Embed Size (px)
Citation preview

Ozone-triggered rapid stomatal response involves theproduction of reactive oxygen species, and is controlledby SLAC1 and OST1
Triin Vahisalu1,2, Irina Puzorjova2, Mikael Brosche1, Ervin Valk2, Martin Lepiku2, Heino Moldau2, Priit Pechter2,
Yuh-Shuh Wang2, Ove Lindgren2, Jarkko Salojarvi1, Mart Loog2, Jaakko Kangasjarvi1 and Hannes Kollist2,*
1Division of Plant Biology, Department of Biosciences, University of Helsinki, PO Box 65 (Viikinkaari 1), FI-00014 Helsinki,
Finland, and2Institute of Technology, University of Tartu, Nooruse 1, 50411 Tartu, Estonia
Received 26 November 2009; revised 16 January 2010; accepted 20 January 2010; published online 12 March 2010.*For correspondence (fax +37 2737 4900; e-mail [email protected]).
SUMMARY
The air pollutant ozone can be used as a tool to unravel in planta processes induced by reactive oxygen species
(ROS). Here, we have utilized ozone to study ROS-dependent stomatal signaling. We show that the ozone-
triggered rapid transient decrease (RTD) in stomatal conductance coincided with a burst of ROS in guard cells.
RTD was present in 11 different Arabidopsis ecotypes, suggesting that it is a genetically robust response. To
study which signaling components or ion channels were involved in RTD, we tested 44 mutants deficient in
various aspects of stomatal function. This revealed that the SLAC1 protein, essential for guard cell plasma
membrane S-type anion channel function, and the protein kinase OST1 were required for the ROS-induced fast
stomatal closure. We showed a physical interaction between OST1 and SLAC1, and provide evidence that
SLAC1 is phosphorylated by OST1. Phosphoproteomic experiments indicated that OST1 phosphorylated
multiple amino acids in the N terminus of SLAC1. Using TILLING we identified three new slac1 alleles where
predicted phosphosites were mutated. The lack of RTD in two of them, slac1-7 (S120F) and slac1-8 (S146F),
suggested that these serine residues were important for the activation of SLAC1. Mass-spectrometry analysis
combined with site-directed mutagenesis and phosphorylation assays, however, showed that only S120 was a
specific phosphorylation site for OST1. The absence of the RTD in the dominant-negative mutants abi1-1 and
abi2-1 also suggested a regulatory role for the protein phosphatases ABI1 and ABI2 in the ROS-induced
activation of the S-type anion channel.
Keywords: stomata, signaling, SLAC1, OST1, ozone, reactive oxygen species.
INTRODUCTION
Stomata, small pores on the aerial parts of plants, control
CO2 influx for photosynthesis and water vapor loss. They
also restrict the entry of ozone (O3) – a major air pollutant
with an increasingly negative impact on crop yields, global
carbon fixation (Hopkin, 2007) and climate change (Sitch
et al., 2007). Ozone degrades immediately to reactive
oxygen species (ROS) in the apoplastic space of plant cells,
and has therefore been used as a tool to study the signaling
role of the apoplastic ROS (Kangasjarvi et al., 2005;
Wrzaczek et al., 2009). ROS are involved in the regulation of
abscisic acid (ABA)- (Lee et al., 1999; Pei et al., 2000),
ethylene- (Desikan et al., 2006), methyl jasmonate-
(Munemasa et al., 2007) and salicylic acid-mediated (Mori
et al., 2001) stomatal signaling. The rapid induction of ROS
during CO2-induced stomatal closure has also been shown
(Kolla et al., 2007). Collectively, ROS are central intermediate
signaling components in plant guard cells, and it is likely that
ozone is a useful tool for the study of ROS-dependent
stomatal signaling.
Stomatal guard cells are among the most studied and best
understood plant signaling systems, yet there remain con-
siderable gaps in the understanding of the signaling that
leads to stomatal movements in response to different
stimuli. For example, the importance of guard cell anion
channels as central regulators of stomatal closure was
demonstrated 20 years ago (Keller et al., 1989; Schroeder
442 ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd
The Plant Journal (2010) 62, 442–453 doi: 10.1111/j.1365-313X.2010.04159.x

and Hagiwara, 1989), but the protein essential for guard cell
anion channel functioning, SLAC1, was identified only very
recently (Negi et al., 2008; Vahisalu et al., 2008). SLAC1 is
essential for stomatal closure in response to ABA, CO2, O3,
light–dark transitions and humidity change, and by second-
ary messengers Ca2+, H2O2 and NO. However, the signaling
cascades upstream of SLAC1, which require the capturing of
very early and probably transient responses, are as yet
unexplored.
Most experiments addressing the molecular details of
guard cell signaling have been performed with epidermal
peels or isolated guard cells. The preparation procedure(s)
are likely to introduce unwanted effects, including an
elevated production of ROS as a result of damage. Thus,
using intact plants and minimum handling of the plant
would help to define the function of ROS in stomatal
regulation more clearly. The simple application of ozone to
intact plants offers the possibility to control the concentra-
tion and duration of the exposure precisely. We have
constructed a gas-exchange system where eight soil-grown
Arabidopsis plants can be enclosed in individual flow-
through exposure vessels, in a non-invasive manner (Kollist
et al., 2007). Using this system we have shown that as little
as 150 nl l)1 of O3 triggers a rapid transient decrease (RTD)
in stomatal conductance (Kollist et al., 2007). The decrease
was induced within a few minutes of O3 exposure, but the
stomata reopened again despite the continuous presence of
ozone. The recovery suggests that the closure was not a
result of physical ozone damage, but instead reflects the
biological action of ROS formed from ozone breakdown in
the apoplast, transduced through a signaling cascade.
The lack of ozone-triggered RTD in the ABA-insensitive
mutant abi2-1 (Kollist et al., 2007), carrying a dominant-
negative mutation in the type-2C protein phosphatase ABI2,
suggests a role for protein phosphorylation in O3/ROS-
induced stomatal signaling. Murata et al. (2001) have also
shown that H2O2-induced stomatal closure was impaired in
the abi2-1 mutant. H2O2-induced stomatal closure was also
disrupted in the recessive ABA-insensitive mutant gca2
(Pei et al., 2000). On the contrary, in abi1-1, another ABA-
insensitive dominant-negative mutant of the protein phos-
phatase ABI1 (Murata et al., 2001), and in mutants of the
protein kinase OPEN STOMATA 1 (OST1) (also referred to as
SRK2E and Snf1-related protein kinase 2.6, SnRK2.6), a
positive regulator of ABA-induced stomatal closure (Mustilli
et al., 2002), H2O2-induced stomatal closure was not dis-
rupted, suggesting a role for these proteins between ABA
perception and ROS production.
We have shown that activation of S-type anion channels is
required for ROS-induced stomatal signaling, as ozone-
triggered RTD was absent in the S-type anion channel
mutant slac1 (Vahisalu et al., 2008). However, other struc-
tural and signaling components involved in relaying the
ROS signal from apoplast to stomatal movements have not
been identified. During recent years, the molecular identities
of many other guard cell transport proteins have been
established (for a review see Pandey et al., 2007; Ward et al.,
2008). Testing the characteristics of ozone-triggered RTD in
plant lines carrying mutated versions of proteins involved
in stomatal regulation could help to understand their role in
ROS-dependent processes.
Here we have explored the ozone-triggered RTD further.
We show the time and concentration dependence of the
process, and provide evidence that stomatal closure coin-
cides with the elevated burst of ROS in guard cells. This
suggests that RTD is induced by the ROS triggered by the
application of ozone. By analyzing RTD in several mutants
carrying mutations in proteins shown to be involved in
stomatal regulation, we show that OST1, ABI1, ABI2 and
SLAC1 are regulators of the ROS-induced rapid stomatal
closure. We demonstrate physical interaction between
SLAC1 and OST1, and provide evidence that SLAC1 is
phosphorylated by OST1.
RESULTS
The ozone-triggered RTD is dependent on exposure time
and ozone concentration
Applying 250 nl l)1 of ozone induced a 40% decrease in
stomatal conductance in wild-type (WT) Arabidopsis Col-0
plants within 5–10 min of exposure, followed by reopening
to the pre-exposure level within the next 40 min (Figure 1a).
The commonly observed sustained ozone-induced decrease
in stomatal conductance (Ahlfors et al., 2004) was visible
90 min after ozone onset. The same stomatal behavior was
seen in the 10 other ecotypes tested (Table S1). To further
elucidate the relationship between the duration of ozone
exposure and the decrease in stomatal conductance, Ara-
bidopsis Col-0 plants were treated with 250 nl l)1 of ozone
for 30, 70, 180, 360 and 720 s. Already a 30-s pulse of
250 nl l)1 ozone caused a clearly detectable decrease in
stomatal conductance (Figure 1b). The decrease reached a
maximum with 180 s of ozone, and longer exposures did not
decrease the conductance any further. To address the effect
of ozone concentration, we applied 3-min ozone pulses with
concentrations ranging from 50 to 600 nl l)1 (Figure 1c). The
response can be separated into three segments: ‘no
response’, ‘response’ and ‘saturation’. In the ‘no response’
segment, essentially no decrease in stomatal conductance
was observed. The threshold for the ‘response’ segment was
80 nl l)1 of ozone (95% confidence interval, shown by the
dashed lines). After the threshold, the decrease in stomatal
conductance increased approximately linearly, by 0.15% per
additional nl l)1 of ozone, with 95% confidence intervals of
�0.04% per nl l)1. The ‘saturated’ segment, where the
decrease in stomatal conductance reached its maximum,
was obtained with concentrations higher than 434 nl l)1 of
ozone.
Ozone-triggered stomatal responses 443
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 442–453
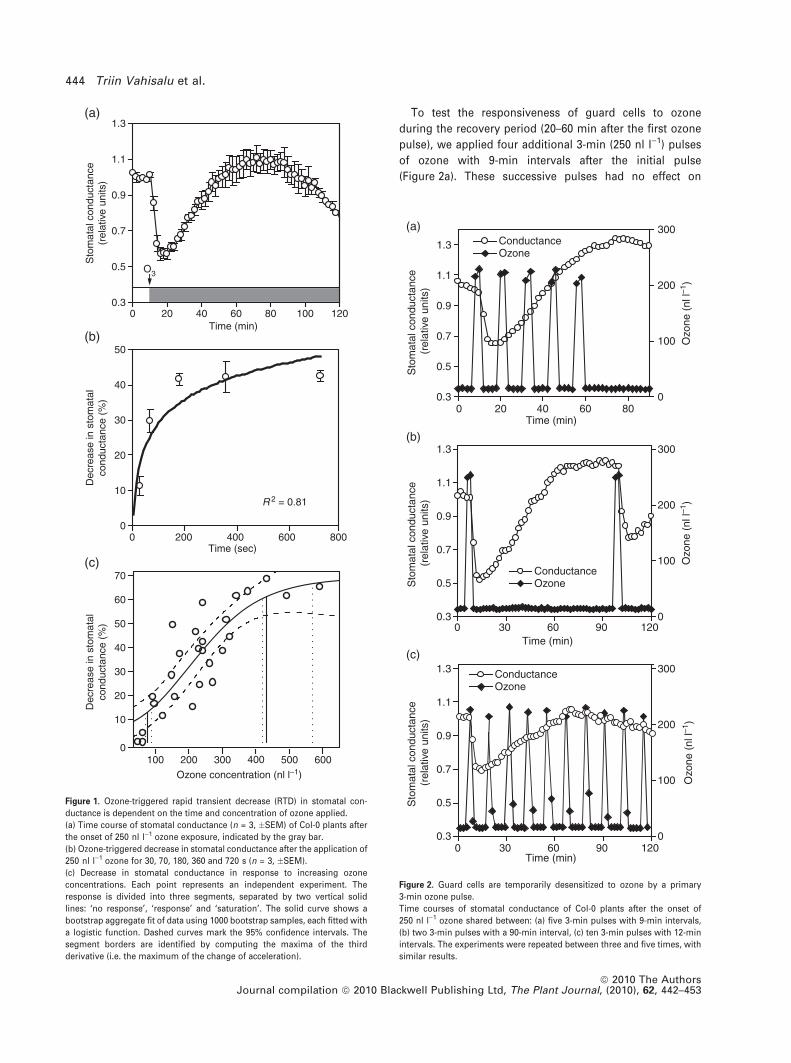
To test the responsiveness of guard cells to ozone
during the recovery period (20–60 min after the first ozone
pulse), we applied four additional 3-min (250 nl l)1) pulses
of ozone with 9-min intervals after the initial pulse
(Figure 2a). These successive pulses had no effect on
R 2 = 0.81
0
10
20
30
40
50
0 200 400 600 800Time (sec)
Dec
reas
e in
sto
mat
alco
nduc
tanc
e (%
)
Ozone concentration (nl l–1)
0
10
20
30
40
50
60
70
200 300 500 600100 400
Dec
reas
e in
sto
mat
alco
nduc
tanc
e (%
)
0.3
0.5
0.7
0.9
1.1
1.3
0 20 40 60 80 100 120Time (min)
Sto
mat
al c
ondu
ctan
ce(r
elat
ive
units
)
O3
(a)
(b)
(c)
Figure 1. Ozone-triggered rapid transient decrease (RTD) in stomatal con-
ductance is dependent on the time and concentration of ozone applied.
(a) Time course of stomatal conductance (n = 3, �SEM) of Col-0 plants after
the onset of 250 nl l)1 ozone exposure, indicated by the gray bar.
(b) Ozone-triggered decrease in stomatal conductance after the application of
250 nl l)1 ozone for 30, 70, 180, 360 and 720 s (n = 3, �SEM).
(c) Decrease in stomatal conductance in response to increasing ozone
concentrations. Each point represents an independent experiment. The
response is divided into three segments, separated by two vertical solid
lines: ‘no response’, ‘response’ and ‘saturation’. The solid curve shows a
bootstrap aggregate fit of data using 1000 bootstrap samples, each fitted with
a logistic function. Dashed curves mark the 95% confidence intervals. The
segment borders are identified by computing the maxima of the third
derivative (i.e. the maximum of the change of acceleration).
Sto
mat
al c
ondu
ctan
ce
(rel
ativ
e un
its)
Ozo
ne (
nl l–1
)
0.3
0.5
0.7
0.9
1.1
1.3
0 30 60 90 120Time (min)
0
100
200
300
ConductanceOzone
Sto
mat
al c
ondu
ctan
ce
(rel
ativ
e un
its)
0.3
0.5
0.7
0.9
1.1
1.3
0 20 40 60 80Time (min)
0
ConductanceOzone
100
200
300
Ozo
ne (
nl l–1
)ConductanceOzone
Sto
mat
al c
ondu
ctan
ce
(rel
ativ
e un
its)
Ozo
ne (
nl l–1
)
0.3
0.5
0.7
0.9
1.1
1.3
0 30 60 90 120Time (min)
0
100
200
300
(a)
(b)
(c)
Figure 2. Guard cells are temporarily desensitized to ozone by a primary
3-min ozone pulse.
Time courses of stomatal conductance of Col-0 plants after the onset of
250 nl l)1 ozone shared between: (a) five 3-min pulses with 9-min intervals,
(b) two 3-min pulses with a 90-min interval, (c) ten 3-min pulses with 12-min
intervals. The experiments were repeated between three and five times, with
similar results.
444 Triin Vahisalu et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 442–453
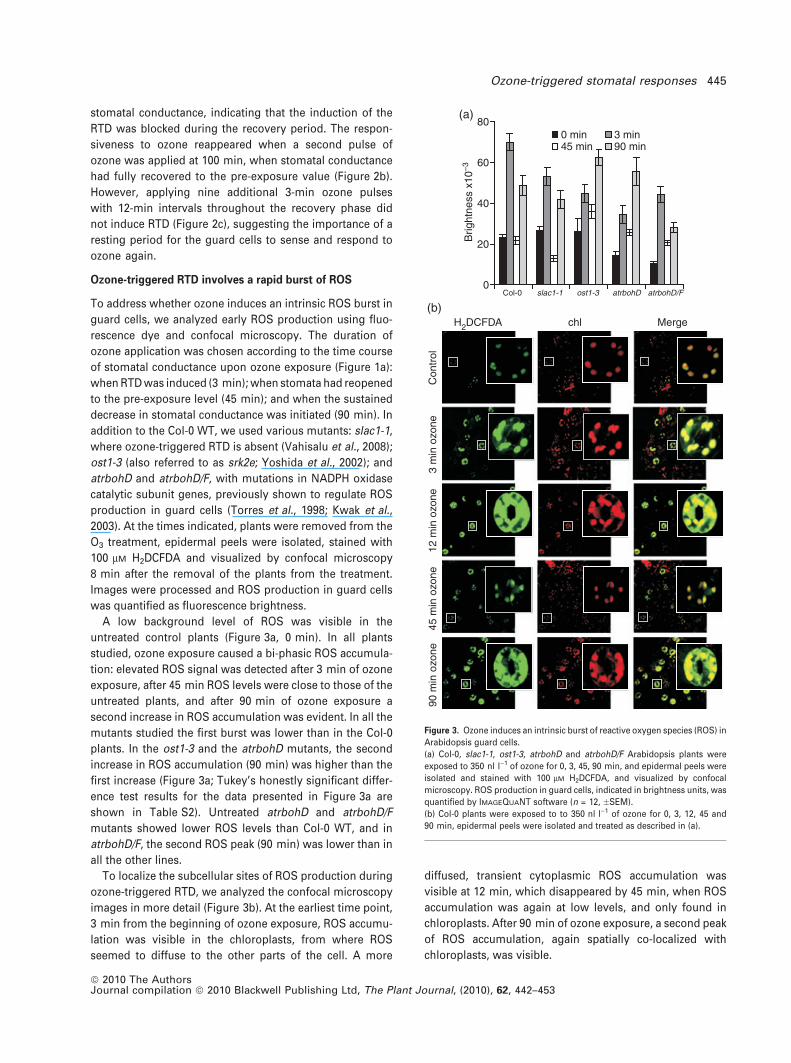
stomatal conductance, indicating that the induction of the
RTD was blocked during the recovery period. The respon-
siveness to ozone reappeared when a second pulse of
ozone was applied at 100 min, when stomatal conductance
had fully recovered to the pre-exposure value (Figure 2b).
However, applying nine additional 3-min ozone pulses
with 12-min intervals throughout the recovery phase did
not induce RTD (Figure 2c), suggesting the importance of a
resting period for the guard cells to sense and respond to
ozone again.
Ozone-triggered RTD involves a rapid burst of ROS
To address whether ozone induces an intrinsic ROS burst in
guard cells, we analyzed early ROS production using fluo-
rescence dye and confocal microscopy. The duration of
ozone application was chosen according to the time course
of stomatal conductance upon ozone exposure (Figure 1a):
when RTD was induced (3 min); when stomata had reopened
to the pre-exposure level (45 min); and when the sustained
decrease in stomatal conductance was initiated (90 min). In
addition to the Col-0 WT, we used various mutants: slac1-1,
where ozone-triggered RTD is absent (Vahisalu et al., 2008);
ost1-3 (also referred to as srk2e; Yoshida et al., 2002); and
atrbohD and atrbohD/F, with mutations in NADPH oxidase
catalytic subunit genes, previously shown to regulate ROS
production in guard cells (Torres et al., 1998; Kwak et al.,
2003). At the times indicated, plants were removed from the
O3 treatment, epidermal peels were isolated, stained with
100 lM H2DCFDA and visualized by confocal microscopy
8 min after the removal of the plants from the treatment.
Images were processed and ROS production in guard cells
was quantified as fluorescence brightness.
A low background level of ROS was visible in the
untreated control plants (Figure 3a, 0 min). In all plants
studied, ozone exposure caused a bi-phasic ROS accumula-
tion: elevated ROS signal was detected after 3 min of ozone
exposure, after 45 min ROS levels were close to those of the
untreated plants, and after 90 min of ozone exposure a
second increase in ROS accumulation was evident. In all the
mutants studied the first burst was lower than in the Col-0
plants. In the ost1-3 and the atrbohD mutants, the second
increase in ROS accumulation (90 min) was higher than the
first increase (Figure 3a; Tukey’s honestly significant differ-
ence test results for the data presented in Figure 3a are
shown in Table S2). Untreated atrbohD and atrbohD/F
mutants showed lower ROS levels than Col-0 WT, and in
atrbohD/F, the second ROS peak (90 min) was lower than in
all the other lines.
To localize the subcellular sites of ROS production during
ozone-triggered RTD, we analyzed the confocal microscopy
images in more detail (Figure 3b). At the earliest time point,
3 min from the beginning of ozone exposure, ROS accumu-
lation was visible in the chloroplasts, from where ROS
seemed to diffuse to the other parts of the cell. A more
diffused, transient cytoplasmic ROS accumulation was
visible at 12 min, which disappeared by 45 min, when ROS
accumulation was again at low levels, and only found in
chloroplasts. After 90 min of ozone exposure, a second peak
of ROS accumulation, again spatially co-localized with
chloroplasts, was visible.
0Col-0 atrbohD atrbohD/Fost1-3slac1-1
20
40
60
80
Brig
htne
ss x
10–3
0 min 3 min45 min 90 min
H2DCFDA chl
Con
trol
3 m
in o
zone
12 m
in o
zone
45 m
in o
zone
90 m
in o
zone
Merge
(a)
(b)
Figure 3. Ozone induces an intrinsic burst of reactive oxygen species (ROS) in
Arabidopsis guard cells.
(a) Col-0, slac1-1, ost1-3, atrbohD and atrbohD/F Arabidopsis plants were
exposed to 350 nl l)1 of ozone for 0, 3, 45, 90 min, and epidermal peels were
isolated and stained with 100 lM H2DCFDA, and visualized by confocal
microscopy. ROS production in guard cells, indicated in brightness units, was
quantified by IMAGEQUANT software (n = 12, �SEM).
(b) Col-0 plants were exposed to to 350 nl l)1 of ozone for 0, 3, 12, 45 and
90 min, epidermal peels were isolated and treated as described in (a).
Ozone-triggered stomatal responses 445
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 442–453

Rapid ozone-triggered RTD is a specific process
A 3-min pulse of ozone triggered RTD in all 11 Arabidopsis
ecotypes tested (Table S1), suggesting that RTD is a genet-
ically robust response. Many mutants deficient in ABA sig-
naling and ROS production in guard cells, in response to
different stimuli, have been identified (Li et al., 2006). We
tested several of them to elucidate the role of these proteins
in the apoplastic ROS-induced RTD (Table S1).
The a and b subunits of the heterotrimeric G protein have
been shown to be necessary for the ozone-induced oxidative
burst in guard cells (Joo et al., 2005). The a subunit is also
involved in regulating stomatal closure in response to ABA
(Wang et al., 2001). Stomata of the Ga mutant gpa1-4, the Gbmutant agb1-2 and the double mutant agb1/gpa1 responded
to ozone like the WT (Table S1), suggesting that heterotri-
meric G proteins were not involved in the signaling from
apoplastic ROS to the activation of anion fluxes under our
conditions.
The AtrbohD, AtrbohE and AtrbohF NADPH oxidase
subunits have a role in guard cell ABA signal transduction
(Kwak et al., 2003). However, atrbohD, atrbohE and atrbohF
single mutants, and all double mutant combinations,
responded to ozone pulse like the WT (Table S1), suggesting
that the O3-derived apoplastic ROS formation can function-
ally mimic the apoplastic ROS production by NADPH
oxidases in guard cells.
The plant stress hormones ethylene, jasmonic acid,
salicylic acid and ABA are important regulators of ozone
responses (Kangasjarvi et al., 2005), and have also been
shown to be regulators of guard cell signaling (Lee et al.,
1999; Pei et al., 2000; Mori et al., 2001; Desikan et al., 2006;
Munemasa et al., 2007). We tested mutants deficient in
biosynthesis and/or essential signal components for each of
these hormones – none of them were required for RTD
(Table S1). Exceptionally, some components of ABA signal-
ing, but not ABA biosynthesis, were required for RTD (see
below).
A mutant deficient in the protein kinase HT1, known to
control stomatal movements in response to CO2 (Hashim-
oto et al., 2006), responded to ozone like the WT (Table
S1). The mutant of the plasma membrane-localized ATP
binding cassette transporter AtMRP5, shown to have
impaired Ca2+ activation of guard cell anion channels
(Suh et al., 2007), had a normal response to ozone (Table
S1), as did the cpk3 and cpk6 (calcium-dependent protein
kinase 3 and 6) mutants required for the activation of
S-type anion currents by ABA and calcium (Mori et al.,
2006) (Table S1). The ABA-insensitive mutant gca2 (Pei
et al., 2000) also responded to ozone like the WT (Table
S1), indicating that the activity of GCA2 is not required for
the apoplastic ROS-induced stomatal closure. These results
suggest that the apoplastic ROS-induced stomatal move-
ments did not operate through the same set of regulatory
components through which CO2, and Ca2+-dependent
signaling act.
There were, however, mutants that did not display the
O3-triggered RTD. We have previously shown that the ABA-
insensitive protein phosphatase type-2C mutant abi2-1
completely lacked the ozone-triggered RTD (Kollist et al.,
2007). Here (Table S1), we show that, in addition to abi2-1,
the ozone-triggered RTD was also absent in abi1-1, ost1-1
and ost1-3, and in two K+-channel mutants the kinetics of
RTD was altered.
Ozone-triggered RTD is modulated by K+ channels, and
requires functional SLAC1, OST1, ABI1 and ABI2
Stomatal movements are facilitated by the activity of ion
channels and transporters in the plasma membrane and
vacuolar membrane of guard cells (Pandey et al., 2007). In
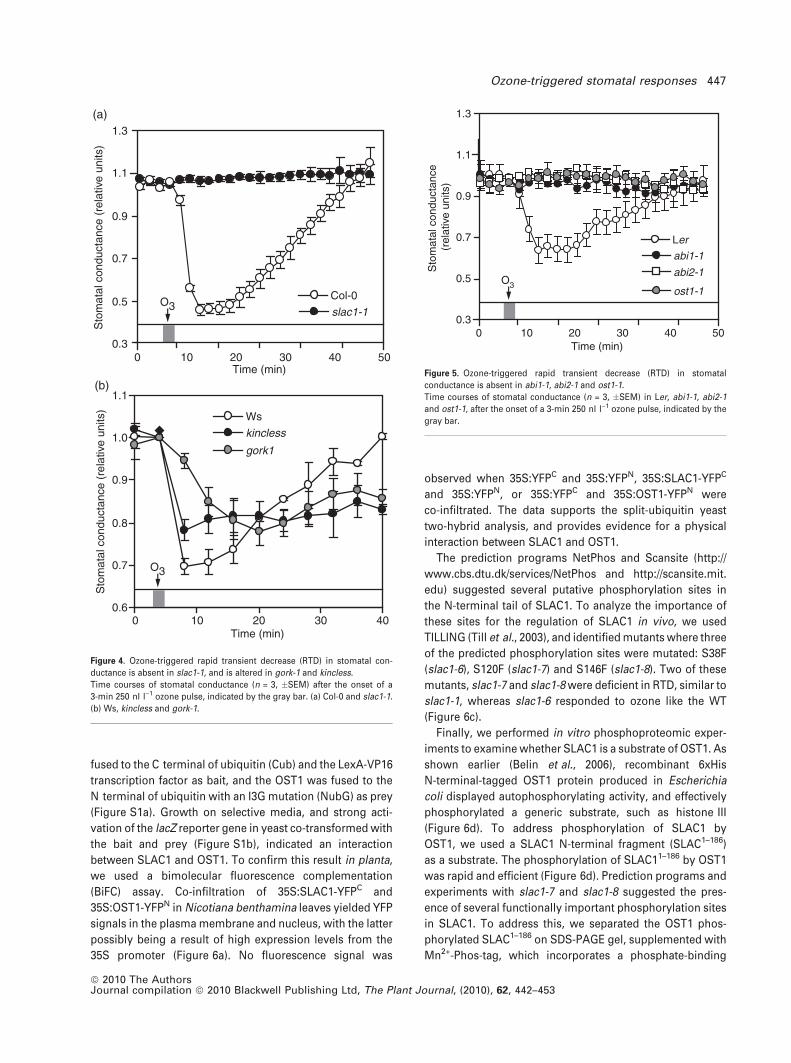
addition to the S-type ion channel mutant slac1-1, where the
ozone-triggered RTD was absent (Figure 4a), the patterns of
RTD were also different in two potassium channel mutants:
gork-1, where the guard cell plasma membrane K+ outward
rectifying channel GORK activity is fully suppressed (Hosy
et al., 2003), and kincless, where the inward rectifying K+
current is abolished (Lebaudy et al., 2008) (Figure 4b). In the
Ws-2 WT, ozone triggered an RTD of 30% within 4 min of
exposure. The response was strongly delayed in gork-1,
where the RTD was only 5% within 4 min, and a maximal
decrease of 23% was only achieved 16 min after the begin-
ning of the exposure (Figure 4b). In kincless, ozone caused a
22% RTD within 4 min, but reopening was almost com-
pletely absent; after 40 min, conductance had fully recov-
ered in the WT, whereas in kincless no recovery was seen
(Figure 4b). These results suggest that, in addition to initial
anion currents (SLAC1), subsequent GORK-mediated K+ flux
is required for the rapid decrease in stomatal conductance.
During the recovery period, additional ozone pulses had no
effect (Figure 2a). This implies that the activity of the inward-
rectifying potassium channel was not directly affected by the
apoplastic ROS induced by O3.
In order to elucidate the role of protein phosphorylation in
the ozone-triggered RTD upstream of SLAC1, we analyzed
RTD in abi1-1, abi2-1 and ost1-1 in more detail (Figure 5).
Whereas a 3-min O3 pulse triggered RTD in both Col-0 and
Ler WTs, stomata of all three protein phosphorylation
mutants were insensitive to the O3 pulse, suggesting that
the protein kinase OST1 and the phosphatases ABI1/ABI2 are
required for the SLAC1-dependent RTD.
Protein interaction, genetic and phosphoproteomic analysis
suggest that SLAC1 is regulated by OST1
The absence of ozone-triggered RTD in ost1 mutants
immediately suggested a model where SLAC1 is regulated
by OST1. Therefore, we first addressed whether SLAC1 and
OST1 interact in a split-ubiquitin membrane yeast two-
hybrid system (Johnsson and Varshavsky, 1994). SLAC1 was
446 Triin Vahisalu et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 442–453
fused to the C terminal of ubiquitin (Cub) and the LexA-VP16
transcription factor as bait, and the OST1 was fused to the
N terminal of ubiquitin with an I3G mutation (NubG) as prey
(Figure S1a). Growth on selective media, and strong acti-
vation of the lacZ reporter gene in yeast co-transformed with
the bait and prey (Figure S1b), indicated an interaction
between SLAC1 and OST1. To confirm this result in planta,
we used a bimolecular fluorescence complementation
(BiFC) assay. Co-infiltration of 35S:SLAC1-YFPC and
35S:OST1-YFPN in Nicotiana benthamina leaves yielded YFP
signals in the plasma membrane and nucleus, with the latter
possibly being a result of high expression levels from the
35S promoter (Figure 6a). No fluorescence signal was
observed when 35S:YFPC and 35S:YFPN, 35S:SLAC1-YFPC
and 35S:YFPN, or 35S:YFPC and 35S:OST1-YFPN were
co-infiltrated. The data supports the split-ubiquitin yeast
two-hybrid analysis, and provides evidence for a physical
interaction between SLAC1 and OST1.
The prediction programs NetPhos and Scansite (http://
www.cbs.dtu.dk/services/NetPhos and http://scansite.mit.
edu) suggested several putative phosphorylation sites in
the N-terminal tail of SLAC1. To analyze the importance of
these sites for the regulation of SLAC1 in vivo, we used
TILLING (Till et al., 2003), and identified mutants where three
of the predicted phosphorylation sites were mutated: S38F
(slac1-6), S120F (slac1-7) and S146F (slac1-8). Two of these
mutants, slac1-7 and slac1-8 were deficient in RTD, similar to
slac1-1, whereas slac1-6 responded to ozone like the WT
(Figure 6c).
Finally, we performed in vitro phosphoproteomic exper-
iments to examine whether SLAC1 is a substrate of OST1. As
shown earlier (Belin et al., 2006), recombinant 6xHis
N-terminal-tagged OST1 protein produced in Escherichia
coli displayed autophosphorylating activity, and effectively
phosphorylated a generic substrate, such as histone III
(Figure 6d). To address phosphorylation of SLAC1 by
OST1, we used a SLAC1 N-terminal fragment (SLAC1–186)
as a substrate. The phosphorylation of SLAC11–186 by OST1
was rapid and efficient (Figure 6d). Prediction programs and
experiments with slac1-7 and slac1-8 suggested the pres-
ence of several functionally important phosphorylation sites
in SLAC1. To address this, we separated the OST1 phos-
phorylated SLAC1–186 on SDS-PAGE gel, supplemented with
Mn2+-Phos-tag, which incorporates a phosphate-binding
0 10 20 30 40 500.3
0.5
0.7
0.9
1.1
1.3
Sto
mat
al c
ondu
ctan
ce (
rela
tive
units
)
slac1-1
Col-0O3
Sto
mat
al c
ondu
ctan
ce (
rela
tive
units
)
0.6
0.7
0.8
0.9
1.0
1.1
0 10 20 30 40
Ws
kincless
gork1
O3
Time (min)
Time (min)
(a)
(b)
Figure 4. Ozone-triggered rapid transient decrease (RTD) in stomatal con-
ductance is absent in slac1-1, and is altered in gork-1 and kincless.
Time courses of stomatal conductance (n = 3, �SEM) after the onset of a
3-min 250 nl l)1 ozone pulse, indicated by the gray bar. (a) Col-0 and slac1-1.
(b) Ws, kincless and gork-1.
0 10 20 30 40 500.3
0.5
0.7
0.9
1.1
1.3
Sto
mat
al c
ondu
ctan
ce
(rel
ativ
e un
its)
abi1-1
ost1-1
abi2-1
Ler
O3
Time (min)
Figure 5. Ozone-triggered rapid transient decrease (RTD) in stomatal
conductance is absent in abi1-1, abi2-1 and ost1-1.
Time courses of stomatal conductance (n = 3, �SEM) in Ler, abi1-1, abi2-1
and ost1-1, after the onset of a 3-min 250 nl l)1 ozone pulse, indicated by the
gray bar.
Ozone-triggered stomatal responses 447
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 442–453
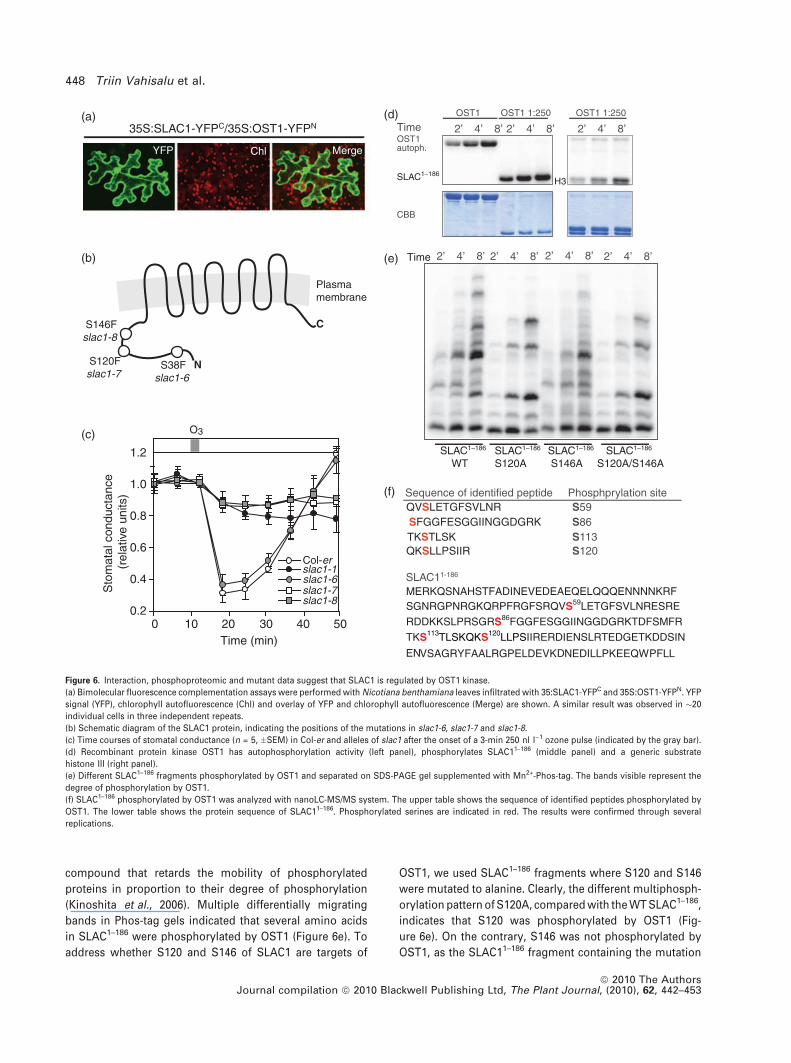
compound that retards the mobility of phosphorylated
proteins in proportion to their degree of phosphorylation
(Kinoshita et al., 2006). Multiple differentially migrating
bands in Phos-tag gels indicated that several amino acids
in SLAC1–186 were phosphorylated by OST1 (Figure 6e). To
address whether S120 and S146 of SLAC1 are targets of
OST1, we used SLAC1–186 fragments where S120 and S146
were mutated to alanine. Clearly, the different multiphosph-
orylation pattern of S120A, compared with the WT SLAC1–186,
indicates that S120 was phosphorylated by OST1 (Fig-
ure 6e). On the contrary, S146 was not phosphorylated by
OST1, as the SLAC11–186 fragment containing the mutation
35S:SLAC1-YFPC/35S:OST1-YFPN
YFP Chl Merge
(a)
N
C
Plasma membrane
S38Fslac1-6
S120Fslac1-7
S146Fslac1-8
(b)
(c)
(d)
(e)
(f)
Sto
mat
al c
ondu
ctan
ce
(rel
ativ
e un
its)
Time (min)
0.2
0.4
0.6
0.8
1.0
1.2
0 10 20 30 40 50
O3
slac1-1
slac1-7
Col-er
slac1-6
slac1-8
SLAC1–186
WTSLAC1–186
S120ASLAC1–186
S146A SLAC1–186
S120A/S146A
Time
S59S86
S120S113TKSTLSK
QVSLETGFSVLNRSequence of identified peptide Phosphprylation site
SFGGFESGGIINGGDGRK
QKSLLPSIIR
Time 2’ 8’4’ 2’ 8’4’ 2’ 8’4’
2’ 8’4’2’ 8’4’2’ 8’4’2’ 8’4’
OST1 OST1 1:250 OST1 1:250
H3
OST1autoph.
CBB
SLAC11-186
MERKQSNAHSTFADINEVEDEAEQELQQQENNNNKRFSGNRGPNRGKQRPFRGFSRQVS59LETGFSVLNRESRE
RDDKKSLPRSGRS86FGGFESGGIINGGDGRKTDFSMFR
TKS113TLSKQKS120LLPSIIRERDIENSLRTEDGETKDDSIN
ENVSAGRYFAALRGPELDEVKDNEDILLPKEEQWPFLL
SLAC1–186H3
Figure 6. Interaction, phosphoproteomic and mutant data suggest that SLAC1 is regulated by OST1 kinase.
(a) Bimolecular fluorescence complementation assays were performed with Nicotiana benthamiana leaves infiltrated with 35:SLAC1-YFPC and 35S:OST1-YFPN. YFP
signal (YFP), chlorophyll autofluorescence (Chl) and overlay of YFP and chlorophyll autofluorescence (Merge) are shown. A similar result was observed in �20
individual cells in three independent repeats.
(b) Schematic diagram of the SLAC1 protein, indicating the positions of the mutations in slac1-6, slac1-7 and slac1-8.
(c) Time courses of stomatal conductance (n = 5, �SEM) in Col-er and alleles of slac1 after the onset of a 3-min 250 nl l)1 ozone pulse (indicated by the gray bar).
(d) Recombinant protein kinase OST1 has autophosphorylation activity (left panel), phosphorylates SLAC11–186 (middle panel) and a generic substrate
histone III (right panel).
(e) Different SLAC1–186 fragments phosphorylated by OST1 and separated on SDS-PAGE gel supplemented with Mn2+-Phos-tag. The bands visible represent the
degree of phosphorylation by OST1.
(f) SLAC1–186 phosphorylated by OST1 was analyzed with nanoLC-MS/MS system. The upper table shows the sequence of identified peptides phosphorylated by
OST1. The lower table shows the protein sequence of SLAC11–186. Phosphorylated serines are indicated in red. The results were confirmed through several
replications.
448 Triin Vahisalu et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 442–453

S146A, or the combined mutations of S120A and S146A,
revealed similar phosphorylation patterns as the WT SLAC1–186
and SLAC1–186 containing S120A, respectively. Additionally,
we analyzed the SLAC11–186 fragment phosphorylated by
OST1 with mass spectrometry. This indicated that Ser59,
Ser86, Ser113 and Ser120, but not Ser146, were phosphor-
ylated by OST1 (Figure 6f and Figure S2).
Taken together, these data indicate that regulation of
SLAC1 by OST1 may involve the phosphorylation of multi-
ple amino acids, and suggests that Ser120 of SLAC1 is one of
the functionally important phosphosites.
DISCUSSION
Reactive oxygen species have a demonstrated role in guard
cell signaling in response to various external and internal
factors (Pei et al., 2000; Kolla et al., 2007). Apoplastic ROS,
formed as a result of the activity of the NADPH oxidases
(Kwak et al., 2003), are essential components in guard cell
ABA signaling. Ozone is known to degrade to various ROS in
the apoplast, thus it is likely that the ozone-triggered RTD
addressed in this study is a result of the action of the apo-
plastic ROS. This ROS from ozone degradation would act in
a similar manner as the ROS produced by NADPH oxidase
activity in ABA- or methyl jasmonate-induced stomatal
closure (Kwak et al., 2003; Munemasa et al., 2007). Hence,
ozone can be used as a tool to simplify the very complex
regulatory network in guard cells, and enables the study of
the role of ROS alone. The fast kinetics of RTD (Figure 1b)
implies that either the ROS are perceived directly in the
apoplast, followed by a rapid signal transmission to guard
cells, or that the ROS formed in the apoplast translocate
(most likely after dismutation to H2O2, as O�2 is impermeable
through a biological membrane) to the inside of guard cells,
where they immediately elicit the response.
Ozone triggered a biphasic ROS accumulation in guard
cells, where chloroplasts were the major source for ROS
formation (Figure 3). Previously, it has been shown that
ozone-induced ROS production was initiated from guard cell
chloroplasts, followed by ROS production in guard cell
membranes, which required NADPH oxidases encoded by
the AtrbohD and AtrbohF genes (Joo et al., 2005). The time
point studied with atrbohD and atrbohF by Joo et al. (2005)
was 1 h. In our experiments, the first phase of O3-induced
ROS accumulation (detected after 3 min) was significantly
reduced, but not abolished, in atrbohD and atrbohD/F
mutants (Figure 3a). This suggests the presence of several
sources for initial ROS production, for example, cell wall
peroxidases, in addition to the NADPH oxidases. Recently, it
has been shown that OST1 can phosphorylate AtrbohF, and
possibly regulates its activity (Sirichandra et al., 2009). The
ost1-3 mutant had lower initial ROS production (Figure 3a),
suggesting that regulation of Atrboh-mediated ROS produc-
tion by OST1 could be functionally relevant during initial
ozone responses. Interestingly, the second ROS peak was
lower in the atrbohD/F double mutant (Figure 3a), which
suggests that the membrane-bound NADPH oxidases have
an influence on the second peak of ROS production. It is
noteworthy that the timing of the first and second peak of
ROS accumulation (Figure 3) coincided with the fast and
slow decrease in stomatal conductance triggered by ozone
(Figure 1a). The decline in ROS production to control levels
at 45 min, despite the plant being continuously exposed to
ozone, favors a model for enzymatic control of ROS
production. However, whether this temporal coincidence
also has a mechanistic grounding needs to be studied
further, as well as the role of chloroplastic ROS in guard cell
signaling.
After the perception of the apoplastic ROS, the signal is
rapidly transduced to the guard cell anion channel SLAC1, as
RTD was absent in the slac1 mutant. However, RTD is not a
result of the activity of the plasma membrane anion channel
only: in the K+ efflux channel mutant gork-1, the ozone
response was delayed, suggesting that after the activation of
the anion fluxes, K+ fluxes through GORK were also required
for the rapid decrease in stomatal conductance (Figure 4b).
Thus, apparently the signal from apoplastic ozone/ROS is
first passed to SLAC1, and the subsequent activation of
anion fluxes and membrane depolarization are required for
GORK activation. The lack of recovery in stomatal conduc-
tance in the kincless mutant indicates a central role for K+
inward-rectifying channels during the recovery period
(Figure 4b).
An intriguing feature of the ozone-triggered RTD is that
during the recovery period further ozone pulses had no
effect (Figure 2b). This suggests that the primary ozone
pulse changes the status of the voltage-dependent channel
assembly in a manner that temporarily desensitizes stomata
to further ozone pulses. Blocking and desensitization have
been shown to occur for several ion channels under different
treatments (Roelfsema and Prins, 1997; Raschke et al., 2003).
Additionally, blue light treatment rapidly induces Ca2+
transients (Baum et al., 1999), and a 30–120-min recovery
period is required to elicit the response again. Similarly,
both H2O2 (Price et al., 1994) and ozone (Clayton et al., 1999;
Evans et al., 2005) induced rapid transient increases in
cytosolic Ca2+, and a recovery period of a few hours was
required before the response could be elicited again. One of
the possible causes of desensitization may be channel
inactivation (Hedrich et al., 1990; Pei et al., 1998), or if Ca2+
is part of the RTD, it might need to be returned to its original
resting state concentration and/or localization.
Several different cellular regulatory and signaling pro-
cesses in stomatal guard cells appear to converge at the
activation of anion fluxes, where SLAC1 is a central compo-
nent. ROS also play a role in several of these regulatory
cascades. The absence or presence of RTD in various guard
cell signaling and ion channel mutants (Table S1) allowed us
to dissect components that are required for apoplastic ROS
Ozone-triggered stomatal responses 449
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 442–453

responses in guard cells. Collectively, our results suggest
that the signal pathway from apoplastic ROS to guard cell
anion channel activation is at least partly independent from
previously described CO2 and ABA signaling pathways.
The use of protein kinase and phosphatase inhibitors has
previously shown that protein phosphorylation is involved
in the activation of anion channels (Schmidt et al., 1995;
Grabov et al., 1997; MacRobbie, 1998; Pei et al., 2000). Some
protein kinases involved in these responses have been
identified. The protein kinase HT1, a central component in
the regulation of stomatal movements by CO2, was not
required for the ROS activation of SLAC1. The activity of the
Ca2+-dependent protein kinases CPK3 and CPK6, which have
been shown to be involved in the Ca2+-dependent ABA
activation of S-type anion currents (Mori et al., 2006),
showed a normal RTD response. Thus, these protein kinas-
es, and other proteins, such as GCA2, previously shown to
be involved in CO2- or ABA- and Ca2+-dependent stomatal
regulation, are not components required for the apoplastic
ROS-induced regulation of stomatal movement. The OXI1
kinase, necessary for some ROS signaling processes (Rentel
et al., 2004), was not required for RTD either (Table S1). In
addition, ABA itself is not part of the signal cascade, as the
ABA-deficient aba1-3 mutant showed WT responses.
However, we have previously shown that in addition to
slac1 (Vahisalu et al., 2008), the ABA-insensitive protein
phosphatase type 2C mutant, abi2-1, completely lacked the
ozone-induced decrease in stomatal conductance (Kollist
et al., 2007). The ozone-triggered RTD was also missing in a
second dominant-negative mutant of the type-2C protein
phosphatases, ABI1, and in two mutant alleles of the OST1,
demonstrating that these proteins are essential in the
apoplastic ROS-induced RTD (Figure 5). It has been shown
recently that ABI1, ABI2 and OST1 interact physically both in
yeast two-hybrid and in planta assays, and that these and
other phosphatases act as negative regulators of OST1 via
PYR/PYL/RCAR proteins (Fujii et al., 2009; Umezawa et al.,
2009; Vlad et al., 2009). Similarly, our genetic data suggested
that ABI1 and ABI2 could be regulators of OST1 in ROS-
induced stomatal closure through dephosphorylation
(Figures 5 and 7). Using the split-ubiquitin yeast two-hybrid
and BiFC assays, we showed a physical interaction between
OST1 and SLAC1 (Figure S1b and 6a). The physical interac-
tion between SLAC1 and OST1, and the requirement for
OST1-dependent phosphorylation for the activation of
SLAC1, was also very recently demonstrated by two other
groups (Geiger et al., 2009; Lee et al., 2009).
Our phosphoproteomic experiments proved that OST1
was able to phosphorylate multiple amino acids of SLAC1,
including Ser120, but not Ser146 (Figure 6d–f), further
supporting that OST1 is responsible for SLAC1 activation.
This also suggests that Ser146 could be the target for a
different protein kinase. An alternative explanation for the
absence of RTD in slac1-8 (Figure 6c) could be a conforma-
tional change of SLAC1 protein, as in the slac1-8 mutant,
Ser146 is substituted with phenylalanine, which is a hydro-
phobic and considerably larger amino acid. SLAC1 has been
shown to be involved in the regulation of stomatal closure in
response to many factors, such as CO2, darkness, humidity,
ABA and Ca2+ (Vahisalu et al., 2008). Thus, it would be of
great interest to study the stomatal responses of slac1-7 and
slac1-8 to other stimuli, in order to address whether different
phosphorylation patterns of SLAC1 exist in response to
different stimuli.
Collectively, our data suggest a model for RTD (Figure 7)
where OST1 activates SLAC1 via phosphorylation, and
suggests the importance of S120 for this regulation. ABI1
and ABI2 keep OST1 inactive by dephosphorylation (Umez-
awa et al., 2009; Vlad et al., 2009). Possibly, the protein
phosphatase activity of ABI1 and ABI2 could also be directed
towards SLAC1: this should be addressed in further studies.
Recently, OST1 has been shown to phosphorylate and
reduce the activity of the inward K+ channel KAT1 (Sato
et al., 2009). Thus, OST1 would activate SLAC1 and at the
same time inactivate KAT1, leading to faster stomatal
closure.
Using ozone we show that the last steps of the ROS-
induced signaling cascade, leading to the activation of
SLAC1 and the induction of stomatal closure, could be as
Figure 7. Schematic model of events during ozone-triggered rapid transient
decrease (RTD) in stomatal conductance.
Ozone induces reactive oxygen species (ROS) production in the guard cells
(1). This leads to the activation of OST1 (2). The lack of RTD in abi1-1 and
abi2-1 suggests that the protein phosphatases ABI1 and ABI2 regulate OST1
activity (Umezawa et al., 2009). Our data suggest that several amino acid
residues of the N-terminal tail of SLAC1 are phosphorylated by OST1, and
highlight the functional importance of S120 (3). Phosphorylation leads to the
activation of S-type anion channels and anion efflux (A) out) from guard cells
(4), which in turn causes plasma membrane depolarization, and activation of
outward-rectifying K+ channel GORK and K+ efflux from guard cells (5). The
overall efflux of anions and K+ contributes to the loss of guard cell turgor,
leading to stomatal closure. For reopening, the activity of K+ uptake channels
are required (6).
450 Triin Vahisalu et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 442–453

simple as the recently described minimal signaling pathway
for regulating ABA-induced gene expression (Fujii et al.,
2009). Furthermore, the two pathways share several com-
mon components: ABI1, ABI2 and OST1. Our next challenge
is to find out how the cells perceive ROS and translate this
into activation of OST1.
EXPERIMENTAL PROCEDURES
Plant material and growth conditions
For the whole-plant gas-exchange experiments, 24–26-day-oldplants were used. Plants were grown as described previously(Kollist et al., 2007). The ost1-1 mutant is in the Ler background(Mustilli et al., 2002). The ost1-3 is a T-DNA knock-out of OST1kinase in the Col-0 background (originally referred to as srk2e;Yoshida et al., 2002). For clarity, we refer to srk2e as ost1-3throughout this report. The source and identity of other mutantsused in the study are given in Table S1.
Whole-plant stomatal conductance measurements and
fluorescence microscopy
The Arabidopsis whole-rosette gas-exchange measurement detailswere described previously (Kollist et al., 2007). Prior to ozoneexposure, plants were acclimated in the measuring cuvettes for atleast 1 h. Plants were exposed to 350 nl l)1 ozone for 3, 12, 45 or90 min. Abaxial epidermal peels were isolated and loaded with100 lM H2DCFDA in 10 mM Tris–HCl, pH 7.2, for 5 min in darkness,and were washed with 10 mM Tris–HCl, pH 7.2. Clean-air controlpeels were isolated and loaded after acclimation in gas-exchangecuvettes.
ROS production was visualized by Nikon TE2000-U C1 confocalmicroscope (Nikon, http://www.nikon.com) using excitation at488 nm and emission at 530 nm. Images were processed usingNikon EZ-C1 FREEVIEWER software (gold version 3.30; Nikon).Brightness values of individual guard cell pairs were obtained aftercorrecting for the brightness of epidermal cells with IMAGEQUANT -v4.2a (Molecular Dynamics, now part of GE Healthcare, http://www.gelifesciences.com).
Isolation of TILLING lines
New ethyl methanesulphonate mutants slac1-6 (S38F), slac1-7(S120F) and slac1-8 (S146F) were identified through TILLING (Tillet al., 2003; http://tilling.fhcrc.org). Details are described in Appen-dix S1.
Split-ubiquitin membrane yeast two-hybrid assay
The split-ubiquitin yeast two-hybrid assay was conducted using theDUALmembrane kit 3 (Dualsystems Biotech, http://www.dualsystems.com). Details are described in Appendix S1.
BiFC interaction experiments
The cDNA of SLAC1 and OST1 was cloned into the pSPYNE andpSPYCE vectors. BiFC experiments were performed using transienttransfection of N. benthamiana leaves with Agrobacteriumtumefaciens, as described by Voinnet et al. (2003), images wereacquired 48–72 h after transfection by confocal microscopy (alsosee Appendix S1).
Protein expression and purification
OST1 and SLAC1 N-terminal fragments encoding amino acids 1–186(SLAC11–186) cDNAs were cloned into the pQE-30 UA (Qiagen,
http://www.qiagen.com) and pET28a (Novagen, now part of Merck,http://www.merck4biosciences.com) vectors, respectively. 6xHIS-OST1 was expressed using the XL-1 blue E. coli strain (Strata-gene, http://www.stratagene.com). 6xHIS-SLAC11–186 variants wereexpressed in Rosetta (DE3) pLysS cells (Novagen). Recombinantproteins were purified using a Chelating Sepharose� FastFlow (Amersham, now part of GE Healthcare, http://www.gelifesciences.com) column chelated with CoCl2 (for details, see Appen-dix S1).
In vitro kinase assays and mass spectrometry
Proteins were separated by SDS-PAGE using a 12% (w/v) acrylam-ide gel or 10% (w/v) acrylamide gel supplemented with Phos-Tag(Kinoshita et al., 2006). Gels were stained with Coomassie brilliantblue R-250 (Sigma-Aldrich, http://www.sigmaaldrich.com), andincorporation of 32P to the proteins was detected and visualized byautoradiography (for further details see Appendix S1). NanoLC-MS/MS analysis for the mapping of OST1 phosphorylation sites inSLAC11–186 was carried out by using LTQ-Orbitrap (Thermo FisherScientific, http://www.thermofisher.com), equipped with a nano-spray source (Proxeon, http://www.proxeon.com) and a 1200 Seriesnano-LC system (Agilent Technologies, http://www.agilent.com)(for details, see Appendix S1).
ACKNOWLEDGEMENTS
We acknowledge several labs (listed in Table S1) for sharing theirmutants. Work in the labs of HK and ML was funded by the EstonianScience Foundation (grants 7763, 6766, 7869, 7361 and EMP24)and Estonian Ministry of Education and Research (themeSF0180071s07). Work in the JK’s lab was supported by the Academyof Finland Centre of Excellence program (2006–2011), MB wassupported by an Academy of Finland Post-Doctoral grant (deci-sion# 108760), and TV was supported by the Finnish GraduateSchool in Plant Biology.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the onlineversion of this article:Figure S1. The split-ubiquitin yeast two-hybrid assay suggests thatSLAC1 interacts with OST1 kinase.Figure S2. Mass spectrometry analysis suggests that OST1 phos-phorylates at least four serines in the SLAC1–186 fragment.Table S1. List of mutants tested for ozone-triggered rapid transientdecrease (RTD) in stomatal conductance.Table S2. Statistical analysis of data presented in Figure 3a (Tukey’shonestly significant difference test).Appendix S1. Supplementary experimental procedures.Please note: As a service to our authors and readers, this journalprovides supporting information supplied by the authors. Suchmaterials are peer-reviewed and may be re-organized for onlinedelivery, but are not copy-edited or typeset. Technical supportissues arising from supporting information (other than missingfiles) should be addressed to the authors.
REFERENCES
Ahlfors, R., Lang, S., Overmyer, K. et al. (2004) Arabidopsis RADICAL-
INDUCED CELL DEATH1 belongs to the WWE protein-protein interaction
domain protein family and modulates abscisic acid, ethylene, and methyl
jasmonate responses. Plant Cell, 16, 1925–1937.
Baum, G., Long, J.C., Jenkins, G.I. and Trewavas, A.J. (1999) Stimulation of
the blue light phototropic receptor NPH1 causes a transient increase in
cytosolic Ca2+. Proc. Natl Acad. Sci. USA, 96, 13554–13559.
Belin, C., de Franco, P.O., Bourbousse, C., Chaignepain, S., Schmitter, J.M.,
Vavasseur, A., Giraudat, J., Barbier-Brygoo, H. and Thomine, S. (2006)
Ozone-triggered stomatal responses 451
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 442–453

Identification of features regulating OST1 kinase activity and OST1 function
in guard cells. Plant Physiol. 141, 1316–1327.
Clayton, H., Knight, M.R., Knight, H., McAinsh, M.R. and Hetherington, A.M.
(1999) Dissection of the ozone-induced calcium signature. Plant J. 17, 575–
579.
Desikan, R., Last, K., Harrett-Williams, R., Tagliavia, C., Harter, K., Hooley, R.,
Hancock, J.T. and Neill, S.J. (2006) Ethylene-induced stomatal closure in
Arabidopsis occurs via AtrbohF-mediated hydrogen peroxide synthesis.
Plant J. 47, 907–916.
Evans, N.H., McAinsh, M.R., Hetherington, A.M. and Knight, M.R. (2005) ROS
perception in Arabidopsis thaliana: the ozone-induced calcium response.
Plant J. 41, 615–626.
Fujii, H., Chinnusamy, V., Rodrigues, A., Rubio, S., Antoni, R., Park, S.Y.,
Cutler, S.R., Sheen, J., Rodriguez, P.L. and Zhu, J.K. (2009). In vitro
reconstitution of an abscisic acid signalling pathway. Nature, doi:10.1038/
nature08599.
Geiger, D., Scherzer, S., Mumm, P. et al. (2009) Activity of guard cell anion
channel SLAC1 is controlled by drought-stress signaling kinase-phospha-
tase pair. Proc. Natl Acad. Sci. USA, 106, 21425–21430.
Grabov, A., Leung, J., Giraudat, J. and Blatt, M.R. (1997) Alteration of anion
channel kinetics in wild-type and abi1–1 transgenic Nicotiana benthamiana
guard cells by abscisic acid. Plant J. 12, 203–213.
Hashimoto, M., Negi, J., Young, J., Israelsson, M., Schroeder, J.I. and Iba, K.
(2006) Arabidopsis HT1 kinase controls stomatal movements in response
to CO2. Nat. Cell Biol. 8, 391–397.
Hedrich, R., Busch, H. and Raschke, K. (1990) Ca2+ and nucleotide dependent
regulation of voltage dependent anion channels in the plasma membrane
of guard cells. EMBO J. 9, 3889–3892.
Hopkin, M. (2007) Carbon sinks threatened by increasing ozone. Nature, 448,
396–397.
Hosy, E., Vavasseur, A., Mouline, K. et al. (2003) The Arabidopsis outward K+
channel GORK is involved in regulation of stomatal movements and plant
transpiration. Proc. Natl Acad. Sci. USA, 100, 5549–5554.
Johnsson, N. and Varshavsky, A. (1994) Split ubiquitin as a sensor of protein
interactions in vivo. Proc. Natl Acad. Sci. USA, 91, 10340–10344.
Joo, J.H., Wang, S., Chen, J.G., Jones, A.M. and Federoff, N.V. (2005) Different
signaling and cell death roles of heterotrimeric G protein a and b subunits in
the Arabidopsis oxidative stress response to ozone. Plant Cell, 17, 957–970.
Kangasjarvi, J., Jaspers, P. and Kollist, H. (2005) Signalling and cell death in
ozone-exposed plants. Plant Cell Environ. 28, 1021–1036.
Keller, U.B., Hedrich, R. and Raschke, K. (1989) Voltage dependent anion
channels in the plasma membrane of guard cells. Nature, 341, 450–452.
Kinoshita, E., Kinoshita-Kikuta, E., Takiyama, K. and Koike, T. (2006) Phos-
phate-binding tag, a new tool to visualize phosphorylated proteins. Mol.
Cell Proteomics, 5, 749–757.
Kolla, V.A., Vavasseur, A. and Raghavendra, A.S. (2007) Hydrogen peroxide
production is an early event during bicarbonate induced stomatal closure
in abaxial epidermis of Arabidopsis. Planta, 225, 1421–1429.
Kollist, T., Moldau, H., Rasulov, B., Oja, V., Ramma, H., Huve, K., Jaspers, P.,
Kangasjarvi, J. and Kollist, H. (2007) A novel device detects a rapid ozone-
induced transient stomatal closure in intact Arabidopsis and its absence in
abi2 mutant. Physiol. Plant. 129, 796–803.
Kwak, J.M., Mori, I.C., Pei, Z.-M., Leonhardt, N., Torres, M.A., Dangl, J.L.,
Bloom, R.E., Bodde, S., Jones, J.D.G. and Schroeder, J.I. (2003) NADPH
oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA sig-
naling in Arabidopsis. EMBO J. 22, 2623–2633.
Lebaudy, A., Hosy, E., Simonneau, T., Sentenac, H., Thibaud, J.B. and Dreyer,
I. (2008) Heteromeric K+ channels in plants. Plant J. 54, 1076–1082.
Lee, S., Choi, H., Suh, S., Doo, I.-S., Oh, K.-Y., Choi, E.J., Schroeder Taylor,
A.T., Low, P.S. and Lee, Y. (1999) Oligogalacturonic acid and chitosan
reduce stomatal aperture by inducing the evolution of reaction oxygen
species from guard cells of tomato and Commelina communis. Plant
Physiol. 121, 147–152.
Lee, S.C., Lan, W., Buchanan, B.B. and Luan, S. (2009) A protein kinase-
phosphatase pair interacts with an ion channel to regulate ABA signaling in
plant guard cells. Proc. Natl Acad. Sci. USA, 106, 21419–21424.
Li, S., Assmann, S.M. and Albert, R. (2006) Predicting essential components of
signal transduction networks: a dynamic model of guard cell abscisic sig-
naling. PLoS Biol. 4, 1732–1748.
MacRobbie, E.A.C. (1998) Signal transduction and ion channels in guard cells.
Philos. Trans. R. Soc. Lond. B, 353, 1475–1488.
Mori, I.C., Pinontoan, R., Kawano, T. and Muto, S. (2001) Involvement of
superoxide generation in salicylic acid-induced stomatal closure in Vicia
faba. Plant Cell Physiol. 42, 1383–1388.
Mori, I.C., Murata, Y., Yang, Y. et al. (2006) CDPKs CPK6 and CPK3 function in
ABA regulation of guard cell S-type anion- and Ca2+-permeable channels
and stomatal closure. PLoS Biol. 4, 1749–1762.
Munemasa, S., Oda, K., Watanabe-Sugimoto, M., Nakamura, Y., Shimoishi, Y.
and Murata, Y. (2007) The coronatine-insensitive 1 mutation reveals the
hormonal signaling interaction between abscisic acid and methyl jasmo-
nate in Arabidopsis guard cells. Specific impairment of ion channel acti-
vation and second messenger production. Plant Physiol. 143, 1398–1407.
Murata, Y., Pei, Z.-M., Mori, I.C. and Schroeder, J.I. (2001) Abscisic acid acti-
vation of plasma membrane Ca2) channels in guard cells requires cytosolic
NAD(P)H and is differentially disrupted upstream and downstream of
reactive oxygen species production in abi1–1 and abi2–1 protein phos-
phatase 2C mutants. Plant Cell, 13, 2513–2523.
Mustilli, A.-C., Merlot, S., Vavasseur, A., Fenzi, F. and Giraudat, J. (2002)
Arabidopsis OST1 protein kinase mediates the regulation of stomatal
aperture by abscisic acid and acts upstream of reactive oxygen species
production. Plant Cell, 14, 3089–3099.
Negi, J., Matsuda, O., Nagasawa, T., Oba, Y., Takahashi, H., Kawai-Yamada,
M., Uchimiya, H., Hashimoto, M. and Iba, K. (2008) CO2 regulator SLAC1
and its homologues are essential for anion homeostasis in plant cells.
Nature, 452, 483–486.
Pandey, S., Zhang, W. and Assmann, S.M. (2007) Roles of ion channels and
transporters in guard cell signal transduction. FEBS Lett. 581, 2325–2336.
Pei, Z.M., Baizabal-Aguirre, V.M., Allen, G.J. and Schroeder, J.I. (1998) A
transient outward-rectifying K+ channel current down-regulated by cyto-
solic Ca2+ in Arabidopsis thaliana guard cells. Proc. Natl Acad. Sci. USA, 95,
6548–6553.
Pei,Z.-M.,Murata,Y.,Benning,G.,Thomine, S.,Klusener,B.,Allen,G.J.,Grill,E.
and Schroeder, J.I. (2000) Calcium channels activated by hydrogen
peroxide mediate abscisic acid signalling in guard cells. Nature, 406, 731–
734.
Price, A.H., Taylor, A., Ripley, S.J., Griffiths, A., Trewavas, A.J. and Knight,
M.R. (1994) Oxidative signals in tobacco increase cytosolic calcium. Plant
Cell, 6, 1301–1310.
Raschke, K., Shabahang, M. and Wolf, R. (2003) The slow and the quick anion
conductance in whole guard cells: their voltage-dependent alternation, and
the modulation of their activities by abscisic acid and CO2. Planta, 217, 639–
650.
Rentel, M.C., Lecourieux, D., Ouaked, F. et al. (2004) OXI1 kinase is necessary
for oxidative burst-mediated signalling in Arabidopsis. Nature, 427, 858–
861.
Roelfsema, M.R.G. and Prins, H.B.A. (1997) Ion channels in guard cells of
Arabidopsis thaliana (L.) Heynh. Planta, 202, 18–27.
Sato, A., Sato, Y., Fukao, Y. et al. (2009) Threonine at position 306 of the KAT1
potassium channel is essential for channel activity and is a target
site for ABA-activated SnRK2/OST1/SnRK2.6 protein kinase. Biochem. J.
doi: 10.1042/BJ20091221.
Schmidt, C., Schelle, I., Liao, Y.J. and Schroeder, J.I. (1995) Strong regulation
of slow anion channels and abscisic acid signaling in guard cells by
phosphorylation and dephosphorylation events. Proc. Natl Acad. Sci. USA,
92, 9535–9539.
Schroeder, J.I. and Hagiwara, S. (1989) Cytosolic calcium regulates ion
channels in the plasma membrane of Vicia faba guard cells. Nature, 338,
427–430.
Sirichandra, C., Gu, D., Hu, H.C. et al. (2009) Phosphorylation of the Arabid-
opsis AtrbohF NADPH oxidase by OST1 protein kinase’’. FEBS Lett. 583,
2982–2986.
Sitch, S., Cox, P.M., Collins, W.J. and Huntingford, C. (2007) Indirect radiative
forcing of climate change through ozone effects on the land-carbon sink.
Nature, 448, 791–794.
Suh, S.J., Wang, Y.F., Frelet, A., Leonhardt, N., Klein, M., Forestier, C.,
Mueller-Roeber, B., Cho, M., Martinoia, E. and Schroeder, J.I. (2007) The
ATP binding cassette transporter AtMRP5 modulates anion and Ca2+
channel activities in Arabidopsis guard cells. J. Biol. Chem. 282, 1916–1924.
Till, B.J., Reynolds, S.H., Greene, A., Codomo, C.A., Enns, L.C., Johnson, J.E.,
Burtner, C., Odden, A.R., Young, K. and Taylor, N.E. (2003) Large-scale
discovery of induced point mutations with high-throughput TILLING.
Genome Res. 13, 524–530.
452 Triin Vahisalu et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 442–453

Torres, M.A., Onouchi, H., Hamada, S., Machida, C., Hammond-Kosac, K.E.
and Jones, J.D.G. (1998) Six Arabidopsis thaliana homologues of the hu-
man respiratory burst oxidase (gp91phox). Plant J. 14, 365–370.
Umezawa, T., Sugiyama, N., Mizoguchi, M., Hayashi, S., Myouga, F.,
Yamaguchi-Shinozaki, K., Ishihama, Y., Hirayama, T. and Shinozaki, K.
(2009) Type 2C protein phosphatases directly regulate abscisic acid-
activated protein kinases in Arabidopsis. Proc. Natl Acad. Sci. USA, 106,
17588–17593.
Vahisalu, T., Kollist, H., Wang, Y.F. et al. (2008) SLAC1 is required for plant
guard cell S-type anion channel function in stomatal signalling. Nature,
452, 487–491.
Vlad, F., Rubio, S., Rodrigues, A., Sirichandra, C., Belin, C., Robert, N., Leung,
J., Rodriguez, P.L., Lauriere, C. and Merlot, S. (2009) Protein phosphatases
2C regulate the activation of the Snf1-related kinase OST1 by abscisic acid
in Arabidopsis. Plant Cell, doi: 10.1105/tpc.109.069179.
Voinnet, O., Rivas, S., Mestre, P. and Baulcombe, D. (2003) An enhanced
transient expression system in plants based on suppression of gene
silencing by the p19 protein of tomato bushy stunt virus. Plant J. 33, 949–
956.
Wang, X.Q., Ullah, H., Jones, A.M. and Assmann, S.M. (2001) G protein reg-
ulation of ion channels and abscisic acid signaling in Arabidopsis guard
cells. Science, 292, 2070–2072.
Ward, J.M., Maser, P. and Schroeder, J.I. (2008) Plant ion channels: gene
families, physiology, and functional genomics analyses. Annu. Rev.
Physiol. 71, 59–82.
Wrzaczek, M., Brosche, M., Kollist, H. and Kangasjarvi, J. (2009) Arabidopsis
GRI is involved in the regulation of cell death induced by extracellular ROS.
Proc. Natl Acad. Sci. USA, 106, 5412–5417.
Yoshida, R., Hobo, T., Ichimura, K., Mizoguchi, T., Takahashi, F., Alonso, J.,
Ecker, J.R. and Shinozaki, K. (2002) ABA-activated SnRK2 protein kinase is
required for dehydration stress signaling in Arabidopsis. Plant Cell Physiol.
43, 1473–1483.
Ozone-triggered stomatal responses 453
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 442–453









![Stomatal index responses of Agrostis canina to CO2 and sulphur dioxide: implications for palaeo-[CO2] using the stomatal proxy](https://img.dokumen.tips/doc/110x75/6351f5c485f87b9f8602ac53/stomatal-index-responses-of-agrostis-canina-to-co2-and-sulphur-dioxide-implications.jpg)



















