers) is also present in all members of the �core platysteir-ids� and even (more or less developed) in other corvid
groups (J. Fjeldsa, personal observation). This characterwas not considered by Raikow et al. (1980), as members
of the Platysteiridae were not included in their study. It
has not been fully documented how broadly this charac-
ter is shared, or whether its development, and adaptative
significance for courtship suggests homology.
Representatives of the core �malaconotids,� �platy-steirids,� and �vangids� formed a well-supported clade
(clade A on Fig. 1) in all our analyses. However, rela-tionships between these three main groups depended
on the analytical methods employed and were generally
poorly supported, so the exact sister-group of the �coremalaconotids� remains undetermined. The individual
analyses of the two nuclear introns showed that the
molecular divergence between the three main groups
was nearly identical to the divergence observed between
genera in the �core malaconotids.� These results agreewith those obtained by Sibley and Ahlquist (1990) in
their large DNA/DNA hybridization study where the
short internodes between the Clade A genera suggest a
rapid speciation burst leading to several lineages of very
divergent appearance. Evolution of foraging behavior,
and thus beak morphology, is particularly known for
its evolutionary plasticity (Cibois et al., 2001; Grant
et al., 2000; Irestedt et al., 2004; Pasquet et al., 1999).
4.2. Relationships within the �core malaconotids�
Nilaus is morphologically divergent from other repre-
sentatives of the �core malaconotids� because its tarsal
scutellation resembles that of Prionops and the nest
building is more Batis-like (Harris and Franklin, 2000;
Mayr, 1943). Our data as well as hind limb musculatureand DNA/DNA hybridization data nest it within the
�core malaconotids� (Raikow et al., 1980; Sibley and

J. Fuchs et al. / Molecular Phylogenetics and Evolution 33 (2004) 428–439 437
Ahlquist, 1990). Traylor (1970) argued that Nilaus is the
link between the �core malaconotids� and the �core platy-steirids,� but this also implies that the �core platysteirids�is the sister-group of the �core malaconotids,� a state-
ment that was not clearly supported by our data. Our
analyses placed it as the sister-group of clade H, but al-ways with low bootstrap values. The SH test did not re-
ject the placement of Nilaus as the first offshoot in the
�core malaconotids� nor did it reject its placement as
the sister-group of clade J (dL = 1.54, p = 0.31 and
dL = 2.38, p = 0.17, respectively). Its position relative
to clades H and J must thus be considered as unresolved.
The remaining malaconotid genera clustered together
in two primary clades that were always supported byhigh bootstrap/posterior probability values. The first
clade (Clade H) included Bocagia, Dryocopus, Malacon-
otus, and Tchagra and the second clade (Clade J) con-
sisted of Chlorophoneus, Laniarius, Rhodophoneus, and
Telophorus.
The genus Malaconotus comprises six little-known
large bush-shrikes, most of which are very locally distrib-
uted (relictual), with unique character combinations.Malaconotus was traditionally believed to be the closest
relative of Chlorophoneus due to their closely similar col-
ors (Hall et al., 1966; Urban et al., 1997). However,Mal-
aconotus differs from Chlorophoneus by the lack of
mouth spots (in nestlings) and by general communica-
tion behavior (Harris and Franklin, 2000). DNA–DNA
hybridization data suggested that Malaconotus is basal
relative to other members of the �core malaconotids� (Sib-ley and Ahlquist, 1990). A sister-group relationship be-
tween Chlorophoneus and Malaconotus was strongly
rejected by our data (SH test, dL = 54.19, p = 0.004).
Our analyses generate some new hypotheses about the
relationships of Malaconotus and suggest an association
with Dryoscopus, Bocagia, and Tchagra. However, the
relative position of Malaconotus within clade J depends
on the phylogenetic reconstruction method employed,and was always moderately supported.
The genus Dryoscopus is morphologically very uni-
form (six small species with black upperparts and white
or grayish underparts). Dryoscopus was monophyletic in
all our analyses, a result that was always highly sup-
ported, and lent further credence by a synapomorphic
deletion. Linear classifications placed it between Nilaus
and Tchagra (including Bocagia) (Rand, 1960; Sibleyand Monroe, 1990), suggesting a close link between Dry-
oscopus and Tchagra, whereas DNA/DNA hybridiza-
tion data could not resolve its relationships (Sibley
and Ahlquist, 1990). Our data also suggest that Tchagra
and Bocagia are very close to Dryoscopus. More taxa
and sequences from species within the genus may be nec-
essary to assess its exact sister-group (Malaconotus or
Bocagia–Tchagra).Due to similar plumage aspect, egg color, and dis-
play-flights, some authors merge Bocagia and Tchagra
in the genus Tchagra (Rand, 1960; Sibley and Monroe,
1990; Wolters, 1975–1982). However, Harris and Frank-
lin (2000) retained two genera and pointed out that Boc-
agia differs from Tchagra in several characters (sexual
dimorphism, eye color, tail color, communication
behavior, nest decoration, and vocalization). They sug-gested that Bocagia may be closer to Dryoscopus than
to Tchagra. This statement is clearly refuted by our data,
suggesting that the characters proposed by Harris and
Franklin (2000) for uniting Bocagia and Dryoscopus
are plesiomorphic within clade H. The diagnostic char-
acters proposed by Harris and Franklin (2000) for
retaining Bocagia are nearly all utilized in courtship dis-
plays and thus could have an accelerated rate of evolu-tion due to sexual selection. While the molecular
evidence seems to support merging Bocagia with Tcha-
gra, a denser taxon sampling is desirable to better quan-
tify sister-group relationships and generic limits.
The genusTchagra (sensu stricto) consists of four open
habitat species of very uniform plumage pattern. Its
monophyly was always well supported (high bootstrap
values and two synapomorphic deletions) by our data.The two species we lack in our taxon sampling (T. jamesi
and T. tchagra) are believed to form a superspecies with
T. australis (Sibley and Monroe, 1990). This assumption
is emphasized by the fact that a hybrid between T. aus-
tralis and T. jamesi has been described (Keith and
Twomey, 1968). However, inclusions of T. jamesi and
T. tchagra in a further study are necessary to determine
the taxonomic status of T. jamesi and eventually to quan-tify the amount of gene flow between the two taxa.
The second clade (clade J) consisted of the genera
Chlorophoneus, Laniarius, Rhodophoneus, and Telopho-
rus. This clade was supported by high bootstrap values
and a synapomorphic deletion (nine nucleotides). An
ethological synapomorphy (body bobbing while calling)
also supports this clade (Harris and Franklin, 2000).
The genus Laniarius comprises 16 species that areuniform in shape and proportions. Although colors
are variable, monophyly was never questioned and the
only contradictory hypotheses concern intra-generic
relationships (Harris and Franklin, 2000; Smith et al.,
1991). Our data also strongly support monophyly of
Laniarius (high bootstrap values and a synapomorphic
deletion of one nucleotide). Laniarius is traditionally
considered closer to Telophorus and Chlorophoneus thanto other representatives of the �core malaconotids� (Har-
ris and Franklin, 2000). Our data suggest that the closest
relatives of the genus Laniarius are the genera Chloro-
phoneus, Rhodophoneus, and Telophorus.
Representatives of the three latter genera formed a
clade in all our analyses but always with moderate to
low bootstrap support. However, we detected support
for monophyly because all its members share a synapo-morphic insertion of one nucleotide. The genus Chloro-
phoneus is not monophyletic (SH test, dL = 39.05,

438 J. Fuchs et al. / Molecular Phylogenetics and Evolution 33 (2004) 428–439
p = 0.009) in our analyses due to C. dohertyi being more
closely related to the genera Rhodophoneus and Telopho-
rus than to other Chlorophoneus species. This result is in
agreement with several morphological characters such
as toe fusion, presence of the Musculus iliofemoralis
externus muscle, presence of a black pectoral bar, andecological differences (Clark, 1981; Raikow et al.,
1980), also supported at the molecular level. We propose
to merge the undergrowth species of the genus Chloro-
phoneus (sensu Howard and Moore, 2003) (dohertyi
and viridis—including quadricolor) and Rhodophoneus
in Telophorus, and to keep the forest, woodland, and
bushland species in the genus Chlorophoneus (remaining
species). Monophyly of the latter genus (sensu stricto)needs to be tested because one of its members, the Crit-
ically Endangered Mount Kupe Bush-shrike (Chloro-
phoneus kupeensis), is quite different from other
Chlorophoneus species and some characters suggest that
it might be related to the �core vangids� (Harris and
Franklin, 2000). These latter authors argue that Chloro-
phoneus is more closely related to Rhodophoneus due to
the presence of mouth spots in nestlings, nest structure,and egg coloration, although some behavioral and mor-
phological data suggest a close relationship between
Rhodophoneus and Telophorus. The characters proposed
by Harris and Franklin (2000) are certainly a melting
pot of plesiomorphic and synapomorphic characters,
but in all cases they support the close affinities of the
genera Chlorophoneus, Rhodophoneus, and Telophorus.
Acknowledgments
We are very grateful to J.M. Bates and D. Willard
(Field Museum of Natural History, Chicago) and C. Co-
hen (FitzPatrick Institute, University of Cape Town) for
providing some of the samples we used. To Annie Tillier,
Celine Bonillo, and Josie Lambourdiere who kindly pro-vided technical help during laboratory works at MNHN,
and to Francois Baillon (IRD) who helped E. Pasquet
while in Cameroon doing field work. Laboratory work
at MNHN was supported by the �Service Commun de
Systematique Moleculaire,� IFR CNRS 101, MNHN.
R.C.K.B. thanks the Skye Foundation and Charitable
trust for providing funding for this project while at the
FMNH. The first version of the manuscript benefitedfrom the comments of two anonymous referees.
References
Baker, R.H., DeSalle, R., 1997. Multiple sources of character
information and the phylogeny of Hawaiian Drosophilids. Sys-
tematic Biology 46, 654–673.
Baker, R.H., Yu, X.B., DeSalle, R., 1998. Assessing the relative
contribution of molecular and morphological characters in simul-
taneous trees. Molecular Phylogenetics and Evolution 9, 427–436.
Barker, F.K., 2004. Monophyly and relationships of wrens (Aves:
Troglodytidae): a congruence analysis of heterogeneous mitochon-
drial and nuclear DNA sequence data. Molecular Phylogenetics
and Evolution 31, 486–504.
Barker, F.K., Barrowclough, G.F., Groth, J.G., 2002. A phylogenetic
hypothesis for passerine birds: taxonomic and biogeographic
implications of an analysis of nuclear DNA sequence data.
Proceedings of the Royal Society of London Series B 269, 295–308.
Bates, J.M., Hackett, S.J., Goerck, J.M., 1999. High levels of
mitochondrial DNA differentiation in two lineages of antbirds
(Drymophila and Hypocnemis). The Auk 116, 1093–1106.
Beecher, W.J., 1953. A phylogeny of the oscines. The Auk 70 (3), 270–
333.
Benson, C.W., 1985. Vangas. In: Poyser, T., Poyser, A.D. (Eds.), A
Dictionary of Birds. Calton, England, p. 619.
Benson, C.W., Brooke, R.K., Dowsett, R.J., Irwin, M.P.S., 1971. The
Birds of Zambia. Collins, London.
Bremer, K., 1994. Branch support and tree stability. Cladistics 10, 295–
304.
Cibois, A., Schulenberg, T.S., Pasquet, E., 2001. An endemic radiation
of Malagasy songbirds is revealed by mitochondrial DNA sequence
data. Evolution 55 (6), 1198–1206.
Cicero, C., Johnson, N.K., 2002. Phylogeny and character evolution in
the Empidonax group of Tyrant-flycatcher (Aves: Tyrannidae): a
test of W.E. Lanyon�s hypothesis using mtDNA sequences.
Molecular Phylogenetics and Evolution 22 (2), 289–302.
Clark Jr., G.A., 1981. Toe fusion in Oscines. Wilson Bulletin 93 (1),
67–76.
Desjardins, P., Morais, R., 1990. Sequence and gene organization of
the chicken mitochondrial genome a novel gene order in higher
vertebrates. Journal of Molecular Biology 212, 599–634.
Dorst, J., 1960. Les caracteres de la scutellation du tarse chez les
Vangides. L�Oiseau et RFO 30 (1), 32–44.
Erard, C., 1987. Ecologie et comportement des gobe-mouches (Aves:
Muscicapinae, Platysteirinae, Monarchidae). Memoire du Museum
National d�Histoire Naturelle Serie A 146, 1–234.
Ericson, P.G.P., Chritidis, L., Cooper, A., Irestedt, M., Jackson, J.,
Johansson, U.S., Norman, J.A., 2002a. A Gondwana origin of
passerine birds supported by DNA sequences of the endemic New
Zealand wrens. Proceedings of the Royal Society of London Series
B 269, 235–241.
Ericson, P.G.P., Christidis, L., Irestedt, M., Norman, J.A., 2002b.
Systematic affinities of the lyrebirds (Passeriformes: Menura), with
a novel classification of the major groups of passerine birds.
Molecular Phylogenetics and Evolution 25, 53–62.
Ericson, P.G.P., Irestedt, M., Johansson, U.S., 2003. Evolution,
biogeography, and patterns of diversification in passerine birds.
Journal of Avian Biology 34, 3–15.
Felsenstein, J., 1985. Confidence limits on phylogenies: an approach
using the bootstrap. Evolution 39, 783–791.
Grant, P.R., Grant, B.R., Petren, K., 2000. The allopatric phase of
speciation: the sharped-beaked ground finch (Geospiza difficilis) on
the Galapagos Islands. Biological Journal of the Linnean Society
69, 287–317.
Hackett, S.J., 1996. Molecular phylogenetics and biogeography of
Tanagers in the genus Ramphocelus (Aves). Molecular Phylogenet-
ics and Evolution 5 (2), 368–382.
Hall, B.P., Moreau, R.E., Galbraith, I.C.J., 1966. Polymorphism and
parallelism in the African Bush-shrikes of the genus Malaconotus
(including Chlorophoneus). The Ibis 108, 161–182.
Harris, T., Arnott, G., 1988. Shrikes of Southern Africa. Struik
Winchester, Cape Town.
Harris, T., Franklin, K., 2000. Shrikes and Bush-Shrikes including
Wood-Shrikes, Helmet-Shrikes, Flycatcher-Shrikes, Philentomas,
Batises and Wattle-Eyes. Helm Editions, London.
Heslewood, M.M., Elphinstone, M.S., Tidemann, S.C., Baverstock,
P.R., 1998. Myoglobin intron variation in the Gouldian Finch

J. Fuchs et al. / Molecular Phylogenetics and Evolution 33 (2004) 428–439 439
Erythrura gouldiae assessed by temperature gradient gel electro-
phoresis. Electrophoresis 19, 142–151.
Hillis, D.M., Bull, J.J., 1993. An empirical test of bootstrapping as a
method for assessing confidence in phylogenetic analysis. System-
atic Biology 42, 182–192.
Howard, R., Moore, A., 2003. A Complete Checklist of the Birds of
the World, third ed. Helm Editions, London.
Huelsenbeck, J.P., Rannala, B., 1997. Phylogenetic methods come of
age: testing hypotheses in an evolutionary context. Science 276,
227–232.
Huelsenbeck, J.P., Ronquis, F., Hall, B., 2001. MrBayes: Bayesian
inference of phylogeny. Bioinformatics 17, 754–755.
Irestedt, M., Fjeldsa, J., Ericson, P.G.P., 2004. Phylogenetic relation-
ships of woodcreepers (Aves: Dendrocolaptinae)—incongruence
between molecular and morphological data. Journal of Avian
Biology 35, 280–288.
Johanson, U.S., Ericson, P.G.P., 2003. Molecular support for a sister
group relationship between Pici and Galbulae (Piciformes sensu
Wetmore 1960). Journal of Avian Biology 34, 184–197.
Keith, S., Twomey, A., 1968. New distributional records of some East
African birds. The Ibis 110, 237–548.
Mayr, E., 1943. What genera belong to the family Prionopidae. The
Ibis 85, 216–218.
Morony Jr., J.J., Bock, W.J., Farrand Jr., J., 1975. Reference List of
the Birds of the World. American Museum of Natural History,
New York.
Pasquet, E., Cibois, A., Baillon, F., Erard, C., 1999. Relationships
between the ant-trushes Neocossyphus and the flycatcher trushes
Stizorhina and their position relative to Myadestes, Entomodestes
and some other Turdidae. Journal of Zoological Systematics and
Evolutionary Research 37 (4), 177–185.
Pocock, T.N., 1966. Contributions to the osteology of African birds.
The Ostrich 6 (Suppl.), 83–94.
Posada, D., Crandall, K.A., 1998. ModelTest 3.04: testing the model
of DNA substitution. Bioinformatics 14, 817–818.
Raikow, R.J., Polumbo, P.J., Borecky, S.R., 1980. Appendicular
myology and relationships of the shrikes (Aves: Passeriformes:
Laniidae). Annals of Carnegie Museum 49, 131–152.
Rand, A.L., 1960. Family Laniidae. In: Mayr, E., Greenway Jr., J.C.
(Eds.), Check-List of Birds of the World A Continuation of the
Work of James L. Peters Volume IX. Museum of Comparative
Zoology, Cambridge, MA, pp. 309–365.
Schulenberg, T.S., 1993. Phylogeny of the Vangidae: inferences from
mitochondrial DNA. Proceedings of the VIII Pan-African Orni-
thological Congress, 23–28.
Shimodaira, H., Hasagawa, M., 1999. Multiple comparisons of Log-
likelihoods with applications to phylogenetic inference. Molecular
Biology and Evolution 16 (8), 1114–1116.
Sibley, C.G., Ahlquist, J.E., 1990. Phylogeny and Classification of
Birds: A Study in Molecular Evolution. Yale University Press, New
Haven, CT.
Sibley, C.G., Monroe, B.L., 1990. Distribution and Taxonomy of the
Birds of the World. Yale University Press, New Haven, CT.
Slade, R.W., Moritz, C., Heideman, A., Hale, P.T., 1993. Rapid
assessment of single-copy nuclear DNA variation in diverse species.
Molecular Ecology 2, 359–373.
Smith, E.F.G., Arctander, P., Fjeldsa, J., Amir, O.G., 1991. A new
species of shrike (Laniidae: Laniarius) from Somalia, verified by
DNA sequence data from the only known individual. The Ibis 133
(2), 227–235.
Sorenson, M.D., 1999. TreeRot ver. 2. Boston University, Boston.
Sorenson, M.D., Ast, J.C., Dimcheff, D.E., Yuri, T., Mindell, D.P.,
1999. Primers for a PCR-based approach to mitochondrial genome
sequencing in birds and other vertebrates. Molecular Phylogenetics
and Evolution 12, 105–114.
Sorenson, M.D., Payne, R.B., 2001. A single origin of brood
parasitism in African finches: implications for host-parasite
coevolution. Evolution 55, 2550–2567.
Sorenson, M.D., Quinn, T.W., 1998. Numts: a challenge for avian
systematics and population biology. The Auk 115, 214–221.
Swofford, D.L., 2001. PAUP: Phylogenetic Analysis Using Parsi-
mony (and other methods) 4.0b10. Sinauer Associates, Sunder-
land, MA.
Traylor, M.T., 1970. Notes on African muscicapidae. The Ibis 112,
395–397.
Urban, E.K., Fry, C.H., Keith, S. (Eds.), 1997. The Birds of Africa,
vol. 5. Academic Press, London.
Urban, E.K., Fry, C.H., Keith, S. (Eds.), 2000. The Birds of Africa,
vol. 6. Academic Press, London.
Wolters, H.E., 1975–1982. Die Vogelarten der Erde. Paul Parey,
Hamburg and Berlin.
Yamagishi, S., Honda, M., Eguchi, K., Thorstrom, R., 2001. Extreme
endemic radiation of the Malagasy vangas (Vangidae). Journal of
Molecular Evolution 53 (1), 39–46.




























